Embed Size (px)
Citation preview

research papers
Acta Cryst. (2017). D73, 349–364 https://doi.org/10.1107/S2059798317002534 349
Received 9 November 2016
Accepted 14 February 2017
Edited by J. L. Martin, Griffith University,
Australia
Keywords: 8-oxoguanine nucleotides;
sanitization of nucleotide pool; Nudix enzyme;
histidine phosphatase domain; binding sites at
intermolecular interfaces; enzyme action;
MutT1; Mycobacterium smegmatis.
PDB references: Mycobacterium smegmatis
MutT1, 5gg5; complex with 8-oxo-dGTP, 5gg6;
complex with 8-oxo-dGTP, 8-oxo-dGMP and
pyrophosphate (I), 5gg7; complex with
8-oxo-dGTP, 8-oxo-dGMP and pyrophosphate
(II), 5gg8; complex with 8-oxo-GTP, 8-oxo-GMP
and pyrophosphate, 5gg9; complex with
8-oxo-GDP, 8-oxo-GMP and pyrophosphate,
5gga; complex with 8-oxo-dGDP, 5ggb;
complex with phosphate and magnesium ions
(excess magnesium) (I), 5ggc; complex with
phosphate and magnesium ions (excess
magnesium) (II), 5ggd
Supporting information: this article has
supporting information at journals.iucr.org/d
Biochemical and structural studies ofMycobacterium smegmatis MutT1, a sanitizationenzyme with unusual modes of association
S. M. Arif,a A. G. Patil,b U. Varshneyb and M. Vijayana*
aMolecular Biophysics Unit, Indian Institute of Science, Bangalore 560 012, India, and bDepartment of Microbiology and
Cell Biology, Indian Institute of Science, Bangalore 560 012, India. *Correspondence e-mail: [email protected]
Mycobacterium smegmatis MutT1, which is made up of a Nudix domain (domain
1) and a histidine phosphatase domain (domain 2), efficiently hydrolyses 8-oxo-
GTP and 8-oxo-dGTP to the corresponding nucleoside diphosphates and
phosphate in the presence of magnesium ions. Domain 1 alone hydrolyses
nucleoside triphosphates less efficiently. Under high concentrations and over
long periods, the full-length enzyme as well as domain 1 catalyses the hydrolysis
of the nucleoside triphosphates to the respective nucleoside monophosphates
and pyrophosphate. The role of domain 2 appears to be limited to speeding up
the reaction. Crystal structures of the apoenzyme and those of ligand-bound
enzyme prepared in the presence of 8-oxo-GTP or 8-oxo-dGTP and different
concentrations of magnesium were determined. In all of the structures except
one, the molecules arrange themselves in a head-to-tail fashion in which domain
1 is brought into contact with domain 2 (trans domain 2) of a neighbouring
molecule. The binding site for NTP (site A) is almost exclusively made up of
residues from domain 1, while those for NDP (site B) and NMP (site C) are at
the interface between domain 1 and trans domain 2 in an unusual instance of
intermolecular interactions leading to binding sites. Protein–ligand interactions
at site A lead to a proposal for the mechanism of hydrolysis of NTP to NDP and
phosphate. A small modification in site A in the crystal which does not exhibit
the head-to-tail arrangement appears to facilitate the production of NMP and
pyrophosphate from NTP. The two arrangements could be in dynamic
equilibrium in the cellular milieu.
1. Introduction
Reactive oxygen species and reactive nitrogen intermediates
produced during cellular metabolism or present as environ-
mental pollutants generate oxidatively damaged nucleoside
triphosphates (NTPs) and deoxynucleoside triphosphates
(dNTPs) in the cell (Cooke et al., 2003; Szabo & Ohshima,
1997; Hayakawa et al., 1999). Excessive accumulation of these
nucleotides in the nucleotide pool impairs the fidelity of DNA
replication (Pursell et al., 2008) and RNA transcription
(Kamiya et al., 2009). Of the nucleic acid bases, guanine, which
is highly susceptible to oxidation, is converted to 8-oxo-7,8-
dihydroguanine (8-oxo-G). Post-incorporation, 8-oxo-G is a
major source of spontaneous mutation because of its ambig-
uous and almost equally efficient pairing with cytosine and
adenine during replication (Kuchino et al., 1987; Moriya et al.,
1991; Shibutani et al., 1991). To deal with such challenging
threats to genomic integrity, cells have developed an elaborate
8-oxo-G (GO) repair system consisting of MutM (Fpg), MutY
and MutT (Kurthkoti & Varshney, 2011; Michaels & Miller,
1992). 8-Oxo-G paired with cytosine is excised by MutM, while
MutY removes adenine or guanine paired with 8-oxo-G
during replication (Au et al., 1989; Jain et al., 2007; Kurthkoti et
ISSN 2059-7983
# 2017 International Union of Crystallography

al., 2010). The MutT protein, on the other hand, works in a
completely different way by hydrolysing 8-oxo-G-containing
nucleotides in the nucleotide pool, thereby minimizing their
direct incorporation into DNA (Michaels & Miller, 1992).
MutT has been reported to have a high specificity for 8-oxo-G-
containing nucleotides. It hydrolyses 8-oxo-dGTP to 8-oxo-
dGMP at a rate three times faster than it hydrolyses dGTP to
dGMP (Maki & Sekiguchi, 1992). Thus, MutT substantially
minimizes errors in DNA replication and RNA transcription
caused by 8-oxo-G-containing nucleotides by sanitizing them
selectively from the nucleotide pool (Ito et al., 2005; Maki &
Sekiguchi, 1992; Taddei et al., 1997). MutT homologue 1
(MTH1), another 8-oxo-dGTPase, also hydrolyses 8-oxo-
dGTP to 8-oxo-dGMP (Nakabeppu et al., 2006).
MutT proteins are Nudix hydrolases, a versatile superfamily
of Mg2+-requiring enzymes that are found in almost every
form of life, including viruses, bacteria, archaea and eukary-
otes, and catalyse the hydrolysis of nucleoside diphosphate
linked to other moieties X, hence the name ‘Nudix’ (Bessman
et al., 1996). These enzymes are recognized by the highly
conserved 23-residue Nudix signature sequence or Nudix
motif, GX5EX7REUXEEXGU, where U is Leu, Ile or Val and
X is any residue (Bessman et al., 1996).
The susceptibility to guanosine modification in myco-
bacteria is very high owing both to the high GC content of
their genome and to the presence of reactive oxygen species
and reactive nitrogen intermediates produced by macrophages
as a host immune response following an infection. To deal with
this situation, mycobacterial genomes encode all of the
proteins specific to the GO repair pathway (Cole et al., 1998).
Of the nine proteins with a Nudix motif predicted by bio-
informatics studies, four putative Nudix hydrolases annotated
as MutT homologues, namely MutT1, MutT2, MutT3 and
MutT4, correspond to Escherichia coli MutT (EcMutT; Cole et
al., 1998; Dos Vultos et al., 2006; Moreland et al., 2009). Gene-
knockout studies have shown that both MutT1 and MutT4
have an antimutator role in Mycobacterium smegmatis (Dos
Vultos et al., 2006), while MutT3 (RenU) is an important
component of the redox homeostatic system, RHOCS, that is
critical for the survival of mycobacteria inside macrophages
and the formation of biofilms (Wolff et al., 2015). Subse-
quently, the involvement of M. tuberculosis MutT1 (MtMutT1)
in decreasing A to C mutations in a MutT-deficient strain of
E. coli was reported, thus suggesting an inherent antimutator
role for the enzyme (Patil et al., 2013). Moreover, MtMutT1
has been suggested to play a crucial role in the survival of the
bacteria under oxidative stress by the virtue of its antimutator
role (Patil et al., 2013). MutT1 and MutT2 have been shown to
have an 8-oxoguanosine triphosphatase activity (Dos Vultos et
al., 2006; Patil et al., 2013; Sang & Varshney, 2013). Interest-
ingly, MtMutT1 has been shown to hydrolyse 8-oxo-GTP and
8-oxo-dGTP to the corresponding nucleoside diphosphates,
but not to nucleoside monophosphates, at normal substrate
and enzyme concentrations (Patil et al., 2013). To understand
the basis of this novel and unusual activity and substrate
specificity, it was desirable to carry out structural studies on
MtMutT1. However, on account of problems with the
expression of MtMutT1, M. smegmatis MutT1 (MsMutT1) was
chosen for structural studies. Our preliminary studies on
MsMutT1 suggested the presence of an N-terminal Nudix
hydrolase domain (MsMutT1-NTD) corresponding to the
single-domain EcMutT and a C-terminal histidine phospha-
tase domain (MsMutT1-CTD) in the protein (Arif et al., 2012).
The presence of a phosphatase domain in MsMutT1 in addi-
tion to a Nudix hydrolase domain of the type that constitutes
EcMutT and human MutT homologue 1 (HsMTH1) was
intriguing. We have now carried out detailed crystallographic
studies of MsMutT1 and its complexes, along with comple-
mentary biochemical studies, as part of a long-range program
on the structural biology of mycobacterial proteins, which
forms a component of a concerted national and international
effort (Arora et al., 2011; Murillo et al., 2007; Terwilliger et al.,
2003). The protein appears to be an enzyme in which the
binding sites are formed primarily by intermolecular inter-
actions of the type that bring the Nudix domain of one
molecule and the phosphatase domain of a neighbouring
molecule into close proximity. The enzyme is capable of
hydrolysing 8-oxo-GTP and 8-oxo-dGTP to the corresponding
nucleoside diphosphates and monophosphates, simulta-
neously, sequentially or both. A detailed examination of the
crystal structures leads to a proposal as to how this is achieved.
