Embed Size (px)
Citation preview

P e rVisionRes., Vol. 36, No. 15, pp. 2303-23]0, 1996
Copyright01996 ElsevierScience Ltd. All rights reserved0042-6989(9s)o0289.8 Printed in Great Britain
0042-6989/96$15,00+ 0,00
Binocular Correlation Detection with OrientedDynamic Random-line StereogramsLAWRENCE K. CORMACK,*~RUSSELL B. RIDDLE*
Received 22 September 1994; in revised form 17July 1995
Stereopsis relies principally on the extraction of horizontal retinal disparities. As such, we assumethat the vertical contours (i.e., horizontal contrast energy) are of principle import for stereopsis.Yet there are theoretical reasons for believing that horizontal contours should be involved inbinocular matching (if not stereopsis proper) as well. First, they would facilitate the computation ofvertical disparities, which are necessary for the control alfdisjunctive eye movements and perhapsthe computation of absolute depth. Second, the process of binocular matching is a two-dimensionalone; its solution requires information along both principle orientations. In this study, we havemeasured the efficacy with which horizontal or vertical contours can be binocularly matched bymeasuring thresholds for the detection of interocular correlation for oriented dynamic random-linestereograms. We find that the slopes of the psychometric functions are almost a factor of twosteeper when matching vertical contours, indicating a narrower noise distribution along thedecision axis associated with these stimuli. Copyright @ 1996 Elsevier Science Ltd.
Stereopsis Orientation Binocular Disparity
INTRODUCTION
The process of extracting disparity information from thetwo eyes’ images can be conceptually divided into twostages, one of binocular combination (solving thebinocular correspondenceproblem) and one of disparitycomputation or extraction. Disparity extraction can beexamined psychophysically by making stereoacuity,disparity incrementand ‘formin depth’judgments,whilebinocular combination can be examined psychophysi-cally by making judgments of interocular correlation(Tyler & Julesz, 1978; Cormack et al., 1991).The logicbehind measuring interocular correlation thresholds issimply that in order to determine how well the visualsystem matches binocular images, one should measurethresholds for the degree to which binocular imagesmatch.
In principle, the process of binocular combinationcould use information at all orientations in order toachievebinocularmatching.Some registrationof verticaldisparities is necessary for the control of vergence eyemovements (e.g., Stevenson & Schor, 1993), whichwould be facilitated by the use of horizontal edgeinformation.It is also possiblethat verticaldisparitiesare
*Department of Psychology and Center for Vision and Image Sciences,The University of Texas at Austin, Austin, TX 78701, U.S.A.
~To whom all correspondence should be addressed [Fax +1-5124715935; Email [email protected]. edu].
registered as a source of absolute distance informationwith which to scale horizontal disparities (e.g., Bishop,1989).Regardless,the process of binocular combinationis inherently two-dimensional, since it is extremelyunlikely that the relative orientation of the visual axescan be known with sufficient precision to drive imageregistrationvia afferent signals alone.
Three hypotheses can be generated by makingassumptions about the degree to which horizontal andvertical edge informationis used for binocularmatching.If observers do, in fact, have access to a mechanism ofbinocular matching which utilizes all of the informationin the stimulus, regardless of its orientation, thenthresholdsfor the detection of interocular correlation indynamic random-line stereograms should not show ahorizontal/verticalanisotropy.
However, if the process of binocular combination towhich we have accesspsychophysicallyexists solelyas aprecursor to stereopsis, correlation detection might beexpected to show a drastichorizontal/vertica]anisotropy.In other words, if observers are using the output ofstereopsis mechanisms to make judgments about theinterocularcorrelationof the stimulus,then the ability tomake these judgments should be dramatically impairedwhen onl,yhorizontal edge informationis present.
The third hypothesisis intermediateto the above two;due to the lesser demands eventually placed on theprecision of horizontal edge matching by disparitycomputations, the process of binocular combination
2303

2304 L. K. CORMACK and R. B, RIDDLE
100%
w
o%
75%
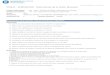
w 03 /FIGURE 1. Examples of single frames of the vertical stimuli. The crosshairs are to aid fusion. The stimuli were dynamic, whichis illustrated here by the additional frames displaced in time, and were surounded in space by darkness and in time by a meanluminance field containing a fixation mark. Top: Fully correlated random line stereogram. Center: Uncorrelated random linestereogram. Bottom: 75~0 correlated random line stereogram after a simulated temporal integration of 90 msec. This more
accurately illustrates the perceived contrast of the stimuli during the dynamic stimulus presentation.
might have evolved to be somewhat better for verticalthan horizontal stimuli. In this case, a horizontal/verticalanisotropywould exist, but would not be as drastic as thesecond hypothesispredicts.
