Embed Size (px)
Citation preview

BIOLOGÍA REPRODUCTIVA DEL PEPINO DE MAR Holothuria (Selenkothuria) glaberrima Selenka, 1867 EN
SANTA MARTA, COLOMBIA.
Erika Paola Ortiz Gómez
1190351
UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE CIENCIAS, DEPARTAMENTO DE BIOLOGÍA – SEDE BOGOTÁ CENTRO DE ESTUDIOS EN CIENCIAS DEL MAR (CECIMAR) – SEDE CARIBE
CONVENIO UNIVERSIDAD NACIONAL DE COLOMBIA - INVEMAR
Santa Marta, D.T.C.H., 2011


BIOLOGÍA REPRODUCTIVA DEL PEPINO DE MAR Holothuria (Selenkothuria) glaberrima Selenka, 1867 EN SANTA MARTA,
COLOMBIA.
Erika Paola Ortiz Gómez
1190351
Tesis presentada como requisito parcial para optar al título de: Magister en Biología – Línea Biología Marina
Directora: Milena Benavides Serrato, M.Sc., INVEMAR
Codirector:
Sven Zea, Ph.D., CECIMAR, Sede Caribe
Línea de Investigación: Biología Marina
Grupo de Investigación: Fauna Marina Colombiana: diversidad y usos
UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE CIENCIAS, DEPARTAMENTO DE BIOLOGÍA – SEDE BOGOTÁ CENTRO DE ESTUDIOS EN CIENCIAS DEL MAR (CECIMAR) – SEDE CARIBE
CONVENIO UNIVERSIDAD NACIONAL DE COLOMBIA – INVEMAR
Santa Marta, D.T.C.H., 2011


A mis padres, Gracias por creer en mí,
Por apoyar cada proyecto de vida que se me ocurre Y brindarme sus consejos y sabiduría.
A Jair, Por acompañarme en esta etapa del camino,
Y hacerla tan feliz, divertida y especial.
A mis abuelos, Enriquetica, Santicos y Mariita,
Por ser el pilar más fuerte De esta gran familia.


AGRADECIMIENTOS A mis padres, Jair y mi familia por brindarme su apoyo incondicional y creer siempre en mí y en mis sueños. Angelina y Euginius, gracias por su amistad, por reír, llorar, convivir, pepiniar y compartir conmigo momentos inolvidables, que hicieron de esta etapa toda una aventura. A Javier, Milena y Sven por creer en este proyecto y apoyarlo, brindarme su tiempo, enseñanzas, consejos y correcciones, sin los que este trabajo no hubiera sido realidad. A la cohorte XVI de la Maestría en Ciencias de la Universidad Nacional, Angélica, Eugenia, Elizabeth y Patricia, por ser tan únicas y especiales. También a Mery y Luisino, que con su sonrisa, amabilidad y diligencia fueron de gran ayuda en este proceso. A Andiecita, que siempre está ahí para recordarme que hay que soñar. Al Instituto de Investigaciones Marinas y Costeras – INVEMAR, especialmente al Programa de Investigación, Valoración y aprovechamiento de recursos marinos vivos (VAR) por su apoyo económico; a todo su personal, principalmente Pocho, Carlitos y el equipo del laboratorio de Bioprospección Marina. A la Dr. Chantal Conand, por sus grandes aportes y consejos que desentramaron en más de una ocasión este proceso. A todos aquellas personas que de una u otra forma directa o indirectamente estuvieron ligadas a este trabajo.


CONTENIDO
RESUMEN ...................................................................................................................... XI
ABSTRACT ................................................................................................................... XII
INTRODUCCION ............................................................................................................. 1
METODOLOGÍA .............................................................................................................. 7
Análisis de datos ..................................................................................................... 10
RESULTADOS............................................................................................................... 11
Características morfológicas de la población local .................................................. 11
Proporción de sexos ................................................................................................ 11
Talla y peso de la pared corporal ............................................................................ 12
Peso mínimo de la primera madurez ....................................................................... 15
Morfología gonadal .................................................................................................. 15
Ciclo reproductivo a nivel de túbulos ....................................................................... 18
Ciclo reproductivo a nivel de individuos: estadios de madurez ................................ 18
Índice gonadosomático............................................................................................ 27
Tipo de ciclo y desove ............................................................................................. 29
DISCUSIÓN ................................................................................................................... 31
CONCLUSIONES Y RECOMENDACIONES ................................................................. 34
BIBLIOGRAFÍA ............................................................................................................. 36
GLOSARIO .................................................................................................................... 44
ANEXOS ........................................................................................................................ 45


Resumen y Abstract
XI
RESUMEN
Los pepinos de mar constituyen un recurso pesquero en muchos países asiáticos; su
explotación indiscriminada ha generado extinciones locales y veda, trasladando la
presión a especies de otras áreas, generando la necesidad de conocer la biología
poblacional de estas nuevas especies susceptibles a la extracción para crear
herramientas de manejo adecuadas. Este es el primer trabajo enfocado a la biología
reproductiva del pepino tropical Holothuria (Selenkothuria) glaberrima. Se recolectaron al
azar 20 individuos mensuales en 4 litorales rocosos de enero a octubre de 2010, en
Santa Marta – Caribe colombiano, área con una época climática seca de afloramiento
costero de aguas frías y otra de lluvias con mayor temperatura. Este holotúrido dioico
presenta una proporción sexual de 1:1. La gónada está constituida por numerosos
túbulos, que pueden estar en diferentes estadios de desarrollo, aunque con predominio
de uno en particular. Observaciones macroscópicas e histología caracterizaron cinco
estadios gonadales: (I) inmaduro, (II) en desarrollo, (III) maduro, (IV) parcialmente
desovado y (V) recuperación. El porcentaje mensual de individuos en cada estadio y el
índice gonadosomático revelaron que la población de H. (S.) glaberrima tiene madurez
sexual continua a lo largo del año (en diferentes individuos) y el desarrollo gonadal no es
totalmente sincrónico entre sexos. Sin embargo, una proporción de la población describe
un ciclo en el cual la madurez parece asociarse al incremento en la temperatura
superficial del agua de mar al final de la época seca y los desoves muestran mayor
asociación con la época mayor de lluvias entre julio y septiembre.
Palabras clave: Reproducción continua o aperiódica, Holothuria (Selenkothuria)
glaberrima, Holoturoidea, Caribe colombiano, Gametogénesis.

Biología Reproductiva de H. (S.) glaberrima
XII
ABSTRACT
Sea cucumbers are a fishery resource in many Asian countries; indiscriminate exploitation
has led to local extinctions and fishing closures, shifting the pressure on species from
other areas, creating the need to understand the population biology of these new species
susceptible to extraction to create appropriate management tools. This is the first study
focused on the reproductive biology of the tropical cucumber Holothuria (Selenkothuria)
glaberrima. Twenty individuals per month were collected randomly from January to
October of 2010 in four rocky shores, in Santa Marta - Colombian Caribbean, area with a
dry climatic period of coastal upwelling of cold waters and a rainy season with higher
temperatures. This dioecious holothurian has a sex ratio of 1:1. The gonad is constituted
by numerous tubules, which may be in different stages of development, although with a
predominance of one in particular. Macroscopic and histological observations
characterized five gonadal stages: (I) immature, (II) developing, (III) mature, (IV) partially
spawned and (V) recovery. The monthly percentage of individuals in each stage and
gonadosomatic index revealed that the population of H. (S.) glaberrima has continuous
sexual maturity throughout the year (different individuals) and gonad development is not
completely synchronous between sexes. However, a proportion of the population
describes a cycle in which the maturity seems to be associated with the increase in
surface sea-water temperature at the end of the dry season and spawning shows a
greater association with the major rainy season between July and September.
Keywords: Aperiodic or continuous reproduction, Holothuria (Selenkothuria) glaberrima,
Holothuroidea, Colombian Caribbean, Gametogenesis.

Introducción
1
INTRODUCCION
Los holotúridos o pepinos de mar son una clase de equinodermos extensamente
distribuidos en ambientes marinos, desde las zonas intermareales hasta las
profundidades oceánicas y en todos los océanos del mundo. Desempeñan un papel
ecológico importante como modificadores de sustrato, al ingerir sedimentos, modificar su
composición y reciclar la materia orgánica (Hendler et al., 1995; Uthicke, 2001; Mangion
et al., 2003).
Aunque no exhiben dimorfismo sexual externo, su sistema de apareamiento ocurre con
sexos separados (macho - hembra) en relación 1:1, a excepción de algunos synáptidos
hermafroditas. El modo reproductivo en la mayoría de los casos implica liberación de
gametos al agua, fertilización externa y larva planctónica (Hendler et al., 1995, Ruppert y
Barnes, 1996). La reproducción es denso-dependiente, presenta bajos índices de
reclutamiento y madurez tardía, características que en conjunto pueden ocasionar alta
vulnerabilidad de las poblaciones frente a cambios ambientales y/o presiones
antropogénicas (Conand, 1993; Chao et al., 1994; Herrero-Pérezrul et al., 1999;
Ramofafia et al., 2000; Uthicke y Benzie, 2000; Uthicke et al., 2004; Abdel-Razek et al.,
2005; Uthicke y Conand, 2005; Muthiga, 2006; Toral-Granda y Martínez, 2007).
Presentan una sola gónada unida a la pared dorsal del cuerpo y formada por uno o dos
mechones de numerosos túbulos, ramificados o no. Un gonoducto simple, corre por el
mesenterio dorsal hasta el gonoporo, ubicado sobre la línea medio dorsal
inmediatamente antes del anillo tentacular (Hendler et al., 1995; Ruppert y Barnes, 1996)
(Figura 1). A medida que avanza la maduración, la capa germinal de la gónada sufre
cambios marcados de espesor que permiten un aumento significativo de la superficie
para el desarrollo de los gametos (Costelloe, 1985; Cameron y Fankboner 1986).
La morfología gonadal presenta modificaciones entre familias, especies de la misma
familia e incluso entre poblaciones, en forma (túbulos, nódulos, sáculos), longitud, grado
de ramificación, espesor, e incluso estadio de desarrollo (Smiley, 1988; Hamel y Mercier,
1996; Sewell et al., 1997; Foglietta et al., 2004). Basados en estas características
morfológicas y observaciones histológicas de los túbulos gonadales, varios autores han
descrito 4 estadios de madurez generales para machos y hembras: inmaduro (I),

Biología Reproductiva H. (S) glaberrima
2
recuperación o reposo (II), maduro (III) y desovado (IV) (Anexo 1). Sin embargo, se
pueden presentar estadios intermedios adicionales como crecimiento, desove parcial,
pre-desove, entre otros (Tuwo y Conand, 1992; Herrero-Perezrul et al., 1999; Ramofafia
et al., 2000; Toral - Granda y Martínez, 2007).
Figura 1. Anatomía interna de un pepino de mar común. (Tomada y modificada de
Ruppert y Barnes, 1996).
Diversos estudios de la biología reproductiva del grupo han señalado que los eventos de
maduración, crecimiento gonadal y desove siguen un modelo sincrónico al interior del
individuo y entre la mayoría de los individuos de la población local. Toda la gónada
madura al mismo tiempo, describiendo un patrón de reproducción cíclica, con
periodicidad anual, estacional o mensual. Esta periodicidad se da presumiblemente como
respuesta a la interacción de factores ambientales, químicos y hormonales, que modulan
la reproducción de la población (Conand, 1993; Ramofafia et al., 2000; Mercier et al.,
2000; Mercier y Hamel, 2009).
Por otra parte, se ha propuesto un patrón de reproducción poblacional ''continuo'' o
“aperiódico” para un número creciente de especies, principalmente las que se encuentran
a lo largo del ecuador o a grandes profundidades, sugiriendo que no hay una periodicidad
detectable, puesto que la mayoría de estas especies presentan individuos maduros a la
lo largo de todo el año. Sin embargo, aunque se describen periodos de reproducción más
extensos y eventos de desove en diferentes momentos del año, estos se asocian solo a
un porcentaje de la población cada vez (Ong Che y Gómez, 1985; Reichenbach, 1999;
Palazón-Fernández, 2001; Guzmán et al., 2003; Ramofafia et al., 2003; Mercier et al.,
2007; Mercier y Hamel, 2009). Adicionalmente, algunos trabajos registran, para algunas