2. Materials and methods
2.1. Cloning, expression and purification of MsMutT1,MsMutT1-NTD and MsMutT1-CTD
PCR amplification of the gene MSMEG_2390 (UniProt
reference No. A0QUZ2) from the M. smegmatis mc2155
genome was carried out by Phusion DNA polymerase using
the primers MsMutT1Fp (50-GATCCATATGTCGAAGGA-
CACCGA-30) and MsMutT1Rp (50-GTTGAAGCTTTACT-
TCTCGTC-30) containing NdeI and HindIII sites, respectively
(indicated in bold), as described previously for cloning of the
M. tuberculosis MutT1 (MtMutT1) gene (Patil et al., 2013).
The PCR amplicon was digested with NdeI and HindIII and
ligated into the pTrcNdeHis vector to generate pTrcNdeHis-
MsMutT1. The sequence of the construct was confirmed by
DNA-sequence analysis and it was then used to generate
clones corresponding to MsMutT1-NTD and MsMutT1-CTD.
The generation of clones corresponding to MsMutT1-NTD
and MsMutT1-CTD involved the PCR insertion of a STOP
codon followed by an NdeI restriction site between the
nucleotide sequences corresponding to the two domains in the
pTrcNdeHis-MsMutT1 construct. Site-directed mutagenesis
of pTrcNdeHis-MsMutT1 involving these insertions was
performed by PCR using Phusion DNA polymerase and the
primers MsMutT1(del)Fp (50-TTCGTAAAACGACCGTA-
ACATATGGTCGACACCAAGACGGT-30) and MsMutT1
(del)Rp (50-ACCGTCTTGGTGTCGACCATATGTTACG-
GTCGTTTTACGAA-30) with insertion sequences corre-
sponding to a STOP codon and an NdeI restriction site
(indicated in bold), employing the QuikChange protocol for
site-directed mutagenesis (Agilent). The mutant plasmid thus
research papers
350 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364

obtained was confirmed by restriction digestion using NdeI
and represents pTrcNdeHis-MsMutT1-NTD, a clone corre-
sponding to the N-terminal domain of MsMutT1. The
pTrcNdeHis-MsMutT1-NTD clone was then treated with the
NdeI enzyme, thereby removing the nucleotide sequence
corresponding to the N-terminal domain. This was followed
by separation and ligation of the digested clone, resulting in
pTrcNdeHis-MsMutT1-CTD, a clone corresponding to the
C-terminal domain of MsMutT1. The sequences of both of the
constructs were confirmed by DNA sequencing. A peptide
stretch, MGSSHHHHHHSSGLVPRGSH, containing the
histidine tag was appended to the N-terminus of all three of
the protein sequences during the cloning procedure.
Proteins were purified by Ni–NTA affinity chromatography
followed by size-exclusion chromatography using a Superdex
200 HiLoad 16/600 column and a buffer consisting of 25 mM
Tris–HCl pH 7.5, 500 mM NaCl, 10% glycerol, 2 mM
�-mercaptoethanol in conditions that were essentially iden-
tical to those described previously for MtMutT1 (Patil et al.,
2013).
2.2. Activity assays for MsMutT1, MsMutT1-NTD andMsMutT1-CTD
All three proteins were dialyzed extensively against a buffer
consisting of 25 mM Tris–HCl pH 7.5, 50 mM NaCl, 2%(v/v)
glycerol, 2 mM �-mercaptoethanol before performing an
activity assay. Activity assays were performed in assay buffers
and conditions similar to those used previously for MtMutT1
and human Nudix type 5 (NUDT5) proteins (Patil et al., 2013;
Ishibashi et al., 2003) with some modifications. Reaction
mixtures (10 ml) consisting of 25 mM Tris–HCl pH 7.5, 8 mM
MgCl2, 50 mM NaCl, 5 mM DTT, 2% glycerol, various NTP or
dNTP substrates (�100–1000 mM) and enzymes were incu-
bated at 310 K. Reactions were terminated by adding 10 ml of
0.1% SDS and the mixtures were analysed by separating them
using a DNAPac column (DNAPac PA200 analytical, 4 �
250 mm), employing an integrated high-pressure liquid-
chromatography (HPLC) system (UltiMate 3000). An elution
system consisting of buffer I (25 mM Tris–HCl pH 9.0) and
buffer II (1 M lithium chloride) with a gradient of 0–50% was
used at an isocratic flow rate of 0.5 ml min�1 for 28 min.
Nucleotides were detected by UV-light absorbance. Peaks
corresponding to guanosine and 8-oxoguanosine nucleotides
were detected at a wavelength of 252 nm.
2.3. Crystallization
The protein solution was dialyzed against a buffer
consisting of 25 mM Tris–HCl pH 7.5, 500 mM NaCl, 10%
glycerol, 2 mM �-mercaptoethanol prior to crystallization.
Crystals of apo MsMutT1 were obtained after extensive
screening using screening kits from Hampton Research
employing the microbatch-under-oil method at 298 K (Arif et
al., 2012). The quality of the crystals was improved by incu-
bating them in a drop containing the precipitant supplemented
with 20% ethylene glycol. Similar crystallization trials were
used to generate crystals of MsMutT1 complexed with the
known substrates 8-oxo-GTP and 8-oxo-dGTP (obtained from
Jena Bioscience) by the co-crystallization method. A mixture
consisting of the protein solution at a concentration of
7 mg ml�1 and the substrates at a molar concentration ten
times that of the protein was incubated for 2 h at 277 K before
crystallization. The above mixture supplemented with 10 mM
MgCl2 was also incubated in a similar fashion for another
set of crystallization experiments. Drops containing equal
volumes of the precipitant solution and the protein–substrate
mixture were used for crystallization. Eight further crystals
of MsMutT1 were generated during these crystallization
attempts using various precipitant solutions. Details of these
crystals, along with that of apo MsMutT1, and the respective
precipitants involved are given in Table 1.
2.4. X-ray data collection, structure solution and refinement
Intensity data from two of the nine crystals (crystal 3 and
crystal 8) were collected using a MAR345 detector mounted
on a Bruker-AXS Microstar Ultra II Cu K� rotating-anode
X-ray generator. Data sets from the remaining seven crystals
were collected using a CCD detector on the synchrotron X-ray
beamline BM14 at the European Synchrotron Radiation
Facility, Grenoble, France. All of the data sets were collected
at a temperature of 100 K using ethylene glycol or glycerol as
research papers
Acta Cryst. (2017). D73, 349–364 Arif et al. � Mycobacterium smegmatis MutT1 351
Table 1Basic data on the different crystals.
See text for details. The values in parentheses in column 3 are the concentrations of magnesium ions in the medium.
Crystal No. LigandPrecipitant†(Mg concentration) Form
Head-to-tailarrangement Site A Site B Site C
1 None 1 I Yes2 8-Oxo-dGTP 2 II No 8-Oxo-dGTP3 8-Oxo-dGTP 2 III Yes 8-Oxo-dGTP PPi 8-Oxo-dGMP4 8-Oxo-dGTP 2 IV Yes 8-Oxo-dGTP PPi 8-Oxo-dGMP5 8-Oxo-GTP 2 IV Yes 8-Oxo-GTP PPi 8-Oxo-GMP6 8-Oxo-GTP (MgCl2 soaking) 2 (10 mM, 2 min) IV Yes 8-Oxo-GDP PPi 8-Oxo-GMP7 8-Oxo-dGTP (MgCl2 co-crystallization) 3 (10 mM) V Yes 8-Oxo-dGDP8 8-Oxo-dGTP 4 (200 mM) VI Yes9 8-Oxo-GTP 4 (200 mM) VI Yes
† 1, 0.2 M ammonium sulfate, 0.1 M MES monohydrate pH 6.5, 30%(w/v) PEG monomethyl ether 5000, 0.01 M TCEP hydrochloride; 2, 0.2 M sodium acetate trihydrate, 0.1 M Tris–HClpH 8.5, 30%(w/v) PEG 4000; 3, 0.2 M sodium acetate, 0.1 M sodium acetate trihydrate pH 4.6, 30%(w/v) PEG 4000; 4, 0.2 M magnesium chloride hexahydrate, 0.1 M Tris–HCl pH 8.5,30%(w/v) PEG 4000.
cryoprotectant. The data were processed and merged using
iMosflm (Battye et al., 2011) and SCALA (Evans, 2006) from
the CCP4 program suite (Winn et al., 2011). Structure-factor
amplitudes were obtained from intensities using TRUNCATE
from the CCP4 program suite.
The crystal structure of apo MsMutT1 was determined by
molecular replacement with Phaser (McCoy et al., 2007) from
the CCP4 program suite using a data set collected previously
(Arif et al., 2012). A value of 2.32 A3 Da�1 for the Matthews
coefficient suggested the presence of one molecule in the
asymmetric unit (Matthews, 1968). The coordinates of puta-
tive MutT1 from Bifidobacterium adolescentis (PDB entry
3fjy; New York SGX Research Center for Structural Geno-
mics, unpublished work), with a sequence identity of 28% to
MsMutT1, were used as the search model to obtain the initial
phases. The solution obtained from Phaser was then built into
the partially interpretable electron-density map using Coot
(Emsley et al., 2010). An initial rigid-body refinement followed
by positional refinement and individual B-factor refinement
was performed on this partially built solution using
REFMAC5 (Murshudov et al., 2011). Cycles of model building
and subsequent refinement followed. The structure thus
obtained was further refined using the high-resolution X-ray
data obtained subsequently. The coordinates of this structure
were then used as a search model to determine the structures
of the various MsMutT1–ligand complexes from the remaining
crystals by the molecular-replacement method. The structures
underwent similar refinement as described previously. The
2Fo � Fc and Fo � Fc maps computed at stages where R and
Rfree had acceptable values showed unambiguous density for
the respective ligands. The ligand molecules were subse-
quently modelled and their locations were further confirmed
by computing simulated-annealing Fo � Fc OMIT maps using
CNS v.1.3 (Brunger et al., 1998). Various components from the
research papers
352 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364
Table 2Details of data collection and refinement.
