Embed Size (px)
Citation preview

12
Biological Decomposition of Solid Wood
T. KENT KIRK
U.S. Department of Agriculture, Forest Service, Forest Products Laboratory,Madison WI 53705
ELLIS B. COWLING
School of Forest Resources, North Carolina State University,Raleigh, NC 27605
Decomposition of wood is an important part of thecarbon cycle of nature. Decomposition is caused byfungi, insects, and marine borers that use the wood asfood or shelter, or both. Lignin in wood provides a phys-ical barrier to enzymatic decomposition of cellulose andhemicelluloses. This barrier is breached mechanically byinsects and marine borers, biochemically by white- andsoft-rot fungi, and possibly by small nonenzyme cata-l y s t s i n t h e c a s e o f b r o w n - r o t f u n g i . C e l l u l o s eis degraded by endo- and exo-glucanases and β-gluco-sidases, hemicelluloses by endo-glycanases and glycosi-dases, lignin by nonspecific enzymes, and perhaps bynonenzymatic, oxidative agents. Rapid strength loss oc-curs with all decay fungi; but especially with brown-rotfungi. Strength loss due to insect attack is roughly pro-portional to the amount of wood removed. Fungal de-composition of wood can be prevented by keeping itbelow its fiber-saturation moisture content (approxi-mately 27% of its dry weight) and by using the heart-wood of naturally durable woods (species) or preser-vative-treated wood. Useful application of wood-decom-posing fungi is limited currently to production of ediblemushrooms. Potential applications include biologicalpulping, pretreatment for enzymatic conversion of woodto sugars, and waste treatment. Many aspects of woodbiodecomposition have not been researched adequately.
Millions of tons of wood are produced every year in theforests of the world. Observation, however, tells us thatthe sum-total of wood upon the surface of the earth re-mains fairly constant from year to year and from century
This chapter not subject to U.S. copyright.Published 1984, American Chemical Society

456 THE CHEMISTRY OF SOLID WOOD
to century. We must, therefore, conclude that there aredestructive agencies at work by which millions of tons ofwood are destroyed annually. Regarded in this light theproblem of what these destructive agencies are, and howthey act, becomes of general scientific and economic in-terest.
A. H. R. Buller,preeminent mycologist ,1906 (1)
B U L L E R R E C O G N I Z E D T H A T W O O D I S T H E M O S T A B U N D A N T O R G A N I C M A T E -
R I A L on ea r th . He a l so recognized tha t the t rees tha t fo rm woodthrough photosynthetic processes, and the fungi and other “destruc-tive agencies” that destroy wood through respiratory processes areengaged in a never-ending cycle of biosynthetic and biodecomposi-tional forces. These relationships are shown by the following simpli-fied reaction for the predominant part of the carbon cycle of the earth:
Photosynthesis by trees andother plants
Respiration by fungi andother destructive organisms
Life as we know it would stagnate for lack of atmospheric carbondioxide in about 20 years if wood destruction were to cease whilephotosynthesis continued unabated (2). Thus, the biological decom-position of wood is both a great blessing and a serious limitation tothe usefulness of wood.
Man has probably always recognized that wood disintegrates onthe forest floor and in other moist environments. But i t was only acentury ago, in 1878, that wood decay was recognized as a biologicalprocess. The pioneering German forest pathologist Robert Hartig wasthe first to prove that fungi are the cause rather than the product ofwood decay (3). Forty years later Oshima (4) demonstrated that someinsects digest the structural polymers of wood.
This chapter presents an overview of the biological decomposi-tion of wood. It begins with a brief description of the major types ofwood destruction and their causal agents, and it continues with adescription of the progressive changes that take place in wood as itis decomposed. Special emphasis is given to the chemistry and bio-

12. K I R K A N D C O W L I N G Biological Decomposition 457
chemistry involved. The chapter ends with a brief treatment of howwood in use can be protected from decomposition and some bene-ficial uses of wood-decomposing organisms.
We have used the terms decomposition and degradation to referto the conversion of one or more of the structural polymers of woodto simpler molecules. Degradat ion can also be used to mean dete-rioration, i.e., to decrease the value of wood for some use; we haveused only deteriorat ion in this narrower sense. Decay and rot referto the fungal decomposition of wood.
Hundreds of research papers on wood biodecomposit ion havebeen published since the pioneering works of Hartig (3) and Buller(1). We aim to introduce the reader to the principles that those in-vestigations have developed. With this in mind we have categorizedand generalized, but we hope that we have not oversimplified. Thebiodecomposit ion processes are quite complex biologically, chemi-cally, and biochemically. We have pointed to some of the unknownsin a field that retains many. In Table I we have listed a number ofreviews that can be consulted for details.
Susceptibility and Resistance
When wood is laid down by the cambium of a l iving tree, twomajor types of wood cells are formed-thick-walled fiber cells thatmake wood strong and thin-walled parenchyma cells in which reservefoods are stored. Wood fiber cells die a few days or weeks after theyare formed and lose their cytoplasmic contents as soon as they be-come functional in water transport . Thus, mature wood fiber cellsconsist almost entirely of cell wall polymers-cellulose, hemicellu-loses, and lignin. For this reason, wood fiber cells can be degradedonly by organisms that have the ability to decompose these structur-ally complex high-polymeric materials.
Table I. Some Reviews on the Biological Decomposition of Wood
Subject
General, historicalInsects, marine borersChemical, physical changes
during decompositionCellulose and hemicellulose
decompositionLignin decompositionControl of biodecompositionUses and potential uses of
biodecomposing organisms
References
5 - 1 06, 11-14
5, 6, 15-M
19-2728-33
Chapter 11 this book, 34-40
26, 41, 42