Introducción
3
especies tropicales y templadas, un desarrollo asincrónico de la gónada, es decir túbulos
en diferente estadio de maduración presentes en la misma gónada (Theel, 1901;
Engstrom, 1980; Smiley, 1988; Tuwo y Conand, 1992; Hamel y Mercier, 1996; Foglietta
et al., 2004; Toral-Granda y Martínez, 2007), implicando que un individuo puede desovar
varias veces dentro de un mismo período.
La temporalidad de la gametogénesis en ambos patrones se ha asociado a la distribución
geográfica y a cambios en las condiciones ambientales, especialmente temperatura,
intensidad lumínica, fotoperiodo, ciclo lunar, disponibilidad de alimento y señales
químicas difusibles (Tuwo y Conand, 1992; Hamel y Mercier, 1995; Kubota y Tomari,
1998; Abdel-Razek et al., 2005; Muthiga, 2006; Asha y Muthiah, 2008; Mercier y Hamel,
2009) (Tabla 1).
Estudios ecológicos o de aspectos específicos de la clase Holothuroidea no se han
realizado hasta el momento en Colombia; los trabajos con el grupo se han limitado a la
realización de inventarios y notas taxonómicas o ecológicas sobre algunas especies de
diferentes áreas del Caribe y Pacífico (Álvarez–Larrauri, 1981; Caycedo, 1978 y 1979;
Pawson y Caycedo, 1980; Neira y Cantera, 1988; Pardo et al., 1988; Benavides-Serrato y
Borrero-Pérez, 2000; Arboleda, 2002; González et al., 2002; Ortiz-Gómez, 2006).
Holothuria (Selenkothuria) glaberrima Selenka, 1867 es un pepino aspidochirótido
pequeño, de cuerpo cilíndrico y piel suave, con gran cantidad de pies ambulacrales en la
superficie ventral. Presenta tentáculos largos, ramificados y dendríticos, los cuales
extiende y retrae en respuesta a la acción de las olas. Su color corporal puede variar de
café desteñido hasta casi negro, contrastando con tentáculos oscuros. Aunque se
distribuye desde las zonas de baja marea hasta los 42 m de profundidad, es una especie
gregaria típica del litoral rocoso, que usualmente se encuentra en áreas expuestas a la
acción del oleaje, aferrándose firmemente a las superficies externas de las rocas y entre
las grietas (Lewis, 1960; Hendler et al., 1995). Lewis (1960) en su estudio cualitativo
sobre la fauna de litorales rocosos de Barbados registró junio y julio como época
reproductiva de la especie, coincidiendo con el verano (valores más altos de temperatura
superficial del agua de mar). Describe huevos de 0,09 mm de diámetro color rosa
brillante, que pasaron al segundo estado celular 2 horas después de la fecundación,
dando lugar a una blástula nadadora en 4 horas y larva auricularia después de 3 días.

Tabla 1. Resumen de la temporalidad de la gametogénesis con algunos posibles factores de control exógeno en holoturoideos del
orden Aspidochirotida. (H.: Holothuria; I.: Isostichopus; P.: Parastichopus; A.: Actinopyga; (H.): Halodeima; (S.): Selenkothuria; (P.):
Panningothuria; (M.): Mertensiothuria; (Mi.): Microthele; (Me.): Metriatyla).
Especie Área Época reproductiva Factor control Ciclo Desarrollo individuos
Desarrollo gónada
Autor
H. (H.) floridana Florida Gametogénesis: primavera y verano - - - Asincrónico^ Engstrom, 1980
H. (H.) mexicana
H. (H.) mexicana Bocas del Toro
Panamá
Gónadas maduras todo el año. Máxima actividad reproductiva febrero-julio y julio-noviembre
respectivamente.
Posiblemente T°, salinidad y régimen
de lluvia Aperiódico*
Sincronía parcial+
- Guzmán, et al., 2003
I. badionotus
I. badionotus Venezuela Gónadas maduras todo el año. Pico reproductivo septiembre-noviembre.
- Aperiódico - - Palazón-Fernández, 2001
I. badionotus Venezuela Gónadas maduras todo el año.
Mayor proporción de julio-agosto - Aperiódico
Sincronía parcial
Asincrónico Foglietta et al.,2004
H. (S.) lubrica Golfo de California México
Maduración junio-agosto. Desove en verano (agosto)
Temperatura Anual** Sincrónico++ Sincrónico^^ López et al., 2010
I. fuscus Golfo de California México
Gametogénesis: enero - junio. Desove: julio-septiembre (verano)
Temperatura Anual Sincronía
parcial - Herrero-Pérezrul et al., 1999
I. fuscus Galápagos
Ecuador
Gónadas maduras todo el año. Cuatro picos: febrero, diciembre,
marzo y agosto Temperatura Aperiódico - Asincrónico
Toral-Granda y Martínez, 2007
*Aperiódico: Ciclo que presenta gónadas maduras todo el año, con la presencia de algunos picos reproductivos variables o no definidos. **Anual: Ciclo en el cual el desarrollo gametogénico ocurre a lo largo del año, con un pico de maduración y uno de desove definidos. +Sincronía Parcial: los estadios de madurez de machos y hembras se presentan simultáneamente solo en algunos meses.
++ Sincrónico: la gametogénesis de los individuos es simultánea entre sexos.
^Asincrónico: Desarrollo de los túbulos gonadales en cohortes, presentando diferentes estadios de desarrollo en una misma gónada. ^^Sincrónico: Todos los túbulos de la gónada presentan el mismo estadio de desarrollo.

……continuación Tabla 1
Especie Área Época reproductiva Factor control Ciclo Desarrollo individuos
Desarrollo gónada
Autor
P. parvimensis Baja California
México Desove febrero a mayo
(invierno/primavera). - Anual Sincrónico Sincrónico Fajardo-León et al., 2008
H. (P.) forskali Bretaña (Francia)
Crecimiento gonadal verano - otoño. Desove en abril.
Temperatura Anual Sincrónico Asincrónico Tuwo y Conand, 1992
H. (H.) atra Egipto
Gametos maduros todo el año. Abundancia máxima machos junio
– noviembre, hembras junio - diciembre.
- Anual Sincrónico Sincrónico Abdel-Razek et al., 2005
H. (M.) arenacava Kenia Crecimiento gonadal febrero-marzo. Desove marzo-mayo.
Intensidad lumínica y temperatura
Anual Sincrónico - Muthiga, 2006
H. (Mi.) fuscogilva Islas Salomón Inicio gametogénesis en marzo.
Desove - octubre Fotoperiodo y ciclo
lunar Anual Sincrónico Sincrónico Ramofafia et al, 2000
H. (H.) atra
Nueva Caledonia
Desove diciembre-febrero (aguas cálidas)
Temperatura
Anual
Sincrónico - Conand, 1993
H. (Mi.) fuscopunctata
A. mauritiana
A. echinites
H. s. versicolor
H. (Mi.) nobilis Desove - temporada fría
H. (Me.) scabra Desove diciembre-febrero,
segundo pico variable Bianual
A. mauritiana Guam Desove - temporada cálida Temperatura y
fotoperiodo Anual Sincrónico - Hooper et al., 2008
A. mauritiana Islas Salomón Desove octubre - diciembre Temperatura y
fotoperiodo Anual - - Ramofafia et al., 2001

Biología Reproductiva de H. (S) glaberrima
6
En Colombia este pepino ha sido registrado formando agrupaciones en la Isla de Tierra
Bomba-Cartagena y en plataforma rocosa de Punta Loma - Santa Marta, con
opistobranquios del género Balcis entre los tentáculos y acompañado de Holothuria
(Halodeima) grisea (Caycedo, 1978). Sin embargo la información sobre su ecología es
escasa y limitada, siendo este trabajo el primero en estudiar la biología reproductiva de la
especie.
Conocer los aspectos básicos de la biología reproductiva de las especies no solo es
importante desde el punto de vista ecológico, sino también como herramienta para
evaluar cómo pueden verse afectadas, y como responden a perturbaciones naturales y
artificiales (Mercier y Hamel, 2009). Los holotúridos han sido consumidos en países
asiáticos por siglos debido a sus propiedades curativas y alimenticias. Una vez
capturados, son eviscerados, hervidos y secados antes de ser comercializados. La pared
corporal seca se conoce como “beche–de–mer” o “trepang” y se consume principalmente
en platos con salsas o sopas. El precio del beche–de–mer varía grandemente entre
especies y entre la misma especie, dependiendo del tamaño del animal y el cuidado con
el que se procesa, alcanzando más de 300 USD/ kg si los animales están en un estado
perfecto y presentable. Sin embargo, especies de bajo valor comercial o animales mal
procesados se venden a una fracción de este precio (Purcell, 2010).
Aunque para H. (S.) glaberrima no se conoce un interés comercial y alimentario
específico, se ha encontrado que en conjunto con otras especies ha sido explotada
indiscriminadamente en las áreas de Santa Marta y la Guajira en el Caribe colombiano, lo
que se ha evidenciado en varios informes y decomisos de pesca ilegal (Borrero et al.,
2010; Benavides-Serrato, 2009). Esto genera preocupación ante el desconocimiento de
su biología y ecología en el área, pues aunque este holotúrido pequeño no sería
aceptado para comercialización como “beche–de–mer”, sí puede estar siendo usado en
fresco para sopas o acuarios (López et al., 2010). Teniendo en cuenta lo anterior, este
estudio describe algunos aspectos importantes de la biología reproductiva del pepino de
mar H. (S.) glaberrima para las condiciones locales de Santa Marta, aportando
información base de su ecología poblacional y posible manejo pesquero. Además, la
ocurrencia en el área de una época de afloramiento costero con temperaturas más bajas
que el normal del Caribe generó la hipótesis de posibles diferencias entre el ciclo
gametogénico local con el de la especie en otras áreas y con otros holotúridos tropicales.

Metodología
7
METODOLOGÍA
El estudio de la biología reproductiva de H. (S.) glaberrima se llevó a cabo durante 10
meses del año 2010 en cuatro litorales rocosos del área de Santa Marta al noreste del
Caribe colombiano: Punta Loma (11° 07’ 33,52’’ N - 74° 13’ 59,90’’ O), El Rodadero (11°
12’ 27,40’’ N – 74° 13’ 47,44’’ O), Inca-Inca (11° 16’ 07,42’’ N – 74° 14’ 02,35’’ O) y
Taganga (11° 16’ 07,42’’ – 74° 11’ 37,16’’ O) (Figura 2).
Figura 2. Área de estudio, señalando (estrellas) los litorales rocosos de Punta Loma,
Rodadero, Inca-Inca y Taganga donde se recolectaron los pepinos (Elaborado por el
Laboratorio de Sistemas de Información LabSIS- INVEMAR).
El área presenta un clima tropical semiárido, determinado por el régimen de los vientos
Alisios, con dos periodos climáticos principales: la época seca, de diciembre a abril, con

Biología Reproductiva de H. (S.) glaberrima
8
fuertes vientos provenientes del noreste que producen un fenómeno local de afloramiento
de aguas frías (20° – 25 °C), y una época húmeda mayor o de lluvias, de agosto a
noviembre, con las aguas superficiales cálidas (27° - 29 °C) y un importante aporte de
materia orgánica e inorgánica por descargas continentales (Márquez, 1982; Ramírez,
1983; Díaz et al., 2000).
Los individuos fueron recolectados en el primer metro de profundidad; debido a sus
hábitos crípticos fue necesario dar vuelta a las rocas para encontrarlos. Durante la
semana de luna llena y cubriendo las dos épocas climáticas, se colectaron 5 individuos al
azar en cada litoral para un total de 20 mensuales. En el día o días de muestreo se
tomaron datos de temperatura y salinidad con una sonda WTW modelo LF320; los datos
de precipitación y temperatura superficial del mar se obtuvieron del Centro de
Observación Marina Tadeista (COMARTA) y del Instituto de Teledetección Marina de la
Universidad del Sur de Florida (Institute for Marine Remote Sensing – IMaRS).
A partir de mayo, la cuota de recolecta se dividió entre la estación del Aeropuerto y
Taganga, puesto que los individuos recolectados en El Rodadero presentaban tallas muy
pequeñas y algunos individuos de Inca - Inca se encontraron muertos con lesiones en la
piel por perturbaciones físicas cuyo origen fue imposible determinar (mar de leva o
prácticas de pesca ilegal). En visitas posteriores no se encontraron individuos en esta
estación.
Los individuos recolectados se empacaron en bolsas herméticas individuales con agua
de mar y sal de Epsom para relajarlos (Solís Marín y Pérez, 1999); se almacenaron en
neveras de icopor y se transportaron a los laboratorios del Instituto de Investigaciones
Marinas y Costeras “José Benito Vives De Andréis” (INVEMAR). La disección se realizó
el mismo día o al día siguiente de la recolecta.
En el laboratorio, uno a uno los individuos se colocaron sobre una bandeja donde se
tomó una fotografía de registro y se midió la longitud total. Usando una balanza
electrónica Acculab v-200 de 0,01 gr de precisión se registró el peso total y el peso de la
pared corporal sin órganos internos (PCV). La gónada (PG) se pesó en una balanza
analítica Acculab ALC-210.4 de 0,0001 gr de precisión. Todos los tejidos se pasaron por
un papel secante aproximadamente un minuto antes de ser pesados con el fin de
disminuir la cantidad de agua en ellos. Con la información obtenida se calculó el índice
gonadosomático (IG) así: IG= (PG/PCV) x 100 %.

