Embed Size (px)
Citation preview

(Aus dem Zoologischen Institut der Universit~t Miinster i. W.)
BEITR XGE ZUR P H Y S I O L O G I E DES HXMOGLOBINS W I R B E L L O S E R TIERE.
I. VERSUCHE UBEt~ DIE ATMUNG ~NORMALER UND MIT KOHLENOXYD VERGIFTETER REGENWURMER BEI TEMPERATUREN UM 200 C.
Von
FRIEDRIGH KR~J'GER.
Mit 4 Textabbildungen.
(Eingegangen am 8. April 1940.)
Den Ausgangpunkt zu vorliegender Untersuchung stellte die eigen- artige, yon THOMAS (1935) beschriebene Form der Abhangigkeit der Atmung des Regenwurmes vom Sauerstoffdruck dar. Durch Beriiek- siehtigung der Kohlensaureabgabe hoffte ich, die Versuche yon THO~_~s erganzen zu kSnnen. Es ergab sich dabei allerdings, dab ich seine Be- funde nicht bestatigen konnte.
THomAs stellte die Sauerstoffdruckabhangigkeit der Regenwurm- atmung so dar, dab zwischen 21% und 15% Sauerstoffgehalt die A~mung mit sinkender Konzentration sehr steil auf 40% fallt, zwischen 15% und 3% konstant bleibt, um erst bei weiterer Verminderung der Sauer- stoffspannung wieder stark abzusinken. T]zo~As wiederum konnte nicht die Ergebnisse bestatigen, die DOLK und van der Paauw (1929) an narko- tisierten Wiirmern erha]ten batten. Nach deren Angabe soll der Sauer- stoffverbraueh zwischen 21% und 2,5% konstant bleiben und erst unterhalb letzterer Konzentration absinken. I m Gegensatz hierzu standen sehliel31ieh die alteren Angaben yon KONOPACKI (1907), der feststellte, dal] der Sauerstoffverbrauch bei allen Konzentrationen vom Sauerstoffdruck abhangig ist, wenn auch zwischen 21% und 10% weniger stark als bei Konzentrationen unter 10 %.
Ieh habe bier zunachst nut die Verhaltnisse bei normalen Tieren beriicksichtigt. In neuerer Zeit sind daneben Versuche durchgeffihrt worden, die bezweekten, durch Vergiftung mit Kohlenoxyd die Funktion des Hamoglobins als Trager des Sauerstoffes auszusehalten und auf diese Weise Einblick in die Bedeutung des roten Blutfarbstoffes beim Regenwurm zu gewinnen. Die von DOLK und v ~ DER I:~AAUW, sowie yon THOMAS durehgeffihrten Versuehe lieferten ebenfalls kein einheit- liches Bild. Erstere, die wie erwahnt, mit narkotisierten Tieren arbei- teten, fanden nach Vergiftung mit Kohlenoxyd als wiehtigsten Unter- schied den, dab bei ihnen das Abfallen der Atmung schon bei etwa 7% Sauerstoff einsetzt, im iibrigen beobachteten sie keinen Unter- schied gegenfiber normalen Tieren. Hieraus schlieBen sie - - wie aueh

44 Friedrich Kriiger:
schon yon Jo~DA~ und SCHWA~Z (1920) gei~uftert wurde - - , daft da s I-I~moglobin nur bei n iederen Sauers to f fkonzen t ra t ionen in F u n k t i o n t r i t t , oberhalb 7 % abe t die Versorgung der Gewebe ausschlieftlich durch physikal isch gelSsten Sauerstoff erfolgt. THOMAS g laub t dagegen bei der sehr s t a rken S t reuung ihrer W e r t e keinen sicheren Unte rsch ied zwischen den normalen und CO-vergif te ten Tieren fests tel len zu kSnnen. Seine eigenen Messungen erweisen ffir die CO-vergif te ten Wi i rmer eine deut l iche Unter legenhei t gegenfiber normalen Tieren bei der Sauerstoff- konzen t ra t ion der Luft . Dagegen is~ un te rha lb 15% Sauerstoff die A t m u n g normale r und vergifteber Wfi rmer gleich grol). Nach seiner Ansicht l~ftt sich nichts Sicheres fiber die Leis tung des H~moglobins auf Grund der vor l iegenden Versuche aussagen.
Die offensichtl ichen Widersprf iche und Unsicherhei ten, die bezfiglich der A t m u n g des Regenwurmes bestehen, rechtfer t igen eine Nachprf i fung und Erg/ inzung der vor l iegenden Angaben.
Versuchstechnik.
Die gesamten Atmungsmessungen ffihrte ich mi t I-Iilfe des yon m i r beschriebenen Mikrorespi rometers aus, der eine gleichzeitige Be s t immung yon Sauers tof fverbrauch und Kohlensi~ureabgabe ges ta t te t . Den im prinzipiel len Aufbau unver~nder ten A p p a r a t babe ich inzwischen in einigen technischen Einze]hei ten wesentl ich verbessern k6nnen.
1. Die wichtigste Verbesserung ist die, da~ ich den Apparat ohne Gummi- verbindungen herstellte. I)ieses geschah einmal dadurch, daI~ ich das Manometer an den Respirationsapparat, sowie an das Kompensationsgef~B und das MeBrohr mit Hilie yon Schliffstficken ansehloB, die durch Federn oder Gummibiinder zu- sammengepreBt werden. Urn mSglichst spannungslosen Zusammenhalt der Schliff- stiicke zu erreichen, wurden nach dem endgiiltigen Zusammenbau des ganzen Apparates die Zuleitungen zu den Schliffen an geeigneter Stelle bis zum Erweichen des Glases erhitzt, bis sich alle Spannungen ausgeglichen haben und dann langsam abgekfihlt. Durch die Schliffstficke ist im Bedarfsfall das manometer leicht ab- zutrennen, was ~r allem beim Reinigen des Apparates yon grS~tem Vorteil ist~ Es erfibrigte sich dadurch auch der Hahn, der urspriinglich zur Fiillung und Reini- gung des manometers vorgesehen war. Da ferner durch die Anbringung der Schliffe die mechanische Beanspruchung des ganzen Apparates bei der yon Zeit zu Zeit notwendigen Reinigung, bei der die Gummiverbindungen ge15st und sparer wieder zusammengesetz~ werden mul3ten, herabgesetzt war, konnte ich auch Pumpe und Objektbeh~lter fest~ aneinander schweil3en lassen. Dadurch kSnnen auch diese Teile leicht mit Chromsehwefelsi~ure gereinigt und mit Alkoho] und _~ther getrocknet werden. Der Fortfall der Gummiverbindungen zeitigte noch den Vorteil grSl~erer Volumenkonstanz, denn es hatte sich erwiesen, da~ diese nach dem Zusammen- setzen des Apparates meist noch mehrere Tage arbeiten, ehe alle Spannungen sich ausgeglichen haben und Volumenkonstanz eingetreten ist. Eine weitere Ver- meidung zu starker mechanischer Beanspruchung bedeutete es fernerhin, dab ich
2. die aus den beiden Dreiwegeh~hnen, sowie aus dem Kompensationsgef~B naeh auBen fiihrenden und fiber den Spiegel des Wassers im Thermostaten heraus- ragenden Glasrohre kurz oberhalb der H~hne abschnitt und entsprechend lange Stiieke Glasrohr elastisch mit einem Stiickchen Gummischlauch ansetzte.