458 T H E C H E M I S T R Y O F S O L I D W O O D
By contrast, wood-storage cells remain alive for many years andonly lose their cytoplasmic contents when sapwood is transformedinto heartwood. The sugars, starch, amino acids, and proteins in thewood-storage cells make sapwood highly susceptible to invasion by ala rge number o f fung i and bac te r ia tha t can use the rese rve foodmaterials but cannot attack the more complicated cell wall polymers.
The heartwood of certain species of trees is moderately to highlyresistant to decomposition even by organisms that can degrade thecell wall polymers. Highly resistant species include trees such ascypress, various cedars, and osage-orange; moderately resistant spe-cies include white oak, Douglas-fir, and certain pines. Resistance iscaused by phenols, terpenes, alkaloids, and other substances that aredeposited in heartwood and are toxic to wood-destroying fungi, bac-teria, insects, and marine borers (43). Because these toxic substancesdo not occur in sapwood, the dead sapwood of all tree species ishighly susceptible to biological decomposition. I t should be notedthat the l iving sapwood in trees is resistant to decay by virtue ofactive defense mechanisms; the heartwood is actually more suscep-tible than the living sapwood.
Although numerous fungi and some insects can cause decom-position of the dead heartwood tissues inside living trees, these or-ganisms rarely continue to cause decomposition of t imber productsafter the trees are harvested. Despite much speculation about whythis is so, no scientific explanation of this phenomenon has beenprovided to date (44).
Types of Wood Deterioration
Table II summarizes the major types of wood deterioration andthe causal organisms. The following discussion deals with deteriora-tion without cell wall decomposition; the bulk of the chapter dealswith deterioration with decomposition of cell walls.
D e t e r i o r a t i o n W i t h o u t D e c o m p o s i t i o n . W h e n f r e s h - c u tlumber or veneer is properly air-seasoned, the stored food materialsin the sapwood are soon depleted by the respiratory processes of thewood parenchyma cells themselves. If drying is delayed, however,the fresh-cut wood can be invaded by so-called sap-stain fungi anda lgae , o r by bac te r ia and molds tha t deve lop over the sur face o rpenetrate deep into the sapwood by growing through the ray cellsfrom one wood storage cell to another. These organisms use the con-tents of the wood storage cells as food and thus do not affect thestrength of wood seriously; they primarily discolor the wood or alterits permeability.
When fresh-cut wood is kiln-dried immediately, the living cellso f the sapwood a re k i l l ed by the hea t and the rese rve foods a re

12. K I R K A N D C O W L I N G Biological Decomposition 459
Table II. Types of Biological Deterioration of Wood and theOrganisms Responsible
Type of Deterioration Organism(s)
Deterioration without decompositionLoss of stored food reserves
Mechanical boring, pecking,cu t t ing
StainsSurface discolorationPit membrane destruction
Decomposition of structural polymersMechanobiochemicalBiochemical (decays)
Living wood cells insapwood
Insects, birds, mammals
F u n g iFungi, algaeBacteria, fungi
Insects, marine borersF u n g i
retained in the wood storage cells. If kiln-dried wood becomes wetagain, these stored foods can again become substrates for growth ofdiscoloring fungi and bacteria.
When fresh-cut logs are converted quickly into large piles ofchips, the l iving cells of the sapwood, together with the fungi andbacteria mentioned above, rapidly convert the stored food reservesinto carbon dioxide (CO2), water, and heat (see Reaction 1 for res-pirat ion) (45). If this metabolic heat is not dissipated, the pile be-comes hot, and under conditions of very poor ventilation can lead tospontaneous combustion (46). For all of these reasons, fresh-cut sap-wood must be considered to be alive and, therefore, must be handledas a perishable raw material.
The mos t common d i sco lor ing organ isms a re o f two genera ltypes -sur face molds wi th co lored spores , and a lgae tha t g row onwood surfaces; and fungi with dark-colored hyphae that discolor thewood interior as they penetrate deep into sapwood. Aspergillus spp.and Penicil l ium spp. are among the most common surface molds.Discolorations caused by these fungi usually are so superficial thatthey can be removed by brushing, planing, or sanding. Ceratocystisspp . a re among the mos t common deep-pene t ra t ing saps ta in fung i(47). These discolorations usually cannot be removed even by vig-orous bleaching chemicals.
Bacillus polymyxa (Prazmowski) Mace and certain other bacteria(48), as well as stain fungi and some molds such as Trichoderma viridePers. ex Fr. (49,50) , degrade the pectin membranes in the borderedpits between wood cells. This degradation greatly increases the per-meability of the wood to water and organic solvents. Increased per-

