- 1. chapterFifteen to twenty billion years ago, the universe
aroseas a cataclysmic eruption of hot, energy-rich
sub-atomicparticles. Within seconds, the simplest elements(hydrogen
and helium) were formed. As the universeexpanded and cooled,
material condensed under the in-fluenceof gravity to form stars.
Some stars becameenormous and then exploded as supernovae,
releasingthe energy needed to fuse simpler atomic nuclei into
themore complex elements. Thus were produced, over bil-lionsof
years, the Earth itself and the chemical elementsfound on the Earth
today. About four billion years ago,life arosesimple microorganisms
with the ability to ex-tractenergy from organic compounds or from
sunlight,which they used to make a vast array of more
complexbiomolecules from the simple elements and compoundson the
Earths surface.Biochemistry asks how the remarkable propertiesof
living organisms arise from the thousands of differ-entlifeless
biomolecules. When these molecules are iso-latedand examined
individually, they conform to all thephysical and chemical laws
that describe the behaviorof inanimate matteras do all the
processes occurringin living organisms. The study of biochemistry
showshow the collections of inanimate molecules that
consti-tuteliving organisms interact to maintain and
perpetu-atelife animated solely by the physical and chemicallaws
that govern the nonliving universe.Yet organisms possess
extraordinary attributes,properties that distinguish them from
other collectionsof matter. What are these distinguishing features
of liv-ingorganisms?A high degree of chemical complexity
andmicroscopic organization. Thousands of differ-entmolecules make
up a cells intricate internalstructures (Fig. 11a). Each has its
characteristicsequence of subunits, its unique
three-dimensionalstructure, and its highly specific selection
ofbinding partners in the cell.Systems for extracting,
transforming, andusing energy from the environment (Fig.11b),
enabling organisms to build and maintaintheir intricate structures
and to do mechanical,chemical, osmotic, and electrical work.
Inanimatematter tends, rather, to decay toward a moredisordered
state, to come to equilibrium with itssurroundings.THE
FOUNDATIONSOF BIOCHEMISTRY1.1 Cellular Foundations 31.2 Chemical
Foundations 121.3 Physical Foundations 211.4 Genetic Foundations
281.5 Evolutionary Foundations 31With the cell, biology discovered
its atom . . . Tocharacterize life, it was henceforth essential to
study thecell and analyze its structure: to single out the
commondenominators, necessary for the life of every
cell;alternatively, to identify differences associated with
theperformance of special functions.Franois Jacob, La logique du
vivant: une histoire de lhrdit(The Logic of Life: A History of
Heredity), 1970We must, however, acknowledge, as it seems to me,
thatman with all his noble qualities . . . still bears in hisbodily
frame the indelible stamp of his lowly origin.Charles Darwin, The
Descent of Man, 187111
2. (b)A capacity for precise self-replication andself-assembly
(Fig. 11c). A single bacterial cellplaced in a sterile nutrient
medium can give riseto a billion identical daughter cells in 24
hours.Each cell contains thousands of different molecules,some
extremely complex; yet each bacterium isa faithful copy of the
original, its constructiondirected entirely from information
containedwithin the genetic material of the original
cell.Mechanisms for sensing and responding toalterations in their
surroundings, constantlyadjusting to these changes by adapting
theirinternal chemistry.Defined functions for each of their
compo-nentsand regulated interactions among them.This is true not
only of macroscopic structures,such as leaves and stems or hearts
and lungs, butalso of microscopic intracellular structures and
indi-vidualchemical compounds. The interplay amongthe chemical
components of a living organism is dy-namic;changes in one
component cause coordinat-ingor compensating changes in another,
with thewhole ensemble displaying a character beyond thatof its
individual parts. The collection of moleculescarries out a program,
the end result of which isreproduction of the program and
self-perpetuationof that collection of moleculesin short, life.A
history of evolutionary change. Organismschange their inherited
life strategies to survivein new circumstances. The result of eons
ofevolution is an enormous diversity of life forms,superficially
very different (Fig. 12) butfundamentally related through their
shared ancestry.Despite these common properties, and the
funda-mentalunity of life they reveal, very few
generalizationsabout living organisms are absolutely correct for
everyorganism under every condition; there is enormous
di-versity.The range of habitats in which organisms live,from hot
springs to Arctic tundra, from animal intestinesto college
dormitories, is matched by a correspondinglywide range of specific
biochemical adaptations, achieved2 Chapter 1 The Foundations of
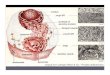
Biochemistry(a)(c)FIGURE 11 Some characteristics of living matter.
(a) Microscopiccomplexity and organization are apparent in this
colorized thin sec-tionof vertebrate muscle tissue, viewed with the
electron microscope.(b) A prairie falcon acquires nutrients by
consuming a smaller bird.(c) Biological reproduction occurs with
near-perfect fidelity.FIGURE 12 Diverse living organisms share
common chemical fea-tures.Birds, beasts, plants, and soil
microorganisms share with hu-mansthe same basic structural units
(cells) and the same kinds ofmacromolecules (DNA, RNA, proteins)
made up of the same kinds ofmonomeric subunits (nucleotides, amino
acids). They utilize the samepathways for synthesis of cellular
components, share the same geneticcode, and derive from the same
evolutionary ancestors. Shown hereis a detail from The Garden of
Eden, by Jan van Kessel the Younger(16261679). 3. within a common
chemical framework. For the sake ofclarity, in this book we
sometimes risk certain general-izations,which, though not perfect,
remain useful; wealso frequently point out the exceptions that
illuminatescientific generalizations.Biochemistry describes in
molecular terms the struc-tures,mechanisms, and chemical processes
shared byall organisms and provides organizing principles
thatunderlie life in all its diverse forms, principles we referto
collectively as the molecular logic of life. Althoughbiochemistry
provides important insights and practicalapplications in medicine,
agriculture, nutrition, andindustry, its ultimate concern is with
the wonder of lifeitself.In this introductory chapter, then, we
describe(briefly!) the cellular, chemical, physical
(thermody-namic),and genetic backgrounds to biochemistry andthe
overarching principle of evolutionthe develop-mentover generations
of the properties of living cells.As you read through the book, you
may find it helpfulto refer back to this chapter at intervals to
refresh yourmemory of this background material.1.1 Cellular
FoundationsThe unity and diversity of organisms become apparenteven
at the cellular level. The smallest organisms consistof single
cells and are microscopic. Larger, multicellularorganisms contain
many different types of cells, whichvary in size, shape, and
specialized function. Despitethese obvious differences, all cells
of the simplest andmost complex organisms share certain
fundamentalproperties, which can be seen at the biochemical
level.Cells Are the Structural and Functional Units of AllLiving
OrganismsCells of all kinds share certain structural features
(Fig.13). The plasma membrane defines the periphery ofthe cell,
separating its contents from the surroundings.It is composed of
lipid and protein molecules that forma thin, tough, pliable,
hydrophobic barrier around thecell. The membrane is a barrier to
the free passage ofinorganic ions and most other charged or polar
com-pounds.Transport proteins in the plasma membrane al-lowthe
passage of certain ions and molecules; receptorproteins transmit
signals into the cell; and membraneenzymes participate in some
reaction pathways. Be-causethe individual lipids and proteins of
the plasmamembrane are not covalently linked, the entire
struc-tureis remarkably flexible, allowing changes in theshape and
size of the cell. As a cell grows, newly madelipid and protein
molecules are inserted into its plasmamembrane; cell division
produces two cells, each with itsown membrane. This growth and cell
division (fission)occurs without loss of membrane integrity.1.1
Cellular Foundations 3Nucleus (eukaryotes)or nucleoid
(bacteria)Contains genetic materialDNA andassociated proteins.
Nucleus ismembrane-bounded.Plasma membraneTough, flexible lipid
bilayer.Selectively permeable topolar substances. Includesmembrane
proteins thatfunction in transport,in signal reception,and as
enzymes.CytoplasmAqueous cell contents andsuspended particlesand
organelles.centrifuge at 150,000 gSupernatant: cytosolConcentrated
solutionof enzymes, RNA,monomeric subunits,metabolites,inorganic
ions.Pellet: particles and organellesRibosomes, storage
granules,mitochondria, chloroplasts, lysosomes,endoplasmic
reticulum.FIGURE 13 The universal features of living cells. All
cells have anucleus or nucleoid, a plasma membrane, and cytoplasm.
The cytosolis defined as that portion of the cytoplasm that remains
in the super-natantafter centrifugation of a cell extract at
150,000 g for 1 hour.The internal volume bounded by the plasma
mem-brane,the cytoplasm (Fig. 13), is composed of anaqueous
solution, the cytosol, and a variety of sus-pendedparticles with
specific functions. The cytosol isa highly concentrated solution
containing enzymes andthe RNA molecules that encode them; the
components(amino acids and nucleotides) from which these
macro-moleculesare assembled; hundreds of small organicmolecules
called metabolites, intermediates in biosyn-theticand degradative
pathways; coenzymes, com-poundsessential to many enzyme-catalyzed
reactions;inorganic ions; and ribosomes, small particles
(com-posedof protein and RNA molecules) that are the sitesof
protein synthesis.All cells have, for at least some part of their
life, ei-thera nucleus or a nucleoid, in which the genome 4. the
complete set of genes, composed of DNAis storedand replicated. The
nucleoid, in bacteria, is not sepa-ratedfrom the cytoplasm by a
membrane; the nucleus,in higher organisms, consists of nuclear
material en-closedwithin a double membrane, the nuclear
envelope.Cells with nuclear envelopes are called eukaryotes(Greek
eu, true, and karyon, nucleus); those with-outnuclear
envelopesbacterial cellsare prokary-otes(Greek pro,
before).Cellular Dimensions Are Limited by Oxygen DiffusionMost
cells are microscopic, invisible to the unaided eye.Animal and
plant cells are typically 5 to 100 m in di-ameter,and many bacteria
are only 1 to 2 m long (seethe inside back cover for information on
units and theirabbreviations). What limits the dimensions of a
cell? Thelower limit is probably set by the minimum number ofeach
type of biomolecule required by the cell. Thesmallest cells,
certain bacteria known as mycoplasmas,are 300 nm in diameter and
have a volume of about1014 mL. A single bacterial ribosome is about
20 nm inits longest dimension, so a few ribosomes take up a
sub-stantialfraction of the volume in a mycoplasmal cell.The upper
limit of cell size is probably set by therate of diffusion of
solute molecules in aqueous systems.For example, a bacterial cell
that depends upon oxygen-consumingreactions for energy production
must obtainmolecular oxygen by diffusion from the surroundingmedium
through its plasma membrane. The cell is sosmall, and the ratio of
its surface area to its volume isso large, that every part of its
cytoplasm is easily reachedby O2 diffusing into the cell. As cell
size increases, how-ever,surface-to-volume ratio decreases, until
metabo-lismconsumes O2 faster than diffusion can supply
it.Metabolism that requires O2 thus becomes impossibleas cell size
increases beyond a certain point, placing atheoretical upper limit
on the size of the cell.There Are Three Distinct Domains of LifeAll
living organisms fall into one of three large groups(kingdoms, or
domains) that define three branches ofevolution from a common
progenitor (Fig. 14). Twolarge groups of prokaryotes can be
distinguished on bio-chemicalgrounds: archaebacteria (Greek arche-,
ori-gin)and eubacteria (again, from Greek eu, true).Eubacteria
inhabit soils, surface waters, and the tissuesof other living or
decaying organisms. Most of the well-studiedbacteria, including
Escherichia coli, are eu-bacteria.The archaebacteria, more recently
discovered,are less well characterized biochemically; most
inhabitextreme environmentssalt lakes, hot springs, highlyacidic
bogs, and the ocean depths. The available evi-dencesuggests that
the archaebacteria and eubacteriadiverged early in evolution and
constitute two separate4 Chapter 1 The Foundations of
BiochemistryEubacteria EukaryotesPurple
bacteriaCyanobacteriaFlavobacteriaThermotogaGram-positivebacteriaExtremehalophilesGreennonsulfurbacteriaMethanogens
Extreme thermophilesMicrosporidiaPlantsFlagellatesFungiAnimals
CiliatesArchaebacteriaFIGURE 14 Phylogeny of the three domains of
life. Phylogenetic relationships are often illustrated by a family
treeof this type. The fewer the branch points between any two
organisms, the closer is their evolutionary relationship. 5.