Beitr~ige zur Physiologie des HAmoglobins wirbelloser Tiere. I. 45
3. Am GlasstSpsel des Objektbeh~lters sowie an diesem selbst liel] ich je ein Paar Glashaken anbringen, die gestatten, mit Gummib~ndern den StSpsel lest einzupressen; die ohne diese Vorsiehtsmal3regel mSgliehe Volumen~nderung des Apparates wird dadurch vermieden.
4. Maehten reich die Versuche am Regenwurm auf eine Mal]nahme aufmerk- sam, die zur Kohlens~urebestimmung notwendig ist. Legt man n~mlich die Wiirmer einfaeh auf den Boden des Objektbehi~lters, so 15st sich anseheinend ein Teil der Kohlens~ure im Tierk0rper und ent0geht der Messung; man erh~lt so zu niedrige respi- r~torisehe Quotienten. Diese Gefahr ist sehr einfach dadurch zu umgehen, dal] man die Versuchsobjekte etwas erhSht in dem Atmungsgef~13 unterbringt. Man kann das einmal dadureh erreiehen, dal~ man in einiger Entfernung fiber dem Boden eine durchlSeherte Glas- oder Porzellanplatte anbringt, auI die man das Objekt legt, oder indem man das Objekt in einem Gazebeutel au~h~ngt. Um beiden MSglieh- keiten leicht gereeht werden zu k6nnen, habe ich bei einem neuen Apparat den Tierbeh~Llter b~rnenf6rmig gestaltet, so dal3 ein rundes Sieb entspreehender GrSl3e leicht in einiger Entfernung fiber dem Boden aufgelegt werden kann, aul3er- dem habe ich den Glasst6psel des Tierbeh~lters mit einem Glashaken versehen lassen, an dem man einen Gazebeutel aufhi~ngen kann.
5. Eine Verbesserung erfuhr aul3erdem die Gestalt des Kompensationsgef~13es, das ieh frfiher in GrSl3e des Objektbeh~lters ausffihrte. In diesem war das Ver- h~ltnis zwisehen Gasraum und Fliissigkeit ein anderes, als im eigentlichen AtmUngs- apparat, was bei Temperaturi~nderungen zu S~Srungen ffihren kann. Eine bessere A~m~herung an die im Atmungsteil herrschenden Verh~ltnisse glaubte ieh dadurch herzustellen, dab ieh dem Kompensationsgef~ eine T-fSrmige Gestalt gab, be- stehend aus einem liingeren horizontalen Tell, der in seinem Volumen etwa dem der Pumpe entspricht und auch eine ~hnliche Flfissigkeitsffillung aufnehmen kann und einem in der Mitre senkreeht darauf aufgesetzten Teil, der etwa dem Ob- jektbeh~lter zuzfiglich der Leitungen entspricht und yon dem der Ansehlul3 zum Manometer und der Hahn des Kompensationsgef~es ausgehen. Man ftillt den horizontalen Tell des Kompensationsgefi~13es nieht ganz mit Fliissigkeit, beim Sehaukeln des Apparates gest~ttet die gro~e FlfissigkeitsoberflAehe in Verbindung mit dem Hin- und HerflieBen einen schnellen Austauseh zwischen der Flfissig- keit und der Gasphase.
Die Abb. 1 mSge ein Bild geben von dem Aussehen eines naeh diesen Ges ieh tspunkten kons t ru ie r ten Appara tes . Aus ihr geht die gegen- seitige Anordnung der Teile, sowie die Vers gegenfiber der urspr i ingl ichen F o r m des Appa ra t e s hervor, die lediglich den Glastei l des Appa ra t e s betreffen, wAhrend der Metal l rahmen, sowie die Antr iebs- vor r ich tung unver~Lndert geblieben sind.
Was die im vorl iegenden Fa l le angewandte Unte rsuchungs technik ~nbetr iff t , so hube ich, wie TI~O~AS und die anderen Autoren, mi t hungern- den Wfi rmern gearbei te t , die auf feuehtem Fliel~p~pier gehal ten wurden. Die Messungen wurden stets an einze]nen Wt i rmern durchgefi ihr t , in der Regel wurde jeder W u r m zu mehreren Versuchsserien herangezogen. Zumeis t h~be ieh die Wfi rmer im Gazesack aufgehAngt, da sie sich versehiedent l ich so eng auf dem Sieb des Atmungsbeh~tlters zusammen- r ingel ten, dal] eine Luf tz i rku la t ion und d a m i t Messungen unmSglich wurden. Die SAekehen wurden zur Desinfekt ion in Alkohol aufbewahr t , um StSrungen durch Bakte r ien zu vermeiden. Die Ss wurden jeweils nur zu einer Versuchsserie gebraucht , naehdem sie nat i i r l ich

46 Friedrich Kriiger:
zuvor sorgf~ltig getrocknet waren. In der Zeit zwischen den Versuchen wurden die Wfirmer in Glasschalen im Dunkeln aufbewahrt.
Auf die Einhaltung yon Standardbedingungen verzichtete ich im Anschlul~ an die Erfahrungen von THOMAS, da mir durch die Anwendung der Narkose nur die Gefahr unfibersehbarer Komplikationen gegeben erschien. Ich achtete darauf, die Wtirmer beim Einbringen in die Gaze-
s~ckchen mSglichst wcnig anzufassen und zu reizen. Es zeigte sich, dab die Wiir- mer einige Zeit naeh dem Einsetzen in den Apparat ziemlich gleichm~l~ig atme- ten, wie ich welter unten bespreehen werde.
Ffir die Herstellung der Gasgemisehe stand mir Bombenstickstoff mit einem Sauerstoffgehalt yon 5 % zur Verifigung. Aus diesem Grunde konnte ich niedri- gere Sauerstoffkonzentra- tionen nicht herstellen und beschri~nkte reich aul]er der Sauerstoffkonzentration der Luft auf die Stufen yon 15%, 10% und 5%. Ftir die reich im Augenblick in- teressierenden Fragen er- schien dieser Bereich genii-
Abb. 1. A n s i e h t des R e s p i r a t i o n s a p p a r a t e s . . 4 0 b - g e n d . Das Kohlenoxyd be- jektbehi~lter , B P u m p e , C K o m p e n s a t i o n s g e f a B , reitete ich aus Ameisen- D Mei~rohr, a P o r z e l l a n - S i e b p l a t t e im Ob jek tbehh l t e r , b S c h l i f f v e r b i n d u n g e n z u m M a n o m e t e r m i t Fede rn , s ~ u r e z u der konzentrierte c G u m m i v e r b i n d u n g e n d e r A u s l e i t u n g s r o h r e , d ~ I i v e a u - Schwefels~ure getropft wur-
gef~B des ]VIe[~rohres, e D r e h l a g e r fi ir die Sehaukelbewegung. de. Ich bewahrte es in
einem kleinen l%ezipienten auf. Abweichend yon THOMAS und DOLE und vA~ D~R PAAVW leitete ich die Gasgemische mit den verschieclenen Sauerstoffkonzentrationen in den Atnmngsapparat ein. Die Wiirmer blieben ws der ganzen Versuchszeit unbehelligt im Apparat liegen. Die Zahlenangaben fiir den Gasstoffwechsel beziehen sicb auf feuchtigkeitsgess Gas yon Zimmertemperatur je 30 Minuten.
In den ersten Versuehen, die ich zum Teil in Gemeinschaft mit Herrn T~oMAs am Inst i tut ffir vergleichende Physiologie zu Utrecht als Stipendiat der Rockefeller-Foundation durchitihrte, benutzte ich