In the psychophysicaland eye movement literature, itseems that a horizontal/vertical anisotropy almostinvariably appears, regardless of the stimuli or the taskemployed (e.g., psychophysics:Volkman, 1859; Fender& Julesz, 1967; Stevenson & Schor, 1994; Mansfield&Parker, 1993; eye movements: Perlmutter & Kertez,1978; cf. Erkelens, 1987 w. Stevenson & Schor, 1993);only Mayhew & Frisby (1978) have reported otherwise.
In the physiologicalliterature, the situationis reversed.Following Barlow et al. (1967), no horizontal/verticalanisotropy had been reported for disparity tuning in thevisual cortex of cat or monkey, either in terms of theamountof receptivefielddisparityor the numbersof cells
tuned to disparity in various directions(e.g., Pettigrewetal., 1968; Nikara et al., 1968; Joshua & Bishop, 1970;von der Heydt et al., 1978; Ferster, 1981; LeVay &Voight, 1988), until DeAngelis et al. (1991) demon-strated that the phase disparity of receptive fields in catvisual cortex was distributed more broadly in thehorizontaldirection.
In this study, we wished to determine whether therewas any difference in psychophysicalcorrelation detec-tion when the stimuli contained only vertical orhorizontal edge information. This was achieved bymeasuring correlation thresholds for vertical and hor-izontal dy:rtamic random-line stimuli. We report apronounceddifference in the slopes of the psychometricfunctions.This difference in slope is consistentwith theplethora of horizontal/vertical anisotropies reported inthepsychophysicalliteratureand the recentphysiologyof

BINOCULAR CORRELATION DETECTION 2305
DeAngeliset al. (1991),but the apparent lack of obviousconcomitant anisotropies in the earlier physiologicalliterature remains somewhat puzzling.
METHODS
Observers
Four observers participated in the main portion of thestudy. Three of these were relatively inexperiencedpsychophysicalobservers,while the fourth had extensiveexperience.The three inexperiencedobserversunderwentextensive training until they seemed comfortable withtheir criterion and their thresholds appeared to stabilizefor a given stimuluscondition.All observershad normalor corrected-to-normal acuity and no known history ofocular motility abnormalities. All observers readilyperceived simple figures portrayed in static anaglyphicrandom-elementstereograms.
Apparatus and stimuli
The experimentswere run on a MacintoshQuadra 950microcomputer. The stimuli were displayed on a single13 inch color monitor driven from the Quadra’s built-invideo and viewed through a mirror haploscope at aviewing distanceof 57.3 cm (as described in Cormack etal., 1994). The haploscope was adjusted using astereoscopic matching procedure such that the conver-gence and accommodativedemand exactly matched theviewing distance.*
Stimuliwere dynamicrandom-linestereograms,singleframe examples of which are shown in Fig. 1.Conceptually,these are simply a single row (or column)of a standard random-elementstereogramrepeated down(or across) the stereogram, producing the random-linestereogram.Each half-imageof a stereo-pairwas 2 deg x2 deg (square) within a dark surround. Each line(analogous to an element in a standard random-elementstereogram) was approximately 2 arcmin across. Thedensity of the stereogramswas 50%; on average, half thelines were light. All stimuli were presented at zerodisparity and centered on the fixationpoint.
The stimuliwere computedand drawn into memoryona trial-to-trial basis using uniformly distributed pseudo-random numbers produced with the Bayes–Durhamalgorithm (Press et al., 1988). During a stimuluspresentation, frames of the stimulus were copied from
*In this procedure, a physical object is placed at the same viewingdistance as the monitor (in this case, 57.3 cm), such that the subjectcan view it and the monitor simultaneously by peering just over theinitial mirror-pair of the haploscope; the object appears in thesubject’s upper visual hemitield and the stereoscopic view of themonitor appears in the lower hemifield (if the initial mirror pair ofthe haploscope is sufficiently close to the pupil plane, as it was inour case, a superimposed view can be obtained). A depth match isthen made between a stereoscopic fixation point cmthe monitor andthe physical object by adjusting the haploscope mirrors. Once astereoscopic match has been made, the absolute disparity (and thusthe vergence demand) of the stereoscopic fixation point on themonitor is that of a physical object at the desired viewing distance.
main memoryto the video memory in synchronywith themonitor’s frame refresh. The stimulus frame rate of thestereograms was 33,3 Hz (30 msec stimulus frameduration; each stimulus frame lasting two video frames)and a total of 40 stimulusframes were presented at eachintervad.This correspondsto a total interval duration of1.2 sec. These parameters were empirically chosen tomaximlizethe performanceof the naive observers.