Heterotrophs(carbon fromorganiccompounds)Examples:Purple
bacteriaGreen bacteriadomains, sometimes called Archaea and
Bacteria. All eu-karyoticorganisms, which make up the third
domain,Eukarya, evolved from the same branch that gave riseto the
Archaea; archaebacteria are therefore moreclosely related to
eukaryotes than to eubacteria.Within the domains of Archaea and
Bacteria are sub-groupsdistinguished by the habitats in which they
live.In aerobic habitats with a plentiful supply of oxygen,some
resident organisms derive energy from the trans-ferof electrons
from fuel molecules to oxygen. Otherenvironments are anaerobic,
virtually devoid of oxy-gen,and microorganisms adapted to these
environmentsobtain energy by transferring electrons to nitrate
(form-ingN2), sulfate (forming H2S), or CO2 (forming CH4).Many
organisms that have evolved in anaerobic envi-ronmentsare obligate
anaerobes: they die when ex-posedto oxygen.We can classify
organisms according to how theyobtain the energy and carbon they
need for synthesiz-ingcellular material (as summarized in Fig. 15).
Thereare two broad categories based on energy sources:
pho-totrophs(Greek trophe-, nourishment) trap and usesunlight, and
chemotrophs derive their energy fromoxidation of a fuel. All
chemotrophs require a source oforganic nutrients; they cannot fix
CO2 into organic com-pounds.The phototrophs can be further divided
intothose that can obtain all needed carbon from CO2
(au-totrophs)and those that require organic
nutrients(heterotrophs). No chemotroph can get its carbon1.1
Cellular Foundations 5atoms exclusively from CO2 (that is, no
chemotrophsare autotrophs), but the chemotrophs may be
furtherclassified according to a different criterion: whether
thefuels they oxidize are inorganic (lithotrophs) or
or-ganic(organotrophs).Most known organisms fall within one of
these fourbroad categoriesautotrophs or heterotrophs among
thephotosynthesizers, lithotrophs or organotrophs amongthe chemical
oxidizers. The prokaryotes have several gen-eralmodes of obtaining
carbon and energy. Escherichiacoli, for example, is a
chemoorganoheterotroph; it re-quiresorganic compounds from its
environment as fueland as a source of carbon. Cyanobacteria are
photo-lithoautotrophs;they use sunlight as an energy sourceand
convert CO2 into biomolecules. We humans, like E.coli, are
chemoorganoheterotrophs.Escherichia coli Is the Most-Studied
Prokaryotic CellBacterial cells share certain common structural
fea-tures,but also show group-specific specializations (Fig.16). E.
coli is a usually harmless inhabitant of the hu-manintestinal
tract. The E. coli cell is about 2 m longand a little less than 1 m
in diameter. It has a protec-tiveouter membrane and an inner plasma
membranethat encloses the cytoplasm and the nucleoid. Betweenthe
inner and outer membranes is a thin but strong layerof polymers
called peptidoglycans, which gives the cellits shape and rigidity.
The plasma membrane and theAutotrophs(carbon
fromCO2)Examples:CyanobacteriaPlantsHeterotrophs(carbon from
organiccompounds)Phototrophs(energy fromlight)Chemotrophs(energy
from chemicalcompounds)All organismsLithotrophs(energy
frominorganiccompounds)Examples:Sulfur bacteriaHydrogen
bacteriaOrganotrophs(energy fromorganiccompounds)Examples:Most
prokaryotesAll nonphototrophiceukaryotesFIGURE 15 Organisms can be
classified according to their sourceof energy (sunlight or
oxidizable chemical compounds) and theirsource of carbon for the
synthesis of cellular material. 6. layers outside it constitute the
cell envelope. In theArchaea, rigidity is conferred by a different
type of poly-mer(pseudopeptidoglycan). The plasma membranes
ofeubacteria consist of a thin bilayer of lipid moleculespenetrated
by proteins. Archaebacterial membraneshave a similar architecture,
although their lipids differstrikingly from those of the
eubacteria.The cytoplasm of E. coli contains about 15,000ribosomes,
thousands of copies each of about 1,000different enzymes, numerous
metabolites and cofac-tors,and a variety of inorganic ions. The
nucleoidcontains a single, circular molecule of DNA, and
thecytoplasm (like that of most bacteria) contains one ormore
smaller, circular segments of DNA called plas-mids.In nature, some
plasmids confer resistance totoxins and antibiotics in the
environment. In the labo-ratory,these DNA segments are especially
amenableto experimental manipulation and are extremely use-fulto
molecular geneticists.Most bacteria (including E. coli) lead
existences asindividual cells, but in some bacterial species cells
tendto associate in clusters or filaments, and a few
(themyxobacteria, for example) demonstrate simple
socialbehavior.Eukaryotic Cells Have a Variety of
MembranousOrganelles, Which Can Be Isolated for StudyTypical
eukaryotic cells (Fig. 17) are much larger thanprokaryotic
cellscommonly 5 to 100 m in diameter,with cell volumes a thousand
to a million times larger thanthose of bacteria. The distinguishing
characteristics ofeukaryotes are the nucleus and a variety of
membrane-boundedorganelles with specific functions:
mitochondria,endoplasmic reticulum, Golgi complexes, and
lysosomes.Plant cells also contain vacuoles and chloroplasts
(Fig.17). Also present in the cytoplasm of many cells aregranules
or droplets containing stored nutrients such asstarch and fat.In a
major advance in biochemistry, Albert Claude,Christian de Duve, and
George Palade developed meth-odsfor separating organelles from the
cytosol and fromeach otheran essential step in isolating
biomoleculesand larger cell components and investigating their6
Chapter 1 The Foundations of BiochemistryRibosomes Bacterial
ribosomes are smaller thaneukaryotic ribosomes, but serve the same
functionprotein synthesis from an RNA message.Nucleoid Contains a
single,simple, long circular DNAmolecule.Pili Providepoints
ofadhesion tosurface ofother cells.FlagellaPropel cellthrough
itssurroundings.Cell envelopeStructure varieswith type
ofbacteria.Outer membranePeptidoglycan layerInner
membraneGram-negative bacteriaOuter membrane;peptidoglycan
layerPeptidoglycan layerInner membraneGram-positive bacteriaNo
outer membrane;thicker peptidoglycan
layerCyanobacteriaGram-negative; tougherpeptidoglycan
layer;extensive internalmembrane system withphotosynthetic
pigmentsArchaebacteriaNo outer membrane;peptidoglycan layer
outsideplasma membraneFIGURE 16 Common structural features of
bacterial cells. Becauseof differences in the cell envelope
structure, some eubacteria (gram-positivebacteria) retain Grams
stain, and others (gram-negativebacteria) do not. E. coli is
gram-negative. Cyanobacteria are alsoeubacteria but are
distinguished by their extensive internal membranesystem, in which
photosynthetic pigments are localized. Although thecell envelopes
of archaebacteria and gram-positive eubacteria looksimilar under
the electron microscope, the structures of the membranelipids and
the polysaccharides of the cell envelope are distinctly
dif-ferentin these organisms. 7. 1.1 Cellular Foundations
7Ribosomes are protein-synthesizingmachinesPeroxisome destroys
peroxidesCytoskeleton supports cell, aidsin movement of
organellsLysosome degrades intracellulardebrisTransport vesicle
shuttles lipidsand proteins between ER, Golgi,and plasma
membraneGolgi complex processes,packages, and targets proteins
toother organelles or for exportSmooth endoplasmic reticulum(SER)
is site of lipid synthesisand drug metabolismNucleus contains
thegenes (chromatin)Ribosomes CytoskeletonGolgicomplexNucleolus is
site of ribosomalRNA synthesisRough endoplasmic reticulum(RER) is
site of much proteinsynthesisMitochondrion oxidizes fuels toproduce
ATPPlasma membrane separates cellfrom environment,
regulatesmovement of materials into andout of cellChloroplast
harvests sunlight,produces ATP and carbohydratesStarch granule
temporarily storescarbohydrate products ofphotosynthesisThylakoids
are site of light-drivenATP synthesisCell wall provides shape
andrigidity; protects cell fromosmotic swellingPlasmodesma provides
path Cell wall of adjacent cellbetween two plant cellsNuclear
envelope segregateschromatin (DNAprotein)from cytoplasmVacuole
degrades and recyclesmacromolecules, storesmetabolites(a) Animal
cellGlyoxysome contains enzymes ofthe glyoxylate cycle(b) Plant
cellFIGURE 17 Eukaryotic cell structure. Schematic illustrations of
thetwo major types of eukaryotic cell: (a) a representative animal
celland (b) a representative plant cell. Plant cells are usually 10
to100 m in diameterlarger than animal cells, which typicallyrange
from 5 to 30 m. Structures labeled in red are unique toeither
animal or plant cells. 8. structures and functions. In a typical
cell fractionation(Fig. 18), cells or tissues in solution are
disrupted bygentle homogenization. This treatment ruptures
theplasma membrane but leaves most of the organelles in-tact.such
as nuclei, mitochondria, and lysosomes differ insize and therefore
sediment at different rates. They alsodiffer in specific gravity,
and they float at differentlevels in a density gradient. The
homogenate is then centrifuged; organelles Centrifugation
Differential centrifugation results in a rough fraction-ationof the
cytoplasmic contents, which may be furtherpurified by isopycnic
(same density) centrifugation. Inthis procedure, organelles of
different buoyant densities(the result of different ratios of lipid
and protein in eachtype of organelle) are separated on a density
gradient. Bycarefully removing material from each region of the
gra-dientand observing it with a microscope, the biochemistcan
establish the sedimentation position of each organelle8 Chapter 1
The Foundations of Biochemistry
FractionationSampleSucrosegradientLess densecomponentMore
densecomponent8 7 6 5 4 3 2 1
Isopycnic(sucrose-density)centrifugation(b) Low-speed
centrifugation(1,000 g, 10 min)Supernatant subjected tomedium-speed
centrifugation(20,000 g, 20 min)Supernatant subjectedto
high-speedcentrifugation(80,000 g, 1 h)Supernatantsubjected tovery
high-speedcentrifugation(150,000 g, 3
h)DifferentialcentrifugationTissuehomogenizationTissuehomogenatePelletcontainsmitochondria,lysosomes,peroxisomesPelletcontainsmicrosomes(fragments
of ER),small vesiclesPellet containsribosomes,
largemacromoleculesPelletcontainswhole
cells,nuclei,cytoskeletons,plasmamembranesSupernatantcontainssolubleproteins(a)FIGURE
18 Subcellular fractionation of tissue. A tissue such as liveris
first mechanically homogenized to break cells and disperse
theircontents in an aqueous buffer. The sucrose medium has an
osmoticpressure similar to that in organelles, thus preventing
diffusion of wa-terinto the organelles, which would swell and
burst. (a) The large andsmall particles in the suspension can be
separated by centrifugationat different speeds, or (b) particles of
different density can be sepa-ratedby isopycnic centrifugation. In
isopycnic centrifugation, a cen-trifugetube is filled with a
solution, the density of which increasesfrom top to bottom; a
solute such as sucrose is dissolved at differentconcentrations to
produce the density gradient. When a mixture oforganelles is
layered on top of the density gradient and the tube iscentrifuged
at high speed, individual organelles sediment until theirbuoyant
density exactly matches that in the gradient. Each layer canbe
collected separately. 9. and obtain purified organelles for further
study. Forexample, these methods were used to establish
thatlysosomes contain degradative enzymes, mitochondriacontain
oxidative enzymes, and chloroplasts containphotosynthetic pigments.