Beitrgge zur Physiologie des Hgmoglobins wirbelloser Tiere. I. 47
Temperaturen von etwa 250 (~, die durch einen Thermoregulator konstant erhalten wurde. In den spgteren Versuchen arbeitete ich im ungeheizten Thermostaten yon Zimmertemperatur. Bei entsprechender Regulierung der Zimmertemperatur lie]]en sich genfigend konstante Temperaturen erzielen. Allerdings liel~en sich auf diese Weise keine bestimmten Wiirme- grade einstellen, diese schwankten daher in den spgteren Versuchen erheblich mit der Umgebungstemperatur.
Die grSgte Schwierigkeit bereitete die Verarbeitung des, wie wir sehen werden, sehr stark schwankenden Zahlenmaterials. Als beste Liisung ergab sieh die Benutzung des arithmetischen Mittels, das in der Regel aueh gut der allgemeinen Verteilung der Werte entspricht. Von der Wiedergabe der mittleren quadratischen Abweichung habe ich abgesehen. Diese setzt einen Mittelwert voraus, um den die Werte entsprechend der Wahrscheinlichkeit sehwanken. Diese Voraussetzung trifft fiir die Atmung des Regenwurmes nicht zu. Durch ihre Abhgngig- keit yon mannigfachen Faktoren zeigt sie eine kompliziertere Vertei- lung der Werte. In dieser Abhgngigkeit der Atmung von verschiedenen Faktoren liegt die Sehwierigkeit, fiberhaupt zu eindeutigen Zahlen zu kommen. Ich bin daher so vorgegangen, dal~ ieh stets zungchst die Atmung in Luft als grundsi~tzlichen Mal~stab festlegte und auf ihre GrSge die Abweichungen bezog, die durch vergnderte Bedingungen hervorgerufen wurden. Da so in den einzelnen Versuchsreihen Werte erhalten wurden, die in ihrem relativen Verh~tltnis zur Atmung in Luft ein m6glichst getreues Bild der vorliegenden Abhi~ngigkeit geben, mug dieses auch fiir die Mittelwerte aus allen Versuchen gelten, unabhgngig yon der absoluten GrSl]e des Ausgangswertes.
Die Yersuchsergebnisse.
a) Die Konstanz der Atmung nicht narkotisierter Wi~rmer.
Da ich auf die Anwendung yon Narkosemitteln zur Herstellung yon Standardbedingungen verzichtete, andererseits die Natur der Ver- suche sehr lang dauernde Messungen verlangte, ergab sich die Frage, wieweit man unter den gegebenen Umstgnden eine gleichmggige Atmung der Wfirmer voraussetzen daft und wieweit die Atmung an aufeinander- folgenden Tagen vergleichbar ist.
Zur Er5rterung dieser Frage habe ich in Tabelle 1 s~tmtliche yon dem Wurm E i m Laufe yon mehr als 6 Wochen bei Atmung in Luft erhaltenen Werte zusammengestellt. Es sind hierbei auch Zahlen aus den Tabellen 2 und 4 enthalten, darfiber hinausgehend noch solche Mel~ergebnisse, die yon abgebrochenen Versuchen stammen oder die in diesen Tabellen keinen Platz finden konnten. Zum Teil sind diese Werte in Einzelversuchen erhalten, die sich fiber eine wesent- lich lgngere Dauer erstreckten, als die im allgemeinen hierfiir einge-

48 Friedrich Krfiger:
haltene Spanne yon etwa 30 Min. In solchen F/~llen wurde zumeist auf die Bestimmung der Kohlens/~ureabgabe verzichtet, da sie unter diesen Umst/~nden nieht unbedingt zuverlassig ist und die Anh~ufung des CO 2 im AtmungsgefiiB zu St6rungen frihren kann. Da nicht alle Versuche unter gleichm~ltigen Bedingungen fortgeffihrt wurden, ist die Dauer der einzelnen MeBserien, w/~hrend der die W/irmer vollkommen unbehelligt im Apparat liegen blieben, nicht gleichm/~6ig. Bei jeder Reihe ist die Zeit vermerkt, welche vom Beginn der ersten bis zum Ende der ]etzten Messung vergangen war. Bei sehr lange dauernden Versuchen wurde die Luft im Apparat gewechselt. Ich begniige mich mit der Wiedergabe der an einem Wurm gewonnenen Zahlen, da weitere Beispiele keine grunds/~tzlieh neuen Erkenntnisse bieten kSnnten.
Betraehten wir zun/~chst den Sauerstoffverbrauch unseres Beispieles, so kSnnen wir erkennen, dab in sehr vielen Fallen die ersten Werte einer Reihe betr~chtlich hSher liegen als die folgenden, so z. B. bei den Ver- suchen vom 31. Juli und vom 3. und 6. August. Es liegt nahe, diese Erscheinung auf die Erregung zurfickzuf/ihren, der die Wiirmer beim Einsetzen in den Apparat ausgesetzt sind. Da mit den Messungen stets erst etwa 1 Stunde nach dem Einsetzen begonnen wurde, kann man erkennen, wie lange sich die Hantierungen mit dem Wurm auf die GrSBe seines Stoffumsatzes auswirkten. Ich habe daher in den sp~teren Versuehen regelm/~Big erst nach etwa 2 Stunden mit den Messungen begonnen, um diese Nachwirkung abklingen zu lassen. I m Gegensatz hierzu fanden sich zum Beginn mancher Versuche auch verminderte Werte ffir den Sauerstoffverbrauch. Die Versuehe vom 17. und 22. September bieten Beispiele hierffir.
Die auf die abweichenden Anfangswerte folgenden Messungen zeigen dagegen wesentlich bessere Konstanz. Wenn st/~rker aus dem Rahmen herausfallende Werte auftreten, sind sie in der Regel gefolgt yon solchen in entgegengesetzter l~ichtung, so dab Mittelwerte ein zuverl/~ssiges Bild geben.
Auf Grund dieser Erfahrungen bin ich bei meinen Versuchen so vor- gegangen, dab ich wartete, bis 2 oder 3 oder auch 4 aufeinandeffolgende Messungen mSgliehst konstant blieben und habe dann erst mit vers ten Versuchsbedingungen gearbeitet. DaB man auch bei nicht narkoti- sierten Wfirmern mit einiger Sieherheit fiir 1/~ngere Zeit mit konstanter Atmung rechnen kann, zeigen etwa die Versuche vom 6. 8. und vom 14, 16., 21. und 22. 9. Jedenfalls sind die auftretenden Schwankungen nieht grSBer, als sie :DoLK und VAN ])~R PAAVW bei narkotisierten Tieren fanden.
Die oft beobaehteten hohen Anfangswerte fiir den Sauerstoffver- braueh haben mir den Gedanken nahegelegt, hierin die Ursache ffir die stark abweiehenden Ergebnisse von THOMAS ZU sehen. THOMAS spricht davon, dab er die Wrirmer in Trill einn/~hte (1935a). Dieses wfirde natur-

Beitri~ge zur Physiologie des H~moglobins wirbelloser Tiere. I. 49
Datum
30.7.
31.7.
3.8.
6.8.
7.8.
9.8.
13.9. 14.9.
15.9.
16.9.
17.9.
21.9.
22.9.
Versuchs- ] dauer Tempe- in Sttmden ratur
41/2
7~/~
3
51/~ 7
5
6~/~
Tabel le 1.
170
18 o
200
210
230
250
16 0 16 0
15 0
150
150
17 o
19 o
O~-Verbr. CO~-Abg. RQ O~-Verbr. C02-Abg. RQ O~-Verbr. CO~-Abg. RQ 02-Verbr. C02-Abg. RQ 02-Verbr. C02-Abg. RQ O~-Verbr. C02-Abg. RQ 02-Verbr. O~-Verbr. C02-Abg. RQ 02-Verbr. C02-Abg. RQ 02-Verbr. CO~-Abg. RQ 02-Verbr. C02-Abg. RQ 02-Verbr. CO 2-Verbr. RQ O2-Verbr. CO~-Abg. RQ
MeBergebnisse
111 100 109 91 91 72
0,82 0,91 0,66 97 82 72 76 87 72 60 65
0,90 0,88 0,86 0,86 87 60 59 51 49 97 64 63 47 46 1,12 1,07 1,07 0,92 0,95 111 76 78 61 75 51 58 59
0,46 0,77 0,76 93 88 80 80 77 99 88 74 70 63
1,07 1,00 0,93 0,87 0,82 113 115 80 87
0,78 0,76 28 26 36 66 52 63 44 - - 57
0,67 - - 0,89 41 46 47 27 33 30
0,66 0,72 0,64 63 55 64 58 55 40 15 48 35
0,64 0,30 0,75 0,60 58 65 60 60 40 41 34 - -
0,69 0,63 0,57 - - 75 64 68 38 - - 37
0,51 - - 0,55 76 s2 s7 s6 45 48
0,59 0,59
50 48
gem/~$ eine sehr starke Reizung darstellen, deren Auswirkung einige Zeit andauer t . Die yon ibm angewandte Technik er laubte eine Pr i i fung der Kons tanz der A t m u n g zu Beginn tier Messungen nicht. Es ist n icht zu erkennen, wie lange Zeit nach dem Einsetzen der Wiirmer in den Atmungsappara t mi t den Versuchen begonnen wurde. T ~ o ~ s spr~cht abet nicht yon besonderen VorsiehtsmaSnahmen, so da$ m a n a n n e h m e n daft, alas er sehr schnell nach dem Einn/~hen der Wiirmer mi t den Messungen angefangen hat . Die zun~chst sehr hohen und alarm schnell abs inkenden Werte fiir den Sauerstoffverbrauch, die den steilen Abfall
Z. f. vergl. Physiologie. Bd. 28. 4

