Embed Size (px)
Citation preview

1
Bioassay Directed Characterization of Selected Indigenous
Medicinal Plants for their Anti-HCV Potential
A thesis submitted
to
University of Sargodha
In partial fulfillment of the requirements for the degree of
Doctor of Philosophy
in
CHEMISTRY
by
Sobia Noreen
Department of Chemistry
University of Sargodha
Sargodha
November 2014

2
CONTENTS
Page
DEDICATION I
DECLARATION II
CERTIFICATE III
APPROVAL CERTIFICATE IV
ACKNOWLEDGEMENT VI
ABSTRACT VII-VIII
ABBREVIATIONS IX-XI
CONTENTS XII-XV
LIST OF FIGURES XVI-XVII
LIST OF TABLES XVIII

3
Dedication
The fruit of my work is dedicated to
My Holy Prophet Hazrat Muhammad (SAW)
&
My Uncle Ch. Javed Iqbal (Late)
His abundant affections, patronizing encouragement and secret prayers have enabled
me to accomplish this task.
DECLARATION

4
I hereby declare that the work described in this thesis was carried out by me under
the supervision of Prof. Dr. Ishtiaq Hussain, Professor, Department of Chemistry,
University of Sargodha, Sargodha. 40100, Pakistan for the degree of Doctor of
Philosophy in Chemistry.
I also hereby declare that the substance of this thesis has neither been submitted
elsewhere nor is being concurrently submitted for any other degree. I further declare
that the work embodied in this thesis is the result of my own research and where work
of any other investigator has been used that has been duly acknowledged.
________________
Sobia Noreen
Registration No.: 09/UOS/ Ph.D/ CHM/01
Department of Chemistry
University of Sargodha

5
CERTIFICATE OF ORIGINALITY OF RESEARCH WORK
In accordance with Revised Rules & Regulations 2008 for MS/MPhil, MS/MPhil
leading to PhD and PhD, I hereby certify that Ms. Sobia Noreen has conducted
research work under my supervision, and is qualified to submit the thesis entitled,
“Bioassay directed characterization of selected indigenous medicinal plants for their
anti-HCV potential’’ for the degree of Doctor of Philosophy. It is further certified that
the research work carried out by the scholar is original and nothing is plagiarized.
Prof. Dr. Ishtiaq Hussain,
Supervisor
Department of Chemistry,
University of Sargodha,Sargodha
Dr. Muhammad Sher
Chairman
Department of Chemistry,
University of Sargodha, Sargodha
Prof. Dr. Muhammad Ilyas Tariq
Supervisor-II
Department of Chemistry,
University of Sargodha, Sargodha

6
APPROVAL CERTIFICATE
This thesis entitled “Bioassay directed characterization of selected indigenous
medicinal plants for their anti-HCV potential” submitted by Sobia Noreen,
Session 2009-2014, in the Partial fulfillment of the requirement for the degree of
Ph.D. in Chemistry is hereby approved.
Supervisors:
Prof. Dr. Muhammad Ilyas Tariq Supervisor-II Department of Chemistry, University of Sargodha, Sargodha.
External Evaluator
Chairman:
Prof. Dr. Ishtiaq Hussain, Supervisor Department of Chemistry, University of Sargodha, Sargodha.
Prof. Dr. Kausar Malik Director ORIC Lahore College for Women University Lahore.
Dr. Muhammad Sher
Department of Chemistry
University of Sargodha, Sargodha.

7
ACKNOWLEDGMENT
Prior to acknowledgment, I glorify, almighty Allah, the most merciful, the
most gracious who gave me the power, health, strength and means to complete my
research. First of all, all my acknowledgements and praises are for the Prophet
Muhammad, thousands salutations and benedictions to his sublime Holiness, the
chosen, the benefactor – the blessing and peace of God be with Him and his AAL A.S
through whose grace the sacred Quran descended.
It is an honor for me to owe my deepest gratitude to my supervisor, Prof. Dr. Ishtiaq
Hussain, his supervision and support, guidance, correction and valuable comments
made me to accomplish present study. I am also highly obliged to my co-supervisor
Prof. Dr. Ilyas Tariq for her excellent guidance and enthusiastic encouragement
throughout this research work. I would also like to express my gratitude to the Vice
Chancellor, Prof. Dr. Muhammad Akram Chaudhary, whose motivation as a role
model geared through the challenges. It is my pleasure to express my thanks to Prof.
Dr. Nazra Sultana, Dean, Faculty of Science and Technology; Dr. Muhammad Sher,
Chairman, Department of Chemistry, University of Sargodha for his support and
encouragement during the course of studies. Special thanks for my honorable
teachers Prof. Dr. Tayyab Husain, Diretor, CEMB, University of the Punjab, Lahore
and Dr. Shahid Iqbal, Dr. Ashraf Shaheen, University of Sargodha for their every
possible cooperation and constructive criticism during this study. Special thanks
reserve to Dr. Jhon M. Gardiner, MIB, University of Manchester, UK for his
generous support of his lab analytical facilities for six months during my research
visit to accomplish this task. I am also grateful to my all other colleagues from my
department or other department of my university for their assistance and guidance for
achievement of this task. I am thankful to all my teachers my mender who taught me
even a single word. I am here only because of you.
My sincere thanks to Mr. Asif, Mr. Afzal, Mr. Farhan Ahmad (Hightech.lab) and
other staff, who was really helpful and provides me every possible facility needed to
accomplish this task.
My special thanks to my all my friends, colleagues and students (Dr. Tusneem, Bushra
Ijaz, Fozia, Usman Ali ashfaq, Dr. Anjum Murtaza, Aqeel khan, Rehana, Sadia,
Farhana, Ayesha and Iffat for their sincere support, guideline and their cheerful
company which can never be forgotten by me. My heartiest acknowledgements are for
the untiring efforts of my family members especially my Mother, my anti, my sisters
and brothers who had been encouraging through entire period of my studies and
because of their prayers I am at this destiny. I owe my special gratitude to my dear
loving husband Qamar-ul-zaman, whose enthusiastic encouragement and sacrifices
made me able to complete this project. At the end I cannot forget my sweet baby Basil
Raffan, whose cheerful company always refresh me and provide me strength to
achieve my goals. My all success is due to prayers of my dear ones.
Sobia Noreen

8
ABSTRACT
Hepatitis C is the most common chronic blood born infection affecting approximately
3 % population of the world and about 6 % population of the Pakistan. About one–
fifth of the individuals with this infection results in the development of chronic liver
diseases, including liver cirrhosis and hepatocellular carcinoma. The present treatment
standard is pegylated INF-a along with ribavirin, direct-acting antivirals (DAAs) like
telaprevir and boceprevir for definite period according to viral genotype. The main
goal of therapy is to achieve a stable virological response along with eradication of
HCV infection and cure of the fundamental HCV induced liver disease.
Unfortunately, only less than 50 % of HCV patients get benefit to some extent from
this therapy due to side effects, resistance and high cost. Hence, there is a need to
develop anti-HCV agents, both from herbal sources and synthetic chemicals which are
non toxic, more efficacious and cost-effective.
Soon-Valley of Pakistan is bestowed with a unique biodiversity, comprising of
different climatic zones and wide range of plant species especially that have
hepatoprotective effect. No doubt phyto-constituents having the remarkable potential
inhibit the replication cycle of various types of DNA or RNA viruses. The present
work is intended to explore plants with anti-HCV potential, leading to natural
chemical entities as lead compounds. In current study, methanolic extracts of shade
air dried selected parts of fifteen medicinal plants of Soon Valley were screened
against HCV NS3 protease (genotype 3a) and as well as whole virus. The cellular
toxicity effects of organic extracts on the viability of Huh-7 were studied through
Trypan blue exclusion method and MTT assay. In serum inhibition assay, liver cells
were infected with high titre of HCV positive serum of genotype 3a for screening of
antiviral plants against whole virus. In in vitro protease inhibition assay, Huh-7 cells
were transfected with HCV NS3 protease by introducing mammalian expression
construct PCR3.1/FLAGtag/HCV NS3 in the presence and absence of plants extracts.
Only the methanolic extract of Caralluma tuberculata (CTS) and Portulaca oleracea
L. (POL) exhibited 57 % and 70 % inhibition. Four fractions of each CTS and POL
extract were obtained through bioassay-guided extraction. Subsequent inhibition of all
organic extract fractions against NS3 serine protease were checked to track the

9
specific target inside the genome of the virus and in this case only ethyl aceatate and
methanolic of CTS extracts inhibit the 69 % & 53 % and the POL extracts inhibit 80
% & 85 % expression of HCV NS3 protease respectively while keeping GAPDH
expression constant. The results showed that the POL and CTS methanolic crude and
ethyl acetate extract specifically abridged the HCV NS3 protease expression in a
dose-dependent fashion. The plant organic extract was screened for the presence of
phytochemical through standard procedures. Phytochemical analysis of selected
fractions confirmed the presence of alkaloids, coumarins, flavonoids, glycosides,
saponins, terpenoids, and tannins etc. In addition, the active fractions of both plant
extracts were also checked for their some other biological activities like antioxidant
potential, in vitro anti thrombotic and antibacterial activities. Antioxidant potential of
CTSM, CTSE, POLM and POLE was determined in term of total phenol, flavonoids,
tannin, carotenoid and free radical scavenging potential by following the different
reported procedure. Free radicals scavenging activities of extracts were determined by
DPPH, ABTS and FRAP assays. Among extracts highest antiradical values against
DPPH were found to be 81.747 by CTSM while lowest value (36.124) revealed by
POLE. Identification and quantification of phenolic acids in methanolic extracts of
CTS and POL were done by HPLC with UV/DAD that confirm the presence of
Sinapic acid, m-Coumaric acid, Gallic acid and Quercetin in CTSM along with Gallic
acid and Quercetin in POLM.
In case of in vitro anti thrombotic potential, highest activity among extract fractions
was shown by CTSM and minimum by POLE while the overall order of antithrombotic
potential was CTSM > CTSE > POLM >POLE. The antibacterial activity of methnolic
extracts of CTS and POL against Gram positive (Bacillus subtilis, Styphylococcus
aureus and Styphylococcus epidermidis) and Gram negative (Pseudomonas
aerugenosa, Escherichia coli and Salmonella typhi) strains of bacteria was
determined by disc diffusion method. Percent inhibition with respect to positive
control (Ciproflaxacin) indicated that both CTSM and POLM exhibited more than 60
% inhibition against Styphylococcus epidermidis.
Furthermore to identify the active ingredients, CTSE extract was also fractioned by
column chromatography. Five compounds CT-EtOAc-01 i.e. Methyl(12-O-benzoyl-
androstan-3-O-(14)-β-D-cymaropyranosyl-(14)-β-Dcymaropyranosyl-(14)-β-

10
D-cymaropyranosyl) hexanoate, CT-EtOAc-02 i.e. Methyl (12-O-benzoyl-androstan-
3-O-(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl--(14)-β-D-
cymaropyranosyl) hexanoate, CT-EtOAc-03 i.e. Methyl (23-O-benzoyl-androst-6-en-
17-O-(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl--(14)-β-D-
cymaropyranosyl)hexanoate, CT-EtOAc-04 i.e. Phenyl (12-O-benzoyl-gonan-6-en-
5’’-O-(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl)oxalate and CT-
EtOAc-05 i.e. Trans-p-ferulylalcohol-4-O-(6-(2-methyl-3-hydroxypropionyl)
glucopyranoside were isolated, purified and characterized by spectral data (UV-VIS,
FTIR, EI-MS and NMR).
The results obtained in this study suggested that methanolic and ethyl acetate extracts
of CTS and POL possess remarkable antiviral, antioxidant, antithrombotic, and
antibacterial effects which might be due to the charisma of their bioactive compounds
like glycosides, flavonoids, tannins, saponins and alkaloids etc. Further studies in
vitro, in vivo and in silico assays are mandatory to decipher the thorough aspects of
these phytocompounds. The novel antiviral activity of compounds from CTS presents
an attractive lead for natural chemical entities meant for the development of
prospective anti-HCV agents. Hence, POL and CTS extracts and its potential
constituents alone or plus with interferon could offer a future option to treat chronic
HCV.
11
ABBREVIATIONS
ABI Applied Biosystem
ABTS 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid)
Amp Ampicillin
AMPK AMP-activated protein kinase
APS Ammonium per sulfate
ATP Adenosin triphosphate
BHT Butylated hydroxytoluene
BMI Body mass index
bp Base pair
CC50 Cytotoxic concentration
CHC Chronic hepatitis C
CHCl3 Chloroform
CHO Chinese hamster ovary
CLDN1 Claudin1
Cox-2 Cyclooxygenase-2
CTS Caralluma tuberculata stem
CTSM Caralluma tuberculata methanolic extract
CTSE Caralluma tuberculata ethyl acetate extract
DAAs Direct acting antivirals
dH2O Distilled water
DMEM Dulbecco's Modification of Eagle's Medium
DNA Deoxyribonucleic acid
DMSO Dimethyl sulfoxide
dNTPs Deoxynucleoside triphosphate
DPPH 2,2-diphenyl-1-picrylhydrazyl
dsRNA Double stranded RNA
E1 Envelope glycoprotein 1
E2 Envelope glycoprotein 2
ECL chemiluminscent
EDTA Ethylenediamine tetra acetic acid
eIF Eukaryotic initiation factors
ELISA Enzyme Linked Immuno Sorbent Assay
ER Endoplasmic reticulum
ESI-MS Electro spray ionization-mass spectrometry
EtOAc ethyl acetate
FAS Fatty acid synthase
FBS Fetal Bovine Serum
GAG Glycosaminoglycans
GAPDH Glyceraldehyde-3-phosphate dehydrogenase
GFP Green fluorescence protein
GTP Guaninosine triphospahte

12
HBV Hepatitis B virus
HCC Hepatocellular carcinoma
HCV Hepatitis C virus
HPLC High performance liquid chromatography
HSC Hepatic stellate cells
Huh-7 Human hepatoma cell line-passage 7
HVR 1 Hypervariable region 1
IC Internal control
IDUs Injection drug users
IFN Interferon
IR Infra-red
IRES Internal ribosomal entry site
ISDR IFN-sensitivity determining region
ISGs Interferon Stimulated Genes
JNK c-Jun N-terminal kinase
Kb Kilo base pair
kbp Kilo base pair
kDa Kilo daltons
MAPK Mitogen activated protein kinase
MAVS Mitochondrial Inhibitor of Viral Signaling
MeOH methanol
mg Microgram
mL Microlitre
M-MLV Moloney murine leukemia virus
mRNA Messenger RNA
NAFLD Non-alcoholic fatty liver disease
NANBH Non-A, non-B hepatitis
ng Nano gram
NMR Nuclear magnetic resonance
NS Nonstructural
nt Nucleotide
NTR Non-translated region
ORF Open reading frame
P53 tumor supressor protein
PAGE polyacrylamide gel electrophoresis
PAPRα Peroxisomal proliferator activator receptor-alpha
PBS Phosphate Buffered Saline
PEG Polyethylene glycol
PEG-INFα Pegylated interferon α
PKR Protein Kinase R
pM Pico-Mole
PO Portulaca oleracea
POL Portulaca oleracea leaves
POLE Portulaca oleracea L ethylacetate

13
POLM Portulaca oleracea methanol
RdRp RNA-dependent RNA polymerase
RISC RNA induced silencing complex
RNA Ribonucleic acid
RNAi RNA interference
RNase Ribonuclease (enzyme)
ROS Reactive oxygen species
rpm Revolution Per Minute
RT Room Temperature
RT-PCR Reverse transcriptase polymerase chain reaction SDS–PAGE Sodium dodecyl (lauryl) sulfate-polyacrylamide gel
electrophoresis
ssRNA Single stranded RNA
SVR Sustained virological response
TAE Tris Acetate EDTA
TBE Tris-boric acid EDTA (buffer)
TGFB Transforming growth factor-B
TMS Trimethylsilane
TNF-α Tumor necrosis factor alpha
TLC Thin layer chromatography
UBC Ubiquitin
WHO World health organization

14
Table of Contents 1.0 Introduction ....................................................................... Error! Bookmark not defined.
1.1 Natural Products from Times of Yore to 21st Millennium Error! Bookmark not defined.
1.2 Status of Medicinal Plants and their Research in Pakistan Error! Bookmark not defined.
1.3 Preface to Genus Caralluma and Caralluma tuberculata . Error! Bookmark not defined.
1.3.1 Active Constituents .................................................. Error! Bookmark not defined.
1.3.2 Pharmacological worth of Caralluma tuberculata and other species of Caralluma
........................................................................................ Error! Bookmark not defined.
1.4 Introduction to Portulaca oleracea ................................. Error! Bookmark not defined.
1.4.1 Phytochemistry of Portulaca oleracea ...................... Error! Bookmark not defined.
1.4.2 Pharmacological attributes of Portulaca oleracea .... Error! Bookmark not defined.
1.5 Hepatitis......................................................................... Error! Bookmark not defined.
1.6 Hepatitis C and its Virus .................................................. Error! Bookmark not defined.
1.6.1 Transmission of Pathogen and Genotype Prevalence Error! Bookmark not defined.
1.6.2 HCV Structure and Genome ..................................... Error! Bookmark not defined.
1.6.3 HCV Non-Structural and Structural Proteins ............. Error! Bookmark not defined.
1.6.4 HCV RNA Replication and Potential Drug Target....... Error! Bookmark not defined.
1.6.5 In vitro Infection Systems for HCV Replication .......... Error! Bookmark not defined.
1.6.6 Available Treatment Options against HCV ................ Error! Bookmark not defined.
1.6.7 NS3 Gene as Candidate Drug Target......................... Error! Bookmark not defined.
1.6.8 Ayruvadic Extract and Plant Derived Compounds as anti HCV Drug ................ Error!
Bookmark not defined.
1.7 Goals/Objectives ............................................................ Error! Bookmark not defined.
2.0 Chemicals, Equipment and Techniques ............................... Error! Bookmark not defined.
2.1 Chemicals ...................................................................... Error! Bookmark not defined.
2.1.1 Materials for Chromatography ................................. Error! Bookmark not defined.
2.1.2 Chemicals and Reagents used for Determination of Antioxidant....Error! Bookmark
not defined.
2.2 Instrument and Apparatus ............................................... Error! Bookmark not defined.
2.3 Materials used for Biological Activities .......................... Error! Bookmark not defined.
2.3.1 Cell Lines ................................................................ Error! Bookmark not defined.
2.3.2 Plasmids .................................................................. Error! Bookmark not defined.
2.3.3 Reagents & Chemicals ............................................. Error! Bookmark not defined.
2.3.4 Designing of Primers ............................................... Error! Bookmark not defined.
2.4 Instrumentation & Techniques ....................................... Error! Bookmark not defined.

15
2.4.1 Physical Constant .................................................... Error! Bookmark not defined.
2.4.2 Spectroscopy ........................................................... Error! Bookmark not defined.
2.4.3 Thin Layer Chromatography .................................... Error! Bookmark not defined.
2.4.4 Column Chromatography (CC) ................................ Error! Bookmark not defined.
2.5 Methodology .................................................................. Error! Bookmark not defined.
2.5.1 Biochemical Analysis of Plants ................................ Error! Bookmark not defined.
2.5.2 Collection, Identification and Preservation of Medicinal Plant Materials ........ Error!
Bookmark not defined.
2.5.3. Plant Extract Preparation......................................... Error! Bookmark not defined.
2.5.4 Screening of Plant Extracts for Cellular Toxicity Analysis via Trypan Blue Dye
Exclusion Method ............................................................ Error! Bookmark not defined.
2.6 Anti-HCV Analysis of Compounds in Liver Cells........... Error! Bookmark not defined.
2.6.1 Antiviral Analysis of Plant Extract against HVC Ns3 Protease via Transfection
Assay ............................................................................... Error! Bookmark not defined.
2.7 Gene Expression studies via Reverse Transcriptase PCR (RT-PCR) ....Error! Bookmark
not defined.
2.7.1 Extraction of Total RNA .......................................... Error! Bookmark not defined.
2.7.2 Synthesis of cDNA .................................................. Error! Bookmark not defined.
2.7.3 Reverse Transcription PCR (RT-PCR) for Gene Expression Analysis Error! Bookmark
not defined.
2.7.4 Expression studies via Real-time PCR...................... Error! Bookmark not defined.
2.8 Western Blotting of PO extracts ..................................... Error! Bookmark not defined.
2.8.1 Protein Isolation ...................................................... Error! Bookmark not defined.
2.8.2 Protein Quantification ............................................. Error! Bookmark not defined.
2.8.3 Protein Samples Preparation ................................... Error! Bookmark not defined.
2.8.4 SDS Gel Electrophoresis ........................................... Error! Bookmark not defined.
2.8.5 Transfer of Protein and Immunoblotting .................. Error! Bookmark not defined.
2.8.6 Enhanced Chemiluminescence (ECL) Detection ........ Error! Bookmark not defined.
2.9 Analysis of the Two Plants Extract That Exhibit Positive Potential against HCV .. Error!
Bookmark not defined.
2.10 Phytochemical Screening .............................................. Error! Bookmark not defined.
2.10.1 Qualitative Analysis of the Extracts ....................... Error! Bookmark not defined.
2.10.2 Quantitative Determination of the Chemical Constituency Specifically
Antioxidants ..................................................................... Error! Bookmark not defined.
2.11 Anti-clotting Properties of CTS and POL Extracts ........... Error! Bookmark not defined.
2.11.1 Sample Preparation ............................................... Error! Bookmark not defined.

16
2.11.2 Preparation of Anti-Clot Test Solution .................... Error! Bookmark not defined.
2.11.3 Clot Lysis ................................................................ Error! Bookmark not defined.
2.12 Antibacterial assay of extracts ...................................... Error! Bookmark not defined.
2.13 Bioassay guided extraction of CTS and POL ................ Error! Bookmark not defined.
2.14 Isolation of Compounds via Column Chromatography .. Error! Bookmark not defined.
2.14.1 Isolation of Compounds ......................................... Error! Bookmark not defined.
2.14.2 Physico-chemical Characteristics of Isolated Compounds..... Error! Bookmark not
defined.
2.14.3 Elucidation of Compounds Structure by spectroscopy .......... Error! Bookmark not
defined.
3.0 Results and Discussion ....................................................... Error! Bookmark not defined.
3.1 Collection of Botanical Samples and Preparation of Extract .......... Error! Bookmark not
defined.
3.2 Percentage yield of plants extract.................................... Error! Bookmark not defined.
3.3. Toxicological Studies of Plant Extracts in CHO and Liver Cell Lines .Error! Bookmark
not defined.
3.3.1 Trypan Blue Dye Exclusion Assay ........................... Error! Bookmark not defined.
3.3.2 Cytotoxic Screening of Phyto Extracts via MTT Assay Error! Bookmark not defined.
3.4 Anti-HCV Assay ............................................................ Error! Bookmark not defined.
3.5 Inhibition of HCV NS3 Gene Expression by Selected Organic ExtractsError! Bookmark
not defined.
3.6 Western Blotting ............................................................ Error! Bookmark not defined.
3.7 Phytochemical analysis................................................... Error! Bookmark not defined.
3.7.1 Qualitative Analysis of CTS and POL .............. Error! Bookmark not defined.
3.7.2 Quantitative Methods for Screening of Phytoconstituents Error! Bookmark not
defined.
3.8 Antioxidant Potential of Biologically Active Plant Extract Fractions Error! Bookmark not
defined.
3.8.1 Total Phenolic, Total Flavonoids and Total Carotenoid Contents ...Error! Bookmark
not defined.
3.8.2 Free Radical Scavenging Activity of Extracts by DPPH, ABTS and FRAP Assays
........................................................................................ Error! Bookmark not defined.
3.8.3 Quantitative Estimation of Phenolic Acid by HPLC . Error! Bookmark not defined.
3.10 Antithrombotic Potential of Extracts ............................. Error! Bookmark not defined.
3.11 Screening of Antibacterial Activity of Samples ............. Error! Bookmark not defined.
3.12 Extraction, Purifcation and identification of Compounds Obtained from CTSE Extract
............................................................................................ Error! Bookmark not defined.

17
3.12.1 Compound CT-EtOAc-01 ...................................... Error! Bookmark not defined.
3.12.2 Compound CT-EtOAc-02 ...................................... Error! Bookmark not defined.
3.12. 3 Compound CT-EtOAc-03 ................................. Error! Bookmark not defined.
3.12.4 Compound CT-EtOAc-04 ........................................ Error! Bookmark not defined.
3.12.5 Compound CTEtOAc-05 ................................... Error! Bookmark not defined.
3.13 Discussion .................................................................... Error! Bookmark not defined.
4.0 Future Prospects ................................................................ Error! Bookmark not defined.
5.0 References ......................................................................... Error! Bookmark not defined.
6.0 Appendix ........................................................................... Error! Bookmark not defined.

18
LIST OF FIGURES
Fig. 1.1: Taxonomy of Caralluma tuberculata N.E.Br. and whole plant along with
flower ………………………………………………………………………………….6
Fig 1.2: Chemical structures of some notable basic compounds of Caralluma genus..9
Fig. 1.3: Pharmacological properties of some significant Caralluma species ………14
Fig. 1.4: Taxonomy of Portulaca oleracea L and whole plant along with flower ….15
Fig. 1.5: Detailed life cycle of hepatitis C virus …………………………………….29
Fig 1.6: The HCV subgenomic replicon system …………………………………….30
Fig. 1.7: Scheme of HCV Pseudo-particle production and infection..………………31
Fig. 2.1: Cycling parameters for expression analysis of HCV NS3 gene in reverse
transcription PCR ……………………………………………………………………52
Fig. 2.2: Thermal cycling conditions for Real-time PCR …………………………...53
Fig. 2.3: Scheme for bioassay-guided fractionation of Caralluma tuberculata and
Portulaca oleracea L methanolic extracts……………………………………………66
Fig. 3.1: Percentage yield of methanolic extracts of plants collected from Soon-
sakaser valley of Pakistan…………………………………………………………….71
Fig. 3.2 (A & B): Cellular toxicity analysis CTS and POL extracts through Trypan
blue exclusion method ………………………………………………………….........73
Fig. 3.3 (A & B): Toxic effect of plant (CTS and POL) crude extracts on Huh-7 and
CHO cell lines by MTT assay …………………………………………………..…...74
Fig 3.4: Anti-HCV activity of methanolic extracts of selected fifteen different
plants………………………………………………………………...........................76
Fig. 3.5 (A&B): Anti-HCV activity of organic extracts of POL and CTS…………..77
Fig. 3.6 (A and B): Antiviral Activity of methanolic and ethyl acetate extracts of POL
against HCV NS3 protease. ………………………………………………................79
Fig. 3.7 (A and B): Qualitative & quantitative inhibition of HCV NS3 protease by
IFN, methanolic and ethyl acetate extract of CTS. ………………………………….79
Fig. 3.8: Antiviral Activity of methanolic and ethyl acetate extracts of POL against
HCV NS3 protease at protein level…………………………………………………..80
Fig. 3.9: Total Phenolic Contents of selected sample extracts. All experiments were
performed in triplicate. Data are presented as the mean ± SEM of n = 3
experiments…………………………………………………………………………..84
Fig. 3.10: Total flavonoid Contents of selected sample extracts. All experiments were
performed in triplicate. Data are presented as the mean ± SEM of n = 3
experiments…………………………………………………………………………..84
Fig. 3.11: Total carotenoid Contents of selected sample extracts. All experiments
were performed in triplicate. Data are presented as the mean ± SEM of n = 3
experiments…………………………………………………………………………..85
Fig. 3.12: DPPH free radical scavenging potential of selected sample extracts. All
experiments were performed in triplicate. Data are presented as the mean ± SEM of n
= 3 experiments………………………………………………………………………86
Fig. 3.13: ABTS free radical scavenging potential of selected sample extracts. All
experiments were performed in triplicate. Data are presented as the mean ± SEM of n
= 3 experiments.……………………………………………………………………...86
Fig. 3.14: Free radical scavenging potential of selected sample extracts by FRAP
assay All experiments were performed in triplicate. Data are presented as the mean ±
SEM of n = 3 experiments. …………………………………………………………..87
Fig. 3.15: Identification and quantification of Phenolic acids in CTSM
………………………………………………………………………………………..88

19
Fig. 3.16: Identification and quantification of Phenolic acids in
POLM…………………………………………………………………………………88
Fig. 3.17: In vitro thrombolytic activity of selected fractions of plant extracts with
respect to positive control (Streptokinase) and negative control (water). All results are
presented as mean and standard deviation for the three individual experiments. ***P
value < 0.001 vs Streptokinase (control). ……………………………………………89
Fig. 3.18: Percent inhibition of bacteria by selected extracts with respect to
ciproflaxacin (positive control)………………………………………………………91

20
LIST OF TABLES
Table 1.1: Historical perspectives of natural products.................................................2
Table 1.2: Apocynaceae Meve classifications into subfamilies with their genera and
species numbers …………………....................................................... ....................5
Table 1.3: Global distribution and pharmacological activities of different species of
Caralluma genus……………………………………………………………………..13
Table 2.1: Primers of GAPDH and HCV non-structural NS3 gene of 1a & 3a
genotype was used in this study …………………………………………………….44
Table 2.2: Botanicals details chosen for screening against total RNA HCV and its
gene NS-3 serine protease…………………………………………………………....47
Table 2.3: Composition of the gels used in SDS-PAGE ……………………………55
Table 3.1: Qualitative phytochemical screening of CTS and POL methanolic extracts.
………………………………………………………………………………………..81
Table 3.2: Percent phytochemical components in CTS and POL methanolic
extracts………………………………………………………………………………..82
Table 3.3: Identification and quantification of Phenolic acids by HPLC……..…….88
Table 3.4: Zones of inhibition (mm) in Disc Diffusion Plate for the antibacterial
activity of successively extracted alcoholic extracts (SAE) of plants that have also
antiviral potential.…………………………………………………………………….91

21
1.0 Introduction
A ‘natural product’ term widely covers the product of natural bio source like plant,
animal and microbes. It may be in the form of extract or single pure compound
(secondary metabolites) like alkaloids, limonoids, steroid, flavonoid, lignana,
terpenoids, organosulfur, tannin, furyl, polylines, chlorophyllins, thiophenes,
saponins, sulphides, coumarins, etc. (Naithani et al., 2008). The worth of medicinal
plants lies in these metabolites that are non-nutritive for plants, but produce certain
physiological action in plants and human against different types of infectious disease
and metabolic disorders (Edeoga et al., 2005). In the field of antiviral compound
search these Phyto-compound are striking targets owing to their ability to hamper
viral entrance, blocking or restricting the replication of the RNA / DNA genome of
the virus (Naithani et al. 2008). Bioactive compounds are also generated by plants as
a result of their own “defence mechanisms” against pathogens (Wedge et al., 2000).
Human interest in natural products constantly spans the history because of their
striking potential as medicines, food, agriculture, cosmetics, etc. (Edeoga et al.,
2005).
1.1 Natural Products from Times of Yore to 21st
Millennium
Phytotherapy history is as old as the history of humankind itself (Ahmad et al., 2006).
System of alternative medicine remained enriched by traditional Ayurveda, Kampo,
Unani and Chinese medicine throughout the history. All these systems are based on
myths and reality based knowledge that decipher frequently modified theory and
practices. All these approaches of natural component formulation are still widely
accepted in this century, owing to their therapeutic strength (Harvey et al., 2000).
Summarized authenticated information about natural product usage as medicine in
historical perspective is appendices in table 1.1.
Mostly the formulation scheme was kept in secret by traditional healers, herbalist and
shaman, so this thing is a major obstacle to check the authenticity of these
formulations on a scientific basis. Hence, flair to access bioactive compounds,
realizing their worth and potential has always remained a main driving force. The

22
incredible resurgence of this research field was noticed over the preceding decade or
so due to their small size (<200 Da), incomparable structure, diversity and drug like
characteristics (absorbed and metabolized easily) (Sarker et al., 2005).
Table 1.1: Historical perspectives of natural products Time period Category Elucidation
Before 320 BC Ayurveda (Chinese old
system of medicine)
Information regarding the medicinal properties of
plants and their products.
1550 BC Ebers papyrus Details about large number of crude drugs from plant
source like gum Arabica, castor seeds etc.
460-377 BC Hippocrates Enlist medicinal properties of 400 plants
370-287 BC Theophrastus Describe usage of plants and animals as medicine
23-79 AD Pliny the Elder Plant and animals usage as medicine
60-80 AD Discorides Author of “De material medica” ( Provide information
about medicinal potential of more than 600 plants)
131-200 AD Galen Author of “Galenicals” who practiced botanical
medicine and familiarized the West to its usage
Before 632AD Tib-E-Nabvi In this book Hazrat Muhammad (PBUH) suggested the
treatment of different disease by natural sources
15th Century Krauterbuch Depicted elaborated information and picture of
medicinal plants.
No doubt, the contribution of natural products is remarkable for the development of
drug is either in its intact form like vincristine (Catharanthus roseus), or act as
‘‘building blocks’’ substance that further modified into more complex molecule like
diosgenin (Dioscorea floribunda) used in oral contraceptives, or the novel analogs
synthesis from them like synthetic analogs of penicillin (Penicillium notatum) (Sarker
et al., 2005). Plant based medicine satisfying the prime health care requirement of 80
% of the populace of developing countries (WHO, 2002).
In the last couple of decades, traditional medicine has been remained appealing target
by academia and the pharmaceutical industry in search of "potential leads" that can be
further utilized in the synthesis of modern-day drugs (De Silva et al., 1997). More
than 40 % of modem drugs have been derived from plants specifically anticancer and
antibacterial drugs. In the beginning of this millennium, eight out of thirty top selling
drugs (amoxicillin, azithromycin, clavulanic acid, cyclosporine, plastitaxel,
ceftriaxone, pravastatin and simvastatin) were either natural product or their
respective derivative (Cragg et al., 1997).
Owing to the ever increasing cost, adverse effects, multiple drug and microbial
resistance of the synthetic drugs; people are now turning to the world of

23
ethnopharmacognosy that is literally more efficacious, safe, and inexpensive with a
alleviated side effect (Sànchez-Lamar et al., 1999). Moreover, these natural products
can also disclose new springs of economic resources such as gums, oils, tannins etc.
and these can be further used as precursors of other synthetic complexes (Farnsworth
et al., 1966). Not only in Asian and African countries but also in Europe and North
America plant based therapies are extensively accepted in the form of food
supplements, nutraceuticals, complementary and alternative medicines.
Along with the expansion of newest molecular targets the stipulation for novel
biologically active molecules has been increased. The previous practice of natural
product research mainly spans around straightforward extraction and recognition of
chemistry instead of its in vitro bioactivity. Sometime, in vivo models were used to
determine their biological activity on the basis of ethno-pharmacological information.
But the modern strategies opted in vitro bioassay-guided separation along with
detection of active ‘‘lead’’ from natural reservoir. More attention is given to
bioactivity and libraries of natural products.
New horizons of interdisciplinary science strengthen the research on natural product
by introducing the concept of genetic manipulation and natural combinatorial
chemistry. Because of the latest updates in the hyphenated techniques of separation
and purification like mass spectrometry, advanced spectroscopic techniques, and
ultrasensitive in vitro micro plate-based assays have revolutionized the research of
pre-isolation of crude extract, online detection, chemical finger printing, and
metabolomic analyses (Sarker et al., 2005).
1.2 Status of Medicinal Plants and their Research in
Pakistan
Pakistan is bestowed with exclusive floral array in nine different ecological zones
owing to its ideal climate conditions. In Pakistan, approximately 600 species out of
6000 wild plants are considered highly significant from the medicinal potential point
of view (Hamayun et al., 2003). Mostly rural and suburban population of Pakistan
primarily depends on plants or their active ingredients for their healthcare (Shinwari

24
et al., 2003). Unani system of medicine is used widely by traditional healers or
Hakims while they suggest plant based remedies (Goodman et al., 1992).
Soon-Valley of Pakistan is blessed by the unique biodiversity of plant species because
of a wide range of unusual climatic precincts. Its climate is subtropical with unique
geological formation due to high concentration of salt as well as other minerals in the
subsoil. In the local vegetation, a least work has been conducted on natural product
with respect to the medicinal point of view (Ahmad et al., 2008). The plant
communities are losing their species richness at high rate due intensive deforestation
and unlimited expansion of urban areas. Despite of ever increasing human knowledge
and folklore regarding usage of natural product, appropriate modern scientific
approaches have only been applied to a very little part of the world's flora. It is a
matter of grave concern to exploit the potential of those plant species that are in
waiting line before they become endangered (Sasidharan et al., 2011).
1.3 Preface to Genus Caralluma and Caralluma
tuberculata
Caralluma is considered in the family Apocynaceae that comprises three subfamilies
Rauvolfioideae, Apocynoideae and Asclepiadaceae holding 424 genera (Liede-
Schumann et al., 2005). Ornamental plants like Allamanda, Frangipani, Oleander,
Vinca of tropical area, large trees with special buttress roots of rainforests and
deciduous or evergreen trees, climbers or shrubs of the world warm and temperate
regions are included in these genera. Milky latex of most of the plants is considered
important for medicinal purpose and rubber production. Later, in 1890 Robert Brown
separated Asclepiadaceae (milkweed family) from Apocynaceae. Recent advances in
genomic, molecular and morphological analysis merge the Asclepiadaceae and
Periploceae into Apocynaceae family (Endress et al., 2005). Meve also categorized
Asclepiadaceae into three tribes as fallow Table no. 1.2.
Caralluma tuberculata is a member of the milkweed family Asclepiadaceae which
include about 2500 species from 200 genera and widely distributed in some tropical
regions of Punjab, Khyber Pakhtoonkhawa and Baluchistan provinces of Pakistan. In
Pakistan, Asclepiadaceae is represented by approximately 23 genera along with 40
25
species. In semi-arid and desert vicinity of the country this Caralluma species have
been used as emergency victuals for centuries.
Table 1.2: Apocynaceae Meve classifications into subfamilies with their genera and
species numbers
Subfamily Total
Genera
Total
Species Distribution
Asclepiadoideae 177 3000 Cosmopolitan particularly in Africa, Asia
and Europe
Periplocoideae 45 190 Restricted to Africa and Asia only
Secamonoideae 9 180 Restricted to Africa and Asia only
(Meve et al., 2005; Verhoeven et al., 2001 & Muller et al., 2002)
The Caralluma turbeculata vernacular name is chunga or chung in Punjab. Plants
belonging to the genus Caralluma are normally leafless (Dawidar et al., 2012). This
perennial herb has 15-45 cm height with a succulent, fleshy, leafless, erect and 4
angled stems having grooves on it. Quadrangular stem bears small flowers in several
varieties of dark colour (Madhuri et al., 2011). Sessile single or many flowers in
lateral cymes having ovate-lanceolate five sepals and deeply divided, glabrous, lobe
lanceolate, dark purple colour corolla of about 9 mm in diameter. Anthers are without
appendages and each having pollen mass one. Glabrous Follicles with size about 9-11
cm are progressively tapering to the tip. The flowering season is June. It mostly grows
widely on rocks after rain, but it is also cultivated in some areas. Its stem and roots are
not only eaten raw as famine food, but also cooked due to its pharmacological
potential. The complete classification of Caralluma tuberculata is deciphered in
Fig.1.1.
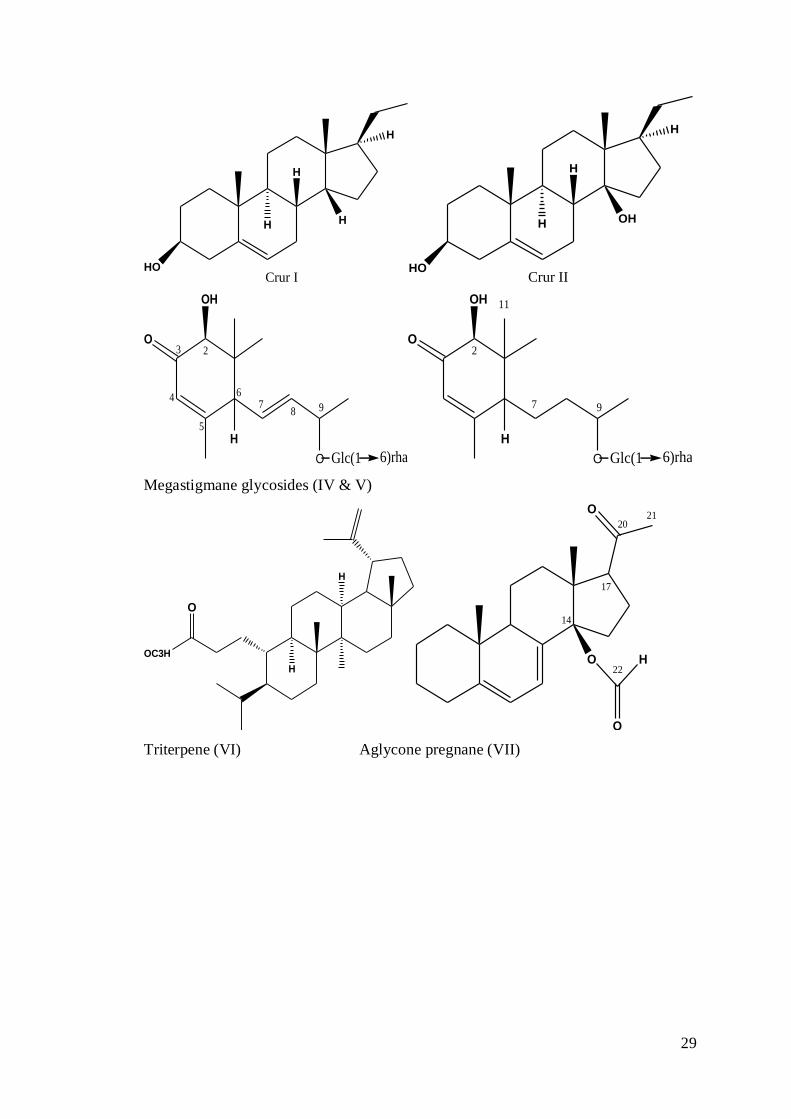
1.3.1 Active Constituents
Caralluma, in general, is famous for the presence of its key component glycosides
specifically pregnane and aglycone, megastigmane glycosides, flavonoids, saponins
and triterpenes. Pregnanes (I) are the C-21 steroids having perhydro-1,2-
cyclopentanophenanthrene ring as backbone that have β-oriented methyl group at two
positions (C-10 and C-13) and two outside chains of carbon atoms at C-17. In
derivative form of pregnane on C-14 atom β confirmation is present along with a
hydroxyl group (Deepak et al., 1989). Genus Caralluma was 1st time explored for its
constituents in 1967 and two compounds (dihydrosarcostin (3, 8, 12, 14, 17,