The isolation of an organelle en-richedin a certain enzyme is often
the first step in thepurification of that enzyme.The Cytoplasm Is
Organized by the Cytoskeletonand Is Highly DynamicElectron
microscopy reveals several types of protein fila-mentscrisscrossing
the eukaryotic cell, forming an inter-lockingthree-dimensional
meshwork, the cytoskeleton.There are three general types of
cytoplasmic filamentsactin filaments, microtubules, and
intermediate filaments(Fig. 19)differing in width (from about 6 to
22 nm),composition, and specific function. All types
providestructure and organization to the cytoplasm and shapeto the
cell. Actin filaments and microtubules also help toproduce the
motion of organelles or of the whole cell.Each type of cytoskeletal
component is composedof simple protein subunits that polymerize to
form fila-mentsof uniform thickness. These filaments are not
per-manentstructures; they undergo constant disassembly1.1 Cellular
Foundations 9into their protein subunits and reassembly into
fila-ments.Their locations in cells are not rigidly fixed butmay
change dramatically with mitosis, cytokinesis,amoeboid motion, or
changes in cell shape. The assem-bly,disassembly, and location of
all types of filamentsare regulated by other proteins, which serve
to link orbundle the filaments or to move cytoplasmic
organellesalong the filaments.The picture that emerges from this
brief surveyof cell structure is that of a eukaryotic cell with
ameshwork of structural fibers and a complex system
ofmembrane-bounded compartments (Fig. 17). The
fila-mentsdisassemble and then reassemble elsewhere.
Mem-branousvesicles bud from one organelle and fuse withanother.
Organelles move through the cytoplasm alongprotein filaments, their
motion powered by energy de-pendentmotor proteins. The endomembrane
systemsegregates specific metabolic processes and providessurfaces
on which certain enzyme-catalyzed reactionsoccur. Exocytosis and
endocytosis, mechanisms oftransport (out of and into cells,
respectively) that involvemembrane fusion and fission, provide
paths between thecytoplasm and surrounding medium, allowing for
secre-tionof substances produced within the cell and uptakeof
extracellular materials.Actin stress
fibers(a)Microtubules(b)Intermediate filaments(c)FIGURE 19 The
three types of cytoskeletal filaments. The upper pan-elsshow
epithelial cells photographed after treatment with antibodiesthat
bind to and specifically stain (a) actin filaments bundled
togetherto form stress fibers, (b) microtubules radiating from the
cell center,and (c) intermediate filaments extending throughout the
cytoplasm. Forthese experiments, antibodies that specifically
recognize actin, tubu-lin,or intermediate filament proteins are
covalently attached to afluorescent compound. When the cell is
viewed with a fluorescencemicroscope, only the stained structures
are visible. The lower panelsshow each type of filament as
visualized by (a, b) transmission or(c) scanning electron
microscopy. 10. Although complex, this organization of the
cyto-plasmis far from random. The motion and the position-ingof
organelles and cytoskeletal elements are undertight regulation, and
at certain stages in a eukaryoticcells life, dramatic, finely
orchestrated reorganizations,such as the events of mitosis, occur.
The interactions be-tweenthe cytoskeleton and organelles are
noncovalent,reversible, and subject to regulation in response to
var-iousintracellular and extracellular signals.Cells Build
Supramolecular StructuresMacromolecules and their monomeric
subunits differgreatly in size (Fig. 110). A molecule of alanine is
lessthan 0.5 nm long. Hemoglobin, the oxygen-carrying pro-teinof
erythrocytes (red blood cells), consists of nearly600 amino acid
subunits in four long chains, folded intoglobular shapes and
associated in a structure 5.5 nm indiameter. In turn, proteins are
much smaller than ribo-somes(about 20 nm in diameter), which are in
turnmuch smaller than organelles such as mitochondria,
typ-ically1,000 nm in diameter. It is a long jump from
sim-plebiomolecules to cellular structures that can be seen10
Chapter 1 The Foundations of Biochemistry(a) Some of the amino
acids of proteinsCOOOC AOHOOH OC AOH OC ACHHNOOHCHNUracil
ThymineNH2CN CHNAdenine GuanineOHOCH2HNH
HOHNH2OHOHH2CHCytosineHOHHONHOCH2 HH H-D-Ribose 2-Deoxy-
-D-riboseHOHOHOHCOOCH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2CH2
CH2CH2 CH3CH2OleateCH2PalmitateCH2OHCHOHCH2OHCH3 CH2CH2OHCH3CH2OHH
HHHOHOOHHOHOH-D-Glucose(b) The components of nucleic acids (c) Some
components of lipids(d) The parent sugarOHO POOHPhosphoric
acidNCholineCH3GlycerolCH2CH3CH2CH2COOCH2CH2CH2CHCHCCCHHCN
NHOCCCHCN NH NCOCHCNHOCHCNHCOCHCNHH2NCH3Nitrogenous
basesFive-carbon sugarsH3NH3NH3NH3NOC ACOO
COOCOOH3NCOOH3NCOOCOOACH3ACH2OHACAH2Alanine SerineAspartateOC
AACASHCysteineHistidineC AH2OHTyrosineOC AACAH2C HCHHCNNHFIGURE 110
The organic compounds from which most cellularmaterials are
constructed: the ABCs of biochemistry. Shown here are(a) six of the
20 amino acids from which all proteins are built (theside chains
are shaded pink); (b) the five nitrogenous bases, two
five-carbonsugars, and phosphoric acid from which all nucleic acids
arebuilt; (c) five components of membrane lipids; and (d)
D-glucose, theparent sugar from which most carbohydrates are
derived. Note thatphosphoric acid is a component of both nucleic
acids and membranelipids. 11. Chromosomewith the light microscope.
Figure 111 illustrates thestructural hierarchy in cellular
organization.The monomeric subunits in proteins, nucleic acids,and
polysaccharides are joined by covalent bonds. Insupramolecular
complexes, however, macromoleculesare held together by noncovalent
interactionsmuchweaker, individually, than covalent bonds. Among
thesenoncovalent interactions are hydrogen bonds (betweenpolar
groups), ionic interactions (between chargedgroups), hydrophobic
interactions (among nonpolargroups in aqueous solution), and van
der Waals inter-actionsall of which have energies substantially
smallerthan those of covalent bonds (Table 11). The natureof these
noncovalent interactions is described in Chap-ter2. The large
numbers of weak interactions betweenmacromolecules in
supramolecular complexes stabilizethese assemblies, producing their
unique structures.In Vitro Studies May Overlook Important
Interactionsamong MoleculesOne approach to understanding a
biological process isto study purified molecules in vitro (in
glassin thetest tube), without interference from other
moleculespresent in the intact cellthat is, in vivo (in the
liv-ing).Although this approach has been remarkably re-vealing,we
must keep in mind that the inside of a cellis quite different from
the inside of a test tube. The in-terferingcomponents eliminated by
purification maybe critical to the biological function or
regulation of themolecule purified. For example, in vitro studies
of pure1.1 Cellular Foundations 11Level 4:The celland its
organellesLevel 3:SupramolecularcomplexesLevel
2:MacromoleculesLevel 1:Monomeric unitsNucleotidesDNA OO P O OO
HAmino acidsProteinCellulosePlasma membraneCell
wallSugarsOCH2NH2NNHH HOH HHH3N C
COOCH3OHHOHCH2OHHHOOHOHHCH2OHOHFIGURE 111 Structural hierarchy in
the molecular organization ofcells. In this plant cell, the nucleus
is an organelle containing severaltypes of supramolecular
complexes, including chromosomes. Chro-mosomesconsist of
macromolecules of DNA and many different pro-teins.Each type of
macromolecule is made up of simple subunitsDNA of nucleotides
(deoxyribonucleotides), for example.enzymes are commonly done at
very low enzyme con-centrationsin thoroughly stirred aqueous
solutions. Inthe cell, an enzyme is dissolved or suspended in a
gel-likecytosol with thousands of other proteins, some ofwhich bind
to that enzyme and influence its activity.TABLE 11 Strengths of
Bonds Commonin BiomoleculesBond Bonddissociation dissociationType
energy* Type energyof bond (kJ/mol) of bond (kJ/mol)Single bonds
Double bondsOOH 470 CPO 712HOH 435 CPN 615POO 419 CPC 611COH 414
PPO 502NOH 389COO 352 Triple bondsCOC 348 CmC 816SOH 339 NmN 930CON
293COS 260NOO 222SOS 214*The greater the energy required for bond
dissociation (breakage), the stronger the bond. 12. Some enzymes
are parts of multienzyme complexes inwhich reactants are channeled
from one enzyme to an-otherwithout ever entering the bulk solvent.