50 Yriedrich Krfiger:
seiner Atmungskurve zwischen 21% und 15% Sauerstoff bewirken, wi~ren demnach der Ausdruck der abklingenden Erregung der Wfirmer beim Einbringen in den Atmungsapparat.
Der Vergleich der AtmungsgrSBe an verschiedenen Tagen ist wegen der verschiedenen angewandten Temperaturen bei dem in Tabelle 1 besprochenen Beispiel nicht ganz leicht. Aber auch in meinen ersten Versuchen, bei denen ich mit einer konstanten Temperatur yon 25 o arbeitete, fand ich Unterschiede im Sauerstoffverbrauch an verschie- denen Tagen. Ich glaubte zun~chst an einen schi~digenden EinfluB der ziemlich hohen Temperatur. Demgegenfiber zeigen die Versuche vom 15, 16., und 17.9., dab auch bei niederen Temperaturen die Atmungs- grSBe nicht identisch zu sein braucht. Es ist also die Atmungs- grSBe durch verschiedene zum Tell unkontrollierbare Faktoren beein- fluBbar, so z. B. durch die Dauer des Hungerzustandes, wie schon GI]~sc~EN beobachtete. In den ersten Versuchstagen war der Sauer- stoffverbrauch nennenswert hSher als im September, auch wenn man yon der niederen Temperatur absieht. Einen anderen Faktor stellt der Wassergehalt dar. Am 13. 9. wurde der Wurm nach l~ngerer Pause wieder zu Versuchen herangezogen. Dutch Austrocknung seines SchM- chens hatte er Wasser verloren und als Folge ergab sich eine betr~cht- lich herabgesetzte Atmung, die abet nach dem Ersatz der Flfissigkeit schon am n~chsten Tage eine normale HShe erreicht hatte. Entgegen den Schwankungen des Sauerstoffverbrauches ffir die einzelnen Tage, stellte er sich innerhalb einer Versuchsserie auf eine gewisse GrSBe ein, die auch erhalten blieb.
Zu erw~hnen ist noch, dab bei zweien meiner Wiirmer in einzelnen Versuchen kein Sauerstoffverbrauch gefunden wurde, sondern eine dariiber noch hinausgehende Gasabscheidung. Eine solche wurde auch yon TEO~AS und DOLK und VAN DER PAAVW beobachtet. Ob die Aus- scheidung dieses Gases, dessen Natur noch unbekannt ist, allen Wiirmern zukommt, vermag ich nicht zu sagen. In diesem Falle wiirden die fiir den Sauerstoffverbrauch ge~undenen Werte in Wirklichkeit hSher sein als sie gefunden wurden. Dagegen wiirde die Kohlens/~urebestimmung durch die Gasabscheidung nicht beeinfluBt werden und daher die respira- torischen Quotienten etwas niedriger anzunehmen sein. Die Wiirmer, bei denen Gasabscheidung gefunden wurde und die zudem sehr unregel- m~Bige Werte lieferten, wurden nicht zu den Versuchen herangezogen. Ich glaube nicht, dab bei den iibrigen Wiirmern die Gasabscheidung ein solches Ausma6 erreicht, dab hierdurch die Versuchsergebnisse grund- s/~tzlich beeinfluBt werden.
Die Kohlens~ureabscheidung bietet etwa dasselbe Bild wie der Sauerstoffverbrauch. Nach einigen abweiehenden Anfangswerten folgen im allgemeinen konstante weitere Zahlen. Auch hier finden sich - - z. B. am 6. und 7. 8. des Beispieles - - erhebliche Untersehiede in der

Beitr~ge zur Physiologie des Hi~moglobins wirbelloser Tiere. I. 51
T a b e l l e 2.
Versuchs- Bezeich-
In cram hung je 30 rain. Datum
Temperatur
a 02-Verbr. 19 .9 .33 C02-Abg.
250 C c O2-Verbr.
21 .9 .33 CO~-Abg. 250 C
d 02-Verbr. 22 .9 .33 CO~-Abg.
25 o C f 02-Verbr.
25 .9 .33 CO~-Abg. 250 C
h O~-Verbr. 27 .9 .33 C02-Abg.
250 C i 02-Verbr.
28 .9 .33 C02-Abg. 250 C
n 3 O~-Verbr. 25 .2 .33 C02-Abg.
250 C 02 O2-Verbr.
13.2- 35 C02-Abg. 25 o C
o 3 02-Verbr. 14. 2.35 C02-Abg.
25 o C A a 02-Verbr.
17.7.37 CO2-Abg. 210 C
A T O~-Verbr. 22. 7.37 C02-Abg.
220 C B 2 O~-Verbr.
15.7.37 C02-Abg. 22 o C
B a 02-Verbr. 21 .7 .37 C02-Abg.
22 o C E~ 02-Verbr.
30 .7 .37 C02-Abg. 170 C
15.E~137 O2-Verbr" CO~-Abg.
15 ~ C E12 02-Verbr.
16.9 .37 CO~-Abg. 15 ~ C
E13 O2-Verbr. 17.9.37 CO~-Abg.
15oc F 1 02-Verbr.
30. 7~ 37 CO~-Abg. 17oc
G 02-Verbr. 8 .10.37 C02-Abg.
19oc
Atmosph~ri- sche Luft 15 % O~ 10 % O~ 5 % 02
1 2 3 1 1 2 1 3 1 1 2 3 1 1
58 4o
39 32
27 31
2 3
38 36 27 26
48 45
16 27 24 31
27 24 37
42
37 25
29 25 14 12
4-*