460 T H E C H E M I S T R Y O F S O L I D W O O D
meabili ty is a problem in wood finishing but can be a help in thepenetration of pulping chemicals and preservatives into sapwood.
Mechanical disintegration of wood is caused by many species ofinsects, birds, and some mammals. In some cases this disintegrationcan be quite serious.
Deterioration with Decomposition. The susceptibili ty of thewood cell wall polymers to biological decomposition is determinedlargely by their accessibili ty to enzymes and other metabolites pro-duced by wood-destroying fungi or, in the case of certain insects andmarine borers, by microorganisms that live in the digestive tracts ofthese animals. Direct physical contact between the enzymes or othermetabolites and the wood cell wall polymers is prerequisite to hy-drolytic or oxidative degradation. Because the cellulose, hemicellu-loses, and lignin are all water-insoluble polymers and are depositedin wood cell walls in intimate physical admixture with each other,this necessary physical contact can be achieved only by diffusion ofthe enzymes or other metabolites into this complex matrix or by finegrinding of the wood prior to digestion.
The crucial structural component of wood governing wood’s bi-ological decomposibility is lignin. In wood, the cellulose microfibrilsare coated or overlayered by hemicelluloses which in turn are undera lignin sheath (51). The lignin is covalently bonded to, and to someextent physically intermixed with, the hemicelluloses; the bonds be-tween lignin and hemicelluloses are probably infrequent (52). What-ever the exact relationship between the hemicelluloses and l ignin,the lignin physically prevents enzyme access to both the hemicellu-loses and cellulose. Digestibility of solid wood and other intact lig-nified tissues (lignocelluloses) is largely a function of lignin content(Figure 1).
Three biological mechanisms have evolved for overcoming thelignin barrier: (1) insects and marine borers physically disrupt thebarrier by grinding the wood very finely; (2) some microorganisms,primarily higher fungi, decompose lignin and thus expose the poly-saccharides; and (3) certain other higher fungi apparently secretenonenzymatic cellulose-depolymerizing agents that are small enoughto penetrate the-l ignin sheath. Mechanism 1 permits mechano-bio-chemical decomposi t ion of solid wood; Mechanisms 2 and 3 permitbiochemical decomposition. We will discuss each mechanism in turn.
Mechanobiochemical Decomposition
Circumventing the l ignin barrier to enzymatic digestion of thepolysaccharides occurs when wood is finely ground. Below a certainwood particle size, the polysaccharides (celluloses and hemicellu-
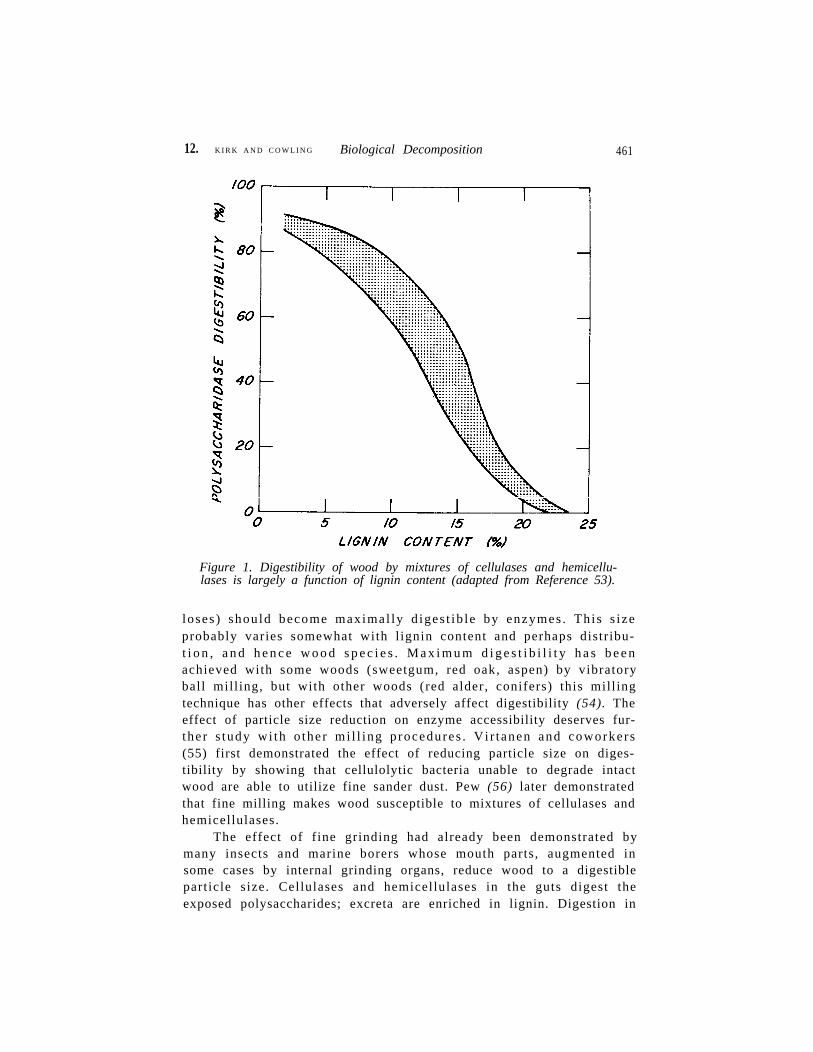
12. K I R K A N D C O W L I N G Biological Decomposition 461
Figure 1. Digestibility of wood by mixtures of cellulases and hemicellu-lases is largely a function of lignin content (adapted from Reference 53).
loses ) should become maximal ly d iges t ib le by enzymes . This s izeprobably varies somewhat with l ignin content and perhaps distribu-t i o n , a n d h e n c e w o o d s p e c i e s . M a x i m u m d i g e s t i b i l i t y h a s b e e nachieved with some woods (sweetgum, red oak, aspen) by vibratoryball mill ing, but with other woods (red alder, conifers) this mill ingtechnique has other effects that adversely affect digestibility (54). Theeffect of particle size reduction on enzyme accessibility deserves fur-the r s tudy wi th o ther mi l l ing p rocedures . Vi r tanen and coworkers(55) first demonstrated the effect of reducing particle size on diges-tibility by showing that cellulolytic bacteria unable to degrade intactwood are able to utilize fine sander dust. Pew (56) later demonstratedthat fine milling makes wood susceptible to mixtures of cellulases andhemicellulases.
The effect of fine grinding had already been demonstrated bymany insects and marine borers whose mouth parts, augmented insome cases by internal grinding organs, reduce wood to a digestibleparticle size. Cellulases and hemicellulases in the guts digest theexposed polysaccharides; excreta are enriched in lignin. Digestion in

462 T H E C H E M I S T R Y O F S O L I D W O O D
many insects is incomplete, reflecting failure to grind the wood finelyenough, absence of a full enzyme complement, insufficient residencetime, or other factors. Certain wood-boring insects, including am-brosia beetles, lyctus beetles, carpenter ants, and carpenter bees donot digest the structural polymers of wood. Wood passes through thegut of the lyctus beetles, but only the easily digested nonstructuralmate r ia l s -p r imar i ly s t a rch in pa renchyma ce l l s -a re removed ( 6 ,11, 12). The ambrosia beetles, bees, and ants do not ingest wood.
Some insects, such as the Indian longhorn beetle, Stromatiumbarbatum Fabricius (57), and the common marine borers Limnoriatripunctata Menzies (an isopod) (58) and Bankia setacea Tryon (amollusk) (59), are thought to have endogenous cellulases (and perhapso ther po lysacchar ide hydro lases ) . Termi tes and mos t o f the o therwood-digesting insects rely on polysaccharolytic microbes in theirguts. The beetle Stromatium barbatum (57) and the marine borerBankia setacea (60) are thought to utilize both their own cellulasesand those of gut microbes.
Wood-decomposing insects and the marine borers commonly di-gest only the cellulose and hemicelluloses. Reports of extensive de-composition of lignin (6, 60) deserve further study with modern tech-niques. Limi ted decompos i t ion of l ign in , however , p robably doesoccur in some insects. One of the most convincing reports is thatconversion of 1 4C lignin to 1 4CO2 in the gut of the termite Nasuti-termes exitiosus (Hill) was demonstrated (61). Because anaerobic de-composition of lignin has not been observed (62, 63) it is probablethat some oxygen is present in the guts of Nasutitermes (61) and inthose of certain other insects that reportedly digest lignin.
Table III lists representative wood-decomposing insects and ma-rine borers and summarizes some features of their action on wood.Figure 2 shows the extensive damage caused by some of these wood-decomposing animals. Only a few detailed studies of the chemistryand b iochemis t ry of wood degrada t ion by the insec t s and mar inebore r s have been made . Because the an imals de r ive nour i shmentfrom the structural polymers of wood, the subject is of practical sig-nificance and deserves more research attention.
Biochemical Decomposition: The Wood Decays
Types of Decay. As shown in Figure 3, wood decay fungi canbe divided into three classes based on the type of decay they cause:white, brown, and soft rots. The white rots can be further separatedinto pocket rots and uniform rots (Figure 3). In North America, thewhite and brown rots are caused by about 1700 species of wood-decaying fungi in the class Basidiomycetes; over 90% of these causethe white rot type of decay (67). Early reports of non-basidiomycete