Metodología
9
La disección se realizó ubicando el individuo sobre su parte lateral, de manera que se
observaba la división dorso-ventral, con los tentáculos hacia el investigador; se hizo un
corte en dirección boca-ano siguiendo la línea imaginaria de la división dorso-ventral; la
gónada apareció del lado izquierdo unida al tejido intestinal. Dependiendo del estado de
madurez de la gónada está fue muy difícil de encontrar cuando era transparente y
pequeña o difícil de extraer cuando estaba tan desarrollada que se extendía a lo largo del
cuerpo enrollándose en el intestino y el sistema hemal. Para facilitar la observación y
manipulación fue necesario agregar agua sobre el individuo o sumergirlo en una pequeña
cantidad, permitiendo que los tejidos flotaran y se diferenciaran (Figura 3). La asignación
de color o tonalidad de los individuos y sus órganos se hicieron con base en la guía de
color para naturalistas (NCG) de Smithe (1975).
Figura 3. Estructuras internas del pepino de mar H. (S.) glaberrima en fresco.
Después de registrar la coloración, peso y longitud de las gónadas, estas se fijaron en
formalina al 4 % (en agua de mar) durante 24 - 48 horas; posteriormente se cambiaron a
alcohol al 70 % para su preservación y procesamiento histológico. La determinación del
sexo, estadio de madurez y ocurrencia de eventos reproductivos se asignó usando la
escala de clasificación de Ramofafia et al. (2000) y Tuwo y Conand (1992) (Anexo I)
basada en criterios de forma, color y longitud de la gónada y las características
microscópicas de los oocitos y el esperma.
Para el análisis histológico se seleccionó el tipo de túbulo más representativo en la
gónada y en ocasiones otros abundantes de diferente estadio. Se tomaron porciones de
aproximadamente 1 a 2 cm y se colocaron dentro de casetes histológicos; cuando el

Biología Reproductiva de H. (S.) glaberrima
10
tejido era muy delgado o pequeño se envolvió en papel de arroz para asegurar su
procesamiento. Una vez empacados, los casetes se llevaron al procesador de tejidos
Shandon Citadel 2000 donde se realizó la deshidratación pasándolo por una serie
sucesiva de alcoholes así: 70 % (2 h), 90 % (2 h), 96 % (dos recambios cada uno de 1 h),
100 % (tres recambios, uno de 1 h y dos de 2 h), Xileno (tres recambios, uno de 1 h y dos
de 1:30 h), y finalmente Parafina liquida (a 56 °C dos recambios de 2 y 3 h
respectivamente) que luego se dejó solidificar (Gómez- León, 2005).
Posteriormente, utilizando un micrótomo rotativo manual Microm HM325 se realizaron
cortes longitudinales a los túbulos de 5 o 7 micrómetros de espesor. Los cortes se tiñeron
con hematoxilina y eosina (Gómez- León, 2005) (Anexo 2) y se observaron al
microscopio. Se realizó un registro fotográfico de los estadios gametogénicos
encontrados con cámara Euromex DC30 y el software de procesamiento de imágenes
ImageFocus v 2.0, para medir el diámetro de los túbulos disponibles en machos y
hembras y 15 oocitos por hembra.
Análisis de datos
Se determinó si la proporción de sexos era 1:1 mediante la prueba Chi cuadrado (X2). Se
estableció la relación talla - peso general y por sexos usando regresión lineal. Para
explicar la distribución de los individuos maduros, dilucidar los estadios de madurez de
acuerdo a las tallas y brindar una base al manejo pesquero de la especie, se estableció
el peso de la primera madurez a partir del 50 % de la población muestreada en una curva
de frecuencia acumulada de los individuos sexuados (general y por sexo) con intervalos
de peso de 3 gr (Conand, 1993).
Para establecer posibles diferencias entre machos y hembras en longitud y peso se
realizaron pruebas Kolmogorov – Smirnov (K-S). La distribución de frecuencias de tallas
y pesos se realizó en primera instancia para todos los individuos juntos, usando
intervalos de 3 cm y 3 gr respectivamente; luego se calculó para hembras y machos por
separado. Además se utilizó una prueba de independencia X2 para estimar la variación
de cada tipo de túbulos a lo largo del muestreo.

Resultados
11
RESULTADOS
Características morfológicas de la población local
Los individuos de H. (S.) glaberrima recolectados presentaron piel verde oliva (greenish
olive NCG49), delgada y suave, en general limpia o cubierta con una ligera capa de
sedimento y algunos trozos de algas adheridos como camuflaje. Los especímenes casi
siempre tenían expuesta la corona de tentáculos dendríticos, con el color de la base
crema y las puntas sepia (cream color NCG54 y sepia NCG219), de aproximadamente 1
cm de largo cuando estaba extendida. La región dorsal era de apariencia lisa, bien
diferenciada de la ventral con pequeñas papilas y abundantes pies ambulacrales
formando una suela que utilizan para aferrarse a los espacios bajo las rocas y resistir el
oleaje. Generalmente se encontraron acompañados de H. (H.) grisea y en algunos
individuos se observó el gasterópodo Balcis sp. adherido a la piel o los tentáculos.
(Figura 4).
Figura 4. Individuos de H. (S.) glaberrima en la estación Punta Loma. A. camuflado en el
sedimento (flecha); profundidad 1,5 m. B. en proceso de alimentación, corona tentacular
expuesta; profundidad 0,5 m. C. gasterópodo Balcis sp. adherido a la región tentacular.
Proporción de sexos
Se recolectaron en total 197 individuos de H. (S.) glaberrima, 76 hembras y 72 machos,
32 con gónadas pero de sexo indeterminado y 17 sin tejido gonadal. El test de X2 no
mostró diferencias significativas en la proporción 1:1 entre sexos (X2= 0,11; gl=1;