52 Friedrich Kriiger:
GrSl~e der Kohlensgureabgabe und des R Q an aufeinanderfolgenden Tagen, fiir die eine Erldgrung nicht gegeben werden kann.
Der respiratorische Quotient ist bei ffisChen Wiirmern hSher als bei l~nger hungernden, bei denen er sich dem Wert 0,6 n~hert. Eine Tat- saehe, die auch schon von GIESCHE~ beschrieben wird. Da ich sowohl mit sehr frischen als auch mit lange hungernden Tieren arbeitete, land ich sehr stark schwankende Werte fiir den t~ Q, die bei dem in Tabelle 1 betrachteten Beispiel zwischen 0,3 und 1,12 lagen. Wenn die friiheren Autoren in engeren Grenzen liegende Quotienten fanden, so hat das seine Ursache darin, dab sie ihre Bestimmungen nicht mit Einzeltieren, sondern mit mehreren Wiirmern durchfiihrten, wodurch solche nur ver- einzelt auftretende Ausnahmewerte ausgeglichen werden. Im iibrigen stimmen die yon mir gefundenen Quotienten grSBenordnungsweise gut mit denen der friiheren Autoren iiberein. Eine eindeutige Festlegung der GrSBe von R Q ffir den Regenwurm erscheint nach meinen Beob- achtungen kaum mSglich, so dab ich darauf verzichtete.
Die geschilderten Erfahrungen zeigen, dab die im Atmungsgef~B befindlichen Wiirmer nach dem Abklingen der ersten Erregung eine ziemlich gleichms Atmung zeigen, so dab man auf die Anwendung der Narkose bei ihnen verzichten kann. Diese schlieBt nur die Gefahr ein, uniibersehbare physiologische Xnderungen auszulSsen. Sauerstoff- aufnahme nnd Kohlens~ureabgabe stellen auch fiir den gleichen Wurm und bei gleiehen Bedingungen keine konstanten GrSBen dar. ,Sie bleiben nur gleichmgBig bei dem unbehelligt im Respirationsapparat liegenden Tier. Vergleichbare Werte lassen sich daher nur unter diesen Bedin- gungen, d. h. unter AussehluB s Reize erhalten. Die im folgenden wiedergegebenen Versuche zeigen die MSglichkeit, unter diesen Umst~nden zu zuverlS, ssigen und reproduzierbaren Ergebnissen zu kommen.
b) Die Abhiingigl~eit der Atmung des Regenwurmes yon der Sauersto]/- konzentration .
Die Frage nach der Abh~ngigkeit der Atmung yon der Sauerstoff- spannung ist fiir den Regenwurm auch yon 5kologisehem Interesse, da seine unterirdische Lebensweise ihn ohne Zweifel zwingt, zeitweilig mit geringeren Sauerstoffspannungen vorlieb zu nehmen, als sie der Mehrzahl der Tiere in der freien Atmosphs zur Verfiigung steht. Wie ich aber einleitend bemerkte, bestand hieriiber bislang in keiner Weise Ubereinstimmung, so dal] sieh die N0twendigkeit neuer Messungen ergab.
Die hierbei yon mir erhaltenen Werte sind in der Tabel]e 2 zusammen- gestellt. Die einzelnen benutzten Wfirmer sind durch Buchstaben ge- kennzeichnet. Soweit mehrere Versuche mit einzelnen Wiirmern vor- liegen, sind diese durch an die Buchstaben angeh~ngte Ziffern unter- schieden. Da die zum Teil recht unterschiedlichen AtmungsgrSBen, sowie

Beitr/~ge zur Physiologie des tt~moglobins wirbelloser Tiere. I. 53
die Vielzahl der in jeder Stufe erhaltenen Werte den Uberblick fiber die Ergebnisse ersehweren, habe ieh in Tabelle 3 das gleiehe Zahlenmaterial in der Weise verarbeitet, dag ieh die in jeder Stufe erhaltenen Ergebnisse in Durehsehnittswerten zusammengefagt habe und fiberdies zu einer weiteren Erleiehterung des Vergleiehes in allen Megreihen die Atmung in atmosph~riseher Luft gleieh 100 setzte und die Atmung bei den fibrigen Sauerstoffkonzentrationen in Prozent der Atmung in Luft berechnete. Augerdem habe ieh in dieser Tabelle nieht die einfachen Kohlensgurewerte aufgenommen, sondern zur Kennzeiehnung der Kohlensgureabgabe reich des respiratorisehen Quotienten bedient. Hier- dureh wird weiterhin der Vergleieh der Versuehe erleiehtert.
Die Kl/~rung der ges~ellten Frage wurde dadureh auBerordentlich ersehwert, dag trotz aller getroffenen Vorsiehtsmal3regeln die Reaktion der Wfirmer auf die in den Versuehen ver/~nderte Sauerstoffkonzentration auf das sts variierte, so dab erst eine gr6gere Zahl yon Messungen ein zuverl~ssiges Bild yon den vorliegenden Verh~ltnissen bieten konnte. Bei Erniedrigung der Sauerstoffspannung auf 15 % antworteten einzelne Wiirmer z. B. mi~ einer Erh6hung der Atmung auf 110%, ws ein anderer seinen Sauerstoffverbraueh auf 63% verminderte, wie die Zahlen der Tabelle 2 erkennen lassen. Diese seheinbare Regellosigkeit betrifft nieht nur die Ergebnisse an versehiedenen Wfirmern, sondern begegnet uns aueh bei dem gleiehen Wurm. Der Wurm B2, clef an dem einen Tage mit einer ErhShung seinerAtmung auf 110% der Atmung in Luft reagiert, antwortet einige Tage sp~tter auf die gleiehe Konzen- trations/~nderung mit einer Verminderung seines Sauerstoffverbrauehes auf 82%. Ein anderes Beispiel bietet der Wurm E, der in 4 Versuchs- reihen 4 sich erheblich unterseheidende Kurven ffir den Sauerstoff- verbraueh zwisehen 21% und 5% Sauerstoffgehalt lieferte.
Es erseheint daher unm6glieh, den einzelnen Sauerstoffspannungen eindeutige AtmungsgrSBen zuzuordnen. Immerhin ergab es sieh, dab bei Berfieksiehtigung einer gr61]eren Zahl yon Versuchen die Bildung yon Durehsehnittswerten gestattet ist, die nieht nut ein zusammenfassendes Bild yon den Einzelversuchen ergeben, sondern in gewissen Grenzen sogar reproduzierbar sind, wie die Ann/~herung meiner Werte an die yon Ko~oPAcxi erhaltenen beweist.
Ein ansehauliehes Bfld yon der Vielgestaltigkeit der Reaktion der Regenwfirmer auf die Anderung der Sauerstoffkonzentration bietet die graphische Darstellung in Abb. 1, in der eine groge Zahl yon normalen Wiirmern erhaltener Kurven fibereinander gezeiehnet sind. Aul3er der im allgemeinen mit sinkender Sauerstoffkonzentration abnehmenden Riehtung des Sauerstoffverbrauches - - eine Regel, die nur in wenigen F/~llen durehbroehen w i r d - kann man wenig aus den Kurven ent- nehmen. Eine weitergehende Analyse erfordert ein Eingehen auf das in Tabelle 3 enthaltene Zahlenmaterial.

54 Friedrieh Kriiger:
Es ist wichtig, hierbei noehmals zu betonen, dab das relative Ver- h~ltnis der Zahlen in einer Reihe, die einer Versuchsserie entsprechen, als mSglichst reine Auswirkung der ver~ndert~n Sauerstoffspannung gewertet werden mul~, denn um mSglichst gleichmal3ige physiologische Stimmung w~hrend des Versuches zu gews wurde jede dieser Serien innerhalb eines Tages durchgeffihrt, ws welcher Zeit der Wurm vollkommen unberfihrt in dem Apparat liegen blieb. Der einzige s Reiz, der auf ihn einwirken konnte, war das EinstrSmen der
i
.]g
Abb. 2. Die Abh~ng igke i t des Saue r s to f fve r - b r auches n o r m a l e r W i i r m e r yon der Sauer- s to f fkonzen t r a t i on . 13 der e rha l t enen K u r -
yen t tbere inander geze ichnet .
Gasgemische. Da diese aber Zim- mertemperatur aufwiesen und Was- serdampf gesi~ttigt waren, konnte der hierdurch gesetzte Reiz nicht sehr stark sein. Wenn aber das relative Verhs der Einzelwerte jeder Serie zueinander innerhalb der gegebenen Schwankungsbreite als zutreffend angesehen werden mu$, so gilt dieses auch ffir das rela- tive Verh~ltnis der Durchschnitts- werte, die aus der Summe der Ein- zelwerte gebildet wurden.
Wenn wir, wie es in Tabelle 3 geschehen ist, den Sauerstoffver- brauch in Luft als 100 bezeichnen, so betr~gt die Atmung bei der auf 15 % herabgesetzten Sauerstoff- spannung zwischen 63% und 110%, schwankt also offensichtlich in sehr weiten Grenzen. Da aber die fiber 100 hinausgehenden Werte Ausnah- men darstellen, die meisten Werte sogar betri~chtlich unter 100 liegen,
mu• man - - ebenso wie wir bei den weiteren Stufen sehen werden - - grunds~tzlich eine Abhs des Sauerstoffverbrauches yon der Sauerstoffspannung feststellen. Steht dieses im Widerspruch zu den Angaben von DOLK und vA~ D~R PAAVW, SO erreicht doch anderer- seits die Atmungsverminderung nicht das Ausmal~, welches THomAs land. Das kommt in dem Durchschnittswert von 91%, der sich aus meinen Zahlen ergibt, zum Ausdruck. Auf die wahrscheinliche Ursache des starken Atmungsabfalles, den dieser Autor fand, bin ich oben zu sprechen gckommen.
Grundsiitzlich den gleichen Verh~ltnissen begegnen wir bei weiterer Herabsetzung der Sauerstoffkonzentration auf 10%. Die von mir ge- fundene Schwankungsbreite liegt hier zwischen 58 % und 116%, ist also