Diffusionis hindered in the gel-like cytosol, and the cytosolic
com-positionvaries in different regions of the cell. In short,a
given molecule may function quite differently in thecell than in
vitro. A central challenge of biochemistry isto understand the
influences of cellular organization andmacromolecular associations
on the function of individ-ualenzymes and other biomoleculesto
understandfunction in vivo as well as in vitro.SUMMARY 1.1 Cellular
Foundations All cells are bounded by a plasma membrane;have a
cytosol containing metabolites,coenzymes, inorganic ions, and
enzymes; andhave a set of genes contained within a
nucleoid(prokaryotes) or nucleus (eukaryotes). Phototrophs use
sunlight to do work;chemotrophs oxidize fuels, passing electrons
togood electron acceptors: inorganic compounds,organic compounds,
or molecular oxygen. Bacterial cells contain cytosol, a nucleoid,
andplasmids. Eukaryotic cells have a nucleus andare
multicompartmented, segregating certainprocesses in specific
organelles, which can beseparated and studied in isolation.
Cytoskeletal proteins assemble into longfilaments that give cells
shape and rigidity andserve as rails along which cellular
organellesmove throughout the cell. Supramolecular complexes are
held together bynoncovalent interactions and form a hierarchyof
structures, some visible with the lightmicroscope. When individual
molecules areremoved from these complexes to be studiedin vitro,
interactions important in the livingcell may be lost.1.2 Chemical
FoundationsBiochemistry aims to explain biological form and
func-tionin chemical terms. As we noted earlier, one of themost
fruitful approaches to understanding biologicalphenomena has been
to purify an individual chemicalcomponent, such as a protein, from
a living organismand to characterize its structural and chemical
charac-teristics.By the late eighteenth century, chemists
hadconcluded that the composition of living matter is
strik-inglydifferent from that of the inanimate world.
AntoineLavoisier (17431794) noted the relative chemical
sim-plicityof the mineral world and contrasted it with
thecomplexity of the plant and animal worlds; the latter,he knew,
were composed of compounds rich in the ele-mentscarbon, oxygen,
nitrogen, and phosphorus.During the first half of the twentieth
century, par-allelbiochemical investigations of glucose breakdown
inyeast and in animal muscle cells revealed remarkablechemical
similarities in these two apparently very dif-ferentcell types; the
breakdown of glucose in yeast andmuscle cells involved the same ten
chemical intermedi-ates.Subsequent studies of many other
biochemicalprocesses in many different organisms have confirmedthe
generality of this observation, neatly summarized byJacques Monod:
What is true of E. coli is true of theelephant. The current
understanding that all organismsshare a common evolutionary origin
is based in part onthis observed universality of chemical
intermediates andtransformations.Only about 30 of the more than 90
naturally occur-ringchemical elements are essential to organisms.
Mostof the elements in living matter have relatively lowatomic
numbers; only five have atomic numbers abovethat of selenium, 34
(Fig. 112). The four most abun-dantelements in living organisms, in
terms of percent-ageof total number of atoms, are hydrogen,
oxygen,nitrogen, and carbon, which together make up morethan 99% of
the mass of most cells. They are the light-estelements capable of
forming one, two, three, and fourbonds, respectively; in general,
the lightest elements12 Chapter 1 The Foundations of Biochemistry1
2H HeBulk elementsTrace elements3 4 5 6 7 8 9 10Li Be B C N O F
Ne11 12 13 14 15 16 17 18Na Mg Al Si P S Cl Ar19 20 21 22 23 24 25
26 27 28 29 30 31 32 33 34 35 36K Ca Sc Ti V Cr Mn Fe Co Ni Cu Zn
Ga Ge As Se Br Kr37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53
54Rb Sr Y Zr Nb Mo Tc Ru Rh Pd Ag Cd In Sn Sb Te I Xe55 56 72 73 74
75 76 77 78 79 80 81 82 83 84 85 86Cs Ba Hf Ta W Re Os Ir Pt Au Hg
Tl Pb Bi Po At Rn87 88Fr RaLanthanidesActinidesFIGURE 112 Elements
essential to animallife and health. Bulk elements (shadedorange)
are structural components of cellsand tissues and are required in
the diet ingram quantities daily. For trace elements(shaded bright
yellow), the requirements aremuch smaller: for humans, a few
milligramsper day of Fe, Cu, and Zn, even less of theothers. The
elemental requirements forplants and microorganisms are similar
tothose shown here; the ways in which theyacquire these elements
vary. 13. form the strongest bonds. The trace elements (Fig.
112)represent a miniscule fraction of the weight of the hu-manbody,
but all are essential to life, usually becausethey are essential to
the function of specific proteins,including enzymes. The
oxygen-transporting capacityof the hemoglobin molecule, for
example, is absolutelydependent on four iron ions that make up only
0.3% ofits mass.Biomolecules Are Compounds of Carbon witha Variety
of Functional GroupsThe chemistry of living organisms is organized
aroundcarbon, which accounts for more than half the dryweight of
cells. Carbon can form single bonds with hy-drogenatoms, and both
single and double bonds withoxygen and nitrogen atoms (Fig. 113).
Of greatest sig-nificancein biology is the ability of carbon atoms
to formvery stable carboncarbon single bonds. Each carbonatom can
form single bonds with up to four other car-bonatoms. Two carbon
atoms also can share two (orthree) electron pairs, thus forming
double (or triple)bonds.The four single bonds that can be formed by
a car-bonatom are arranged tetrahedrally, with an angle of1.2
Chemical Foundations 13(a) (b)109.5C109.5(c)CC120XCCABYabout 109.5
between any two bonds (Fig. 114) and anaverage length of 0.154 nm.
There is free rotationaround each single bond, unless very large or
highlycharged groups are attached to both carbon atoms, inwhich
case rotation may be restricted. A double bondis shorter (about
0.134 nm) and rigid and allows littlerotation about its
axis.Covalently linked carbon atoms in biomolecules canform linear
chains, branched chains, and cyclic struc-tures.To these carbon
skeletons are added groups ofother atoms, called functional groups,
which conferspecific chemical properties on the molecule. It
seemslikely that the bonding versatility of carbon was a
ma-jorfactor in the selection of carbon compounds for themolecular
machinery of cells during the origin and evo-lutionof living
organisms. No other chemical elementcan form molecules of such
widely different sizes andshapes or with such a variety of
functional groups.Most biomolecules can be regarded as
derivativesof hydrocarbons, with hydrogen atoms replaced by a
va-rietyof functional groups to yield different families oforganic
compounds. Typical of these are alcohols, whichhave one or more
hydroxyl groups; amines, with aminogroups; aldehydes and ketones,
with carbonyl groups;and carboxylic acids, with carboxyl groups
(Fig. 115).Many biomolecules are polyfunctional, containing twoor
more different kinds of functional groups (Fig. 116),each with its
own chemical characteristics and reac-tions.The chemical
personality of a compound is de-terminedby the chemistry of its
functional groups andtheir disposition in three-dimensional space.H
C H HCOCCOOC O C ON C NNC N NC CC C C C C C C CCCCCCCCOCCCNCC
CCCFIGURE 113 Versatility of carbon bonding. Carbon can form
cova-lentsingle, double, and triple bonds (in red), particularly
with othercarbon atoms. Triple bonds are rare in
biomolecules.FIGURE 114 Geometry of carbon bonding. (a) Carbon
atoms havea characteristic tetrahedral arrangement of their four
single bonds.(b) Carboncarbon single bonds have freedom of
rotation, as shownfor the compound ethane (CH3OCH3). (c) Double
bonds are shorterand do not allow free rotation. The two doubly
bonded carbons andthe atoms designated A, B, X, and Y all lie in
the same rigid plane. 14. Carbonyl(aldehyde)HHR
COCarbonyl(ketone)HCHHCHC1 R2R COCarboxyl R COEther R1 O R2FIGURE
115 Some common functional O O Rgroups of biomolecules. In this
figureand throughout the book, we use R torepresent any
substituent. It may be assimple as a hydrogen atom, but typicallyit
is a carbon-containing moiety. Whentwo or more substituents are
shown in amolecule, we designate them R1, R2, andso forth.Cells
Contain a Universal Set of Small MoleculesDissolved in the aqueous
phase (cytosol) of all cells isa collection of 100 to 200 different
small organic mole-cules(Mr ~100 to ~500), the central metabolites
in themajor pathways occurring in nearly every cellthemetabolites
and pathways that have been conservedthroughout the course of
evolution. (See Box 11 for anexplanation of the various ways of
referring to molecu-larC R2NC CHCHO P R2Mixed anhydride R C
OPPweight.) This collection of molecules includes thecommon amino
acids, nucleotides, sugars and theirphosphorylated derivatives, and
a number of mono-,di-, and tricarboxylic acids. The molecules are
polar orcharged, water soluble, and present in micromolar
tomillimolar concentrations. They are trapped within thecell
because the plasma membrane is impermeable tothemalthough specific
membrane transporters cancatalyze the movement of some molecules
into and out14 Chapter 1 The Foundations of BiochemistryHHydroxyl R
O H(alcohol)OO OOMethyl R CHHEthyl R CHCHHEster R1 COO R2Sulfhydryl
R S HDisulfide R1 S S R2Phosphoryl R O POOHThioester R1 COS
R2Anhydride R1 CO O(two car-boxylicacids)OImidazole RNHNHGuanidino
R NCNHHHAmino R NHHAmido R CONHHPhenyl R C CHCH(carboxylic acid
andOphosphoric acid;also called acyl phosphate)OHPhosphoanhydride
R1OOOO 15. P amidoCH3O C O CH2OO A OOOCH2OCH2ONHOCBOof the cell or
between compartments in eukaryotic cells.The universal occurrence
of the same set of compoundsin living cells is a manifestation of
the universality ofmetabolic design, reflecting the evolutionary
conserva-tionof metabolic pathways that developed in the
earli-estcells.There are other small biomolecules, specific to
cer-taintypes of cells or organisms. For example, vascularplants
contain, in addition to the universal set, smallmolecules called
secondary metabolites, which playa role specific to plant life.
These metabolites includecompounds that give plants their
characteristic scents,and compounds such as morphine, quinine,
nicotine,and caffeine that are valued for their physiological
ef-fectson humans but used for other purposes by plants.The entire
collection of small molecules in a given cellhas been called that
cells metabolome, in parallel withthe term genome (defined earlier
and expanded on in1.2 Chemical Foundations
15CH3OOOPOOOCH2NH2NEimidazolePONSection 1.4). If we knew the
composition of a cellsmetabolome, we could predict which enzymes
and meta-bolicpathways were active in that cell.Macromolecules Are
the Major Constituents of CellsMany biological molecules are
macromolecules, poly-mersof high molecular weight assembled from
rela-tivelysimple precursors. Proteins, nucleic acids,
andpolysaccharides are produced by the polymerization ofrelatively
small compounds with molecular weights of500 or less. The number of
polymerized units can rangefrom tens to millions. Synthesis of
macromolecules isa major energy-consuming activity of cells.