Values in parentheses are for the highest resolution shell.
Crystal No. 1 2 3 4 5 6 7 8 9
Data collectionSpace group P212121 P21 P21 P21 P21 P21 P212121 P21 P21
Unit-cell parametersa (A) 37.45 57.11 42.00 42.05 42.01 41.73 42.49 38.99 38.95b (A) 87.66 89.50 85.35 84.81 83.95 84.30 87.89 87.67 87.60c (A) 101.28 59.93 87.43 43.52 44.00 43.58 95.20 90.68 90.53� (�) 90.0 99.1 94.1 93.6 93.0 94.0 90.0 99.9 100.1
Resolution range (A) 43.85–1.64(1.73–1.64)
49.36–1.75(1.84–1.75)
29.90–1.70(1.79–1.70)
43.43–1.85(1.95–1.85)
43.94–1.60(1.69–1.60)
43.47–1.75(1.84–1.75)
64.59–1.10(1.16–1.10)
31.97–1.85(1.95–1.85)
44.57–1.70(1.79–1.70)
VM (A3 Da�1) 2.18 1.98 2.05 2.03 2.03 2.0 2.3 2.0 2.0Solvent content (%) 43.5 37.9 40.0 39.4 39.4 38.6 47.2 38.5 38.3No. of molecules in
asymmetric unit1 2 2 1 1 1 1 2 2
Unique reflections 41476 (5726) 59643 (8608) 66623 (9503) 26082 (3818) 40162 (5870) 30343 (4381) 145101 (20940) 51169 (7409) 65728 (9517)Multiplicity 3.6 (3.2) 5.0 (5.0) 4.5 (4.4) 4.2 (4.2) 4.5 (4.5) 5.9 (5.8) 7.9 (7.0) 5.3 (5.2) 3.2 (3.1)Completeness (%) 99.4 (96.3) 99.4 (98.8) 98.5 (96.5) 100.0 (100.0) 99.9 (100.0) 100.0 (100.0) 100.0 (99.9) 99.8 (99.3) 99.9 (100.0)hI/�(I)i 11.4 (3.6) 9.8 (1.9) 7.9 (1.8) 9.5 (2.0) 10.8 (2.1) 9.1 (1.6) 10.2 (1.9) 7.6 (1.9) 6.6 (1.5)Rmerge† (%) 7.1 (33.7) 8.7 (67.5) 8.6 (78.0) 8.6 (68.3) 7.3 (65.3) 11.2 (98.0) 9.8 (96.2) 13.1 (79.8) 7.9 (68.0)CC1/2 0.996 (0.832) 0.996 (0.803) 0.998 (0.593) 0.993 (0.770) 0.996 (0.800) 0.993 (0.706) 0.996 (0.720) 0.997 (0.744) 0.996 (0.789)
Refinement and model statisticsR factor (%) 15.7 16.2 19.6 17.6 16.5 17.8 16.0 22.3 21.6Rfree (%) 19.7 20.4 23.6 22.0 20.6 22.9 18.5 26.7 25.9R.m.s. deviation from ideal
Bond lengths (A) 0.018 0.019 0.015 0.017 0.016 0.018 0.02 0.015 0.017Bond angles (�) 2.0 2.0 2.0 2.0 2.0 2.0 2.0 1.7 1.9
No. of non-H atomsProtein atoms 2441 4686 4617 2334 2357 2352 2511 4598 4594Ligands (NTP/NDP/
NMP/PPi)— 64 178 65 67 63 28 — —
Solvent atoms 457 562 644 262 339 239 486 484 527Average B factors (A2)
Protein atoms 15.2 30.8 26.7 37.0 27.5 35.9 15.9 29.4 30.7Ligands (NTP/NDP/
NMP/PPi)— 27.7 38.3 46.1 28.8 45.7 17.5 — —
Solvent atoms 32.7 39.0 35.6 42.8 38.2 42.8 31.6 34.2 36.6Ramachandran plot statistics‡ (residues in, %)
Favoured region 91.5 90.9 91.7 91.6 90.7 91.1 91.9 89.8 91.4Allowed region 8.5 9.1 7.7 8.4 8.9 8.9 7.7 10.0 8.2Generously allowed
region0.0 0.0 0.6 0.0 0.4 0.0 0.4 0.2 0.4
Disallowed region 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0PDB code 5gg5 5gg6 5gg7 5gg8 5gg9 5gga 5ggb 5ggc 5ggd
† Rmerge =P
hkl
Pi jIiðhklÞ � hIðhklÞij=
Phkl
Pi IiðhklÞ, where Ii(hkl) is the ith observation of reflection hkl and hI(hkl)i is the weighted average intensity for all i observations of
reflection hkl. ‡ Calculated for nonglycine and nonproline residues using PROCHECK.

research papers
Acta Cryst. (2017). D73, 349–364 Arif et al. � Mycobacterium smegmatis MutT1 353
Figure 1HPLC separation of the nucleotide substrates and the products formed by the action of the enzyme using a DNAPac column (DNAPac PA200 analytical,4 � 250 mm). (a) Activity assay using full-length MsMutT1. (i)–(iii) are controls for (iv), (v) for (vi), (vii) for (viii) and (ix) for (x). (b) Activity assayusing MsMutT1-NTD (i, ii) and MsMutT1-CTD (iii, iv). (c) Effect on catalysis of a higher concentration and longer incubation of full-length MsMutT1(ii) and MsMutT1-NTD (iii) with the substrates. (i) is the common control for (ii) and (iii). (d) Analysis of the crystallization solution used during co-crystallization of MsMutT1 with 8-oxo-GTP/8-oxo-dGTP. (i) is the control for (ii). The MgCl2 concentration was 8 mM in (a) and (b), while it was200 mM in (c). MgCl2 was not added in (d). In all of the figures, the x axis is the retention time (min) of the nucleotides and the y axis is intensity of thenucleotide peak in milli-absorbance units (mAU) at a wavelength (WVL) of 252 nm.

precipitant and the cryoprotectant were also modelled into the
electron density when appropriate. O atoms of water mole-
cules were successively added to the structures in the final
cycles of refinement using peaks with heights greater than 3.0�and 1.0� in Fo � Fc and 2Fo � Fc maps, respectively. Alter-
native conformations were assigned to the side chains of a few
of the amino-acid residues and one ligand molecule in one of
the crystals during the course of model building and refine-
ment. In the cases where the data set had a resolution better
than 1.5 A, anisotropic B factors were used in the final stage of
refinement. Refinement parameters, along with data-collec-
tion and data-processing statistics, are summarized in Table 2.
2.5. Analysis of the structures
The refined structures were evaluated using PROCHECK
(Laskowski et al., 1993). Secondary structure was assigned
using STRIDE (Heinig & Frishman, 2004). Interatomic
distances were calculated using
CONTACT from the CCP4
program suite. Hydrogen bonds
were assigned based on a distance
less than or equal to 3.6 A
between the donor (D) and the
acceptor (A) atom and a D–
H� � �A angle greater than 90�.
Structural superpositions were
performed using ALIGN (Cohen,
1997). The buried surface area
was calculated using NACCESS
(Hubbard & Thornton, 1996).
Figures for molecular repre-
sentations were generated using
PyMOL (DeLano, 2002).
Topology and parameter files for
various nucleotides were gener-
ated by ACPYPE (AnteChamber
PYthon Parser interfacE; Sousa
da Silva & Vranken, 2012).
Energy minimizations of the
protein–ligand complexes were
performed using the GROMACS
v.5.0.7 package employing the
AMBER99SB force field
(Hornak et al., 2006). Free ener-
gies of ligand binding to the
protein were estimated using
AutoDock 4.2 (Morris et al.,
2009).
2.6. Dynamic light-scatteringmeasurements
A dynamic light-scattering
(DLS) experiment on MsMutT1
at three different concentrations
including one corresponding to
that used for crystallization
experiments (7 mg ml�1) was carried out at the Centre for
Cellular and Molecular Biology (CCMB), Hyderabad using a
SpectroSize 300, a cuvette-based DLS instrument employing a
laser diode operating at a wavelength of 660 nm, a scattering
angle of 90� and an avalanche photodiode detector. The
protein solutions were centrifuged at 13 000 rev min�1 prior to
the DLS measurements. 20 measurements were made for each
sample during the DLS experiments. The collected data were
analysed using the SpectroSize 300 software, resulting in
average hydrodynamic size-distribution profiles. Deconvolu-
tion of the autocorrelation function was performed using the
CONTIN algorithm (Provencher, 1982).