12. K I R K A N D C O W L I N G Biological Decomposition 463
white rots (e.g., decay by Xylaria spp.) may be incorrect (26). White-and brown-ro t fungi a re c lose ly re la ted , which makes the evo lu-tionary basis for their very different effects on wood most intriguing.
Soft rots are caused by fungi in the classes Ascomycetes andFungi imperfecti . Most are normally found in the soil or in marineenvi ronments (68 ) . The l i s t o f sof t - ro t fungi i s long and growinglonger. A streptomycete has been shown to cause at least limited softrot in beech (69); it is likely that other soil bacteria will be shown tocause soft rot. We have observed that certain litter-decomposing ba-sidiomycetes, such as Collybia butyrucea, cause a slow decomposi-tion that resembles soft rot macroscopically.
Macroscopic and microscopic characterist ics of white, brown,and soft rots are summarized in Table IV
Progressive Changes in Chemical Composition. The chemicalcomposition of wood begins to change as soon as it is colonized bythe hyphae of wood-destroying fungi. Initial invasion of the ray cellsand vessels (70) is primarily at the expense of soluble sugars, starch,and o ther ca rbohydra tes . The hyphae then pene t ra te and becomeestablished in virtually every cell of the wood in which environmentaland other factors are favorable. This at tack results in a progressivedepletion of all three structural polymers during white and soft rots,and in the polysaccharides during brown rot (Figure 4). Depletion oflignin roughly parallels loss of polysaccharides in white rot, lags be-hind polysaccharide depletion in soft rot, and is insignificant in brownrot. A slight initial increase in lignin is frequently seen during brownrot (see Figure 4). This increase is probably due to partial oxygenationof the lignin polymer (31, 72).
Some variation in the relative rates of cellulose vs. hemicellulosedepletion is observed among various fungi and woods. Marked se-lectivity for the hemicelluloses has been achieved through geneticmanipulation of several white-rot fungi (73) and occurs naturally insome species, e.g., Polyporus pargamenus Fr. (74). Selective removalof cellulose is not observed, a consequence of its location within thel ign in-hemice l lu lose shea th . Se lec t ive removal o f l ign in does no toccur, apparently because polysaccharides provide energy necessaryfor lignin decomposition (28, 75–77).
Progressive Changes in Strength Properties. Decay of woodhas profound effects on strength properties. Of the various measuresof wood strength (78), toughness and the related property of work tomaximum load are most sensitive to decay. Toughness, which is alsocalled impact bending strength, is relatively easy to measure and hasbeen the most widely studied.
All three types of decay cause losses in toughness and relatedproperties that far exceed their losses in weight. Wood decayed to

Table III. Decomposition of Wood by Some Insects and Marine Borers
Classification(example) Description of Damage Notes and References
Coleoptera, Anobiidae[Xestobium rufovillosum(De G.), “deathwatchbeetle”]
Coleoptera, Cerambycidae[Hyalotrupes bajulus(L.) “old houseborer”]
[Stromatium burbutum(Fabricius) “Indianlonghorn beetle”]
Insects
Round, powder-filled galleries (<3 mmin diameter) ; wood becomes r iddled(Figure 2)
“ P o w d e r p o s t ” b e e t l e s ; d a m a g edone primarily by larvae; limiteddigestion of cellulose and hemicel-luloses (6)
Oval-shaped galleries winding irreg-u l a r ly t h rough the wood , ga l l e r i e spowder-filled (6-10 mm in diameter)(Figure 2)
As above
Longhorn beetles; larvae (“round-headed borers”) do damage; limiteddigestion of cellulose and hemicel-luloses (11)
Larvae do the damage; extensivedigestion of cellulose and hemicel-luloses; lignin decomposition re-ported (57)

Isoptera, Kalotermitidae(Kulotermes flavicollisFabr.)
Isoptera, Rhinotermitidae(Reticulitermes spp.)
Hymenoptera, Siricidae(Sirex phantoma)
Adapedonta, Teredinidae(bivalve mollusks)(Bankia, Teredo spp.)
Isopoda, Limnoridae(Limnoria tripunctataMenzies)
Galleries, chambers within wood; honey-combing; wood structure thoroughly de-stroyed (Figure 2)
First attack is on less dense part of wood,so that earlywood is removed preferen-tially; wood structure is thoroughly de-stroyed (Figure 2)
Small (4-6 mm in diameter) round gal-leries tightly packed with frass
Marine Borers
Small entry holes on wood surface; in-terior riddled with much larger boreholes (Figure 2)
Maze of tunnels from surface inward, re-sulting in progressive erosion (Figure 2)
“Drywood” termites; polysaccha-r ides are extensively digested;lignin may be partly decomposed(6, 64)“Subterranean” termites; polysac-charides are extensively digested;lignin at least slightly altered (65)
“Wood wasps”; some digestion ofcellulose (6)
Digest cellulose extensively, andprobably hemicelluloses, but notlignin; Bankia secretes own cellu-lase, and also has cellulolytic gutmicrobes (59)
Preferentially digest cellulose (andprobably hemicelluloses); lignin un-altered (66); thought to secrete owncellulases; gut is sterile (58)

466 T H E C H E M I S T R Y O F S O L I D W O O D

12. K I R K A N D C O W L I N G Biological Decomposition 467

Table IV. Macroscopic and Microscopic Features of White, Soft, and Brown Rots of Wood
Type of Decay (representativecausal organisms) Macroscopic Featuresa Microscopic Feuturesb
White rot[Coriolus versicolor (L. exFr.) Quel., Phanerochaetechrysosporium Burds.]
Soft rot(Chaetomium globosum Kunze,Paecilomyces spp., Allescheria
Usually not discernible in earlystages, w o o d l a t e r b e c o m e sbleached or discolored; mottlingor flecking is common; some spe-cies form white pockets in wood;dark zone lines often observed innature ; wood re ta ins s ize andshape
O n l y w a t e r - s o a k e d w o o d a t -tacked; decay even in early stagescharacterized by softening and
Hyphae colonize lumens of cells,penetrate first from cell to cell viapits, later directly via bore holes;progressive decay observed asthinning of cell walls; (in “pocketrots,” decay is limited to the de-veloping pockets)
Hyphae grow longitudinally with-in the secondary walls; character-istic catenate, spindle-, or dia-

terrestris Apinis)
Brown rot[Gloeophyllum trabeum (Pers. exFr.) Murr., Poria plaenta (Fr.)Cke., Merulius lacrymans(Wulf.) Fr.]
a See Figure 3 and References 7 and 8.b See Reference 70.
minute checking of surfaces; dullgray to brown color. Decay pro-
mond-shaped cavities, orientedwith cellulose microfibrils
gresses gradually from the surfaceinward
Often not discernible during earlystages, but wood rapidly becomesbrash; later, wood becomes dis-colored, finally brown and soft;drying causes extensive checkingacross the grain, giving a cubicalpattern; mycelial fans may bepresent on surfaces or within thechecks
Hyphae colonize lumens of cells,penetrate from cell to cell via pitsand often directly via bore holes.As decay progresses, cell wallsmay shrink and collapse

Figure 4. Cellulose (glucan) and hemicelluloses (xylan, pentosan) are depleted during decay by white-,brown-, and soft-rot fungi; lignin is decompose efficiently only by white-rot fungi. Key: white rot,sweetgum by Coriolus versicolor (L. ex Fr.) Quel. (15); brown rot, sweetgum by Poria placenta (Fr.) Cke.
(15); and soft rot, beech by Chaetomium globosum Kunze (71).