26
(20S) -hexahydroxypregnane and tomentogenin (3, 12, 14, 17, (20S)-
pentahydroxypregnane) (II &III) were extracted from C. dalzielii (Deepak et al.,
1997).
Domain Eukaryota
Kingdom Plantae
Subkingdo
m
Viridaeplantae
Phylum Tracheophyta
Subphylum Euphyllophytina
Infraphylu
m
Radiatopses
Class Magnoliopsida
Subclass Asteridae Caralluma tuberculata: Whole plant
& flower
Superorder Gentiananae
Order Gentianales
Family Apocynaceae
Subfamily: Asclepiadoideae
Tribe Ceropegieae
Genus Caralluma
Specific
epithet
tuberculata - N.E.Br.
Botanical
name
Caralluma tuberculata
N.E.Br.
Fig. 1.1: Taxonomy of Caralluma tuberculata N.E.Br. and whole plant along with
flower
The literature survey indicates that the pregnane glycosides were isolated from
various parts of the plant. With polar solvents like ethylacetate, n-butanol, methanol,
ethanol and water, the compounds like C21 class of steroidal glycosides with high
molecular weight compounds were obtained but on other hand compounds having low
weight like cumarin, sterol, steroid, terpenoid were obtained when the same parts
were extracted with non-polar solvents like n-hexane, benzene etc. In a couple of
studies, two new pregnane glycosides carumbellosides I and II and steroidal
glycosides i.e. carumbellosides III-V were isolated from the extract of Caralluma
umbellate whole plant first reported the isolation and identification of (Kishore et al.,
2010; Sheng-Xiang et al., 1997). New steroidal glycosides, stalagmosides I-V and
indicosides I and II together with the known compounds Carumbelloside III,
lasianthoside A and lasianthoside B were isolated from the whole plant of Caralluma

27
stalagmifera. The genus is also characterized by the presence of flavones glycosides
(Sreelatha et al., 2010).
Two novel steroid glycosides were isolated from Caralluma tuberculata that
exhibited fairly in vitro cytotoxic activity on breast cancer cells even in micromolar
concentration. Pregnanes are C21 steroids and often found in nature conjugated as
glycosides. A flurry of pregnane glycosides and esterified polyhydroxypregnane were
extracted from Caralluma and other family member of Asclepiadaceae. Some of these
glycosides appear as novel potential template for drug development against cancer
and tumor (Waheed et al., 2011).
Chemical investigation of Caralluma tuberculata indicates that it was endowed with
the flavone glycosides and several pregnane glycosides (Abdel-Sattar et al., 2011;
Oyama et al., 2007). Five pregnane glycosides were isolated from Caralluma
tuberculata, in addition to a known one (russelioside E, 6). All these six compounds
were checked for their action against malaria and trypanosome. Moreover, their
cytotoxic effect was also tested on the growing human MRC5 embryonic cell line
(Abdel-Sattar et al., 2008). Caratuberoside C (I), and D (II), two new pregnane
glycosides were isolated from Caralluma tuberculata (Rizwani et al., 1993). A
medicinal herb, Caralluma tuberculata, furnished a pregnane type compound,
caratuberside A2. The sugar linkage was at C-14 which is a rare site of substitution
(Rizwani et al., 1993). Key phytochemical ingredients of Caralluma umbellate
include pregnane glycosides, flavone glycosides, bitter principles, saponins and
various flavonoids (Al-Massarani et al., 2012; Ray et al., 2012). Some acylated
pregnane glycosides like russeliosides E–H were obtained from chloroform extract of
Caralluma russeliana (Abdel-Sattar et al., 2008).
The aglycone pregnane (Pregnane-14β -formyl-5, 7-dien-20-one) (VII), with unusual
formyl group at C-14, were extracted from of the non polar (petroleum ether) extract
Caralluma umbellate stem (Babu et al., 2008). Similarly two more aglycone steroids
Cur I (3-hydroxy-pregn-5-ene) and Cur II (3, 14-dihydroxy pregn-5-ene) were
extracted from the non polar extract of the roots of same plant (Kishore et al., 2010).
Furthermore, in other studies, one more compound, i.e. 20S-epimer of boucerin was

28
extracted 1st time from the toluene extract of the above said specie of Caralluma
(Kunert et al., 2009).
The Phytochemistry of genus Caralluma is characterized by many pregnane
glycosides, but recently megastigmane glycosides (IV & V) have also been isolated
from Caralluma negevensis with few flavones. Saponins and flavonoids
predominantly found in the Caralluma are of great interest because of the wide range
of immunostimulating activities. As Caralluma attenuata, the fresh whole plant
contains luteolin-4-O-neohesperidoside, a flavonoid (flavones glycoside) identified as
the major chemical constituents of the plant. Caralluma adscendens is declared to
contain saponin glycosides, bitters, pregnane glycosides (caratubersides A and B and
various boucerosides) (Tatiya et al., 2010).
Several members of the genus Caralluma are rich in triterpenes, pregnane
glycosides or their esters which may have found medicinal uses (Dawidar et al.,
2012). Triterpenes squalene, lupeol, lupenone, lupeol acetate, β -sitosterol acetate and
guimarenol were extracted from Caralluma buchardii. Glycone moieties include
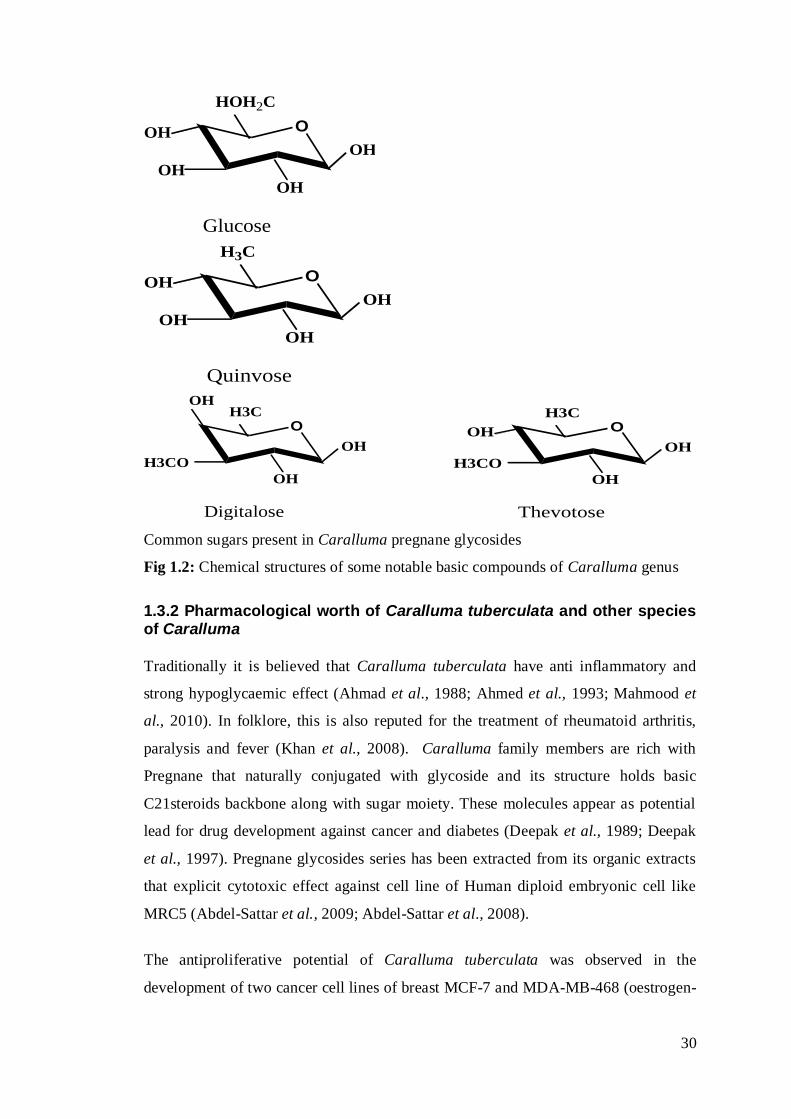
different sugars like glucose, digitalose, thevotose, quinvose etc.
HOH
H
H
OH
6
1
2
3
4
57
8
9
10
11
12
13
14
15
16
17
18
19
20
21
AB
CD
HO
CH3
H
H
CH3
OHOH
CH3
OH
OH
R
Pregnane (I) R= H, Tomentogin (II) R=OH, Dihydrosarcostin (III)
29
HO
H
H H
H
Crur IHO
H
H OH
H
Crur II
H
O
OH
O
23
4
5
67
8 9
Glc(1 6)rha
H
O
OH
O
2
7 9
Glc(1 6)rha
11
Megastigmane glycosides (IV & V)
OC3H
O
H
H
2021
O
14
O
H
O
17
22
Triterpene (VI) Aglycone pregnane (VII)
30
O
OH
OH
HOH2C
Glucose
OH
OH
O
OH
OH
H3C
Quinvose
OH
OH
O
OH
OH
OHH3C
H3CO
Digitalose
O
OH
OH
H3C
H3CO
Thevotose
OH
Common sugars present in Caralluma pregnane glycosides
Fig 1.2: Chemical structures of some notable basic compounds of Caralluma genus
1.3.2 Pharmacological worth of Caralluma tuberculata and other species of Caralluma
Traditionally it is believed that Caralluma tuberculata have anti inflammatory and
strong hypoglycaemic effect (Ahmad et al., 1988; Ahmed et al., 1993; Mahmood et
al., 2010). In folklore, this is also reputed for the treatment of rheumatoid arthritis,
paralysis and fever (Khan et al., 2008). Caralluma family members are rich with
Pregnane that naturally conjugated with glycoside and its structure holds basic
C21steroids backbone along with sugar moiety. These molecules appear as potential
lead for drug development against cancer and diabetes (Deepak et al., 1989; Deepak
et al., 1997). Pregnane glycosides series has been extracted from its organic extracts
that explicit cytotoxic effect against cell line of Human diploid embryonic cell like
MRC5 (Abdel-Sattar et al., 2009; Abdel-Sattar et al., 2008).
The antiproliferative potential of Caralluma tuberculata was observed in the
development of two cancer cell lines of breast MCF-7 and MDA-MB-468 (oestrogen-

31
dependent and estrogen-independent), and colonic cells (U937 and Caco-2) (Waheed
et al., 2011). Androstan and pregnane glycosides stimulate caspase mediated
apoptosis. Caspase enzyme, dependent on Calcium, is activated via two possible
pathways, intrinsic (e.g. mitochondria damage) and extrinsic (e.g. Cell surface death
receptor ligation). This activated caspase along with caspase 3 triggers the DNA
fragmentation by the cleavage of PARP (poly-ADP Ribose polymers). After PARP
cleavage, cell irrevocably prone to apoptosis. Due to structural homology of Pregnane
glycosides(12-O-benzoyl-20-O-acetyl-3,12,14,20-tetrahydroxy-pregnan3-ylO-D-
glucopyranosyl-(1→4)-d-glucopyranosyl-(1→4)-3-methoxy-d-ribopyranoside) with
estrogen agonist inhibits Calcium exchangers, as a result calcium concentration
increased that stimulate caspases and apoptosis (Deepak et al., 1997). Another
possibility is activation of the xenobiotic and steroid receptors that also induce the
apoptosis process in cancerous cells of the breast (Verma et al., 2009). This process is
not fully cleared hence further research is mandatory to decipher the activation of
xenobiotic receptors.
Treatment of mice with Caralluma tuberculata extract stimulates composite
biochemical and cytological variations in it (Al-Bekairi et al., 1992). The ethanolic
extract of Caralluma tuberculata afforded potential protection against gastric mucosa
injuries caused by ethanol (80 %), sodium hydro oxide (0.2 M), hypertonic salt
solution and indomethacin in a dose-dependent manner (Abdel-Sattar et al., 2011;
Alharbi et al., 1994).
Many flavonoids such as Quercetin, rutin, Kaempferol, flavone glycoside and
hypolaetin-8-glucoside have been reported to have gastric ulcer protective effects.
Similarly saponins, such as the derivatives of glycyrrhetinic acid and triterpenoid
saponins were also reported to have antiulcer effects in rats. It therefore appears
responsible to suggest that flavonoids and saponins in Caralluma tuberculata may be
totally or partially responsible for its antigastric ulcer activity (Alharbi et al., 1994).
Pregnane glycosides, Penicilloside E displayed the maximum selectivity index (SI
12.04) which was demonstrated by its highest antitrypanosomal potential trailed by
caratuberside C. it was also proved that acylation is required for the antitrypanosomal
activity instead of the of glycosylation process at C-20 (Abdel-Sattar et al., 2009).

32
The methanolic extracts of Caralluma tuberculata N. E. Br. as well as its n-butanol,
chloroform and petroleum ether soluble portion were examined for cytotoxicity
against human diploid embryonic cell line (MRC5) and different other
pharmacological activities. Overall six compounds were extracted from the
chloroform fraction and were further tested for their antimalarial, antitrypanosomal
and antiprotozoal activity. As for the anti-malarial activity, only petroleum ether
soluble fraction demonstrated moderate inhibitory effect (IC507.94l g/mL), this
fraction showed high cytotoxicity on MRC5 (IC50 0.8l g/mL) (Waheed, 2001).
Methanolic extract did not exhibited any potential against the trypanosoma as
compare to the petroleum ether extract that demonstrated reasonable activity (IC50
0.5l g/mL) along with 1.6 selectivity index. In contrast, modest activity i.e. (IC50 3.5l
g/mL) along with selectivity index (17.9) was shown by the extract of chloroform.
Chloroform extract was chosen further for extraction of biologically active six
compounds that exhibit strong potential against malarial and trypanosome (Abdel-
Sattar et al., 2008).
Other members of this genus like Caralluma fimbriata and Caralluma siniaca were
considered to reduce body weight and blood glucose level respectively (Habibuddin et
al., 2008; Lawrence et al., 2004). In “The Wealth of India” (1992) Caralluma
fimbriata was recorded as very useful medicinal plant because as indean tribes used it
act as hypoglycemic agent, weight loss stimulator and suppressant of appetite, pain,
fever and inflammation (Abdel-Sattar et al., 2007; Sreelatha et al., 2010).
Caralluma edulis is also famous for its anti-diabetic potential (Wadood et al., 1989).
Similarly hypoglycemic synergistic effect was noticed when C. edulis and C.
attenuate were used in combination along with extract of phlorizin for reducing blood
and urine glucose level in conjunction with weight loss (Venkatesh et al., 2003). In
Nepal and Sri Lanka, stem of wild Caralluma umbellata are used in stomach
disorders and abdominal pains (Kishore et al., 2010).
Caralluma adscendens is a thick, succulent perennial herb found wild in Africa,
Afghanistan, Ceylon, India, Southern Europe and Saudi Arabia. In India, it grows
naturally in the dry hills of Andhra Pradesh, Warangal and various other districts. It
may also consumed in form of pickle and eaten as vegetable during famine (Tatiya et
al., 2010). It is locally known as “Makadshenguli/Shengulmakad”. The local people
33
use it in raw form for treatment of diabetes and it is also utilized as a vegetable (Mali
et al., 2009). Caralluma attenuata is eaten raw because it acts remedy for diabetes
while its juice along with black pepper is suggested for the cure of migraine (Kumar
et al., 2011). It usually exhibits antinociceptive activity owing to its luteolin-49-O-
neohesperidoside components (Kumar et al., 2011).
Caralluma dalzielii N. E. Br. is a succulent herb occurring wildly on the Sahelian
region of West Africa from Senegal to Nigeria where is used in folk medicine as
antispasmodic and analgesic remedy. Plant latex in Bandiagara area of Mali is used
for wound healing, while the stems are grounded and eaten row as tonic remedy and
for bodily exhaustion and cardiac problems. A previous phytochemical study on this
plant reported the isolation of two tomentogenin esters (De Leo et al., 2005). In short,
worldwide distribution and pharmacological properties of some significant species of
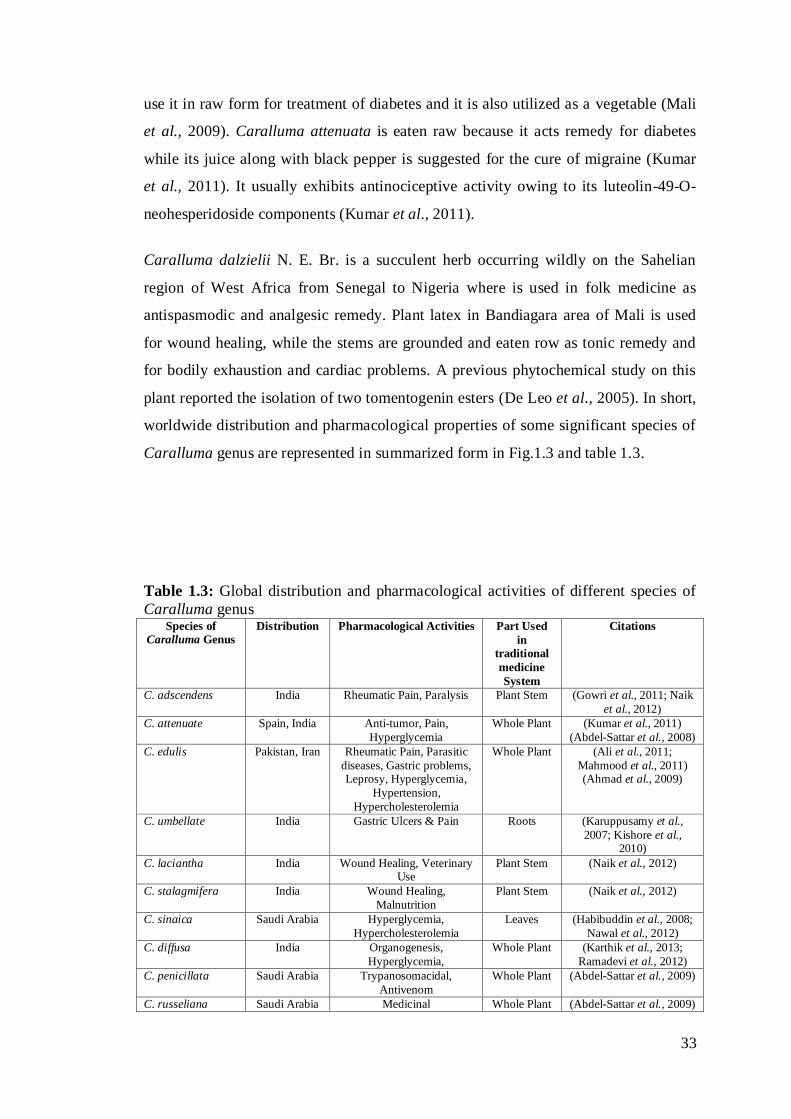
Caralluma genus are represented in summarized form in Fig.1.3 and table 1.3.
Table 1.3: Global distribution and pharmacological activities of different species of
Caralluma genus Species of
Caralluma Genus
Distribution Pharmacological Activities Part Used
in
traditional
medicine
System
Citations
C. adscendens India Rheumatic Pain, Paralysis Plant Stem (Gowri et al., 2011; Naik
et al., 2012)
C. attenuate Spain, India Anti-tumor, Pain,
Hyperglycemia
Whole Plant (Kumar et al., 2011)
(Abdel-Sattar et al., 2008)
C. edulis Pakistan, Iran Rheumatic Pain, Parasitic
diseases, Gastric problems, Leprosy, Hyperglycemia,
Hypertension,
Hypercholesterolemia
Whole Plant (Ali et al., 2011;
Mahmood et al., 2011) (Ahmad et al., 2009)
C. umbellate India Gastric Ulcers & Pain Roots (Karuppusamy et al.,
2007; Kishore et al., 2010)
C. laciantha India Wound Healing, Veterinary Use
Plant Stem (Naik et al., 2012)
C. stalagmifera India Wound Healing,
Malnutrition
Plant Stem (Naik et al., 2012)
C. sinaica Saudi Arabia Hyperglycemia,
Hypercholesterolemia
Leaves (Habibuddin et al., 2008;
Nawal et al., 2012)
C. diffusa India Organogenesis,
Hyperglycemia,
Whole Plant (Karthik et al., 2013;
Ramadevi et al., 2012)
C. penicillata Saudi Arabia Trypanosomacidal,
Antivenom
Whole Plant (Abdel-Sattar et al., 2009)
C. russeliana Saudi Arabia Medicinal Whole Plant (Abdel-Sattar et al., 2009)
34
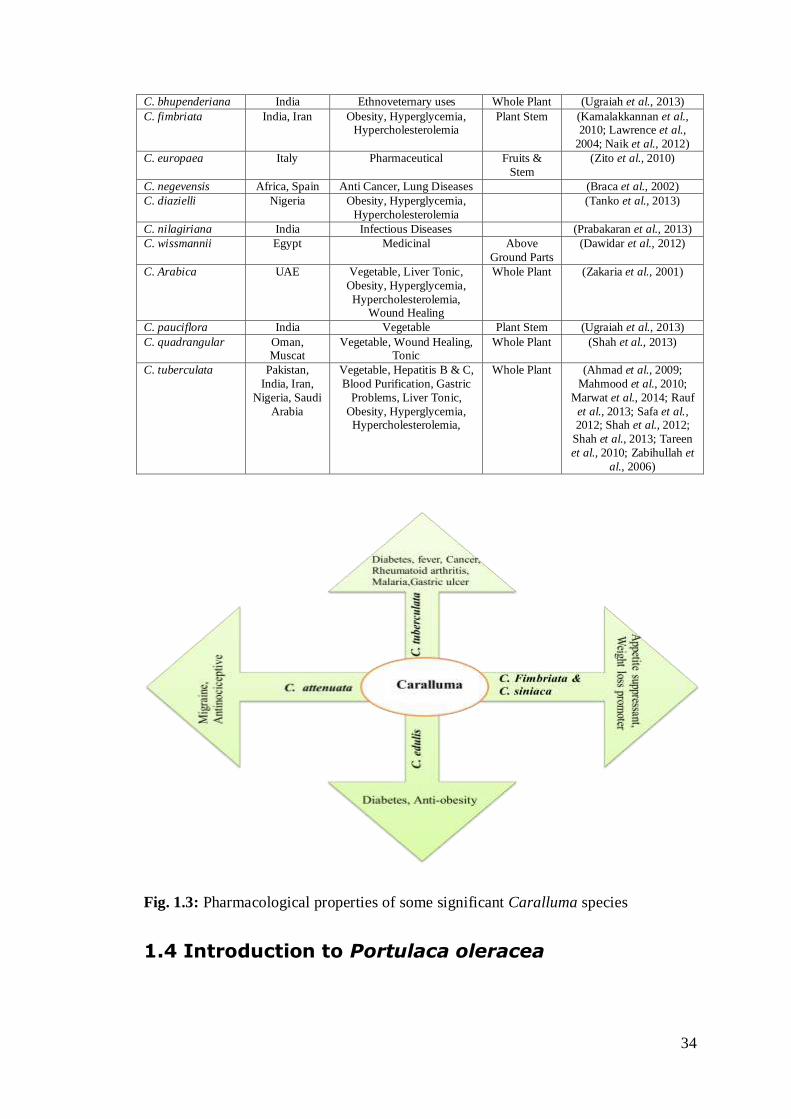
C. bhupenderiana India Ethnoveternary uses Whole Plant (Ugraiah et al., 2013)
C. fimbriata India, Iran Obesity, Hyperglycemia, Hypercholesterolemia
Plant Stem (Kamalakkannan et al., 2010; Lawrence et al.,
2004; Naik et al., 2012)
C. europaea Italy Pharmaceutical Fruits &
Stem
(Zito et al., 2010)
C. negevensis Africa, Spain Anti Cancer, Lung Diseases (Braca et al., 2002)
C. diazielli Nigeria Obesity, Hyperglycemia,
Hypercholesterolemia
(Tanko et al., 2013)
C. nilagiriana India Infectious Diseases (Prabakaran et al., 2013)
C. wissmannii Egypt Medicinal Above
Ground Parts
(Dawidar et al., 2012)
C. Arabica UAE Vegetable, Liver Tonic,
Obesity, Hyperglycemia,
Hypercholesterolemia, Wound Healing
Whole Plant (Zakaria et al., 2001)
C. pauciflora India Vegetable Plant Stem (Ugraiah et al., 2013)
C. quadrangular Oman, Muscat
Vegetable, Wound Healing, Tonic
Whole Plant (Shah et al., 2013)
C. tuberculata Pakistan,
India, Iran,
Nigeria, Saudi
Arabia
Vegetable, Hepatitis B & C,
Blood Purification, Gastric
Problems, Liver Tonic,
Obesity, Hyperglycemia, Hypercholesterolemia,
Whole Plant (Ahmad et al., 2009;
Mahmood et al., 2010;
Marwat et al., 2014; Rauf
et al., 2013; Safa et al., 2012; Shah et al., 2012;
Shah et al., 2013; Tareen
et al., 2010; Zabihullah et
al., 2006)
Fig. 1.3: Pharmacological properties of some significant Caralluma species
1.4 Introduction to Portulaca oleracea

35
Portulaca oleracea, frequently well-known as Purslane in English and Kurfa in
Arabic and Persian, is found all the temperate zones of the globe (Burkill et al., 1997).
The Portulaca name is considered to narrate from the Latin language word “porto”
denotes to hold or carry and “lac” denotes milk overall milky juice holding plant and
it is mentioned in official pharmacopoeias of Mexico, France and Spain (Eduardo et
al., 1995). In traditional Chinese medicine (TCM), Purslane is a natural herb that may
also be considered food during the famine years. Chinese usually dig this feral
vegetable and consume it, even presently still procure it from herbal shops. Numerous
authors elaborated the botanical morphology of Portulaca oleracea from time to time
(Matthews et al., 1993; Mitich et al., 1997; Rydberg et al., 1932). Generally, purslane
is depicted as an annual succulent with prevailing prostrate growth. This annual
herbaceous weed is succulent having 15-20 cm long stem and 6 mm long green flashy
leaf (Chan et al., 2000). Stems are reddish in color, glabrous, and branch radially from
the central axis while Leaves position is alternate or sub-alternate and are succulent
glabrous. Roots consist of many fibrous lateral roots and a long thick taproot. Flowers
are few along with sessile terminal head; Flowers are yellow in colour and noticed in
sunny morning. It is a self pollinated plant whose reproduction is either via fragments
of stem or seeds (Zimmerman et al., 1976).
Domain Eukaryota
Kingdom Plantae Subkingdom
Tracheobionta
Phylum Spermatophyta Subphylum Magnoliophyta Class Magnoliopsida Subclass Caryophyllidae
Order Caryophyllales Portulac oleracea: Whole plant and its flowers Family Portulacaceae
Genus Portulaca L. Specific epithet
oleracea L.
Botanical name
Portulaca
oleracea L.
Fig. 1.4: Taxonomy of Portulaca oleracea L and whole plant along with flower

36
1.4.1 Phytochemistry of Portulaca oleracea
Pursulane is also considered as a good source of alkaloids and five oleraceins
alkaloids (A- E) are reported in this regard (Xiang et al., 2014). Similarly sixteen
phenolic including N-cinnamoyl phenylethylamides, pyrrole alkaloid
(portulacaldehyde), amides and five phenylpropanoid acids were secluded from its
polar extract (Kokubun et al., 2012). In one study five compounds (caffeic acid,
hesperidin, oleracein A, oleracein B, and oleracein E) were extracted from the
ethanolic extract with the help of a range of column chromatography (Yang et al.,
2007). Four monoterpene glycosidic compounds including two novel ((3S)-3-O-(-
D-glucopyranosyl)-3,7-dimethyl-7-hydroperoxyocta-1,5-dien-3-ol and portuloside A)
and two already reported compounds ((3S)-3-O-(-D-glucopyranosyl)-3,7-
dimethylocta-1,5-dien-3,7-diol and (3S)-3-O-(-Dglucopyranosyl)-3,7-dimethylocta-
1,6-dien-3-ol ) were isolated from methanolic extract of Portulac oleracea (Sakai et
al., 1996; Seo et al., 2003). In addition to these perence of allantoin, N,N`-
dicyclohexylurea, β-sitosterol and β-sitosterol-glucoside in the extract of plant aerial
part was also confirmed with the help of spectral and crystallographic data of these
constituents (Rasheed et al., 2004). A notable discovery in phytochemistry was seen
in the form of three types of polysaccharides that exhibited promising antiviral
activities against influenza and herpes simplex viruses. These are an acidic
polysaccharide (1,3-,1,6- and 1,3,6-linked galactopyranosyl & 1,5-linked
arabinofuranosyl), a neutral polysaccharides (arabinoglucomannan) and a pectin
polysaccharide (having galacturonic acid, Galactose, rhamnose and Arabinose) (Dong
et al., 2010). High performance liquid chromatography (HPLC) was also used to
determine the presence of four ccompounds like caffeic acid, p-coumaric acid, ferulic
acid and hesperidin in this plant (Wang et al., 2011).
1.4.2 Pharmacological attributes of Portulaca oleracea
Ancients from most civilizations considered it as a holy herb possessing spiritual,
anti-magical and marvelous medicinal attributes (Grieve et al., 1997). Chinese assume
that it carries “vegetable mercury” (Cantwell et al., 1993). It is consumed as a
vegetable in cooked form and in salad dressing in raw form nearly in all the countries.
Purslane is famous for its unusual therapeutic attributes and has been used as a

37
remedy for oral ulcer, diabetes and urinary disorders. It has powerful analgesic,
antibacterial and antifungal activities (Chan et al., 2000; Oh et al., 2000).
It has been reported that because of high concentration of omega 3 fatty acid it can be
used relaxant for skeletal muscle (Parry et al., 1993). Mechanism of action of
Portulaca oleracea aqueous extract was studied and proved to possess the relaxant
potential of skeletal muscle that may be considered by the interference of Ca2+
recruitment in skeletal muscle. Oral administration of this extract produces relaxation
in rat’s skeletal muscle (Okwuasaba et al., 1987). Of course, Purslane holds high
concentration of poly-phenols and unsaturated fatty acids that are excellent
scavengers and denote a promising antitumor potential (Ebrahimzadeh et al., 2009)
(Ebrahimzadeh et al., 2009; Liu et al., 2000).
Purslane seed oil (PSO) exhibited convincing antitumor characteristic in albino mice
bearing Ehrlich ascites carcinoma (EAC) due to the presence of flavonoids content.
Hussein et al. administered PSO and 5-Flourourasil, both separately and in mixture to
the EAC induced mice for 21days and observed reduction in tumor volume along with
notable improvement in life span and different biochemical parameters (Hussein et
al., 2014).
Omega-3 fatty acid occupy a significant position for anti-carcinogenic prospective of
Portulaca oleracea due to its different role. It can rationalize the accessibility of
Linoleic acid. Moreover it also acts as a competitive inhibitor of Ω-6. Both LA and Ω-
6 are essential materials required directly or indirectly in cancer metabolism. Ω-3 fatty
acid causes the un-saturation in membrane of cancerous cell that cause disintegration
and also slow down the growth of cancerous cell by the activation of gene that start
the process of apoptosis. Ω-3 (fatty acid) has inhibiting upshot on spur of ras
P21cancer genes. This put a stop to the proliferation by preventing the attachment of
cancerous cells onto the basement membrane and production of collagenases. Clinical
findings decipher that Ω-3 fatty acid can also prop up the effect of chemo and
radiotherapy and enhance recovery process in cancer patients after operation.
According to Simopoulos and Robinson, who mentioned in their book “The Omega
Diet”, that people having modern unhealthy lifestyle consuming diet especially
modern western diet having Ω-6 fatty acid contents 20 times more than that of Ω-3

38
fatty acids in contrast to the conventional asian diet. Ratio of cancer mortality is
higher in western countries than eastern countries (Simopoulos & Robinson, 2002).
The functioning of peripheral and central nervous system is directly affected by the
concentration of Portulaca oleracea. When the Portulaca oleracea ethanolic extract
was intra peritonealy administrated in mice, it reduced the locomotive action and
increases the muscle relaxant potential and antinociceptive activity (Radhakrishnan et
al., 2001).
In an automatic single-cell bioassay system the alcoholic and aqueous extracts of this
plant exhibited antimicrobial potential especially against different species of gram
negative bacteria and antifungal action against the growth of selected fungal hyphae
like fungi Aspergillus and Trichophyton and the yeast Candida (Banerjee et al., 2002;
Oh et al., 2000). Fungitoxicity of aqueous and organic solvent (e.g. hexane, ethanol
and chloroform) extracts were tested against different species of fungi (Banerjee et
al., 2002). Ethanolic extracts of aerial parts of Portulaca oleracea also exhibited
noteworthy analgesic and anti-inflammatory attributes (Chan et al., 2000; Islam et al.,
1998; Zakaria et al., 1998). It also has the potential to stimulate the healing process of
the wound by minimizing the wound surface area and enhancing its tensile strength
(Rashed et al., 2003). The aqueous extracts of the aerial part of Portulaca oleracea
resulted relaxation of rabbit jejunum and guinea pig fundus in a dose dependent
manner. Moreover extract also created a dosage dependent pressure reaction on rat
blood pressure (Parry et al., 1993). In contrast daily uptake of Portulaca oleracea
seeds extract showed anti-fertility action on male albino rat’s reproductive organs. It
induced an effective impairment of spermatogenesis (Verma et al., 1982).
In the present decade, Purslane polysaccharides were also utilized to treat burns,
headaches, liver, stomach and intestinal ailments, cough, shortness of breath and
arthritis (Chen et al., 2010; Dong et al., 2010; Li et al., 2009). Polysaccharide from
pursulane can also assuage fatigue persuaded by mice forced swimming. The plant
positive potential was noticed to enhance the activity, timing of mouse by increasing
concentration of hepatic glycogen along with decreasing concentration of blood lactic
acid and serum urea nitrogen (Jingrong et al., 2009).

39
Portulaca oleracea also has the ability to alleviate the oxidative stress provoking by
the deficiency of vitamin A. Phenolic alkaloids are the latest class of antioxidants
discovered in this plant (Yang et al., 2009). Alcoholic and aqueous extracts of the
Portulaca oleracea has also the potential to hamper the gastric abrasions persuaded
by ethanol or hydrochloric acid. By enhancing the dose of both extracts in mice, the
severity of ulcers and reduces their effects on the secretion of gastric acid were also
calculated. Therefore, Portulaca oleracea is used in folk medicines due to its
gastroprotective action (Karimi et al., 2004).
Portulaca oleracea boiled aqueous extract causes a momentous increase in pulmonary
action. The results of the study showed that it is relatively potent, but transitory
bronchodilatory outcome on the asthmatic airways (Malek, Boskabady, Borushaki, &
Tohidi, 2004). The plant is commonly utilized to remove intestinal worms due to the
bioactive compounds present in it. This plant provides efficacious treatment for
controlling intestinal parasite loads (Quinlan et al., 2002). Traditionally Portulaca
oleraceae is seen as one of the most familiar herb for solving urinary problems. It is
safe and effectual to facilitate stable clinical trials (Lans et al., 2006).
The portulaca oleracea extract has hypoxia neuroprotective effects (Wang et al.,
2007). It enhances the expression of protein in the mouse cortices by raising levels of
ATPs in cortices. In mouse it also decreases the brain inflammation (Dong et al.,
2005). An aqueous extract of this plant improves the learning behaviour and memory
ability in mice (Hongxing et al., 2007). Portulaca oleracea is also helpful to improve
the disorder of lipid (Xiao et al.,2005). Its highly polar extract shows that, in high
doses, it reduces the activity of blood urea nitrogen (nephrotoxicity indicators)
(Karimi et al., 2010). Hydroalcoholic extract of Portulaca oleracea can also be
considered for the cure of hypercholesterolemia (Movahedian et al., 2007). An
Ethanolic extract of this plant also shows hepatoprotective activity in rats (Elkhayat et
al., 2008). The oral administration of the homogenates of Portulaca oleracea reduces
the blood-sugar level of alloxan-diabetic rabbits to normal (Akhtar et al.,).
Another exiting application of Portulaca oleracea is its land remedial action as it is
used for landfill leachates and removal of nerotoxic compounds like bisphenol A
(industrial effluent contaminant) from the waste water (Imai et al., 2007).

40
1.5 Hepatitis
The term hepatitis generally refers to the condition of liver inflammation that may
self- limiting to severe chronic form along with liver cirrhosis and carcinoma (Ryder
et al., 2001). Different types of viruses responsible for different types of Hepatitis
along with some other contributing factors like autoimmune diseases, alcohol,
nonalcoholic fatty liver disease (diabetes, obesity, hyperlipidemia and metabolic
disorders, etc.), certain drugs (acetaminophen, antibiotics and central nervous system
drugs), organic solvents and some herbal medicines etc. (Ghabril et al., 2010). There
are six categories of hepatitis depending upon the type of hepatic virus like hepatitis
viruses A to E and G.
In addition to this, hepatitis type may be Ischemic and Giant cell hepatitis. Ischemic
hepatitis appears due to any sort of liver injuries caused by insufficient supply of
oxygen or blood. This condition is frequently associated with heart failure, sepsis or
shock. As a result the level of Alanine transaminase (ALT) and aspartate transaminase
(ASP) enzymes elevated (Masuoka et al., 2013). A rare type of Giant cell hepatitis is
usually observed among newly born babies in which multinucleate giant cells present
in liver. The actual cause of this hepatitis is not known, but it is said to be associated
with autoimmune disorders, drug toxicity and viral infections (Raj et al., 2011).
1.6 Hepatitis C and its Virus
Hepatitis C virus (HCV), as a causative driving force of non-A, non-B hepatitis, was
first time discovered in 1989 which widely spread throughout the world population.
HCV contagion is a global health dilemma in both developed and developing
countries. More than 190 million individuals are infected worldwide and more than 70
% of the personnel’s’ results in the development of the liver diseases which are
chronic, more than 15 % develops hepatocellular carcinoma and about 20 % develop
liver cirrhosis. The rate of chronicity of HCV in humans is rationally increasing from
50 % to 80 % and it depends upon the age when infection appears. It was observed in
both the strains (heterologous and homologous) of animal model (chimpanzee) that in
certain cases an immune response was seen in animals during HCV infectivity that
cannot resist against the development of chronic infection (Farci et al., 1994).

41
The prevalence frequency of HCV diversifies significantly throughout the globe; the
rate is low at Western hemisphere while higher at Eastern hemisphere. Overall, highly
HCV prevalent areas are Cameroon, Central African region, South Asia and Western
African (Hamid et al., 2004). Egypt is the country where the occurrence of this
infection was noticed up to 20 % of its total population. Even the highly developed
areas like the United States of America and Europe have 1-2 % of the population that
carries HCV genes. It becomes very hard to find the HCV new infections because of
lack of diagnosis of this infection at acute level.
1.6.1 Transmission of Pathogen and Genotype Prevalence
HCV has a narrow range of tissue tropism and host specificity. In humans, HCV
transmition occurs by direct contact to blood. There are various courses of HCV
transmission for infection like injectable drug users, sexual transmission especially
Gay, people with multiple partners, sex workers, and partners of HIV/HCV co-
infected individuals in developed countries (Sulkowski et al., 2002). On other side,
international standards are not applied in developing countries about blood
transfusion, shaving from barbers, injecting drug users, reuse of syringes, reuse of
needles for ear and nose piercing, tattooing, unsterilized dental and surgical
instruments are the focal cause to transmit HCV. Sharing eating or drinking utensil,
coughing, hugging, sneezing, usual social contact, water or food are not the causative
ways of transmit HCV (Mast et al., 1998). On the other way transmission through
mother to her baby is rare but currently documented (Alter et al., 2007).
On the bases of heterogeneity of genomic sequences, classification of HCV is
categorized into eleven main genotypes (designated 1-11), about 100 different strains
(numbered 1, 2, 3, etc.) and many subtypes (designated a, b, c, etc.) (Simmonds et
al.,1999). The variability is distributed throughout the genome especially non-coding
regions at either end of the genome (5'-UTR and 3'-UTR; UTR-untranslated region)
(Simmonds et al., 1999). Genotype 1-3 is globally distributed. Most common of all
are 1b and 1a which is approximately 60 % of all world infectivity and prevail
abundantly in North America and Northern Europe, then in Eastern and Southern
Europe and subsequently in Japan. Type 1 is more commonly represented than type 2.
Genotype 3 is distributed diversely in various countries and widespread in South East
Asia. Type 4 primarily found in central Africa, Egypt and Middle East. Genotype 5 is

42
discovered in vicinity of South Africa, and type 6-11are spread in Asia (Houghton et
al., 1991; Mondelli et al., 1999; WHO, 1999). It is debate able point that weather the
genotype of virus has any effect on pathogenicity of disease or not. There are other
factors observed in patients and are characteristic of disease, such as immunological,
genetic, and environmental that cause variation in development of disease (Mondelli
et al., 1999). For the treatment of viral disease, it is significant to know about the
genotype and its specific response against medication. Type 2 and 3 give more
efficient responses, type 1 connected with less effective output solely for interferon.
In advanced gold parameters of therapy - the combined form of ribavirin and
pegylated interferon-α, all types give actively enhanced response.
1.6.2 HCV Structure and Genome
The structural details of HCV infectious viruses remained insufficient for a longer
period of time because it was very hard to maintain the cultivation of HCV viruses in
cell culture systems which are important for producing adequate numbers of virions
for thorough exploration under electron microscope. Furthermore, virus particles
derived from serum or plasma are connected with low density lipoprotein (LDL) of
serum, due to this reason the isolation of viruses from centrifugation becomes so
difficult (Thomssen et al., 1992). It has been observed that those HCV viruses that
were extracted from the cell cultures are having spherical envelope comprising dimer
or tetramer of two HCV envelope glycoproteins (E1 and E2) (Heller et al., 2005;
Wakita et al., 2005; Yu et al., 2007). A spherical shaped structure has been observed
on inner side of virions which represents core (nucleocapsid) that harbors core
proteins and viral genome (Wakita et al., 2005).
The positive RNA genome contains 3’ non-translated region (NTR), a single open
reading frame (ORF), internal ribosome entry site (IRES) and 5’NTR are significant
for efficient RNA replication. The 5' untranslated region (UTR) of HCV having 341
nt is highly conserved part of genome because of stem loop and pseudo-knot
containing four structural domains and is placed at upstream of open reading frame
(ORF) translation starting codon (Brown et al., 1992). A pseudo knot and number of
step loops are present in four advanced structural domains, named I –IV domains.
Very first core coding part of 12-30 nt form the IRES along with II to IV domains
(Honda et al., 1996).