Macromol-eculesthemselves may be further assembled
intosupramolecular complexes, forming functional unitssuch as
ribosomes. Table 12 shows the major classesof biomolecules in the
bacterium E. coli.SOCH2OCH2ONHOCBOBOOC A HAOHO C AACH3BOABOOHNKC H
B CANAOOPOHaminophosphoanhydrideAcetyl-coenzyme AHNCHCC
HOOHmethylhydroxylmethylthioester amidophosphorylC CCHCHAAOO
OFIGURE 116 Several common functional groupsin a single
biomolecule. Acetyl-coenzyme A (oftenabbreviated as acetyl-CoA) is
a carrier of acetylgroups in some enzymatic reactions.BOX 11
WORKING IN BIOCHEMISTRYMolecular Weight, Molecular Mass, and
TheirCorrect UnitsThere are two common (and equivalent) ways to
de-scribemolecular mass; both are used in this text. Thefirst is
molecular weight, or relative molecular mass,denoted Mr. The
molecular weight of a substance is de-finedas the ratio of the mass
of a molecule of that sub-stanceto one-twelfth the mass of
carbon-12 (12C).Since Mr is a ratio, it is dimensionlessit has no
asso-ciatedunits. The second is molecular mass, denotedm. This is
simply the mass of one molecule, or the mo-larmass divided by
Avogadros number. The molecu-larmass, m, is expressed in daltons
(abbreviated Da).One dalton is equivalent to one-twelfth the mass
ofcarbon-12; a kilodalton (kDa) is 1,000 daltons; a
mega-dalton(MDa) is 1 million daltons.Consider, for example, a
molecule with a mass1,000 times that of water. We can say of this
moleculeeither Mr18,000 or m18,000 daltons. We can alsodescribe it
as an 18 kDa molecule. However, the ex-pressionMr18,000 daltons is
incorrect.Another convenient unit for describing the massof a
single atom or molecule is the atomic mass unit(formerly amu, now
commonly denoted u). Oneatomic mass unit (1 u) is defined as
one-twelfth themass of an atom of carbon-12. Since the
experimen-tallymeasured mass of an atom of carbon-12 is1.99261023
g, 1 u1.66061024 g. The atomicmass unit is convenient for
describing the mass of apeak observed by mass spectrometry (see Box
32). 16. Molecular Components of anTABLE 12E. coli CellProteins,
long polymers of amino acids, constitutethe largest fraction
(besides water) of cells. Some pro-teinshave catalytic activity and
function as enzymes;others serve as structural elements, signal
receptors, ortransporters that carry specific substances into or
outof cells. Proteins are perhaps the most versatile of
allbiomolecules. The nucleic acids, DNA and RNA, arepolymers of
nucleotides. They store and transmit geneticinformation, and some
RNA molecules have structural andcatalytic roles in supramolecular
complexes. The poly-saccharides,polymers of simple sugars such as
glucose,have two major functions: as energy-yielding fuel storesand
as extracellular structural elements with specificbinding sites for
particular proteins. Shorter polymers ofsugars (oligosaccharides)
attached to proteins or lipidsat the cell surface serve as specific
cellular signals. Thelipids, greasy or oily hydrocarbon
derivatives, serve asstructural components of membranes,
energy-rich fuelstores, pigments, and intracellular signals. In
proteins,nucleotides, polysaccharides, and lipids, the number
ofmonomeric subunits is very large: molecular weights inthe range
of 5,000 to more than 1 million for proteins,up to several billion
for nucleic acids, and in the millionsfor polysaccharides such as
starch. Individual lipid mol-eculesare much smaller (Mr 750 to
1,500) and arenot classified as macromolecules. However, large
num-bersof lipid molecules can associate noncovalently intovery
large structures. Cellular membranes are built ofenormous
noncovalent aggregates of lipid and proteinmolecules.Proteins and
nucleic acids are informationalmacromolecules: each protein and
each nucleic acidhas a characteristic information-rich subunit
sequence.Some oligosaccharides, with six or more different
sug-arsconnected in branched chains, also carry informa-tion;on the
outer surface of cells they serve as highlyspecific points of
recognition in many cellular processes(as described in Chapter
7).Three-Dimensional Structure Is Describedby Configuration and
ConformationThe covalent bonds and functional groups of a
biomol-eculeare, of course, central to its function, but so alsois
the arrangement of the molecules constituent atomsin
three-dimensional spaceits stereochemistry. Acarbon-containing
compound commonly exists asstereoisomers, molecules with the same
chemicalbonds but different stereochemistrythat is,
differentconfiguration, the fixed spatial arrangement of
atoms.Interactions between biomolecules are invariably
stereo-specific,requiring specific stereochemistry in the
in-teractingmolecules.Figure 117 shows three ways to illustrate the
stereo-chemicalstructures of simple molecules. The
perspec-tivediagram specifies stereochemistry unambiguously,but
bond angles and center-to-center bond lengths arebetter represented
with ball-and-stick models. In space-16 Chapter 1 The Foundations
of BiochemistryApproximatenumber ofPercentage of differenttotal
weight molecularof cell speciesWater 70 1Proteins 15 3,000Nucleic
acidsDNA 1 1RNA 6 3,000Polysaccharides 3 5Lipids 2 20Monomeric
subunitsand intermediates 2 500Inorganic ions 1
20MCOH2N#CDOHHOCAH!HOH(a)(c)(b)FIGURE 117 Representations of
molecules. Three ways to representthe structure of the amino acid
alanine. (a) Structural formula in per-spectiveform: a solid wedge
(!) represents a bond in which the atomat the wide end projects out
of the plane of the paper, toward thereader; a dashed wedge (^)
represents a bond extending behind theplane of the paper. (b)
Ball-and-stick model, showing relative bondlengths and the bond
angles. (c) Space-filling model, in which eachatom is shown with
its correct relative van der Waals radius. 17.
HGDHDGCOOHGHDGCOOHfilling models, the radius of each atom is
proportionalto its van der Waals radius, and the contours of
themodel define the space occupied by the molecule (thevolume of
space from which atoms of other moleculesare
excluded).Configuration is conferred by the presence of either(1)
double bonds, around which there is no freedom ofrotation, or (2)
chiral centers, around which substituentgroups are arranged in a
specific sequence. The identi-fyingcharacteristic of
configurational isomers is thatthey cannot be interconverted
without temporarilybreaking one or more covalent bonds. Figure
118ashows the configurations of maleic acid and its isomer,fumaric
acid. These compounds are geometric, or cis-trans,isomers; they
differ in the arrangement of theirsubstituent groups with respect
to the nonrotating dou-blebond (Latin cis, on this sidegroups on
the sameside of the double bond; trans, acrossgroups on
op-positesides). Maleic acid is the cis isomer and fumaricacid the
trans isomer; each is a well-defined compoundthat can be separated
from the other, and each has itsown unique chemical properties. A
binding site (on anenzyme, for example) that is complementary to
one ofthese molecules would not be a suitable binding site forthe
other, which explains why the two compounds havedistinct biological
roles despite their similar chemistry.1.2 Chemical Foundations
1711In the second type of configurational isomer, fourdifferent
substituents bonded to a tetrahedral carbonatom may be arranged two
different ways in spacethatis, have two configurations (Fig.
119)yielding twostereoisomers with similar or identical chemical
proper-tiesbut differing in certain physical and biological
prop-erties.A carbon atom with four different substituentsis said
to be asymmetric, and asymmetric carbons arecalled chiral centers
(Greek chiros, hand; somestereoisomers are related structurally as
the right handis to the left). A molecule with only one chiral
carboncan have two stereoisomers; when two or more (n)
chi-ralcarbons are present, there can be 2n stereoisomers.Some
stereoisomers are mirror images of each other;they are called
enantiomers (Fig. 119). Pairs ofstereoisomers that are not mirror
images of each otherare called diastereomers (Fig. 120).As Louis
Pasteur first observed (Box 12), enan-tiomershave nearly identical
chemical properties butdiffer in a characteristic physical
property, their inter-actionwith plane-polarized light. In separate
solutions,two enantiomers rotate the plane of plane-polarizedlight
in opposite directions, but an equimolar solutionof the two
enantiomers (a racemic mixture) shows nooptical rotation. Compounds
without chiral centers donot rotate the plane of plane-polarized
light.CHOOCPCMaleic acid
(cis)CH3OJC11-cis-RetinallightCHOOCDHPCFumaric acid
(trans)All-trans-RetinalH3CH3GCH910 12H3CH3C9121011CH3CJOGHCH3 CH3
GD(b)(a)CH3 CH3 GDFIGURE 118 Configurations of geometric isomers.
(a) Isomers suchas maleic acid and fumaric acid cannot be
interconverted withoutbreaking covalent bonds, which requires the
input of much energy.(b) In the vertebrate retina, the initial
event in light detection is theabsorption of visible light by
11-cis-retinal. The energy of the absorbedlight (about 250 kJ/mol)
converts 11-cis-retinal to all-trans-retinal,triggering electrical
changes in the retinal cell that lead to a nerveimpulse. (Note that
the hydrogen atoms are omitted from the ball-and-stickmodels for
the retinals.) 18. Given the importance of stereochemistry in
reac-tionsbetween biomolecules (see below), biochemistsmust name
and represent the structure of each bio-moleculeso that its
stereochemistry is unambiguous.For compounds with more than one
chiral center, themost useful system of nomenclature is the RS
system.In this system, each group attached to a chiral carbonis
assigned a priority. The priorities of some commonsubstituents
areOOCH2OOHONH2OCOOHOCHOOCH2OHOCH3OHFor naming in the RS system,
the chiral atom is viewedwith the group of lowest priority (4 in
the diagram onthe next page) pointing away from the viewer. If the
pri-orityof the other three groups (1 to 3) decreases inclockwise
order, the configuration is (R) (Latin rectus,right); if in
counterclockwise order, the configuration18 Chapter 1 The
Foundations of BiochemistryYYCB(a)Mirrorimage
oforiginalmoleculeChiralmolecule:Rotatedmoleculecannot
besuperimposedon its mirrorimageOriginalmoleculeAACX
BYACBXXXACXBACX BX(b)Achiralmolecule:Rotatedmoleculecan
besuperimposedon its mirrorimageMirrorimage
oforiginalmoleculeOriginalmoleculeXACXBFIGURE 119 Molecular
asymmetry: chiral and achiral molecules.(a) When a carbon atom has
four different substituent groups (A, B,X, Y), they can be arranged
in two ways that represent nonsuperim-posablemirror images of each
other (enantiomers). This asymmetriccarbon atom is called a chiral
atom or chiral center. (b) When a tetra-hedralcarbon has only three
dissimilar groups (i.e., the same groupoccurs twice), only one
configuration is possible and the molecule issymmetric, or achiral.