Beitrage zur Physiologie des I4iimoglobins wirbelloser Tiere. I. 55
Tabe l l e 3.
Atmospharische 15 % O~ 10 % O3 5 % O3 Versuchs- Luft
Bezeichnung O~-Ver- RQ RQ branch
a
c
d f h i n 3
0 2
03 Aa A7 B2 B3 E1 En E~2 El8 F1 G
Durchschnittl. O2-Verbrauch
Resp.- Quotient
0 3 - V e t - brauch R Q
100 0,84 100 0,77 100 0,70 100 0,83 lO0 0,64 100 0,67 100 0,71 100 0,78 100 0,75 100 0,70 100 0,93 100 0,71 100 0,74 100 0,79 100 0,67 100 0,54 100 0,63 100 0,76 100 1,06
100
0,75
100 0,70 63 0,73 88 0,69 98 0,75 95 0,71 95 0,70 98 0,70 92 0,70 97 0,69
70 1,07 110 0,52 82 0,78 75 0,81 93 0,78 98 0,62 79 0,48 95 0,70
108 0,91
91
0,72
O~-Ver- braueh I
108 78 74 84 89 62 84 96 78
116 103 100 74 58 84 72 79 92
100
86
0,63 0,73 0,83 0,70 0,62 1,00 0,91 0,63 0,84 0,86 0,84 0,56 0,82 0,66 0,74 0,62 0,54 0,58 0,92
0,74
o 3-Ver- brauch R Q
70 0,81 51 0,78 51 1,18 49 0,95
41 1,32 38 1,90
34 1,62 88 0,96 95 0,76
63 0,97 48 0,71
71 0,74 67 0,61 67 0,51 80 1,00
61
0,99
noch betr / icht l icher als in der vorigen Stufe. Da aber nur in 4 Versuchen eine Steigerung des Sauers toffverbrauches gegenfiber der vorigen Stufe auf t r i t t , muf~ m a n auch hier wieder die grunds/~tzliche Abh/ ingigkei t yon der Sauers tof fkonzent ra t ion feststellen, was in dem Durchschni t t s - wert , der zu 86% gefunden wurde, zum Ausdruck kommt .
Die A t m u n g des Wurmes bei 5 % Sauerstoff lag in meinen Versuchen zwischen 95% und 34% der A t m u n g in Luf t und war ausn~hmslos - - zum Teil sogar sehr betr / icht l ich - - gegeniiber der vor igen Stufe herab- gese tz t . .~As Mit te lwer t ergibt sich 61%. W/~hrend zwischen 21% und 15% der durchschni t t l iche Atmungsabfa l l 9% und zwischen 15% und 10% nur 5% betrug, f/illt zwischen 10% und 5% - - also bei gleicher Konzen t r a t ionsve rminde rung - - der Sauers tof fverbrauch um 25%. Das k o m m t besonders deut l ich in der graphischen Dars te l lung der Mit te lwer te in Abb. 4 (S. 61) zum Ausdruck, wo der Abfa l l des Sauer- s toffverbrauches zwischen 21% und 10% sich als n~hezu geradl in ig erweist, urn un te rha lb 10% wesentl ich stei ler abzusinken.
Sowohl die Mehrzahl der Einzelwerte als aueh die Durchschni t t s - werte erweisen die grunds/~tzliche Abhs des Sauers toffverbrauches des Regenwurmes yon der Sauers tof fkonzent ra t ion . Wenn entgegen dieser Tendenz sich verha l t ende Wer t e auf t re ten, so kSnnen sie nur der Ausdruck yon regul ierenden F a k t o r e n sein und ich werde sie daher

56 Friedrich Kriiger:
auch als ,,Regulationen" bezeichnen. Da wir die Eigenschaften des Blutes als konstant annehmen miissen, kSnnen die ,,Regulationen" ihren Sitz nur in den Geweben des Wurmes haben und stellen deren Reaktion auf die ver~nderte Sauerstoffspannung dar. Es ist wohl anzunehmen, dab wir auf diese ,,Regulationen" die Mannigfaltigkeit der beobachteten Atmungskurven zurfickffihren miissen.
Wie immer wieder betont sei, haben die yon mir angegebenen Durch- schnittszahlen nur bedingten Wert, da bei der starken Schwankungs- breite der Ergebnisse die zuf~llige Zusammenstellung der verwandten Wiirmer auf das Endergebnis einen sehr grofien Einflu] hat. Unter diesen Umst~nden mug man meine Versuchsergebnisse als in bester ~bereinstimmung mit denen yon KONOPAOKI bezeichnen, der einen vollkommen gleichartigen Kurvenverlauf land. Es erhalten so meine Ergebnisse eine wertvolle Best~tigung durch diese ~ltere Arbeit. Dagegen konnten weder die Ergebnisse yon DOLK und vA~ D]~ PAAVW, noch die yon Ttto~As best~tigt werden.
Wenn hier vielleicht der Einwand erhoben wird, da~ artliche Unter- schiede eine Rolle spielen kSnnten, so mSchte ich dazu betonen, dab nach meiner Erfahrung hSchstens geringfiigige Unterschiede zwischen den Regenwurmgattungen bestehen, die bei cler relativ geringen Zahl von Versuchen iiberdeckt werden yon individuellen Schwankungen.
Die Kohlenss bietet bei den normalen Wfirmern recht einfache Verh~ltnisse, da sie sich im Durchschnitt entsprechend dem Sauerstoffverbrauch einstellt; eine Ausnahme bietet nur die Sauerstoff- konzentration yon 5%. W~hrend n~mlich zwischen 21% und 10% der respiratorische Quotient praktisch unver~ndert bleibt, steigt er bei 5% Sauerstoff betr~chtlich an. Die Einzelwerte schwanken zwar, wie die Tabelle 2 zeigt, ffir die Sauerstoffkonzentrationen zwischen 21% und 10% zwischen 0,48 und 1,35. Die Mittelwerte fiir diese drei Stufen liegen dagegen sehr nahe beieinander (0,75--0,72--0,74). Der allgemein hieraus berechnete Mit~elwert ws 0,74. Auf den genauen Zahlenwert lege ich jedoch kein grol]es Gewicht, da bei der grol~en Schwankungs- breite auch hier zuf~lligen Einfliissen eine sehr starke Rolle zukommt. Wie wir sehen werden, lieferten im wesentlichen die gleichen Wfirmer in den Kontrollversuchen mit den C0-vergifteten Tieren einen betr~cht- lich hSheren Durchschnittswert fiir den respiratorischen Quotienten. Dal~ aber das relative Verh~ltnis der Quotienten zueinander unter den eingehaltenen Versuchsbedingungen als zuverl~ssig angesehen werden kann, zeigte gerade die in diesen Versuchen hervortretende und kaum besser zu erwartende Konstanz fiir die Sauerstoffkonzentrationen zwischen 21% und 10%.
Gegeniiber dieser Konstanz von R Q ist die bei 5% Sauerstoff in fast allen Einzelversuchen zutage tretende ErhShung von R Q sehr auff~llig und finder daher auch in dem Durchschnittswert yon 0,99