43
The HCV IRES form the smooth pre-initiation complex by the direct attachment of
40S subunit of the ribosome, without any involvement of initiation factors of
canonical translation. Binding of the subunit is the crucial stage, which contributes to
formation of HCV polyprotein translation (Forton et al., 2002; Lerat et al., 2002).
About 225 nt are present in 3'UTR and these are arranged in three regions as
following, a highly conserved 3'-terminal stretch of 98 nt (3'X region) containing
three stem-loop structures SL1 to SL3 and an elongated poly(U)-poly(U/UC) tract and
a variable part of about 30-40 nt. (Kolykhalov et al., 1996). The 3'UTR in addition to
the four stable stem-loop structures is located at the 3' terminal coding sequence of the
NS5B and with the NS5B RNA dependant RNA polymerase (RdRp). While the 52
upstream nt of the poly (U/C) tract and the 3'X region were seen significant for RNA
replication, although viral replication increased by remaining sequence of the 3'UTR
(Friebe et al., 2002; Ito et al., 1996).
Proteolytically progression of polyprotein is done by enzymes; viral and cellular
proteases, into ten particular proteins, comprising of four structural proteins such as
NS5B, NS5A, NS4B, NS4A, NS3 and NS2 and remaining non structural (Lindenbach
et al., 2005). Four viral enzymes encoded by these NS genes are as follows: NS3.4A
serine protease, NS2.3 cysteine protease, NS5B RNA-dependent RNA polymerase
and NS3 RNA helicase. These all four enzymes are vital for infectivity (HCV
replication) in animal model (Kolykhalov et al., 2000). In general two enzymes
considered as striking goals for developing an oral drug for HCV, these enzymes are
NS5B RNA-dependent RNA polymerase and NS3.4A serine protease.
1.6.3 HCV Non-Structural and Structural Proteins
1.6.3.1 Structural proteins
1.6.3.1.1 Core protein
Core protein is a membrane bounded present in the cytosol and found to be coupled
with mitochondria, lipid droplets endoplasmic reticulum and nucleus. As a result of
these interactions of core protein, it is considered to be involved in various pathways
like apoptosis, cell signalling, and transcription of genes and metabolism of lipid
(Tellinghuisen et al., 2002). Core proteins have direct or indirect relation with
hepatocellular carcinoma and hepatitis (Hope et al., 2002; Lerat et al., 2002).

44
Nucleocapsid of HCV comprises conserved core protein, which is folded into three
domains. All domains differ on the basis of hydrophobic interactions like domain one
has two small hydrophobic regions of 117 amino acids. Domain two is comparatively
less basic while 3rd domain is highly hydrophobic among all and also carrying signal
sequences of E1 (envelope) protein (Bukh et al., 1994).
1.6.3.1.2 Envelope protein
Next to the core protein, highly glycosylated two envelope proteins E1 and E2 are
present that have significant roles during cell entry. The conserved glycosylated site
of envelope proteins is rich in mannose concentration. The weight of E1 is 35 kDa
and has five N-linked glycpsylation sites. Its role is as fusogenic subunit (Bartosch et
al., 2003). The molecular weight of E2 is 72 kDa and having eleven N-linked
glycosylation points that directly interlink with cell surface receptors which support
entrance the virus in the host cell (Nielsen et al., 2004). Moreover owing to its hyper
variable regions of amino acids (HVR1 and HVR2) it is highly susceptible to
mutations and thus acts as one of the contributing factors of HCV genotype
variability. Conserved epitopes of B-cell and T-cell in E2 proteins of HCV will a
potential target for developing anti-HCV vaccine in the future (Idrees et al., 2013).
1.6.3.1.3 P7 Protein
P7 is a transmembrane protein, having 63 amino acids, whose N-terminal domain is
toward cytoplasmic region and C-terminal domin is suspended in Endoplasmic
reticulum. It forms a bridge between the genes of envelope protein (E2) and
nonstructural protein (NS2). Its role considered as signal sequence which stimulate
NS2 translocation into the lumen of endoplasmic reticulumn. Furthermore, ion
channels formed by this protein facilitate the virus infection (Griffin et al., 2003).
Some characteristics of P7 are similar to viroporin. P7 role also considers significant
during assembly and the liberation of virons according to specific genotype
(Steinmann et al., 2007).
1.6.3.2 HCV non-structural proteins
1.6.3.2.1 NS2

45
HCV non structural protein NS2 is a trasmembrane protein of 23kDa which is
considered very important for the in vitro and in vivo completion of the HCV
replication cycle (Pietschmann et al., 2006). N-terminus hydrophobic residues of NS3
forming 3-4 transmembrane domains enter into the membrane of the endoplasmic
reticulum. The N- and C-terminus of NS2 (having two active sites and reside in the
cytoplasm) plays an important role autoproteolytic action of NS2/NS3 (Grakoui et al.,
1993; Lorenz et al., 2006). NS2-3 also called metalloprotease because Zinc metal
stabilizes the structure of NS3 at its active site. Moreover Zinc stimulates the protease
activity (Reed et al., 1995).
1.6.3.2.2 NS3
The NS3 is chymotrypsin-like serine protease that has the catalytic triad of amino
acids (Ser-1165, Asp-1107 and His- 1083). It belongs to the trypsin/chymotrypsin
protease superfamily. Replacement of Ser-1165 and His-1083 with another amino
acid such as alanine stops the cleavage of NS3 of HCV polyprotein without
interfering with NS3 structure (Bartenschlager et al., 1994; Grakoui et al., 1993b).
NS3 can cleave consensus sequence of decapeptide (DXXXXC-SXXX) due to its
wide range of specificity for the substrate (Grakoui et al., 1993b). The substrate-
binding pocket can keep six amino acid residues. Sometimes, when the substrate adds
10 amino acid residues, its cleavage proficiency becomes best which suggest that at
the surface of NS3 the substrate interactions occur. At the N-terminus, the last 185
amino acids of HCV NS3 protease engaged in the cleavage involving NS3, 4A, 4B,
5A and 5B (Bartenschlager et al., 1994). The short consensus sequence of NS3
protease connected with the catalytic subunit of protein kinase A (PKA). This leads to
PKA catalytic subunit retention in cytoplasm that prevents its entrance in nucleus.
PKA alters the target protein function by adding phosphate groups, thus regulating
intracellular signaling (Borowski et al., 1997).
NS4A is a 54 amino acid viral protein which binds to the protease domain as a
cofactor (Satoh et al., 1995). NS4A has two roles. It supports NS3 to membranes and
it stabilizes the structure of NS3, as a result protease is activated (Urbani et al., 1999).
NS3 has a structural zinc ion that plays a significant role for domain of protease NS3
as well as auto-protease (NS2-3) (Love et al., 1996). This zinc ion is coordinated with

46
NS3 by three cysteine residues and one water molecule (Satoh et al., 1995). It is
demonstrated through studies that only one fragment is needed for full protease
activation. In vitro which is small, predominantly hydrophobic 13 amino acid long
fragment of NS4A that binds directly to NS3 (Tomei et al., 1993). All of the
evaluated studies have shown that the substrate specificity of NS3 protease has used
steady-state kinetics and parameters such as kcat, Km, and kcat/Km to define this
specificity.
It is important to note through studies by De Francesco and co-workers that some Pn-
P1 product peptides show substantial inhibitory activity which can be enhanced by
amino acid substitution. Thus, it is feasible that for the N-terminal product peptide,
the NS3 protease affinity might amend the rate-limiting step in fixed circumstances.
The base of catalytic/chemical mechanism of NS3 protease-catalyzed cleavage of
peptide bonds is to maintain the catalytic triad of NS3, similar to another enzyme
which is serine proteases. It was studied that the NS4A is present usually in complex
form with 3-dimensional structure of NS3 serine protease (Kim et al., 1996; Love et
al., 1996; Yan et al., 1998). The presence Oxyanio hole and catalytic triads are the
main requirements of protease activity. Oxyanion hole basically constitutes the
backbone amides of Ser-139 along with Gly-137 while catalytic triad is a combination
of Asp-81,Ser-139 and His- 57. In the proper location of this catalytic triad and the
substrate, there exists major involvement of NS4A (Bartenschlager et al., 1993;
Grakoui et al., 1993a; Tomei et al., 1993). His-57 is known to act as a general base to
activate the Ser-139 nucleophile. When the carbonyl carbon of the scissile bond of
Ser-139 exemplify nucleophilic attack, it escorts the development of tetrahedral
transitional state having an oxyanion which becomes stable by this oxyanion hole of
the enzyme at once when a substrate binds to it.
The NS3-NS4A protease is regarded as the most peculiar target for anti-HCV drugs.
The HCV antiviral drugs posses two enzymatic components of the replicase that are
the most popular targets for their development; the NS5B RdRp and NS3- NS4A
serine protease.
These are attractive in part because both these enzymes are indispensable for HCV
replication and used for the development of biochemical assays for inhibitor
screening, which is an important consideration in the era particularly before cell-based

47
systems (Pawlotsky et al., 2006; Pawlotsky et al., 2004). In recent times, to provoke
the dsRNA-dependent interferon regulatory factor 3 (IRF-3) pathways, the HCV NS-
3NS4A were shown in vitro that is essential arbitrator of interferon associated
response during viral infection (Foy et al., 2003).
1.6.3.2.3 NS4A and NS4B
NS4A protein of HCV RNA comprises 54 amino acids and work as cofactor for the
activity of NS3 due to its N-terminal hydrophobic residues (Wolk et al., 2000). It is
assumed that last twenty amino acid residues of NS4A make a helix that provide an
anchoring surface for binding of the complex of NS3/NS4A onto the membrane of
endoplasmic reticulumn (Kim et al., 1996). The posphorylation activity of NS5A is
also modulated by NS4A which is confirmed by deletion analysis of NS5A central
amino acids (2135 to 2139) (Asabe et al., 1997).
NS4B is considered very important because recruitment of different proteins of HCV
dependent upon it. NS4B interlinked with NS4A thus indirectly regulate its function
(Lin et al., 1997). Molecular weight of NS4B is 27 kDa and it comprises four
transmembrane helixes of hydrophobic amino acids (Lundin et al., 2006). Being an
essential membrane protein it is found to be restricted along with remaining non
structural proteins inside the endoplasmic reticulum (Hugle et al., 2001). Inside
endoplasmic reticulum a membranous plexus is formed because of the action of
NS4B. All the viral proteins were gathered here and form the replication complex
(Egger et al., 2002).
1.6.3.2.4 NS5A and NS5B
NS5A always remain a potential target for anti-HCV therapies due to its significant
role in replication, apoptosis, modulation of different pathways of cell signaling and
enhancement of interferon response. Cell signaling pathways like ROS (Reactive
oxygen species), MAPK (mitogen-activated protein kinase) and
PIK(phosphatidylinositol 3-kinase signaling) pathways lead to hepatocellular
carcinoma and modulation of hepatocytes are regulated directly and indirectly by
NS5A (Macdonald et al., 2004). NS5A have interferon-α sensitivity-determining
region (ISDR) which interacts with protein kinase receptor i.e. gene product
stimulated by interferon- α (Enomoto et al., 1995). This region causes resistance

48
during treatment with interferon (Gale et al., 1997). Being a hydrophilic phospho-
protein it does not span across the membrane like other non structural proteins of
RNA. A distinctive amphipathic α-helix present at the N-teminal of NS5A helps its
association with membrane and presence in replication complex (Penin et al., 2004).
Any sort of mutation in α-helix region totally disrupts the replication of HCV. Such
sort of mutations is essential for creating replicon cell line (Elazar et al., 2003;
Lohmann et al., 1999).
An important RNA dependent RNA polymerse (RdRp) i.e. NS5B involves the
production of new strand of RNA. It molecular weight is 65 KDa. It is prone to error
due to absence of proof reading ability and thus contributes in genetic diversity of
HCV. NS5B hydrophobic profile indicates that at its c-terminus sequence of 21 amino
acids present that involve in membrane penetration (Yamashita et al., 1998).
Structural confirmation decipher that this polymerase having the shape like ‘right
hand’ in which active cleft of palm completely encircled by two sub domains appear
with shape like thumb and finger (Lesburg et al., 1999). Its aspartic acid residues
combine with metallic divalent ion in order to facilitate the attachment of primer and
polymerization of priming nucleotide along with the release of pyrophosphate
product. It is also proficient for RNA ‘de novo’ synthesis in the absence of primer.
Due to the central role of NS5B it appears as notable target along with NS3 Serine
protease for antiviral therapies of this millennium (De Francesco et al., 2005;
Moradpour et al., 2005).
1.6.4 HCV RNA Replication and Potential Drug Target
The high level of diversity is a result of both the error prone RNA dependent RNA
polymerase (RdRp), which is lacking a proofreading functional, and higher replication
rate of HCV, together leading to a very high mutation rate (Lindenbach et al., 2005).
This remarkable degree of variability manifests itself in that within an infected host
HCV exists not as a single genetic homogenous virus, but rather a population of
closely related but yet distinct virions, referred to as the “quasispecies swarm”
(Pawlotsky et al., 2006). Within the swarm exist many preformed variants that often
resistant to DAAs and these emerge when the drugs are present. Resistance to DAAs
will likely be the major challenge of HCV therapy in years to come.
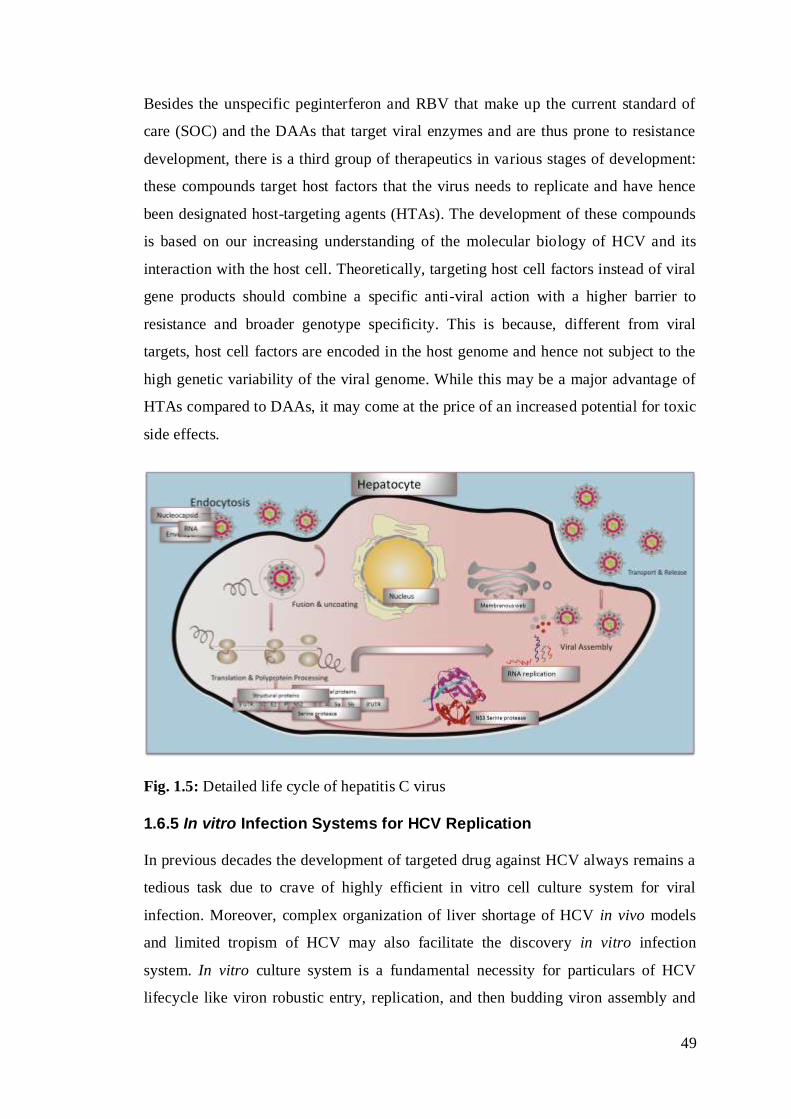
49
Besides the unspecific peginterferon and RBV that make up the current standard of
care (SOC) and the DAAs that target viral enzymes and are thus prone to resistance
development, there is a third group of therapeutics in various stages of development:
these compounds target host factors that the virus needs to replicate and have hence
been designated host-targeting agents (HTAs). The development of these compounds
is based on our increasing understanding of the molecular biology of HCV and its
interaction with the host cell. Theoretically, targeting host cell factors instead of viral
gene products should combine a specific anti-viral action with a higher barrier to
resistance and broader genotype specificity. This is because, different from viral
targets, host cell factors are encoded in the host genome and hence not subject to the
high genetic variability of the viral genome. While this may be a major advantage of
HTAs compared to DAAs, it may come at the price of an increased potential for toxic
side effects.
Fig. 1.5: Detailed life cycle of hepatitis C virus
1.6.5 In vitro Infection Systems for HCV Replication
In previous decades the development of targeted drug against HCV always remains a
tedious task due to crave of highly efficient in vitro cell culture system for viral
infection. Moreover, complex organization of liver shortage of HCV in vivo models
and limited tropism of HCV may also facilitate the discovery in vitro infection
system. In vitro culture system is a fundamental necessity for particulars of HCV
lifecycle like viron robustic entry, replication, and then budding viron assembly and

50
their release and afterward development of therapeutic accordingly (Wilson &
Stamataki, 2012). However in present decade notable efforts have been made in this
regard e.g. HCV Replicons, HCV Pseudoparticles, Cell-Culture-Derived HCV
(HCVcc), Primary Cell Culture and Immortalized Primary Hepatocytes, Hepatoma
Cell Lines (Hep3B cells, PLC/PRF/5 cell, Huh-6 cells, Huh-7 clones : Huh-7, Huh-
7.5, & Huh-7.5.1) and liver slices.
1.6.5.1 Sub-genomic replicons system of HCV
Sub-genomic replicons (total RNA or genome elements of structural & non structural
proteins) are high level replicas of HCV genomes that were studied in a Huh-7 cell
system (Blight et al., 2000). Replicon constructs of HCV having the ability to
replicate independently in various hepatoma cell lines and used for identification of
adaptive mutation and resistant cell types. All types having a gene for neomycin
phosphotransferase used for selection.
Bi-cistronic replicon having Encephalo myo crditis virus (EMCV) internal ribosome
entry site (IRES) prior to the onset of genes of non-structural protein. T7 promoter
position is next to all these above mentioned genes in RNA transfected in Huh-7 cell
lines. In the presence of G418 antibiotic, RNA replicates rapidly to form colonies
(Fig. 1.6). Interferon resistance originated by adaptive mutations that boost up the rate
of HCV RNA replication (Bartenschlager, 2002). Transit assay replicon system is
used for identification of these mutations in which gene of luciferase or neomycin
phosphotransferase replaced the gene of phosphoyransferase. Such two mutations are
noticed in NS3 and one mutation in NS5A (Blight et al., 2000; Lohmann et al., 2003).
In short, viral and host signaling require for replication can be studied by this in vitro
system but this system fail to answer that how virus enter into host cell and its
assembly process.

51
Fig. 1.6: The HCV subgenomic replicon system
1.6.5.2 Hepatitis C Virus pseudoparticals (HCVpp)
This model system helps to study initial phases of viral life cycle like attachment to
host cell membrane and then internalization. HCVpp are produced as a result of
transfection of 293T cells (embryonic cells of human kidney) by three vectors. The 1st
vector encode genes of Pol and Gag proteins and liable for budding of particles at
plasma membrane and encapsulation of RNA while 2nd vector encode genes of
luciferase reporter protein. The 3rd vector encodes E1 and E2 envelope glycoproteins
that are essential for HCV entry into host cell membrane. These tranfected kidney
cells secrete pseudoparticles that can be utilized to infect Huh-7 cells. Level of
infectivity can be checked by quantification of luciferase or expression of GFP in
hepatoma cells. Furthermore HCVpp also use to recognize Claudin-1 and Occludin
(HCV coreceptors) and neutralize the monoclonal antibody action against heteroogus
HCV glycoprotein (E1, E2) and sera of HCV infected person (Cai et al., 2005; Hsu et
al., 2003). HCV pseudoparticles are also essential to find out fusion mechanism of
virus and also utilized to identify the inhibitors that can block/hamper entry of HCV
(Fig. 1.7).
NEOR 3 4A 4B 5A 5B
HCV IRES EMCV IRES
T7
In vitro Transcription
Removal of DNA
NEOR 3 4A 4B 5A 5B
Transfection
Huh-7 Hepatoma
No RNA No Replication Replication
G418 Selection
Cell Death Cell Death Colony
The HCV The HCV Subgenomic Replicon Subgenomic Replicon SystemSystem
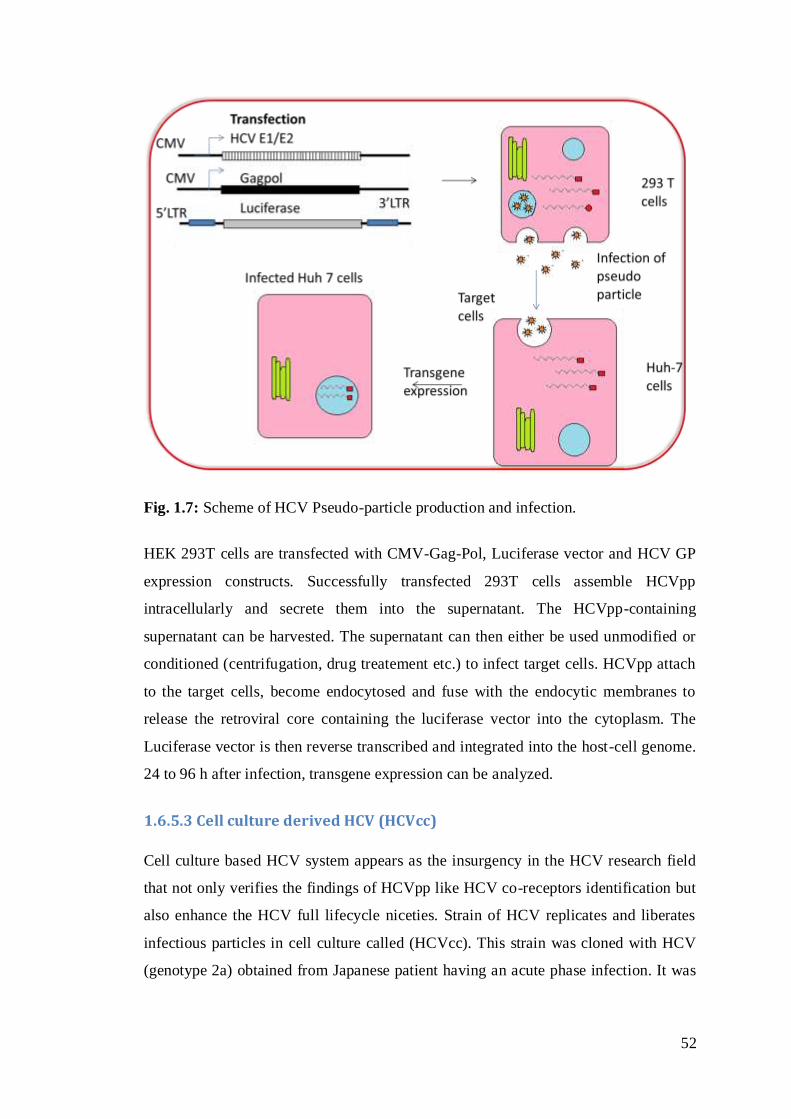
52
Fig. 1.7: Scheme of HCV Pseudo-particle production and infection.
HEK 293T cells are transfected with CMV-Gag-Pol, Luciferase vector and HCV GP
expression constructs. Successfully transfected 293T cells assemble HCVpp
intracellularly and secrete them into the supernatant. The HCVpp-containing
supernatant can be harvested. The supernatant can then either be used unmodified or
conditioned (centrifugation, drug treatement etc.) to infect target cells. HCVpp attach
to the target cells, become endocytosed and fuse with the endocytic membranes to
release the retroviral core containing the luciferase vector into the cytoplasm. The
Luciferase vector is then reverse transcribed and integrated into the host-cell genome.
24 to 96 h after infection, transgene expression can be analyzed.
1.6.5.3 Cell culture derived HCV (HCVcc)
Cell culture based HCV system appears as the insurgency in the HCV research field
that not only verifies the findings of HCVpp like HCV co-receptors identification but
also enhance the HCV full lifecycle niceties. Strain of HCV replicates and liberates
infectious particles in cell culture called (HCVcc). This strain was cloned with HCV
(genotype 2a) obtained from Japanese patient having an acute phase infection. It was

53
also appear infectious within in vivo animal models (chimpanzee and mice) that were
transplanted with human hepatocyte.
1.6.5.4 Immortalized Primary Hepatocytes and Primary Cell Culture
Prime hepatocyte cell cultures obtained from chronically infected chimpanzees or
humans by HCV were exploited for HCV infectivity studies. But such primary culture
are considered best option because of its diverse viral population, reduced level of
HCV RNA replication, presence of HCV specific Abs in patient serum and poor
reproducibility of data.
1.6.5.5 Hepatoma Cell Lines
These hepatic cell clone (Huh-7 clones) are considered as best ideal in vitro system to
date that having high susceptibility for viral infection. The discovery of Huh-7 clones
(Huh-7, Huh-7.5, and Huh-7.5.1) opened a new horizon in research on HCV.
Proficient replication viral genome in Huh-7.5 cells may fairly credited to a major
defected retinoic-acid-inducible gene-I (RIG-I) pathway of antiviral immune
response. Human hepatocellular carcinoma derived Huh-7 clones are poorly
differentiated; exhibit atypical proliferation, deviant gene regulation, and tainted
signaling pathways initially raised the questions about their physiological implication
to the in vivo setting.
1.6.6 Available Treatment Options against HCV
Interferon boosts up the action innate immune response because it acts like cytokine
that after binding of receptor against HCV regulate tyrosine kinase mediated
phosphorylation cascade. As a result transcriptional factor in nucleus persuade the
synthesis of various antiviral proteins. There are chances that autoimmune diseases
(autoimmune hepatitis or diabetes, etc.) may also be provoked due to immune
modulator action of interferon.
First successful chemotherapeutic against HCV is a guanosine analogue called
ribavirn. This broad spectrum drug reduced the infectivity of HCV not only by acting
against RNA dependent RNA polymerase but also ensuing mutations in HCV
genome. It also acts in synergism by enhancing the immune modulation process of
Interferon by affecting the signalling pathways.

54
During HCV infection oxidative stress, increased that minimizes the antiviral effect of
interferon and ribavirin by jamming the JNK/STAT signalling pathways. But the
limitations that are associated with the protracted use of ribavirin are its side effects
like haemolytic anaemia, insomnia and low grade fever, etc. Other additional
contributing factors are immense viral titer baseline and fibrosis stage, age, alcohol
intake, gender, African and American race, baby boomers, and changed immune
response of the host due to raised levels of interleukin-8 (IL-8) and interleukin-10 (IL-
10) in serum. Moreover vaccine development against still facing hindrance owing to
the capacity to HCV genome for undergoes sequence variation during evasion
strategies.
Further progress is looming on the horizon. Knowledge of the molecular structure of
the hepatitis C proteins has allowed the design of new drugs targeting the sites of
HCV-encoded enzymes that are important for the replication of the virus.
1.6.7 NS3 Gene as Candidate Drug Target
The NS3 is chymotrypsin-like serine protease (Hahm et al., 1995) of 69 kDa that has
the catalytic triad of His- 1083, Asp-1107 and Ser-1165. It belongs to the
trypsin/chymotrypsin protease superfamily. When His-1083 and Ser-1165 are
replaced with alanine, it stopped NS3 cleavage of the HCV polyprotein without
affecting the NS3 protein structure (Bartenschlager et al., 1994; Grakoui et al., 1993).
NS3 can cleave consensus sequence of decapeptide (DXXXXC-SXXX) due to its
wide range of specificity for the substrate (Grakoui et al., 1993a). The substrate-
binding pocket can keep six amino acid residues. Sometimes, when the substrate adds
10 amino acid residues, its cleavage proficiency becomes best which suggest that at
the surface of NS3 the substrate interactions occur. At the N-terminus, the last 185
amino acids of HCV NS3 protease engaged in the cleavage involving NS3, 4A, 4B,
5A and 5B (Bartenschlager et al., 1993). The short consensus sequence of NS3
protease connected with the catalytic subunit of protein kinase A (PKA). This results
in the retention of the catalytic subunit of PKA in the cytoplasm that prevents it
entering the nucleus. PKA alters the target protein function by adding phosphate
groups, thus regulating intracellular signaling (Borowski et al., 1997).

55
NS4A is a 54 amino acid viral protein which binds to the protease domain as a
cofactor (Satoh et al., 1995). NS4A has two roles. It supports NS3 to membranes and
it stabilizes the structure of NS3, as a result protease is activated (Tanji et al., 1995;
Urbani et al., 1999). NS3 has a structural zinc ion that is of significant for the NS3
protease domain and NS2-3 auto-protease (Love et al., 1996). This zinc ion is
coordinated with NS3 by three cysteine residues and one water molecule (Satoh et al.,
1995). It is demonstrated through studies that only one fragment is required for full
activation of the protease in vitro which is small, predominantly hydrophobic 13
amino acid long fragment of NS4A that binds directly to NS3. It is important to note
that some Pn-P1 product peptides show substantial inhibitory activity which can be
enhanced by amino acid substitution. Thus, it is feasible that for the N-terminal
product peptide, the NS3 protease affinity might amend the rate-limiting step in fixed
circumstances. The catalytic/chemical mechanism of NS3 protease-catalyzed cleavage
of peptide bonds is based on the conservation of the catalytic triad which likely
similar to that observed for other serine proteases (De Francesco et al., 2003).
The 3-dimensional structure of the NS3 serine protease domain complexed with
NS4A has been determined (Kim et al., 1996; Love et al., 1996; Yan et al., 1998).
Protease activity requires an oxyanion hole (backbone amides of Gly-137 and Ser-
139) and a catalytic triad (Ser-139, His-57 and Asp-81). In the proper location of this
catalytic triad and the substrate, there exists major involvement of NS4A
(Bartenschlager et al., 1993; Grakoui et al., 1993b; Tomei et al., 1993). His-57 is
known to act as a general base to activate the Ser-139 nucleophile. When the carbonyl
carbon of the scissile bond of Ser-139 exemplify nucleophilic attack, it escorts the
development of tetrahedral transitional state having an oxyanion which becomes
stable by this oxyanion hole of the enzyme at once when a substrate binds to it (Pause
et al., 2003).
The NS3-NS4A protease is one of the most popular viral targets for anti-HCV
therapeutics. The HCV antiviral drugs have been two enzymatic components of the
replicase that are the most popular targets for their development, the NS3/4A serine
protease and the NS5B RNA-dependent RNA polymerase. These are attractive in part
because both these enzymes are required for HCV replication and for the development
of biochemical assays for inhibitor screening which is an important consideration in

56
the era particularly before cell-based systems (Pawlotsky, 2006; Pawlotsky &
McHutchison, 2004). In recent times, to provoke the dsRNA-dependent interferon
regulatory factor 3 (IRF-3) pathways, the HCV NS-3NS4A were shown in vitro that is
essential mediator of interferon induction in response to a viral infection (Foy et al.,
2003). Furthermore, NS3-NS4A comes into view to avert dsRNA signaling through
the toll-like receptor 3 upstream of IRF-3 (Li et al., 2005).
1.6.7.1 NS3 Serine protease as potential HCV inhibitors
Finally, two 1st generation direct acting antiviral (DAA) protease inhibitors
(boceprevir and telaprevir) with their marvelous effect in chronically HCV affected
patent comes in market twenty year following the discovery of virus responsible for
Hepatitis C.
In 2011 FDA approved two peptidomimetic ketoamide protease inhibitors, Telaprevir
(VX-950) by Vertex Pharmaceuticals and Tibotec and Boceprevir, (SCH 503034) by
Schering-Plough, as orally administered DAA drugs that selectively inhibit NS3/4A
protease of HCV (Venkatraman et al., 2006). This peptidomimetic linear inhibitor
along with gold standard care of therapy exhibits marvelous viral cure results in
African-American Hepatitis C patients (62 %) in term of ideal effectiveness and
tolerability. These regime results of these small molecule inhibitors along with
interferon are >70 % in patients affected with genotype II and III and 40-70 % in
remaining other genotypes of HCV. But certain confines like daily three time dose
along with food, side effects like anemia, rashes on skin etc. interaction with other
drugs, genetic barrier aligned with resistance etc. These precincts signify the need of
2nd generation of inhibitors that improvement chances in pharmacokinetics along with
minor side effect.
Second wave of protease inhibitors include Simeprevir (TMC435); BI201335 of
Boehringer Ingelheim Pharmaceuticals, Ingelheim, Germany; Danoprevir/r (RG7277)
of Intermune Pharmaceuticals, Brisband, CA; Asunaprevir (BMS-650032; Bristol-
Myers Squibb, New York, NY; ABT-450/r of Abbott, Abbott Park, IL; Enanta
Pharmaceuticals, Watertown, MA; Sovaprevir (ACH-1625; Achillion
Pharmaceuticals, New Haven, CT; MK-5172 (Merck & Co., Inc, Whitehouse Station,
NJ) and ACH-2684 of Achillion Pharmaceuticals etc. These inhibitors are at the 2nd or

57
3rd phase of clinical trial and offering potent effect with better dosage options along
with bearable side effects.
1.6.8 Ayruvadic Extract and Plant Derived Compounds as anti HCV Drug
Ethnopharmacological data shows that large number of natural products of plant
origin has been used throughout the history for infections caused by viruses like
Hepatitis, Herpes simplex virus, Human immune deficiency virus and influenza virus.
High Throughput screening of the products show the way to the discovery of potential
lead compounds as inhibitors of viral growth during in vitro assays.
In the current decade, numerous traditional plants were analysed scientifically in order
to confirm their utilization in anti HCV therapy. For example Herbal extract from
Acacia nilotica and Phyllanthus amarus abrogate the replication of HCV RNA
(Ravikumar et al., 2011; Rehman et al., 2011). Similarly, some other plant extracts
like Acacia confusa and San-Huang-Xie-Xin-Tang exhibited anti-HCV potential via
suppressing the expression of cyclooxygenase 2 (Lee et al., 2000). Glycyrrhizin
obtained from roots extract of licorice showed potent effect in chronically suffered
hepatic patients by plummeting hepatic cancer (Ashfaq et al., 2011).
Different plant secondary metabolites like flavonoids, lignin and polyphenols were
found to have potential to abrogate the growth of HCV via in vitro and in vivo
analysis. Different flavonoids like Apigenin, Luteolin, Ladanein-BJ486K, Silibinin,
Naringenin, Quercetin, (−)-Epigallocatechin-3-gallate (EGCG) Honokiol etc. and
lignin compounds like Honokiol and 3-Hydroxy Caruilignan C were reported to have
the potential against HCV.
Apigenin and Luteolin appear as inhibitors of NS5B polymerase in cell culture assay
as well as in notional depiction of its molecular attributes used for biological molecule
and ligand interaction. Another flavnoid molecule, Ladanein, that was extracted from
the plant Marrubium peregrinum L. was reported to have anti HCV activity when
used in combination therapy with cyclosporin A. But the mechanism how it prevent
the iron from entry and how it inhibit its replication is not still clear (Haid et al.,
2012).

58
Silibinin of Silybum marianum seed appear as inhibitor of HCV at different stages of
virus life cycle in cell culture like NS3 serine protease and RNA-dependent RNA
polymerase (NS5B) (Ashfaq et al., 2011). Another significant inhibition response was
noticed at place of entry with HCVpp and fusion assay of liposome (Wagoner et al.,
2010). Silibinin Low bioavailability problem was overcome by the injectable
formulation of succinate-conjugated silibinin that exhibited considerable inhibition
along with reduced viral titer (Rutter et al., 2011). Thus the intravenous formulation
of silymarin instead of oral is thought to be very effective (Payer et al., 2010).
The use of Naringenin in nutraceutical is already famous due to its both in vitro and in
vivo attributes like anti-carcinogenic, anti-inflammatory and anti-oxidant. This
flavonoid predominantly present in Grapefruit. As assembly and secretion of HCVs
have some association with metabolism of lipoprotein. An obvious Naringenin dose
dependent inhibition of core protein, HCV + RNA strand and ApoB was noticed in
Huh-7 cells (Nahmias et al., 2008).
Quercetin is derived from fruits, grains, and vegetable and having direct link with
HCV virulence instead of sub-genomic direct inhibition. It was confirmed by co-
immuno precipitation assay and colacalization assay that interact directly with heat
shock proteins-40 (HSP-40) and heat shock proteins-70 (HSP-70). Another significant
effect of Quercetin (50 μM) was noticed in cell culture-based bicistronic reporter
method is the reduced action of Internal ribosomes entry sites (IRES) of HCV both in
presence and absence of NS5A (Bachmetov et al., 2012; Gonzalez et al., 2009).
EGCG, a catechin molecule, of green tea come as another inhibitor of HCV entry into
host cell. Pseudo particles of HCV were utilized to confirm the EGCG effect on HCV
entry. It inhibits the entry of HCV polyprotein regardless to its genotype in human
primary hepatocytes and cell line derived from liver. Another potential of EGCG is its
ability to inhibit cell-to-cell spread, a most important route of transmission of HCV in
the infected liver. For efficient results of complete viral eradication EGCG was used
along with direct acting drugs like cyclosporin A or boceprevir.
Liginin compounds like honokiol extracted from Magnolia officinalis inhibit the viral
entry and NS5A, NS5B and NS3 serine protease during sub genomic replication in
Huh-7 cells culture system along with HCVpp, HCVcc, and replicons of subgenom of

59
HCV 1a and 2a genotype. Similarly 3-hydroxy caruilignan C compound of Swietenia
macrophylla was used for anti-HCV therapy owing to its ability to decrease the level
of HCV RNA. It was also having synergistic effect and utilized in combination
therapy along with IFN α and Telapvir (Calland et al., 2012).
Polyphenol inhibitors of HCV like Excoecariphenol D, Corilagin separated
Excoecaria agallocha L extract and 3-hydroxy caruilignan C extracted from
Swietenia macrophylla inhibited NS3 serine protease with IC50 (in vitro) value 13.5
μM , 37.5 μM and 12.6 μM respectively (Wu et al., 2012).
Natural medicinal flora have been explored for their antiviral potential against
numerous viruses specifically hepatitis B and C viruses, human immunodeficiency
virus, herpes simplex virus and influenza virus because it appear potent in vitro
inhibitor of viral growth in the cell culture. But there are still flurry plants that are in
queue to assess and exploited for therapeutic role against functionally and genetically
diverse and worse families of virus. Nevertheless, these plants can be used to reduce
Hepatitis C virus titer in mammalian cells.
Keeping in view the worth of medicinal flora the present study was designed to screen
the widely growing selected varieties plants against HCV. There are different parts of
this study. Firstly, for this purpose plants was collected from the Soon sexier valley of
Punjab, Pakistan on the basis of available ethnopharmacological data. Alcoholic
extracts of selected plants were first utilized to check their in vitro toxicological
effects in hepatoma cell line by cell counting via hamocytometer and trypan blue
exclusion methods. Then HCV (3a genotype) infected liver cells were employed for
in vitro antiviral screening of these phytoextracts at their non toxic dose and viral titer
was counted with quantitative RT-PCR. After this dose response of promising extracts
were studied in order to find out 50% inhibitory concentration (IC50) and their
synergistic effect if any with interferon alpha 2α. After this results were also verified
against subgenomic non structural protein (NS3). In second half potential extracts
were examined thoroughly for their chemical constituents by qualitative and
quantitative analysis. Then different fractions of these potential extracts on the basis
of polarity were obtained through bioassay guided extraction and theses fractions
were again screen against HCV whole genome and specifically its gene NS3.

60
Thirdly active fractions of extracts were subjected to column chromatography in order
to extract the compounds present in them. Presence of sigle and pure compound in
eluted fractions was confirmed by thin layer chromatography (TLC). Copounds were
identified by the spectral data obtained from UV-Vis spectroscopy, FTIR,
Electrospray ionization mass spectrometry (ESI-MS), one and two dimensional
Nuclear magnetic resonance (NMR).
Findings of present study depicts that two out of one thirteen plants designated as
CTF and POL exhibit anti-HCV activity in a dose-dependent fashion at non toxic
concentration and had synergistic effect when combined with interferon alpha 2a.
Moreover the domino effect corroborate with data, using sera from HCV patients and
HCV sub-genomic replicon. Furthermore, after bioassay guided extraction and
identification of phytochemicals the compounds were isolated from active extract
fractions of both plants by column chromatography. Purified fractions were tested
again for activity against HCV in our in-vitro assay.
1.7 Goals/Objectives
The purpose of this study was to explore new therapeutic agents against target
Hepatitis C virus RNA from natural flora in order to elucidate the control of
pathogenesis of HCV infection. Overall objectives of this project are following:
Main goal of project is in vitro antiviral screening of undocumented Soon-
Valley medicinal plants against HCV whole genome and its NS-3 serine
protease.
Toxicological analysis of all the selected plant extracts in hepatoma cell line.
Investigation of dose response and synergistic studies along with IFNα against
HCV RNA.
Bioassay guided extraction followed by the purification and characterization
of biologically active constituents from selected flora that inhibits HCV RNA
in vitro.