Biología Reproductiva de H. (S.) glaberrima
12
p>0,05). A lo largo de los 10 meses de muestreo, hembras, machos e indeterminados
estuvieron presentes, mientras que los individuos sin gónada solo se presentaron en
enero, febrero y mayo (Figura 5).
Figura 5. Porcentaje de sexos mensual de H. (S.) glaberrima en Santa Marta, Colombia
durante el periodo de muestreo. (H: Hembras, M: Machos, I: Indeterminados, SG: Sin
gónada).
Talla y peso de la pared corporal
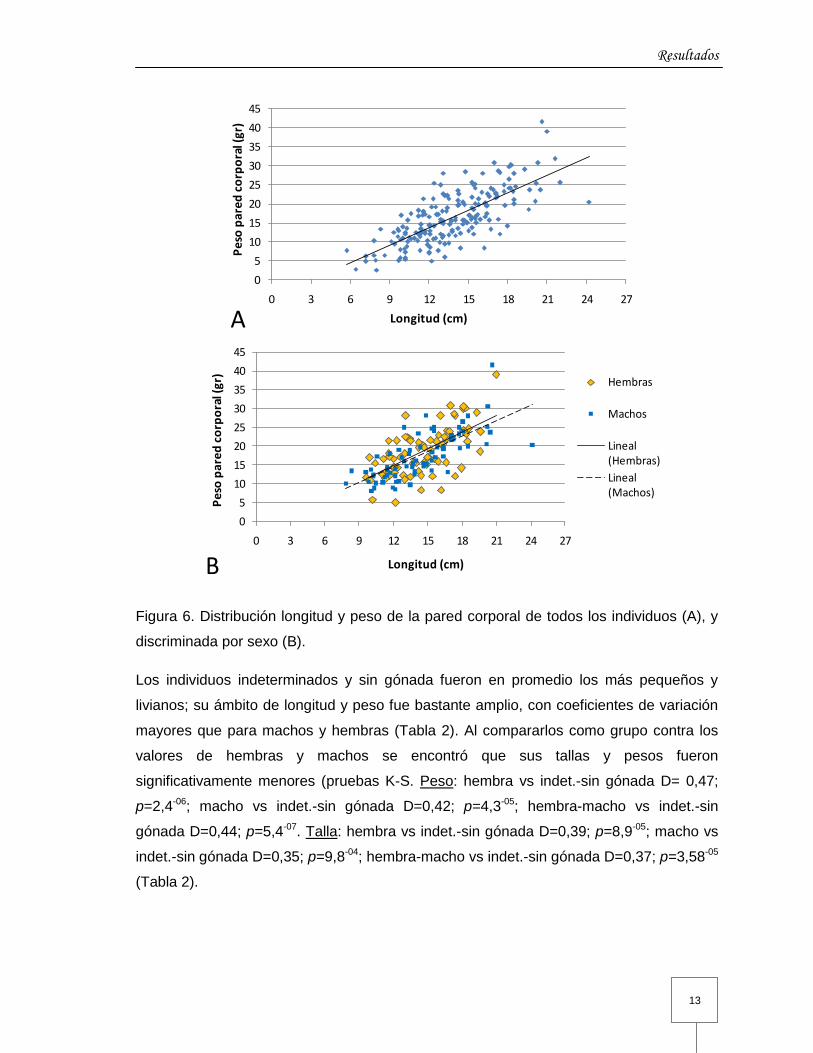
El análisis de regresión lineal mostró que existe una relación significativa entre longitud
del individuo y el peso de su pared corporal (p= 0,00) (Figura 6). La longitud de los
individuos muestreados varió entre 5,7 y 24,2 cm (promedio 13,8±3,30 cm). En promedio
las hembras fueron ligeramente más largas que los machos (Tabla 2), pero las
diferencias no fueron significativas (prueba K-S, D=0,10453; p=0,7919). La distribución
de frecuencias de longitud fue unimodal para todos (moda=12,6 cm) al igual que para los
sexos por separado (moda hembras=14,2 cm; machos=12,1 cm) (Figura 7).
En cuanto al peso de la pared corporal, este varió entre 2,6 y 41,6 g (promedio 16,7±6,89
g). El peso promedio de la pared corporal de las hembras fue más alto que el de los
machos (Tabla 2), pero sin diferencias significativas (prueba K-S, D=0,15789; p=0,2899).
La distribución de frecuencias de peso de la pared corporal fue unimodal (13,6 g), pero al
discriminarla por sexos fue bimodal en ambos casos (hembras=21,2 y 14,9 g;
machos=17,9 y 12,5 g) (Figura 7).
0
10
20
30
40
50
60
70
80
90
100
Ene Feb Mar Abr May Jun Jul Ago Sep Oct
Po
rce
nta
je d
e in
div
idu
os
%
SG I M H
Resultados
13
Figura 6. Distribución longitud y peso de la pared corporal de todos los individuos (A), y
discriminada por sexo (B).
Los individuos indeterminados y sin gónada fueron en promedio los más pequeños y
livianos; su ámbito de longitud y peso fue bastante amplio, con coeficientes de variación
mayores que para machos y hembras (Tabla 2). Al compararlos como grupo contra los
valores de hembras y machos se encontró que sus tallas y pesos fueron
significativamente menores (pruebas K-S. Peso: hembra vs indet.-sin gónada D= 0,47;
p=2,4-06; macho vs indet.-sin gónada D=0,42; p=4,3-05; hembra-macho vs indet.-sin
gónada D=0,44; p=5,4-07. Talla: hembra vs indet.-sin gónada D=0,39; p=8,9-05; macho vs
indet.-sin gónada D=0,35; p=9,8-04; hembra-macho vs indet.-sin gónada D=0,37; p=3,58-05
(Tabla 2).
0
5
10
15
20
25
30
35
40
45
0 3 6 9 12 15 18 21 24 27
Pes
o p
ared
co
rpo
ral (
gr)
Longitud (cm)A
0
5
10
15
20
25
30
35
40
45
0 3 6 9 12 15 18 21 24 27
Pe
so p
are
d c
orp
ora
l (g
r)
Longitud (cm)
Hembras
Machos
Lineal (Hembras)
Lineal (Machos)
B
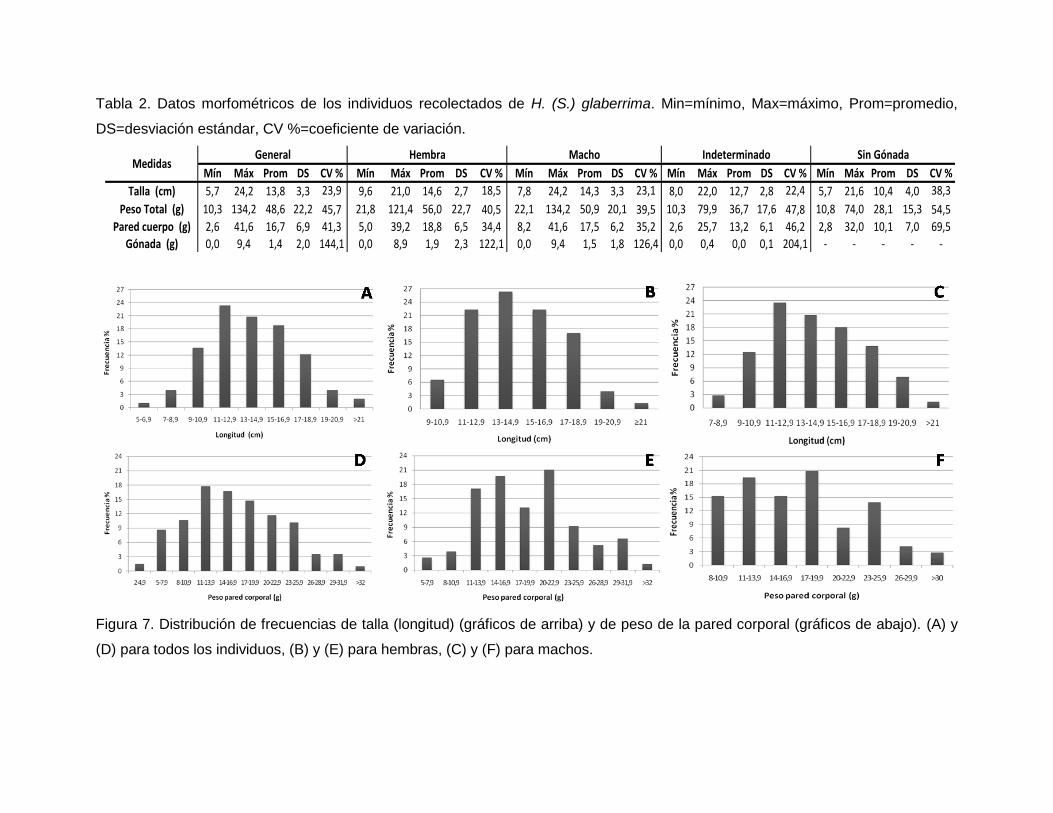
Tabla 2. Datos morfométricos de los individuos recolectados de H. (S.) glaberrima. Min=mínimo, Max=máximo, Prom=promedio,
DS=desviación estándar, CV %=coeficiente de variación.
Figura 7. Distribución de frecuencias de talla (longitud) (gráficos de arriba) y de peso de la pared corporal (gráficos de abajo). (A) y
(D) para todos los individuos, (B) y (E) para hembras, (C) y (F) para machos.
Mín Máx Prom DS CV % Mín Máx Prom DS CV % Mín Máx Prom DS CV % Mín Máx Prom DS CV % Mín Máx Prom DS CV %
Talla (cm) 5,7 24,2 13,8 3,3 23,9 9,6 21,0 14,6 2,7 18,5 7,8 24,2 14,3 3,3 23,1 8,0 22,0 12,7 2,8 22,4 5,7 21,6 10,4 4,0 38,3
Peso Total (g) 10,3 134,2 48,6 22,2 45,7 21,8 121,4 56,0 22,7 40,5 22,1 134,2 50,9 20,1 39,5 10,3 79,9 36,7 17,6 47,8 10,8 74,0 28,1 15,3 54,5
Pared cuerpo (g) 2,6 41,6 16,7 6,9 41,3 5,0 39,2 18,8 6,5 34,4 8,2 41,6 17,5 6,2 35,2 2,6 25,7 13,2 6,1 46,2 2,8 32,0 10,1 7,0 69,5
Gónada (g) 0,0 9,4 1,4 2,0 144,1 0,0 8,9 1,9 2,3 122,1 0,0 9,4 1,5 1,8 126,4 0,0 0,4 0,0 0,1 204,1 - - - - -
Hembra Macho Indeterminado Sin GónadaMedidas
General
Resultados
15
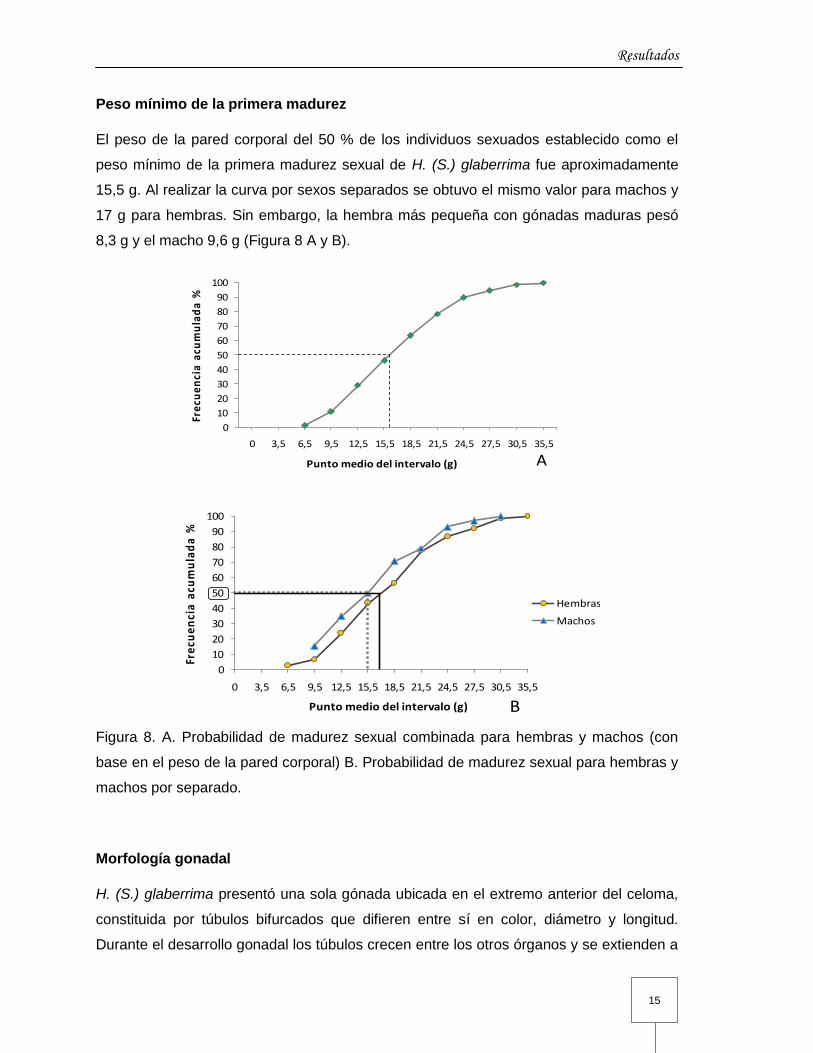
Peso mínimo de la primera madurez
El peso de la pared corporal del 50 % de los individuos sexuados establecido como el
peso mínimo de la primera madurez sexual de H. (S.) glaberrima fue aproximadamente
15,5 g. Al realizar la curva por sexos separados se obtuvo el mismo valor para machos y
17 g para hembras. Sin embargo, la hembra más pequeña con gónadas maduras pesó
8,3 g y el macho 9,6 g (Figura 8 A y B).
Figura 8. A. Probabilidad de madurez sexual combinada para hembras y machos (con
base en el peso de la pared corporal) B. Probabilidad de madurez sexual para hembras y
machos por separado.
Morfología gonadal
H. (S.) glaberrima presentó una sola gónada ubicada en el extremo anterior del celoma,
constituida por túbulos bifurcados que difieren entre sí en color, diámetro y longitud.
Durante el desarrollo gonadal los túbulos crecen entre los otros órganos y se extienden a
0
10
20
30
40
50
60
70
80
90
100
0 3,5 6,5 9,5 12,5 15,5 18,5 21,5 24,5 27,5 30,5 35,5
Fre
cue
nci
a a
cum
ula
da
%
Punto medio del intervalo (g) A
0
10
20
30
40
50
60
70
80
90
100
0 3,5 6,5 9,5 12,5 15,5 18,5 21,5 24,5 27,5 30,5 35,5
Fre
cue
nci
a a
cum
ula
da
%
Punto medio del intervalo (g)
Hembras
Machos
B

Biología Reproductiva de H. (S.) glaberrima
16
lo largo de la cavidad perivisceral ocupándola casi en su totalidad cuando están maduros.
Los túbulos de las gónadas se clasificaron en diferentes tipos de acuerdo con sus
características morfométricas, asociadas a estados de desarrollo gonadal, así:
Túbulos primarios (P): de color blanco translucido y generalmente con una sola
bifurcación, miden entre 0,2 y 2,9 cm aunque en pocas ocasiones alcanzan más de 2
cm. Cuando están formando parte de una gónada desarrollada tienen una longitud de
0,3 – 2,8 cm (1,47±0,81) en hembras y 0,2- 2,9 cm (1,64±0,74) en machos (Figura 9
A, J y K).
Túbulos medios (Md): usualmente presentan un color rosa claro (rose pink NCG108D)
en hembras y crema claro (cream color NCG54) en machos, permitiendo establecer el
sexo de los individuos a simple vista. Pueden estar bifurcados tres o cuatro veces, con
longitud entre 3,0 y 9,0 cm (5,0±1,39) en hembras y 3,0 a 8,2 cm (4,7±1,50) en
machos (Figura 9 B).
Túbulos maduros (M): con tres a cinco bifurcaciones, son rosa vivido en hembras
(poopy red NCG108A) y beige o naranja pastel en machos (chamois NCG123D y
warm buff NCG118); presentan tonalidades más intensas al estar bien desarrollados.
En hembras miden 4,6 a 17,7 cm (9,7±3,23) y en machos 5,9 a 18,5 cm (10,0±3,21)
(Figura 9 B, D y F).
Túbulos desovados (D): dependiendo del proceso de reabsorción varían de color
naranja claro a café (flesh ocher NCG132D o yellow ocher NCG123C y burnt sienna
NCG132) y frecuentemente tienen a lo largo puntos rojos o café oscuro; en algunos se
observó un canal interno más claro. Tanto en hembras (0,9–11,5 cm; 3,9±2,46) como
en machos (0,3–13,2 cm; 3,3±3,06) la longitud promedio de los túbulos no superó los
4 cm pero algunos pocos alcanzaron los 13 cm; los túbulos pueden encontrarse
formando gran parte de la gónada o como un pequeño penacho (Figura 9 G, H e I).
La gónada puede presentar uno, dos o tres tipos de túbulos al mismo tiempo y en
desarrollo consecutivo, generando una considerable superposición de estadios.

Resultados
17
Figura 9. Gónadas de H. (S.) glaberrima. A-B. Tipos de túbulos para ambos sexos. C-D.
Gónada hembras. E-F. Gónada machos. G-H-I. Gónadas con túbulos desovados. J-K.
Gónadas indeterminadas. (Todas las escalas 1 cm). D: Túbulos desovados; G: gónada;
In: intestino; M: Túbulos maduros; Md: Túbulos medios; P: Túbulos primarios; SH:
sistema hemal.

Biología Reproductiva de H. (S.) glaberrima
18
Ciclo reproductivo a nivel de túbulos
El análisis de presencia–ausencia de cada tipo de túbulo a lo largo del muestreo reveló
que los túbulos medios y maduros están presentes en todos los meses. De enero a abril
los pequeños o medios tuvieron la mayor proporción. Los túbulos desovados están
ausentes en marzo y abril, alcanzando su mayor representación entre julio y octubre
(Figura 10). La prueba de independencia X2 (p=0,00) mostró que la proporción de cada
tipo de túbulos varió significativamente a lo largo del año.
Figura 10. Gráfica de presencia - ausencia de túbulos pequeños o medios, maduros y
desovados en la gónada a lo largo de los meses de muestreo.
Ciclo reproductivo a nivel de individuos: estadios de madurez
El desarrollo general de los individuos a través del tiempo se estableció categorizando 5
estadios de desarrollo gonadal en los que se tuvieron en cuenta las características
morfológicas e histológicas de los túbulos más representativos encontrados en la gónada
(Tabla 3), así:
Indeterminado (I): gónada muy pequeña (0,3-3,1 cm, 1,5±0,68 cm) formada
exclusivamente por túbulos primarios; histológicamente pared gonadal gruesa, se
observa tejido conectivo en el lumen y no hay células sexuales diferenciables que
permitan asignar el sexo (Figura 11).
0
20
40
60
80
100
120
140
ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT
Fre
cu
en
cia
Re
lati
va
(%
)
Presencia - Ausencia
Peq-med
Desov
Maduros

Resultados
19
Figura 11. Características microscópicas del estadio Indeterminado de H. (S.) glaberrima.
A. 5x, B. 40x (Escalas 100 µm). L: lumen; PG: pared gonadal; TC: tejido conectivo.
En desarrollo (II): gónadas con túbulos medios principalmente, aunque en algunas se
presentan túbulos primarios en pequeñas cantidades. En las hembras el diámetro1 de los
túbulos fue de 58–621 µm (274±146 µm). Paredes gonadales gruesas y ligeramente
onduladas, una gran producción de oocitos previtelogenéticos basófilos forman un borde
interno oscuro en la capa germinal, mientras que oocitos tempranos de núcleo grande
comienzan a migrar hacia el lumen; diámetro de los oocitos entre 9,6 y 103 µm
1Diámetro medido en el microscopio sobre los túbulos disponibles en la placa histológica

Biología Reproductiva de H. (S.) glaberrima
20
(46,3±32,5 µm) (Figura 12 A y B). En los machos, los túbulos de la gónada presentan
paredes con pliegues del epitelio germinal bordeados por una capa densa de
espermatocitos que forman columnas hacia el centro del lumen; a medida que el estado
avanza los pliegues se reducen y los espermatozoides se hacen abundantes en el lumen.
Túbulos con un diámetro de 47 a 704 µm (331±179 µm) (Figura 13 A y B).
Figura 12. Características microscópicas del estadio En desarrollo en hembras de H. (S.)
glaberrima. A. 10x, B. 40x (Escalas 100 µm). L: lumen; Opv: Oocito previtelogenético;
Ovt: Oocito vitelogenético temprano; PG: pared gonadal.