In this case the molecule is superimposable onits mirror image: the
molecule on the left can be rotated counter-clockwise(when looking
down the vertical bond from A to C) to cre-atethe molecule in the
mirror.CH3CH3Diastereomers (nonmirror images)CH3CCH3HC
HXYEnantiomers (mirror images) Enantiomers (mirror images)CCH3XC
YHHCCH3HC Y YXHCH3CCH3HCXHFIGURE 120 Two types of stereoisomers.
There are four different2,3-disubstituted butanes (n2 asymmetric
carbons, hence 2n4stereoisomers). Each is shown in a box as a
perspective formula anda ball-and-stick model, which has been
rotated to allow the reader toview all the groups. Some pairs of
stereoisomers are mirror images ofeach other, or enantiomers. Other
pairs are not mirror images; theseare diastereomers. 19. BOX 12
WORKING IN BIOCHEMISTRYis (S) (Latin sinister, left). In this way
each chiral car-bonis designated either (R) or (S), and the
inclusionof these designations in the name of the compound
pro-videsan unambiguous description of the stereochem-istryat each
chiral center.142 3Counterclockwise(S)143 2Clockwise(R)Another
naming system for stereoisomers, the D and Lsystem, is described in
Chapter 3. A molecule with a sin-glechiral center (glyceraldehydes,
for example) can benamed unambiguously by either system.4COOHH OHHC
3OHH(2R,3R)-Tartaric acid (2S,3S)-Tartaric acid(dextrorotatory)
(levorotatory)CHO1.2 Chemical Foundations 19C
3(2)2CHOOC1OH2CHOOC1HO4COHOHHO C H H OHCH2OHCHO(4)
(1)CH2OH(3)L-Glyceraldehyde (S)-GlyceraldehydeDistinct from
configuration is molecular confor-mation,the spatial arrangement of
substituent groupsthat, without breaking any bonds, are free to
assumedifferent positions in space because of the freedom
ofrotation about single bonds. In the simple hydrocarbonethane, for
example, there is nearly complete freedomof rotation around the COC
bond. Many different, in-terconvertibleconformations of ethane are
possible,depending on the degree of rotation (Fig. 121).
Twoconformations are of special interest: the staggered,which is
more stable than all others and thus predomi-nates,and the
eclipsed, which is least stable. We cannotisolate either of these
conformational forms, becauseLouis Pasteur and Optical Activity:In
Vino, VeritasLouis Pasteur encountered the phenome-nonof optical
activity in 1843, during hisinvestigation of the crystalline
sedimentthat accumulated in wine casks (a form oftartaric acid
called paratartaric acidalsocalled racemic acid, from Latin
racemus,bunch of grapes). He used fine forcepsto separate two types
of crystals identicalin shape but mirror images of each other.Both
types proved to have all the chemi-calproperties of tartaric acid,
but in solu-tionone type rotated polarized light to theleft
(levorotatory), the other to the right(dextrorotatory). Pasteur
later described the experi-mentand its interpretation:In isomeric
bodies, the elements and the propor-tionsin which they are combined
are the same, onlythe arrangement of the atoms is different . . .
Weknow, on the one hand, that the molecular arrange-mentsof the two
tartaric acids are asymmetric, and,on the other hand, that these
arrangements are ab-solutelyidentical, excepting that they exhibit
asym-metryin opposite directions. Are the atoms of thedextro acid
grouped in the form of a right-handedspiral, or are they placed at
the apex of an irregu-lartetrahedron, or are they disposed
according tothis or that asymmetric arrangement? We do notknow.*Now
we do know. X-ray crystallo-graphicstudies in 1951 confirmed that
thelevorotatory and dextrorotatory forms oftartaric acid are mirror
images of eachother at the molecular level and establishedthe
absolute configuration of each (Fig. 1).The same approach has been
used todemonstrate that although the amino acidalanine has two
stereoisomeric forms (des-ignatedD and L), alanine in proteins
existsexclusively in one form (the L isomer; seeChapter 3).FIGURE 1
Pasteur separated crystals of two stereoisomers of tartaricacid and
showed that solutions of the separated forms rotated
po-larizedlight to the same extent but in opposite directions.
Thesedextrorotatory and levorotatory forms were later shown to be
the(R,R) and (S,S) isomers represented here. The RS system of
nomen-clatureis explained in the text.Louis Pasteur18221895*From
Pasteurs lecture to the Socit Chimique de Paris in 1883,quoted in
DuBos, R. (1976) Louis Pasteur: Free Lance of Science,p. 95,
Charles Scribners Sons, New York. 20. mol)kJ/(12energy 84Potential
12.1kJ/molthey are freely interconvertible. However, when one
ormore of the hydrogen atoms on each carbon is replacedby a
functional group that is either very large or elec-tricallycharged,
freedom of rotation around the COCbond is hindered. This limits the
number of stable con-formationsof the ethane
derivative.Interactions between BiomoleculesAre
StereospecificBiological interactions between molecules are
stereo-specific:the fit in such interactions must be
stereo-chemicallycorrect. The three-dimensional structure
ofbiomolecules large and smallthe combination of con-figurationand
conformationis of the utmost impor-tancein their biological
interactions: reactant withenzyme, hormone with its receptor on a
cell surface,antigen with its specific antibody, for example
(Fig.122). The study of biomolecular stereochemistry withprecise
physical methods is an important part of mod-ernresearch on cell
structure and biochemical function.In living organisms, chiral
molecules are usuallypresent in only one of their chiral forms. For
example,the amino acids in proteins occur only as their L
iso-mers;glucose occurs only as its D isomer. (The con-ventionsfor
naming stereoisomers of the amino acidsare described in Chapter 3;
those for sugars, in Chap-ter7; the RS system, described above, is
the mostuseful for some biomolecules.) In contrast, when a
com-poundwith an asymmetric carbon atom is chemicallysynthesized in
the laboratory, the reaction usually pro-ducesall possible chiral
forms: a mixture of the D and Lforms, for example. Living cells
produce only one chiralform of biomolecules because the enzymes
that syn-thesizethem are also chiral.Stereospecificity, the ability
to distinguish betweenstereoisomers, is a property of enzymes and
other pro-teinsand a characteristic feature of the molecular
logicof living cells. If the binding site on a protein is
com-plementaryto one isomer of a chiral compound, it willnot be
complementary to the other isomer, for the samereason that a left
glove does not fit a right hand. Twostriking examples of the
ability of biological systems todistinguish stereoisomers are shown
in Figure 123.SUMMARY 1.2 Chemical Foundations Because of its
bonding versatility, carbon canproduce a broad array of
carboncarbonskeletons with a variety of functional groups;these
groups give biomolecules their biologicaland chemical
personalities. A nearly universal set of several hundred
smallmolecules is found in living cells; the interconver-sionsof
these molecules in the central metabolicpathways have been
conserved in evolution. Proteins and nucleic acids are linear
polymersof simple monomeric subunits; their sequencescontain the
information that gives eachmolecule its three-dimensional structure
andits biological functions.20 Chapter 1 The Foundations of
Biochemistry0 60 120 180 240 300 3600Torsion angle (degrees)FIGURE
121 Conformations. Many conformations of ethane are
pos-siblebecause of freedom of rotation around the COC bond. In
theball-and-stick model, when the front carbon atom (as viewed by
thereader) with its three attached hydrogens is rotated relative to
the rearcarbon atom, the potential energy of the molecule rises to
a maximumin the fully eclipsed conformation (torsion angle 0, 120,
etc.), thenfalls to a minimum in the fully staggered conformation
(torsion angle60, 180, etc.). Because the energy differences are
small enough toallow rapid interconversion of the two forms
(millions of times per sec-ond),the eclipsed and staggered forms
cannot be separately isolated.FIGURE 122 Complementary fit between
a macromolecule and asmall molecule. A segment of RNA from the
regulatory region TAR ofthe human immunodeficiency virus genome
(gray) with a bound argin-inamidemolecule (colored), representing
one residue of a protein thatbinds to this region. The argininamide
fits into a pocket on the RNAsurface and is held in this
orientation by several noncovalent interac-tionswith the RNA. This
representation of the RNA molecule is pro-ducedwith the computer
program GRASP, which can calculate theshape of the outer surface of
a macromolecule, defined either by thevan der Waals radii of all
the atoms in the molecule or by the solventexclusion volume, into
which a water molecule cannot penetrate. 21. NH3H3 Molecular
configuration can be changed only bybreaking covalent bonds. For a
carbon atomwith four different substituents (a chiralcarbon), the
substituent groups can bearranged in two different ways,
generatingstereoisomers with distinct properties. Onlyone
stereoisomer is biologically active.Molecular conformation is the
position of atomsin space that can be changed by rotation
aboutsingle bonds, without breaking covalent bonds. Interactions
between biological molecules arealmost invariably stereospecific:
they require acomplementary match between the
interactingmolecules.1.3 Physical FoundationsLiving cells and
organisms must perform work to stayalive and to reproduce
themselves. The synthetic reac-tionsthat occur within cells, like
the synthetic processesin any factory, require the input of energy.
Energy is alsoconsumed in the motion of a bacterium or an
Olympicsprinter, in the flashing of a firefly or the electrical
dis-chargeof an eel. And the storage and expression ofinformation
require energy, without which structuresrich in information
inevitably become disordered andmeaningless.In the course of
evolution, cells have developedhighly efficient mechanisms for
coupling the energyobtained from sunlight or fuels to the many
energy-consumingprocesses they must carry out. One goal of1.3
Physical Foundations 21(R)-Carvone (S)-Carvone(spearmint)
(caraway)(a)CH3O CCH2CCCH3 CCH2CHCH2HCH3O CCH2CCCHCH2H
CCH2CH3(b)CHCH2CH2biochemistry is to understand, in quantitative
and chem-icalterms, the means by which energy is
extracted,channeled, and consumed in living cells. We can
considercellular energy conversionslike all other energy
con-versionsin the context of the laws of thermodynamics.Living
Organisms Exist in a Dynamic Steady State,Never at Equilibrium with
Their SurroundingsThe molecules and ions contained within a living
or-ganismdiffer in kind and in concentration from those inthe
organisms surroundings. A Paramecium in a pond,a shark in the
ocean, an erythrocyte in the human blood-stream,an apple tree in an
orchardall are different incomposition from their surroundings and,
once theyhave reached maturity, all (to a first
approximation)maintain a constant composition in the face of
con-stantlychanging surroundings.Although the characteristic
composition of an or-ganismchanges little through time, the
population ofmolecules within the organism is far from static.
Smallmolecules, macromolecules, and supramolecular com-plexesare
continuously synthesized and then brokendown in chemical reactions
that involve a constant fluxof mass and energy through the system.