12. K I R K A N D C O W L I N G Biological Decomposition 471
weight losses of less than 3% by white-, soft- , or brown-rot fungifrequently has lost over 50% of i ts strength measured as toughness(18, 79-85). Microscopic observations reveal that this loss is not dueto a localized destruction of part of the test piece, but rather to ageneral decomposition.
It is not known why toughness is so sensitive to decay by allthree groups of fungi. Campbell (5) concluded that the reduction isdue to a “shortening of the cellulose chain molecules in wood.” Thisis reasonable for brown rot, but unlikely for white rot or soft rot .Cowling (15) demonstrated that the brown-rot fungus Poria monticolaMurr. [now Poria placenta (Fr.) Cke.] causes a sharp reduction int h e d e g r e e o f p o l y m e r i z a t i o n ( D P ) o f c e l l u l o s e d u r i n g d e c a y o fsweetgum wood (Figure 5). In contrast , the white-rot fungus Poly-porus versicolor L. ex Fr. [now Corioius versicolor (L. ex Fr.) Quel.]only gradually reduces cellulose DP (Figure 5). Levi and Preston (71)showed that the residual cellulose in beech wood is also only grad-ua l ly decreased in DP dur ing decay by the sof t - ro t fungus Chae-tomium globosum Kunze. Thus the basis for toughness loss in whiteand soft rots lies elsewhere and deserves further research.
The effect of brown-rot fungi on other wood strength propertiesis much more pronounced than that of white-rot fungi (see Reference18), and reflects cellulose depolymerization. Interestingly, wood spe-c ies vary subs tan t ia l ly in s t reng th loss a t a g iven weigh t loss bybrown-rot fungi (18). The basis for this variation is not known anddeserves addit ional research.
Mechanisms of Wood Decay. Microscopy indicates that theenzymes or other agents that degrade the wood cell wall polymersdi f fuse away f rom the hyphae , and tha t they are secre ted by thelaterals as well as by the growing tips (70, 86, 87). Gelatinous sheathsencase both hyphae and substrate when brown- or white-rot fungiattack isolated cellulose (88). The nature and actual importance ofsuch sheaths in the decay of solid wood need further investigation.The shea ths might p rovide an op t imum envi ronment for enzymeaction and might also help prevent loss of enzymes with their pre-cious nitrogen. Because the nitrogen content of wood is quite low,this important element is l imiting in the nutrit ion of wood-decom-posing organisms. I ts importance in wood decay has been discussed
(89).Studies of the biochemical mechanism of wood decay have, with
very few exceptions, been conducted with isolated cellulose, hemi-celluloses or l ignin, or with appropriate model compounds, ratherthan with solid wood. The few studies with enzymes produced duringgrowth on or in wood have revealed no peculiarities that would bringinto question the results of studies with the isolated components.

472 THE CHEMISTRY OF SOLID WOOD
Figure 5. Cellulose in wood is depolymerized early during brown rot, butonly gradually during white rot. The data are for sweetgum sapwooddecayed by the brown-rot fungus Poria placenta (Fr.) Cke. (formerly P.monticola Murr.) and for the white-rot fungus Coriolus versicolor (L. ex
Fr.) Quel. (formerly Polyporus versicolor L. ex Fr.) (15).
Isolated cellulose was not degraded by the brown-rot fungus Poriaplacenta unless the fungus was in contact with whole wood (90).Further study indicated, however, that the wood could be replacedwi th s imple sugars , i . e . , tha t nu t r i en t s in the wood were s implyserving as starter carbon/energy sources (90).

12. KIRK AND COWLING Biological Decomposition 473
In the following discussion, we have assumed that wood decayis due to the concerted action of the individual enzyme systems re-sponsible for cellulose, hemicellulose, and lignin decomposition.
CELLULOSE DECOMPOSITION. Decomposition of crystall ine cel-lulose by white-rot fungi and various soil fungi (including the soft-rotfungi), results from the synergistic action of three types of hydrolyticenzymes: e n d o-1,4-β-glucanases, exo-1,4-β-glucanases, and β-gluco-s idases . P resumably , bac te r i a l and an imal ce l lu lases a re s imi la r .Brown-rot fungi also decompose crystalline cellulose, but most pos-s e s s n o e x o-glucanase; their unique mechanism of cellulose degra-dation is discussed later.
The mechanism of cellulose degradation by fungi has been thesub jec t o f ex tens ive resea rch ( 1 9 – 2 1 , 2 3 – 2 5 ) , pr imar i ly wi th themold Tr i choderma v i r i de and the whi te - ro t fungus S p o r o t r i c h u mpulverulentum Novo. (now Phanerochaete chrysosporium Burds.) .Results indicate that the endo-1,4-β-glucanases (Cx enzymes) act ran-domly over the exposed surfaces of cellulose microfibrils. Nonre-ducing termini generated by this action are then hydrolyzed by exo-1,4-β-glucanases (cellobiohydrolases, C1 enzymes), releasing cello-biose. Cellobiose may be cleaved by a β-glucosidase to yield glucose,or in white rot and perhaps other fungi it may be oxidized to cello-bionic acid and then cleaved. The endo- and exo-glucanases act syn-ergist ically, perhaps as a loose complex (91). Generally, the endo-glucanases and probably the exo-glucanases are repressed by highconcentrations of monosaccharides (23). Both types of glucanaseshave molecular weights ranging up to about 75,000 (92, 93), whereast h e β-glucosidases are considerably larger (94).
In addition, oxidizing enzymes probably are involved in cellulosedecompos i t ion by cer ta in whi te - ro t fungi . Phanerochae t e chry so -sporium possesses cellobiose oxidase, which converts cellobiose tocellobiono-δ-lactone, with O2 serving as electron acceptor (23). Thisenzyme i s respons ib le fo r the more rap id hydro lys i s o f ce l lu loseunder aerobic conditions than under anaerobic conditions, presum-ably because i t removes cellobiose and prevents the transglycosy-lation reactions and the inhibition of endo-glucanase activity that oc-curs when cellobiose accumulates (23, 73). Similar oxidizing activityis not found in the species of Fungi Imperfecti examined (91). An-other enzyme, cellobiose:quinone oxidoreductase, has the same ac-tion, but requires quinones as electron acceptors (23). A glucose ox-idase also has been implicated in the overall process (23); it oxidizesglucose to gluconolactone with O2 or quinones as electron acceptors.Presumably, these oxidizing activities regulate the amounts of glucoseand cellobiose, ultimately coordinating the rates of cellulose hydro-lysis and the metabolism of end products. Phenols, which are inter-