Resultados
21
Figura 123. Características microscópicas del estadio En desarrollo en machos de H. (S.)
glaberrima. A. 10x, B. 40x (Escala 100 µm). Et: espermatozoides tempranos; Ec:
Espermatocito; L: lumen; Flecha: columnas hacia el lumen.
Maduro (III): la gónada formada en su mayoría por túbulos maduros que pueden estar
acompañados por una cantidad variable de primarios o medios; en algunas se puede
observar un penacho muy pequeño de túbulos desovados en un estado avanzado de
reabsorción. Histológicamente la pared gonadal es muy delgada y el lumen está lleno de
células sexuales bien desarrolladas. En las hembras oocitos eosinófilos que tienen
núcleo y nucléolo bien definidos, con células foliculares rodeándolos; el diámetro de
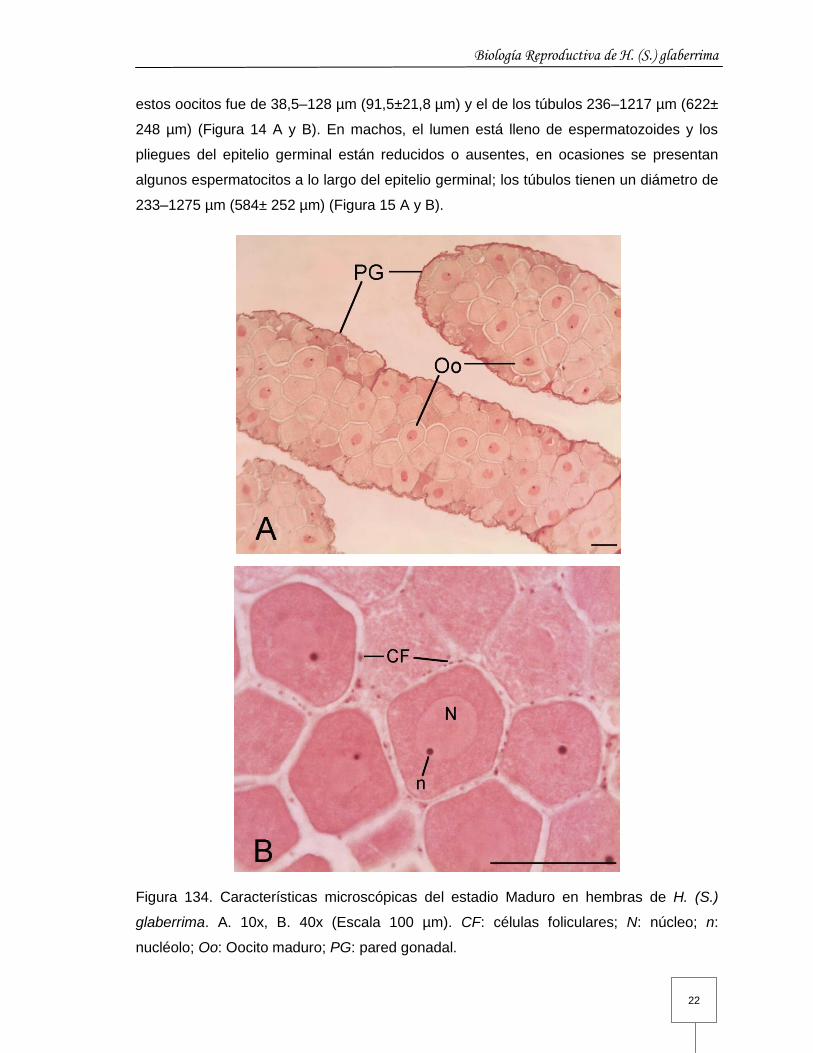
Biología Reproductiva de H. (S.) glaberrima
22
estos oocitos fue de 38,5–128 µm (91,5±21,8 µm) y el de los túbulos 236–1217 µm (622±
248 µm) (Figura 14 A y B). En machos, el lumen está lleno de espermatozoides y los
pliegues del epitelio germinal están reducidos o ausentes, en ocasiones se presentan
algunos espermatocitos a lo largo del epitelio germinal; los túbulos tienen un diámetro de
233–1275 µm (584± 252 µm) (Figura 15 A y B).
Figura 134. Características microscópicas del estadio Maduro en hembras de H. (S.)
glaberrima. A. 10x, B. 40x (Escala 100 µm). CF: células foliculares; N: núcleo; n:
nucléolo; Oo: Oocito maduro; PG: pared gonadal.

Resultados
23
Figura 14. Características microscópicas del estadio Maduro en machos de H. (S.)
glaberrima. A. 10x, B. 40x (Escala 100 µm). E: espermatozoides; Ec: Espermatocitos.
Parcialmente desovado (IV): principalmente túbulos desovados acompañados de túbulos
maduros y medios; presentan lúmenes vacíos o con algunas células sexuales
remanentes y fagocitos. En las hembras algunos oocitos remanentes dispersos rodeados
de tejido sobrante, con un diámetro de 72–134 µm (101±15,5 µm) y túbulos de 113–800
µm (433±171 µm) (Figura 16 A). En los machos, se observan muchos lúmenes vacíos,
aunque en algunos de ellos se ven parches difusos de espermatozoides remanentes;
túbulos con diámetro de 58–861 µm (355±167 µm) (Figura 16 B).

Biología Reproductiva de H. (S.) glaberrima
24
Figura 16. Características microscópicas del estadio Parcialmente desovado en H. (S.)
glaberrima. A. Hembra 10x, B. Macho 5x (Escala 100 µm). F: fagocitos.
Recuperación (V): las gónadas en este estadio están formadas por túbulos muy
pequeños desovados y primarios, el lumen se ve vacío y las paredes gonadales gruesas.
En ambos sexos se observan algunos oocitos o espermatocitos tempranos en la capa
germinal; también pueden presentarse células sexuales remanentes. El diámetro de los

Resultados
25
oocitos observados en este estadio fue de 10–111 µm (33,5±30,3 µm) y el diámetro de
los túbulos para hembras y machos fue de 55–360 µm (207±89 µm) y 113–271 µm
(188±61 µm) respectivamente (Figura 17 A y B).
Figura 17. Características microscópicas del estadio Recuperación en H. (S.) glaberrima.
A Hembra 5x, B. Macho 10x (Escala 100 µm). Or: Oocito remanente.
El porcentaje mensual de individuos en cada estadio de madurez reveló que H. (S.)
glaberrima es sexualmente activa a lo largo del año; 33 % de los individuos muestreados
estaban maduros, 21 % parcialmente desovados y 14 % inmaduros.
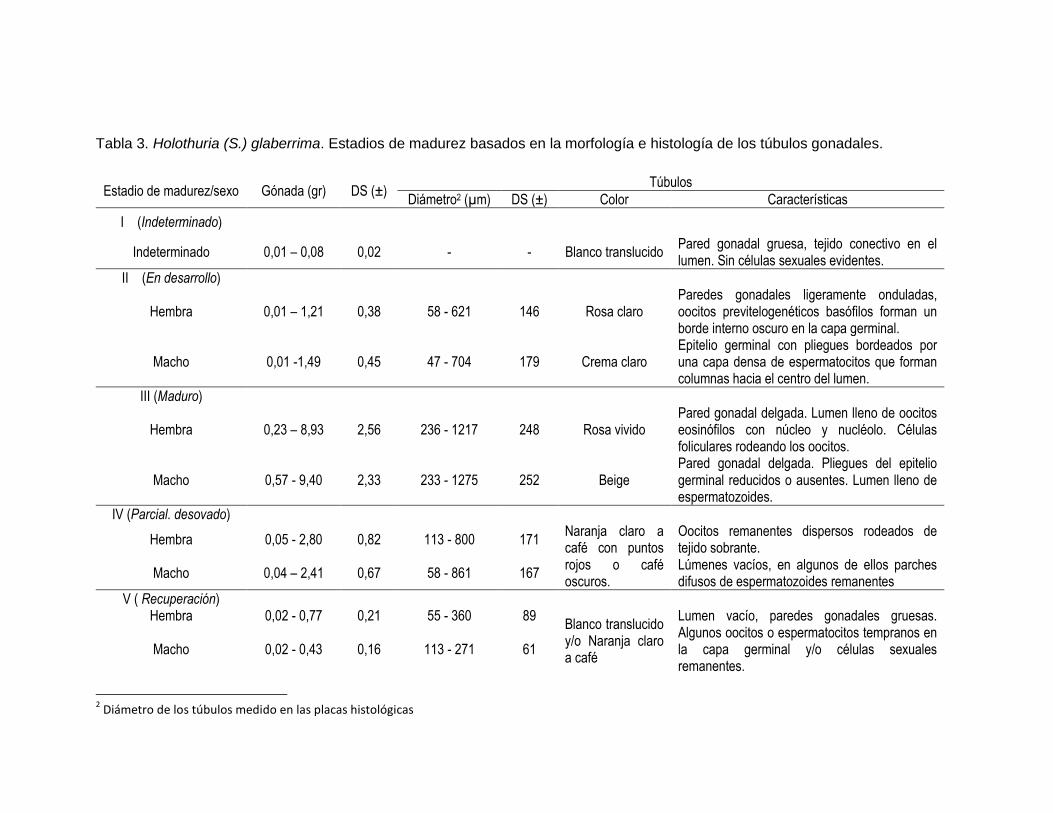
Tabla 3. Holothuria (S.) glaberrima. Estadios de madurez basados en la morfología e histología de los túbulos gonadales.
Estadio de madurez/sexo Gónada (gr) DS (±) Túbulos
Diámetro2 (µm) DS (±) Color Características
I (Indeterminado)
Indeterminado 0,01 – 0,08 0,02 - - Blanco translucido Pared gonadal gruesa, tejido conectivo en el lumen. Sin células sexuales evidentes.
II (En desarrollo)
Hembra 0,01 – 1,21 0,38 58 - 621 146 Rosa claro Paredes gonadales ligeramente onduladas, oocitos previtelogenéticos basófilos forman un borde interno oscuro en la capa germinal.
Macho 0,01 -1,49 0,45 47 - 704 179 Crema claro Epitelio germinal con pliegues bordeados por una capa densa de espermatocitos que forman columnas hacia el centro del lumen.
III (Maduro)
Hembra 0,23 – 8,93 2,56 236 - 1217 248 Rosa vivido Pared gonadal delgada. Lumen lleno de oocitos eosinófilos con núcleo y nucléolo. Células foliculares rodeando los oocitos.
Macho 0,57 - 9,40 2,33 233 - 1275 252 Beige Pared gonadal delgada. Pliegues del epitelio germinal reducidos o ausentes. Lumen lleno de espermatozoides.
IV (Parcial. desovado)
Hembra 0,05 - 2,80 0,82 113 - 800 171 Naranja claro a café con puntos rojos o café oscuros.
Oocitos remanentes dispersos rodeados de tejido sobrante.
Macho 0,04 – 2,41 0,67 58 - 861 167 Lúmenes vacíos, en algunos de ellos parches difusos de espermatozoides remanentes
V ( Recuperación) Hembra 0,02 - 0,77 0,21 55 - 360 89
Blanco translucido y/o Naranja claro a café
Lumen vacío, paredes gonadales gruesas. Algunos oocitos o espermatocitos tempranos en la capa germinal y/o células sexuales remanentes.
Macho 0,02 - 0,43 0,16 113 - 271 61
2 Diámetro de los túbulos medido en las placas histológicas