The hemoglo-binmolecules carrying oxygen from your lungs to
yourbrain at this moment were synthesized within the pastmonth; by
next month they will have been degradedand entirely replaced by new
hemoglobin molecules.The glucose you ingested with your most recent
mealis now circulating in your bloodstream; before the dayis over
these particular glucose molecules will have beenOOC CH2CCON
HCHOOCH3OOCCH2CCONCHCOOCH3HCCHCHHCCCHNHCCHCHHCCCHH
HL-Aspartyl-L-phenylalanine methyl ester(aspartame)
(sweet)L-Aspartyl-D-phenylalanine methyl ester(bitter)FIGURE 123
Stereoisomers distinguishable by smelland taste in humans. (a) Two
stereoisomers of carvone:(R)-carvone (isolated from spearmint oil)
has thecharacteristic fragrance of spearmint; (S)-carvone
(fromcaraway seed oil) smells like caraway. (b) Aspartame,the
artificial sweetener sold under the trade nameNutraSweet, is easily
distinguishable by taste receptorsfrom its bitter-tasting
stereoisomer, although the twodiffer only in the configuration at
one of the two chiralcarbon atoms. 22. converted into something
elsecarbon dioxide or fat,perhapsand will have been replaced with a
fresh sup-plyof glucose, so that your blood glucose concentrationis
more or less constant over the whole day. The amountsof hemoglobin
and glucose in the blood remain nearlyconstant because the rate of
synthesis or intake of eachjust balances the rate of its breakdown,
consumption,or conversion into some other product. The constancyof
concentration is the result of a dynamic steady state,a steady
state that is far from equilibrium. Maintainingthis steady state
requires the constant investment of en-ergy;when the cell can no
longer generate energy, itdies and begins to decay toward
equilibrium with its sur-roundings.We consider below exactly what
is meant bysteady state and equilibrium.Organisms Transform Energy
and Matterfrom Their SurroundingsFor chemical reactions occurring
in solution, we can de-finea system as all the reactants and
products present,the solvent that contains them, and the immediate
at-mospherein short, everything within a defined regionof space.
The system and its surroundings together con-stitutethe universe.
If the system exchanges neithermatter nor energy with its
surroundings, it is said to beisolated. If the system exchanges
energy but not mat-terwith its surroundings, it is a closed system;
if it ex-changesboth energy and matter with its surroundings,it is
an open system.A living organism is an open system; it
exchangesboth matter and energy with its surroundings. Living
or-ganismsderive energy from their surroundings in twoways: (1)
they take up chemical fuels (such as glucose)from the environment
and extract energy by oxidizingthem (see Box 13, Case 2); or (2)
they absorb energyfrom sunlight.The first law of thermodynamics,
developed fromphysics and chemistry but fully valid for biological
sys-temsas well, describes the principle of the conservationof
energy: in any physical or chemical change, thetotal amount of
energy in the universe remains con-stant,although the form of the
energy may change.Cells are consummate transducers of energy,
capable ofinterconverting chemical, electromagnetic, mechanical,and
osmotic energy with great efficiency (Fig. 124).The Flow of
Electrons Provides Energy for OrganismsNearly all living organisms
derive their energy, directlyor indirectly, from the radiant energy
of sunlight, whicharises from thermonuclear fusion reactions
carried outin the sun. Photosynthetic cells absorb light energyand
use it to drive electrons from water to carbon di-oxide,forming
energy-rich products such as glucose(C6H12O6), starch, and sucrose
and releasing O2 into theatmosphere:light6CO26H2O 888n
C6H12O66O2(light-driven reduction of CO2)Nonphotosynthetic cells
and organisms obtain the en-ergythey need by oxidizing the
energy-rich products ofphotosynthesis and then passing electrons to
atmos-22 Chapter 1 The Foundations of
Biochemistry(a)(b)(c)(d)(e)Potential
energyEnergytransductionsaccomplishwork Nutrients in
environment(complex molecules such assugars, fats) SunlightChemical
transformationswithin cellsCellular work: chemical synthesis
mechanical work osmotic and electrical gradients light production
genetic information transferHeatIncreased randomness(entropy) in
the surroundingsMetabolism produces compoundssimpler than the
initialfuel molecules: CO2, NH3,H2O, HPO42Decreased
randomness(entropy) in the systemSimple compounds polymerizeto form
information-richmacromolecules: DNA, RNA,proteinsFIGURE 124 Some
energy interconversion in living organisms. Dur-ingmetabolic energy
transductions, the randomness of the system plussurroundings
(expressed quantitatively as entropy) increases as the
po-tentialenergy of complex nutrient molecules decreases. (a)
Living or-ganismsextract energy from their surroundings; (b)
convert some ofit into useful forms of energy to produce work; (c)
return some en-ergyto the surroundings as heat; and (d) release
end-product mole-culesthat are less well organized than the
starting fuel, increasing theentropy of the universe. One effect of
all these transformations is (e) in-creasedorder (decreased
randomness) in the system in the form ofcomplex macromolecules. We
return to a quantitative treatment of en-tropyin Chapter 13. 23.
pheric O2 to form water, carbon dioxide, and other endproducts,
which are recycled in the environment:C6H12O6O2 888n
6CO26H2Oenergy(energy-yielding oxidation of glucose)Virtually all
energy transductions in cells can be tracedto this flow of
electrons from one molecule to another,in a downhill flow from
higher to lower electrochem-icalpotential; as such, this is
formally analogous to theflow of electrons in a battery-driven
electric circuit. Allthese reactions involving electron flow are
oxidation-reductionreactions: one reactant is oxidized
(loseselectrons) as another is reduced (gains electrons).Creating
and Maintaining Order Requires Workand EnergyDNA, RNA, and proteins
are informational macromole-cules.In addition to using chemical
energy to form thecovalent bonds between the subunits in these
polymers,the cell must invest energy to order the subunits in
theircorrect sequence. It is extremely improbable that aminoacids
in a mixture would spontaneously condense into asingle type of
protein, with a unique sequence. This wouldrepresent increased
order in a population of molecules;but according to the second law
of thermodynamics, thetendency in nature is toward ever-greater
disorder in theuniverse: the total entropy of the universe is
continu-allyincreasing. To bring about the synthesis of
macro-moleculesfrom their monomeric units, free energy mustbe
supplied to the system (in this case, the cell).The randomness or
disorder of the components of achemical system is expressed as
entropy, S (Box 13).Any change in randomness ofthe system is
expressed asentropy change, S, which byconvention has a positive
valuewhen randomness increases.J. Willard Gibbs, who devel-opedthe
theory of energychanges during chemical reac-tions,showed that the
free-energycontent, G, of anyclosed system can be definedin terms
of three quantities:enthalpy, H, reflecting thenumber and kinds of
bonds;entropy, S; and the absolute temperature, T (in
degreesKelvin). The definition of free energy is GHTS.When a
chemical reaction occurs at constant tempera-ture,the free-energy
change, G, is determined bythe enthalpy change, H, reflecting the
kinds and num-bersof chemical bonds and noncovalent
interactionsbroken and formed, and the entropy change, S,
de-scribingthe change in the systems randomness:GHT S1.3 Physical
Foundations 23OOOHCNH2A process tends to occur spontaneously only
if Gis negative. Yet cell function depends largely on
mole-cules,such as proteins and nucleic acids, for which thefree
energy of formation is positive: the molecules areless stable and
more highly ordered than a mixture oftheir monomeric components. To
carry out these ther-modynamicallyunfavorable, energy-requiring
(ender-gonic)reactions, cells couple them to other reactionsthat
liberate free energy (exergonic reactions), so thatthe overall
process is exergonic: the sum of the free-energychanges is
negative. The usual source of freeenergy in coupled biological
reactions is the energy re-leasedby hydrolysis of phosphoanhydride
bonds suchas those in adenosine triphosphate (ATP; Fig. 125;
seealso Fig. 115). Here, each P represents a phosphorylgroup:Amino
acids 888n polymer G1 is positive (endergonic)OPOP 888nOP P G2 is
negative (exergonic)When these reactions are coupled, the sum of G1
andG2 is negativethe overall process is exergonic. Bythis coupling
strategy, cells are able to synthesize andmaintain the
information-rich polymers essential to life.Energy Coupling Links
Reactions in BiologyThe central issue in bioenergetics (the study
of energytransformations in living systems) is the means by
whichenergy from fuel metabolism or light capture is coupledto a
cells energy-requiring reactions. In thinking aboutenergy coupling,
it is useful to consider a simple me-chanicalexample, as shown in
Figure 126a. An objectat the top of an inclined plane has a certain
amount ofpotential energy as a result of its elevation. It tends
toslide down the plane, losing its potential energy of po-sitionas
it approaches the ground. When an appropri-atestring-and-pulley
device couples the falling object toanother, smaller object, the
spontaneous downward mo-tionof the larger can lift the smaller,
accomplishing aJ. Willard Gibbs,18391903O POO POO PO O CH2OHOHOHHH
HP P P Ribose AdenineCHCNN C NNFIGURE 125 Adenosine triphosphate
(ATP). The removal of the ter-minalphosphoryl group (shaded pink)
of ATP, by breakage of a phos-phoanhydridebond, is highly
exergonic, and this reaction is coupled tomany endergonic reactions
in the cell (as in the example in Fig. 126b). 24. certain amount of
work. The amount of energy availableto do work is the free-energy
change, G; this is al-wayssomewhat less than the theoretical amount
of en-ergyreleased, because some energy is dissipated as theheat of
friction. The greater the elevation of the largerobject, the
greater the energy released (G) as the ob-jectslides downward and
the greater the amount ofwork that can be accomplished.How does
this apply in chemical reactions? In closedsystems, chemical
reactions proceed spontaneously un-tilequilibrium is reached. When
a system is at equilib-rium,the rate of product formation exactly
equals the7 moleculesO2(a gas)rate at which product is converted to
reactant. Thusthere is no net change in the concentration of
reactantsand products; a steady state is achieved. The energychange
as the system moves from its initial state to equi-librium,with no
changes in temperature or pressure, isgiven by the free-energy
change, G. The magnitude ofG depends on the particular chemical
reaction and onhow far from equilibrium the system is initially.
Eachcompound involved in a chemical reaction contains a
cer-tainamount of potential energy, related to the kind andnumber
of its bonds. In reactions that occur sponta-neously,the products
have less free energy than the re-24 Chapter 1 The Foundations of
BiochemistryBOX 13 WORKING IN BIOCHEMISTRYEntropy: The Advantages
of Being DisorganizedThe term entropy, which literally means a
changewithin, was first used in 1851 by Rudolf Clausius, oneof the
formulators of the second law of thermody-namics.A rigorous
quantitative definition of entropyinvolves statistical and
probability considerations.However, its nature can be illustrated
qualitatively bythree simple examples, each demonstrating one
aspectof entropy. The key descriptors of entropy are ran-domnessand
disorder, manifested in different ways.Case 1: The Teakettle and
the Randomization of HeatWe know that steam generated from boiling
water cando useful work. But suppose we turn off the burnerunder a
teakettle full of water at 100 C (the sys-tem)in the kitchen (the
surroundings) and allowthe teakettle to cool. As it cools, no work
is done, butheat passes from the teakettle to the
surroundings,raising the temperature of the surroundings
(thekitchen) by an infinitesimally small amount until
com-pleteequilibrium is attained. At this point all parts ofthe
teakettle and the kitchen are at precisely the sametemperature. The
free energy that was once concen-tratedin the teakettle of hot
water at 100 C, poten-tiallycapable of doing work, has disappeared.