Beitrgge zur Physiologie des Hgmoglobins wirbelloser Tiere. I. 57
ihren Ausdruck. Wir kSnnen hieraus wohl nur schliel]en, dab bei dieser Sauerstoffkonzentration die Sauerstoffversorgung der Gewebe unzu- reichend bleibt, so dab anaerobe Kohlens~urebildung einsetzt. Aueh K0~0rACKI beobachtete, bei etwa 10% Sauerstoffgehalt der Luft be- ginnend, eine ErhShung des respiratorischen Quotienten.
c) Die Abhdngigkeit der Atmung CO-vergifleter Wi~rmer vonder Sauer- sto//konzentration.
Um einen sicheren Vergleich mit der normalen Atmung zu erm6glichen, wurde stets vor Beginn der Versuche der Sauerstoffverbrauch in reiner Luft bis zur Konstanz geprfift und erst dann durch Einleiten yon CO- haltiger Luft mit den eigentlichen Versuchsserien begonnen. Hierbei ergab sich eindeutig in allen Versuchen eine deutliche Verminderung der Sauerstoffaufnahme, wie aus den Tabcllen 4 und 5 hervorgeht, yon denen erstcre wieder die rohen Versuchswerte enth~lt, ]etztere die umgerechneten Werte, bezogen auf die als 100 angenommene Atmung nicht vergifteter Wfirmer. Die Kohlens~ureabgabe ist auch hier wieder durch die GrSBe des respiratorischen Quotienten gekennzeichnet.
Die Herabsetzung der Atmung nach Vergiftung mit Kohlenoxyd ist eine Best~tigung der Ergebnisse yon T~o~As. Es betrug die Atmung der CO-Wtirmer nur 92% bis 46% der Atmung normaler Wfirmer in Luft. Als Durchschnittswert ergaben meine Versuche 73%. Es ist nicht m5glich, die beobachtete Atmungsverminderung auf die durch den CO-Zusatz herabgesetzte Sauerstoffkonzentratlon zurfickzuffihren, da die hierdurch bewirkte Konzentrations~nderung nur etwa 1% be- tr~g~ und, wie die im vorigen Abschnitt besprochenen Versuche zeigen, die Konzentrationsabh~ngigkeit der Atmung in diesem Bereich nur schwach ist.
Verminderung der Sauerstoffkonzentration auf 15% bewirkt eine weitere Herabsetzung der Atmung, die nur in einem Falle ausbleibt und sogar zu einer Steigerung fiber den Luftwert des normalen Tieres hinausffihrt. Dieses beweist, daB auch bei den CO-vergifteten Wfirmern ,,Regulationen" vorkommen. Dem HSchstwert yon 115% steht als niederster Wert 26% gegeniiber, der Durchschnitt aller Zahlen ergibt fiir diese Stufe den Wert von 60 % der Ausg~ngsatmung.
Auch bei 10% Sauerstoff wurde ein Fall beobachtet (At), bei d'em der Sauerstoffverbrauch oberhMb der Atmung in Luft liegt. Im fibrigen lassen sowohl die Einzelwerte als auch der Durchschnitt yon 55 % ein weiteres Absinken der Atmung erkennen. Entsprechendes gilt auch fiir die letzte Stufe yon 5% Sauerstoff, fiir welche sich der Durchschnitts- wert zu 42% der Ausgangsatmung ergibt.
Die Atmung der CO-Wiirmer unterscheidet sich yon der normaler Wiirmer einmal dadurch, dab sie bei allen untersuchten Sauerstoff- konzentrationen vermindert ist. Ferner ist zwischen 21% und 10%
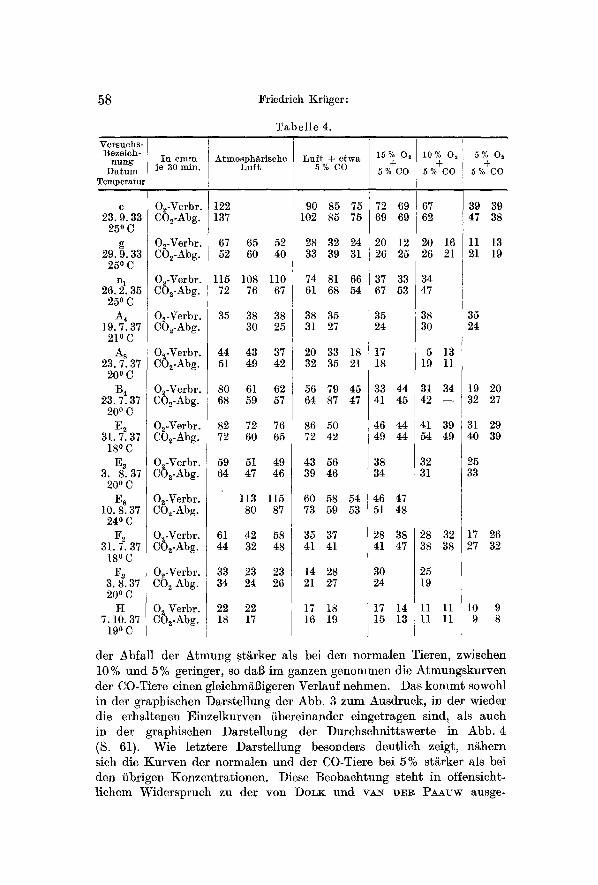
58 Friedrich Kriiger:
Tabe l le 4.
Versuchs- I Bezeich- I
h u n g / Datum
Temperat~r
e
23.9.33 250 C
g 29.9.33
25 ~ C
n 1 26.2.35
250 C
A4 19.7.37 210 C
As 23.7.37
20 ~ C
B4 23.7.37
200 C
E2 31.7.37
180 C E~
3. 8.37 200 C
Es 10.8.37 24 o C
Fs 31.7.37
]8 o C F~
3.8.37 200 C
H 7.10.37
190 C
In cram je 30 min.
O~-Verbr. C02-Abg.
Oo-Verbr. C~)2-Abg.
Oe-Verbr. COz-Abg.
O~-Verbr. C02-Abg.
O~-Verbr. C02-Abg.
O~-Verbr. C02-Abg.
02-Verbr. C02-Abg.
O~-Verbr. C02-Abg.
O~-Verbr. C02-Abg.
Os-Verbr. CO S Abg.
02 Verbr. C02-Abg.
Atmospharische Luft
122 137
67 65 52 52 60 40
115 108 110 72 76 67
35 38 38 30 25
44 43 37 51 49 42
80 61 62 68 59 57
82 72 76 72 60 65
59 51 49 64 47 46
113 115 80 87
61 42 58 44 32 48
33 23 23 34 24 26
22 22 18 17
Luft + etwa 5% CO
90 85 75 102 85 75
28 32 24 33 39 31
74 81 66 61 68 54
38 35 31 27
20 33 18 32 35 21
56 79 45 64 87 47
86 50 72 42
43 56 39 46
60 58 54 73 59 53
35 37 41 41
14 28 21 27
17 18 16 19
15 o~ o3
5% co
72 69 69 69
20 12 26 25
37 33 67 53
35 24
17 18
33 44 41 45
46 44 49 44
38 34
46 47 51 48
28 38 41 47
3O 24
17 14 15 13
10 o~ o2
5% co
67 62
20 16 26 21
34 47
38 3O
13 1 11
31 34 42 - -
41 39 54 49
32 31
28 32 38 38
25 19
11 11 11 11
5% O~ +
5% CO
39 39 47 38
11 13 21 19
35 24
19 20 32 27
31 29 40 39
25 33
17 26 27 32
10 9 9 8
der Abfall der A tmung starker als bei den norm~len Tieren, zwischen 10% und 5% geringer, so dab im ganzen genommen die Atmungskurven der CO-Tiere einen gleichms Verlauf nehmen. Das k o m m t sowohl in der graphischen Darstel lung der Abb. 3 zum Ausdruck, in der wieder die erhal tenen Einze lkurven i ibereinander e ingetragen sind~ als auch in der graphischen Darstel lung der Durchschni t t swer te in Abb. 4 (S. 61). Wie letztere Darstel lung besonders deutl ich zeigt, ns sich die K u r v e n der normalen und der CO-Tiere bei 5% sti~rker als bei den fibrigen Konzentra t ionen. Diese Beobachtung steht in offensicht- l ichem Widerspruch zu der yon DOLK und v~N ~ ) ~ PAACW ausge-