2.7. Small-angle X-ray scattering (SAXS) measurements
SAXS measurements of MsMutT1 in a buffer as used for
crystallization were recorded, again at CCMB, using an
S3-MICRO Point-Focus system (Hecus X-ray Systems),
research papers
354 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364
Figure 2Overall structure of MsMutT1. (a) Domain architecture of MsMutT1 and structural features of (b)MsMutT1-NTD (domain 1) and (c) MsMutT1-CTD (domain 2). Domain 1 is in red and domain 2 is ingreen. The linker region is in blue. The highly conserved Nudix motif in MsMutT1-NTD, which adopts acharacteristic SLHL (strand–loop–helix–loop) structure, and the conserved RHG motif in MsMutT1-CTDare shown in yellow.

employing an X-ray wavelength of 1.5418 A and a PILATUS
100K detector with a pixel size of 172 � 172 mm at a distance
of 300 mm from the sample. Scatterings were measured with
an exposure time of 2 h. Averaging of the raw data was carried
out using Fit2D (Hammersley, 1997). The data were analysed
using the ATSAS (Konarev et al., 2006) program suite and
scattering plots were generated using PRIMUS (Konarev et al.,
2003) from the same suite.
3. Results and discussion
3.1. Enzyme activity
The enzymatic activity of MsMutT1 against GTP and dGTP
and their corresponding oxidized forms, namely 8-oxo-GTP
and 8-oxo-dGTP, were analysed in identical assay conditions
involving an assay buffer similar to that used previously for
MtMutT1 and human Nudix type 5 (NUDT5) proteins (Patil et
al., 2013; Ishibashi et al., 2003), consisting of 25 mM Tris–HCl
pH 7.5, 8 mM MgCl2, 50 mM NaCl, 5 mM DTT, 2% glycerol
and an enzyme concentration of 0.27 mM (0.01 mg ml�1).
MsMutT1 hydrolyses 8-oxo-GTP and 8-oxo-dGTP to 8-oxo-
GDP and 8-oxo-dGDP, respectively (Fig. 1a); hydrolysis of
GTP and dGTP was not observed (Fig. 1a). 8-Oxo-GDP or
8-oxo-dGDP did not hydrolyse any further even after incu-
bation with the enzyme for 12–24 h.
The presence of a C-terminal histidine phosphatase domain
(MsMutT1-CTD) in addition to the N-terminal Nudix
hydrolase domain in MsMutT1 (MsMutT1-NTD) necessitated
an examination of the role of the phosphatase domain in the
enzyme. Thus, the enzymatic activities of the two domains
individually against 8-oxo-GTP and 8-oxo-dGTP were also
checked. Surprisingly, MsMutT1-NTD could hydrolyse both
the substrates to the corresponding nucleoside diphosphates.
However, it showed a diminished activity compared with the
full-length enzyme (Fig. 1b). For instance, 2.7 pmol of
MsMutT1 took approximately 30 min to hydrolyse 1 nmol of
8-oxo-dGTP completely. The same amount of substrate,
however, required 21.7 pmol of MsMutT1-NTD (approxi-
mately eight times that of MsMutT) and almost 4 h, rather
than just the 30 min required by the full-length enzyme, for
complete hydrolysis. MsMutT1-CTD, on the other hand, did
not show any activity towards either 8-oxo-GTP or 8-oxo-
dGTP (Fig. 1b). In another experiment, a mixture of the two
domains instead of the full-length protein was used in the
assay. The result of the experiment was identical to that
obtained when MsMutT1-NTD alone was used (Supplemen-
tary Fig. S1). Thus, it appears that although it is inactive
against either of the substrates, the presence of the C-terminal
histidine phosphatase domain is necessary as an integral part
of the enzyme molecule for efficient catalysis by the N-
terminal Nudix hydrolase domain.
3.2. Effect of enzyme concentration and incubation period
The results of crystallographic studies (see below) necessi-
tated an examination of the effects of enzyme concentration
and the incubation period on activity. Enzyme activity was
previously measured, as is usually performed, at compara-
tively low protein concentration and short incubation periods.
Crystallization, however, involves high protein concentrations
and long incubation periods. Varying amounts (0–200 mM) of
research papers
Acta Cryst. (2017). D73, 349–364 Arif et al. � Mycobacterium smegmatis MutT1 355
Figure 3Arrangement of molecules and generation of ligand-binding sites. (a) Head-to-tail arrangement of MsMutT1 molecules that brings domain 1 and transdomain 2 into periodic proximity, leading to the generation of ligand-binding sites, as observed in most of the crystals. (b) Disposition of neighbouringmolecules in crystal 2. (c) Various ligand-binding sites in domain 1 and at the interface between domain 1 and trans domain 2. Ligands from differentcrystals are mapped approximately on the protein structure involving domain 1 and trans domain 2 of crystal 5.
magnesium were added in the crystallization experiments.
Crystals grown from solutions containing no externally added
magnesium were of great use in defining the different binding
sites and they probably represented intermediate stages in the
reaction process. Therefore, it was important to explore the
reaction over long periods in solutions containing a high
protein concentration and no externally added magnesium as
well.
MsMutT1 at a concentration of approximately 22 mM
(0.8 mg ml�1) converted 8-oxo-dGTP completely to 8-oxo-
dGMP after incubation for approximately 12 h in the presence
of 200 mM MgCl2 (Fig. 1c). A similar result was obtained
when 8-oxo-GTP was used instead of 8-oxo-dGTP. When the
experiment involving 8-oxo-dGTP was repeated with an
incubation period of 3 h, 8-oxo-dGDP and 8-oxo-dGMP were
obtained, with the former as the major product. A similar
result was obtained when 42.56 mM (0.8 mg ml�1) MsMutT1-
NTD was incubated for 12 h with 8-oxo-dGTP (Fig. 1c). An
identical experiment involving MsMutT1-CTD did not yield
any product. It would thus appear that in the presence of
magnesium the enzyme converts 8-oxoguanine trinucleotides
to the corresponding dinucleotides at low protein concentra-
tions and short periods of incubation. With increasing
concentrations and times of incubation, the mononucleotides
also begin to appear. When the time of incubation is further
increased, the trinucleotide is converted completely into the
mononucleotide. Thus, it seems that the trinucleotide is
converted first to the dinucleotide, which is further converted
to the mononucleotide. However, a direct conversion of
trinucleotide to mononucleotide simultaneously cannot be
ruled out. Furthermore, MsMutT1-NTD, but not MsMutT1-
CTD, is by itself capable of converting an NTP into an NDP as
well as an NMP, although sluggishly.
Crystallization experiments were carried out at a still higher
concentration of the enzyme, in most cases with no externally
added magnesium. Trace amounts of magnesium probably
copurified with the protein during preparation. Therefore,
most of the crystals were grown in conditions in which reac-
tions could presumably take place very slowly or partially on
account of the very limited availability of magnesium. The
formation of 8-oxo-GMP/8-oxo-dGMP along with 8-oxo-
GDP/8-oxo-dGDP was observed when appropriate solutions
used for co-crystallization of MsMutT1 with 8-oxo-GTP/
8-oxo-dGTP were analysed by HPLC (Fig. 1d).
3.3. Molecular structure: overall features
The structures of apo MsMutT1 and eight of its complexes,
determined at resolutions ranging from 1.85 to 1.10 A,
belonged to six somewhat related crystal forms (Tables 1 and
2). Four of the crystals have two molecules with nearly iden-
tical structures in the asymmetric unit. Thus, the structures
provide 13 independent copies of the MsMutT1 molecule. No
electron density was observed for the 20–22 N-terminal resi-
dues in all of the structures. Residues 35–46 are also ill-defined
to different extents in different molecules. Thus, the refined
models of the molecule consist of residues 289–302 of the 322
residues in the sequence.
All 13 copies of the molecule have essentially the same
structure, with a N-terminal domain (NTD) extending to
residue 157 and a C-terminal domain (CTD; residues 163–322)
connected by a linker region (residues 158–162) (Fig. 2a). The
NTD, which is made up of three �-helices (�1–�3) and six
�-strands (�1–�6), has an �/�/� sandwich fold (Fig. 2b). The
three long strands (�1, �4 and �5) form a curled mixed
�-sheet, which is sandwiched between a short �-helix (�3) on
one side and a long �-helix (�1) on the other side. This
corresponds to the main frame of the Nudix fold. The three
short antiparallel �-strands �2, �3 and �6 together with the
helix �2 form an additional lobe attached to this frame.
Residues 66–88 involving a short strand (�30), a loop, �1 and
research papers
356 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364
Figure 4Aggregation of MsMutT1 in solution. Scattered intensity versushydrodynamic radius in DLS experiments at protein concentrations of(a) 1 mg ml�1 and (b) 7 mg ml�1 in the crystallization buffer. (c) Guinierplot of a SAXS experiment at a protein concentration of 18 mg ml�1 inthe crystallization buffer. A departure from linearity indicates aggrega-tion (Putnam et al., 2007).
another loop (SLHL) constitute the highly conserved
23-residue MutT signature sequence or Nudix motif. The
CTD, which is primarily made up of a six-stranded �-sheet
(�1–�6) flanked by two helices on one side (�2 and �3) and
another set of two helices (�5 and �6) on the other side, has an
�/� architecture (Fig. 2c) of the type found in the members of
the phosphoglycerate mutase family (histidine phosphatase
superfamily, branch 1). The structure contains two more short
helices, namely �1 and �4. The loop connecting �1 and �1
harbours the conserved RHG (arginine–histidine–glycine)
motif, which is a distinctive feature of the histidine phospha-
tase superfamily.
Not only the geometries of the two domains but also their
mutual orientation remains nearly the same in all of the
crystals, presumably on account of the strong interaction
between the two domains in the molecule. The interface
between the NTD and CTD involves the burial of 1201.7 A2 of
surface area, which is nearly equally distributed between the
two domains, of which 714.5 A2 is nonpolar. In addition, the
two domains are also connected by eight distinct hydrogen
bonds.