The stereograms all consisted of two gray levels, themean luminanceof which was 60 cd/m2.The contrast(asmeasured on a single stimulus frame) was jitteredbetween 99 and 74% on an interval-by-intervalbasis.~The effective contrast was somewhat lower than thesevalues due to the temporal integration of the visualsysteml; Fig. 1 (bottom) illustrates the approximateperceived contrast of the stimuli.
Psychophysicalprocedure
A temporal two-alternative forced-choice method ofconstant stimuli procedure was used. Five hundredmillisecondsbefore the first alternativewas presented, awarning tone occurred. Tones also occurred coincidentwith the onset of the first and second intervals, whichwere separated by 500 msec. As stated above, eachstimulus interval lasted 1.2 sec.
A uniform field at mean luminance, containing a 20arcmin square fixation box flanked vertically by 20arcmin longnoniuslineswas presentbetween the trialsaswell as during the interval between the two alternatives.While the stimulus intervals were much too long toobviatedisjunctiveeye movements,the fixationmark didensure that vergence posturewas at a consistentpositionat stimulusonset.
The observer judged whether the first or secondintervalcontained a non-zero correlation,and respondedvia the keyboard;a tone signaleda correct response.Eachobserver performed 15 runs per orientation. A “run”consistedof 10blocksof five trials each, one trial at eachof the five possible interocular correlation levels. Theorder of the trialswas randomizedwithin blocks (methodof constant stimuli). The interocular correlation levelswere chosen to bracket the observer’sthresholdbased onpilot data.
Some additionaldata were collected using five runs of30 blclckseach (for LC); in all cases, a threshold wasbased on 150 trials at each of five stimulus levels.
RESULTS
Figure 2 showsthe psychometricfunctionsfor the fourobservers. The main graphs plot percent correct as a
~The contrast was jittered to prevent observers from simply selectingthe interval with the higher binocular contrast as the correlatedinterval, as this strategy would require no binocular matching. Thisis an innocuous safeguard, as it is known that stimulus contrast hasno effect on correlation thresholds when contrast is above five orsix threshold multiples (Cormack et al., 1991). Moreover, thethresholds reported here are the same as those that were collectedusing constant contrast.

100
90
80
70
60
50
40
L. K. CORMACK and R. B. RIDDLE
n
Lc
1...I...I...., ,.4() 0.25 ().5 0.75 1
1
0 0.25 0.5 ().75 1
1:T
O 0.25 0.5 ().75 140 I
o 0,25 ().5 ().75 1
Int e ro c uIar
100
90
80
70
60
50
40
A AI I + + I 9
. DL z?—
0$4
d’ ‘2
~o
~H;-0 0.25 0.5 0.75 1
() 0.25 0.5 0.75 1
100 1 I I
BH I90
80
70
60
5’0
4!()
1-
() 0.25 0.5 0.75 1
Correlation
FIGURE 2, Percent correct t 1 SE obtained with the horizontal (diamonds) and vertical (squares) stimuli for the four observers.Also shown are best-fit sigmoidal (Weibull) functions for illustrative purposes. Small diamonds for LC and DL are dataconfirming an upper asymptote of 100% for the horizontal stimuli. The inset graphs show the functions plotted as d’ vsinterocular correlation with best-fit straight lines, from which estimates of threshold values and psychometric slopes were
obtained, Three of the four observers show a markedly higher sensitivity to the vertical stimuli.
function of interocular correlation for both the vertical(square symbols) and horizontal (diamond symbols)stimuli. The solid lines are best-fit sigmoidal (Weibull)functions,which are shown to help illustratethe trends inthe data. The smallerdiamondsshownfor LC and DL areadditional data that were collected to confirm thatperformance asymptote at 100% for the horizontalstimuli (RR was unavailablefor further data collection).