Resultados
27
Al calcular el porcentaje de machos y hembras en los estadios II, III, IV y V, se observó
que, aunque el desarrollo no es sincrónico entre sexos, se presenta una tendencia en la
cual los individuos maduros parecen ser más abundantes en los primeros meses del año,
mientras que de junio a octubre son más representativos los individuos parcialmente
desovados. Las mayores proporciones de hembras maduras (III) se presentaron en
enero (100 % - 4 ind.) y mayo (86% - 6 ind.) y estuvieron ausentes en septiembre. Los
machos maduros predominaron en abril y mayo (60 % – 6 ind. y 71% - 5 ind.
respectivamente); en general su proporción mensual fue más baja que la de las hembras.
Los individuos parcialmente desovados tuvieron las proporciones más bajas de enero a
mayo (0% hembras; machos 20% - 1 ind. y 29% - 2 ind.) aumentando sustancialmente de
junio a octubre y alcanzando sus mayores valores en las hembras para junio (71% - 5
ind.) y septiembre (50% - 5 ind.); en el caso de los machos en julio (57% - 4 ind.) y
agosto (45% - 5 ind.). Cabe resaltar que el mes de octubre fue el tercer valor más alto de
desovados para ambos sexos (44% hembras – 4 ind. y 43% machos – 3 ind.) (Figura 18).
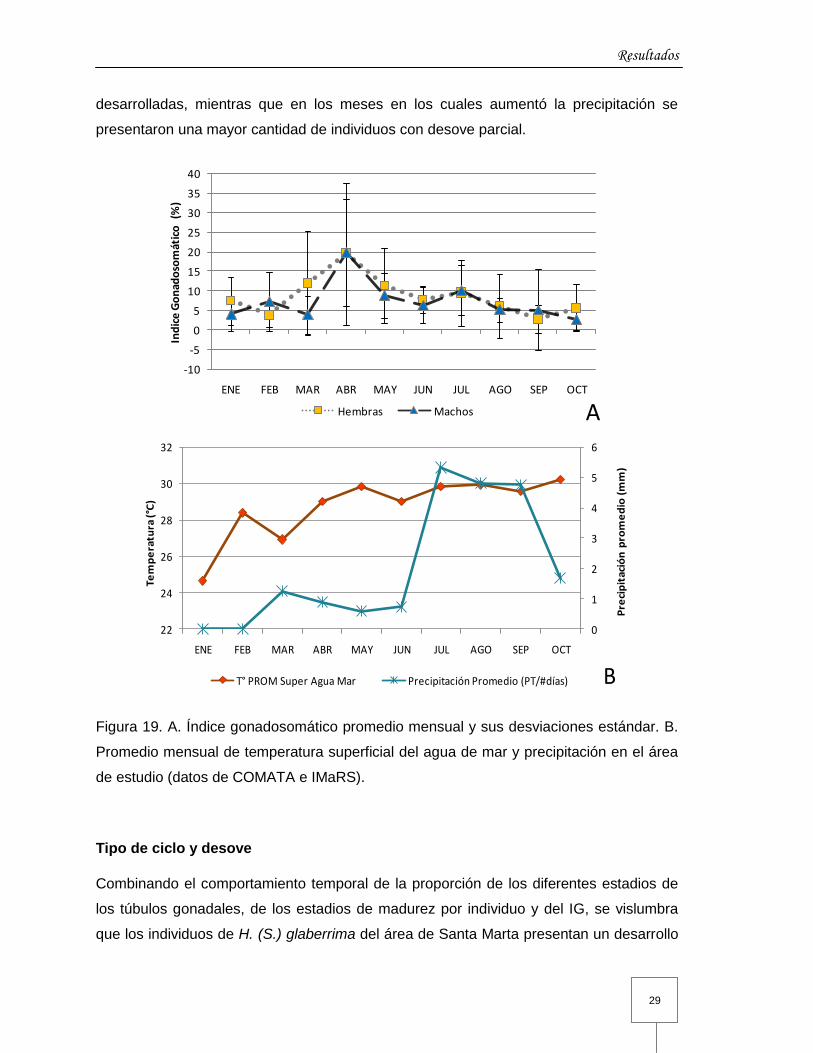
Índice gonadosomático
El índice gonadosomático (IG) osciló entre 0 y 56 % y presentó individuos con valores de
0 en todos los meses, con una evolución similar en los valores promedio de ambos
sexos, aunque generalmente mayor en hembras (Figura 19 A). Aunque la evolución de
los estadios no es del todo sincrónica entre sexos, los valores promedio más altos del IG
(hembras 19,6± 8,2 % y machos 19,8±13,6 %) se presentan en abril y mayo,
coincidiendo con las proporciones altas de hembras y machos maduros anteriormente
mencionadas. En julio el IG muestra un pico pequeño (9,5±8,4 % hembras y 10,2±6,6 %
machos) asociado a la presencia de una menor proporción de gónadas maduras con
respecto a abril. En septiembre y octubre se presentaron los valores promedio más bajos
de IG, asociados a la presencia de una gran cantidad de gónadas parcialmente
desovadas, siendo en hembras 2,8±3,7 % y en machos 2,6±2,8 %. La gran variabilidad
de estadios gametogénicos entre individuos se refleja en los valores altos de la
desviación estándar en todos los meses (Figura 19 A).

Biología Reproductiva de H. (S.) glaberrima
28
Figura 18. Distribución mensual de los porcentajes de estadios de madurez durante los
meses de muestreo para hembras y machos.
En cuanto a las condiciones ambientales se observó que la temperatura superficial del
agua de mar se incrementó entre marzo y abril, y se mantuvo relativamente alta hasta
octubre. Por su parte, la precipitación en la zona describió un aumento de julio a
septiembre, época descrita para el área como lluviosa mayor, y se vio intensificada por
el fenómeno de “La Niña”, que incremento las lluvias en toda Colombia (Figura 19 B). Las
variaciones de estas condiciones ambientales se vieron asociadas a los eventos del ciclo
gametogénico, puesto que en los meses en los cuales se incrementó la temperatura, se
presentaron las proporciones más altas de individuos con gónadas maduras y
Resultados
29
desarrolladas, mientras que en los meses en los cuales aumentó la precipitación se
presentaron una mayor cantidad de individuos con desove parcial.
Figura 19. A. Índice gonadosomático promedio mensual y sus desviaciones estándar. B.
Promedio mensual de temperatura superficial del agua de mar y precipitación en el área
de estudio (datos de COMATA e IMaRS).
Tipo de ciclo y desove
Combinando el comportamiento temporal de la proporción de los diferentes estadios de
los túbulos gonadales, de los estadios de madurez por individuo y del IG, se vislumbra
que los individuos de H. (S.) glaberrima del área de Santa Marta presentan un desarrollo
-10
-5
0
5
10
15
20
25
30
35
40
ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT
Ind
ice
Go
nad
oso
mát
ico
(%
)
Hembras Machos A
0
1
2
3
4
5
6
22
24
26
28
30
32
ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT
Pre
cip
itac
ión
pro
me
dio
(m
m)
Tem
pe
ratu
ra (
°C)
T° PROM Super Agua Mar Precipitación Promedio (PT/#días) B

Biología Reproductiva de H. (S.) glaberrima
30
progresivo de la gónada, con cierta sincronía entre túbulos y entre sexos, mostrando
individuos maduros a lo largo de todo el muestreo. Esto sugiere que la población puede
tener un ciclo de reproducción continua o aperiódica. Sin embargo, se observó que una
parte representativa de la población parece estar respondiendo a cambios ambientales,
asociando así, la época de maduración (abril – mayo) con un incremento en la
temperatura superficial del agua de mar, y el desove (junio- octubre) con la época
lluviosa mayor. Sin embargo, la presencia de hembras maduras y machos en
recuperación para el mes de enero pueden indicar la existencia de otro pico reproductivo
entre noviembre y enero que no fue cubierto en este estudio.

Discusión
31
DISCUSIÓN
En este estudio se confirmó que H. (S.) glaberrima es una especie gonocórica que no
presenta dimorfismo sexual externo y su proporción de sexos de 1:1. Como aporte
novedoso se describen por primera vez para esta especie varias características de su
ciclo reproductivo. Se registra la maduración progresiva y parcialmente sincrónica de los
túbulos gonadales, encontrando en la población individuos maduros durante todo el año.
Sin embargo, aunque no parece haber un ciclo definido y sincrónico y teniendo en cuenta
que el periodo de octubre a enero no fue muestreado, se observa que una proporción
importante de la población parece estar respondiendo a algunos cambios en las
condiciones ambientales, mostrando cierta asociación del proceso de maduración con el
incremento de la temperatura al final de la época seca (abril-mayo) y desoves a partir de
julio durante la época de lluvias, con sincronía parcial entre sexos.
Al igual que otras especies de la familia Holothuroidea, H. (S.) glaberrima tiene una
distribución de sexos 1:1. Esto parece ser un esquema general en holotúridos dioicos
(Engstrom, 1980; Conand 1981; Conand 1993; Herrero-Perezrul et al., 1999). No se
presentaron organismos hermafroditas, pero si individuos sin gónada. La presencia de
estos individuos en otras especies se ha atribuido al proceso de reabsorción del tejido
gonadal (Ramofafia, 2000; López et al., 2010). Aunque en este trabajo no se pudo
determinar la causa de esta ausencia, se puede asociar, en individuos pequeños, a que
aún no hubieran alcanzado la primera madurez sexual y en individuos de mayor peso a
un posible proceso de reabsorción.
La capacidad de elongación y retracción de los pepinos de mar les permite modificar su
tamaño en gran proporción, haciendo que este sea muy variable, mientras que el peso
húmedo de la pared corporal es una medida más confiable de la biomasa (Conand, 1981;
Tuwo y Conand, 1992; Fajardo et al., 2008). H. (S.) glaberrima mostró que existe una
relación positiva entre la longitud y el peso. Sin embargo, la dispersión de puntos indica
que hay variaciones en peso que no se explican solo por la talla y que dos individuos de
la misma talla puede pesar diferente debido a otras causas como factor de condición,
reproducción etc. Esto sugiere que, aunque el proceso de relajación disminuye la
variabilidad en los datos de longitud, el peso húmedo de la pared corporal refleja mejor el
estado de los individuos, además de hacer comparables los datos con otros autores.

Biología Reproductiva de H. (S.) glaberrima
32
El promedio de longitud y peso de la pared corporal de H. (S.) glaberrima fue 13,8±3,3
cm y 16,7± 6,9 g respectivamente, ubicándola como una especie pequeña comparada
con otras de la familia Holothuridae (Hendler, et al., 1995). El peso de la primera madurez
sexual del 50 % de la población fue alrededor de 15,5 g, mucho menor que el de las
especies de interés comercial (Conand, 1993; Guzmán et al., 2003; Toral-Granda y
Martínez, 2007). De hecho, Holothuria (Selenkothuria). lubrica, otra especie que es
también considerada pequeña, alcanza su primera madurez en 27,3 g (López et al.
2010). Las especies pequeñas crecen más rápido, tienen ciclos de vida más cortos y se
reproducen a edades tempranas (ver López, et al., 2010), lo cual puede explicar que el
peso promedio aquí encontrado sea cercano al de maduración.
La gónada (ovario y teste) de H. (S.) glaberrima, al igual que la de otros holotúridos
aspidochirotas es un ramillete de túbulos que se extiende a través del celoma a medida
que madura (Tuwo y Conand, 1992; Chao et al., 1995; Abdel-Razek et al., 2005; Toral –
Granda y Martínez, 2007; López et al., 2010). Sin embargo, se diferencia de otras
especies del orden por presentar simultáneamente en la misma gónada túbulos de
diferente grado de desarrollo, mostrando que la maduración tubular no procede a la
misma velocidad en todos generando una considerable superposición de los estadios. La
gónada puede presentar uno, dos o tres tipos consecutivos al mismo tiempo, por lo cual
las características de cada túbulo y su predominancia en la gónada fueron utilizadas para
adjudicar el estadio de desarrollo gonadal para el individuo. El proceso de desarrollo de
la gónada en diferentes clases de túbulos ha sido descrito también en varias especies y
por varios autores (Theel, 1901; Engstom, 1980; Smiley, 1988; Tuwo y Conand, 1992;
Hamel y Mercier, 1996; Belova y Viktorovskaya, 2007). Smiley (1988) describe el modelo
de desarrollo tubular (Tubule Recruitment Model) en el cual la gónada funcionaria como
una “banda transportadora” que permitiría el desarrollo y la reabsorción de los túbulos
año tras año. Así, en el año N, la gónada tiene túbulos maduros y en desarrollo, y para el
año N+1, los túbulos maduros serian desovados, los que estaban en desarrollo serían los
maduros y se generaría una nueva cohorte de túbulos en crecimiento, por lo cual nunca
se presenta reabsorción completa de la gónada y los individuos más viejos, presentarían
todas las clases de túbulos. Cabe destacar, que aunque H. (S.) glaberrima presentó una
gónada con clases de túbulos y oocitos sincrónicos en las distintas clases como lo
sugiere el modelo anterior, sí presento ciertas diferencias. En primera instancia, las
clases presentes entre individuos no siempre fueron las mismas; además, se presentaron