Itsequivalent in heat energy is still present in the teaket-tle
kitchen (i.e., the universe) but has becomecompletely randomized
throughout. This energy is nolonger available to do work because
there is no tem-peraturedifferential within the kitchen. Moreover,
theincrease in entropy of the kitchen (the surroundings)is
irreversible. We know from everyday experiencethat heat never
spontaneously passes back from thekitchen into the teakettle to
raise the temperature ofthe water to 100 C again.Case 2: The
Oxidation of GlucoseEntropy is a state not only of energy but of
matter.Aerobic (heterotrophic) organisms extract free en-ergyfrom
glucose obtained from their surroundingsby oxidizing the glucose
with O2, also obtained fromthe surroundings. The end products of
this oxidativemetabolism, CO2 and H2O, are returned to the
sur-roundings.In this process the surroundings undergoan increase
in entropy, whereas the organism itself re-mainsin a steady state
and undergoes no change inits internal order. Although some entropy
arises fromthe dissipation of heat, entropy also arises from
an-otherkind of disorder, illustrated by the equation forthe
oxidation of glucose:C6H12O66O2 On 6CO26H2OWe can represent this
schematically asThe atoms contained in 1 molecule of glucose plus
6molecules of oxygen, a total of 7 molecules, are morerandomly
dispersed by the oxidation reaction and arenow present in a total
of 12 molecules (6CO26H2O).Whenever a chemical reaction results in
an in-creasein the number of moleculesor when a solidsubstance is
converted into liquid or gaseous products,which allow more freedom
of molecular movementthan solidsmolecular disorder, and thus
entropy,increases.Case 3: Information and EntropyThe following
short passage from Julius Caesar, ActIV, Scene 3, is spoken by
Brutus, when he realizes thathe must face Mark Antonys army. It is
an information-richnonrandom arrangement of 125 letters of
theEnglish alphabet:CO2(a gas)H2O(a liquid)Glucose(a solid)12
molecules 25. abddfisactants, thus the reaction releases free
energy, which isthen available to do work. Such reactions are
exergonic;the decline in free energy from reactants to products
isexpressed as a negative value. Endergonic reactions re-quirean
input of energy, and their G values are posi-tive.As in mechanical
processes, only part of the energyreleased in exergonic chemical
reactions can be used toaccomplish work. In living systems some
energy is dissi-patedas heat or lost to increasing entropy.In
living organisms, as in the mechanical example inFigure 126a, an
exergonic reaction can be coupled toan endergonic reaction to drive
otherwise unfavorable1.3 Physical Foundations 25(a) Mechanical
exampleG0 G0Endergonic ExergonicReaction 2:ATP ADPPi Reaction
3:reactions. Figure 126b (a type of graph called a
reac-tioncoordinate diagram) illustrates this principle for
theconversion of glucose to glucose 6-phosphate, the firststep in
the pathway for oxidation of glucose. The sim-plestway to produce
glucose 6-phosphate would be:Reaction 1: GlucosePi On glucose
6-phosphate(endergonic; G1 is positive)2.(Pi is an abbreviation for
inorganic phosphate, HPO4Dont be concerned about the structure of
these com-poundsnow; we describe them in detail later in thebook.)
This reaction does not occur spontaneously; GThere is a tide in the
affairs of men,Which, taken at the flood, leads on to
fortune;Omitted, all the voyage of their lifeIs bound in shallows
and in miseries.In addition to what this passage says overtly, it
hasmany hidden meanings. It not only reflects a complexsequence of
events in the play, it also echoes the playsideas on conflict,
ambition, and the demands of lead-ership.Permeated with
Shakespeares understandingof human nature, it is very rich in
information.However, if the 125 letters making up this
quota-tionwere allowed to fall into a completely random,chaotic
pattern, as shown in the following box, theywould have no meaning
whatsoever.In this form the 125 letters contain little or no
infor-mation,but they are very rich in entropy. Such
con-siderationshave led to the conclusion that informationis a form
of energy; information has been called neg-ativeentropy. In fact,
the branch of mathematics calledinformation theory, which is basic
to the programminglogic of computers, is closely related to
thermodynamictheory. Living organisms are highly ordered,
nonran-domstructures, immensely rich in information and
thusentropy-poor.c
defghiIklmnoOraefhilmnoradefhilmnoradefhilnoraefhilnoradefhilno
aehilnoaehinoaeinoaeioeeieeeestTuvwWyustststtstststtttWorkdoneraisingobjectLoss
ofpotentialenergy ofposition(b) Chemical exampleFree energy,
GReaction 1:GlucosePi glucose 6-phosphateGlucoseATP glucose
6-phosphateADPG1G2 G3G3 = G1G2Reaction coordinateFIGURE 126 Energy
coupling in mechanical and chemicalprocesses. (a) The downward
motion of an object releases potentialenergy that can do mechanical
work. The potential energy made avail-ableby spontaneous downward
motion, an exergonic process (pink),can be coupled to the
endergonic upward movement of another ob-ject(blue). (b) In
reaction 1, the formation of glucose 6-phosphatefrom glucose and
inorganic phosphate (Pi) yields a product of higherenergy than the
two reactants. For this endergonic reaction, G is pos-itive.In
reaction 2, the exergonic breakdown of adenosine triphos-phate(ATP)
can drive an endergonic reaction when the two reactionsare coupled.
The exergonic reaction has a large, negative free-energychange
(G2), and the endergonic reaction has a smaller, positive
free-energychange (G1). The third reaction accomplishes the sum of
re-actions1 and 2, and the free-energy change, G3, is the
arithmeticsum of G1 and G2. Because G3 is negative, the overall
reactionis exergonic and proceeds spontaneously. 26. is positive. A
second, very exergonic reaction can occurin all cells:Reaction 2:
ATP On ADPPi(exergonic; G2 is negative)These two chemical reactions
share a common inter-mediate,Pi, which is consumed in reaction 1
and pro-ducedin reaction 2. The two reactions can therefore
becoupled in the form of a third reaction, which we canwrite as the
sum of reactions 1 and 2, with the commonintermediate, Pi, omitted
from both sides of the equation:Reaction 3: GlucoseATP Onglucose
6-phosphateADPBecause more energy is released in reaction 2 than
isconsumed in reaction 1, the free-energy change for re-action3,
G3, is negative, and the synthesis of glucose6-phosphate can
therefore occur by reaction 3.The coupling of exergonic and
endergonic reactionsthrough a shared intermediate is absolutely
central to theenergy exchanges in living systems. As we shall see,
thebreakdown of ATP (reaction 2 in Fig. 126b) is the
ex-ergonicreaction that drives many endergonic processesin cells.
In fact, ATP is the major carrier of chemicalenergy in all
cells.Keq and G Are Measures of a Reactions Tendencyto Proceed
SpontaneouslyThe tendency of a chemical reaction to go to
completioncan be expressed as an equilibrium constant. For
thereactionaAbB 888n cCdDthe equilibrium constant, Keq, is given
by[ACeeqc[[]aq]Keq[DBeeqd]bq]where [Aeq] is the concentration of A,
[Beq] the concen-trationof B, and so on, when the system has
reachedequilibrium. A large value of Keq means the reactiontends to
proceed until the reactants have been almostcompletely converted
into the products.Gibbs showed that G for any chemical reaction isa
function of the standard free-energy change,G a constant that is
characteristic of each specificreactionand a term that expresses
the initial concen-trationsof reactants and
products:[ACic[[]ai]GGRT ln [DB]]iidb (11)where [Ai] is the initial
concentration of A, and so forth;R is the gas constant; and T is
the absolute temperature.When a reaction has reached equilibrium,
no driv-ingforce remains and it can do no work: G0. Forthis special
case, [Ai][Aeq], and so on, for all reactantsand products,
and[ACeeqc[[]aq] [DBee]]qqdbKeq[Ci]c[Di]d[Ai]a[Bi]bSubstituting 0
for G and Keq for [Ci]c[Di]d/[Ai]a[Bi]b inEquation 11, we obtain
the relationshipGRT ln Keqfrom which we see that G is simply a
second way (be-sidesKeq) of expressing the driving force on a
reaction.Because Keq is experimentally measurable, we have away of
determining G, the thermodynamic constantcharacteristic of each
reaction.The units of G and G are joules per mole (orcalories per
mole). When Keq1, G is large andnegative; when Keq1, G is large and
positive.From a table of experimentally determined values of
ei-therKeq or G, we can see at a glance which reactionstend to go
to completion and which do not.One caution about the interpretation
of G: ther-modynamicconstants such as this show where the
fi-nalequilibrium for a reaction lies but tell us nothingabout how
fast that equilibrium will be achieved. Therates of reactions are
governed by the parameters of ki-netics,a topic we consider in
detail in Chapter 6.Enzymes Promote Sequences of Chemical
ReactionsAll biological macromolecules are much less
thermody-namicallystable than their monomeric subunits, yetthey are
kinetically stable: their uncatalyzed break-downoccurs so slowly
(over years rather than seconds)that, on a time scale that matters
for the organism, thesemolecules are stable. Virtually every
chemical reactionin a cell occurs at a significant rate only
because of thepresence of enzymesbiocatalysts that, like all
othercatalysts, greatly enhance the rate of specific
chemicalreactions without being consumed in the process.The path
from reactant(s) to product(s) almost in-variablyinvolves an energy
barrier, called the activationbarrier (Fig. 127), that must be
surmounted for any re-actionto proceed. The breaking of existing
bonds andformation of new ones generally requires, first, the
dis-tortionof the existing bonds, creating a transitionstate of
higher free energy than either reactant or prod-uct.The highest
point in the reaction coordinate dia-gramrepresents the transition
state, and the differencein energy between the reactant in its
ground state andin its transition state is the activation energy,
G.An enzyme catalyzes a reaction by providing a morecomfortable fit
for the transition state: a surface thatcomplements the transition
state in stereochemistry, po-larity,and charge. The binding of
enzyme to the transi-tionstate is exergonic, and the energy
released by thisbinding reduces the activation energy for the
reactionand greatly increases the reaction rate.A further
contribution to catalysis occurs when twoor more reactants bind to
the enzymes surface close toeach other and with stereospecific
orientations that fa-26 Chapter 1 The Foundations of Biochemistry
27. vor the reaction. This increases by orders of