61
Preliminary analysis (qualitative and quantitative) of phytoconstituents in the
selected extracts.
Antioxidant scavenging potential of extracts through DPPH free radical
scavenging assay, ABTS radical cation scavenging assay, Ferric reducing
antioxidant power assay and quantification of phenolic acids by HPLC.
Other pharmacological analysis like antithrombitic and antibacterial potential
of the extracts.
Extraction of compounds from active fractions of plant extract through
column chromatography.
Identification of compounds on the basis of spectral analysis obtained by UV-
Vis, FTIR, and NMR analysis.
2.0 Chemicals, Equipment and Techniques
2.1 Chemicals
The majority of chemicals used were obtained from commercial sources and used as
received without more purification. Analytical grade solvents petroleum ether (60-80
°C), n-hexane, chloroform (CHCl3), ethyl acetate (EtOAc), methanol (MeOH),
acetone and dimethylsulfoxide (DMSO) were used for extraction and chromatography
procedures. For NMR experiments, deuterated solvents were used such as
Chloroform-d1 (CHCl3-d1) and methanol-d4 (CH3OH-d4) were purchased from Armar
Chemicals (Buchs, Switzerland).
2.1.1 Materials for Chromatography
a. Thin layer chromatography (TLC): TLC analyses were carried out on pre-
coated Aluminium-backed TLC plates with 0.2 mm thickness (Merck,
Germany). Different solvent systems were selected for this purpose like C6H6:
EtOAc (90:10, 80:20 and 70:30), C6H6: EtOAc: MeOH (70:20:10),
Chloroform : EtOAc (50:50), EtOAc : MeOH (50:50), EtOAc : MeOH

62
(30:70), CHCl3 : MeOH (95: 5, 90: 10 and 85: 15), EtOAc: MeOH: H2O (80:
10:10) etc.
b. Column chromatography: Silica gel 60 F245 particle size 0.04-0.63 mm 230 -
400 mesh, (Fluka, Buchs, Switzerland) and activated by heating at 105 °C for
one hour prior to utilize for column chromatography.
c. Reagents for identification
Anisaldehyde-sulfuric acid spray reagent: p-Anisaldehyde (0.5
mL) was mixed with 10 mL of glacial acetic acid, followed by the addition of
methanol (85.0 mL) and of concentrated sulfuric acid (5.0 mL). The reagent
was sprayed on TLC plates, which were then heated in an oven at 105 °C until
color development was completed.
Phosphomolybedic acid reagent: Phosphomolybedic acid (10 g) was
dissolved in ethanol (100.0 mL). This solution was sprayed on TLC plates,
which were then dried and heated till the color appeared.
Vanillin reagent: Vanillin (1.00 g) was dissolved in concentrated
sulfuric acid (100 mL). Reagent was sprayed on TLC plates, dried and then
heated in an oven at 105 °C until color development was completed.
Ceric sulfate spray reagent Ce(SO4)2: Ceric sulfate (1.0 g) was
dissolved in concentrated sulfuric acid (10 mL ). The mixture was added to
distilled water to a final volume of 250 mL. The reagent was sprayed on TLC
plates, which were then heated in an oven at 105 °C until colour development
was completed.
Ammonia reagent: Ammonium chloride (1g) was dissolved in
distilled water (100 mL). TLC plates sustaining tests spot were exposed to the
vapors of ammonia appearance of yellow colour spot indicates the presence of
the flavonoid.
2.1.2 Chemicals and Reagents used for Determination of Antioxidant
All sort of the chemicals and reagents utilized in this project were of analytical grade
and procured from Sigma-Aldrich or E. Merck or else specified. Acetone,
Chloroform, Ethanol, Glacial Acetic acid, Hexane, Hydrochloric acid, Iso-octane,
Methanol, Sodium carbonate, Folin-Ciocalteu reagent, 2,2'-azino-bis(3-
ethylbenzothiazoline-6-sulphonic acid)(ABTS), Aluminium chloride, Ascorbic acid,
Butylated hydroxytoluene (BHT), Catechin, 2,2-diphenyl-1-picrylhydrazyl (DPPH),

63
Disodium hydrogen phosphate, Ferric chloride, Gallic acid, Manganese dioxide,
Phenolphthalein, Potassium iodide, Potassium ferricyanide, Sodium dihydrogen
phosphate, Sodium hydroxide, Sodium thiosulphate, Starch, Sodium nitrite,
Trichloroacetic acid.
2.2 Instrument and Apparatus
Analytical balance (Sheimadzu Ay 220), Rotary evaporator, Oven (LabTech. Korea),
Heating mantle, UV-visible spectrophotometer (Perkin-Elmer Lambda-2
Spectrophotometer), Orbital shaker. Soxhlet assembly, conical flasks, beakers, and all
other glassware used in this research work were of A-grade Pyrex.
2.3 Materials used for Biological Activities
2.3.1 Cell Lines
Human hepatocyte derived cellular carcinoma (Huh-7) cell line cells and Chinese
hamster ovary (CHO) cell lines were received as gift from Dr. Zafar Nawaz
(Department of Biochemistry and Molecular Biology, University of Miami, USA) and
Dr. Ahmed Usman Zafar (Biopharmaceuticals lab, CEMB, Pakistan) respectively.
Huh-7 cells were cultured and reserved in Dulbecco's modified Eagle medium
(DMEM) (Sigma, USA) having high concentration of glucose (4500 mg/mL). DMEM
high glucose supplemented with fetal bovine serum (12 %) (Sigma, USA). Media was
made highly selective by the use of antibiotics like streptomycin (100 μg/mL) along
with penicillin (100 IU/mL) (Sigma, USA) prior to use for culturing of Huh-7 cell.
Complete culture medium having inoculums’ of cells was incubated at 37 oC in an
atmosphere of 5 % CO2 in humidified incubator. Similarly CHO cells were cultured in
DMEM Hams F-12 supplemented with fetal bovine serum (10 %), penicillin (100
IU/mL) and streptomycin (100 μg/mL).
2.3.2 Plasmids
Nonstructural genes (NS2, NS3/4A) of HCV genotype 3a along with FLAGtag were
cloned in mammalian expression vector pCR3.1. HCV NS3 gene of genotype 1a

64
along with FLAGtag was also cloned in pCR3.1 expression vector. These vectors
were transformed into competent Top10 cells of E.coli and plasmids were purified by
mini-prep and maxi-prep by using Qiagen DNA purification kit. These vectors
(pCR3.1) were compassionately offered by Functional Genomics laboratory, CEMB.
2.3.3 Reagents & Chemicals
Solvent dimethyl sulfoxide (DMSO) of Sigma Aldrich (Cat # D2650, Sigma) and
MTT reagent (Cat # M565) of PhytoTechnology Laboratories (USA) were used.
Maxima SYBR Green/ROX qPCR Master Mix (2X) (Cat # K0221) and HCV Real-
time PCR Quantitative kit were procured from Thermo Scientific Inc. and Sacace
biotechnologies (Italy) respectively. Housekeeping gene of human tissues i.e.
Glyceraldehydes-3-phosphate dehydrogenase (GAPDH) (Cat # sc25778) was
purchased from Santa Cruz Biotechnology.
2.3.4 Designing of Primers
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene and HCV specific non-
structural (NS3) gene sequences of genotype 1a and 3a were retrieved from NCBI.
The NCBI sequence was transformed into the FASTA format and then primer 3
software, (http://gmdd.shgmo.org/primer3/?seqid=47) was used for designing of
primers. The oligonucleotide primer sequences for selected genes are presented in
table 2.1.
Table 2.1: Primers of GAPDH and HCV non-structural NS3 gene of 1a & 3a
genotype was used in this study Sr. No. Primers Sequences 5’-3’ Length
1 GAPDH F ACCACAGTCCATGCCATCAC 20
2 GAPDH R TCCACCACCCTGTTGCTGTA 20
3 NS3-1a F GGACGACGATGACAAGGACT 20
4 NS3-1a R CCTCGTGACCAGGTAAAGGT 20
5 NS3-3a F GACCATTGTGACCAGCTTGA 20
6 NS3-3a R GCGGGTGACCAAGTACAAGT 20
2.4 Instrumentation & Techniques

65
2.4.1 Physical Constant
Melting points were measured on Stuart Scientific SMP10 apparatus and are
uncorrected. Optical rotation was calculated via JASCO-DIP360 digital polarimeter
2.4.2 Spectroscopy
Infrared spectra were obtained by using ALPHA-P FTIR spectrometer and Mass
spectra (MS) were recorded using a Micromass Platform II spectrometer using an
electrospray ionization source. Nuclear magnetic resonance (1H NMR, 13C, 13C-
DEPT, COSY, HSQC, and HMBC NMR) spectra spectra were recorded at 300 MHz
and 400 MHz NMR on a Bruker DPX 400 spectrometer, expressed in ppm (δ) relative
to trimethylsilane (TMS) as an internal standard. Spectra were recorded after
dissolving sample in CHCl3-d1 and CH3OH-d4 on the basis of their solubility. These
facilities were availed from the Manchester Institute of Biotechnology and
Department of Chemistry, University of Manchester, United Kingdom.
2.4.3 Thin Layer Chromatography
Thin layer chromatography (TLC) is an economical technique utilized for the
separation, preliminary qualitative detection or partly a quantitative visual analysis of
samples. The TLC chromatography technique gives a clue as to how many
components are in a mixture or an extract. It is also used to determine the purity of the
isolated compound. Commercially prepared TLC aluminum sheets of 20 x 20cm
Silica gel 60 F254 was used. The plate was cut to size of 5 x 5cm and the extract was
spotted at the bottom the TLC plate (about 0.5cm from the base). The plate was then
placed in a developing tank containing chosen solvent system. The spots were
developed using DV light, anasaldehyde and iodine vapor.
2.4.4 Column Chromatography (CC)
The preparatory role of column chromatography helps to separate desired pure
compound from crude mixture. The concentration of extracting the compound may
vary from very low high concentration depending upon the column size and type of
mobile phase. Cotton wool plug and small amount of sand was put at the column

66
bottom in order to provide a uniform base to stationary phase otherwise stationary
phase gel will block the column.
The adsorbent which is the silica gel (7~230 mesh size) was weighed using a ratio of
30 gm of the adsorbent to 1.00 gm of the crude. The column was filled with a little
quantity of hexane. Slurry of weighted adsorbent was prepared using 100% hexane
and then carefully poured into the column. Air bubbles were prevented from forming.
A solution of the extract was then pipetted on top of the stationary phase. This layer
was topped with a small layer of sand or with cotton or glass wool to protect the shape
of the organic layer from the flow of the eluting solvents. The eluting solvent initially
was 100 % hexane and the polarity was gradually increase at a 95:5 hexane:ethyl
acetate ratio until a 50:50 ratio was used. About 25 mL of eluting mixture was collect
until there appears to be no more solute in the column. The fractions collected were
monitored by TLC and similar fractions were combined together. Solvent was then
removed from the bulked fraction, allowed to dry and weighed. TLC analysis was
carried out on the semi dry bulked fraction using various polar solvent to ensure the
purity of the fraction.
2.5 Methodology
2.5.1 Biochemical Analysis of Plants
Pharmacological attributes of flora are owing to the potential of its bioactive
constituents. In Pakistan, more than fifty percent of the population relies on plant
directly and indirectly for the treatment of different disorders. By considering the
ethno pharmacological usage, plant assessment for their biochemical constituency is
highly crucial.
2.5.2 Collection, Identification and Preservation of Medicinal Plant Materials
Fifteen plants having medicinal value in folkary was collected from the vicinity of the
Soon-Sakaser valley of Punjab, Pakistan in September to November 2010. Taxonomic
identification was authenticated by Mr. Amin Ullah Shah, Department of biological
science, University of the Sargodha, Sargodha. Voucher specimens were deposited
also at Herbarium, University of the Sargodha, Sargodha. All sorts of information

67
concerning plant scientific names, family names, vernacular uses and parts used from
fifteen medicinal plants are shown in Table 1. Freshly collected plant samples were
rinsed with tap water and then with de-ionized water in order to make it free from dust
and any metal contamination. Then roots and leaves were separated manually from
the stem. All sample materials were chopped into smaller pieces with a sharp steel
knife and dried under shade at room temperature till dryness. Air dried plant samples
were pulverized with an electric grinder into fine, coarse powder, and then the dried,
powdered samples were stored in polyethylene air sealed bags at 4°C for further in
vitro and in vivo assessment.
Table 2.2: Botanicals details chosen for screening against total RNA HCV and its
gene NS-3 serine protease. Sr.
No.
Botanical
Name
Common
Name
Family Local use Part(s)
used
Plant’s
Code
1 Achyranthus
aspera
Prickly
Chaff
Flower
Amaranthaceae Diuretic, purgative,
hepato-protective,
laxative, anti-allergic
Leaves AAL
2 Arjuna
Terminalia
Arjun Combretaceae cardiovascular
ailments, wounds
healing, hemorrhages
and ulcers
Bark ATB
3 Bacopa
monnieri
Brahmi boti Plantagenaceae Ulcers, tumors,
ascities, leprosy,
enlarged spleen,
inflammations,
anemia, biliousness
Leaves BMB
4 Caralluma
Tuberculta
Chunga Asclepadaceae Diabetes, Cancer, Rheumatoid arthritis
Ariel part i.e. stem
CTS
5 Caralluma
Tuberculta
Asclepadaceae Diabetes, Cancer, Rheumatoid arthritis
Roots CTR
6 Chichorium
intybus
Kasni Compositae Gallstone, jaundice,
hypertension
Leaves CIL
7 Chlorophytum
borivilianum
Safed
moosli
Liliaceae Diabetes, sex tonic,
pre and post natal
problems, arthritis
Roots CBR
8 Corchorus
depressus
(Linn.)
Bauphali,
Munderi
Tiliaceae. treachery troubles,
antipyretic, diabetes
laxative, gonorrhea,
Leaves CDL
68
chronic cystitis
9 Myristica
fragrans
Houtt.
Nutmeg,
Jaiphal,
Javitri
Myristiceae Carminative, liver
diseases, respiratory
ailments, tuberculosis
Leaves MFHL
10 Portulacea
oleracea L
Purslane,
Hurfa
Portulacaceae Anticancer, boils,
haemorrhoids,
dysentery
Leaves POL
11 Santalum
album
Sandal Santalaceae colds,
bronchitis, ailments of
gallbladder and liver
Bark SAB
12 Sesamum
indicum
Sesame, Til Pedaliaceae Anticancer, laxative,
emollient
Seed SIS
13 Sonchus
Arvensis
Field milk
thistle,
Dodhak
Asteraceae Gallstone, Gout
haemorrhoid,
hypertension, wound
healings
Leaves SAL
14 Rheum emodi Raiwand
cheeni
Polygonaceae liver diseases, diuretic,
Purgative
Roots RER
15 Zizyphus
Jujuba
Chinese
date, Unab
Rhamnaceae Antiulcer, sedative,
immunostimulant,
anti-inflammatory
Seeds ZJS
2.5.3. Plant Extract Preparation
All samples were weighed and immersed in methanol (70 %) separately for overnight
at 38 °C which is the most suitable temperature for enzymatic activity. After 24 hours,
they were filtered through Whatmann filter paper No.42 (125 mm) and cotton wool;
the residue was soaked over again in fresh methanol (70 %). Process of successive
extraction and filtrations was carried out three times. After final filtration, samples
were concentrated in vacuum under reduced pressure using a rotary flask evaporator.
Greenish brown gummy solid of crude methanolic extracts were obtained which is
then dried in desiccator. The final percentage yield of each extract was calculated. All
extracts were stored in pre weighed air tight glass vials for phytochemical analysis at
4 oC in refrigerator.
2.5.4 Screening of Plant Extracts for Cellular Toxicity Analysis via Trypan Blue
Dye Exclusion Method
Hepatoma liver cell line (Huh-7) and standard CHO fibroblast cells were employed as
in vitro models for numerical elucidation of cell viability under the effect of plant
crude extracts. CHO cells were utilized as positive control for cytotoxic analysis of
Phyto extracts of the body cells additional to liver cells. Cell counting was carried out
to determine the 50 % cytotoxic concentration (CC50) of phyto extracts as illustrated
earlier (Ashfaq et al., 2011).

69
2.5.4. 1 Preparation of plant extract stock solution
About 50 mg of every plant organic extract was mixed meticulously in 1.00 mL of
Dimethyl sulfoxide (DMSO) that confers 50 µg/µL stock concentrations. All stock
solutions were sieved by means of 0.22 µm filter and stocked up at -20 ºC.
2.5.4.2 Culturing of Huh-7 and CHO Cell
The cell lines (Huh-7 and CHO) were cultured in Dulbecco's modified Eagle medium
(DMEM) along with Fetal bovine serum (10 %), streptomycin (100 µg/mL) and
penicillin (100 IU/mL), afterward nurtured at 37 ºC along with continuous supply of
CO2 (5 %).
2.5.4.3 Cell viability assay
The crude extracts of all selected plants were scrutinized for their toxicological
assessment against Huh-7 and CHO cell lines by using Trypan blue dye. Liver and
fibroblast cells were seeded in 24 well plates with density of 8×104/well. By
considering the first three wells of culture plate as control, in the remaining wells
different concentrations of the test samples (1.5 µg/mL to 200 µg/mL) were added in
triplica. After 24 h incubation, cells were washed with phosphate buffer saline (1 %
PBS) and then treated with the solution of trypsin EDTA. After trypsanization, fresh
culturing media was added to each week in order to mimick the trypsin activity. Then
suspension of these cells along with trypan blue dye was prepared with 1:1 ratio. 10
µL of the cell suspension was dispensed onto a glass slide and viable cells were
calculated using haemocytometer.
2.5.4.4 Cell proliferation & toxicity assay
Cellular toxicity of plant extracts was also assessed by utilizing enzyme based
methods, one of them known as MTT assay as described previously (Ashfaq et al.,
2011). ‘MTT [3-{4, 5-dimethylthiazol-2-yl}-2, 5-diphenyltetrazolium bromide] assay
is a well-known in vitro method to analyze mitochondrial dehydrogenase activities in
live cells. In metabolically active cells, yellow tetrazoluim dye, MTT, is reduced to
water insoluble purple formazon crystals by NADH. These insoluble farmazon
crystals were dissolved in organic solvents such as DMSO or isopropanol before
measuring absorbance. Absorbance in the visible region is proportional to the number

70
of metabolically active cells. Briefly, in order to valuate cytotoxicity, Huh-7 cells
were plated in 96 wells plate at a density of 2×104 cells/well. After 24 h, cells were
serially diluted in triplicate with different concentrations of plant extracts ranges from
0.8 µg/mL to 200 µg/mL. Control was considered in triplicate by adding DMSO
instead of plant extracts. Plate was incubated for 20 h at 37 oC in an atmosphere of 5
% CO2. After 20 h of treatment, medium was replaced by fresh medium (100 μL) and
30 µL MTT solution (5.00 mg/mL in 1X PBS) was added in each well. Plate was
wrapped with aluminum foil and kept for another 3-4 h of incubation at 37 oC in
humidified atmosphere. After carefully removing medium, 100 µL of DMSO was
added to solubilize formazon crystals in each well. Absorbance measurements of
reduced MTT were taken by microtitre plate reader at wavelength of 570 nm and a
reference wavelength of 620 nm. Proliferation was analyzed as the fraction of
survived treated cells relative to untreated.
Cell viability was calculated by using following formula;
% Cell viability = Treated 570 nm – 620 nm X 100
Control 570 nm – 620 nm
2.6 Anti-HCV Analysis of Compounds in Liver Cells
Huh-7 cells (3×105 cells per well) were cultured in six well culture plate. Cells were
swabbed two times with 1XPBS after 24 hrs. Each well was inoculated with 1×105
IU/ml of HCV-3a genotype infected serum. Specific dose of test compound (least
amount of cell killing) was added in the wells. Total RNA extraction was carried out
by using Gentra RNA extraction kit (Gentra System Pennsylvania, USA) according
to the manufacturer’s directions of 24 h post infection. Afterward, cells were lysed
with cell lysis solution having 5 µL internal control (Sacace Biotechnologies Caserta,
Italy). RNA pallet was dissolved thoroughly in 1 % DEPC (Diethyl pyrocarbonate)
treated water. Quantifications of HCV RNA were carried out by Real Time PCR
Smart Cycler II system (Cepheid Sunnyvale, USA) using Sacace HCV quantitative
analysis kit (Sacace Biotechnologies Caserta, Italy).

71
2.6.1 Antiviral Analysis of Plant Extract against HVC Ns3 Protease via
Transfection Assay
Huh-7 cells were cultured at the density of 3× 105 in 6-well plates for 24 hrs designed
for transfection studies. . After the removal of media, cells were washed thoroughly
with 1×PBS and transfected immediately with the expression vector
pCR3.1/Flag/NS3-NS4A containing NS3 full length gene (Accession # HCV-PK-
HQ202536) and NS4A (Accession # HCV-PK-HM135518) gene kindly provided by
Applied and Functional Genomics lab, CEMB. The cells were co transfected with
pCR3.1/Flag/NS3-NS4A in the presence and absence of 100 μg of plant extract and
interferon along with Lipofectamine™ 2000 (Invitrogen life technologies, Carlsbad,
CA) as per the company's standard procedure. In brief, in 300 μL of DMEM culturing
media LipofectamineTM (9.00 μL for 6-wells) was added and then this media was
incubated at 37 oC for 5.00 mins. Mammalian expression plasmid
(pCR3.1/FLAGtag/HCV NS3) construct (800 ng/well) was added into DMEM (300
μL) separately and incubated at 37 oC for 5 mins. After incubation, both solutions
were mixed thoroughly and again incubated for 25 mins at 37 oC. When culturing
medium in six well plate attain 80 % confluence of cells, it was removed from each
well of the plate and wells were rinsed with 1XPBS. Afterward, 900 uL of fresh
media was poured into each well. After 25 mins, DMEM holding transfection
complex (100 μL) was added carefully to all wells and plate was again incubated in
humidified incubator at 37 oC for 24 h. For ideal transfection outcome, cells were
collected usually after 24 h to 48 h for future gene transcription and expression
studies.
2.7 Gene Expression studies via Reverse Transcriptase PCR (RT-
PCR)
2.7.1 Extraction of Total RNA
After 24 h total RNA was extracted using Trizol reagent (Invitrogen life technologies,
Carlsbad, CA) in accordance with the manufacturer's protocol. In brief, Cells in each
well were washed thoroughly with 1X PBS, lysed by TRIzol® (1.00 mL) and then
transferred to eppendorfs (1.5 mL). About 200 μL chloroform was added into each
well having cell lysate. Samples were overturned carefully with hand for 5.00 mins

72
and centrifuged for 15 mins at 12,000 rpm. After the centrifugation, uppermost
aqueous layer holding RNA was pipette out cautiously into an uncontaminated
eppendorfs that already having absolute isopropanol (500 μL). Material was again
inverted for 3 mins with hand and centrifuged for 10 mins at 12,000 rpm. A little
noticeable lucent RNA pellet become visible and that was washed with ethanol (70
%) and dried up at room temperature. RNA pellet was immersed in 40 μL of Diethyl
pyrocarbonate (DEPC) treated water. Purified RNA was stored for 30 mins at -20 oC.
RNA quantification was carried out by Nanodrop spectrophotometer (Nanodrop ND-
1000, Optiplex, USA).
2.7.2 Synthesis of cDNA
The RNA template (1.00 μg) was utilized to synthesize the cDNA strand by 200 U/μL
M-MLV Reverse Transcriptase (Invirtogen, Life technologies, Carlsbad, CA)
according to company’s protocol via Reverse Transcription System. Shortly, 1.00 μL
of oligodT primer, 1.00 μL of dNTPs and 1.00 μg of RNA were added into DEPC
treated H2O in order to make 12 μL final volume. Reaction mixture was incubated for
5.00 mins at 65 oC and then for 1.00 min at -20 oC. About 4.00 μL of 5X first strand
buffer, 2.00 μL of 0.1M DTT and 1.00 μL RNase free water were added into every
ice-cold reaction mixture tube and then incubated this mixture at 37 oC for 2 mins.
Afterward, 1.00 μL of M-MLV was added, furthermore incubated for 1.00 h at 37 oC
and 70 oC for 15 mins subsequently. Finally sample solution was stored at -20 oC.
2.7.3 Reverse Transcription PCR (RT-PCR) for Gene Expression Analysis
Specific sequence forward and reverse primers for GAPDH (internal control) and
HCV NS3 genes were designed to investigate the effect of plant extract on the
expression of genes through RT-PCR (Applied Biosystems Inc. USA). Primers were
used for the amplification of HCV NS3 and GAPDH genes and their sequence is
already depicted in Table 2.1. Semi-quantitative and quantitative gene expression data
is mostly normalized to expression level of internal control/housekeeping genes.
Housekeeping genes (GAPDH) expression remains unaffected in cells during this
study. GAPDH and β-actin are the most commonly used internal
control/housekeeping genes in comparing gene expression data (Barber et al., 2005).
PCR reaction was carried out by using 5.00 μL of 2X PCR master mix (Fermentas),

73
1.00 μL of cDNA template, 10 pmol of reverse and forward primers and water (DEPC
treated) to make final 10 μL volume. Thermal cycling parameters were expressed in
the fig no. 3.1. The reaction conditions of thermocycler for expression study of both
GAPDH and HCV NS3 gene were same. Amplified products of PCR were analyzed
by Agarose gel electrophoresis using agarose gel (2.00 %). The image of DNA
products were captured in gel documentation apparatus under the luminescence of
short wavelength ultra violet light.
Fig. 2.1: Cycling parameters for expression analysis of HCV NS3 gene in reverse
transcription PCR
2.7.4 Expression studies via Real-time PCR
Reverse transcription tagged along by real-time PCR (RT-qPCR) is considered as the
gold standard for the quantification of gene expression because of its remarkable
sensitivity and sequence specificity characteristics. Relative Quantification was also
conceded out by means of quantitative Real-time PCR on ABI 7500 Real time PCR
machine (Applied Biosystems Inc. USA) using SYBR Green Chemistry (Fermentas,
USA). Reported protocol of expression analysis was followed with little
modifications (Jahan et al., 2011). Briefly in 7.5 µL of SYBR Green/ROX qPCR
Master Mix (2X) (Fermantas), relevant forward and reverse primers (1.00 µL each)
and RNase free water (5.00 µL) were mixed and reserved in ice. After mixing all
regeants cDNA template (0.5 µL) was added in the end and final reaction mixture was
added in the reaction wells of PCR plate. Final data was acquired in extension step.
The assay was done in triplicate and comparative CT method (ΔΔ Ct value) was
utilized for targeted gene relative quantification against GAPDH (internal control).

74
Fig. 2.2: Thermal cycling conditions for Real-time PCR
2.8 Western Blotting of PO extracts
A specific protein in any given sample could be detected by using specific antibody
for that protein & this analytical technique is known as western blotting. It involves
the separation of denatured proteins by using SDS gel electrophoresis, transfer of
those proteins on a nitrocellulose membrane & finally treating with specific
monoclonal antibodies.
2.8.1 Protein Isolation
Cell culture was prepared in 6 well plate/ 60 mm culture dish to conduct transfection
assay. A transfection test was executed by either including or excluding the plant
extracts after 24 h of cell culturing. Then, after 48 h, cells were rinsed with 1XPBS
(phosphate buffer saline). A cocktail of cell lytic buffer comprising of 10 µL (PMSF)
along with 1.00 µL protease inhibitor (Sigma Aldrich, USA) mixed with 1.00 mL of
cell lyses buffer was poured in each well. This cell lystae was kept at -20 0C for 30
min after transferring to the eppendorf, then centrifuged for 25 min at the rate of
13,000 rpm for 4 0C. The supernatant containing protein was collected & stored in
new eppendorfs at -20 0C.

75
2.8.2 Protein Quantification
Quantification of the isolated protein was done by spectrophotometer using Bradford
reagent (Fermentas, ready-to-use, #R1271). This was done by mixing of 10 µL
protein with 990 µL Bradford reagent & subjected to quantification at 595 nm.
2.8.3 Protein Samples Preparation
For protein sample preparation, the pre-calculated sample volume (containing 60 µg
of total protein as per yield of protein extraction quantified by Bradford assay) was
mixed with 5X protein loading buffer (Genei) plus 20X of reducing dye (Fermantas).
After 3-4 min heating of reaction tubes in hot water, those tubes were kept on ice for
subsequent 2-3 mins. Then, after a short spin, samples along with 10 µL of the protein
marker (Fermentas) were loaded into protein gel.
2.8.4 SDS Gel Electrophoresis
After heat shock of samples, 60 µg protein along with the sample buffer (5X) &
reducing agent (20X to remove unwanted structures) were loaded onto SDS-PAGE
for analysis of HCV NS3 protein in presence or absence of plant extracts. The
acrylamide gel apparatus, used for electrophoresis, was positioned vertical in holder
assembly. Except 10 % APS & TEMED (used as polymerization catalyst), all other
reagents were thoroughly mixed. Resolving gel was poured onto the 2/3 portion of
apparatus followed by 70 % ethanol to avoid any bubble formation.
Table 2.3: Composition of the gels used in SDS-PAGE
Solutions Resolving gel 10 % Stacking gel 5.00 %
Acrylamide/bis-acrylamide (30 %) 3.00 mL 0.65 mL
Tris buffer 0.1 M (pH 8.8) 0.5 M (pH 6.8)
10% SDS 0.15 mL 0.08mL
10% APS 0.15mL 0.08 mL
dH2O 2.5 mL 2.75 mL
TEMED 0.006 mL 0.008 mL
After the gel polymerization, ethanol was removed followed by washing with water.
Water was wiped with a filter or tissue paper before inserting combs. After loading
stacking gel, 1XSDS-PAGE tank buffer was poured into Western blot tank (Bio-Rad,

76
Germany). Initially, gel was run at 60 V till samples passed from the stacking gel,
then voltage was switched up to 80 V to allow complete separation of protein
samples.
2.8.5 Transfer of Protein and Immunoblotting
To detect by antibody, proteins were made available at nitrocellulose membrane
(Hybond C extra, Amersham). To acquire this goal, six sheets of blotting paper & one
sheet of nitrocellulose was cut as per the gel dimension. After soaking these papers in
transfer buffer, the gel was carefully placed over nitrocellulose sheet & was covered
by the blotting papers. Then the whole set was placed in electro blot and run at 16 V
for 1.5 h. By the use of 5 % skimmed milk (prepared in TBST at 4 0C overnight), the
membrane was blocked. 1X TBST was used to wash the membrane & then membrane
was incubated for 1h at room temperature with the monoclonal antibody(Primary)
specific to HCV NS3 gene along with GAPDH (Santa cruz, Biotechnology). Again,
1X TBST was used for three times washing of membrane and each washing continued
for 5.0 minutes. The membrane was incubated with anti-mouse antibody (Secondary)
for 45 minutes. Finally, the membrane was washed for 3 more times by using 1X
TBST.
2.8.6 Enhanced Chemiluminescence (ECL) Detection
After removal of excess buffer, the membrane was kept over cling film/flat surface.
Both ECL solutions (chemiluminescence peroxidase substrate; Sigma, Cat # CPS-
3500) were mixed as 1:2 ratios & poured over the membrane for about 4-5 minutes.
The membrane was wrapped with cling film after draining of excessive substrate. The
membrane was exposed to X-ray film (Kodak) for 5.00 minutes on a cassette. Kodak
developer & fixer solutions were used for the film development. Initially, the X-ray
film was dipped in developer solution for about 30-40 seconds, then washing with
distilled water, followed by dipping & hand shaking of the film in fixer solution for
about 30 Sec.

77
The exposed & developed X-ray film showed the protein bands. A comparative study
of protein bands between sample (with plant extracts) and control (without plant
extracts) indicated the inhibition. The protein expression levels were checked using
chemiluminescence’s detection kit (Sigma Aldrich, USA). The protein was quantified
from X-ray images using freely available software Gel-quant-express (http://gel-
quant-express.software.informer.com/4.1/).
2.9 Analysis of the Two Plants Extract That Exhibit Positive
Potential against HCV
All fifteen plant methanolic extract were screened for their activity against the
expression vector (pCR3.1/FLAGtag/HCV NS2-NS3/4A) of genotype 3a under the
uniform conditions explained above. Only two plants (plant codes CTS and POL) out
of exhibiting positive potential during antiviral screening against HCV. So these
plants were further selected for bioassay directed extraction in different solvents and
then qualitative and quantitative analysis of the phytoconstituents of all extract and
their screening against HCV RNA and its protein NS-3 serine protease. In addition to
this antioxidant, antithrombic and antibacterial potential of these promising extracts
methanolic extracts was also carried out. Finally the active fractions were also subject
to the column chromatography for extraction and purification of some compounds
present in them. These compounds were then identified with the help of different
techniques like FTIR, Mass spectrometry and NMR etc.
2.10 Phytochemical Screening
Preliminary qualitative and quantitative phytochemical screening was carried out to
determine the presence of various biochemically active compounds in methanolic
extract.
2.10.1 Qualitative Analysis of the Extracts
Various known chemical tests were used to detect the presence of bioactive
constituents like alkaloids, fatty acids, steroids, glycosides, terpenids, saponins,
flavonoids, resins, phenols, tannin, reducing sugars, carbohydrates, and proteins etc.
in each extract of both plants by using standard procedures.

78
2.10.1.1 Glycosides
Presence of glycoside in the extract was detected by Keller-Killani test. For this
purpose extracts solutions (5.00 mL each) were treated with glacial acetic acid (2.00
mL) having one drop of ferric chloride solution. This mixture was underlaid with of
concentrated sulphuric acid (1.00 mL). A violet ring may appear below the brown
ring, while in the acetic acid layer, a greenish ring may form just gradually throughout
thin layer that signify the presence of glycosides.
2.10.1.2 Steroid
Methanolic extract (0.5 g) of each sample was added into the solution of acetic
anhydride (2.00 mL) and sulphuric acid (2.00 mL). The colour changed from violet to
blue or green in some samples indicating the presence of steroids.
2.10.1.3 Alkaloids
Alkaloids in various fractions were detected by adding the extract (0.4 gm) into of
1.00 % HCl (8.00 mL), then warmed and filtered. Each filtrate (2.00 mL) was titrated
separately by means of (a) potassium mercuric iodide (Mayer’s reagent) and (b)
potassium bismuth (Dragendroff’s reagent). Turbidity indicated the presence of
alkaloids (Harborne et al., 1984).
2.10.1.4 Terpenoids
Extract solution (1.00 mL) was added in of chloroform (10 mL). Concentrated H2S04
(3.00 mL) was carefully added along the sides of the test tube in order to form a layer.
A reddish brown colouration of the interface indicated the presence of terpenoids
(Gibbs et al., 1974).
2.10.1.5 Fatty Acids
Extract solution (0.5 mL) was mixed with ether (5.00 mL). These extract allowed to
evaporate on filter paper and then dried the filter paper. The appearance of
transparence on filter paper indicates the presence of fatty acids (Ayoola et al., 2008).

79
2.10.1.6 Saponins
Detection of saponin was conceded by dissolving the extract solution (5.00 mL) into
distilled water (20 mL) and then agitated in a graduated cylinder for 15 minutes till
the formation of steady persistent foam. Few drops of olive oil were added into the
frothing. Formation of emulsion points toward the existence of saponins (Kumar et
al., 2012).
2.10.1.7 Flavonoids
Three methods were used to test for flavonoids. First, dilute ammonia (5.00 mL) was
added to a portion of an aqueous filtrate of the extract. Concentrated sulphuric acid
(1.00 mL) was added. A yellow colouration that disappears on standing indicates the
presence of flavonoids. Second, a few drops of 1.00 % aluminium solution were
added to a portion of the filtrate. A yellow colouration indicates the presence of
flavonoids. Third, a portion of the extract was heated with ethyl acetate (10 mL) over
a steam bath for 3 min. The mixture was filtered and the filtrate (4.00 mL) was shaken
with dilute ammonia solution (1.00 mL). Appearnce of yellow colouration indicates
the presence of flavonoids (Ayoola et al., 2008; Harborne et al., 1984).
2.10.1.8 Tannins
Sample extract (0.5 mL) was taken along with one ml of water and 1-2 drops of ferric
chloride solution. The result was in the form of blue colour for gallic tannins and for
catecholic green black for catecholic tannins (Iyengar et al., 1995).
2.10.1.9 Coumarins
In extract solution (2.00 mL) of selectected plants 10 % NaOH (3.00 mL) was added
and observed the formation of yellow colour that indicated the presence of coumarins
(Rizk et al., 1982).
2.10.1.10 Anthraquinone (Bornatrager’s test)
In a dry test tube extract (0.5 mg) was taken and chloroform (5.00 mL) was added.
After shaking of 5 min the extract was filtered and equal volume of 10 % ammonium

80
solution was added. Appearance of pink, violet or red colour confirmed the presence
of anthraquinone.
2.10.1.11 Anthocyanins
A mixture of 2 N HCl and ammonia (2.00 mL) was added in the extract solution (2.00
mL) of the plant. The emergence of pinkish red colour and then its conversion into
blue-violet colour indicated the occurrence of anthocyanins (Paris et al., 1969).
2.10.1.12 Emodins
Solutions of NH4OH (2.00 mL) and Benzene (3.00 mL) were added into the extract
solution and observed the emergence of red colour that indicates the presence of
emodins (Rizk et al., 1982).
2.10.1.13 Resin Identification
For the confirm presence of resin in selected plant, distilled water (5.00 mL) was
added into the extract solution and observed the appearance of turbidity.
2.10.2 Quantitative Determination of the Chemical Constituency Specifically
Antioxidants
2.9.2.1 Determination of alkaloids
For alkaloids determination, 5.00 gm of each sample was weighed into a 250 mL
beaker, and 200 mL of 20 % acetic acid in ethanol was added and was covered to
stand for 4 hrs. This was filtered and the extract was concentrated using a water bath
to evaporate one-quarter of the original volume. The concentrated ammonium
hydroxide solution was added drop-wise to the extract until the precipitation was
completed. The entire solution was allowed to settle and the precipitate was collected
by filtration, after which it was weighed (Harborne et al., 1984).
2.10.2.2 Determination of Pectin
To determine pectin, 5.00 gm of the plant sample was dispersed in 40 mL water in a
beaker and boiled gently for one hour. The content of the beaker was transferred to a

81
50 mL volumetric flask and diluted up to the mark with distilled water. The contents
of the flask were thoroughly mixed and filtered into a 50 mL volumetric flask, after
which 10 mL of the filtrate was taken into 80 mL beaker, where 30 mL de-ionized
water and 1.00 mL sodium hydroxide were added to it along with a constant string. It
was then allowed to stand overnight. To this solution, 5.00 mL of acetic acid (1.00 N)
was added with continuous stirring and was allowed to stand for 5 mins. Then 2.5 mL
of the calcium chloride solution were allowed to stand for one hour and the solution
was boiled for one minute. The solution was filtered through Whatman No. 41 filter
paper and washed with hot de-ionized water until the solution become free from
chloride. The residue was transferred to the previously weighed watch glass. The
watch glass, along with the residue, was placed on a water bath and dried in the oven
at 100 °C until a constant weight was obtained (Horwitz et al., 2000).
2.10.2.3 Determination of saponins
For the saponins determination, 5.00 gm of each plant samples was weighed and was
dispersed in 100 mL of 20 % ethanol. The suspension was heated over a hot water
bath for 4.00 h with continuous stirring at about 55 °C. The filtrate and the residue
were re-extracted with another 100 mL of 20 % ethanol. The combined extracts were
reduced to 40 mL over water bath at about 90 °C. The concentrate was transferred
into a 250 mL separating funnel and 20 mL of diethyl ether was added and shaken
vigorously. The aqueous layer was recovered while the ether layer was discarded. The
purification process was repeated and about 30 mL of n-butanol was added. The
combined n-butanol extracts were washed twice with 10 mL of 5.00 % aqueous
sodium chloride. The remaining solution was heated in a water bath. After
evaporation, the samples were dried in the oven to a constant weight. The saponin
content was calculated in percentage (Obadomi et al., 2001).
2.10.2.4 Total phenolic contents
Evaluation of total phenolic contents was carried out by modifying the reported
method (Kalpna et al., 2011). Shortly, freshly prepared 0.5 N Folin-Ciocalteu reagent
(0.1 mL) was added into 0.5 mL of diluted extract sample (10 mg/5mL distilled
water) and incubated for 15 minutes. Then the solution was again incubated in dark
for half an hour after the addition of 2.5 mL of 7 % Na2CO3 solution. Finally noticed

82
the absorbance of at 700 nm and expressed the results as Gallic Acid Equivalent mg/g
dry weight of the plant extract.
2.10.2.5 Total flavonoid contents
Total flavonoid contents were determined the reported method (Zhishen et al., 1999).
In brief, 0.3 mL of fresh prepared NaNO2 solution (1:20) was added into the 1.00 mL
of plant extract aqueous solution (10 mg/mL). Then, after 5 minutes of room
temperature incubation, 0.3 mL of freshly prepared AlCl3 (1:10) was poured into that
solution. Then wait for six minutes for the accomplishment of reaction and then added
2.00 mL of 1.00 M NaOH. Finally, make the total volume upto 10 mL by the
addition of distilled water and immediately measured the absorbance at 510 nm. Final
results were articulated as Catechin Equivalent mg/g of the plant dry weight.
2.10.2.6 Total carotenoid contents
Total carotenoids were determined following a method reported by (Lichtenthaler et
al., 2001). In brief, plant powder samples (5.0 gm) were immersed in 80 % acetone
separately and incubated at 4 0C in dark for three days. After incubation samples were
filtered and centrifuged at 2500 rpm for 15 minutes and separate supernatant. Finally
measure the supernatant absorbance at three different wavelength i.e. 663.2 nm, 646.8
nm and 470 nm. Concentrations of total carotenoid and Chlorophyll a, Chlorophyll b
were calculated via following equations and results were articulated in µg/g of leaf.
Ca= 12.25A663.2- 2.79A646.8
Cb = 21.50A646.8 – 5.10A663.2
Cx+c =𝟏𝟎𝟎𝟎𝐀𝟒𝟕𝟎 – 𝟏.𝟖𝟐𝐂𝐚 – 𝟖𝟓.𝟎𝟐𝐂𝐛
𝟏𝟗𝟖
2.10.2.7 Total tannin contents
Total tannin contents were determined according to the reported protocol (Hosu et al.,
2014) with a few modifications. Plant extract (10 mg) was diluted with 4.00 mL
distilled water, which was divided into two equal fractions. In both factions
concentrated HCl (3.00 mL) and distilled water (1.00 mL) were added. In one fraction
ethanol (0.5 mL) was added while the second one in which ethanol was not added was
incubated at 100 0C for 30 min. Then measured the absorbance difference of these

83
two fractions at three different wavelength 470, 520 and 570 nm and calculated the
total tannin contents with the help of following formula.
∆A520 = 1.1 x ∆A470
∆A520 = 1.57 x ∆A570
TTC (g/l) = 15.7 x minimum (∆A520)
2.10.2.8 DPPH radical scavenging activity
Reported DPPH assay along with certain modification was used to determine the
DPPH radical scavenging capacity of plant extracts (Proestos et al., 2013). DPPH
radical exhibits utmost absorbance at 517 nm while drop in absorbance values is
calculated as radical scavenging potential of phyto extract. For this assay, 0.2 mM
DPPH (2.00 mL) was added into 2.00 ml of methanolic solution of plant extracts
(5:1). Similarly in case of control, DPPH solution (2.00 mL) was added into methanol
(2.00 mL) and incubated the reaction mixtures for one hour in dark. Then measure the
absorbance at 517 nm against blank (methanol). All measurements were made in
triplicate and DPPH radical percent inhibition was calculated with the aid of
subsequent formula:
% Inhibition = 𝑨𝒄𝒐𝒏𝒕𝒓𝒐𝒍−𝑨𝒔𝒂𝒎𝒑𝒍𝒆
𝑨𝒄𝒐𝒏𝒕𝒓𝒐𝒍×𝟏𝟎𝟎
2.10.2.9 ABTS radical cation scavenging activity
Reported ABTS assay along with slight modification was used to determine the
ABTS radical cation scavenging capacity of plant extracts (Proestos, et al., 2013). In
brief, 7.00 mM solution of ABTS and 2.45 mM solution of MnO2 were prepared
separately and mixed with ratio of 1:1 and incubated in dark for dark for 48 hours.
Then diluted the solution of ABTS with aqueous methanol (1:25) and finally of this
solution (2.00 mL) was added in 200 μL of plant extract prepared in aqueous
methanolic (1:10). Similarly control was prepared and absorbance was measured at
734nm. Percent scavenging of ABTS radical cation was calculated using the relation:
% Inhibition = 𝑨𝒄𝒐𝒏𝒕𝒓𝒐𝒍−𝑨𝒔𝒂𝒎𝒑𝒍𝒆
𝑨𝒄𝒐𝒏𝒕𝒓𝒐𝒍×𝟏𝟎𝟎
All experiments and recordings were performed thrice and results were averaged.