*An unfortunate aspect of fitting percent correct data with sigmoidalfunctions such as the Weibrrll or cumulative normal is that the slope(or steepness) parameter of these functions causes them to “pivot”about the 50Y0 correct point on the function. As such, the slopeparameter does not map to the amount of noise along the decisionaxis in a direct fashion. This can be appreciated intuitively byconsidering that increasing the noise on the decision axis mustcause performance at all non-zero signal levels to deteriorate.
Errol’bars show ~ one standard error; where omitted,they are smaller than the plot symbols.It shouldbe notedthat much of the variability is inherent in the stimuli;correlation models and “ideal observers” show compar-able amountsof variability (cf. Cormack et al., 1994). Itis readily seen that the rate of improvementas a functionof interocular correlation is greater for the verticalstimuli.
For purposesof further analysis,percentcorrectvalueswere converted to d’ values and the resulting psycho-metric data were fitted with linear functions. Thesetransformeddata and best-fitlines are plotted in the insetgraphsof Fig. 2.* Regardlessof the methodof plotting,itcan be readily seen that performance improves morequickly as a function of interocular correlation for thevertical stimuli in three of the four observers. For thefourth observer, performance was identical for both

BINOCULAR CORRELATION DETECTION 2307
a.
1I Veiiical Horizontal I 1
0
LC RR DL BH
‘5 ~
10
5
0
T
LC
ml❑ WRTICAI>
❑ IIORIZONI’AL
RR 1)1,
subject
FIGURE 3. Slopes and “threshold” values from the straight line fits tothe psychometric data (d’ vs interocular correlation) shown in Fig. 1.(a) Correlation thresholds (d’=1) for vertical (solid bars) and horizontal(open bars) for four observers. Hatched bars show d’=2performance.Afl observers except BH performed consistently better for verticalrandom-line stimuli. (b) Slopes of the psychometric functions forvertical (solid bars) and horizontal (open bars) for four observers. Errorbars show the formal standard errors (68% confidence intervals) of thebest-fit slopes. For all observers except BH, the slopes of thepsychometric functions are much shallower for the horizontalcondition, indicating that the internal representation of the correlation
signal is noisier for these stimuli.
orientations and substantially worse than for the otherthree observers.
Figure 3(a) shows the interocularcorrelation at whichperformance reached both the d’= 1 (unhatched) andd’= 2 (hatched) levels for both the vertical (solid) andhorizontal (open) stimuli. It is clear from this graph thatthe difference in performance is not capturedby definingand examining a single “threshold” point on thepsychometric functions; the difference in performancebetween the conditions is not simply an offset along thesignal axis.
Perhaps then, a more important comparison is thatbetween the slopes of the psychometric functionsobtained for horizontal and vertical stimuli. Figure 3(b)showsthe slopesof thebest-fittinglinear functionsfor thefour observersin both conditions.The error bars show tone formal standarderror (i.e., 68% confidenceintervals)from the line fits. For three of the four observers, thedifference in the slopes between the two conditions isquitepronounced.For these three observers,the slope forthevertical stimuliwas about 1.8 timesas steep as that forthe hcmizontalstimuli. As the slopes of these lines areessentially a mapping from a stimulus strength axis to adecision axis, we can conclude that, at some level, theinternidnoisedistributionsassociatedwith the horizontalstimuli are almost a factor of two broader than thoseassociatedwith the vertical stimuli.
The upper asymptotes of the psychometric functionswere (examinedfor any systematic trend by fitting thepercent correct data with Weibull functionsand allowingthe upper asymptote to vary along with the offset andsteepnessparameters.However,they were alwaysnear orequal to 100% (the data were also fit with the upperasymptotesof the Weibull functions fixed at 100%, andthe fit:sdid not change substantially).We also collectedfurthe:rdata using the horizontal stimuli at high signallevelsfor observersLC and DL. These data are plotted asthe small diamond symbols in Fig. 2, and they confirmthat the upper asymptoteis indeed 100%correct. This isan indicationthat the observerswere not simplyattendingto irrelevant aspects of the horizontal stimuli, a strategythatwould result in less than perfectperformance,even atarbitrarilyhigh signal levels (see e.g., Swanson & Birch,1992).