3.4. Intermolecular interactions and generation of bindingsites
Among the crystals studied, all except crystal 2 exhibit
nearly the same type of aggregation of molecules. The central,
common feature of this aggregation pattern is a head-to-tail
arrangement around a 21 screw axis, in which domain 1 of one
molecule is brought into close periodic proximity to domain 2
of a neighbouring molecule (trans domain 2; Fig. 3a). The
packing of molecules in crystal 2 is very different and involves
proximity of the same domains from neighbouring molecules
(Fig. 3b). Domain 1 and trans domain 2, as they occur in the
head-to-tail arrangement, probably form the predominant
functional unit of the enzyme. The results of dynamic light-
scattering experiments on MsMutT1 at a protein concentra-
tion of 1 mg ml�1 (Fig. 4a) indicate a dimer (radius 39 A and
molecular weight 75.3 kDa) as the predominant species. This
observation is compatible with the occurrence of dimeric units
in a head-to-tail arrangement. The experiments were repeated
at protein concentrations of 4 and 7 mg ml�1. Both experi-
ments lead to an indication of the same pattern of higher order
aggregation (Fig. 4b). The pattern involves two broad peaks.
The first peak, with an approximate radius of 76 A, roughly
corresponds to a decamer. The second broadest peak, with an
approximate radius of 1330 A, corresponds to aggregates
involving a few thousand molecules. The occurrence of two
peaks cannot be readily explained in terms of the structure.
However, the results of light-scattering experiments indicate
that the protein forms larger aggregates, presumably involving
dimers, at higher concentrations. This observation is in
consonance with the aggregation pattern observed in the
crystal structure. SAXS analysis
of the protein was also carried
out. Interpretable intensities
could be obtained only at a very
high concentration (18 mg ml�1)
of the protein. The results of the
SAXS analysis again indicate
aggregation (Fig. 4c).
In the head-to-tail arrange-
ment in crystals of the apoenzyme
(crystal 1), the interaction
between domain 1 and trans
domain 2 involves the burial of
1422 A2 of surface area, of which
706 A2 is hydrophobic. The
extent of burial is comparable to
the burial of surface area
observed in the association of
subunits in some multimeric
proteins (Arif & Vijayan, 2012;
Jones et al., 2000; Jones &
Thornton, 1996). The interaction
also encompasses 12 hydrogen
bonds involving the main-chain
and side-chain atoms of Arg57,
Tyr58, Asp59, Met140, Asp141,
Leu143, Gln144, Tyr145 and
Arg149 of domain 1 and those of
Arg169, Gly174, Arg175, Arg176,
Lys273, Glu280, Asn295, Arg296
and Lys297 of trans domain 2.
research papers
Acta Cryst. (2017). D73, 349–364 Arif et al. � Mycobacterium smegmatis MutT1 357
Figure 5Simulated-annealing Fo� Fc OMIT maps of representative ligands observed at sites A, B and C in differentcrystals. The OMIT maps are contoured at the 3� level. The crystals in which a particular ligand is observedare given in parentheses.
research papers
358 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364
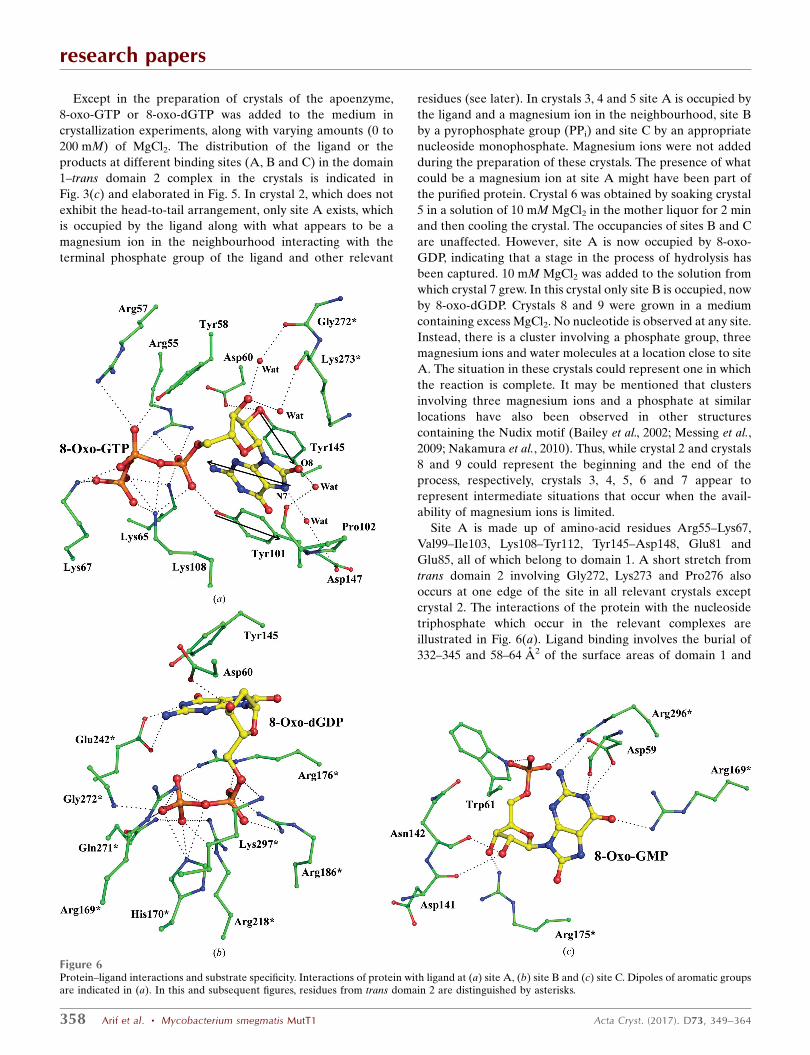
Except in the preparation of crystals of the apoenzyme,
8-oxo-GTP or 8-oxo-dGTP was added to the medium in
crystallization experiments, along with varying amounts (0 to
200 mM) of MgCl2. The distribution of the ligand or the
products at different binding sites (A, B and C) in the domain
1–trans domain 2 complex in the crystals is indicated in
Fig. 3(c) and elaborated in Fig. 5. In crystal 2, which does not
exhibit the head-to-tail arrangement, only site A exists, which
is occupied by the ligand along with what appears to be a
magnesium ion in the neighbourhood interacting with the
terminal phosphate group of the ligand and other relevant
residues (see later). In crystals 3, 4 and 5 site A is occupied by
the ligand and a magnesium ion in the neighbourhood, site B
by a pyrophosphate group (PPi) and site C by an appropriate
nucleoside monophosphate. Magnesium ions were not added
during the preparation of these crystals. The presence of what
could be a magnesium ion at site A might have been part of
the purified protein. Crystal 6 was obtained by soaking crystal
5 in a solution of 10 mM MgCl2 in the mother liquor for 2 min
and then cooling the crystal. The occupancies of sites B and C
are unaffected. However, site A is now occupied by 8-oxo-
GDP, indicating that a stage in the process of hydrolysis has
been captured. 10 mM MgCl2 was added to the solution from
which crystal 7 grew. In this crystal only site B is occupied, now
by 8-oxo-dGDP. Crystals 8 and 9 were grown in a medium
containing excess MgCl2. No nucleotide is observed at any site.
Instead, there is a cluster involving a phosphate group, three
magnesium ions and water molecules at a location close to site
A. The situation in these crystals could represent one in which
the reaction is complete. It may be mentioned that clusters
involving three magnesium ions and a phosphate at similar
locations have also been observed in other structures
containing the Nudix motif (Bailey et al., 2002; Messing et al.,
2009; Nakamura et al., 2010). Thus, while crystal 2 and crystals
8 and 9 could represent the beginning and the end of the
process, respectively, crystals 3, 4, 5, 6 and 7 appear to
represent intermediate situations that occur when the avail-
ability of magnesium ions is limited.
Site A is made up of amino-acid residues Arg55–Lys67,
Val99–Ile103, Lys108–Tyr112, Tyr145–Asp148, Glu81 and
Glu85, all of which belong to domain 1. A short stretch from
trans domain 2 involving Gly272, Lys273 and Pro276 also
occurs at one edge of the site in all relevant crystals except
crystal 2. The interactions of the protein with the nucleoside
triphosphate which occur in the relevant complexes are
illustrated in Fig. 6(a). Ligand binding involves the burial of
332–345 and 58–64 A2 of the surface areas of domain 1 and
Figure 6Protein–ligand interactions and substrate specificity. Interactions of protein with ligand at (a) site A, (b) site B and (c) site C. Dipoles of aromatic groupsare indicated in (a). In this and subsequent figures, residues from trans domain 2 are distinguished by asterisks.

trans domain 2, respectively. The dominant interactions are
between the triphosphate tail and the basic side chains of
Arg55, Arg57, Lys65, Lys67 and Lys108. The side-chain
hydroxyls of Tyr58 and Tyr101 also interact with the tail. O20
of the sugar interacts with the side chains of Asp60 and Tyr145
when the ligand is 8-oxo-GTP. Only Asp148 interacts with the
base through hydrogen bonds. The base is, however, sand-
wiched between the aromatic side chains of Tyr101 and
Tyr145. Residues from trans domain 2 only make water-
mediated interactions with the NTP or NDP ligand. These
water bridges do not exist in crystal 2, which does not exhibit
domain 1–trans domain 2 interactions. This difference in
molecular association does not affect the geometry of site A in
crystal 2, except for a slight shift in the location of the ligand.