Discusión
33
individuos en recuperación y sin gónada; y el proceso no parece ser anual ni sincrónico
entre individuos. Estas diferencias con el modelo pueden deberse a la distribución
geográfica de la especie, en este caso su cercanía al Ecuador, y a los factores
ambientales que la están regulando, situación que se ha presentado también en
Holothuria (Halodeima) floridana y Holothuria (Halodeima) mexicana en la Florida (USA)
(Engstom, 1980); Isostichopus badionotus en las costas noroccidentales de Venezuela
(Foglietta et al., 2004) y en Panamá (Guzman et al., 2003); Holothuria (Panningothuria)
forskali en Bretaña (Francia) (Tuwo y Conand, 1992); Cucumaria frondosa y C. japonica
en Canadá y Japón (Hamel y Mercier, 1996; Belova y Viktorovskaya, 2007).
En el área de Santa Marta, se presentaron individuos maduros de H. (S.) glaberrima a
través de todo el periodo de muestreo, sugiriendo que la especie presenta una
reproducción continua o asincrónica. Empero, la producción continua de gametos a lo
largo del año es probablemente rara en un mismo individuo. La gametogénesis puede ser
escalonada entre diferentes individuos durante el año, por lo que la reproducción parece
ocurrir continuamente en la población entera. Aun no es claro si estas especies generan
y sintetizan gametos de forma “oportunista” (o continua) o si parte de la población sigue
un patrón cíclico (ej. mensual o bianual) que no es detectable fácilmente por las técnicas
y análisis convencionales (ver Mercier y Hamel, 2009).
Varios holotúridos tropicales tienen ciclos reproductivos continuos, anuales o
semianuales, regidos por la influencia de alguna variable ambiental como la temperatura
superficial del mar, las fases lunares, el fotoperiodo, o la disponibilidad de alimento, entre
otras (Harriott, 1985; Conand, 1993; Tuwo y Conand, 1992; Chao et al., 1995; Hopper et
al., 1998). En Santa Marta, a pesar de tener individuos con gónadas maduras a lo largo
de todo el muestreo y aunque la evolución de los estadios no fue del todo sincrónica
entre sexos, se manifestó un incremento en la actividad gametogénica (madurez) en abril
y mayo, asociada a la época de baja pluviosidad e incremento de temperatura, mientras
que el desove pareció coincidir con las mayores precipitaciones y altas temperaturas
(julio-septiembre). No obstante, la presencia de hembras maduras en enero, hace pensar
que puede presentarse otro pico en el periodo de octubre a enero que no se pudo
determinar en este estudio por falta de muestreo y que puede estar asociado a otros
cambios en las condiciones ambientales. Sin embargo, es posible que un gran número
de individuos de la población estén madurando en respuesta a un incremento en la
temperatura superficial del mar y los desoves ocurran en la época de mayores

Biología Reproductiva de H. (S.) glaberrima
34
temperaturas y precipitaciones (junio - octubre). Esto no implica que este sea el único
evento de madurez o desove, puesto que se encontraron al mismo tiempo individuos
maduros, parcialmente desovados y desovados en varios meses de estudio. Para esta
especie en Barbados el desove también estuvo asociado a los valores más altos de
temperatura (junio y julio) (Lewis, 1960).
Aunque en esta especie los individuos maduros se presentan a lo largo de todo el año,
siguiendo el patrón esperado para un holotúrido tropical, al analizar la dinámica de los
cambios estacionales en las gónadas de H. (S.) glaberrima, puede pensarse que
estudios más extensos permitirán dilucidar un posible ciclo semianual o bianual, que se
esté manifestando en una porción de la población y este asociado a los cambios
marcados de temperatura superficial del agua de mar y la precipitación que se presentan
en el área de Santa Marta. Eventos reproductivos asociados a la época de mayor
temperatura y lluvias mayores se han descrito para organismos sésiles como corales
(Montastrea cavernosa) (Acosta y Zea, 1997), esponjas (Zea, 1993), crustáceos (Balanus
trigonus), anélidos (Pomatoceros triqueter) (García y Salzwedel, 1993), entre otros.
CONCLUSIONES Y RECOMENDACIONES
En este estudio se encontró que la población de H. (S.) glaberrima de Santa Marta
presenta una gónada formada por numerosos túbulos en diferentes estadios de
desarrollo, lo cual sugiere un ciclo reproductivo poblacional continuo, con al menos un
desove anual por individuo, principalmente asociado a la época de lluvias.
Las características macroscópicas y microscópicas de la gónada de H. (S.) glaberrima
permitieron establecer cinco estadios de madurez, que brindan una guía para futuros
estudios de esta y otras especies de holotúridos.
Holothuria (S.) glaberrima del área de Santa Marta presentó una proporción de sexos 1:1
y una talla de primera madurez de ≈ 15,5 g de pared corporal.
Aunque este estudio refleja que la mayoría de los individuos de H. (S.) glaberrima
presentan un posible patrón de desarrollo gonadal en respuesta a variables ambientales,
esto no es concluyente, por lo que es necesario hacer investigaciones más largas que
permitan establecer el patrón completo a lo largo de dos o tres años de observación.

Discusión
35
Basados en los resultados obtenidos en este estudio se recomienda ante posibles
eventos de extracción la capturar se realice sobre individuos con una talla mayor o igual
a 13 cm (Equivalente a 15,5 g de la pared corporal).
De ser necesario tomar medidas de manejo pesquero para controlar el recurso, debe
ponerse especial atención a la época de lluvias pues está coincide con el periodo de
desove.
Es importante realizar futuros estudios de crecimiento y dinámica poblacional que
complementen la información obtenida en esta investigación.
Para un muestreo exitoso y seguro de esta especie de litoral, es recomendable tener un
equipo de dos o tres personas buscando simultáneamente, ya que es tedioso y
complicado encontrar los individuos; además se debe usa pantalón, camisa manga larga
y guantes, que brinden protección contra el ambiente rustico del litoral y su fauna
acompañante.

Bibliografía
36
BIBLIOGRAFÍA
Abdel-Razek, F.A., S.H. Abdel-Rahman, N.A. El-Shimy y H.A. Omar. 2005. Reproductive
biology of the tropical sea cucumber Holothuria atra (Echinodermata: Holothuroidea) in
the red sea coast of Egypt. Egy. J. Aquat. R., 31 (2): 383-402.
Acosta, A. y S. Zea. 1997. Sexual reproduction of teh reef coral Montastrea cavernosa
(Scleractinia: Faviidae) in the Santa Marta area, Caribbean coast of Colombia. Mar. Biol.,
128: 141-148.
Álvarez-Larrauri, L.R. 1981. Listado preliminar de los equinodermos de la costa Atlántica
de Colombia. Bol. Mus. Mar, 10: 24-39.
Arboleda, E. 2002. Estado actual del conocimiento y riqueza de peces, crustáceos
decápodos, moluscos, equinodermos y corales escleractíneos del océano Pacifico
colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá. 125 p.
Asha, P.S. y P. Muthiah. 2008. Reproductive biology of the comercial sea cucumber
Holothuria spinifera (Echinodermata: Holothuroidea) from Tuticorin, Tamil Nadu, India.
Aquacult. Int., 16: 231-242.
Belova G. V. y G. I. Viktorovskaya. 2007. Morphometric characteristic of sex cells and
seasonal changes of female gonads of Cucumaria japonica (Semper, 1868)
(Echinodermata: Holothuroidea) from Peter the Great Bay, Sea of Japan. Rus. J. Mar.
Biol., 33 (3): 173–180
Benavides-Serrato, M. 2009. Decomiso de pepinos de mar en la zona costera de Santa
Marta. Concepto técnico tipo c. Instituto de investigaciones marinas y costeras “José
Benito Vives de Andréis”. Santa Marta. 15p.
Benavides-Serrato, M. y G.H. Borrero-Pérez. 2000. Equinodermos de la franja superior
del talud continental del Caribe Colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano,
Santa Marta. 527p.

Biología Reproductiva de H. (S.) glaberrima
37
Borrero-Pérez G. H, M. Santos-Acevedo, E. Ortiz-Gómez. 2010. Perspectives and
present situation of sea cucumber fisheries in the Colombian Caribbean Sea. In: Harris
L.G., S.A. Böttger, C.W. Walker, M.P. Lesser (eds) Echinoderms: Durham. Proc 12th Int
Echinoderm Conf. CRC Press, Taylor and Francis group, Balkema, Leiden, pp 588.
Cameron, J.L. y P.V. Fankboner. 1986. Reproductive biology of the comercial sea
cucumber Parastichopus californicus (Stimpson) (Echinodermata: Holothuroidea). I.
Reproductive periodicity and spawning behavior. Can. J. Zool., 64: 168 – 175.
Caycedo, I. 1978. Holothurioidea (Echinodermata) de aguas someras en la costa norte de
Colombia. An. Inst. Inv. Mar. – Punta Betín, 10: 149 – 198.
Caycedo, I. 1979. Observaciones de los equinodermos en las Islas del Rosario. An. Inst.
Inv. Mar. – Punta Betín, 11: 39 – 47.
Chao, S. M., C. E. Chen y P. S. Alexander. 1994. Reproduction and growth of Holothuria
atra (Echinodermata: Holothuroidea) at two contrasting sites in southern Taiwan. Mar.
Biol., 119: 565-570.
Chao, S. M., C. E. Chen y P. S. Alexander. 1995. Reproductive cycles of tropical sea
cucumbers (Echinodermata: Holothuroidea) in southern Taiwan. Mar. Biol., 122:289-295
Conand, C. 1981. Sexual cycle of three commercially important holothurian species
(Echinodermata) from the lagoon of New Caledonia. Bull. Mar. Sci., 31(3): 523-543
Conand, C. 1993. Reproductive biology of the holothurians from the major communities of
the New Caledonian Lagoon. Mar. Biol., 116: 439-450.
Costelloe, J. 1985. The anual reproductive cycle of the holothurian Aslia lefevrei
(Dendrochirota: Echinodermata). Mar. Biol., 88: 155-165.
Díaz, J. M., L.M. Barrios, M. H. Cendales, J. Garzón-Ferreira, J. Geister, M. López-
Victoria, G.H. Ospina, F. Parra-Velandia, J. Pinzón, B. Várgas-Angel, F. Zapata y S. Zea.

Bibliografía
38
2000. Áreas coralinas de Colombia. Colombia: Invemar-Serie Publicaciones Especiales
No. 5. 176 pp.
Engstrom, N. A. 1980. Reproductive cycles of Holothuria (Halodeima) floridana, H. (H.)
mexicana and their hybrids (Echinodermata:Holothuroidea) in southern Florida, U.S.A. Int.
J. Invertebr. Repr., 2: 237-244.
Fajardo León, M. C., Suárez Higuera, M. C. L., A. Del Valle Manríquez, A. Hernández
López. 2008. Biología reproductiva del pepino de mar Parastichopus parvimensis
(Echinodermata: Holothuroidea) de Isla Natividad y Bahía Tortugas, Baja California Sur,
México. Cienc. Marinas, 34 (002): 165-177.
Foglietta L. M., M. I. Camejo, L. Gallardo, F. C. Herrera. 2004. A maturity index for
holothurians exhibiting asynchronous development of gonad tubules. J. Exp. Mar. Biol.
Ecol., 303: 19– 30.
García, C. y H. Salzwedel. 1993. Recruitment patterns of sessile invertebrates onto
fouling plates in the bay of Santa Marta, Colombian Caribbean. Bol. Invemar, 22 (1): 30-
44.
Giraldo, R. y N. H. Campos. 1997. Una aplicación del análisis de varianza de medidas
repetidas en un experimento con metales pesados. Rev. Acad. Colomb. Cienc. Vol. XXI.
(81): 467 – 472.
Gómez-León, J. 2005. Estudio Epizoótico de Agentes Patógenos en Almeja Fina
(Ruditapes decussatus) Cultivada en Galicia. Tesis Doctorado Cienc. Biol., Univ. de
Santiago de Compostela, España. 220 p.
González, D.N., O.D. Solano y G.R. Navas. 2002. Equinodermos colectados por la
expedición CIOH-INVEMAR-SMITHSONIAN desde Cartagena hasta el Golfo de Urabá,
Caribe Colombiano. Bol. Inst. Inv. Mar. Cost., 31: 85-132.