Beficsrewe can conclude that the visual system iscapablle of matching horizontal contours, albeit lesseffectively than it matches vertical contours, there is oneartifact in the horizontalstimuli that must be considered:the correlation carried by the (vertical) terminations ofthe horizontallines.It is possiblethatobserverswere ableto base their judgments on the correlation of thesevertical ends, even thoughall observersfixatedthe centerof the stimulus. To eliminate this possibility, a controlcondition was run in which a border of uncorrelateddynamic noise flanked the horizontal line stimulus oneither side. This border prevented the interocularcorrelationimposedon the signal intervalfrom manifest-ing itself at the line-ends.Thus, any and all detection ofthe signal must based on the information carried by thehorizcmtalcontours of the stimulus.
Figure 4 shows the psychometric functions from oneobserver. In the figure, percent correct is plotted as afunctionof correlationfor three conditions:vertical lines,horizcmtallines and horizontallines with an uncorrelatedborder. Overall, thresholdswere slightly higher as thesedata were collected using a brief (180 msec) stimulusdurationto obviateeye movements,but they are elevatedby exactly the amount one would expect, given thereduced informationcontent in the stimulus(Cormack et

2308 L. K. CORMACK and R. B. RIDDLE
100 . 1 I I I I 1 I I 1
95
90
85
80
75wu~ ~()L~ 65@
60
50 0 IIorizontal
+ IlorizontalWIBorder45
40. .
35 1 I I 1 1 I 1 1 I0 m q *, m a y cc, m o
0 0 0 0 0 0 0 0 0
Correlation
FIGURE 4. Psychometric functions for subject LC for vertical lines (open squares), horizontal lines (open diamonds) and horizontal lines with anuncorrelated border (solid diamonds). The psychometric functions for the latter two conditions are virtually identical. Overall, the thresholds areslightly higher than in the main experiment due to the brief presentation duration (frame rate was 66.7 Hz and the stimuli were 15 lines across in
space x 12 video frames in time).
al., 1994).Clearly, the correlationof the vertical ends ofthe horizontallineswere not contributingto performance.
DISCUSSION
Stereopsiscan be conceptualizedas a multi-stageserialprocess comprising many parallel subunits at each stage(e.g., Tyler, 1983). By tailoring the stimulus and theobservers’ task, one can emphasize the importance ofinformation in various hypotheticalstage/subunitcombi-nations in the hope of revealing the role that the variousstages and subunitsplay in the process of stereopsis.
In this study, we have used random-line stimuli toinvestigate the relative roles of horizontal and verticalinformation. The observers’ task was to discriminate apartially correlated stimulus interval from an uncorre-lated one in order to emphasize the stage of binocularmatching, rather than the subsequent stage of disparityextraction.
There were two major findings of this study. First,correlation detection can be done using stimuli thatcontain only horizontallyoriented edges, in the absenceof both binocular contrast cues and the binocular rivalrynormally associated with static decorrelated stimuli.Second, while thresholds, as typically defined, weresimilar for vertical and horizontal stimuli, the slopes ofthe psychometric functions were much steeper for thevertical stimuli. Taken together, these findings demon-strate that the visual system is capable of matchinghorizontalcontours,but not with the same efficacythat it
matches vertical contours. Moreover, the fact that thepsychometricslopes are shallower for horizontal stimuliindicates that the internal noise distribution along thedecision axis associatedwith these stimuli is broader.
If we assumethat the decisionstage is commonto boththe vertical and horizontal stimulus conditions, then thebroadler(noisier) representationof the horizontal stimulialong;the decision axis must reflect a greater amount ofnoise in the internal representation of the correlationsignal from the horizontalstimuli relative to the verticalstimuli. As the assumptionof a common decision stageseems to be a safe one, it becomes necessary to considerpossiblesourcesfor the additionalnoise. It is not intrinsicto the stimuli; the correlation signals they carry arestatistically identical (as the stimuli themselves areidentical except for an effective 90 deg rotation of thehalf-images). It is not due to observers attending toirrelevant aspects of the stimuli, as this would affect theupper asymptotes of the psychometric functions muchmore drastically than the slopes (cf. Swanson & Birch,1992).It is also unlikelythat the additionalnoise is due toany clifferencein the propertiesof cortical cells tuned todifferentorientationsor directionsof disparities,or to theabsolute number of cells tuned to horizontal or verticalstimulusorientation.Corticalcells have a variance that isproportionalto their mean firing rate (cf. Tolhurst et al.,1983; Geisler & Albrecht, 1995). When measured overbrief durations, the constant of proportionality variesfrom 0.75 to 6.0 and averages about 1.5. No systematicrelationshiphas been foundbetween the variabilityof the

BINOCULAR CORRELATION DETECTION 2309
firing rate of cortical neurons and any of their tuningproperties, such as spatial frequency. This having beensaid, it must be pointed out that the relationshipbetweenresponse variability and preferred orientation has notbeen explicitly addressed. Nonetheless, it remainsunlikely that a reliable relationshipexists between thesevariables.Finally, it is unlikelythat the additionalnoiseisdue to a smallernumberof neuronsprocessinghorizontalstimuli (which would increase the variability of anensemble average). While there do seem to be fewerneurons available for the processing of oblique stimuli,the number of neurons tuned to horizontal and verticalorientations is quite similar (DeValois et al., 1982).