However, in crystal 2 the 124–128 stretch, particularly Glu127,
moves close to the ligand. This movement appears to have
functional implications (see below).
The charge distribution in and hence the electrostatic
potential around the base appears to be important in discri-
minating between 8-oxo-G and G for binding at site A. The
orientation of dipoles, of the type involved in the case of
human OGG1 (David et al., 2007), could be central in this
discrimination. As illustrated in Fig. 6(a), the orientation of
the dipoles in 8-oxo-G and the flanking tyrosine side chains
are such as to promote strong interaction of the base with the
side chains. The interaction is weakened when the base is G, in
which the direction of the dipole is inclined with respect to
that in 8-oxo-G. The substitution of an O atom at C8 promotes
the protonation of N7, which facilitates the formation of a
water bridge with Asp147. Furthermore, the observed water
bridge between O8 and Pro102 O is possible only when 8-oxo-
G, instead of G, is the ligand.
The bulk of the residues which constitute site B are from
trans domain 2. In particular, residues 169–176, which form
part of a long loop, and the short loop 271–273 are involved in
interactions with PPi or the diphosphate tail of 8-oxo-dGDP
(Fig. 6b). These stretches are rich in basic residues. The indi-
vidual residues His170, Arg186 and Arg218 also interact with
the phosphates. The planar guanidinium group of Arg176
makes a stacking interaction with the nitrogen base of 8-oxo-
dGDP. Lys297 and Glu242 interact with the sugar and the
base, respectively. Interestingly, the short 144–146 stretch in
domain 1, which is also part of site A, is close to the nucleoside
at site B. Tyr145 of this stretch stacks against the base. Another
residue from domain 1, Asp60, forms a hydrogen bond to a
sugar hydroxyl group. Binding of NDP involves the burial of
133 and 200 A2 of the surface area of domain 1 and trans
domain 2, respectively.
Site C, which spans domain 1 and trans domain 2, overlaps
to an extent with sites A and B, particularly the latter. The
electron densities of the ligand at this site are not as good as
those at the other two. The atoms also exhibit high tempera-
ture factors. However, the densities are good enough to place
the ligand unambiguously. The NMP ligand at the site buries
146–170 and 168–177 A2 of the surface areas of domain 1 and
trans domain 2, respectively. The 141–144 stretch of domain 1
is at the boundary of all three sites. Two residues in this stretch
interact with the sugar at site C (Fig. 6c). Arg175 of trans
domain 2, a residue involved in site B, also interacts with the
sugar. Trp61 of domain 1 and Arg296 of trans domain 2 are
involved in interactions with the phosphate group at site C.
Asp59 of domain 1, which is at the boundary of site A, and
Arg169 of trans domain 2, which forms part of site B, interact
with the base at site C. PPi at site B and the nucleoside
research papers
Acta Cryst. (2017). D73, 349–364 Arif et al. � Mycobacterium smegmatis MutT1 359
Figure 7Ligand-induced intermolecular and intramolecular movements. (a) Movement of trans domain 2 with respect to domain 1 owing to the binding ofligands. (b) Ligand-induced movements within domain 1.

monophosphate at site C can coexist. However, the sugar
component of the ligand at site B would have serious steric
clashes with the base component of the ligand at site C. Thus,
site C is unoccupied in crystal 7, in which site B is occupied by
a nucleoside diphosphate.
3.5. Ligand-induced movements
Although the overall molecular structure remains the same
in all crystals, specific movements related to ligand binding are
discernible on close examination. As illustrated in Fig. 7(a),
trans domain 2 rotates by 9.6–11.9� in liganded molecules
(crystals 3, 4, 5, 6 and 7) with respect to its position in the
crystals of the unliganded protein
(crystal 1). When the reaction is
complete, as in crystals 8 and 9,
the rotation is reversed, but not
fully. The angle of rotation of
trans domain 2 in these crystals
with respect to the position in
crystal 1 is 3.4–4.6�.
Movements take place within
the domain as well. For instance,
substantial movements, especially
of aromatic residues, take place at
site A on ligand binding (Fig. 7b).
Tyr101 and Tyr145, the side
chains of which stack on either
side of the base, understandably
move towards the base in the
complexes. Tyr101, in addition,
forms a hydrogen bond to a
phosphate group. Tyr58 and
Lys108 also move towards the
ligand to facilitate hydrogen
bonding. In fact, the peptide
stretches to which these residues
belong move, as a whole, towards
the ligand in the complexes. Here
again, these residues and the
peptide stretches that carry them
do not entirely move back to their
position in the crystals of the
apoenzyme (crystal 1) on
completion of the reaction (crys-
tals 8 and 9).
3.6. Comparison with similarenzymes
The NTD of MsMutT1 has low
sequence identities of 18.3 and
13.2% to EcMutT and HsMTH1,
respectively. The three, however,
have remarkably similar core
structures, with r.m.s.d.s of 1.79
and 1.76 A in C� positions
when the structures of EcMutT (Nakamura et al., 2010) and
HsMTH1 (Svensson et al., 2011) are superposed on that of
MsMutT1-NTD. The structures also exhibit significant differ-
ences in the loops and other structural elements surrounding
the active sites (Fig. 8a). Unlike MsMutT1-NTD, insertions in
one such loop in EcMutT and HsMTH1 and an additional
�-hairpin in HsMTH1 help in the formation of closed pockets
by covering the active site. The site in the MsMutT1-NTD
is relatively open. Furthermore, in the nucleotide complexes
the ligands are located deeper in the pocket in EcMutT and
HsMTH1 compared with that in MsMutT1-NTD (Fig. 8b).
Therefore, there could be hitherto unappreciated differences
in substrate specificities. The core of the NTD of the
research papers
360 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364
Figure 8Structure of and ligand location in MsMutT1-NTD and its homologues. (a) Structures of MsMutT1-NTD,E. coli MutT (EcMutT), human MTH1 (HsMTH1) and Bifidobacterium adolescentis MutT1 (BaMutT1).Longer loops in all of these, except MsMutT1-NTD, and an additional �-hairpin in HsMTH1 (blue), shownin boxes, cover their active sites. (b) An overlay of ligand-bound structures of MsMutT1 (NTD with ligand),EcMutT (PDB entry 3a6u; Nakamura et al., 2010) and HsMTH1 (PDB entry 3zr0; Svensson et al., 2011),showing the locations of the ligands.

putative MutT1 of B. adolescentis (BaMutT1-NTD), the
structure of which is available in the PDB (PDB entry 3fjy),
has a fold similar to that of MsMutT1-NTD. Here again, a
larger insertion in another loop seems to provide a lid for
what appears by analogy to be the ligand-binding site. A
description of the structure of this protein has yet to be
published.
CTD is structurally similar to several of the branch 1
members of the histidine phosphatase superfamily. In parti-
cular, it is structurally close to the SixAs from E. coli and
Nakamurella multipartitia, the putative SixA from Agro-
bacterium tumefaciens and the alr0221 protein from Nostoc sp.
For instance, the structures of the SixAs from E. coli and
N. multipartitia superpose on the structure of MsMutT1-CTD
with r.m.s.d.s of 2.01 and 1.80 A, respectively. All of these,
along with MsMutT1-CTD, represent structures with the
minimal core of branch 1 of the histidine phosphatase super-
family and differ from the rest of the branch 1 members and all
of the branch 2 members of the histidine phosphatase super-
family in that they lack an extra subdomain that is found in the
latter (Fig. 9). This extra subdomain seems to cover the active
site of these enzymes and helps in the formation of a deeper
pocket. The active sites of SixA and MsMutT1-CTD are
relatively shallow and open owing to the absence of this extra
domain. However, the mycobacterial enzyme exhibits some
similarity to both groups. His170 in MsMutT1-CTD is found at
a homologous position in both of the groups. Asp183 in
MsMutT1-CTD occupies the same spatial location as the
catalytic aspartate residue in the homologues with a shallow
pocket. Glu242, on the other hand, occupies a location similar
to the catalytic glutamate residue in the structural homologues
with a deeper active-site pocket. Both of these residues have
been proposed to act as a proton donor in the phospho-
histidine-mediated phosphatase activity of these structural
homologues. However, the orientation of the nucleoside
diphosphate at site B is not such as to enable catalysis invol-
ving these residues.
MsMutT1 is an example of a protein in which a Nudix
hydrolase domain occurs along with a histidine phosphatase
domain. The crystal structure of only one other protein
containing histidine phosphatase and Nudix hydrolase
domains has so far been determined (PDB entry 3fjy). It may
be mentioned that the histidine phosphatase domain often
occurs along with other domains (Rigden, 2008). The other
domains which are known to occur along with the histidine
phosphatase domain include kinase domains (for example
phosphofructokinase, inositol hexakisphosphate kinase and
Vip1 kinase domains), the 2H phosphoesterase domain,
reductase or dehydrogenase domains, carbohydrate-binding
modules, ankyrin repeats, SH3 (Src homology 3) domains
and the UBASH (ubiquitin-associated and SH3 domain-
containing) domain. In the light of the roles suggested for the
histidine phosphatase domain in many proteins, a role for it
beyond catalysis cannot be ruled out in MsMutT1.