84
2.10.2.10 Ferric reducing antioxidant power
Plant extracts reducing power was determined according to the method reported by
Zhuang (Zhuang et al., 2012). Briefly, plant extract (10.00 mg) was added into 3.5
mL phosphate buffer (0.2 M, pH 6.6), followed by subsequent addition of 1.00 %
potassium ferricyanide (2 mL) and incubation at room temperature for 20 minutes.
Afterward, 10 % trichloroacetic acid (2.5 mL) was added into the mixture and the
mixture was centrifuged at 3000 rpm for 10 minutes. Then distilled water (2.5 mL)
and 0.1 % ferric chloride (0.5 mL) was added into the 2.5 mL solution of supernatant
and finally noted the absorbance of mixtures at 700 nm. Results were expressed as
Ascorbic Acid Equivalent mg/g dry weight.
2.10.2.11 Identification and Quantification of Phenolic Acids by HPLC
High performance liquid chromatographic method was utilizes for identification and
quantification of phenolic acids in selected plant extracts. In brief, sample was
prepared by dissolving the plant extract (50 mg) into of distilled water (15 mL)
followed by subsequent addition of 24 mL methanol (Merck, HPLC grade) and
shaken the mixture for 5 minutes in shaker. Then 10 mL of 6.00 M HCl (Merck,
Analytical grade) was added and mixture was incubated in oven at 950 C for two
hours. Samples were filtered through 0.2 μ syringe filter paper and 5.00 mL of each
extract was injected on a Shim-Pack CLC-ODS (C-18), (25 cm x 4.6 mm, 5 μm) for
gradient elution. Working solvent were A(H2O:Acetic Acid 94:6, pH=2.27),
B(Acetonitirle 100 %). The gradient profile was 0-15 min= 45 % B, 30-45=100 %B
with flow rate of 1ml/min. UV/Visible detector was set at 280nm for quantitative
determination. The temperature of the column was 25 ◦C. Concentrations of phenolic
acids in each sample were determined with the help of calibration curve.
2.11 Anti-clotting Properties of CTS and POL
Extracts
In vitro thrombolytic potential of methanolic and ethylacetate fractions of CTS and
POL were determined by following the already reported method (Prasad et al., 2007))

85
2.11.1 Sample Preparation
Positive control: Stock of positive control was prepared from commercially available
Streptokinase (SK) vial (15, 00,000 I.U) (Polamin Werk GmbH, Herdecke, Germany)
that was dissolved in 5.00 mL of sterile water and from which 100 μL (30,000 I.U)
was used for in vitro thrombolysis.
Specimens: Whole blood (5.00 mL) was drawn from healthy human volunteers (n =
10) without a history of oral contraceptive or anticoagulant therapy. Blood sample
(500 μL) was transmitted to pre weighed alpine tube for the formation of clot.
2.11.2 Preparation of Anti-Clot Test Solution
Selected plant extract (100 mg) was suspended in 10 mL of distilled water, mixed
vigorously on vortex mixer and kept overnight at room temperature. Then mixture
was filtered through 0.22 micron syringe filter and used to check in vitro thrombolytic
activity.
2.11.3 Clot Lysis
After the formation of clot, serum was completely aspirated without disturbing the
colt and each tube having clot is again weighed for determination of clot weight.
Weight of clot = weight of clot containing tube –weight of empty tube
Each tube having clot was labeled properly and to each tube containing pre weighed
clot, 100 µL of aqueous plant extract added separately. As a positive control, 100 µL
of Streptokinase and as a negative control, 100 µL of distilled water were separately
added to the control tubes. All the tubes were then incubated at 37 °C for 90 minutes
and observed for clot lysis. After incubation, fluid obtained was removed and tubes
were again weighed to examine the difference in weight before and after clot
disruption. Difference obtained in weight taken before and after clot lysis was
expressed as percentage of clot lysis. The experiment was repeated numerous times
with the blood samples of different volunteers (Prasad et al., 2006).
% clot lysis = (Weight of the lysis clot / Weight of clot before lysis) × 100
2.12 Antibacterial assay of extracts

86
The antibacterial action of selected phyto extract was performed on active cultures of
different bacterial strain by following the Kirby Bauer disc diffusion method (Bauer et
al., 1966). For this purpose six bacterial strains (three gram positive and three gram
negative strains) were procured from the microbiology laboratory of department of
pharmacy, University of Sargodha and were as follows: Bacillus subtilis 6633,
Staphylococcus aureus ATCC 25923, Styphylococcus epidermidis ATCC 12228,
Pseudomonas aerugenosa ATCC 27853, Escherichia coli ATCC3411 and Salmonella
typhi ATCC 26823. The bacterial stock cultures were preserved on nutrient agar
plates. A loop full of bacterial cells from the nutrient agar plates was inoculated into
10 mL nutrient broth and incubated at 37 oC for 24 h. After incubation, the culture
suspension was compared with the McFarland standard. In the presence of visible
light visually compared the turbidity of bacterial suspension to the 0.5 McFarland
standards and adjusted the turbidity of bacterial suspension. Nutrient agar media of
(Oxoid, UK) was prepared and sterilized by autoclaving at 121 oC for 15 minutes at
15 psi pressure and petri plates were prepared. Then Wicks paper disk (6.00 mm
diameter) that was carrying sample solution was placed smoothly on the respective
growth media. Afterward 100 µL of innoculum was added, mixed thoroughly and
poured into the petri plates using a sterile glass spreader. The plates were incubated
for 24 h at 37 °C and the diameter of the inhibition zones was recorded in millimeters
by using zone reader. Each antimicrobial assay was performed in triplicate & mean
values were taken (NCCLS. 2003).
2.13 Bioassay guided extraction of CTS and POL
For bioassay-directed extraction, crude methanolic extracts of each plant was
dissolved in 70 % n-hexane and then separate the aqueous layer from non polar
hexane layer with the help of separating funnel. In aqueous faction add the ethyl
acetate repeated the similar procedure for separation of ethyl acetate soluble
compounds. Then chloroform and n-butanol was added respectively in order to obtain
the chloroform and butanol extract. Final remaining aqueous extract was not used in
any of the further analysis. The bioassay guided extraction scheme used for the
separation of polar, slightly polar and non polar compounds is shown in fig. 3. The
polar and non polar organic layers were concentrated in vacuum to give four fractions
(H, C, E and B) which were screened for HCV NS3-SP inhibition activities.

87
Fig. 2.3: Scheme for bioassay-guided fractionation of Caralluma tuberculata and
Portulaca oleracea L methanolic extracts.
2.14 Isolation of Compounds via Column Chromatography
2.14.1 Isolation of Compounds
Flash chromatography was executed to isolate the compounds from Ethyl acetate
fraction on a glass column (60 x 3.5cm) manually packed with silica gel 60. After the
column packing it was equilibrated with CHCl3 that remove all the trapped air
bubbles from silica gel.10 gm of EtOAc extract was adsorbed on silica and this slurry
was loaded on the top of the packed column. Sample elution was carried out stepwise
using a gradient CHCl3-MeOH solvent system. 110 fractions of 250 mL were
collected successively with increasing concentrations of methanol in chloroform and
concentrated. Presence of compounds in fractions was monitored by thin layer
chromatography (TLC) by using commercially available TLC plate coated with
polyamide film (China). Spots were observed directly under ultraviolet light (254 nm)
and then beneath visible light after using different reagents. Fractions possessing
component with same Rf values were combined together. Fractions that have show
two spots on TLC again chromatographed on silica column using Ethyle actate:
methanol solvent system.

88
2.14.2 Physico-chemical Characteristics of Isolated Compounds
After examining the physical appearance and state of extracted compounds and their
melting points were determined on Stuart Scientific SMP10 apparatus. Optical
rotation was calculated via JASCO-DIP360 digital polarimeter.
To verify the steroidal character of the compounds, the EtOAc fractions and the
isolated Compounds were examined by Liebermann-Burchard test (Burke et al., 1974;
Halim et al., 1996). Among the many colour reactions for steroids, the Liebermann-
Burchard procedure is perhaps the most widely used test. Liebermann- Burchard
colour reactions for steroids have oxidative mechanisms, yielding oxidation products.
This reaction was illustrated firstly by in 1885 by Liebermann and then applied to
cholesterol analysis soon after by Burchard. In the early studies Chloroform was used
as a solvent, but the Liebermann-Burchard (L-B) reaction, as carried out today, is
done in an acetic acid, acetic anhydride medium and sulphuric acid (Halim et al.,
1996). Ten drops of 1.00 % solution of the EtOAc fraction in chloroform were added
to a dry test tube. Then 5 drops acetic anhydride was added to the test tube and the
solution was mixed well. Then three drops of concentrated sulphuric acid was added
and the solution was mixed thoroughly. Immediate change in colour was recorded and
even after passage of 10 minutes change is noticed and recorded.
2.14.3 Elucidation of Compounds Structure by spectroscopy
Spectroscopy is the study of the absorption and emission of radiation by matter. In
physical and analytical chemistry, spectrometry is often used in the identification of
substances through the spectrum emitted from or absorbed by them. Fractions having
confirmed single compound were dried thoroughly in vacuum line for 4-6 hrs in order
to make sure that complete evaporation of solvent. Then the structures of these
purified compounds were characterized by FTIR, ESI-MS and NMR.
2.14.3.1. Electro Spray Ionisation (ESI)
Electro Spray Ionisation (ESI) is perhaps the most common method of choice for
analysis of molecular weight of broad range of chemical and biological analyte. It is
considered more sophisticated ionization that does not cause extensive fragmentation

89
of molecular ions. Here ions are generated in form of charge droplet from the liquid
and then subsequently evaporated. The analyte was prepared in methanol (80 %)
along with formic acid (0.1 %) in order to enhance protonation and increase
sensitivity. Mass spectra (MS) were recorded using a Micromass Platform II
spectrometer using an electrospray ionization source.
2.14.3.2 Infra-Red spectroscopy
Infra-red (IR) spectroscopy offers an approach of unearthing the functional moieties
in any molecule, as it identifies the bending and stretching vibrations of bonds
specifically the asymmetrical bonds stretching of the functional groups such as C=O,
NO2, NH2, and OH. Infrared spectra were obtained by using ALPHA-P FTIR
spectrometer
2.14.3.3 Nuclear Magnetic Resonance (NMR) spectroscopy
NMR spectroscopy work on the basis of nuclear magnetic resonance that provides full
insight not only about the structure of unknown compounds but also for reaction
dynamics and environment of molecule. Resonance frequency due to intramolecular
magnetic field created around atoms of molecule. Compound absorbs electromagnetic
rays under suitable conditions of applied magnetic field. Absorption is the basic
property of the atom’s nuclei. NMR spectrum is obtained from peak intensities plot
derived from absorption peaks and frequencies.
Extracted purified compounds were dissolved in CDCl3 or MeOH-D3 (CD3OD)
depending upon the nature of compounds and then their structure were interpreted
from the spectra obtained from one dimensional (1H, 13C, and 13C DEPT) and two
dimensional (COSY, HMBC and HSQC) NMR spectras determined on 300 MHz and
400 MHz NMR on a Bruker DPX 400 spectrometer, expressed in ppm (δ) relative to
trimethylsilane (TMS) as an internal standard.

90
3.0 Results and Discussion
In recent era, research on medicinal plants focuses not only the documentation of their
ethnopharmacological detail but also deciphering the characteristics of their
potentially active principals (Fennell et al., 2004). As we know that plants are
complex mixture of the active and inactive components and the separation of active
from inactive is always remain arduous and tedious task (Sticher, 2008). Five
different approaches usually opted for phyto research like random, ecological,
phylogenetic or chemotaxonomic, ethno-guided and bioassay directed (Albuquerque

91
et al., 2006; Elisabetsky et al., 1994). Different aspects like evidence based
information about ethno pharmacological use by natives; plant part along with disease
name and its availability in natural habitat etc were kept under consideration when
medicinal plants were chosen for scientific assessment of their biological potential
and rationalizing their ethno pharmacological usage (Matos, 2000). This information
should be complete enough to fill the gap between traditional knowledge of plant and
present modern scientific utilization.
3.1 Collection of Botanical Samples and Preparation
of Extract
Different samples from fifteen plants were collected from Soon-sakaser valley of
Khushab, Pakistan on the basis of conventional knowledge of local community and
herbal practioners. All indigenous botanical samples were used as primary source of
health care against viral and liver disease specifically for hepatitis viruses. Clean air
dried botanicals were pulverized and their methanolic extracts were obtained by
maceration method.
Keeping under consideration the limited resources and huge amount medicinal plant
sample a proficient system of extraction is needed for the separation and the
biological screening of compounds from the cellular matrix of plant (Sticher, 2008).
Preferred method (solid-liquid extraction) should be convenient, meticulous, swift,
efficient, pollution free and inexpensive etc. Drawbacks associated with such method
of extraction yield are laborious, prolonged extraction time, high consumption of
solvent and poor yield. A wide variety of the solvents can be used ranging from non
polar to slightly polar and highly polar solvents depending upon the nature of
compound we want to extract. As usually we are working with unknown compounds
and we should know that extractant nature effect the composition of crude extract.
During traditional practice usually water and ethanol are solvent of choice. But in
modern scientific research petroleum ether, hexane, ethyl acetate, chloroform,
acetone, methanol, propanol, butanol etc. some factors like temperature, light and
storage conditions also affect the process of extraction and composition of
constituents.

92
3.2 Percentage yield of plants extract
Method of extraction is an important step for any type of natural product studies
because it has direct effect on the extractant capacity, amount of yield and quality of
extract (Berlin et al., 2005). Non polar and slightly polar solvent like petroleum ether,
hexane, dichloromethane, ethyl acetate extract sterol, terpenoids and some phenolics
while highly polar solvent like methanol, acetone extract wide range of polar
compound for example phenolics. After the calculation of percentage yield of all
samples separately all extracts were stored in contamination free vials and stored in
refrigerator at 4 ᵒC. Percentage yield of methanolic extracts of plants is shown in Fig.
3.1.
Fig. 3.1: Percentage yield of methanolic extracts of plants collected from Soon-
sakaser valley of Pakistan
3.3. Toxicological Studies of Plant Extracts in CHO and Liver Cell
Lines
Due to the ever increasing interest of man natural thing utilization and emphases of
research on medicinal plant has focus the dire need of gathering information regarding
quality, therapeutic efficacy, safety and toxicity potential of botanical extracts before
the treatment of any disease. Therefore cytotoxic trials become the gist of any
medicinal plant research that focus on its biological activity either the crude extract of
plant or any of its bioactive constituent.
CTSBML
CILCBR
CDLMFHL
SABSIS
CTRAAL
SALRER
ZJSPOL
ATB
0
10
20
30
Methanolic extract of plants
% Y
ield

93
For this purpose stock solutions (40 mg/mL) of each extract were prepared in DMSO
(Cell culture grade) separately for in vitro toxicological and antiviral studies in CHO
and hepatoma cell line (Huh-7).
3.3.1 Trypan Blue Dye Exclusion Assay
Prior to antiviral screening against Hepatitis C virus, cytotoxic effects of all the
samples were determined on liver Huh-7 cells through Trypan blue exclusion method.
After 24 h incubation of Huh-7 with different concentrations (ranging from 1.5 µg/mL
to 200 µg/mL) of all extract separately, cell viability was evaluated using trypan blue
dye and cells were counted using haemocytometer. Concentration of CTS and POL
did not exert any effect on viability of CHO and Huh-7 cells (Fig. 3.2 A & B). Similar
results were observed for other 13 extracts most of them appeared as non toxic (data
not shown).
3.3.2 Cytotoxic Screening of Phyto Extracts via MTT Assay
Another In vitro assay performed to check the viability of cell under the effect of
plant extract was MTT assay. In this assay, a cellular enzyme (oxidoreductase) in the
presence of NADPH reduce the substrate tetrazolium dye MTT (3-(4,5-
dimethylthiazol-2-yl)-2,5-diphenyltetrazolium) bromide into purple colour insoluble
formazan crystals in the alive cells. Crystals of formazon was dissolved adding the
solution of DMSO. Absorption of solution in visible range of light directly reflected
the number of alive cells. MTT assay results confirmed that both cell lines remain
viable to the concentration (200 μg/mL) of plant extract. Toxic effects of plant (CTS
and POL) crude extracts are shown in Fig. 3.3 (A & B).

94
Fig. 3.2 (A & B): Cellular toxicity analysis CTS and POL extracts through
Trypan blue exclusion method
For toxicological analysis of CTS (A) and POL (B), Huh-7 liver cells were seeded at a
density of 3 × 105 in six well plate. After considering first well as control, varying
concentrations of the CTS/POL were added in wells. Subsequent to 24 h incubation,
viable cells were counted with haemocytometer via Trypan blue dye exclusion
method. For the treatments, mean (n=3) + SD (n=3) with P value < 0.05 were found
significant with control.
CHO cell lines, derived from Chinese hamster ovary, were chosen usually for toxicity
screening of different products, gene expression, genetics and nutrition studies
because of their ability of fast growth, long-lasting stable expression of gene and
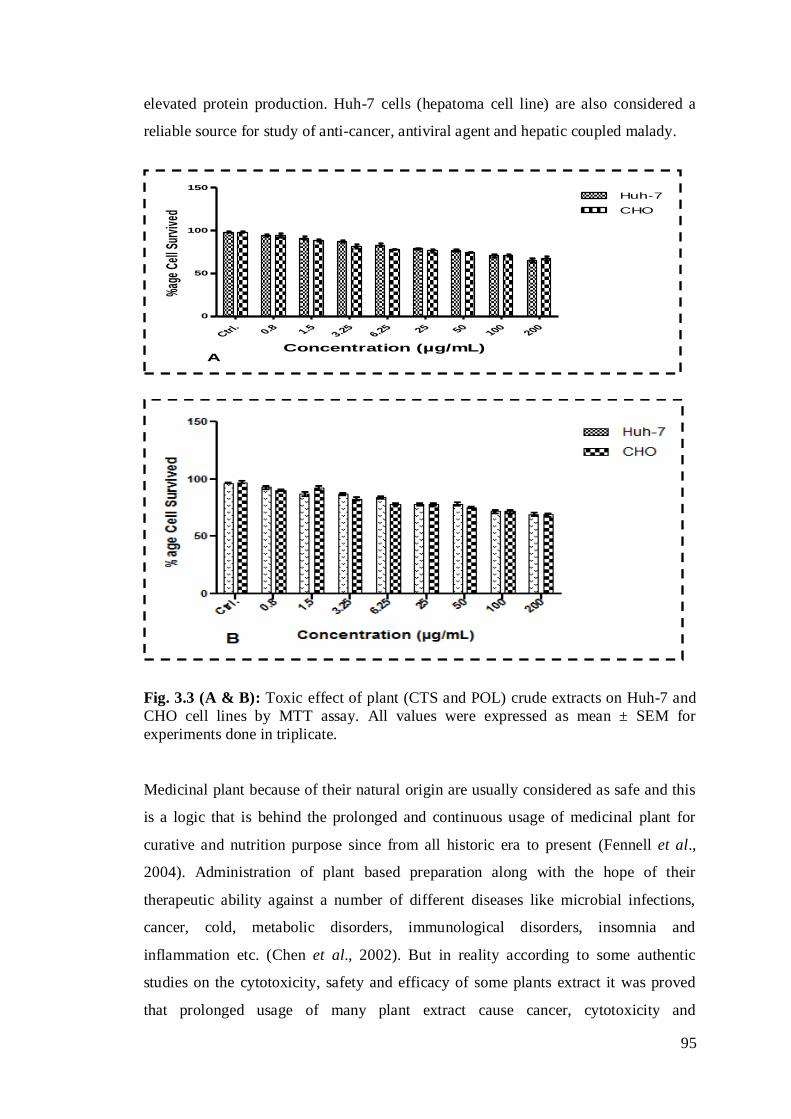
95
elevated protein production. Huh-7 cells (hepatoma cell line) are also considered a
reliable source for study of anti-cancer, antiviral agent and hepatic coupled malady.
Fig. 3.3 (A & B): Toxic effect of plant (CTS and POL) crude extracts on Huh-7 and
CHO cell lines by MTT assay. All values were expressed as mean ± SEM for
experiments done in triplicate.
Medicinal plant because of their natural origin are usually considered as safe and this
is a logic that is behind the prolonged and continuous usage of medicinal plant for
curative and nutrition purpose since from all historic era to present (Fennell et al.,
2004). Administration of plant based preparation along with the hope of their
therapeutic ability against a number of different diseases like microbial infections,
cancer, cold, metabolic disorders, immunological disorders, insomnia and
inflammation etc. (Chen et al., 2002). But in reality according to some authentic
studies on the cytotoxicity, safety and efficacy of some plants extract it was proved
that prolonged usage of many plant extract cause cancer, cytotoxicity and
Ctrl.
0.8 1.53.25
6.25 25 50100
200
0
50
100
150
Huh-7
CHO
AConcentration (µg/mL)
%ag
e Ce
ll Su
rviv
ed

96
genotoxicity (Ernst et al., 2011). Medicinal plant adverse reactions are obvious in
form of adulteration, heavy metal contamination, herb drug interaction and toxicity of
organs like liver, heart, gastrointestinal tract, central nervous system and kidney etc.
(Jordan et al., 2010). Most of the herbal products or formulation of so called herbal
industries do not fulfill the criteria of quality assurance, cytotoxic trials and good
quality manufacturing practice (Palombo, 2006). Furthermore, solvent choice used
for extraction can also cause problem of cytotoxicity. Sometime cytotoxic effect
becomes severe when botanicals are mixed with synthetic drug, microbial byproducts
like toxins etc without any standard clinical trial (Palombo, 2006). Especially when
botanical extract is administered along with narrow range spectra drug it potentially
hampers the response of drug and result cytotoxicity (Chen et al., 2011). In fact
during herbal-drug interaction the active constituent of herb exhibit their action
because these constituent having potential to undergo different enzymatic
transformations and produce toxic and non toxic metabolites (Chen et al., 2011).
Keeping in mind the significance of extract toxicity effect these studies were carried
out and all selected plant extracts appear non toxic and suitable for further biological
studies.
3.4 Anti-HCV Assay
Medicinal plants are utilized for premier health care need against various dreadful
infections throughout the globe. Therapeutic potential of medicinal plants lie behind
the separate or synergistic action of complex mixture of secondary metabolites
(Voravuthikunchai et al., 2006).
It has been reported that HCV can replicate in Huh-7 cells by the detection of viral
genes as well as viral copy number by Real Time PCR in both cells and supernatant,
whereas, HCV replication in cell culture is strictly limited to human hepatocytes and
their derivatives. In the present study, methanolic extracts from 15 different plants
were tested to determine the antiviral activity against HCV. Real time RT-PCR results
showed that only methanolic extracts of CTS and POL out of 15 medicinal plants
exhibited convincing potential (57 % and 70 % respectively) against HCV-RNA at
non-toxic concentration (Figure 3.4). This Methanolic extract further on the basis of
biassay-guided directions subdivided into four different solvent fractions. Both plant

97
methanolic extracts (POLM and CTSM) suspended in water further fractioned with n-
hexane (POLH and CTSH), ethyl acetate (POLE and CTSE), chloroform (POLC and
CTSC) and n-butanol (POLB and CTSB). Among these only ethyl acetate fraction
(CTSE & POLE) in both cases significantly reduced (68 % and 74 %) of the viral titer
(Figure 3.5 (A&B).
Fig. 3.4: Anti-HCV activity of methanolic extracts of selected fifteen different
plants
Huh-7 cells infected with HCV serum was cultured and incubated with 1.5 µg/well
concentration of different plants extracts for 24 h. After incubation total RNA was
extracted and the levels of HCV RNA were determined by Quantitative RT-PCR
assay and are shown as percentage to the levels of HCV RNA in cells in comparison
of control (without treatment of plant extract). Control (Ctrl.) Achyranthus aspera
(AAL), Bacop monnieri (BMB), Chichorium intybus (CIL), Chlorophytum
borivilianum (CBR), Corchorus depressus (Linn.) (CDL), Myristica fragrans Houtt
(MFHL), Portulaca oleracea L (POL), Sesamum indicum (SIS), Sonchus Arvensis
(SAL), Caralluma Tuberculta stem (CTS), Caralluma Tuberculta root (CTR),
Santalum album (SAB), Rheum emodi (RER), Zizyphus Jujuba (ZJB). All the
treatments were made in triplicate (n=3) and results are exemplified as mean ±
standard deviation. ***P value < 0.001 vs. control.
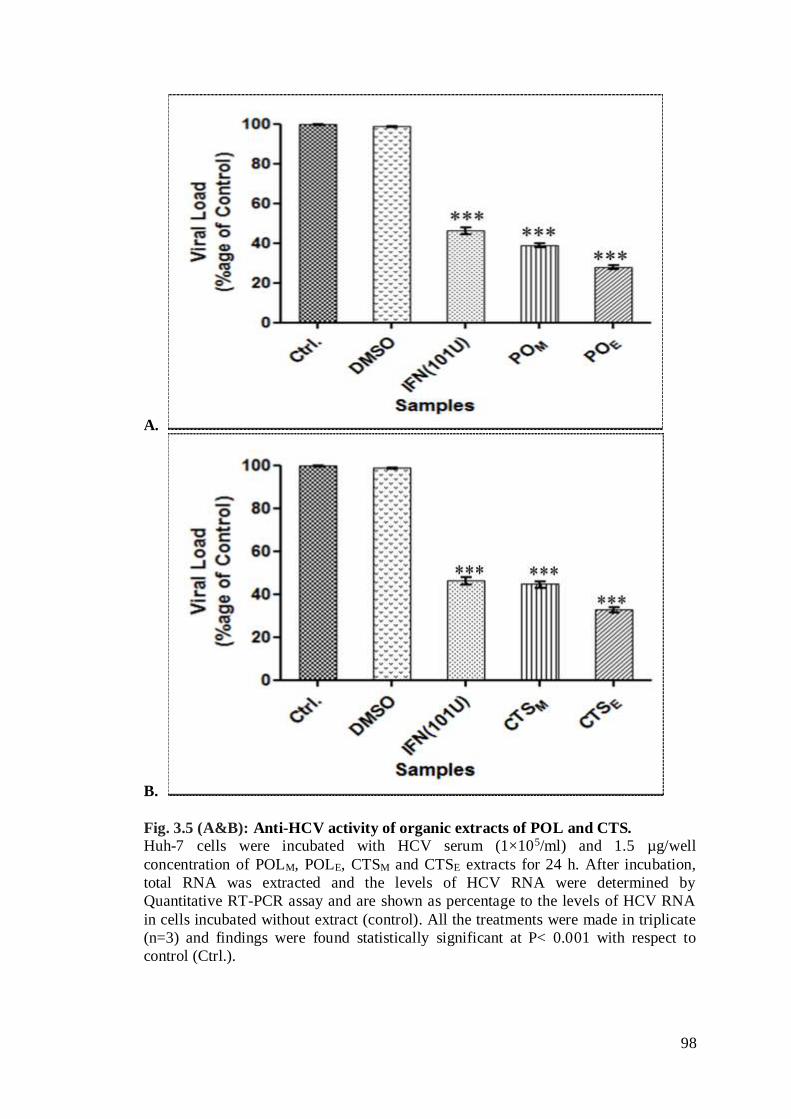
98
A.
B.
Fig. 3.5 (A&B): Anti-HCV activity of organic extracts of POL and CTS.
Huh-7 cells were incubated with HCV serum (1×105/ml) and 1.5 µg/well
concentration of POLM, POLE, CTSM and CTSE extracts for 24 h. After incubation,
total RNA was extracted and the levels of HCV RNA were determined by
Quantitative RT-PCR assay and are shown as percentage to the levels of HCV RNA
in cells incubated without extract (control). All the treatments were made in triplicate
(n=3) and findings were found statistically significant at P< 0.001 with respect to
control (Ctrl.).

99
3.5 Inhibition of HCV NS3 Gene Expression by Selected Organic
Extracts
In order to determine the effect of CTS and POL extracts against HCV NS3 protease,
Huh-7 cells were transfected with NS3 protease plasmid in the presence and absence
of herbal extracts. After 48 h incubation, cells were harvested, RNA was extracted
and cDNA were generated by oligo dT primers. cDNA was amplified by PCR using
primers specific to the HCV NS3 protease. Fig. 3.6 (A & B) and Fig. 3.7 (A & B)
represent the qualitative and quantitative inhibition of HCV NS3 gene by POL and
CTS extracts.
It is obvious from the figures that ethyl aceatate and methanolic extract respectively
inhibit the 69 % and 53 % expression of HCV NS3 protease. Similarly the POLM
reduced the NS3 gene expression to 75 %, whereas the ethyl acetate extract reduced
the level up to 85 %. The effect of POL on NS3 gene was also compared to INF (101
U) and it is shown that POL is more active in the reduction of NS3 gene expression
Fig. 3.6 (A & B). GAPDH mRNA expression was used as internal control, which
remained unaffected after the addition of extract. Present response is comparable to
the in vitro inhibition of HCV NS3-SP with Rhodiola kirilowii ethanolic extract of
and Camellia sinensis Epigallocatechin-3-gallate compound.
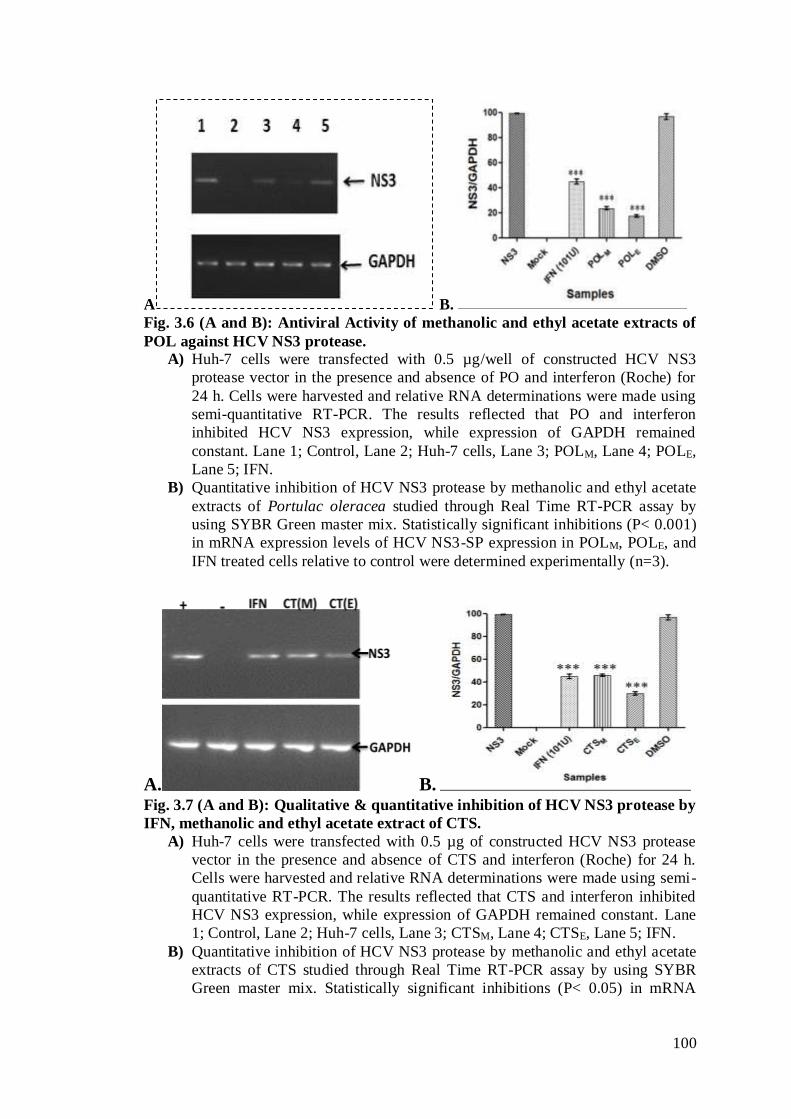
100
A B. Fig. 3.6 (A and B): Antiviral Activity of methanolic and ethyl acetate extracts of
POL against HCV NS3 protease. A) Huh-7 cells were transfected with 0.5 µg/well of constructed HCV NS3
protease vector in the presence and absence of PO and interferon (Roche) for
24 h. Cells were harvested and relative RNA determinations were made using
semi-quantitative RT-PCR. The results reflected that PO and interferon
inhibited HCV NS3 expression, while expression of GAPDH remained
constant. Lane 1; Control, Lane 2; Huh-7 cells, Lane 3; POLM, Lane 4; POLE,
Lane 5; IFN.
B) Quantitative inhibition of HCV NS3 protease by methanolic and ethyl acetate
extracts of Portulac oleracea studied through Real Time RT-PCR assay by
using SYBR Green master mix. Statistically significant inhibitions (P< 0.001)
in mRNA expression levels of HCV NS3-SP expression in POLM, POLE, and
IFN treated cells relative to control were determined experimentally (n=3).
A. B. Fig. 3.7 (A and B): Qualitative & quantitative inhibition of HCV NS3 protease by
IFN, methanolic and ethyl acetate extract of CTS.
A) Huh-7 cells were transfected with 0.5 µg of constructed HCV NS3 protease
vector in the presence and absence of CTS and interferon (Roche) for 24 h.
Cells were harvested and relative RNA determinations were made using semi-
quantitative RT-PCR. The results reflected that CTS and interferon inhibited
HCV NS3 expression, while expression of GAPDH remained constant. Lane
1; Control, Lane 2; Huh-7 cells, Lane 3; CTSM, Lane 4; CTSE, Lane 5; IFN.
B) Quantitative inhibition of HCV NS3 protease by methanolic and ethyl acetate
extracts of CTS studied through Real Time RT-PCR assay by using SYBR
Green master mix. Statistically significant inhibitions (P< 0.05) in mRNA

101
expression levels of HCV NS3-SP expression in CTSM, CTSE, and IFN
treated cells relative to control were determined experimentally (n=3).
3.6 Western Blotting
The effect of inhibition of Portulaca oleracea extract against HCV NS3 protease,
Huh-7 cells were transfected with NS3 protease plasmid in the presence and absence
of herbal extracts was also checked at protein level. Ethyl acetate and methanol
extracts of POL having antiviral potential for the inhibition of HCV NS3 protein were
also studied. Huh-7 cells were transiently transfected with pCR3.1/FLAGtag/HCV
NS3 of genotype 3a. Subsequent to 48h of incubation, the transfected cells were lysed
& western blotting technique along with monoclonal antibodies of HCV NS3 and
GAPDH was used to study the extracted proteins. For setting internal control,
GAPDH was used (Fig. 3.8). There was a dramatic reduction in the protein expression
of the NS3 gene after treatment of Portulaca oleracea extracts (Fig. 3.8). There were
60 % and 70 % inhibition in Huh-7 cells treated POM and POE respectively.
Fig. 3.8: Antiviral Activity of methanolic and ethyl acetate extracts of POL
against HCV NS3 protease at protein level Huh-7 cells were transiently transfected with NS3 gene of HCV and subsequently treated with POL
methanolic (POLM) and ethyl acetate (POLE) extracts DMSO and interferon. Total protein was isolated
24h post transfection and subjected to SDS-PAGE, and western blot. Results shows that POLM and
POLE significantly reduced the expression of NS3 gene as compared to Control, DMSO and interferon
treated cells. Lane 1; Control, Lane 2; DMSO, Lane 3 IFN; Lane 4; POLM, Lane 5; POLE, Lane 6;
Mock. Blots were also normalized by measuring the expression of GAPDH used as internal control.
The quantitative expression of protein on blots was measured using freely available software Gel-

102
quant-express. All experiments were performed in triplicate. Error bars indicate mean S.D,
***P≤0.001.
3.7 Phytochemical analysis
Plants exhibit their therapeutic potential by the charisma of secondary metabolites.
Secondary metabolite concentration of plant is greatly affected by climate conditions,
geographic and genetic variations, soil composition, pollution, season of plant
collection, treatment of plant before extraction, etc. From socioeconomic point of
view phytotherapy is also preferable over costly synthetic drug (Nunes et al., 2003).
Biochemical investigation of the extracts attained with various solvents is the utmost
significant relating to a variety of diseases. Results acquired in this perspective are
given as under.
3.7.1 Qualitative Analysis of CTS and POL
Standard procedures were used for the phytochemical screening of plants.
Phytochemical screening of the extract shows that many natural products were present
in these extracts. Qualitative screening of methanolic extracts of CTS and POL
pointed toward the occurrence of various constituents as revealed in Table 3.1. CTS
extract was found to be rich with glycosides, steroids, saponin, coumarine and resins,
but on other hand POL extract was rich in anthraquinones, fatty acids, flavonoids,
tannin and resins.
Table 3.1: Qualitative phytochemical screening of CTS and POL methanolic extracts
Sr. No.
Constituents
CTS
POL
01 Glycosides +++ +
02 Steroids +++ ++
03 Alkaloids ++ +
04 Terpenoids + ++
05 Fatty acids + +++
06 Saponin +++ +
07 Flavonoids + +++
08 Tannins + ++
09 Coumarine +++ ++
10 Anthraquinones - +++
11 Anthocyanin - -
12 Endomine - +
13 Resins +++ ++ +++, high concentration; ++, medium concentration; +, low concentration; -, not detected

103
3.7.2 Quantitative Methods for Screening of Phytoconstituents
Screening assay disclose presence of plant primary or secondary metabolites
(phytochemicals). Phyto-constituent screening assays, either the qualitative or
quantitative, are easy and inexpensive protocols used for rapid clue about the presence
of particular phyto compound in plant matrix. Table 3.2 demonstrates the percentage
of alkaloids, pectin, soluble pigments, saponin, flavonoids and tannins in methanolic
crude extracts of CTS and POL. The concentration of alkaloids was found to be very
high i.e. 15.82+0.06 in POL extract and while the CTS methanolic extract also has the
highest percentage of alkaloids. Pectin and saponins was also found in significant
quantity.
Table 3.2: Percent phytochemical components in CTS and POL methanolic extracts.
3.8 Antioxidant Potential of Biologically Active Plant
Extract Fractions
The antioxidant potential of metanolic and ethyl acetate extracts of CTS and POL was
determined by following the different reported procedure for total phenol, flavonoids,
tannin, carotenoid and free radical scavenging potential etc.
3.8.1 Total Phenolic, Total Flavonoids and Total Carotenoid Contents
Phenolic compounds are produced in plants in response various factors like UV
radiation, stress like drought, nutrition deficit, harsh temperature and pressure
condition and defensive action against pathogens. These compounds are considered as
Sr. No. Constituents % CTS POL
1 Alkaloids 7.84± 0.001 15.82+0.06
2 Pectin 5.4± 0.431 6.61± 0.031
3 Soluble pigments 2.9740± 0.004 1.035+0.013
4 Saponin 3.08± 0.011 1.76± 0.008
5 Tannins 0.169± 0.004 0.898+0.001

104
significant therapeutically active constituent that deciphered a wide range of activities
like anticancer, antiviral, antibacterial, antifungal, anti-allergic etc. The total phenolic
content of the different fractions of CTS and POL were determined by a modified
Folin-Ciocalteu method and results were expressed in graphical forms (Fig. 3.9). TPC
were resolute in this study weighed against with standard (gallic acid) and final results
are articulated in term of milligrams of gallic acid equivalent (mg GAE/g dry sample).
In present study phenolic contents were found to be highest in concentration
methanolic extract of POL and CTS while their concentration is reduced in case of
ethyl acetate extracts of both plants. So the methanol was found to be the best
extraction solvent for phenols in this study.
Flavonoids being phenolic glycosides donate elevated antioxidant bustle of plant
owing to their metal chelating and scavenging attribute thus act to control
pathophysiology related to oxidative stress. Structure of flavonoid contain a central
backbone of three rings and sometime occur naturally in conjugated form along with a
glycone moiety. Oligomeric form of flavonoid is Proanthocyanidin. Due to the
presence of phenolic group both they operate like antioxidant, phytoestrogen, reduce
the risk of inflammation and cancer (Middleton et al., 2000; Noroozi et al., 1998).
Flavonoids concentration in sample STSM, STSE, POLM, and POLE is 42.15, 18.35,
47 and 35 respectively present as mg/g Catechin equivalents. The projected
flavonoids content of CTS and POL extracts are expressed as mg/g Catechin
equivalents in fig. 3.10.
In plants the carotenoid play an important role in photosynthesis process. The
concentration of carotenoid is also increased during ripening of fruits. These are main
precursor required for the formation of vitamin A and β-carotene in mammals. Owing
to their free radical scavenger and antioxidant potential they have the capacity to
reduce cancer by modulating immune response. In the present study carotenoid
contents were found in agreement with the previous findings. Results of total
carotenoids contents are expressed as μg/g of the samples also revealed a correlation
among antioxidant potential and total carotenoid contents of plant samples which is
also evident from the graphs (Fig. 3.11). All measurements were done in triplicate and
results were averaged.
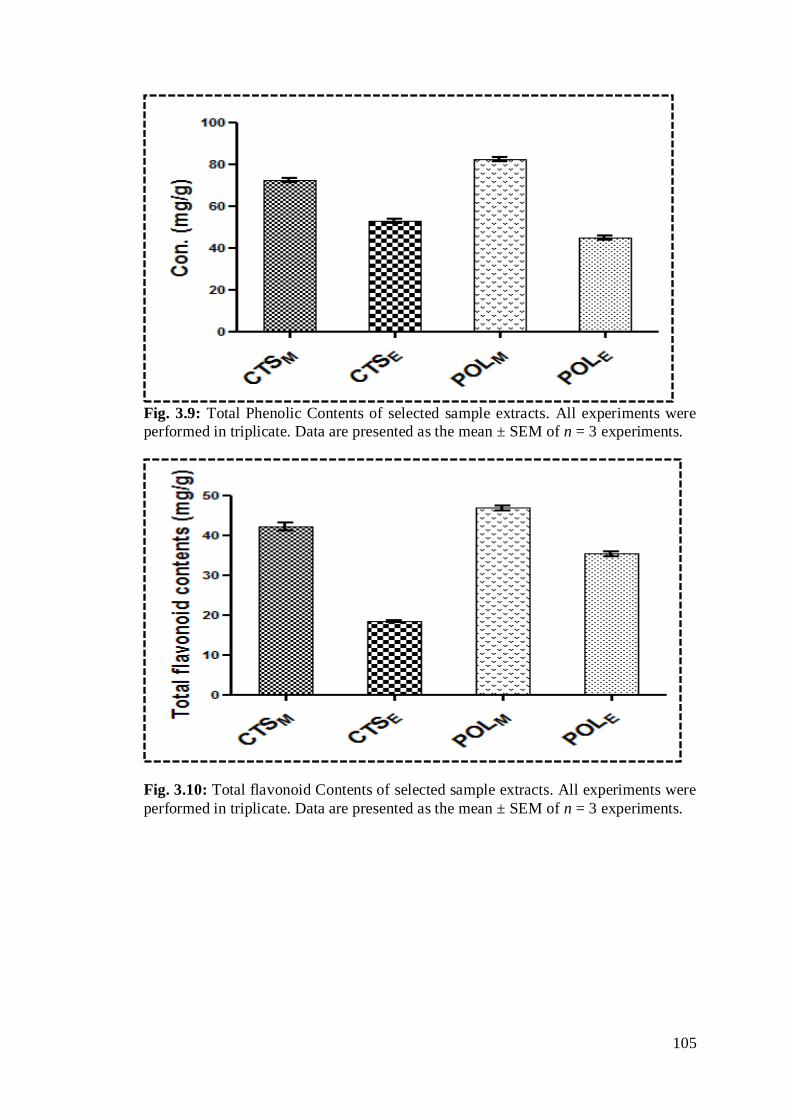
105
Fig. 3.9: Total Phenolic Contents of selected sample extracts. All experiments were
performed in triplicate. Data are presented as the mean ± SEM of n = 3 experiments.
Fig. 3.10: Total flavonoid Contents of selected sample extracts. All experiments were
performed in triplicate. Data are presented as the mean ± SEM of n = 3 experiments.