When formulatingan explanation for our findings,wemust be mindful of other behavioral findings, whichcome from studies on fusion (Volkman, 1859;Fender &Julesz, 1967; Schor & Tyler, 1981), eye movements(Perlmutter & Kertez, 1978; cf. Erkelens, 1987 w.Stevenson & Schor, 1993), stereopsis (Fender & Julesz,1967; Stevenson & Schor, 1994; Mansfield & Parker,1993) and correlation detection (Stevenson & Schor,1994). A pervasive conclusion is that the range ofbinocular matching is larger in the horizontal than thevertical direction. Thus, it would be desirable to relatethis seemingly ubiquitousfindingwith the results of ourown experiment.
We must also, however, view the psychophysicsagainst the backdrop of the physiology, which is thatno horizontal/vertical anisotropy seems to be presentwhen positional disparity tuning is analyzed (e.g.,Pettigrew et al., 1968; Nikara et al., 1968; Joshua &Bishop, 1970;von der Heydt et al., 1978;Ferster, 1981;LeVay & Voight, 1988). The only anisotropy that hasbeen reported is that of the interocular phase differencebetween the monocularreceptive fieldsof binocularcellsin visual cortex by DeAngelis et al. (1991).* Until boththe phase disparity tuning and the positional incongruityof receptivefieldsare measured,the relationshipbetweenpsychophysicsand physiologywill be highlyspeculative.
Two possible schemes can be considered, however.
*These authors found that a phase disparity anisotropy exists in whichcells selective to vertical orientations have a broad distribution ofphase tuning, whereas cells tuned to horizontal orientations tend tohave little interocular phase difference between the monocularreceptive fields. However, the interpretation of this result must betempered by the fact that the positional disparities of the receptivefields were not measured. Since it has been established that there isa broad distribution of positional disparities of cells tuned to allorientations (e.g., von der Heydt et al., 1978; Ferster, 1981), it isreasonable to assume that many of the cells with a zero phasedisparity in the DeAngelis et al. (1991) study actually had a netdisparity tuning due to the (unmeasured) positional disparity. It isequally reasonable to assume that many of the cells had a netdisparity tuning either larger or smaller than the phase disparityalone suggested. Thus, it seems that there are perhaps twomechanisms by which single cells may be capable of respondingto disparate stimuli, by possessing either, or both, a positional offsetand a phase difference between the right and left monocularreceptive fields. Currently, models that seek to exploit theseproperties are fairly unconstrained, as no group has measured boththe positional and phase disparities in a sample of cortical units.
For both schemes, the horizontal/verticalanisotropythatwe report is viewed as a manifestation of a largerdisparity range in the horizontaldirection relative to thevertical direction,as found in the psychophysicalstudiescited ;above.The firstschemeis that a comparisonis madeacross disparities, either at the level of primary visualcortex (possibly in the form of disparity domaininhibition) or at a subsequent processing stage. Whenwe manipulate the correlation (at zero disparity) of arandom-line (or random-element)stimulus, some subtlechanges also occur at non-zero disparities. In a fullycorrelated stimulus comprising n binocular elements,there will be rz2possible binocular matches, n of whichare “valid” matches at zero disparity and rz2—n ofwhich are “false” or “ghost” matches occurring atrandom, non-zero disparities. In an uncorrelated stimu-lus, all rZ2matches are distributed randomly such thatthere is a slight average increase in the overallnumberofmatches at non-zero disparities. Moreover, in a fullycorrelated stimulus, the “false” matches are distributedsymmetrically about the horopter (as originally pointedout b,y Tyler, 1977). It follows that a system that candetect changes over a larger disparity range will haveaccess to more information concerning changes incorrelation, and thus perform better, than will a systemthat has a range restricted to near-zero disparities.Thus,our fi:ndingswould be compatiblewith a larger disparityrange encompassed by binocular cells tuned to verticalstimuli,which is precisely what DeAngelis et al. (1991)found. There is one caveat, however; it is conceivablethat a simple implementationof a larger disparity rangewould actuallydecrease,not increase, the signal-to-noiseratio :sincethe strongest signal under these conditions isobviouslyat zero disparity.This potentialdifficultycouldperhaps be overcome with mechanisms sensitive tochanges in the presence of matches symmetric about thehoropter (Tyler, 1977).