3.7. Insights into the mechanism of action
The biochemical and crystallographic results presented
above lead to a plausible proposal for the mechanism of action
of this unusual enzyme. In most of the crystals obtained under
conditions with a limited supply of magnesium ions, a
magnesium ion coordinated by the main chain of Lys65, the
side chain of Glu85 and water molecules is close to P� of the
NTP (Fig. 10a). The situation is analogous to that in an NTP
complex of EcMutT (Lin et al., 1997), except that the ion in
this case is close to P� and the reaction product of the E. coli
enzyme is a nucleoside monophosphate. One of the water
molecules that coordinate to the magnesium is also close to
the �-phosphate group and could be involved in nucleophilic
attack on P� in an associative mechanism. Glu81, the side
research papers
Acta Cryst. (2017). D73, 349–364 Arif et al. � Mycobacterium smegmatis MutT1 361
Figure 9Comparison of MsMutT1-CTD (the CTD of apo MsMutT1) with its structural homologues E. coli SixA (EcSixA; PDB entry 1ujc; Hamada et al., 2005)and Bacillus subtilis PhoE (BsPhoE; PDB entry 1h2e; Rigden et al., 2003), both of which belong to branch 1 of the histidine phosphatase superfamily.MsMutT1-CTD and EcSixA lack an extra subdomain (in blue) that covers the active site in the BsPhoE structure. The conserved histidine residueindicating the location of the active site is shown in pink.

chain of which is hydrogen-bonded to the water molecule,
could act as a general base to deprotonate the attacking water
ligand. The side chain of Arg80 appears to be involved in
maintaining the correct orientation of the glutamyl side chain.
The side chain of Arg55 is positioned appropriately to
protonate the leaving O atom (O3B) and thus might serve as a
general acid. Lys65, Lys67 and Lys108, which are hydrogen-
bonded to the phosphoryl O atoms of NTP, help to hold the
substrate in the appropriate position. Crystal 6 represents a
situation in which the conversion of 8-oxo-GTP to 8-oxo-GDP
is complete but the product is still located at site A.
Long exposure to magnesium ions, as in crystal 7, appears to
have helped to take the reaction to the next step and trans-
locate the NDP from site A to site B. Energetics appear to
favour this translocation. Energy calculations indicate that the
free energies of interaction of the NDP at site A and site B to
be �65.3 and �70.4 kJ mol�1, respectively. From the pictures
at the two extreme positions, it is difficult to work out the path
between the two. However, from their location at site A and
site B in the relevant crystals, Tyr58, Tyr145, Arg169, Gly272
and Lys273 appear to be involved in the translocation of the
NDP from site A to site B.
The next issue to be addressed is the mechanism of
conversion of 8-oxo-GTP or 8-oxo-dGTP to the corresponding
nucleoside monophosphates. The available crystal structures
with a head-to-tail arrangement do not provide an explanation
for the hydrolysis of the NTP to a nucleoside monophosphate
and a pyrophosphate. However, the structure of MsMutT1 in
crystal 2 provides clues as to how catalysis might proceed.
Notably, Glu127 in this crystal moves closer to the substrate
compared with its position in the other crystals. Both of the
side-chain O atoms of this residue, in addition to the main-
chain O atom of Lys65, a side-chain O atom of Glu85 and
water molecules, coordinate to a magnesium ion (Fig. 10b).
One of these is hydrogen-bonded to a water molecule, which is
in a position to attack P�, with the side chain of Glu127 acting
as a general base to activate the water molecule. Lys108 is
appropriately positioned to donate a proton to the leaving O
atom (O3A) and it appears to facilitate the departure of the
NMP product. Lys67, which is hydrogen-bonded to the
�-phosphate and �-phosphate, might facilitate the departure
of pyrophosphate. The proposed mechanism applies equally
well to the conversion of NDP to NMP when the enzyme has a
conformation similar to that found in crystal 2.
The available results lead to a proposal that the arrange-
ment involving the characteristic juxtaposition of domain 1
and trans domain 2, which is observed in all but one crystal,
and that which does not involve such juxtaposition (crystal 2)
contribute to the catalytic machinery of the enzyme. One
could lead to NDP and PO4 as the products, while the other
could help to produce NMP and PPi. The two could be in
dynamic equilibrium in the cellular milieu with the arrange-
ment which brings together domain 1 and trans domain 2 as
the predominant species. As indicated earlier, energy calcu-
lations and the location of relevant residues suggest the
possibility of translocation of NDP from site A to site B. The
NMP in solution produced by the enzyme in the structural
state found in crystal 2 could have been picked up at site C by
the protein in the dominant arrangement found in all other
crystals.
The obvious structural role of the second domain is in
the generation of sites B and C through the head-to-tail
arrangement of MsMutT1 molecules. It also provides a lid to
the relatively open pocket at site A, thus forming a cave-like
research papers
362 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364
Figure 10Enzyme-catalysis schemes. Hydrolysis of (a) 8-oxo-GTP to 8-oxo-GDP and a phosphate and (b) of 8-oxo-dGTP to 8-oxo-dGMP and a pyrophosphate.Arrows in red indicate the activation of a water nucleophile with the aid of a general base followed by its attack on the phosphate centres. The arrows inpurple direct the leaving O atom towards proton donors (general acids).

structure. By doing so, it might provide additional stability to
the substrate bound at site A, which would otherwise have its
sugar exposed to the solvent, and thus might aid in efficient
catalysis. Thus, trans domain 2 might, in part, play a role
similar to that played by the extra loop produced by the 3–4-
residue insertions in EcMutT and HsMTH1 and the additional
�-hairpin in HsMTH1 (Mishima et al., 2004). Additionally,
trans domain 2 might be involved in promoting product
release via its interactions with the loops surrounding the
active site. Trigger events, mediated by structural elements
that are not involved in direct interactions with the substrate
or product, have been proposed to facilitate substrate binding
(Ge et al., 2013) and product release (Bailey et al., 2002) in
other Nudix hydrolases. However, considering the many roles
that the histidine phosphatase domain assumes in biological
systems, it is unlikely that its role in MsMutT1 is only as a
structural component of the catalytic machinery. Its additional
role(s), perhaps not necessarily involving catalysis, will
hopefully become clear through further studies of the
enzyme.
The above observations on MsMutT1 are likely to be
relevant to MtMutT1 as well. The enzymes from the two
mycobacterial species exhibit a sequence identity of 60.8%.
Perhaps more importantly, 27 of the 32 residues involved in
interactions with the ligands are identical in the two enzymes.
Two of the remaining residues interact with a nucleotide
through main-chain atoms and a third is only involved in a
water bridge. There are only two differences of consequence,
one of which involves a conservative substitution (Arg to Lys).
Neither of them is involved in the proposed mechanism of
action.
4. Conclusions
MsMutT1, which is made up of a Nudix hydrolase domain
(domain 1) and a histidine phosphatase domain (domain 2),
efficiently converts 8-oxo-GTP and 8-oxo-dGTP to the
respective nucleoside diphosphates. At higher concentrations
the enzyme can also catalyse the conversion of the NTPs to
NMPs and pyrophosphate. The same reaction can be cata-
lysed, albeit much less efficiently, by domain 1 alone, but not
by domain 2. Thus, the role of domain 2 in in vitro experiments
is one of speeding up the reaction.
In eight of the nine crystals studied, one of the apoenzyme
and the others grown under different conditions in the
presence of 8-oxo-GTP and 8-oxo-dGTP, the molecules are
arranged in a head-to-tail fashion, with domain 1 of one
molecule and domain 2 of a neighbouring molecule (trans
domain 2) in periodic proximity. Domain 1 and trans domain 2
together form a functional unit and contain three binding sites.
Site A, which binds NTPs, is almost exclusively made up of
residues from domain 1. Site B, for NDPs or a pyrophosphate,
and site C, for NMPs, are at the interface between domain 1
and trans domain 2, in an unusual case in which binding sites
are produced by intermolecular interactions. The disposition
of residues, a magnesium ion and a water molecule at site A,
and the location of NTP and NDP at the site, lead to a plau-
sible proposal for the mechanism of hydrolysis of 8-oxo-GTP
and 8-oxo-dGTP to the respective NDPs.
The structure of the active site in the lone crystal in which
domain 1 and trans domain 2 are not in proximity is very
similar to that in which they are. In the former, however, a
neighbouring peptide stretch bearing a glutamyl residue
moves close to the ligand. This residue, along with a water
molecule, appears to facilitate the hydrolysis of the NTP to
NMP and pyrophosphate. The more frequently observed
arrangement which leads to the production of NDP and
phosphate and the arrangement that facilitates the hydrolysis
into NMP and pyrophosphate could be in dynamic equili-
brium in the cellular milieu. This proposal is in agreement with
the results of biochemical experiments on MsMutT1.
Acknowledgements
X-ray data were collected at the X-ray facility for Protein
X-ray Structure Determination and Protein Design at this
institute, supported by the Department of Science and Tech-
nology (DST), and at ESRF, Grenoble through an arrange-
ment made by the Department of Biotechnology (DBT). The
Graphics Facility supported by the DBT was used to perform
some of the computations. The DLS and SAXS measurements
were carried out in the laboratory of Dr R. Sankarnarayanan
at the Center for Cellular and Molecular Biology, Hyderabad.
UV is a J. C. Bose fellow of the DST. MV is Albert Einstein
Professor of the Indian National Science Academy. This work
was supported by a research grant from the DBT.
References
Arif, S. M., Patil, A. G., Varshney, U. & Vijayan, M. (2012). Acta Cryst.F68, 1214–1216.
Arif, S. M. & Vijayan, M. (2012). Methods Mol. Biol. 922, 23–35.Arora, A., Chandra, N. R., Das, A., Gopal, B., Mande, S. C., Prakash,
B., Ramachandran, R., Sankaranarayanan, R., Sekar, K., Suguna,K., Tyagi, A. K. & Vijayan, M. (2011). Tuberculosis, 91, 456–468.