106
Fig. 3.11: Total carotenoid contents of selected sample extracts. All experiments were
performed in triplicate. Data are presented as the mean ± SEM of n = 3 experiments.
3.8.2 Free Radical Scavenging Activity of Extracts by DPPH, ABTS and FRAP
Assays
H-ion-donating power of polyphenols is considered an imperative biologically
property that convert potentially damaging species of reactive oxygen into the
nontoxic one. Particularly, DPPH• appear quick in action in determining antioxidant
potential for rapid conversion of labile hydrogen atom into radicals. CTSM, CTSE,
POLM and POLE extracts antiradical values against DPPH were found to be 81.747,
58.407, 45.511 and 36.124 respectively.
In vitro antioxidant potential like free radical scavenging action of CTS and POL
methanolic and ethyl acetate extracts was also resolute by the ABTS•+ assay. The
ABTS•+ cation radical is preferred over DPPH free radical based assay when sample
interference or solubility come up. The ABTS•+ scavenging data (Fig 3.13) propose
that extracts components have the potential to scavenge free radicals of ABTS and
thus protect susceptible subject from free radical mediated oxidative damage.
Among all the assays use for determination of antioxidant potential of compounds and
phytoextract, the FRAP assay is the most simple, fast, inexpensive, reproducible and
highly reliable method (Benzie & Strain, 1999). Antioxidant compounds can

107
effectively reduce Fe (III) to Fe (II) exhibiting high reducing power values. Ferric
reducing antioxidant power of selected plant extracts is shown in Fig.3.14.
Fig. 3.12: DPPH free radical scavenging potential of selected sample extracts. All
experiments were performed in triplicate. Data are presented as the mean ± SEM of n
= 3 experiments.
Fig. 3.13: ABTS free radical scavenging potential of selected sample extracts. All
experiments were performed in triplicate. Data are presented as the mean ± SEM of n
= 3 experiments.
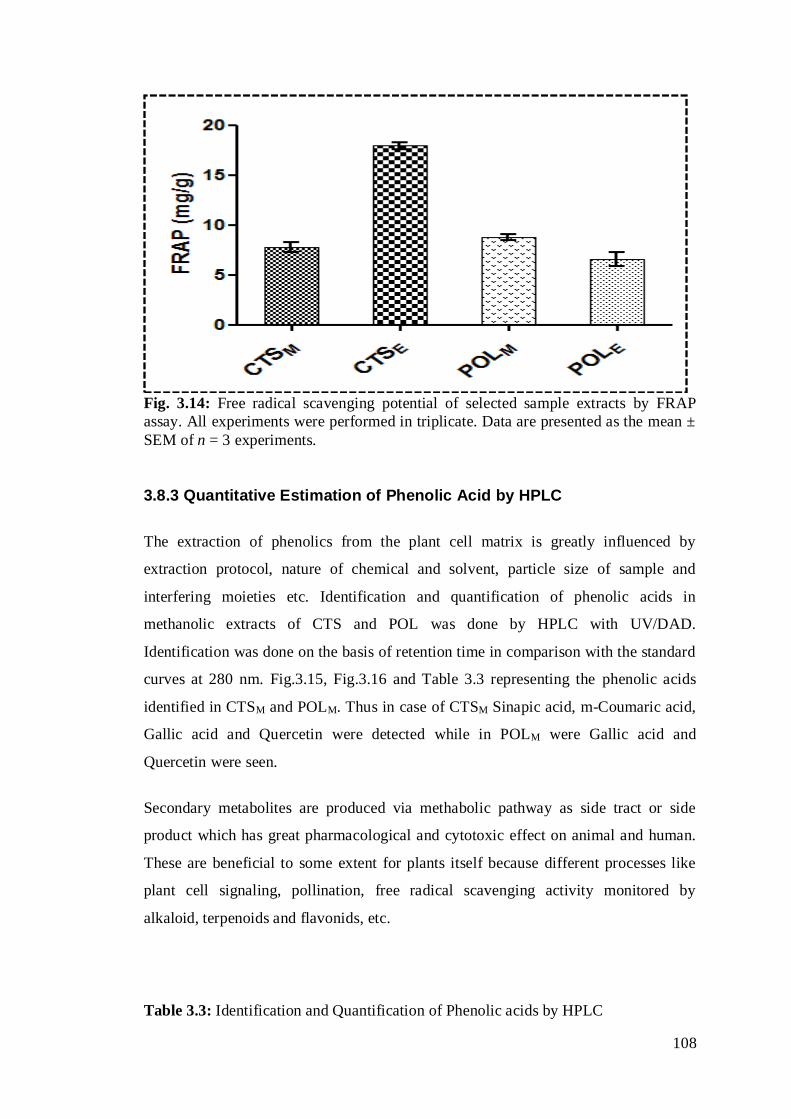
108
Fig. 3.14: Free radical scavenging potential of selected sample extracts by FRAP
assay. All experiments were performed in triplicate. Data are presented as the mean ±
SEM of n = 3 experiments.
3.8.3 Quantitative Estimation of Phenolic Acid by HPLC
The extraction of phenolics from the plant cell matrix is greatly influenced by
extraction protocol, nature of chemical and solvent, particle size of sample and
interfering moieties etc. Identification and quantification of phenolic acids in
methanolic extracts of CTS and POL was done by HPLC with UV/DAD.
Identification was done on the basis of retention time in comparison with the standard
curves at 280 nm. Fig.3.15, Fig.3.16 and Table 3.3 representing the phenolic acids
identified in CTSM and POLM. Thus in case of CTSM Sinapic acid, m-Coumaric acid,
Gallic acid and Quercetin were detected while in POLM were Gallic acid and
Quercetin were seen.
Secondary metabolites are produced via methabolic pathway as side tract or side
product which has great pharmacological and cytotoxic effect on animal and human.
These are beneficial to some extent for plants itself because different processes like
plant cell signaling, pollination, free radical scavenging activity monitored by
alkaloid, terpenoids and flavonids, etc.
Table 3.3: Identification and Quantification of Phenolic acids by HPLC
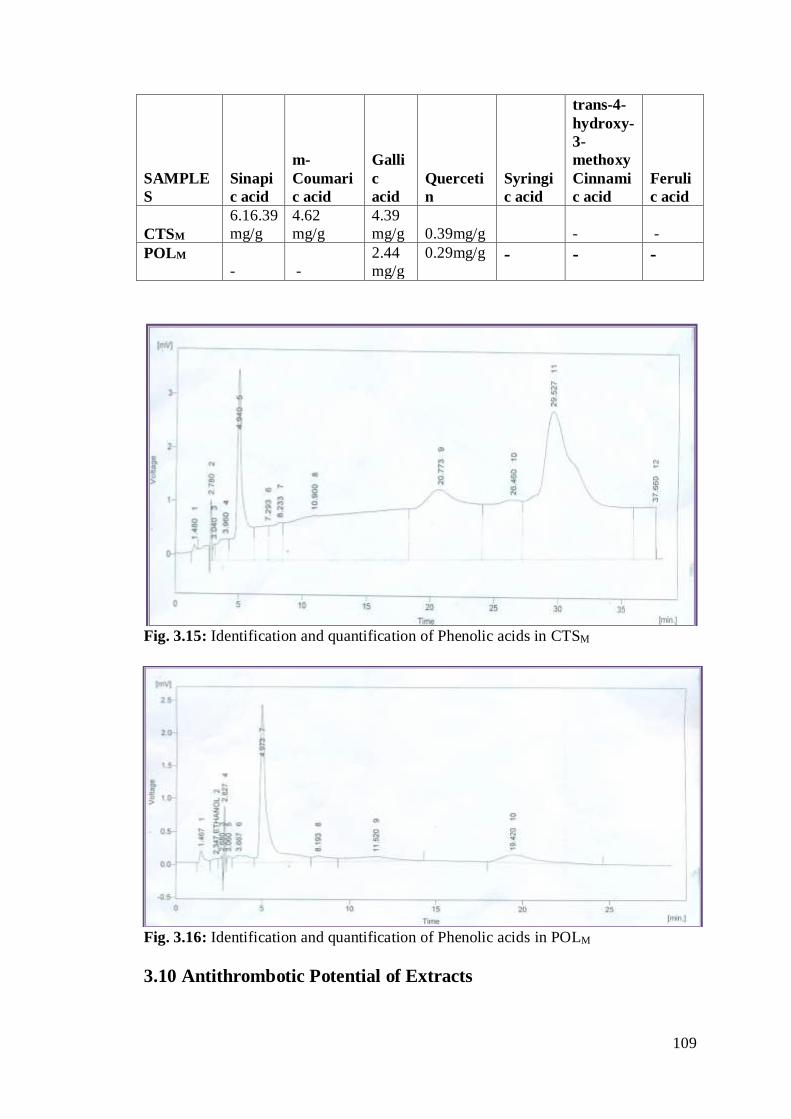
109
SAMPLE
S
Sinapi
c acid
m-
Coumari
c acid
Galli
c
acid
Querceti
n
Syringi
c acid
trans-4-
hydroxy-
3-
methoxy
Cinnami
c acid
Feruli
c acid
CTSM
6.16.39
mg/g
4.62
mg/g
4.39
mg/g 0.39mg/g - -
POLM
- -
2.44
mg/g
0.29mg/g - - -
Fig. 3.15: Identification and quantification of Phenolic acids in CTSM
Fig. 3.16: Identification and quantification of Phenolic acids in POLM
3.10 Antithrombotic Potential of Extracts
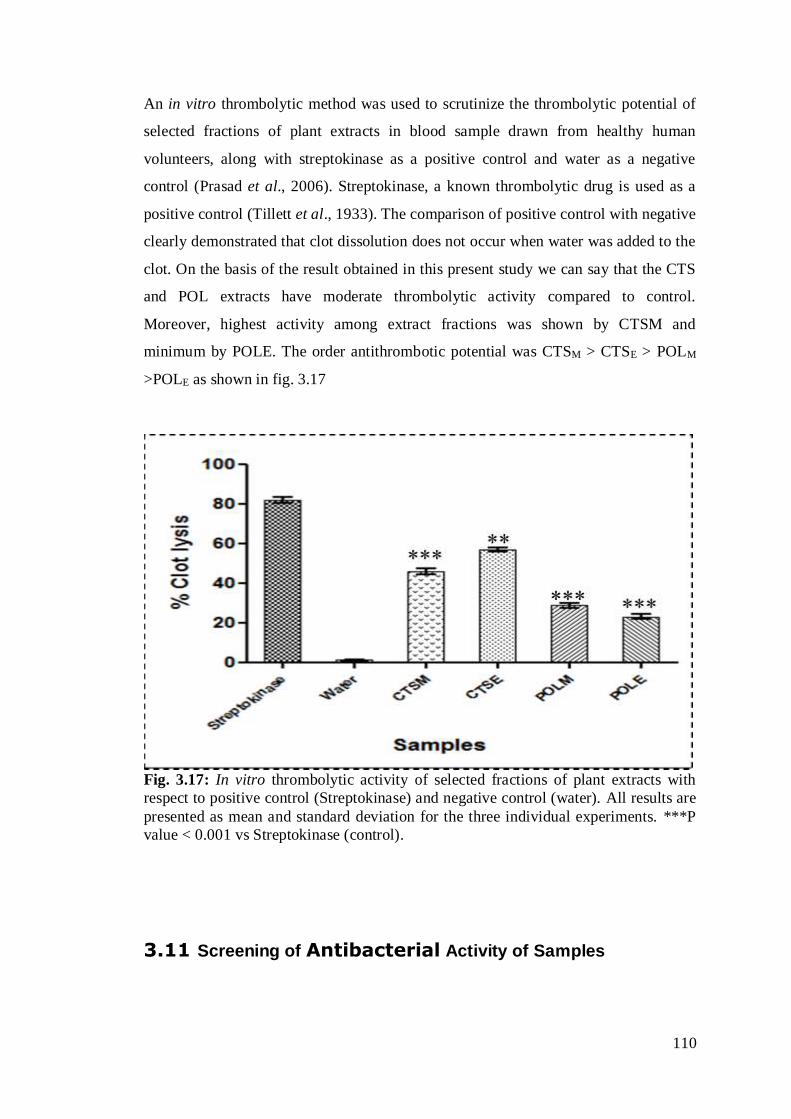
110
An in vitro thrombolytic method was used to scrutinize the thrombolytic potential of
selected fractions of plant extracts in blood sample drawn from healthy human
volunteers, along with streptokinase as a positive control and water as a negative
control (Prasad et al., 2006). Streptokinase, a known thrombolytic drug is used as a
positive control (Tillett et al., 1933). The comparison of positive control with negative
clearly demonstrated that clot dissolution does not occur when water was added to the
clot. On the basis of the result obtained in this present study we can say that the CTS
and POL extracts have moderate thrombolytic activity compared to control.
Moreover, highest activity among extract fractions was shown by CTSM and
minimum by POLE. The order antithrombotic potential was CTSM > CTSE > POLM
>POLE as shown in fig. 3.17
Fig. 3.17: In vitro thrombolytic activity of selected fractions of plant extracts with
respect to positive control (Streptokinase) and negative control (water). All results are
presented as mean and standard deviation for the three individual experiments. ***P
value < 0.001 vs Streptokinase (control).
3.11 Screening of Antibacterial Activity of Samples

111
Infectious diseases are ailment caused by the attack of particular pathogens like
viruses or bacteria or fungus or protozoan parasite and their byproducts like toxin,
capsule etc. These infections may also appear as a result of pathogen transmission as
causative agent from infected person to the other host including human, animal etc.
Host susceptibility for infection, severity and progression of infection, depend several
factors like age, gene, gender, status of immunity and nutrition etc. Infectious disease
considered leading cause of mortality in developing and underdeveloped countries
accounting above 26 % of total reported deaths (Becker et al., 2006). No doubt after
the discovery of antibiotic in 1926 and later development of its various kinds totally
revolutionize the therapeutic of infections. Although marvelous effect of antibiotic is
to cure the infection but on other hand it create resistance problem in microbe.
Horrible examples of resistant mutated microbe emerge that become serious problem
of today because they become difficult to treat (Barbour et al., 2004). Resistance
problem in microbe come into sight as a result of microbe gene transformation
between different species of host, gene mutagenesis or by the involvement of
transposons, cryptic genes activation and chromosomal aberration as result of
interaction between antibiotic induce oxidative stress and chromosome. Moreover,
another main issue linked with the use of antibiotic is side effects that appear after its
use. The domino effect of the arbitrary misuse of antibiotic is the development of
resistant strains in animals and human (Barton, 2000). The cost issue is also another
hindrance that cannot be ignored, especially in low income community.
The antimicrobial activity of methnolic extracts of CTS and POL against Gram
positive and Gram negative bacteria was determined by disc diffusion method. Zones
of inhibition were measured and tabulated in Table 3.5. Gram negative strains of
bacteria such as Pseudomonas aerugenosa was inhibited by both extracts while
Escherichia coli was not inhibited by extract of POL. Similarly CTS does not exhibit
potential against Salmonella typhi. While on other hand, both CTS and POL exhibit
significant potential against all selected strains of gram positive bacteria (Bacillus
subtilis, Styphylococcus aureus, and Styphylococcus epidermidis). Percent inhition
with respect to the positive control (Ciproflaxacin) is also expressed in Fig. 3.18.
Table 3.4: Zones of Inhibition (mm) in Disc Diffusion Plate for the antibacterial
activity of successively extracted alcoholic extracts (SAE) of plants that have also
antiviral potential.

112
Bacterial
Strains
Test organism Zone of inhibition (M+SD) mm
CTS POL Ciproflaxacin
(Positive
control)
Gram positive Bacillus subtilis 8.5 ± 0.1 12± 0.4 23
Styphylococcus aureus 7.33 ±
0.05
11± 0.02 19
Styphylococcus
epidermidis
12.66 ±
0.02
12± 0.16 18
Gram
negative
Pseudomonas
aerugenosa
8.30 ±
0.06
11.86 ±
0.05
20
Escherichia coli 6.23±
0.04
- 20
Salmonella typhi - 8.35±
0.03
25
The results are mean ± S.D. diameter of average of growth inhibition zone of three
replicate. SD= Standard deviation
Fig. 3.18: Percent inhibition of bacteria by selected extracts with respect to
ciproflaxacin (positive control)
3.12 Extraction, Purifcation and identification of
Compounds Obtained from CTSE Extract

113
3.12.1 Compound CT-EtOAc-01
OO
O
O
O
OO
OO
O
OOO
1
3 5 7
8
10
11 12
15
1713
1'
3'
5'
1'2''6''
4''
1'''2'''
4'''1'''
4'''
2'''
1'''2'''
5'''
Cym ICym IICymIII
1H-NMR 1H-NMR (400 MHz, CDCl3) δ (ppm): 8.10-8.12 (m, 1H, H-5’’), 7.85-7.86 (dd, J =
1.6, 9.6 Hz (2H, H-3’’ & H-7’’), 7.83-7.84 (m, 2H, H-4’’ & H-6’’), 4.59-4.61 (m, 1H,
H-3), 3.86-3.90 (m, 1H, H-12), 3.67 (s, 3H, CO2CH3), 2.17-2.72 (m, 1H, H-1’), 2.12-
2.16 (m, 1H, H-17), 2.01-2.03 (m, 4H, H-6 & H-7), 1.85-1.89 (m, 1H, H-8/H-14),
1.387 (m, 1H, H-8/H-14), 1.78-1.87 (m, 2H, H-16), 1.71-1.76 (m, 2H, H-15), 1.63-
1.66 (m, 2H, H-2’), 1.68 (br. d, J = -10 Hz, 2H, H-11), 1.55-1.61 (m, 1H, H-14), 1.50
(dddd, J = -12.0, -12.0, 4.5, 4.5 Hz, 1H, H-5), 1.45 (m, 3H, H-1 & H-9), 1.24-1.28 (m,
2H, H-2), 1.23 (s, 3H, H-18), 1.12-1.15 (m, 2H, H-3’), 1.098 (s, 3H, H-19), 1.03-1.00
(3, 3H, H-5’)
Cym-I: 4.48-4.51 (dd, J =1.6, 9.6 Hz, 8H, H-1’’', H-2’’’, H-4’’’), 3.40-3.55 (m, 4H, H-
3’’’), 3.39 (s, 3H, O-CH3), 1.30-1.33 (m, 1H, H-5’’’), 1.45-1.60 (m, 9H, CH3 at C-5’’’)
Cym-II: 4.48-4.51 (dd, J=1.6, 9.6 Hz, 8H, H-1’’’, H-4’’’, H-5’’’), 3.40-3.55 (m, 4H, H-
6’’’), 3.43 (s, 3H, O-CH3), 1.45-1.60 (m, 9H, CH3 at C-2’’’)
Cym-III: 4.48-4.51 (dd, J=1.6, 9.6 Hz, 8H, H-1’’', H-2’’’), 3.45 (s, 3H, O-CH3), 2.28-
2.32 (m, 1H, H-5),1.82-1.89 (m, 4H, H-3’’’, H-4’’’), 1.45-1.60 (m, 9H, CH3 at C-5’’’).
13C-NMR 13C-NMR (300MHz, CDCl3) δ (ppm): 170.2 (CO2Me), 160.0(C-1’’), 133.0(C-5’’),
132.0(C-1’’), 129.6(C-3’’ & C-7’’), 129.7 (C-4’’ & C-6’’), 82.8 (C-3), 68.3 (C-12), 40.7
(C-5), 49.9 (C-1’), 55.0 (C-13), 46.1(C-14), 46.1(C-8), 46.1 (C-9), 43.7 (C-17), 40.7
(C-5), 39.0 (C-10), 37.1(C-7), 35.7 (C-4, C-11), 31.8 (C-1), 29.1(C-6), 28.5 (C-2, C-
16), 27.5 (C-2’, C-15), 26.2(C-3’), 25.6 (C-4’), 17.9 (C-5’), 12.30 (C-19), 10.0 (C-18)
Cym-I
101.2 (C-1’’’), 76.9 (C-2’’’), 72.8 (C-4’’’), 71.3 (C-5’’’), 55.7 (O-CH3), 34.3 (C-3’’’),
18.6 (CH3 at C-5’’’)

114
Cym-II
99.1(C-1’’’), 77.0 (C-4’’’), 73.9 (C-5’’’), 71.4 (C-2’’’), 58.1 (O-CH3), 34.3 (C-5’’’), 19.2
(CH3 at C-2’’’)
Cym-III
95.6(C-1’’’), 81.0(C-2’’’), 71.7(C-5’’’), 62.0(O-CH3), 35.6(C-3’’’), 35.6 (C-4’’’), 21.5
(CH3 at C-5’’’)
The compound CT-EtOAc-01 was isolated as colorless amorphous powder (24mg)
with melting point 217-220°C, Rf value of 0.42 and [α]D25 of 18.61° (MeOH, C =
0.05). The appearance of absorption signals at 1710, 1608, 1504, 1235 cm-1 in IR
indicates the presence of C=O (ester), C=C (aromatic) and C-O functionalities
respectively.
The positive Lieberman-Burchard’s Molisch and Keller Kiliani tests confirmed the
compound to be steroidal glycoside with 2-deoxysugars. The HR ESI MS exhibited
[M+Na] at 964 amu which was assigned to molecular formula C54H84O13 with 13
double bond equivalent (DBE). The 13C NMR revealed the presence of 54 carbon
resonances that appeared as: 10 methyl, 24 methine, 15 methylene and 5 quaternary
carbons. The Broadband (BB), DEPT (45°, 90° and 135°) and COSY analyses
confirmed aglycon to be comprised of 33 carbons. The detailed NMR analyses also
revealed the presence of two ester carbonyl carbons which appeared at 170.2 and 160
ppm in 13C NMR and were rationalized to be methyl ester moiety and benzoylic
carbonyl respectively. The ester linkages were located to be at C-12 and C-1’ due to a
significant downfield shift of the methine protons to which these ester functionalities
were bound. C-17 appeared at 2.12-2.16 ppm while C-12 appeared at 3.86-3.90 ppm
which is quite low field as compared to other methine protons.
The structure of the aglycone was solved from NMR analyses. The three methyls of
the aglycone appeared at: 1.23 (s, H-18), 1.098 (s, H-19) and 1.03-1.00 (m, H-5’). The
side chain at C-17 was identified by HSQC analysis which indicated presence of one
methyl, one oxygenated methyl, 3 methylene, 1 methine and one quaternary carbon
(carbonyl carbon). The HSQC confirmed the assignment of these protons which were
found correlate with methyl carbons at 12.0, 9.5 and 17.9 ppm respectively. The
HSQC spectrum confirmed C-10 and C-13 to be quaternary carbon while the C1’ was
inferred to be attached with a triplet carbon (i.e. CH2) and a methine (C-17). The
oxygenated methine carbons of the aglycon moiety (i.e., C-3 & C-12) appeared at

115
3.86-3.90 ppm and 4.59-4.61 ppm which were correlated with oxygenated methine
carbons at 82.8 and 68.3 ppm respectively by HSQC experiment.
The UV spectrum showed λmax 227, 268 and 283 nm which indicated presence of
benzoyl system. The appearance of 5 protons (as three sets of multiplets with 2, 2 and
1 H integration), at 7.85-7.86, 7.83-7.84 and 8.09-8.12 ppm indicated the presence of
a deshielded aromatic system which correlated with carbon resonances at 129.6, 129.7
and 133.0 ppm respectively. The singlet aromatic carbon appeared at 132.0 ppm. The
HMBC assignment showed a clear 3J correlation between the benzoyl carbonyl (δC:
160.0) and C-12 (δC: 82.8; δH: 3.86-3.90 ppm) which confirmed the benzoyl moiety to
be attached at C-12. Similarly the location of methyl ester was confirmed by the
correlation with C-1’. The absolute configuration of the isolated compound was left
unresolved.
In addition to the aglycone signals, the 13C NMR also exhibited 21 signals which were
assigned to the sugar part which was found to comprise of three hexoses. The 13C
NMR showed the presence of three anomeric carbons which appeared at 95.6, 99.1
and 101.2 ppm. These values were correlated in HSQC experiment with the multiplet
signal of 8H integration at 4.48-4.51 ppm. Since the peaks were not well resolved,
exact and detailed assignment could not be performed.
The attachment of sugar moiety was confirmed to be at C3 by the significant
downfield shift of H3 in 1H NMR (δH: 4.59-4.61 ppm) and its corresponding carbon
resonance (δC: 68.3 ppm). The long range correlation between C3 and Cym-I also
confirmed the attachment of sugar to be at C17. Therefore the compound was
determined to be methyl(12-O-benzoyl-androstan-3-O-(14)-β-D-cymaropyranosyl-
(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl)hexanoate.
3.12.2 Compound CT-EtOAc-02
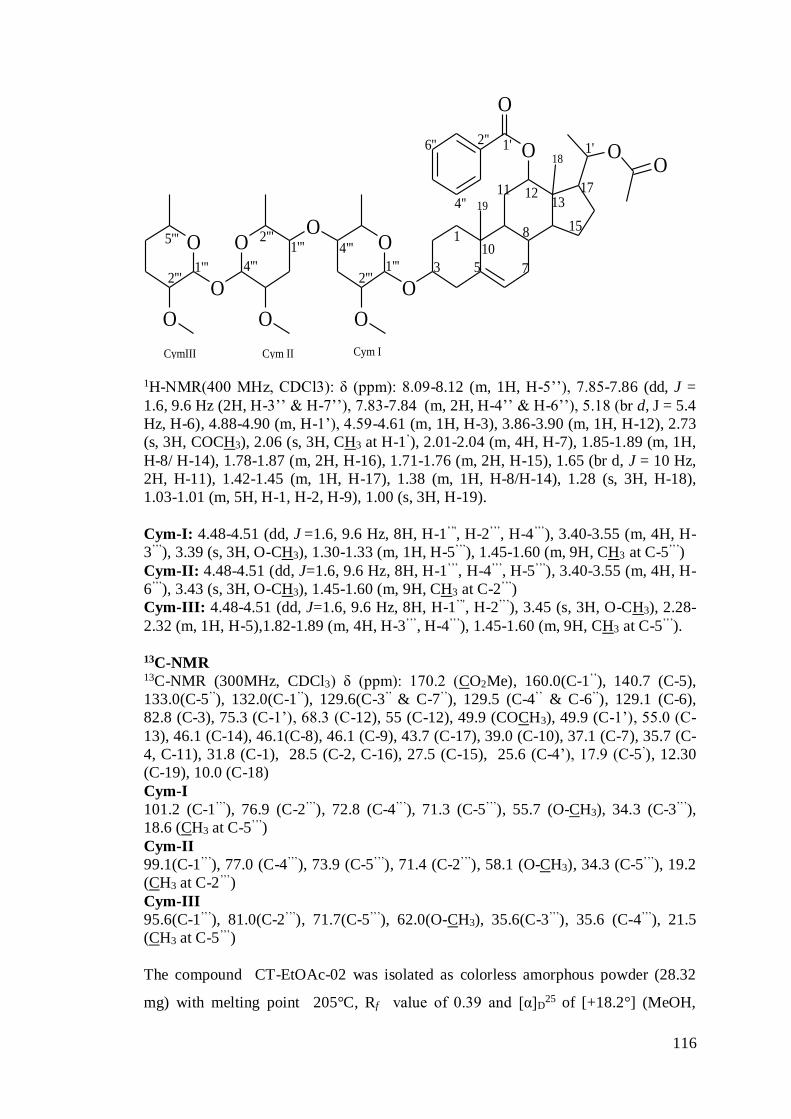
116
OO
O
O
OO
OO
O
OOO
1
3 5 7
8
10
11 12
15
1713
1'1'2''6''
4''
1'''2'''
4'''1'''
4'''
2'''
1'''2'''
5'''
18
19
Cym ICym IICymIII
O
1H-NMR(400 MHz, CDCl3): δ (ppm): 8.09-8.12 (m, 1H, H-5’’), 7.85-7.86 (dd, J =
1.6, 9.6 Hz (2H, H-3’’ & H-7’’), 7.83-7.84 (m, 2H, H-4’’ & H-6’’), 5.18 (br d, J = 5.4
Hz, H-6), 4.88-4.90 (m, H-1’), 4.59-4.61 (m, 1H, H-3), 3.86-3.90 (m, 1H, H-12), 2.73
(s, 3H, COCH3), 2.06 (s, 3H, CH3 at H-1’), 2.01-2.04 (m, 4H, H-7), 1.85-1.89 (m, 1H,
H-8/ H-14), 1.78-1.87 (m, 2H, H-16), 1.71-1.76 (m, 2H, H-15), 1.65 (br d, J = 10 Hz,
2H, H-11), 1.42-1.45 (m, 1H, H-17), 1.38 (m, 1H, H-8/H-14), 1.28 (s, 3H, H-18),
1.03-1.01 (m, 5H, H-1, H-2, H-9), 1.00 (s, 3H, H-19).
Cym-I: 4.48-4.51 (dd, J =1.6, 9.6 Hz, 8H, H-1’’', H-2’’’, H-4’’’), 3.40-3.55 (m, 4H, H-
3’’’), 3.39 (s, 3H, O-CH3), 1.30-1.33 (m, 1H, H-5’’’), 1.45-1.60 (m, 9H, CH3 at C-5’’’)
Cym-II: 4.48-4.51 (dd, J=1.6, 9.6 Hz, 8H, H-1’’’, H-4’’’, H-5’’’), 3.40-3.55 (m, 4H, H-
6’’’), 3.43 (s, 3H, O-CH3), 1.45-1.60 (m, 9H, CH3 at C-2’’’)
Cym-III: 4.48-4.51 (dd, J=1.6, 9.6 Hz, 8H, H-1’’', H-2’’’), 3.45 (s, 3H, O-CH3), 2.28-
2.32 (m, 1H, H-5),1.82-1.89 (m, 4H, H-3’’’, H-4’’’), 1.45-1.60 (m, 9H, CH3 at C-5’’’).
13C-NMR 13C-NMR (300MHz, CDCl3) δ (ppm): 170.2 (CO2Me), 160.0(C-1’’), 140.7 (C-5),
133.0(C-5’’), 132.0(C-1’’), 129.6(C-3’’ & C-7’’), 129.5 (C-4’’ & C-6’’), 129.1 (C-6),
82.8 (C-3), 75.3 (C-1’), 68.3 (C-12), 55 (C-12), 49.9 (COCH3), 49.9 (C-1’), 55.0 (C-
13), 46.1 (C-14), 46.1(C-8), 46.1 (C-9), 43.7 (C-17), 39.0 (C-10), 37.1 (C-7), 35.7 (C-
4, C-11), 31.8 (C-1), 28.5 (C-2, C-16), 27.5 (C-15), 25.6 (C-4’), 17.9 (C-5’), 12.30
(C-19), 10.0 (C-18)
Cym-I
101.2 (C-1’’’), 76.9 (C-2’’’), 72.8 (C-4’’’), 71.3 (C-5’’’), 55.7 (O-CH3), 34.3 (C-3’’’),
18.6 (CH3 at C-5’’’)
Cym-II
99.1(C-1’’’), 77.0 (C-4’’’), 73.9 (C-5’’’), 71.4 (C-2’’’), 58.1 (O-CH3), 34.3 (C-5’’’), 19.2
(CH3 at C-2’’’)
Cym-III
95.6(C-1’’’), 81.0(C-2’’’), 71.7(C-5’’’), 62.0(O-CH3), 35.6(C-3’’’), 35.6 (C-4’’’), 21.5
(CH3 at C-5’’’)
The compound CT-EtOAc-02 was isolated as colorless amorphous powder (28.32
mg) with melting point 205°C, Rf value of 0.39 and [α]D25 of [+18.2°] (MeOH,

117
C=0.05). The IR absorption signals at 1734, 1724, 1456, 1377, 1274 cm-1 in indicates
the presence of two C=O (ester), C=C (aromatic) and C-O functionalities.
Positive Molisch, Lieberman-Burchard, and Keller Kiliani tests confirmed the
compound to be steroidal glycoside comprising 2-deoxysugars. The HR ESI MS
exhibited [M+Na] at 919 amu which was assigned to molecular formula C51H76O13
with 14 double bond equivalent (DBE). The 13C NMR and the DEPT spectras of
compound revealed the presence of 51 carbon resonances that appeared as: ten
methyl, twenty two methine, eleven methylene and six quaternary carbons. The
Broadband (BB), DEPT (45°, 90° and 135°) and COSY analyses confirmed aglycon
to be comprised of 33 carbons. The detailed NMR analyses also revealed the presence
of two ester carbonyl carbons which appeared at 171.2 and 161 ppm in 13C NMR and
were rationalized to be acetoxy group at C-1’ and benzylic carbonyl respectively. The
ester linkages were located to be at C-12 and C-1’ due to a significant downfield shift
on the methine protons to which these ester functionalities were bound. C-1’ appeared
as multiplet at 4.88-4.90 ppm or δ while C-12 appeared at 3.86-3.90 ppm which is
quite low field as compared to other methine protons. Moreover, appearance of
singlet for three protons of at 2.73 ppm also specifies the presence of methyl of
acetoxy group. Appearance of signal of olefinic proton at 5.18 δ (br d) in 1H NMR
spectrum point toward the confirm presence of Δ5 pregnane glycoside. Presence of
aromatic rings in aglycone backbone was also confirmed by UV which exhibited max
at 270 nm.
The structure of aglycone part was solved from NMR analyses. The three methyls of
the aglycone appeared at 1.00 (methyl of C19), 2.73 ppm (COCH3) and 1.24-1.28 ppm
(methyl of C18). The side chain at C-17 was identified by HSQC analyses which
indicated presence of one methine to which one methyl is bound. The HSQC
spectrum confirmed C-10 and C-13 to be attached with quaternary carbon while the
C-1’ was inferred to be attached with acetoxy moiety. The oxygenated methine
carbons of the aglycon moiety appeared at 3.86-3.90 ppm (H-12) and 4.59-4.61 ppm
(H-3) which was correlated with oxygenated methine carbons at 68.3 and 82.8 ppm
respectively by HSQC experiment.

118
In addition to the aglycone signals, the 13C NMR also exhibited 21 signals which were
assigned to the sugar part which was found to comprise of three hexoses. The 13C
NMR showed the presence of three anomeric carbons which appeared at 95.6, 99.1
and 101.2 ppm. These values were correlated in HSQC experiment with the multiplet
signal of 8H integration at 4.48-4.51 ppm. Since the peaks were not well resolved,
exact and detailed assignment could not be performed.
The attachment of sugar moiety was confirmed to be at C-3 by the significant
downfield shift of H3 in 1H NMR (δH: 4.59-4.61 ppm) and its corresponding carbon
resonance (δC: 68.3 ppm). The long range correlation between C3 and Cym-I also
confirmed the attachment of sugar to be at C17. Therefore the compound was
determined to be methyl (12-O-benzoyl-androstan-3-O-(14)-β-D-cymaropyranosyl-
(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl) hexanoate.
3.12. 3 Compound CT-EtOAc-03
O
O
O
OO
OO
O
OOO
1
3 5 7
810
11 12
15
1713
1'1'2''6''
4''
1'''2'''
4'''1'''
4'''
2'''
1'''2'''
5'''
18
19
Cym ICym IICymIII
3'
5'
2''''
4''''
Compound CT-EtOAc-03 (C55H86O11) was obtained from the ethyl acetate extract of
Caralluma tuberculata as colourless, amorphous powder in 33 mg along with [α]D25
[+17.2°] (MeOH, C=2). Rf value calculated from TLC was 0.41.
Significant peaks obtained from IR spectra are at 1720, 1507, 1200 indicate the
presence of ester, and aromatic functional groups. The molecular weight of the
compound was determined by obtained by high resolution ESI-MS which exhibited
[M+Na] at 946 amu which suggested molecular formula of compound to be

119
C54H84O13 with 13 double bond equivalent (DBE). In this structure total number of
hydrogen atoms are eighty six and carbon atoms are fifty five among which
quaternary carbons are four, tertiary carbon (methine group) are nineteen, secondary
carbon (methylene group) are fifteen and primary carbon (methyl group) are seven.
Moreover three methoxy groups are also present.
Most of spectral signal appeared on the same position as in the previous one with a
little bit variation only the difference is of the disappearance of singlet peak of methyl
ester protons at 3.37 δ. And this position was occupied by appearance of additional
protons due to isopropylic group indicate the given compound. A multiplet peak
ranging from 1.01 to 0.98 indicates H-1’. Presence of double bond between C-6 & C-
7 is indicated by doublets at position 5.73 & 6.12 δ for H-6 & H-7.
In addition to the aglycone signals, the 13C NMR also exhibited 21 signals which were
assigned to the sugar part which was found to comprise of three hexoses. The 13C
NMR showed the presence of three anomeric carbons which appeared at 95.6, 99.1
and 101.2 ppm these values were correlated in HSQC experiment with the multiplet
signal of 8H integration at 4.48-4.51s ppm. Since the peaks were not well resolved,
exact and detailed assignment could not be performed. Therefore the compound was
determined to be Methyl (23-O-benzoyl-androst-6-en-17-O-(14)-β-D-
cymaropyranosyl-(14)-β-D-cymaropyranosyl--(14)-β-D-cymaropyranosyl)
hexanoate.
3.12.4 Compound CT-EtOAc-04
OO
O
1
3 5 7
8
10
11 12
15
1713
1'
2''6''
4''
2'
1''O
O
OO
O
OO
1'''2'''
4'''1'''
4'''
2'''
Cym ICym II
4'
6'

120
The compound CT-EtOAc-04 (C45H58O10 ) was isolated as colorless amorphous
powder (28.3 mg) with melting point 196°C, Rf value of 0.54 and [α]D25 of [+24.6°]
(MeOH, C=0.1). The IR absorption signals at 1716, 1698, 1444, 1298 cm-1 in
indicates the presence of two C=O (ester), C=C (aromatic) and C-O functionalities.
Positive Molisch, Lieberman-Burchard, and Keller Kiliani tests confirmed the
compound to be steroidal glycoside comprising 2-deoxysugars. The HR ESI MS
exhibited [M+Na] at 782 amu which was assigned to molecular formula C45H58O10
with 17 double bond equivalent (DBE). The 13C NMR and the DEPT spectras of
compound revealed the presence of 45 carbon resonances that appeared as: four
methyl, twenty five methine, nine methylene and five quaternary carbons. The
Broadband (BB), DEPT (45°, 90° and 135°) and COSY analyses confirmed aglycon
to be comprised of 31 carbons. The detailed NMR analyses also revealed the presence
of two ester carbonyl carbons which appeared at 168.5 and 161 ppm in 13C NMR and
were rationalized to be benzoyl group at C-12 and benzyloxy carbonyl at C-1’
respectively. The ester linkages were located to be at C-12 and C-1’ due to a
significant downfield shift on the methine protons to which these ester functionalities
were bound. Therefore the compound was determined to be Phenyl (12-O-benzoyl-
gonan-6-en-5’’-O-(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl)
oxalate.
3.12.5 Compound CTEtOAc-05
OHH3CO
OO
O
O
HO
OHHO
HO3
5
7
89
1'
1''
3''

121
1H-NMR (400 MHz, CDCl3): δ (ppm): 8.09 (d, j=1.45 HZ, 2H), 7.84 (d, j=7.2Hz,
1H), 7.63-7.45 (m, 2H), 7.50-7.40 (m, 3H), 7.24-7.105 (m, 1H), 4.89-4.72 (m, 5H),
4.47 (d, j=10.43, 1H), 3.79 (t), 3.66 (s, 3H, OMe),2.33 (t, 5H, J=7.63H), 1.99-2.11 (m,
1H),1.55-1.68 (m, 2H). 13C-NMR
Aglycone 13C-NMR (300MHz,CDCl3): δ (ppm): 144 (C-4), 138 (C-1), 137 (C-3), 132 (C-5),
130 (C-7), 127 (C-8), 126 (C-6), 122 (C-2), 60 (C-9), 58 (OME)
Glycone 13C-NMR (300MHz,CDCl3): 103 (C-1’), 80 (C-3’), 72 (C-5’), 71.8 (C-2’), 68 (C-4’),
65.2 (C-6’)
Glycone substituent: 13C-NMR (300MHz,CDCl3): 174 (C-1’’), 60 (C-3’’), 58 (C-2’’), 18 (C-4’’)
Compound 5 was obtained from the initial fractions of ethyl acetate extract of
Caralluma tuberculata during column chromatography using Hexane and ethyl acetate
as mobile fraction. This compound was colourless, amorphous powder of 22.51 mg
along with [α]D25 [+17.2ο] (MeOH, C=2). Rf value calculated from TLC was 0.61.
Significant peaks obtained from IR spectra are at 3500, 2810, 1740, 1470, 1230
indicate the presence of alcoholic, methane, ester, carbonyl and aromatic functional
groups. The molecular weight of the compound was determined by molecular ion
peak obtained by high resolution ESI-MS and it was [M]= 414 amu. Empirical
formula i.e C20H26O10 was calculated after the interpretation of spectral data of NMR
and with the unsaturation 13 double bond equivalent (DBE). The presence of total
twenty carbons in the glycone and aglycone moieties of the structure was confirmed
by 13CNMR. Presence of substituent at position 6’ was confirmed due to downfield
shift observed in 13CNMR at 65.2 ppm. Present compound was already reported but in
different plant species Capsicum annuum L.(Materiska et al., 2003) and its name was
determined to be 3-Hydroxy-2-methyl-propionic acid 3,4,5-trihydroxy-6-[4-(3-
hydroxy-propenyl)-2-methoxy-phenoxy]-tetrahydro-pyran-2-ylmethyl ester or
commonly called Trans-p-ferulylalcohol-4-O-(6-(2-methyl-3-hydroxypropionyl)
glucopyranoside.