The second scheme employs an elliptical integrationarea for binocular correlation (operating across allorientations),which would result in a noisier signal forour horizontal stimuli than for our vertical stimuli,because of the relative reduction in the amount ofinformation-the number of samples—availableto thevisual system(Cormack et al., 1994).In this scheme, thedisparity-tuned neurons observed in primary visualcortex would function as the multiplicationelements ofan operation analogous to a cross-correlation. Thisoperationwould then be completedat a subsequentstageby a summationacrosselements tuned to like disparities,resultingin a cross-correlationfunctionfor a givenvisualdirection. The effect of horizontally elongated spatialneighborhoods over which the cross-correlation isconductedwould be two-fold. First it would result in anincreased disparity range in the horizontal direction.Second, it would result in a signal with relatively lessnoise arisingfrom the vertical randomline stimulidue tothe increased number of samples in the horizontaldirection.
Although it is conceivable that the first scheme might

2310 L. K. CORMACK and R. El. RIDDLE
be implemented in the form of inhibitory interactionsamong disparity-selectiveneurons,both schemesseem tosuggest that a stage of stereoscopic processing mightoccur beyond that represented by the initial leveldisparity-sensitiveneurons. In other words, we shouldentertain the notion that the disparity-specificneuronsofprimary visual cortex might not be the physiologicalsubstrateof stereopsis,but rather the firststageof a rathercomplex process.
CONCLUSION
We have investigatedthe ability of observersto detectinterocular correlation in oriented dynamic random-linestereograms, in which potential cues such as binocularrivalry and binocular contrast had been eliminated. Wefound that the slopes of the psychometricfunctionswerealmost a factor of two steeper for the vertically orientedstimuli.This differencein slope indicatesthat the internaldistribution along the decision axis associated with thehorizontally oriented stimuli is much broader. Thispresumably arises from an additional amount of noisein the internal representation of the correlation signalfrom the horizontal stimuli. This additional noise is notintrinsicto the stimuli,is not due to observersattendingtoirrelevant aspects of the stimuli, and is not likely to beaccounted for by the response properties or numbers ofcells tuned to various orientations in the striate cortex.The noise might, however, be a result of a narrowerpooling of information in the vertical relative to thehorizontal direction, possibly at a stage of stereoscopicprocessing beyond that represented by single disparity-tuned units of striate cortex.
REFERENCES
Barlow, H. B., Blakemore, C. & Pettigrew, J. D. (1967). The neuralmechanism of binocular depth discrimination. Journal of Physiol-ogy, 193, 327–342.
Bishop, P. O. (1989) Vertical disparity, egocentric distance andstereoscopic depth constancy: A new interpretation. Proceedtrrgsofthe Royal SocieQ of LondonB, 237, 445-469.
Cormack, L. K., Stevenson, S. B. & Schor, C. M. (1991). Interocularcorrelation, luminance contrast and cyclopean processing. VisionResearch, 31, 2195–2207.
Cormack, L. K., Stevenson, S. B. & Schor, C. M. (1994). An upperlimit to the binocular combination of stimuli. VisionResearch, 34,2599-2608.
DeAngelis, G. C., Ohzawa, I. & Freeman, R. D. (1991). Depth isencoded in the visual cortex by a specialized receptive fieldstmcture. Nature, 352, 15&159.
DeValois, R. L., Yund, E. W. & Hepler, N. (1982). The orientation anddirection selectivity of cells in macaque visual cortex. VisiorIResearch, 22, 532–544.
Erkelens, C. J. (1987). Adaptation of ocular vergence to stimulationwith large disparities. ExperimentalBrain Research, 66, 507–516.