474 T H E C H E M I S T R Y O F S O L I D W O O D
mediates in lignin decomposition, also affect the amounts of endo-glucanase activity in white-rot fungi (23). Figure 6 summarizes theinterconversions and regulatory interactions in cellulose hydrolysisby white-rot fungi (23).
Cellulose decomposition by brown-rot fungi is an unusual pro-cess. In the early stages of their decay of wood, and in contrast todecay by white-rot fungi, the cellulose is severely depolymerized (seeFigure 5). This discovery (15) explained the highly destructive effectsof these fungi on wood, as described in the section entit led “Pro-gressive Changes in Strength Properties” (page 463), but it presentedan enigma because only a very small percentage of the cellulose inwood is accessible to cellulolytic enzymes. Enzymatic degradationmust cause a gradual loss in cellulose integrity, as seen in whiterot (15).
Because the depolymerizing agent of the brown-rot fungi com-pletely penetrates the crystalline microfibrils, only very small mol-ecules can be responsible (15). In discussing this, Cowling and Brown(92) noted that G. Halliwell (95) had described experiments on thedepolymerization of cellulose under physiological conditions withFenton’s reagent (H2 O 2 and ferrous salts) . Subsequent studies dem-onstrated that brown-rot fungi secrete H2 O 2 and that wood containsenough i ron fo r a poss ib le invo lvement o f an Fe–H2 O 2 sys tem incellulose depolymerization (96). Cellulose is in fact oxidized duringattack by the brown-rot fungus Poria placenta (90). Oxalic acid, whichis secreted by brown-rot fungi, can reduce the Fe3 + normally presentin wood to Fe2 +, the active form in Fenton’s reagent (97, 98). Figure7 shows the proposed mechanism for the depolymerization of cellu-lose. Nicholas et al . (personal communication) demonstrated degra-dation of 1 4C-cellulose by Gloeophyllum trabeum (Pers. ex Fr.) Murr.through a membrane with a nominal molecular weight limit of 1000.
This in i t i a l ox ida t ive depolymer iza t ion of ce l lu lose ev iden t lyopens up the wood cell wall structure so that cellulolytic and hemi-cellulolytic enzymes can reach their substrates despite the presenceof l ign in . So lubi l i ty o f wood in 1% NaOH increases markedly onbrown-rot attack (15) and reflects cellulose depolymerization and theopening up of the wood structure.
Interestingly, enzyme preparations from most brown-rot fungip o s s e s s e n d o - b u t n o t e x o-1,4-β-glucanase activity (99–101). Thesefungi differ from other cellulolytic organisms, too, in that endo-glu-canase production is not repressed by monosaccharides (102). Stil lanother unusual feature is a large enzyme complex that hydrolyzescarboxymethyl cellulose, xylans, glucomannans, and various glyco-sides (103).

Figure 6. Both hydrolytic and oxidative enzymes participate in cellulose decomposition by the white-rotfungus Phanerochaete chrysosporium Burds. Hydrolytic enzymes: 1, endo-1,4-β-glucanases; 2, exo-1,4-β-glucanase; and 3, β-glucosidase. Oxidative enzymes: 4, glucose oxidase, 5, cellobiose oxidase; and 6,
cellobiose:quinone oxidoreductase. Adapted from Reference 23.

476 T H E C H E M I S T R Y O F S O L I D W O O D
Figure 7. Depolymerization of cellulose in wood by brown-rot fungi mightinvolve Fe2+ and H2 O2 (92, 96, 97).
Although white-rot fungi also secrete H2O2 (104), they have notbeen found to depolymerize cellulose oxidatively. One reason mightbe that they possess oxalate decarboxylase, which decomposes oxa-late, whereas brown-rot fungi apparently do not (98). This problemdeserves further investigation.
The cellulolytic system of soft-rot fungi has received relativelylit t le attention. It is probably similar to that of the closely relatedAscomycetes and Fungi Imperfec t i ( 1 9 - 2 5 , 2 7 ) , which apparen t lydiffers little from that of white-rot fungi, except perhaps in the ab-sence of oxidizing enzymes.
HEMICELLULOSE DECOMPOSITION. Dekker and Richards have re-viewed the microbial hemicellulases (22). Wood-rotting fungi pro-duce enzymes capable of hydrolyzing a variety of β-(1 g 4)-linkedglycan (mannan and xylan) substrates, as well as various glycosides(94, 101, 105-8) Judging from the relatively l i t t le research done,corresponding enzymes from the three groups of decay organismsappear to be similar, and are also similar to those from other microbes(22, 106). To our knowledge, the enzymes responsible for completedepolymerization of a wood hemicellulose have not been describedin any microbe. This is another subject that needs additional study.endo-Glycanases from white-, brown-, and soft-rot fungi apparentlyall act randomly and produce dimeric and higher oligomeric products(22, 108-11). Uronic acid-substituted oligosaccharides are producedfrom glucuronoxylan substrates (107-9, 111) but enzymes catalyzing