122
3.13 Discussion
The momentous spread of life threatening disease like HCV, insufficient virus
structure targeted treatment, dearth of vaccination, greater side effects and high cost
of authenticated therapy prompted the researchers and health professional to explore
some suitable alternative but better targeted approaches to control this dilemma.
These alternatives must be efficacious, economical and free of such side effects.
Natural products either plant extract or the isolated pure compounds or the
standardized herbal extract offers the indefinite prospects for novel drug discovery
owing to their matchless chemical miscellany (Cos et al., 2006). Botanical extracts in
conjunction with their flurry of active constituents occupied a central position and
were used successfully against dreadful infectious diseases, various cancers and
metabolic disorders (Duraipandiyan et al., 2006). Turning toward
ethnopharmacognosy is helping us to discover numerous plant based constituents
having antioxidant, anticancer, antifungal, antibacterial and antiviral potential.
On the basis of folk knowledge, eastern system of medicine and exploration of plants
from the valleys and unexplored areas is the focus of research now a day (Ahmad et
al., 2008). The plant communities are losing their species richness at a high rate due
to intensive deforestation and unlimited expansion of urban areas. Through bioassay-
directed screening, structure–activity relationship, and biochemical investigation,
identification of bioactive compounds as potential molecular targets of HCV-NS3 is a
hot issue, which may offer impending expansion of potential anti-HCV regimens.
In the present study, fifteen indigenous medicinal plants (listed in Table 2.1) were
scrutinized for their therapeutic potential like anti-HCV activities (HCV whole virus
and its non-structural proteins NS3/NS4). Recently non-structural proteins of HCV
(NS3 and NS5B) are centres of attraction as promising targets for drug development
against HCV owing to their key role in HCV replication. The binding of anti HCV
agents to the extremely conserved domain at the junction of NS3 protease and
helicase domain results the discovery of distinctive class of DAAs (Saalau-Bethell et
al., 2012).
In first part of the study, plants were collected from the Soon-sakaser valley of
Punjab, Pakistan, after primer folkare information. The pre washed and dust free plant

123
samples were shadow dried at room temperature and ground in the form of fine,
homogeneous, coarse powder and their methanolic extracts were obtained after
successive extraction. Proper operation must be followed in order to obtain the
maximum surface area of the sample for solvent action and finally improve the
kinetics of analytical extraction. Extraction is the essence of medicinal plant research
that helps to obtain maximum desirable constituents from plants to utilize further for
mining and depiction of all details. Extraction should be cautious enough to assure
that no active principal or compound was lost or damaged or distorted during this
procedure (Farnsworth, 1966). Selection of solvent type depends upon the nature of
targeted compound, for example polar solvents (ethanol, methanol and ethyl acetate)
are usually preferred for isolation of hydrophilic compounds (Cos et al., 2006).
Similarly, sorts of extraction schemes were used to obtain crude extracts depending
upon the type of test material, i.e. fresh or dry plants and their powder.
In vitro cellular toxicity studies were done for all plant extracts using Huh-7 and CHO
cells by Trypan blue exclusion method and MTT assay. Phyto extracts exhibited a
broad range of cytotoxic amount varying from the 1.5 µg/mL to the 200 µg/mL.
Cytotoxicity data of only two promising plants POL and CTS, which latter exhibit
strong anti HCV potential in activity guided assay, is reported in Fig. 3.2 (A & B)
while Cytotoxicity data through MTT cell proliferation method is reported in Fig. 3.3
(A & B). Data from both assays show that concentration of CTS and POL did not
exerted any effect on viability of CHO and Huh-7 cells. Cellular toxicity data of other
plants that did not show convincing potential against HCV is not presented. After the
toxicological analysis of plant extracts, the in vitro screening bioassays of plants
against HCV facilitate to explore the anti HCV agents from these phyto sources. In
case of serum inhibition assay, hepatic cell line was infected with positive serum
(high titre of HCV) of 3a genotype for screening of plants against whole virus. Crude
methanolic extracts of selected plants were screened against HCV infected liver cell
line and the results are displayed in Fig. 3.4. Only two methanolic extracts (CTS and
POL) exhibit potent and convincing anti HCV potential as shown in Fig. 3.5 (A&B).
The viral load was reduced to more than 60 % in case of POL while 57 % in case of
CTS which confirm that both possess appreciable antiviral potential. Both plant’s
methanolic extracts (POLM and CTSM) were suspended in water then further
fractionated with n-hexane (POLH and CTSH), ethyl acetate (POLE and CTSE),

124
chloroform (POLC and CTSC) and n-butanol (POLB and CTSB). These different
solvent fractions were screened for their anti HCV activity as well. Among these, only
the ethyl acetate fractions (CTSE & POLE) from both plants significantly reduced the
viral titer (68 % and 74 %) as presented in Fig. 3.5 (A&B).
For the purpose of in vitro protease inhibition, Huh-7 cells were transfected with
mammalian expression vector (PCR3.1/FLAGtag/HCV NS3) in the absence and
presence of selected plant extracts. Fig. 3.6 (A&B) and Fig. 3.7 (A&B) represents the
qualitative & quantitative inhibition of HCV NS3 protease by IFN, methanolic and
ethyl acetate extract of CTS and POL. Moreover in case POL, its inhibitory effect on
the protein expression level of NS3 transfected into Huh-7 cells was also confirmed
by Western blot. There was a dramatic reduction in the protein expression of the NS3
gene after treatment of POL extracts (Fig. 3.8). There were 60 % and 70 % inhibition
in Huh-7 cells treated POM and POE respectively. This response is likely to be
comparable to in vitro inhibition of HCV NS3-SP with ethanolic extract of Rhodiola
kirilowii (Chinese herb) and Camellia sinensis Epigallocatechin-3-gallate compound
(Chen et al., 2012; Zuo et al., 2007). NS3-mediated processing of protein junctions
among the non-structural proteins is indispensible for HCV replication and hence
recognized as striking target for antiviral agents’ development (Pause, et al., 2003).
Moreover, NS3 may also interfere with the functions host cell like protein kinase A
arbitrated signal transduction and inhibition of cell transformation (Borowski et al.,
1996; Sakamuro et al., 1995). Besides POL and CTS, a number of studies have
explored bioactive products from Galla Chinese, Rhodiola kirilowii, Lonicera
hypoglauca Miq. Swietenia macrophylla, Solanum nigrumand Camellia sinensis with
anti-NS3 protease activity (Duan et al., 2004; Wang et al., 2009; Zuo et al., 2007).
Previous reports have verified that Epicatechin and Epigallocatechin compounds from
rhizomes of the Chinese therapeutic herb have remarkable activity against NS3 serine
protease of HCV (Chen et al., 2012).
The phytochemical analysis of methanolic extracts of CTS and POL confirms the
presence of specific constituents in significant amounts as shown in table Table 3.1
and Table 3.2. Qualitative screening of methanolic extracts of CTS and POL pointed
toward the occurrence of various constituents as revealed in Table 3.1. CTS extract
was found to be rich with glycosides, steroids, saponin, coumarine and resins, but on

125
other hand, POL extract was rich in anthraquinones, fatty acids, flavonoids, tannin
and resins. The Preliminary phytochemical screening demonstrated that hexane
fractions were rich in non-polar compounds like steroids etc., while ethyl acetate
fractions sustained high concentration of flavonoids. Presence of these compounds
provided a foundation for identification of unknown but excellent therapeutic agents
and to structure those through spectral data.
As we know that antioxidants exhibit strong anti cancer potential and hepatocellular
carcinoma appear in 15 % cases of patients of HCV, which leads us for the
verification of antioxidant potential of POL and CTS extracts. Fig 3.9 to 3.11 contain
the data for antioxidant potential in term of total phenolic, flavonoids and total
carotenoids contents of biologically active plant extract fractions while Fig. 3.12-3.14
represent the free radical scavenging activity of extracts by DPPH, ABTS and FRAP
assays. In case of in vitro anti thrombotic potential, highest activity among extract
fractions was shown by CTSM and minimum by POLE while the overall order of
antithrombotic potential was CTSM > CTSE > POLM >POLE (Fig. 3.17). The
antibacterial activity of methnolic extracts of CTS and POL against Gram positive
(Bacillus subtilis, Styphylococcus aureus and Styphylococcus epidermidis) and Gram
negative (Pseudomonas aerugenosa, Escherichia coli and Salmonella typhi) strains of
bacteria was determined by disc diffusion method (Table 3.4). Percent inhibition with
respect to the positive control (Ciproflaxacin) indicated that both CTSM and POLM
exhibited more than 60 % inhibition against Styphylococcus epidermidis.
Carlluma tuberculata (CTS) is widely grown succulent perennial herb that is not only
eaten as raw famine food, but also cooked as vegetable in some area. It is considered
valuable due to its potential use against diabetes, cancer and inflammation. Prior to
our work, CTS and POL antiviral specifically anti HCV potential was not reported.
The most potent ethyl acetate fraction of CT was further utilized to isolate the
compounds through flash chromatography using silica gel 60. The structure of
steroidal glycosides was deduced by 1H,13C-NMR, DEPT, COSY and mass spectral
data. Five different compounds CT-EtOAc-01 i.e. ethyl(12-O-benzoyl-androstan-3-O-
(14)-β-D-cymaropyranosyl-(14)-β-Dcymaropyranosyl-(14)-β-D-
cymaropyranosyl) hexanoate, CT-EtOAc-02 i.e. Methyl (12-O-benzoyl-androstan-3-
O-(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl--(14)-β-D-

126
cymaropyranosyl) hexanoate, CT-EtOAc-03 i.e. Methyl (23-O-benzoyl-androst-6-en-
17-O-(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl--(14)-β-D-
cymaropyranosyl)hexanoate, CT-EtOAc-04 i.e. Phenyl (12-O-benzoyl-gonan-6-en-
5’’-O-(14)-β-D-cymaropyranosyl-(14)-β-D-cymaropyranosyl)oxalate and CT-
EtOAc-05 i.e. Trans-p-ferulylalcohol-4-O-(6-(2-methyl-3-hydroxypropionyl)
glucopyranoside were isolated, purified and characterized. which have potential to be
further utilized for docking studies with the structure of NS3-SP to confer further
mechanism.
The potency of bioactive compounds from CTS and POL is very important and well
proven with an additional edge of being wildly grown natural source. The novel
antiviral activity of compounds from these plants presents an attractive lead for
natural chemical entities for the development of potential anti-HCV agents.
Additionally, if natural molecules cannot be replaced in the present anti-HCV
treatment, these could be used in combination or supplementation to interferon thus
reducing overall treatment cost. The above data also proposes that the therapeutic
importance of extracts may lead to an alternative, more potent oral treatment for
chronic HCV.
4.0 Future Prospects
Findings of the present project “Bioassay directed characterization of selected
indigenous medicinal plants for their anti-HCV potential” indicates that both plant

127
species having code CTS and POL are very useful and non toxic medicinal plants that
have strong therapeutic potential against HCV specifically its protein NS3. In short, it
can be speculated that therapeutic spur of antiviral plant extracts or purified fractions
alone or in mixture with IFN could offer an effective therapy alternative against HCV
whole virus and it NS3 protease. Structural analysis of correspondent antiviral active
fraction might prove supportive for identification of new antiviral compounds.
As both species fall mostly in the category of uncultivated and uncommon vegetables,
so these offer a very inexpensive source of medication, especially to poor rural
peoples who live in such vicinity where these grow. There is dire need of hour to
grow these plants commercially in order to attain the benefit from their medicinal
potential and to fulfill the global demand.
Although very few efforts has been made to explore biologically active constituent of
these flora, but these are thoroughly known for their therapeutic potential against
infectious and metabolic disorders. Focus was only given to isolate compound and
identify them but not explored the effect of these compounds at molecular level and
their possible mechanism of action.
Docking studies are considered very important in rational drug designing as it decide
the preferred orientation of drug or compound toward the target (NS3 serine
protease). So such finding would strengthen our goal to develop target based drugs
and then their examination on in vivo animal model. Docking of antiviral compounds
will determine the preferred orientation of antiviral compounds towards NS3 protease.
Docking is important in rational drug designing. Designing structures by docking
marvel ligands to create potent active sites and then compare docked compounds to
known active compounds is useful for designing novel candidate drugs.
The climate of the Soon-valley is subtropical with unique geological formation due to
salt as well as other minerals in the area. In the local vegetation, at least work has
been conducted with respect to medicinal point of view. Intensive efforts needed to
devote toward the discovery of innovative HCV antiviral agents. Owing to the
marvellous therapeutic potential of these extracts there is a possibility to develop such
drugs from these. There are innumerable plants in this valley waiting to be explored

128
for their therapeutic applications against genetically and functionally diverse virus
families.
5.0 References
Abdel-Sattar, E., Ahmed, A. A., Hegazy, M.-E. F., Farag, M. A., & Al-Yahya, M. A.-A. (2007). Acylated pregnane glycosides from Caralluma russeliana. Phytochemistry, 68(10), 1459-1463.
Abdel-Sattar, E., Harraz, F. M., Al-Ansari, S. M., El-Mekkawy, S., Ichino, C., Kiyohara, H., et al. (2009). Antiplasmodial and antitrypanosomal activity of plants from the Kingdom of Saudi Arabia. Journal of natural medicines, 63(2), 232-239.
Abdel-Sattar, E., Harraz, F. M., Al-Ansari, S. M. A., El-Mekkawy, S., Ichino, C., Kiyohara, H., et al. (2008). Acylated pregnane glycosides from Caralluma tuberculata and their antiparasitic activity. Phytochemistry, 69(11), 2180-2186.
Abdel-Sattar, E., Harraz, F. M., Ghareib, S. A., Elberry, A. A., Gabr, S., & Suliaman, M. I. (2011). Antihyperglycaemic and hypolipidaemic effects of the methanolic extract of Caralluma tuberculata in streptozotocin-induced diabetic rats. Natural product research, 25(12), 1171-1179.
Ahmad, I., Aqil, F., & Owais, M. (2006). Modern phytomedicine: Turning medicinal plants into Drugs: John Wiley & Sons.
Ahmad, K., Khan, Z. I., Ashraf, M., Hussain, M., Ibrahim, M., & Valeem, E. E. (2008). Status of plant diversity at Kufri (Soon valley) punjab, pakistan and prevailing threats therein. Pak. J. Bot, 40(3), 993-997.
Ahmad, K., Khan, Z. I., Ashraf, M., Hussain, M., Ibrahim, M., & Valeem, E. E. (2008). Status of plant diversity at Kufri (Soone valley) Punjab, Pakistan and prevailing threats therein. Pak. J. Bot, 40(3), 993-997.
Ahmad, M., Qureshi, R., Arshad, M., Khan, M. A., & Zafar, M. (2009). Traditional herbal remedies used for the treatment of diabetes from district Attock (Pakistan). Pakistan Journal of Botany, 41(6), 2777-2782.
Ahmad, V. U., Usmanghani, K., & Rizwani, G. H. (1988). New pregnane glycosides from Caralluma tuberculata. Journal of natural products, 51(6), 1092-1097.

129
Ahmed, M., Qureshi, S., Al-Bekairi, A., Shah, A., Rao, R., & Qazi, N. (1993). Anti-inflammatory activity of Caralluma tuberculata alcoholic extract. Fitoterapia, 64(4).
Akhtar, M. S., Khan, Q. M., & Khaliq, T. Effects of Portulaca oleracae (Kulfa) and Taraxacum officinale (Dhudhal) in normoglycaemic and alloxan-treated hyperglycaemic rabbits.
Al-Bekairi, A., Qureshi, S., Ahmed, M., Qazi, N., Khan, Z., & Shah, A. (1992). Effect of Caralluma tuberculata on the cytological and biochemical changes induced by cyclophosphamide in mice. Food and chemical toxicology, 30(8), 719-722.
Al-Massarani, S. M., Bertrand, S., Nievergelt, A., El-Shafae, A. M., Al-Howiriny, T. A., Al-Musayeib, N. M., et al. (2012). Acylated pregnane glycosides from< i> Caralluma sinaica</i>. Phytochemistry, 79, 129-140.
Albuquerque, U. P. d., & Hanazaki, N. (2006). As pesquisas etnodirigidas na descoberta de novos fármacos de interesse médico e farmacêutico: fragilidades e pespectivas. Rev Bras Farmacogn, 16(sSupl).
Alharbi, M. M., Qureshi, S., Raza, M., Ahmed, M. M., Afzal, M., & Shah, A. H. (1994). Evaluation of Caralluma tuberculata Pretreatment for the Protection of Rat Gastric Mucosa Against Toxic Damage. Toxicology and applied pharmacology, 128(1), 1-8.
Ali, H., Sannai, J., Sher, H., & Rashid, A. (2011). Ethnobotanical profile of some plant resources in Malam Jabba valley of Swat, Pakistan. Journal of Medicinal Plants Research, 5(18), 4676-4687.
Ali, H. A., Hussein, M. A., & Mohammed, W. E. (2014). Comparative Effects of Purslane seed oil (PSO) and 5-Flourourasil on Ehrlich ascites carcinoma (EAC) in female albino mice. [Research Article]. International Journal of Pharma Sciences, 4(1), 424-430.
Alter, M. J. (2007). Epidemiology of hepatitis C virus infection. World J Gastroenterol, 13(17), 2436-2441.
Asabe, S. I., Tanji, Y., Satoh, S., Kaneko, T., Kimura, K., & Shimotohno, K. (1997). The N-terminal region of hepatitis C virus-encoded NS5A is important for NS4A-dependent phosphorylation. J Virol, 71(1), 790-796.
Ashfaq, U. A., Masoud, M. S., Nawaz, Z., & Riazuddin, S. (2011). Glycyrrhizin as antiviral agent against Hepatitis C Virus. J Transl Med, 9(112), 10.1186.
Ayoola, G. A., Ipav, S. S., Sofidiya, M. O., Adepoju-Bello, A. A., Coker, H. A., & Odugbemi, T. O. (2008). Phytochemical screening and free radical scavenging activities of the fruits and leaves of Allanblackia floribunda Oliv (Guttiferae). International journal of health research, 1(2), 87-93.
Bachmetov, L., Gal‐Tanamy, M., Shapira, A., Vorobeychik, M., Giterman‐Galam, T., Sathiyamoorthy, P., et al. (2012). Suppression of hepatitis C virus by the flavonoid quercetin is mediated by inhibition of NS3 protease activity. Journal of viral hepatitis, 19(2), e81-e88.
Banerjee, G., & Mukherjee, A. (2002). Biological activity of a common weed-Portulaca oleracea L. II. Antifungal activity. Acta Botanica Hungarica, 44(3), 205-208.
Barbour, E. K., Al Sharif, M., Sagherian, V. K., Habre, A. N., Talhouk, R. S., & Talhouk, S. N. (2004). Screening of selected indigenous plants of Lebanon for antimicrobial activity. Journal of ethnopharmacology, 93(1), 1-7.

130
Bartenschlager, R. (2002). Hepatitis C virus replicons: potential role for drug development. Nat Rev Drug Discov, 1(11), 911-916.
Bartenschlager, R., Ahlborn-Laake, L., Mous, J., & Jacobsen, H. (1993). Nonstructural protein 3 of the hepatitis C virus encodes a serine-type proteinase required for cleavage at the NS3/4 and NS4/5 junctions. Journal of virology, 67(7), 3835-3844.
Bartenschlager, R., Ahlborn-Laake, L., Mous, J., & Jacobsen, H. (1994). Kinetic and structural analyses of hepatitis C virus polyprotein processing. J Virol, 68(8), 5045-5055.
Barton, M. D. (2000). Antibiotic use in animal feed and its impact on human healt. Nutrition research reviews, 13(02), 279-299.
Bartosch, B., Bukh, J., Meunier, J. C., Granier, C., Engle, R. E., Blackwelder, W. C., et al. (2003). In vitro assay for neutralizing antibody to hepatitis C virus: evidence for broadly conserved neutralization epitopes. Proc Natl Acad Sci U S A, 100(24), 14199-14204.
Bauer, A., Kirby, W., Sherris, J. C., turck, & Turck, M. (1966). Antibiotic susceptibility testing by a standardized single disk method. American journal of clinical pathology, 45(4), 493.
Becker, R., Silvi, J., Ma Fat, D., L'Hours, A., & Laurenti, R. (2006). A method for deriving leading causes of death. Bulletin of the World Health Organization, 84(4), 297-304.
Benzie, I. F., & Strain, J. (1999). Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods in enzymology, 299, 15-27.
Berlin, E. A., & Berlin, B. (2005). Some field methods in medical ethnobiology. Field Methods, 17(3), 235-268.
Blight, K. J., Kolykhalov, A. A., & Rice, C. M. (2000). Efficient initiation of HCV RNA replication in cell culture. Science, 290(5498), 1972-1974.
Borowski, P., Heiland, M., Oehlmann, K., Becker, B., Kornetzky, L., Feucht, H., et al. (1996). Non-structural protein 3 of hepatitis C virus inhibits phosphorylation mediated by cAMP-dependent protein kinase. Eur J Biochem, 237(3), 611-618.
Borowski, P., Oehlmann, K., Heiland, M., & Laufs, R. (1997). Nonstructural protein 3 of hepatitis C virus blocks the distribution of the free catalytic subunit of cyclic AMP-dependent protein kinase. J Virol, 71(4), 2838-2843.
Braca, A., Bader, A., Morelli, I., Scarpato, R., Turchi, G., Pizza, C., et al. (2002). New pregnane glycosides from Caralluma negevensis. Tetrahedron, 58(29), 5837-5848.
Brown, C. J., Hendrich, B. D., Rupert, J. L., Lafreniere, R. G., Xing, Y., Lawrence, J., et al. (1992). The human XIST gene: Analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. Cell, 71(3), 527-542.
Bukh, J., Purcell, R. H., & Miller, R. H. (1994). Sequence analysis of the core gene of 14 hepatitis C virus genotypes. Proc Natl Acad Sci U S A, 91(17), 8239-8243.

131
Burke, R., Diamondstone, B., Velapoldi, R., & Menis, O. (1974). Mechanisms of the Liebermann-Burchard and Zak color reactions for cholesterol. Clinical chemistry, 20(7), 794-801.
Burkill H.M. (1997). The useful plants of West Tropical Africa (Vol. 4): The Whitefriars Press Limited, Tonbridge, Kent TN9 IQR, Great Britain.
Cai, Z., Zhang, C., Chang, K. S., Jiang, J., Ahn, B. C., Wakita, T., et al. (2005). Robust production of infectious hepatitis C virus (HCV) from stably HCV cDNA-transfected human hepatoma cells. J Virol, 79(22), 13963-13973.
Calland, N., Dubuisson, J., Rouillé, Y., & Séron, K. (2012). Hepatitis C virus and natural compounds: a new antiviral approach? Viruses, 4(10), 2197-2217.
Cantwell, M. I., & Reid, M. S. (1993). Postharvest physiology and handling of fresh culinary herbs. Journal of Herbs, Spices & Medicinal Plants, 1(3), 93-127.
Castro, V., Garcia, C., Gonzalez, A., Hernandez, R., & Suarez, E. (1980). A 3, 4-seco-triterpene from Caralluma buchardii. Phytochemistry, 19(10), 2210-2212.
Chan, K., Islam, M., Kamil, M., Radhakrishnan, R., Zakaria, M., Habibullah, M., et al. (2000). The analgesic and anti-inflammatory effects of Portulaca oleracea L. subsp. sativa (Haw.) Celak. Journal of ethnopharmacology, 73(3), 445-451.
Chan, K., Islam, M. W., Kamil, M., Radhakrishnan, R., Zakaria, M. N., Habibullah, M., et al. (2000). The analgesic and anti-inflammatory effects of Portulaca oleracea L. subsp. Sativa (Haw.) Celak. J Ethnopharmacol, 73(3), 445-451.
Chen, C., Qiu, H., Gong, J., Liu, Q., Xiao, H., Chen, X. W., et al. (2012). (-)-Epigallocatechin-3-gallate inhibits the replication cycle of hepatitis C virus. Arch Virol, 157(7), 1301-1312.
Chen, J., Chen, L., An, Z., Shi, S., & Zhan, Y. (2002). Non-technical causes of fakes existing in Chinese medicinal material markets. Zhong yao cai= Zhongyaocai= Journal of Chinese medicinal materials, 25(7), 516-519.
Chen, T., Wang, J., Li, Y., Shen, J., Zhao, T., & Zhang, H. (2010). Sulfated modification and cytotoxicity of Portulaca oleracea L. polysaccharides. Glycoconjugate journal, 27(6), 635-642.
Cos, P., Vlietinck, A. J., Berghe, D. V., & Maes, L. (2006). Anti-infective potential of natural products: How to develop a stronger in vitro ‘proof-of-concept’. Journal of ethnopharmacology, 106(3), 290-302.
Cragg, G. M., Newman, D. J., & Snader, K. M. (1997). Natural products in drug discovery and development. Journal of natural products, 60(1), 52-60.
Dawidar, A. M., Mubarak, A. Y., Abdel-Mogib, M., & Abdelsattar, E. (2012). Phytochemical investigation of Caralluma wissmannii Research Journal of Pharmaceutical, Biological and Chemical Sciences, 3(2), 884-892.
De Francesco, R., & Migliaccio, G. (2005). Challenges and successes in developing new therapies for hepatitis C. Nature, 436(7053), 953-960.
De Francesco, R., Tomei, L., Altamura, S., Summa, V., & Migliaccio, G. (2003). Approaching a new era for hepatitis C virus therapy: inhibitors of the NS3-4A serine protease and the NS5B RNA-dependent RNA polymerase. Antiviral Res, 58(1), 1-16.
De Leo, M., De Tommasi, N., Sanogo, R., Marzocco, S., Pizza, C., Morelli, I., et al. (2005). New pregnane glycosides from Caralluma dalzielii. Steroids, 70(9), 573-585.

132
De Silva, T. (1997). Industrial utilization of medicinal plants in developing countries. Medicinal plants for forest conservation and health care. FAO, Rome, 34-44.
Deepak, D., Khare, A., & Khare, M. P. (1989). Plant pregnanes. Phytochemistry, 28(12), 3255-3263.
Deepak, D., Srivastav, S., & Khare, A. (1997). Pregnane glycosides Fortschritte der Chemie organischer Naturstoffe/Progress in the Chemistry of Organic Natural Products (pp. 169-325): Springer.
Dong, C.-X., Hayashi, K., Lee, J.-B., & Hayashi, T. (2010). Characterization of structures and antiviral effects of polysaccharides from Portulaca oleracea L. Chemical and Pharmaceutical Bulletin, 58(4), 507-510.
Dong, L., Wang, W., Yue, Y., & Li, M. (2005). [Effects of flavones extracted from Portulaca oleracea on ability of hypoxia tolerance in mice and its mechanism]. Zhong xi yi jie he xue bao= Journal of Chinese integrative medicine, 3(6), 450-454.
Duan, D., Li, Z., Luo, H., Zhang, W., Chen, L., & Xu, X. (2004). Antiviral compounds from traditional Chinese medicines Galla Chinese as inhibitors of HCV NS3 protease. Bioorganic & medicinal chemistry letters, 14(24), 6041-6044.
Duraipandiyan, V., Ayyanar, M., & Ignacimuthu, S. (2006). Antimicrobial activity of some ethnomedicinal plants used by Paliyar tribe from Tamil Nadu, India. BMC Complementary and Alternative Medicine, 6(1), 35.
Ebrahimzadeh, M., Nabavi, S., & Nabavi, S. (2009). Essential oil Composition and Antioxidant Activity of Pterocaryafivaxinifolia. Pakistan Journal ofBiological Sciences, 12(13), 957-963.
Edeoga, H., Okwu, D., & Mbaebie, B. (2005). Phytochemical constituents of some Nigerian medicinal plants. African journal of biotechnology, 4(7), 685-688.
Eduardo, H. R., Estela, R., Luciana, G., & Margutti, L. (1995). Edible Weeds: A Scarcely Used Resource. Ecological Society of America, 76(3), 163-166.
Egger, D., Wolk, B., Gosert, R., Bianchi, L., Blum, H. E., Moradpour, D., et al. (2002). Expression of hepatitis C virus proteins induces distinct membrane alterations including a candidate viral replication complex. J Virol, 76(12), 5974-5984.
Elazar, M., Cheong, K. H., Liu, P., Greenberg, H. B., Rice, C. M., & Glenn, J. S. (2003). Amphipathic helix-dependent localization of NS5A mediates hepatitis C virus RNA replication. J Virol, 77(10), 6055-6061.
Elisabetsky, E., & Shanley, P. (1994). Ethnopharmacology in the Brazilian Amazon. Pharmacology & therapeutics, 64(2), 201-214.
Elkhayat, E. S., Ibrahim, S. R., & Aziz, M. A. (2008). Portulene, a new diterpene from Portulaca oleracea L. Journal of Asian natural products research, 10(11), 1039-1043.
Endress, M. E., & Bruyns, P. V. (2000). A revised classification of the Apocynaceae sl. The Botanical Review, 66(1), 1-56.
Enomoto, N., Sakuma, I., Asahina, Y., Kurosaki, M., Murakami, T., Yamamoto, C., et al. (1995). Comparison of full-length sequences of interferon-sensitive and resistant hepatitis C virus 1b. Sensitivity to interferon is conferred by amino acid substitutions in the NS5A region. J Clin Invest, 96(1), 224-230.
Ernst, E., Hung, S., & Clement, Y. (2011). NCCAM-funded RCTs of herbal medicines: An important critical assessment. Perfusion, 24(3), 89-102.

133
Farci, P., Alter, H. J., Wong, D. C., Miller, R. H., Govindarajan, S., Engle, R., et al. (1994). Prevention of hepatitis C virus infection in chimpanzees after antibody-mediated in vitro neutralization. Proceedings of the National Academy of Sciences, 91(16), 7792-7796.
Farnsworth, N. R. (1966). Biological and phytochemical screening of plants. Journal of pharmaceutical Sciences, 55(3), 225-276.
Fennell, C., Lindsey, K., McGaw, L., Sparg, S., Stafford, G., Elgorashi, E., et al. (2004). Assessing African medicinal plants for efficacy and safety: pharmacological screening and toxicology. Journal of ethnopharmacology, 94(2), 205-217.
Forton, D. M., Thomas, H. C., Murphy, C. A., Allsop, J. M., Foster, G. R., Main, J., et al. (2002). Hepatitis C and cognitive impairment in a cohort of patients with mild liver disease. Hepatology, 35(2), 433-439.
Foy, E., Li, K., Wang, C., Sumpter, R., Ikeda, M., Lemon, S. M., et al. (2003). Regulation of interferon regulatory factor-3 by the hepatitis C virus serine protease. Science, 300(5622), 1145-1148.
Friebe, P., & Bartenschlager, R. (2002). Genetic analysis of sequences in the 3′ nontranslated region of hepatitis C virus that are important for RNA replication. Journal of virology, 76(11), 5326-5338.
Gale, M. J., Jr., Korth, M. J., Tang, N. M., Tan, S. L., Hopkins, D. A., Dever, T. E., et al. (1997). Evidence that hepatitis C virus resistance to interferon is mediated through repression of the PKR protein kinase by the nonstructural 5A protein. Virology, 230(2), 217-227.
Ghabril, M., Chalasani, N., & Björnsson, E. (2010). Drug-induced liver injury: a clinical update. Current opinion in gastroenterology, 26(3), 222.
Gibbs, R. (1974). Chemotaxonomy of Flowering plant Morinda Citrifolia L. Queen’s University Press. Montreal and London.
Gonzalez, O., Fontanes, V., Raychaudhuri, S., Loo, R., Loo, J., Arumugaswami, V., et al. (2009). The heat shock protein inhibitor Quercetin attenuates hepatitis C virus production. Hepatology, 50(6), 1756-1764.
Goodman, S. M., & Ghafoor, A. (1992). The Ethnobotany of Southern Balochistan, Pakistan, with particular reference to medicinal plants: [Chicago, IL] : Field Museum of Natural History.
Gowri, S., & Chinnaswamy, P. (2011). Evaluation of invitro antimutagenic activity of Caralluma adscendens Roxb. in bacterial reverse mutation assay. J Nat Prod Plant Resour, 1, 27-34.
Grakoui, A., McCourt, D. W., Wychowski, C., Feinstone, S. M., & Rice, C. M. (1993a). Characterization of the hepatitis C virus-encoded serine proteinase: determination of proteinase-dependent polyprotein cleavage sites. J Virol, 67(5), 2832-2843.
Grakoui, A., McCourt, D. W., Wychowski, C., Feinstone, S. M., & Rice, C. M. (1993b). A second hepatitis C virus-encoded proteinase. Proc Natl Acad Sci U S A, 90(22), 10583-10587.
Grakoui, A., Wychowski, C., Lin, C., Feinstone, S. M., & Rice, C. M. (1993). Expression and identification of hepatitis C virus polyprotein cleavage products. Journal of virology, 67(3), 1385-1395.
Grieve, C., & Suarez, D. (1997). Purslane (Portulaca oleracea L.): a halophytic crop for drainage water reuse systems. Plant and Soil, 192(2), 277-283.

134
Griffin, S. D., Beales, L. P., Clarke, D. S., Worsfold, O., Evans, S. D., Jaeger, J., et al. (2003). The p7 protein of hepatitis C virus forms an ion channel that is blocked by the antiviral drug, Amantadine. FEBS Lett, 535(1-3), 34-38.
Habibuddin, M., Daghriri, H. A., Humaira, T., Qahtani, M. S. A., & Hefzi, A. A. H. (2008). Antidiabetic effect of alcoholic extract of< i> Caralluma sinaica</i> L. on streptozotocin-induced diabetic rabbits. Journal of ethnopharmacology, 117(2), 215-220.
Hahm, B., Han, D. S., Back, S. H., Song, O.-K., Cho, M.-J., Kim, C.-J., et al. (1995). NS3-4A of hepatitis C virus is a chymotrypsin-like protease. Journal of virology, 69(4), 2534-2539.
Haid, S., Novodomská, A., Gentzsch, J., Grethe, C., Geuenich, S., Bankwitz, D., et al. (2012). A plant-derived flavonoid inhibits entry of all HCV genotypes into human hepatocytes. Gastroenterology, 143(1), 213-222. e215.
Halim, A. F., & Khalil, A. T. (1996). Pregnane glycosides from Caralluma retrospiciens. Phytochemistry, 42(4), 1135-1139.
Hamayun, M. (2003). Ethnobotanical studies of some useful shrubs and trees of District Buner, NWFP, Pakistan. Ethnobotanical Leaflets, 2003(1), 12.
Hamid, S., Umar, M., Alam, A., Siddiqui, A., Qureshi, H., & Butt, J. (2004). PSG consensus statement on management of hepatitis C virus infection-2003. Journal-Pakistan Medical Association, 54(3), 146-149.
Harborne, J. B. (1984). Phytochemical methods (Vol. 278): Springer. Harvey, A. (2000). Strategies for discovering drugs from previously unexplored
natural products. Drug discovery today, 5(7), 294-300. Heller, T., Saito, S., Auerbach, J., Williams, T., Moreen, T. R., Jazwinski, A., et al.
(2005). An in vitro model of hepatitis C virion production. Proceedings of the National Academy of Sciences of the United States of America, 102(7), 2579-2583.
Honda, M., Brown, E., & Lemon, S. (1996). Stability of a stem-loop involving the initiator AUG controls the efficiency of internal initiation of translation on hepatitis C virus RNA. Rna, 2(10), 955.
Hongxing, Z., Nancai, Y., Guofu, H., Jianbo, S., Yanxia, W., Hanju, H., et al. (2007). Neuroprotective effects of purslane herb aquenous extracts against D-galactose induced neurotoxicity. Chemico-biological interactions, 170(3), 145-152.
Hope, R. G., Murphy, D. J., & McLauchlan, J. (2002). The domains required to direct core proteins of hepatitis C virus and GB virus-B to lipid droplets share common features with plant oleosin proteins. J Biol Chem, 277(6), 4261-4270.
Horwitz, W. (2000). Official Methods of Analytical Chemistry. AOAC International Suite 500, 481 North Frederick Avenue Gaithers Burg, Maryland USA.
Hosu, A., Cristea, V.-M., & Cimpoiu, C. (2014). Analysis of total phenolic, flavonoids, anthocyanins and tannins content in Romanian red wines: Prediction of antioxidant activities and classification of wines using artificial neural networks. Food chemistry, 150, 113-118.
Houghton, M., Weiner, A., Han, J., Kuo, G., & Choo, Q. L. (1991). Molecular biology of the hepatitis C viruses: implications for diagnosis, development and control of viral disease. Hepatology, 14(2), 381-388.

135
Hsu, M., Zhang, J., Flint, M., Logvinoff, C., Cheng-Mayer, C., Rice, C. M., et al. (2003). Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proc Natl Acad Sci U S A, 100(12), 7271-7276.
Hugle, T., Fehrmann, F., Bieck, E., Kohara, M., Krausslich, H. G., Rice, C. M., et al. (2001). The hepatitis C virus nonstructural protein 4B is an integral endoplasmic reticulum membrane protein. Virology, 284(1), 70-81.
Huie, C. W. (2002). A review of modern sample-preparation techniques for the extraction and analysis of medicinal plants. Analytical and bioanalytical chemistry, 373(1-2), 23-30.
Idrees, S., Ashfaq, U. A., & Khaliq, S. (2013). HCV Envelope protein 2 sequence comparison of Pakistani isolate and In-silico prediction of conserved epitopes for vaccine development. J Transl Med, 11, 105.
Imai, S., Shiraishi, A., Gamo, K., Watanabe, I., Okuhata, H., Miyasaka, H., et al. (2007). Removal of phenolic endocrine disruptors by Portulaca oleracea. Journal of bioscience and bioengineering, 103(5), 420-426.
Islam, M., Zakaria, M., Radhakrishnan, R., Habibullah, M., & Chan, K. (1998). Evaluation of analgesic activity of the aerial parts of Portulaca oleracea v. sativa and its comparison with two related species. Journal of Pharmacy and Pharmacology, 50(S9), 226-226.
Ito, T., Mukaigawa, J., Zuo, J., Hirabayashi, Y., Mitamura, K., & Yasui, K. (1996). Cultivation of hepatitis C virus in primary hepatocyte culture from patients with chronic hepatitis C results in release of high titre infectious virus. J Gen Virol, 77 ( Pt 5), 1043-1054.
Iyengar, M. A. (1995). Study of Crude Drugs (8th ed. ed.). Manipal, India: Manipal Power Press.
Jahan, S., Khaliq, S., Ijaz, B., Ahmad, W., & Hassan, S. (2011). Role of HCV Core gene of genotype 1a and 3a and host gene Cox-2 in HCV-induced pathogenesis. Virology journal, 8, 155.
Jingrong, L. T. H., & Ramesh, P. (2009). Compounds of Purslane extracts and effects of anti-kinetic fatigue. Journal of Medicinal Plants Research, 3(7), 506-510.
Jordan, S. A., Cunningham, D. G., & Marles, R. J. (2010). Assessment of herbal medicinal products: challenges, and opportunities to increase the knowledge base for safety assessment. Toxicology and applied pharmacology, 243(2), 198-216.
Kalpna, R., Mital, K., & Sumitra, C. (2011). Vegetable and fruit peels as a novel source of antioxidants. Journal of Medicinal Plants Research, 5(1), 63-71.
Kamalakkannan, S., Rajendran, R., Venkatesh, R. V., Clayton, P., & Akbarsha, M. A. (2010). Antiobesogenic and antiatherosclerotic properties of Caralluma fimbriata extract. Journal of nutrition and metabolism, 2010.
Karimi, G., Hosseinzadeh, H., & Ettehad, N. (2004). Evaluation of the gastric antiulcerogenic effects of Portulaca oleracea L. extracts in mice. Phytotherapy Research, 18(6), 484-487.
Karimi, G., Khoei, A., Omidi, A., Kalantari, M., Babaei, J., Taghiabadi, E., et al. (2010). Protective effect of aqueous and ethanolic extracts of Portulaca oleracea against cisplatin induced nephrotoxicity. Iranian Journal of Basic Medical Sciences.