Fender, D. & Julesz, B. (1967). Extension of Panum’s fusional area inbinocularly stabilized vision. Journal of the Optical Society ofAmerica, 57, 819+30.
Ferster, D. (1981). A comparison of binocular depth mechanisms in
area:s 17 and 18 of the cat visual cortex. Journal of Physiology,311,623--655.
Geisler, W. S. & Albrecht, D. G. (1995). Bayesian analysis ofidentification performance in monkey visual cortex: Nonlinearmechanisms and stimulus uncertainty. VisionResearch, 35, 2723–2730.
Joshua, D. E. & Bishop, P. O. (1970). Binocular single vision anddepth discrimination: Receptive field disparities for central andperipheral vision and binocular interaction on peripheral single unitsin cat striate cortex. ExploratoryBrain Research, 10, 391416.
LeVay, S. & Voight, T. (1988). Ocular dominance and disparity codingin ca( visual cortex. VisualNeuroscience, 1, 395414.
Mansfield, J. S. & Parker, A. J. (1993). An orientation-tunedcomponent in the contrast masking of stereopsis. VisionResearch,33, 1.535-1544,
Mayhew, J. E. W. & Frisby, J. P. (1978). Stereopsis masking is notorientationa]ly tuned. Perception, 7, 431-436.
Nikara, T., Bishop, P. O. & Pettigrew, J. D. (1968). Analysis of retinalcorrespondence by studying receptive fields of binocular single unitsin cat striate cortex. ExperimentalBrain Research, 6, 353–372.
Perlmutter, A. L. & Kertez, A. E. (1978). Measurement of humanvertical fusional response. VisionResearch, 18, 219–223.
Pettigrew, J. D., Nikara T. & Bishop, P. O. (1968). Binocularinteraction on single units in cat striate cortex: Simultaneousstimulation by single moving slit with receptive fields incorrespondence. ExperimentalBrain Research, 6, 391JI1O.
Press, ‘W.H., Flannery, B. P., Teukolsky, S. A. and Vetterling, W. T.(1988). Numericalrecipes in C: Theart of scientificcomputing.NewYork: Cambridge University Press.
Schor, C. M. & Tyler, C. W. (1981). Spatio-temporal properties ofPanulm’s fusional area. VisionResearch, 21, 683+92.
Stevenson, S. B. & Schor, C. M. (1993). Human responses tointerocular correlation on the vertical disparity axis. InvestigativeOphthalmologyand VisualScience (suppl.),34, 1439.
Stevenson, S. B. & Schor, C. M. (1994). Horizontal and verticaldisparity range of matching and stereoscopic processes. hrvestiga-tive t9phthalmo[ogyand VisualScience (suppl.),35, 1916.
Swanson, W. H. & Birch, E. E. (1992). Extracting thresholds fromnoisy psychophysical data. PerceptionandPsychophysics,51, 409–422.
Tolhur:st, D. J., Movshon, J. A. & Dean, A. F. (1983). The statisticalreliability of signals in single neurons in cat and monkey visualcortex. VisionResearch, 23, 775–785.
Tyler, C. W. (1977). Stereomovement from interocular delay indynamic visual noise: A random spatial disparity hypothesis.American Journal of Optometry and Physiological Optics, 54,374-386.
Tyler, C. W. (1983) Sensory processing of binocular disparity. InSchclr, C. M. & Ciuffreda, K. J. (Eds), Vergence eye movements:Basi,cand clinical aspects (pp. 199–295). London: Butterworths.
Tyler, C. W. & Julesz, B. (1978). Binocular cross-correlation in timeand !space. VisionResearch, 18, 101–105.
Volkman, A. W. (1859). Die stereoskoopischen erscheinungen in ihrerbezichung zu der Iehre von den identischen netzhautpunkten. Arch.f Ophth., 5, 1-100. As cited in Ogle, K. N. (1950). Researches inbinocularvision. Philadelphia: WB Saunders.
von der Heydt, R., Adorjani, C., Hanny, P. & Baumgartner, G. (1978).Disparity sensitivity and receptive field incongruity of units in thecat striate cortex. ExperimentalBrain Research, 31, 523–545.
Acknowledgements-This work was supported by NIH/NEI EY103O3and AFOSR F49620-93-1-0307. The authors wish to thank Drs DuaneG. Albrecht, Gregory C. DeAngelis and an anonymous reviewer fortheir very helpful comments on the manuscript.