Au, K. G., Clark, S., Miller, J. H. & Modrich, P. (1989). Proc. NatlAcad. Sci. USA, 86, 8877–8881.
Bailey, S., Sedelnikova, S. E., Blackburn, G. M., Abdelghany, H. M.,Baker, P. J., McLennan, A. G. & Rafferty, J. B. (2002). Structure, 10,589–600.
Battye, T. G. G., Kontogiannis, L., Johnson, O., Powell, H. R. & Leslie,A. G. W. (2011). Acta Cryst. D67, 271–281.
Bessman, M. J., Frick, D. N. & O’Handley, S. F. (1996). J. Biol. Chem.271, 25059–25062.
Brunger, A. T., Adams, P. D., Clore, G. M., DeLano, W. L., Gros, P.,Grosse-Kunstleve, R. W., Jiang, J.-S., Kuszewski, J., Nilges, M.,Pannu, N. S., Read, R. J., Rice, L. M., Simonson, T. & Warren, G. L.(1998). Acta Cryst. D54, 905–921.
Cohen, G. H. (1997). J. Appl. Cryst. 30, 1160–1161.Cole, S. T. et al. (1998). Nature (London), 393, 537–544.Cooke, M. S., Evans, M. D., Dizdaroglu, M. & Lunec, J. (2003).
FASEB J. 17, 1195–1214.David, S. S., O’Shea, V. L. & Kundu, S. (2007). Nature (London), 447,
941–950.DeLano, W. L. (2002). PyMOL. http://www.pymol.org.Dos Vultos, T., Blazquez, J., Rauzier, J., Matic, I. & Gicquel, B. (2006).
J. Bacteriol. 188, 3159–3161.Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. (2010). Acta
Cryst. D66, 486–501.Evans, P. (2006). Acta Cryst. D62, 72–82.
research papers
Acta Cryst. (2017). D73, 349–364 Arif et al. � Mycobacterium smegmatis MutT1 363

Ge, H., Chen, X., Yang, W., Niu, L. & Teng, M. (2013). Biochem.Biophys. Res. Commun. 432, 16–21.
Hamada, K., Kato, M., Shimizu, T., Ihara, K., Mizuno, T. &Hakoshima, T. (2005). Genes Cells, 10, 1–11.
Hammersley, A. P. (1997). FIT2D: An Introduction and Overview.ESRF Internal Report ESRF97HA02T. ESRF, Grenoble, France.http://www.esrf.eu/computing/scientific/FIT2D/FIT2D_INTRO/fit2d.html.
Hayakawa, H., Hofer, A., Thelander, L., Kitajima, S., Cai, Y., Oshiro,S., Yakushiji, H., Nakabeppu, Y., Kuwano, M. & Sekiguchi, M.(1999). Biochemistry, 38, 3610–3614.
Heinig, M. & Frishman, D. (2004). Nucleic Acids Res. 32, W500–W502.
Hornak, V., Abel, R., Okur, A., Strockbine, B., Roitberg, A. &Simmerling, C. (2006). Proteins, 65, 712–725.
Hubbard, S. J. & Thornton, J. M. (1996). NACCESS: A ComputerProgram for Calculating Accessibilities. Department of Biochem-istry and Molecular Biology, University College London.
Ishibashi, T., Hayakawa, H. & Sekiguchi, M. (2003). EMBO Rep. 4,479–483.
Ito, R., Hayakawa, H., Sekiguchi, M. & Ishibashi, T. (2005).Biochemistry, 44, 6670–6674.
Jain, R., Kumar, P. & Varshney, U. (2007). DNA Repair (Amst.), 6,1774–1785.
Jones, S., Marin, A. & Thornton, J. M. (2000). Protein Eng. 13, 77–82.
Jones, S. & Thornton, J. M. (1996). Proc. Natl Acad. Sci. USA, 93,13–20.
Kamiya, H., Suzuki, A., Yamaguchi, Y., Handa, H. & Harashima, H.(2009). Free Radic. Biol. Med. 46, 1703–1707.
Konarev, P. V., Petoukhov, M. V., Volkov, V. V. & Svergun, D. I.(2006). J. Appl. Cryst. 39, 277–286.
Konarev, P. V., Volkov, V. V., Sokolova, A. V., Koch, M. H. J. &Svergun, D. I. (2003). J. Appl. Cryst. 36, 1277–1282.
Kuchino, Y., Mori, F., Kasai, H., Inoue, H., Iwai, S., Miura, K.,Ohtsuka, E. & Nishimura, S. (1987). Nature (London), 327, 77–79.
Kurthkoti, K., Srinath, T., Kumar, P., Malshetty, V. S., Sang, P. B., Jain,R., Manjunath, R. & Varshney, U. (2010). Microbiology, 156, 88–93.
Kurthkoti, K. & Varshney, U. (2011). Tuberculosis, 91, 533–543.Laskowski, R. A., Moss, D. S. & Thornton, J. M. (1993). J. Mol. Biol.
231, 1049–1067.Lin, J., Abeygunawardana, C., Frick, D. N., Bessman, M. J. & Mildvan,
A. S. (1997). Biochemistry, 36, 1199–1211.Maki, H. & Sekiguchi, M. (1992). Nature (London), 355, 273–275.Matthews, B. W. (1968). J. Mol. Biol. 33, 491–497.McCoy, A. J., Grosse-Kunstleve, R. W., Adams, P. D., Winn, M. D.,
Storoni, L. C. & Read, R. J. (2007). J. Appl. Cryst. 40, 658–674.
Messing, S. A., Gabelli, S. B., Liu, Q., Celesnik, H., Belasco, J. G.,Pineiro, S. A. & Amzel, L. M. (2009). Structure, 17, 472–481.
Michaels, M. L. & Miller, J. H. (1992). J. Bacteriol. 174, 6321–6325.Mishima, M., Sakai, Y., Itoh, N., Kamiya, H., Furuichi, M., Takahashi,
M., Yamagata, Y., Iwai, S., Nakabeppu, Y. & Shirakawa, M. (2004).J. Biol. Chem. 279, 33806–33815.
Moreland, N. J., Charlier, C., Dingley, A. J., Baker, E. N. & Lott, J. S.(2009). Biochemistry, 48, 699–708.
Moriya, M., Ou, C., Bodepudi, V., Johnson, F., Takeshita, M. &Grollman, A. P. (1991). Mutat. Res. 254, 281–288.
Morris, G. M., Huey, R., Lindstrom, W., Sanner, M. F., Belew, R. K.,Goodsell, D. S. & Olson, A. J. (2009). J. Comput. Chem. 30, 2785–2791.
Murillo, A. C. et al. (2007). Infect. Disord. Drug Targets, 7, 127–139.Murshudov, G. N., Skubak, P., Lebedev, A. A., Pannu, N. S., Steiner,
R. A., Nicholls, R. A., Winn, M. D., Long, F. & Vagin, A. A. (2011).Acta Cryst. D67, 355–367.
Nakabeppu, Y., Kajitani, K., Sakamoto, K., Yamaguchi, H. &Tsuchimoto, D. (2006). DNA Repair (Amst.), 5, 761–772.
Nakamura, T., Meshitsuka, S., Kitagawa, S., Abe, N., Yamada, J.,Ishino, T., Nakano, H., Tsuzuki, T., Doi, T., Kobayashi, Y., Fujii, S.,Sekiguchi, M. & Yamagata, Y. (2010). J. Biol. Chem. 285, 444–452.
Patil, A. G., Sang, P. B., Govindan, A. & Varshney, U. (2013). J. Biol.Chem. 288, 11252–11262.
Provencher, S. W. (1982). Comput. Phys. Commun. 27, 229–242.Pursell, Z. F., McDonald, J. T., Mathews, C. K. & Kunkel, T. A. (2008).
Nucleic Acids Res. 36, 2174–2181.Putnam, C. D., Hammel, M., Hura, G. L. & Tainer, J. A. (2007). Q.
Rev. Biophys. 40, 191–285.Rigden, D. J. (2008). Biochem. J. 409, 333–348.Rigden, D. J., Littlejohn, J. E., Henderson, K. & Jedrzejas, M. J.
(2003). J. Mol. Biol. 325, 411–420.Sang, P. B. & Varshney, U. (2013). J. Bacteriol. 195, 1552–1560.Shibutani, S., Takeshita, M. & Grollman, A. P. (1991). Nature
(London), 349, 431–434.Sousa da Silva, A. W. & Vranken, W. F. (2012). BMC Res. Notes, 5,
367.Svensson, L. M., Jemth, A. S., Desroses, M., Loseva, O., Helleday, T.,
Hogbom, M. & Stenmark, P. (2011). FEBS Lett. 585, 2617–2621.Szabo, C. & Ohshima, H. (1997). Nitric Oxide, 1, 373–385.Taddei, F., Hayakawa, H., Bouton, M., Cirinesi, A., Matic, I.,
Sekiguchi, M. & Radman, M. (1997). Science, 278, 128–130.Terwilliger, T. C. et al. (2003). Tuberculosis, 83, 223–249.Winn, M. D. et al. (2011). Acta Cryst. D67, 235–242.Wolff, K. A., de la Pena, A. H., Nguyen, H. T., Pham, T. H., Amzel,
L. M., Gabelli, S. B. & Nguyen, L. (2015). PLoS Pathog. 11,e1004839.
research papers
364 Arif et al. � Mycobacterium smegmatis MutT1 Acta Cryst. (2017). D73, 349–364