12. K I R K A N D C O W L I N G Biological Decomposition 477
hydrolysis of the xylose–uranic acid linkage have not been reported.A mannanase purified from a brown-rot fungus hydrolyzed both man-nose-(1 g 4)-glucose and glucose-(1 g 4)-mannose linkages (110).Glycosidases active on hemicellulose-derived disaccharides also arep r o d u c e d b y t h e w o o d - r o t t i n g f u n g i ( 9 4 ) , b u t e x o-hydrolases de-grading hemicellulose oligosaccharides have not been reported inthese organisms.
The enzyme complex with multiple glycan and glycoside hydro-lase activity in the brown-rot fungus Poria placenta, mentioned above(103), has a molecular weight of 1.85 × 10 5 ; other hemicellulases(endo-glycanases) of wood-rotting fungi have molecular weights of 3× 104–6 × 104 (94).
Information regarding regulation of the synthesis of the hemi-cellulases is somewhat contradictory (22). Regulation in wood-decom-posing fungi, however, has not been the subject of the detailed studythat i t deserves. Multiple hemicellulase activity is found in culturefiltrates of both white- and brown-rot fungi after growth on a varietyof substrates (107). Hemicellulase production by the two types offungi is different, however, in that brown-rot fungi exhibit good hemi-cellulase (as well as cellulase) activit ies during growth on simplesugars, whereas white-rot fungi do not (107, 112). Production of theenzymes on simple sugars might be induced in response to hyphalwall constituents following substrate depletion, as is an endo-xylanasein the white-rot fungus Stereum sanguinolentum Fr. (94). Hemicel-lulases in the soft-rot fungus Chaetomium globosum are induced spe-cifically by their substrates (108).
LIGNIN DECOMPOSITION. Research on the fungal decompositionof lignin has accelerated greatly in recent years. Some of the reactionscomprising degradation have been elucidated, and the unusual bio-chemical and physiological features are being described. Several re-views (28-33) provide specific literature references.
Research with the white-rot fungi has shown that the process isoxidative, that the ligninolytic system is nonspecific, that i ts rate-limiting component is not induced by lignin, and that depolymeriza-tion may not be an obligatory first step. Lignin degradation, there-fore, is distinct from cellulose and hemicellulose degradation; indeed,it differs from the biodegradation of all other biopolymers studied.Prominent reactions of lignin polymer degradation are oxidations andoxidative cleavages in the propyl side chains, demethylations of meth-oxyl groups, and even cleavages in aromatic rings. The chemistry ofdegradation is being investigated both by characterization of the par-t ia l ly degraded polymer and of low molecular weight degrada t ionproducts, and through studies of the metabolism of low molecularweight dirneric model compounds representing substructures in the

478 T H E C H E M I S T R Y O F S O L I D W O O D
polymer. With the model compounds, the specific reactions that com-prise degradation are rapidly being described; for example, Scheme1 shows the fate of the nonphenolic diarylpropane-type of substruc-ture model in cultures of the white-rot fungus Phanerochaete chryso-s p o r i u m ( 1 1 3 - 1 5 ) . Models r ep resen t ing o the r impor tan t subs t ruc -tures have been studied similarly.
Many past studies on the enzymes of lignin decomposition fo-cused on phenol-oxidizing enzymes, such as lactase and peroxidase,produced by white-rot fungi. It is unlikely, however, that this typeof activity is important in structural decomposition (33), although theenzymes may have some other role in lignin decomposition (28).
Because the lignin polymer is attacked by extracellular nonspe-cific oxidizing agents, it is possible that enzymes may not be involveddirectly, just as cellulose apparently is nonenzymatically oxidized bybrown-rot fungi. Hall (116) suggested that diffusible species such assuper-oxide (O2
.- ), derived from molecular oxygen, may participate inlignin decomposition. Research has since indicated that activated ox-ygen species apparently are commonly produced by the lignin-de-grading fungi. Singlet oxygen, an excited state of O 2 reportedly in-
Scheme 1. Model compounds are being used to elucidate the specific reactionsof lignin decomposition by white-rot fungi. The fate of a nonphenolic diaryl-propane-type of model in ligninolytic cultures of Phanerochaete chrysosporium
Burds is shown (115–15).

12. K I K K A N D C O W L I N G Biological Decomposition 479
volved in other biological reactions, was implicated in l ignin bio-degradation (117), but was later shown not to be involved (118). Hy-droxyl rad ica l ( •OH) has now been impl ica ted ( 1 1 9 - 2 1 ) , but no tproven to be involved. Whether it is or not, good evidence has beengained for participation by its immediate precursor, H2O2 (119, 121,144) . Hydroxyl radical is formed by metal reduction of H2 O 2 (seeFigure 7).
Findings in this very active research area indicate that enzymesare directly involved. An extracellular enzyme in ligninolytic cul-tures of P. chrysosporium has been discovered (122). It requires H2O2
for activity and catalyzes the cleavage of nonphenolic model com-pounds of both the diarylpropane and the arylglycerol-β-aryl ethertypes. In the former type of model, the degradative reaction is theinitial cleavage shown in Scheme 1. The new enzyme catalyzes thepartial depolymerization of l ignin. It is not the same as earl ier re-ported “lignin-degrading enzymes” (123-26) .
Fur ther metabol i sm of low molecu la r weigh t p roduc ts o f theinitial degradation of lignin, however it occurs, is probably via clas-sical modes. Vanillic acid, a prominent product of the fungal degra-dation of lignin (127), is degraded by substrate-inducible enzymes inP. chrysosporium (128, 129).
Although brown-ro t fung i a re poor degraders o f l ign in ( seeFigure 4) they do apparently possess the basic machinery. The maineffect they have on lignin is demethylation of arylmethoxyl groups(130), although oxidative changes occur, including some cleavage ofaromatic rings (72). Indeed, limited oxidation of aromatic and propylside chain carbon as well as methoxyl carbon to CO2 has been dem-onstrated (131, 132). Extensive depolymerization apparently does notoccur (133, 134), and it seems unlikely that the limited degradationof lignin by brown-rot fungi is sufficient to open up the wood struc-ture to polysaccharidases. The nonenzymatic oxidative depolymeri-zation of cellulose, discussed in the section entit led “Cellulose De-composition” (page 473), is evidently what opens up the structure.
Analyses of soft-rotted wood have revealed limited depletion oflignin (71, 135). That these fungi can oxidize lignin to CO2 was shownby using 1 4 C-lignins (132). Rates did not approach those seen withwhi te - ro t fungi , bu t op t imiza t ion s tud ies have no t ye t been con-duc ted . Clear ly , the wood-decompos ing machinery of the sof t - ro tfungi has received too little attention.
Control and Uses of Wood-Decomposing Organisms
Control. When used properly, wood will retain its strength andother desirable properties for many centuries. When wood is used