136
Karthik, P., Samydurai, P., Subbaiyan, B., Thangapandian, V., & Binu, T. (2013). In vitro propagation of a rare succulent medicinal plant Caralluma diffusa (Wight) NE Br. Research in Plant Biology, 3(1), 8-17.
Karuppusamy, S. (2007). Medicinal plants used by Paliyan tribes of Sirumalai hills of Southern India. Natural Product Radiance, 6(5), 436-442.
Khan, S. W., & Khatoon, S. (2008). Ethnobotanical studies on some useful herbs of Haramosh and Bugrote valleys in Gilgit, northern areas of Pakistan. Pakistan Journal of Botany, 40(1), 43.
Kim, J. L., Morgenstern, K. A., Lin, C., Fox, T., Dwyer, M. D., Landro, J. A., et al. (1996). Crystal structure of the hepatitis C virus NS3 protease domain complexed with a synthetic NS4A cofactor peptide. Cell, 87(2), 343-355.
Kishore, M., Surendrababu, K., Hanumantharao, Y., Naga hima bindu, G., & Janardhan, M. (2010). Chemical examination of medicinal plant “Caralluma umbellate” (asclepiadaceae) roots. International Journal of Applied Biology and Pharmaceutical Technology, 1(2), 545-549.
Kokubun, T., Kite, G. C., Veitch, N. C., & Simmonds, M. (2012). Amides and an alkaloid from Portulaca oleracea. Natural product communications, 7(8), 1047-1050.
Kolykhalov, A. A., Feinstone, S. M., & Rice, C. M. (1996). Identification of a highly conserved sequence element at the 3'terminus of hepatitis C virus genome RNA. Journal of virology, 70(6), 3363-3371.
Kolykhalov, A. A., Mihalik, K., Feinstone, S. M., & Rice, C. M. (2000). Hepatitis C virus-encoded enzymatic activities and conserved RNA elements in the 3' nontranslated region are essential for virus replication in vivo. Journal of virology, 74(4), 2046-2051.
Kumar, A. S., Kavimani, S., & Jayaveera, K. (2011). A review on medicinal Plants with potential antidiabetic activity. International journal of phytopharmacology, 2(2), 53-60.
Kumar, S., Sharma, U., Sharma, A., & Pandey, A. (2012). Protective efficacy of Solanum xanthocarpum root extracts against free radical damage: phytochemical analysis and antioxidant effect. Cellular and Molecular Biology, 58(1), 171-178.
Kunert, O., Simic, N., Ravinder, E., Adavi Rao, B. V., Ravi Kumar, B., Michael Alex, R., et al. (2009). Steroidal glycosides from< i> Caralluma umbellata</i>. Phytochemistry Letters, 2(3), 134-138.
Lans, C. (2006). Ethnomedicines used in Trinidad and Tobago for urinary problem and Diabetes mellitus. J Ethnobiol Ethnomedicine, 13, 45-51.
Lawrence, R., & Choudhary, S. (2004). Caralluma fimbriata in the Treatment of Obesity. Paper presented at the 12th Annual World Congress of Anti-Aging Medicine.
Lee, K. H. (2000). Research and future trends in the pharmaceutical development of medicinal herbs from Chinese medicine. Public Health Nutr, 3(4A), 515-522.
Lerat, H., Honda, M., Beard, M. R., Loesch, K., Sun, J., Yang, Y., et al. (2002). Steatosis and liver cancer in transgenic mice expressing the structural and nonstructural proteins of hepatitis C virus. Gastroenterology, 122(2), 352-365.

137
Lesburg, C. A., Cable, M. B., Ferrari, E., Hong, Z., Mannarino, A. F., & Weber, P. C. (1999). Crystal structure of the RNA-dependent RNA polymerase from hepatitis C virus reveals a fully encircled active site. Nat Struct Biol, 6(10), 937-943.
Li, F., Li, Q., Gao, D., Peng, Y., & Feng, C. (2009). Preparation and antidiabetic activity of polysaccharide from Portulaca oleracea L. African Journal of Biotechnology, 8(4).
Li, K., Foy, E., Ferreon, J. C., Nakamura, M., Ferreon, A. C., Ikeda, M., et al. (2005). Immune evasion by hepatitis C virus NS3/4A protease-mediated cleavage of the Toll-like receptor 3 adaptor protein TRIF. Proceedings of the National Academy of Sciences of the United States of America, 102(8), 2992-2997.
Lichtenthaler, H. K., & Buschmann, C. (2001). Chlorophylls and Carotenoids: Measurement and Characterization by UV‐VIS Spectroscopy. Current protocols in food analytical chemistry.
Liede-Schumann, S., Rapini, A., Goyder, D. J., Chase, M. W., & Plunkett, G. M. (2005). Phylogenetics of the new world subtribes of Asclepiadeae (Apocynaceae-Asclepiadoideae): Metastelmatinae, Oxypetalinae, and Gonolobinae. Systematic Botany, 30(1), 184-195.
Lin, C., Wu, J. W., Hsiao, K., & Su, M. S. (1997). The hepatitis C virus NS4A protein: interactions with the NS4B and NS5A proteins. J Virol, 71(9), 6465-6471.
Lindenbach, B. D., & Rice, C. M. (2005). Unravelling hepatitis C virus replication from genome to function. Nature, 436(7053), 933-938.
Liu, L., Howe, P., Zhou, Y.-F., Xu, Z.-Q., Hocart, C., & Zhang, R. (2000). Fatty acids and β-carotene in Australian purslane Portulaca oleracea varieties. Journal of Chromatography A, 893(1), 207-213.
Lohmann, V., Hoffmann, S., Herian, U., Penin, F., & Bartenschlager, R. (2003). Viral and cellular determinants of hepatitis C virus RNA replication in cell culture. J Virol, 77(5), 3007-3019.
Lohmann, V., Korner, F., Koch, J., Herian, U., Theilmann, L., & Bartenschlager, R. (1999). Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science, 285(5424), 110-113.
Lorenz, I. C., Marcotrigiano, J., Dentzer, T. G., & Rice, C. M. (2006). Structure of the catalytic domain of the hepatitis C virus NS2-3 protease. Nature, 442(7104), 831-835.
Love, R. A., Parge, H. E., Wickersham, J. A., Hostomsky, Z., Habuka, N., Moomaw, E. W., et al. (1996). The crystal structure of hepatitis C virus NS3 proteinase reveals a trypsin-like fold and a structural zinc binding site. Cell, 87(2), 331-342.
Love, R. A., Parge, H. E., Wickersham, J. A., Hostomsky, Z., Habuka, N., Moomaw, E. W., et al. (1996). The crystal structure of hepatitis C virus NS3 proteinase reveals a trypsin-like fold and a structural zinc binding site. Cell, 87(2), 331-342.
Lundin, M., Lindstrom, H., Gronwall, C., & Persson, M. A. (2006). Dual topology of the processed hepatitis C virus protein NS4B is influenced by the NS5A protein. J Gen Virol, 87(Pt 11), 3263-3272.

138
Macdonald, A., Crowder, K., Street, A., McCormick, C., & Harris, M. (2004). The hepatitis C virus NS5A protein binds to members of the Src family of tyrosine kinases and regulates kinase activity. J Gen Virol, 85(Pt 3), 721-729.
Madhuri, V., Amrutha.V.A., & Murthy,K.S.R. (2011). Biochemical study of Caralluma species to understand species homology. International Journal of Applied Biology and Pharmaceutical Technology, 2(1), 139-144.
Mahmood, A., & Tabassum, A. (2011). Ethnomedicinal survey of plants from District Sialkot. Pakistan. J. Appl. Pharm, 2(03), 212-220.
Mahmood, T., Muhammad, S., & Shinwari, Z. K. (2010). Molecular and morphological characterization of Caralluma species. Pak. J. Bot, 42(2), 1163-1171.
Malek, F., Boskabady, M., Borushaki, M., & Tohidi, M. (2004). Bronchodilatory effect of Portulaca oleracea in airways of asthmatic patients. Journal of ethnopharmacology, 93(1), 57-62.
Mali, K., Dias, R., Hawaldar, V., & Mahajan, N. (2009). Hypoglycemic activity of Caralluma adscendens in alloxan induced diabetic rats. Int J Chem Sci, 7, 517-522.
Marwat, S. K., Rehman, F., Khan, E. A., Khakwani, A. A., Ullah, I., Khan, K. U., et al. (2014). Useful ethnophytomedicinal recipes of angiosperms used against diabetes in South East Asian Countries (India, Pakistan & Sri Lanka). Pakistan journal of pharmaceutical sciences, 27(5), 13338-11358.
Mast, E. E., Alter, M. J., Holland, P. V., & Purcell, R. H. (1998). Evaluation of assays for antibody to hepatitis E virus by a serum panel. Hepatology, 27(3), 857-861.
Masuoka, H. C., & Chalasani, N. (2013). Nonalcoholic fatty liver disease: an emerging threat to obese and diabetic individuals. Annals of the New York Academy of Sciences, 1281(1), 106-122.
Matos, F. d. A. (2000). Plantas medicinais: guia de seleção e emprego de plantas usadas em fitoterapia no Nordeste do Brasil. Plantas medicinais: guia de seleção e emprego de plantas usadas em fitoterapia no Nordeste do Brasil.
Matthews, J. F., Ketron, D. W., & Zane, S. F. (1993). The biology and taxonomy of the Portulaca oleracea L.(Portulacaceae) complex in North America. Rhodora, 95(882), 166-183.
Meve, U., & Heneidak, S. (2005). A morphological, karyological and chemical study of the Apteranthes (Caralluma) europaea complex. Botanical Journal of the Linnean Society, 149(4), 419-432.
Middleton, E., Kandaswami, C., & Theoharides, T. C. (2000). The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacological reviews, 52(4), 673-751.
Mitich, L. W. (1997). Common purslane (Portulaca oleracea). Weed technology, 11(2), 394-397.
Mondelli, M. U., & Silini, E. (1999). Clinical significance of hepatitis C virus genotypes. Journal of Hepatology, 31, 65-70.
Moradpour, D., Brass, V., & Penin, F. (2005). Function follows form: the structure of the N-terminal domain of HCV NS5A. Hepatology, 42(3), 732-735.
Movahedian, A., Ghannadi, A., & Vashirnia, M. (2007). Hypocholesterolemic Effects of Purslane Extract on Serum Lipids in Rabbits Fed with High Cholesterol Levels. International Journal of Pharmacology, 3(3).

139
Nahmias, Y., Goldwasser, J., Casali, M., van Poll, D., Wakita, T., Chung, R. T., et al. (2008). Apolipoprotein B–dependent hepatitis C virus secretion is inhibited by the grapefruit flavonoid naringenin. Hepatology, 47(5), 1437-1445.
Naik, R. M., Venugopalan, V., Kumaravelayutham, P., & Krishnamurthy, Y. (2012). Nutritive value and mineral composition of edible Caralluma and Boucerosia species from the arid areas of Karnataka. International Journal of Agriculture, Environment and Biotechnology, 5(2), 117-125.
Naithani, R., Huma, L. C., Holland, L. E., Shukla, D., McCormick, D. L., Mehta, R. G., et al. (2008). Antiviral activity of phytochemicals: a comprehensive review. Mini reviews in medicinal chemistry, 8(11), 1106-1133.
Nawal M. A. M., Mothana, R. A., Al-Massarani, S., Matheeussen, A., Cos, P., & Maes, L. (2012). Study of the in vitro antiplasmodial, antileishmanial and antitrypanosomal activities of medicinal plants from Saudi Arabia. Molecules, 17(10), 11379-11390.
Nielsen, S. U., Bassendine, M. F., Burt, A. D., Bevitt, D. J., & Toms, G. L. (2004). Characterization of the genome and structural proteins of hepatitis C virus resolved from infected human liver. Journal of General Virology, 85(6), 1497-1507.
Noroozi, M., Angerson, W. J., & Lean, M. (1998). Effects of flavonoids and vitamin C on oxidative DNA damage to human lymphocytes. The American journal of clinical nutrition, 67(6), 1210-1218.
Nunes, G., Silva, M. d., Resende, U., & Siqueira, J. d. (2003). Plantas medicinais comercializadas por raizeiros no Centro de Campo Grande, Mato Grosso do Sul. Rev Bras Farmacogn, 13(2), 83-92.
Obadomi, B., & Ochuko, P. (2001). Phytochemical studies comparative efficacy of the crude extracts of some homeostatic plants in Edo and Delta States of Nigeria. Global J. Pure Appl. Sci, 8, 203-208.
Oh, K. B., Chang, I. M., Hwang, K. J., & Mar, W. (2000). Detection of antifungal activity in Portulaca oleracea by a single‐cell bioassay system. Phytotherapy Research, 14(5), 329-332.
Okwuasaba, F., Ejike, C., & Parry, O. (1987). Effects of extracts of Portulaca oleracea on skeletal muscle in vitro. Journal of ethnopharmacology, 21(1), 55-63.
Oyama, M., Iliya, I., Tanaka, T., & Iinuma, M. (2007). Five new steroidal glycosides from Caralluma dalzielii. Helvetica chimica acta, 90(1), 63-71.
Palombo, E. A. (2006). Phytochemicals from traditional medicinal plants used in the treatment of diarrhoea: modes of action and effects on intestinal function. Phytotherapy Research, 20(9), 717-724.
Paris, R., & Moyse, H. (1969). Precis de matiere medicinale: Paris: Masson. Parry, O., Marks, J., & Okwuasaba, F. (1993). The skeletal muscle relaxant action of
Portulaca oleracea: role of potassium ions. Journal of ethnopharmacology, 40(3), 187-194.
Pause, A., Kukolj, G., Bailey, M., Brault, M., Do, F., Halmos, T., et al. (2003). An NS3 serine protease inhibitor abrogates replication of subgenomic hepatitis C virus RNA. J Biol Chem, 278(22), 20374-20380.
Pawlotsky, J. M. (2006). Therapy of hepatitis C: from empiricism to eradication. Hepatology, 43(S1), S207-S220.

140
Pawlotsky, J. M., & McHutchison, J. G. (2004). Hepatitis C. Development of new drugs and clinical trials: promises and pitfalls. Summary of an AASLD hepatitis single topic conference, Chicago, IL, February 27–March 1, 2003. Hepatology, 39(2), 554-567.
Payer, B., Reiberger, T., Rutter, K., Beinhardt, S., Staettermayer, A., Peck-Radosavljevic, M., et al. (2010). Successful HCV eradication and inhibition of HIV replication by intravenous silibinin in an HIV–HCV coinfected patient. Journal of Clinical Virology, 49(2), 131-133.
Penin, F., Dubuisson, J., Rey, F. A., Moradpour, D., & Pawlotsky, J. M. (2004). Structural biology of hepatitis C virus. Hepatology, 39(1), 5-19.
Pietschmann, T., Kaul, A., Koutsoudakis, G., Shavinskaya, A., Kallis, S., Steinmann, E., et al. (2006). Construction and characterization of infectious intragenotypic and intergenotypic hepatitis C virus chimeras. Proc Natl Acad Sci U S A, 103(19), 7408-7413.
Prabakaran, R., & Kalimuthu, K. (2013). Antibacterial activity of the whole plant of Caralluma nilagiriana Kumari et Subba Rao–an endemic medicinal plant species. Int J Pharm Bio Sci, 4, 42-48.
Prasad, S., Kashyap, R. S., Deopujari, J. Y., Purohit, H. J., Taori, G. M., & Daginawala, H. F. (2006). Development of an in vitro model to study clot lysis activity of thrombolytic drugs. Thromb J, 4(14), 1-4.
Prasad, S., Kashyap, R. S., Deopujari, J. Y., Purohit, H. J., Taori, G. M., & Daginawala, H. F. (2007). Effect of Fagonia Arabica (Dhamasa) on in vitro thrombolysis. BMC Complementary and Alternative Medicine, 7(1), 36.
Proestos, C., Lytoudi, K., Mavromelanidou, O. K., Zoumpoulakis, P., & Sinanoglou, V. J. (2013). Antioxidant capacity of selected plant extracts and their essential oils. Antioxidants, 2(1), 11-22.
Quinlan, M. B., Quinlan, R. J., & Nolan, J. M. (2002). Ethnophysiology and herbal treatments of intestinal worms in Dominica, West Indies. Journal of ethnopharmacology, 80(1), 75-83.
Radhakrishnan, R., Zakaria, M., Islam, M., Chen, H., Kamil, M., Chan, K., et a l. (2001). Neuropharmacological actions of Portulaca oleraceae L v. sativa (Hawk). Journal of ethnopharmacology, 76(2), 171-176.
Raj, S., Stephen, T., & Debski, R. F. (2011). Giant cell hepatitis with autoimmune hemolytic anemia: a case report and review of pediatric literature. Clinical pediatrics, 50(4), 357-359.
Ramadevi, T., Ugraiah, A., & Pullaiah, T. (2012). In vitro shoot multiplication from nodal explants of Boucerosia diffusa Wight--An endemic medicinal plant. Indian Journal of Biotechnology, 11(3), 344-347.
Rashed, A., Afifi, F., & Disi, A. (2003). Simple evaluation of the wound healing activity of a crude extract of Portulaca oleracea L. (growing in Jordan) in Mus musculus JVI-1. Journal of ethnopharmacology, 88(2), 131-136.
Rasheed, A., Afifi, F. U., Shaedah, M., & Taha, M. O. (2004). Investigation of the active constituents of Portulaca oleraceae L.(Portulacaceae) growing in Jordan. Pak J Pharm Sci, 17(1), 37-45.
Rauf, A., Jan, M., Rehman, W., & Muhammad, N. (2013). Phytochemical, phytotoxic and antioxidant profile of Caralluma tuberculata NE Brown. Wudpecker J Pharm Pharmacol, 2(2), 21-25.

141
Ravikumar, Y., Ray, U., Nandhitha, M., Perween, A., Raja Naika, H., Khanna, N., et al. (2011). Inhibition of hepatitis C virus replication by herbal extract: Phyllanthus amarus as potent natural source. Virus research, 158(1), 89-97.
Reed, K. E., Grakoui, A., & Rice, C. M. (1995). Hepatitis C virus-encoded NS2-3 protease: cleavage-site mutagenesis and requirements for bimolecular cleavage. J Virol, 69(7), 4127-4136.
Rehman, S., Ashfaq, U. A., Riaz, S., Javed, T., & Riazuddin, S. (2011). Antiviral activity of Acacia nilotica against Hepatitis C Virus in liver infected cells. Virology journal, 8(1), 220.
Rizk, A. M. (1982). Fitoterapia (Vol. 52). Rizwani, G. H., Ahmad, M., Usmanghani, K., & Ahmad, V. U. (1993). Caratuberside
A2: a new pregnane from Caralluma tuberculata. Spectroscopy letters, 26(8), 1427-1434.
Rizwani, G. H., Usmanghani, K., Ahmad, M., & Ahmad, V. U. (1993). Pregnane glycosides from Caralluma tuberculata. Natural Product Letters, 2(2), 97-104.
Rutter, K., Scherzer, T.-M., Beinhardt, S., Kerschner, H., Stättermayer, A. F., Hofer, H., et al. (2011). Intravenous silibinin as' rescue treatment'for on-treatment non-responders to pegylated interferon/ribavirin combination therapy. Antiviral therapy, 16(8), 1327.
Rydberg, P. A., & Howe, M. A. (1932). Flora of the prairies and plains of central North America.
Ryder, S., & Beckingham, I. (2001). Acute hepatitis. Bmj, 322(7279), 151-153. Saalau-Bethell, S. M., Woodhead, A. J., Chessari, G., Carr, M. G., Coyle, J., Graham, B.,
et al. (2012). Discovery of an allosteric mechanism for the regulation of HCV NS3 protein function. Nature chemical biology, 8(11), 920-925.
Safa, O., Soltanipoor, M. A., Rastegar, S., Kazemi, M., Nourbakhsh Dehkordi, K., & Ghannadi, A. (2012). An ethnobotanical survey on hormozgan province, Iran. Avicenna Journal of Phytomedicine, 3(1), 64-81.
Sakai, N., Inada, K., Okamoto, M., Shizuri, Y., & Fukuyama, Y. (1996). Portuloside A, a monoterpene glucoside, from Portulaca oleracea. Phytochemistry, 42(6), 1625-1628.
Sakamuro, D., Furukawa, T., & Takegami, T. (1995). Hepatitis C virus nonstructural protein NS3 transforms NIH 3T3 cells. J Virol, 69(6), 3893-3896.
Sànchez-Lamar, A., Fiore, M., Cundari, E., Ricordy, R., Cozzi, R., & De Salvia, R. (1999). Phyllanthus orbicularis Aqueous Extract: Cytotoxic, Genotoxic, and Antimutagenic Effects in the CHO Cell Line. Toxicology and applied pharmacology, 161(3), 231-239.
Sarker, S. D., Latif, Z., & Gray, A. I. (2005). Natural product isolation Natural Products Isolation (pp. 1-25): Springer.
Sasidharan, S., Chen, Y., Saravanan, D., Sundram, K., & Latha, L. Y. (2011). Extraction, isolation and characterization of bioactive compounds from plants’ extracts. African Journal of Traditional, Complementary and Alternative Medicines, 8(1).
Satoh, S., Tanji, Y., Hijikata, M., Kimura, K., & Shimotohno, K. (1995). The N-terminal region of hepatitis C virus nonstructural protein 3 (NS3) is essential for stable complex formation with NS4A. Journal of virology, 69(7), 4255-4260.

142
Ray, S., Sharma, A., & Nagaiah, K. (2012). Phytochemical and pharmacological re-evaluation of Caralluma umbellate. International journal of comprehensive pharmacy, 3(1), 1-2.
Seo, Y., Shin, J., Cha, H. J., Kim, Y.-A., Ahn, J.-W., Lee, B.-J., et al. (2003). A new monoterpene glucoside from Portulaca oleracea. BULLETIN-KOREAN CHEMICAL SOCIETY, 24(10), 1475-1477.
Shah, A., Hussain, S., Din, N. U., Bhatti, K. H., Khan, A., Marwat, S. K., et al. (2012). Sacredjungle: a traditional way of conserving endangered ecosystem and biodiversity in semi tribal are, Kurd sharif & SHO (Dis-trict Karak, Khyber Pakhtunkhwa), Pakistan. . Sci Technol Dev 31, 312-326.
Shah, A., Marwat, S. K., Gohar, F., Khan, A., Bhatti, K. H., Amin, M., et al. (2013). Ethnobotanical study of medicinal plants of semi-tribal area of Makerwal & Gulla Khel (lying between Khyber Pakhtunkhwa and Punjab Provinces), Pakistan.
Sheng-Xiang Qiua, Long-Ze Lina, Geoffrey A. Cordella, Mullangi Ramesh, Bobbala Ravi Kumar, Marupaka Radhakrishna, et al. (1997). Acylated C-21 steroidal bisdesmosidic glycosides from Caraluma umbellata. Phytochemistry, 46(2), 333-340.
Shinwari, Z. K., & Gilani, S. S. (2003). Sustainable harvest of medicinal plants at Bulashbar Nullah, Astore (Northern Pakistan). Journal of ethnopharmacology, 84(2), 289-298.
Simmonds, P., & Smith, D. (1999). Structural constraints on RNA virus evolution. Journal of virology, 73(7), 5787-5794.
Simopoulos, P. A., & Robinson, J. (2002). The Omega Diet. Chinese edition. . Shanghai, China: Shanghai Popular Science Press.
Sreelatha, V., & Pullaiah, T. (2010). Induction of somatic embryogenesis and plant regeneration from intermodal explants of Caralluma stalagmifera. Bot. Res. Int, 3(1), 17-20.
Steinmann, E., Penin, F., Kallis, S., Patel, A. H., Bartenschlager, R., & Pietschmann, T. (2007). Hepatitis C virus p7 protein is crucial for assembly and release of infectious virions. PLoS Pathog, 3(7), e103.
Sticher, O. (2008). Natural product isolation. Natural product reports, 25(3), 517-554. Sulkowski, M. S., Ray, S. C., & Thomas, D. L. (2002). Needlestick transmission of
hepatitis C. Jama, 287(18), 2406-2413. Babu,S. K., Subba R. R. V., Radhakrishnan, V. S., Rao, M. J., & Rambabu, S. S. (2008). A
new pregnane steroid from the stems of Caralluma umbellata. Journal of Asian Natural products research, 10(11), 1013-1016.
Tanji, Y., Hijikata, M., Satoh, S., Kaneko, T., & Shimotohno, K. (1995). Hepatitis C virus-encoded nonstructural protein NS4A has versatile functions in viral protein processing. Journal of virology, 69(3), 1575-1581.
Tanko, Y., Daniel, P., Mohammed, K., Jimoh, A., Yerima, M., & Mohammed, A. (2013). Effect of ethanolic extract of Caralluma diazielli on serum lipid profiles on fructose induced diabetes in wistar rats. Scholar Res, 4, 162-166.
Tareen, R. B., Bibi, T., Khan, M. A., Ahmad, M., & Zafar, M. (2010). Indigenous knowledge of folk medicine by the women of Kalat and Khuzdar regions of Balochistan, Pakistan. Pak J Bot, 42(3), 1465-1485.

143
Tatiya, A., Kulkarni, A., Surana, S., & Bari, N. (2010). Antioxidant and hypolipidemic effect of Caralluma adscendens Roxb. ib alloxanized diabetic rats.
Tellinghuisen, T. L., & Rice, C. M. (2002). Interaction between hepatitis C virus proteins and host cell factors. Curr Opin Microbiol, 5(4), 419-427.
Thomssen, R., Bonk, S., Propfe, C., Heermann, K.-H., Köchel, H., & Uy, A. (1992). Association of hepatitis C virus in human sera with β-lipoprotein. Medical microbiology and immunology, 181(5), 293-300.
Tillett, W. S., & Garner, R. L. (1933). The fibrinolytic activity of hemolytic streptococci. The Journal of experimental medicine, 58(4), 485-502.
Tomei, L., Failla, C., Santolini, E., De Francesco, R., & La Monica, N. (1993). NS3 is a serine protease required for processing of hepatitis C virus polyprotein. Journal of virology, 67(7), 4017-4026.
Ugraiah, A., Sreelatha, V. R., Reddy, P. K., Rajasekhar, K., Rani, S. S., Karuppusamy, S., et al. (2013). In vitro shoot multiplication and conservation of Caralluma bhupenderiana Sarkaria-an endangered medicinal plant from South India. African Journal of Biotechnology, 10(46), 9328-9336.
Urbani, A., Biasiol, G., Brunetti, M., Volpari, C., Di Marco, S., Sollazzo, M., et al. (1999). Multiple determinants influence complex formation of the hepatitis C virus NS3 protease domain with its NS4A cofactor peptide. Biochemistry, 38(16), 5206-5215.
Venkatesh, S., Reddy, G. D., Reddy, B. M., Ramesh, M., & Rao, A. (2003). Antihyperglycemic activity of Caralluma attenuata. Fitoterapia, 74(3), 274-279.
Verma, O., Kumar, S., & Chatterjee, S. (1982). Antifertility effects of common edible Portulaca oleracea on the reproductive organs of male albino mice.
Verma, S., Tabb, M. M., & Blumberg, B. (2009). Activation of the steroid and xenobiotic receptor, SXR, induces apoptosis in breast cancer cells. BMC cancer, 9(1), 3.
Voravuthikunchai, S. P., & Limsuwan, S. (2006). Medicinal plant extracts as anti–Escherichia coli O157: H7 agents and their effects on bacterial cell aggregation. Journal of Food Protection®, 69(10), 2336-2341.
Wadood, A., Wadood, N., & Shah, S. (1989). Effects of Acacia arabica and Caralluma edulis on blood glucose levels of normal and alloxan diabetic rabbits. J Pak Med Assoc, 39(8), 208-212.
Wagoner, J., Negash, A., Kane, O. J., Martinez, L. E., Nahmias, Y., Bourne, N., et al. (2010). Multiple effects of silymarin on the hepatitis C virus lifecycle. Hepatology, 51(6), 1912-1921.
Waheed, A. (2001). Pharmacognostic and phytopharmacological studies of'Caralluma tuberculata'and'Fagonia indica'. Gomal University, Dera Ismail, NWFP Pakistan.
Waheed, A., Barker, J., Barton, S. J., Khan, G.-M., Najm-us-Saqib, Q., Hussain, M., et al. (2011). Novel acylated steroidal glycosides from Caralluma tuberculata induce caspase-dependent apoptosis in cancer cells. Journal of ethnopharmacology, 137(3), 1189-1196.
Wakita, T., Pietschmann, T., Kato, T., Date, T., Miyamoto, M., Zhao, Z., et al. (2005). Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat Med, 11(7), 791-796.

144
Wang MB, W., Gu, L., Dong, L., Wang, X., Ling, C., & Li, M. (2007). Protective effect of Portulaca oleracea extracts on hypoxic nerve tissue and its mechanism. Asia Pac J Clin Nutr, 16(1), 227-233.
Wang, S. Y., Tseng, C. P., Tsai, K. C., Lin, C. F., Wen, C. Y., Tsay, H. S., et al. (2009). Bioactivity-guided screening identifies pheophytin a as a potent anti-hepatitis C virus compound from Lonicera hypoglauca Miq. Biochem Biophys Res Commun, 385(2), 230-235.
Wang, Y., Li, H., Ying, X., & Kang, T. (2011). Liquid chromatographic (LC) determination of four bioactive compounds in the Portulaca oleracea L. Journal of Medicinal Plants Research, 5(31).
Wedge, D. E., & Camper, N. D. (2000). Connections between agrochemicals and pharmaceuticals. Biologically Active Natural Products: Pharmaceuticals, 1-15.
WHO. (1999). Hepatitis C: global prevalence (update). Weekly Epidemiological Record, 74, 421-428.
WHO. (2002). Bulletin of the World Health Organization. 80(7), 610. Wilson, G. K., & Stamataki, Z. (2012). In vitro systems for the study of hepatitis C
virus infection. International journal of hepatology, 2012. Wolk, B., Sansonno, D., Krausslich, H. G., Dammacco, F., Rice, C. M., Blum, H. E., et al.
(2000). Subcellular localization, stability, and trans-cleavage competence of the hepatitis C virus NS3-NS4A complex expressed in tetracycline-regulated cell lines. J Virol, 74(5), 2293-2304.
Wu, S. F., Lin, C. K., Chuang, Y. S., Chang, F. R., Tseng, C. K., Wu, Y. C., et al. (2012). Anti-hepatitis C virus activity of 3-hydroxy caruilignan C from Swietenia macrophylla stems. J Viral Hepat, 19(5), 364-370.
Xiang, C., Xiaowei, Z., Xiaojuan, L., Yong, H., Guohai, Z., Xueqi, G., et al. (2014). Effects of crude polysaccharides from Purslane on fatigue induced by forced swimming. African Journal of Biotechnology, 11(4), 923-929.
Xiao, F., Lu, F., & Xu, L. (2005). Effect of different parts of Portulace oleracea on the levels of TNF-alpha and IL-6 in the supernatant of cultured adipose cell. Zhongguo Zhong yao za zhi= Zhongguo zhongyao zazhi= China journal of Chinese materia medica, 30(22), 1763-1766.
Yamashita, T., Kaneko, S., Shirota, Y., Qin, W., Nomura, T., Kobayashi, K., et al. (1998). RNA-dependent RNA polymerase activity of the soluble recombinant hepatitis C virus NS5B protein truncated at the C-terminal region. J Biol Chem, 273(25), 15479-15486.
Yan, Y., Li, Y., Munshi, S., Sardana, V., Cole, J. L., Sardana, M., et al. (1998). Complex of NS3 protease and NS4A peptide of BK strain hepatitis C virus: a 2.2 Å resolution structure in a hexagonal crystal form. Protein Science, 7(4), 837-847.
Yang, Z., Liu, C., Xiang, L., & Zheng, Y. (2009). Phenolic alkaloids as a new class of antioxidants in Portulaca oleracea. Phytotherapy Research, 23(7), 1032-1035.
Yang, Z., Zheng, Y., & Xiang, L. (2007). Study on chemical constituents of Portulaca oleracea. Zhong yao cai= Zhongyaocai= Journal of Chinese medicinal materials, 30(10), 1248-1250.
Yu, X., Qiao, M., Atanasov, I., Hu, Z., Kato, T., Liang, T. J., et al. (2007). Cryo-electron microscopy and three-dimensional reconstructions of hepatitis C virus particles. Virology, 367(1), 126-134.

145
Zabihullah, Q., Rashid, A., & Akhtar, N. (2006). Ethnobotanical survey in kot Manzaray Baba valley Malakand agency, Pakistan. Pak J Plant Sci, 12, 115-121.
Zakaria, M., Islam, M., Radhakrishnan, R., Chen, H., Kamil, M., Al-Gifri, A., et al. (2001). Anti-nociceptive and anti-inflammatory properties of Caralluma arabica. Journal of ethnopharmacology, 76(2), 155-158.
Zakaria, M., Islam, M., Radhakrishnan, R., Habibullah, M., & Chan, K. (1998). Evaluation of anti‐inflammatory activity of Portulaca species. Journal of Pharmacy and Pharmacology, 50(S9), 227-227.
Zhishen, J., Mengcheng, T., & Jianming, W. (1999). The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food chemistry, 64(4), 555-559.
Zhuang, Y., Chen, L., Sun, L., & Cao, J. (2012). Bioactive characteristics and antioxidant activities of nine peppers. Journal of Functional Foods, 4(1), 331-338.
Zimmerman, C. A. (1976). Growth characteristics of weediness in Portulaca oleracea L. Ecology, 964-974.
Zito, P., Sajeva, M., Bruno, M., Maggio, A., Rosselli, S., Formisano, C., et al. (2010). Essential oil composition of stems and fruits of Caralluma europaea NE Br.(Apocynaceae). Molecules, 15(2), 627-638.
Zuo, G., Li, Z., Chen, L., & Xu, X. (2007). Activity of compounds from Chinese herbal medicine Rhodiola kirilowii (Regel) Maxim against HCV NS3 serine protease. Antiviral research, 76(1), 86-92.

146
6.0 Appendix
Table of Contents 1.0 Introduction .................................................................................................................. 21
1.1 Natural Products from Times of Yore to 21st Millennium........................................... 21
1.2 Status of Medicinal Plants and their Research in Pakistan .......................................... 23
1.3 Preface to Genus Caralluma and Caralluma tuberculata ............................................ 24
1.3.1 Active Constituents ............................................................................................. 25
1.3.2 Pharmacological worth of Caralluma tuberculata and other species of Caralluma
................................................................................................................................... 30
1.4 Introduction to Portulaca oleracea ............................................................................ 34
1.4.1 Phytochemistry of Portulaca oleracea................................................................. 36
1.4.2 Pharmacological attributes of Portulaca oleracea ............................................... 36
1.5 Hepatitis.................................................................................................................... 40
1.6 Hepatitis C and its Virus ............................................................................................. 40
1.6.1 Transmission of Pathogen and Genotype Prevalence .......................................... 41
1.6.2 HCV Structure and Genome ................................................................................ 42
1.6.3 HCV Non-Structural and Structural Proteins ........................................................ 43
1.6.4 HCV RNA Replication and Potential Drug Target.................................................. 48
1.6.5 In vitro Infection Systems for HCV Replication ..................................................... 49
1.6.6 Available Treatment Options against HCV ........................................................... 53
1.6.7 NS3 Gene as Candidate Drug Target.................................................................... 54
1.6.8 Ayruvadic Extract and Plant Derived Compounds as anti HCV Drug ..................... 57
1.7 Goals/Objectives ....................................................................................................... 60
2.0 Chemicals, Equipment and Techniques .......................................................................... 61
2.1 Chemicals ................................................................................................................. 61
2.1.1 Materials for Chromatography ............................................................................ 61
2.1.2 Chemicals and Reagents used for Determination of Antioxidant.......................... 62
2.2 Instrument and Apparatus .......................................................................................... 63
2.3 Materials used for Biological Activities ..................................................................... 63
2.3.1 Cell Lines ........................................................................................................... 63
2.3.2 Plasmids ............................................................................................................. 63
2.3.3 Reagents & Chemicals ........................................................................................ 64
2.3.4 Designing of Primers .......................................................................................... 64

147
2.4 Instrumentation & Techniques .................................................................................. 64
2.4.1 Physical Constant ............................................................................................... 65
2.4.2 Spectroscopy ...................................................................................................... 65
2.4.3 Thin Layer Chromatography ............................................................................... 65
2.4.4 Column Chromatography (CC) ........................................................................... 65
2.5 Methodology ............................................................................................................. 66
2.5.1 Biochemical Analysis of Plants ........................................................................... 66
2.5.2 Collection, Identification and Preservation of Medicinal Plant Materials ............. 66
2.5.3. Plant Extract Preparation.................................................................................... 68
2.5.4 Screening of Plant Extracts for Cellular Toxicity Analysis via Trypan Blue Dye
Exclusion Method ....................................................................................................... 68
2.6 Anti-HCV Analysis of Compounds in Liver Cells...................................................... 70
2.6.1 Antiviral Analysis of Plant Extract against HVC Ns3 Protease via Transfection
Assay .......................................................................................................................... 71
2.7 Gene Expression studies via Reverse Transcriptase PCR (RT-PCR) .......................... 71
2.7.1 Extraction of Total RNA ..................................................................................... 71
2.7.2 Synthesis of cDNA ............................................................................................. 72
2.7.3 Reverse Transcription PCR (RT-PCR) for Gene Expression Analysis ...................... 72
2.7.4 Expression studies via Real-time PCR................................................................. 73
2.8 Western Blotting of PO extracts ................................................................................ 74
2.8.1 Protein Isolation ................................................................................................. 74
2.8.2 Protein Quantification ........................................................................................ 75
2.8.3 Protein Samples Preparation .............................................................................. 75
2.8.4 SDS Gel Electrophoresis ...................................................................................... 75
2.8.5 Transfer of Protein and Immunoblotting ............................................................. 76
2.8.6 Enhanced Chemiluminescence (ECL) Detection ................................................... 76
2.9 Analysis of the Two Plants Extract That Exhibit Positive Potential against HCV ....... 77
2.10 Phytochemical Screening ......................................................................................... 77
2.10.1 Qualitative Analysis of the Extracts .................................................................. 77
2.10.2 Quantitative Determination of the Chemical Constituency Specifically
Antioxidants ................................................................................................................ 80
2.11 Anti-clotting Properties of CTS and POL Extracts ...................................................... 84
2.11.1 Sample Preparation .......................................................................................... 85
2.11.2 Preparation of Anti-Clot Test Solution ............................................................... 85
2.11.3 Clot Lysis ........................................................................................................... 85

148
2.12 Antibacterial assay of extracts ................................................................................. 85
2.13 Bioassay guided extraction of CTS and POL ........................................................... 86
2.14 Isolation of Compounds via Column Chromatography ............................................. 87
2.14.1 Isolation of Compounds .................................................................................... 87
2.14.2 Physico-chemical Characteristics of Isolated Compounds.................................. 88
2.14.3 Elucidation of Compounds Structure by spectroscopy ....................................... 88
3.0 Results and Discussion .................................................................................................. 90
3.1 Collection of Botanical Samples and Preparation of Extract ....................................... 91
3.2 Percentage yield of plants extract............................................................................... 92
3.3. Toxicological Studies of Plant Extracts in CHO and Liver Cell Lines ....................... 92
3.3.1 Trypan Blue Dye Exclusion Assay ...................................................................... 93
3.3.2 Cytotoxic Screening of Phyto Extracts via MTT Assay .......................................... 93
3.4 Anti-HCV Assay ....................................................................................................... 96
3.5 Inhibition of HCV NS3 Gene Expression by Selected Organic Extracts ..................... 99
3.6 Western Blotting ..................................................................................................... 101
3.7 Phytochemical analysis............................................................................................ 102
3.7.1 Qualitative Analysis of CTS and POL ....................................................... 102
3.7.2 Quantitative Methods for Screening of Phytoconstituents ........................... 103
3.8 Antioxidant Potential of Biologically Active Plant Extract Fractions .......................... 103
3.8.1 Total Phenolic, Total Flavonoids and Total Carotenoid Contents ....................... 103
3.8.2 Free Radical Scavenging Activity of Extracts by DPPH, ABTS and FRAP Assays
................................................................................................................................. 106
3.8.3 Quantitative Estimation of Phenolic Acid by HPLC .......................................... 108
3.10 Antithrombotic Potential of Extracts ...................................................................... 109
3.11 Screening of Antibacterial Activity of Samples ...................................................... 110
3.12 Extraction, Purifcation and identification of Compounds Obtained from CTSE Extract
..................................................................................................................................... 112
3.12.1 Compound CT-EtOAc-01 ............................................................................... 113
3.12.2 Compound CT-EtOAc-02 ............................................................................... 115
3.12. 3 Compound CT-EtOAc-03 .......................................................................... 118
3.12.4 Compound CT-EtOAc-04 ................................................................................. 119
3.12.5 Compound CTEtOAc-05 ............................................................................ 120
3.13 Discussion ............................................................................................................. 122
4.0 Future Prospects ......................................................................................................... 126
5.0 References .................................................................................................................. 128

149
6.0 Appendix .................................................................................................................... 146

150

151
152
153