Biología Reproductiva de H. (S.) glaberrima
39
Guzmán, H.M., C.A. Guevara, I.C. Hernández. 2003. Reproductive cycle of two
commercial species of sea cucumber (Echinodermata: Holothuroidea) from Caribbean
Panama. Mar. Biol., 142: 271-279.
Hamel, J.F. y A. Mercier. 1995. Spawning of the sea cucumber Cucumaria frondosa in the
St Lawrence Estuary, eastern Canada. SPS Beche-de-mer Info. Bull., 7: 12-18.
Hamel, J.F. y A. Mercier. 1996. Studies on the reproductive biology of the Atlantic sea
cucumber Cucumaria frondosa. SPS Beche-de-mer Info. Bull., 8: 22-33.
Harriott, V. J. 1985. Reproductive Biology of three congeneric Sea Cucumber Species,
Holothuria atra, H. impatiens and H. edulis, at Heron Reef, Great Barrier Reef. Aust. J.
Mar. Freshw. Res., 36: 51-7
Hendler G., J. Miller, D. Pawson y M. Porter. 1995. Echinoderms of Florida and the
Caribbean Sea. Stars, Sea Urchins and allies. Smithsonian Institution. Hong Kong, 390
pp.
Herrero-Pérezrul, M. D., H. Reyes Bonilla, F. García-Domínguez y C. E. Cintra-
Buenrostro. 1999. Reproduction and growth of Isostichopus fuscus (Echinodermata:
Holothuroidea) in the southern Gulf of California, México. Mar. Biol., 135: 521-532.
Hooper, D. R., C. L. Hunter y R. H. Richmond. 1998. Sexual reproduction of the tropical
sea cucumber, Actinopyga mauritana, in Guam. Bull. Mar. Sci., 63 (1): 1-9.
Kubota, T. y M. Tomari. 1998. Reproduction in the apodid sea cucumber Polycheira
rufescens: semilunar spawning rhythm and sex change. J. Mar. Biol. Ass. U.K., 78: 249-
267.
Lewis, J. B. 1960. The fauna of rocky shores of Barbados, West Indies. Can. J. Zool., 38:
391- 435.
López, J. S., D. Herrero Pérezrul, H. Reyes Bonilla, F. García Domínguez, J. Turrubiates
Morales. 2010. Reproductive cycle of Holothuria (Selenkothuria) lubrica Selenka, 1867

Bibliografía
40
(Echinodermata: Holothuroidea) in Bahía de La Paz, México. In: Harris L.G., S.A. Böttger,
C.W. Walker, M.P. Lesser (eds) Echinoderms: Durham. Proc 12th Int Echinoderm Conf.
CRC Press, Taylor and Francis group, Balkema, Leiden, 487 – 493.
Mangion, P., D. Taddei, P. Frouin y C. Conand. 2003. Feeding rate and impact of
sediment reworking by two deposit feeders Holothuris leucospilota and Holothuria atra on
a fringing reef (Reunion Island, Indian Ocean). En: Heinzeller T. y J.H. Nebelsick, E.
2004. Echinoderms, München, Taylor & Francis Group, London. p 311-317.
Márquez C., G. E. 1982. Los sistemas ecológicos marinos del sector adyacente a Santa
Marta, Caribe colombiano I: Generalidades. Ecología Tropical, 2 (1): 5-18.
Mercier, A., S. C. Battaglene y J. F. Hamel. 2000. Periodic movement, recluitment and
size-related distribution of the sea cucumber Holothuria scabra in Solomon Islands.
Hydrobiologia, 440: 81-100.
Mercier, A., R. H. Icaza y J.F. Hamel. 2007. Long-term study of gamete release in a
broadcast-spawning holothurian: predictable lunar and diel periodicities. Mar. Ecol. Prog.
Ser., 329: 179-189.
Mercier, A. y J. F Hamel. 2009. Advances in Marine Biology Endogenous and Exogenous
Control of Gametogenesis and Spawning in Echinoderms. Elsevier, London. 297pp.
Morgan, A. D. 2000. Induction of spawning in the sea cucumber Holothuria scabra
(Echinodermata: Holothuroidea). J. Wld. Aquacult. Soc., 31 (2): 186-194.
Muthiga, N. A. 2006. The reproductive biology of a new species of sea cucumber,
Holothuria (Mertensiothuria) arenacava in a Kenyan marine protected area: the possible
role of light and temperature on gametogenesis and spawning. Mar. Biol., 149: 585–593.
Neira, R. y J. Cantera. 1988. Notas taxonómicas y ecológicas de los equinodermos de
Bahía Málaga (Costa Pacífica Colombiana). Memorias del VI Seminario Nacional de las
Ciencias y Tecnologías del Mar. Bogotá. p.121-129.

Biología Reproductiva de H. (S.) glaberrima
41
Ong Che, R. y E. Gómez, 1985. Reproductive periodicity of Holothuria scabra Jaeger
at Calatagan, Batangas, Philippines. Asian Mar. Biol., 2: 21-30.
Ortiz-Gómez, E.P. 2006. Cambio espícular en algunos pepinos de mar del género
Holothuria Linnaeus, 1758 (Echinodermata: Holothuroidea) presentes en el Caribe y
pacífico colombianos. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá. 109 p.
Palazón Fernandez J. L. 2001. Reproducción del pepino de mar, Isostichopus badionotus
Selenka (Echinodermata: Holothuroidea). IX Congreso Latinoamericano sobre Ciencias
del Mar. San Andres Isla. p. 419.
Pardo, R., R. Neira, y J. Cantera. 1988. Taxonomía y ecología de los equinodermos
litorales del Pacífico Colombiano. Memorias del VI Seminario Nacional de las Ciencias y
Tecnologías del Mar. Bogotá. p. 115-120.
Pawson, D. y E. Caycedo. 1980. Holothuria (Thymiosycia) thomasi new species, a large
Caribbean coral reef inhabiting sea cucumber (Echinodermata: Holothuroidea). Bull. Mar.
Sci., 30 (2): 454 – 459.
Purcell, S. W. 2010. Manejo de las pesquerías de pepino de mar con un enfoque
ecosistémico. Lovatelli, A., M. Vasconcellos y Y. Yimin (eds). FAO Documento técnico de
Pesca y Acuicultura. No. 520. Roma. 169 p.
Ramírez, G. 1983. Características fisicoquímicas de la Bahía de Santa Marta (agosto
1980-julio 1981). An. Inst. Inv. Mar. Punta de Betín, 13: 111-121.
Ramofafia, C., S.C. Battaglene, J.D. Bell y M. Byrne. 2000. Reproductive biology of the
commercial sea cucumber Holothuria fuscogilva in the Solomon Islands. Mar. Biol., 136:
1045-1056.
Ramofafia, C., M. Byrne y S. Battaglene. 2001. Reproductive biology of the intertidal sea
cucumber Actinopyga mauritana in the Solomon Islands. J. Mar. Biol. Ass. U.K., 81: 523-
531

Bibliografía
42
Ramofafia, C., M. Byrne y C. S. Battaglene. 2003. Reproduction of the commercial sea
cucumber Holothuria scabra (Echinodermata: Holothuroidea) in the Solomom Islands.
Mar. Biol., 142: 281-288.
Reichenbach, N. 1999. Ecology and fishery biology of Holothuria fuscogilva
(Echinodermata: Holothuroidea) in the Maldives, Indian Ocean. Bull. Mar. Sci., 64 (1):
103–113.
Ruppert, E. E. y R. D. Barnes. 1996. Zoología de los Invertebrados. McGraw-Hill
Interamericana, México. 1114 pp.
Sewell, M. A., P. A. Tyler, C. M. Young, C. Conand. 1997. Ovarian Development in the
Class Holothuroidea: a Reassessment of the "Tubule Recruitment Model". Biol. Bull. 192:
17-26.
Smiley, S. 1988. The Dynamics of Oogenesis and the Annual Ovarian Cycle of Stichopus
californicus (Echinodermata:Holothuroidea). Biol. Bull. 175: 79-93.
Smithe, F. B. 1975. Naturalist's Color Guide. American Museum of Natural History (New
York), New York.
Solís-Marín, F. A. y E. M. Pérez. Taxonomía de Equinodermos. ICM y L-UNAM.
CONACYT, ECOSUR. México. 78 pp. 1999.
Theel, H. 1901. A singular case of Hermaphodism in Holothurids. Bihang Svenska
Vetensk Acad.Handl 27: Afd.4., No. 6. 4-38.
Toral-Granda, M. V. y P. C. Martínez. 2007. Reproductive biology and population
structure of the sea cucumber Isostichopus fuscus (Ludwig, 1875) (Holothuroidea) in
Caamaño, Galápagos Islands, Ecuador. Mar. Biol., 151: 2091–2098.
Tuwo, A. y C. Conand, 1992. Reproductive Biology of the Holothurian Holothuria forskali
(Echinodermata). J. Mar. Biol. Ass. UK., 72: 745-758.

Biología Reproductiva de H. (S.) glaberrima
43
Uthicke S. 2001. Interactions between sediment-feeders and microalgae on coral reefs:
grazing losses versus production enhancement. Mar. Ecol. Prog. Ser., 210: 125-138.
Uthicke S, y J. Benzie. 2000. Effect of beche-de-mer fishing on densities and size
structure of Holothuria nobilis (Echinodermata: Holothurioidea) populations on the great
Barrier Reef. Coral Reefs, 19: 271-276.
Uthicke S. y C. Conand. 2005. Local examples of beche-de-mer overfishing: An initial
summary and request for information. SPS Beche-de-mer. Info. Bull., 21: 9-14.
Uthicke S., D. Welch y J. Benzie. 2004. Slow growth and lack of recovery in overfished
holothurians on the Great Barrier Reef: Evidence from DNA fingerprints and repeated
large-scale surveys. Conservation Biology, 18: 1395-1404.
Zea, S. 1993. Recruitment of Demosponges (Porifera, Demospongiae) in rocky and coral
reef habitats of Santa Marta, Colombian Caribbean. Mar. Ecol., 14 (1): 1-21.

Glosario
44
GLOSARIO
Basado en Morgan (2000)
Células foliculares (CF): célula de apoyo alrededor del huevo en desarrollo.
Epitelio germinal: pared interna de los túbulos gonadales.
Espermatozoides (E): espermatozoides maduros en el lumen de los túbulos de las
gónadas, cabeza pequeña y flagelo largo.
Espermatocitos (Ec): núcleo pequeño con cromatina de color oscura, generalmente en
los bordes del lumen.
Fagocitos (F): célula que absorben los oocitos vitelogéneticos.
Lumen (L): espacio interior del túbulo.
Oocito Maduro (Oo): oocito desarrollado, con la cromatina condensada. Presenta
núcleo y nucléolo.
Oocito previtelogenético (Opv): oocitos previos al desarrollo del núcleo. Generalmente
ubicados cerca del epitelio germinal.
Oocito remanente (Or): Oocitos desarrollados que no son expulsados en el momento
del desove y que sufren proceso de reabsorción.
Oocito vitelogenético temprano (Ovt): estado del oocito entre previtelogenético y
maduro. Usualmente se observan migrando del epitelio germinal hacia l lumen.
Vitelogénesis: Proceso de formación de los materiales de reserva del huevo.
.

Anexos
45
ANEXOS
ANEXO 1. Estadios de madurez generales propuestos para holotúridos (Tomados y
modificados de Tuwo y Conand, 1992; Herrero–Perezrul et al., 1999; Ramofafia, 2000;
Toral –Granda y Martínez, 2007; Fajardo et al., 2008)
Inmaduro (I): Gónadas sin células sexuales diferenciables o con gametos que no
presentan estructuras celulares definidas. Abundante tejido conectivo en el lumen y entre
los folículos, que pueden estar colapsados, pared gonadal gruesa. Se presentan algunos
fagocitos pequeños. (Escala 100 µm)
Recuperación o reposo (II): Ovarios de paredes gruesas. En algunas secciones del
epitelio germinal pueden observarse ocasionalmente oocitos remanentes y fagocitos de
la previa estación reproductiva. Oocitos basófilos. Gónadas masculinas solo pueden ser
identificadas con histología. Una capa de espermatogonias está presente en el epitelio
germinal. La pared gonadal tiene su máximo grosor. El lumen está vacío. (Escala 84 µm)

Biología Reproductiva de H. (S.) glaberrima
46
Maduro (III): Ovarios maduros llenos de oocitos eosinófilos y pared gonadal delgada. Los
oocitos mantienen su folículo y la vesícula germinal empieza a tomar posición hacia la
protuberancia basal. En los machos, pliegues del epitelio germinal reducidos o ausentes
y el lumen lleno de espermatozoides. Algunos espermatocitos pueden estar presentes a
lo largo del epitelio. La pared gonadal tiene su mínimo espesor. (Escala 84 µm)
Desovado (IV): Ovarios arrugados y encogidos, de pared gruesa. Oocitos residuales
ocasionalmente presentes en el lumen. Fagocitos evidentes. Las gónadas masculinas
con los túbulos encogidos, y generalmente el lumen vacio o con pocos espermatozoides
remanentes. Espermatogonias dispersas a lo largo del epitelio germinal. La pared
gonadal es gruesa y el epitelio una vez más toma su apariencia con volutas. (Escala 320
µm y 185 µm)

Anexos
47
ANEXO 2. Proceso de tinción, método Hematoxilina - Eosina (Gómez-León, 2005)
Xileno (Caliente en la plancha 35 o 36°)…...5 min (dos recambios)
Etanol 100%.………………………………… 5 min (dos recambios)
Etanol 96%.………………………………….. 5 min
Etanol 10%………………………………….. 5 min
Agua destilada………………………………. 5 min
Hematoxilina………………………………… 8 min
Agua destilada………………………………. 2 min
Alcohol ácido………………………………… 5 inmersiones
Agua corriente………………………………. 5 min
Agua destilada………………………………. 3 min
Etanol 96%…………………………………... 3 min
Eosina………………………………………… 2 min
Etanol 96%…………………………………… 25 inmersiones
Etanol 96%…………………………………… 25 inmersiones
Etanol 96%…………………………………… 3 min
Etanol 100%…………………………………. 1 min 30 seg
Etanol 100%…………………………………. 3 min
Xileno………………………………………… 5 min
Xileno………………………………………… 5 min o hasta montar la muestra
El montaje de las muestras se realizó usando el pegante Shandon Consul-Mount a base
de xileno.





