480 THE CHEMISTRY OF SOLID WOOD
improperly, however, i t can be decomposed by various organisms,as discussed in this chapter.
The cardinal rules for proper use of wood can be stated simply:keep wood dry, and, if you can’t keep it dry, use naturally durableor preservative-treated wood. The first rule is based on a simplebiological principle: liquid water is needed in wood cells to providea medium for diffusion of the enzymes or other metabolites by whichwood-decomposing organisms digest the wood substance. If there isno liquid water present inside the wood cells, there will be no me-dium for diffusion, and therefore no biological decomposition exceptfor certain insects of relatively minor importance. Thus, as long aswood is kept below its f iber-saturation point (about 27% of i ts dryweight), it will never decay.
A few brown-rot fungi, notably Serpula incrassata (Berk. andCurt .) Dank. and Merulius lucrymans (Wulf.) Fr. have the unusualabili ty to conduct l iquid water from moist soil or other sources ofmoisture into dry wood (36). Similarly, subterranean termites canattack very dry wood and, if they have access to water, can transporti t through the tubes they construct between moist soil and wood.But chemical soil treatments around wood buildings can prevent at-tack by even these organisms. Construction practices that thwart bothfungal decay and insect attack have been described (36, 38).
The second cardinal rule is based on an equally simple biologicalprinciple: some chemicals inhibit living organisms. When wood mustbe used in moist environments, use of naturally durable or preser-vative-treated timber will provide long-lasting protection against bi-ological decomposition. Although the heartwood of many tree speciesis naturally durable (36, 43, 136, 137), sapwood of all tree species ishighly susceptible to decomposition. Most construction t imbers oftemperate regions are sapwood and require preservative treatmentfor use in moist environments.
The chemicals that have found wide use and acceptance as woodprese rva t ives , p r imar i ly c reoso te , pen tach lo ropheno l , ch romatedcopper arsenate, and ammoniacal copper arsenate, are broad-spec-trum pesticides. To achieve greater specificity, advantage could betaken of the unique physiological features of the causal organisms.One such feature is digestion of wood. Treatments that make wooda nonsubstrate (Chapter 4), or chemicals that interfere with the syn-thesis, secretion, or activity of the wood-decomposing enzymes canbe envisioned. A chemical that prevents the synthesis of chitin, whichis an essential component of both insects and fungi, is a goal of currentresearch (138). Many other possibilities for more specific interferencewith the growth or activity of the wood-decomposing organisms couldresult from a better understanding of the physiology and biochem-

12. K I R K A N D C O W L I N G Biological Decomposition 481
istry of the decomposition processes and of the organisms respon-sible.
Uses and Potential Uses (26, 139). Although wood partly de-graded by fungi or attacked by insects sometimes has increased aes-thetic appeal for use in decorative paneling, wooden bowls, or otherhousehold items, a decrease in attractiveness and usefulness is moreoften the case. Thus biological decomposition usually leads to a de-crease rather than an increase in the value of wood.
However, deliberate conversion of solid wood by the agents ofbiodecomposition is done commercially and has considerable addi-t iona l po ten t ia l . The wor ld ’s second mos t impor tan t commerc ia lmushroom, comprising 20% of world sales (140-42), is shiitake (Len-tinus edodes) (Figure 8). Cultivated in Asia, primarily on oak logs,shiitake is a major food in Japan and that nation’s largest agriculturalexport. Several other commercial mushrooms are also grown on solidwood (Table V). These mushrooms have much potential in the Westwhere they are currently of minor importance.
Most, and perhaps all , of these edible mushroom-forming fungicause the white-rot type of wood decomposition. Lent inus edodesand some of the others increase the ruminant digestibili ty of wood-somet imes to over 60% ( 2 6 )–because they remove the lignin andhemicelluloses before the cellulose. Thus, they have potential fordirect conversion of wastewood into feed for ruminants. Although theresidue from shiitake production is sometimes fed to cattle in Japan,a process aimed at converting wood to cattle feed by this fungus orother fungi apparently has not been developed.
The possibil i ty of using brown-rot fungi to open up the woodstructure for ruminants or for conversion via enzymatic hydrolysis ordirect fermentation has received virtually no research attention.
Another potential use of white-rot fungi that has received someresearch attention is in biomechanical pulping. Treatment of wood toa weight loss of less than 3% can lower the energy requirements forsubsequent mechanical pulping by more than 20% (139). Mechanicalpulps make up a growing share-now about 10%—of U.S. pulp pro-duction. Energy consumption is high and makes any treatment at-tractive that decreases energy demand. We are struck by the simi-larity in the rapid loss in toughness (see the “Progressive Changes inStrength Properties” section) and the rapid decrease in energy re-quirements for mechanical pulping during white rot . The physical-chemical basis for this decrease should be investigated; it might ac-tua l ly have l i t t l e to do wi th l ign in degrada t ion , which has beenassumed to be the bas i s (139) . Biomechan ica l pu lp ing dese rvesadditional investigation.
Produc t ion o f microb ia l chemica l p roduc ts dur ing growth on

482 T H E C H E M I S T R Y O F S O L I D W O O D

12. KIRK AND COWLING Biological Decomposition 483
Table V. Some Commercial Mushrooms Cultivatedon Solid Wood (140, 143)
Species Substrates
Auricularia spp.Flamulina velutipes (Fr.) Sing.Pholiota nameko (T. Ito)
S. Ito et ImaiPleurotus spp.Tremella fuciformis Berk.Lentinus edodes (Berk.) Sing.
Hardwood logsSawdust
Hardwood logsHardwood logs, sawdustHardwood logsHardwood logs, sawdust
NOTE: Adapted from References 140 and 143.
solid wood, although possible, seems unlikely to become important,because of the availability of more practical substrates.
Altering the properties of wood components for particular usesis another possible use of wood-decomposing microbes. As an ex-ample, in their attack on solid wood, brown-rot fungi leave a ligninresidue that is enriched in phenolic hydroxyl groups (72, 130); suchlignin might serve well in phenolic adhesives.
One additional potential application of wood-decomposing fungii s in was te t rea tment . Al though no t d i rec ted a t so l id wood , suchapplications may be possible simply because the fungi have evolvedthe capacity to degrade such a complex solid substrate. The potentialuse of the lignin-decomposing system of white-rot fungi to decolorizelignin-derived wastes from pulp bleaching has been investigated withpromising results (139). The lack of specificity of the oxidative lig-ninolytic system suggests a broader applicabili ty than just to wood-derived wastes.
Acknowledgments
We thank T . L . Amburgey , Fores t Produc ts Ut i l i za t ion Labo-ratory, Mississippi State Universi ty, Mississippi State, Mississippi ,and G. R. Esenther, W. E. Eslyn, T. L. Highley, B. R. Johnson, andG. F. Leatham, all of the U.S. Forest Products Laboratory, Madison,Wisconsin, for critically reviewing portions of the manuscript. Sam-ples used in Figures 2 and 3 were provided by Esenther and Eslyn.Leatham provided Figure 8. The expert clerical assistance of JaneD. Kohlman is gratefully acknowledged.
Literature Cited

484 THE CHEMISTRY OF SOLID WOOD

12. KIRK AND COWLING Biological Decomposition 485

486 THE CHEMISTRY OF SOLID WOOD

12. K I R K A N D C O W L I N G Biological Decomposition 487





