Beitr~ge zur Physiologie des H~moglobins wirbelloser Tiere. I. 59
T a b e l l e 5.
Versuchs- Atmosph~ri-
sche L~ft Bezeichnung
e I 100 g 100 n i 100 A 4 100 A s 100 B 4 100 Ee 100 E 3 100 E s 100 F 2 100 F~ ioo H 100
O ~-V(~r- brauch R Q
1,12 0,82 0,65 0,72 1,15 0,91 0 , 8 6 0 , 9 8 0,73 0,77 1,07 0 , 7 9
5% O.. + CO Luft + CO
O:-Ver- O~-Ver- brauch RQ brauch RQ
15% O~+CO 10% O~+CO
0 ~-Ver- O 2-Ver- brauch 1~ Q brauch R Q
57 26 32 95 41 56 58 72 40 61
115 68
0,98 I 55 1,69 ! 29 1,71 31 0,69 103 1,06 22 1,13 47 1,03 52 0,90 60 0,94
!1,35 56 0,80 96 1,03 50
55
1,11
0,93 32 1,31 20 1,38 - - - 0,79 95 2,50 - - - 1,35 28 1,29 38 0,97 47
1,28 39 0,76 - - - 1,00 41
42
1,23
I)urchschnittl. 100 O2-Verbrauch
Resp.- Quotient 0,88
68 1,05 46 1,24 67 0,831 97 0,80 59 1,28 88 ]1,09 88 0,84 92 0,87 50 1,07 67 1,14 81 1,23 77 1,00
60 7, I 1,04
1,09 1,69
0,69
1,51 1,32 1,32
1,41
0,90
1,24
s p r o c h e n e n V e r m u t u n g , dab das Hs m o g l o b i n des R e g e n w u r m e s die Aus-
n u t z u n g n i ede re r Saue r s to f fkonzen-
t r a t i o n e n e r l e i ch t e r t ; es g i l t dieses also s icher n i c h t f i ir die a n g e w a n d -
t e n Ve r suchsbed ingungen . F i i r e ine w e i t e r g e h e n d e Ana ly se
d e r W i r k u n g de r CO-Verg i f t ung is t die Be r i i cks i ch t igung der K o h l e n -
s~ureaussche idung uner l~Blich. I c h h a t t e schon oben d a r a u f h ingewie-
sen, d a b die A u s g a n g s v e r s u c h e dieser Re ihen , obwohl sie in g le icher Weise
m i t L u f t u n d u n v e r g i f t e t e n u n d z u m groBen Tel l sogar den g le ichen Wii r -
m e r n anges te l l t wurden , e inen er- heb l i ch hShe ren W e r t fiir R Q - - n~m- l ich 0,88 i m D u r c h s c h n i t t - - e rgaben .
D a a b e t a u c h diese Versuche so durchgef t ih r t wurden , dab das rela- t i v e Verh~l tn i s der Z a h l e n zue inan-
de r als zu t r e f f end angesehen w e r d e n
//\
f
\
\
Abb. 3. Die Abh~ngigkeit des Sauerstoffver- brauches CO-vergifteter Wtkrmer yon tier Sauerstoffkonzentration. 11 der erhaltenen ]~urven sind iibereinander gezeichnet (ab- weichend yon Tabelle 5 ist die Atmung in
Luft + CO als 100 angenommen).
l o 20
Oz - IF~#nung

60 Yriedrich Krtiger:
kann, bleiben die wohl etwas hSher als ,,normal" liegenden Quotienten ohne EinfluB.
Bei der Betrachtung der Kohlens~ureabgabe fs die fast aus- nahmslos festzustellende ErhShung yon I~Q als Folge der Vergiftung mit dem Kohlenoxyd ins Auge. Dieses kommt auch im Durchschnitts- weft zum Ausdruck, der fiir CO-Wiirmer in Luft zu 1,04 gefunden wurde, ws der Ausgangswert 0,88 betrug. Bei Verminderung der Sauer- stoffkonzentration steigt der Wert yon R Q weiterhin an, so findet sich fiir 15% Sauerstoff ein RQ von 1,11 im Durchschnitt, fiir 10% Sauerstoff ein RQ. yon 1,23 und schlieBlich fiir 5% Sauerstoff ein RQ yon 1,24.
Es bleibt die Frage zu kl~ren, ob diese Steigerung des respiratorischen Quotienten Ausdruck einer Schs der Gewebsatmung oder einfach
eine Folge der Ausschaltung des Hgmoglobins Tabelle 6.
Atmung tier CO-Tiere
in Luft
97 92 88 88 81 77 68 67 67 59 50 46
Erh6hung des I~Q gegeniiber
normalen Tieren in %
q- 11,1 - - 11,2 q- 19,8
- - 22,3 q- 15,0 q- 26,0
- - 6,3 § 48,0 q- 27,7 q~ 11,3 q- 46,7 q- 51,2
ist. Wenn letzteres zutrifft, dann muf~ die Steigerung des RQ um so grSBer sein, je starker die Atmung gegeniiber normalen Tie- ren herabgesetzt ist, da ja in diesem Falle ein urs~tchlicher Zusammenhang zwischen der Anderung beider Gr61~en besteht. Von einer Schs der Gewebsatmung wfirde man dagegen erwarten, dab sie sich ziemlich gleich- ms bei allen Tieren auswirkt.
Zur Priifung habe ich in Tabelle 6 die Sauerstoffverbrauchswerte der Wfirmer in Luft q- CO der GrSl~e nach geordnet und ihnen die in Prozenten ausgedrfickte Erh6- hung der R Q-Werte, bezogen auf die in
reiner Luft gefundenen Quotienten, gegeniibergestellt. Wenn man zwar auch keine volle Parallele zwischen der Atmungsverminderung durch das CO und der ErhShung des R Q feststellen kann, so war eine solche bei der immer wieder beobachteten Regulationsf~higkeit der Atmung des Regenwurmes nicht zu erwarten. Im groBen und ganzen aber kann man feststellen, dab der RQ um so starker erhSht ist, je starker die Atmung vermindert ist. W~hrend in der oberen H~lfte der Tabelle als HSchstwert der Steigerung 27% vorkonamt, erreicht in der unteren I-I~lfte der Tabelle, die die Atmungsverminderungen fiber 30% erfaBt, die ErhShung des R Q Wertes um 50%.
Hinzuzuffigen ist noch, dad die durch das CO bedingte Atmungs- verminderung ein solches Ausmal~ erreicht, dab sie auch bei normalen Wiirmern entsprechende Quotienten bedingen wiirde. Eine allenfalls vorhandene Sch~digung der Gewebsatmung kann daher hSchstens geringfiigig sein.

BeitrAge zur Physiologie des tt~moglobins wirbelloser Tiere. L 6t
d) Besprechung der Versuchsergebnisse.
Diesem Abschn i t t mSchte ich eine kurze Zusammenfassung der Ergebnisse vorausschicken. Zu diesem Zweck sind in Tabel le 7 noch e inmal die aus meinen Versuchen sich ergebenden Durchschni t t swer te zusammengeste l l t . Ich ha t t e schon oben be~ont, dab es sich hierbei n icht um ganz e indeut ig festgelegte Zahlen ~0~
r - " ~ [ I I hande l t , sondern dab sie s ta rk yon I ~ Z e r s individuel len und physiologischen Schwankungen abh/~ngig sind. Da- gegen ist das re la t ive Verhgltnis der zu einer Reihe gehSrenden Wer t e als in den gegebenen Grenzen zuverl/~s- sig anzusehen. Besonders deut l ich k o m m t der EinfluB solcher unbe- s t immte r F a k t o r e n in der Ges ta l tung des resp i ra tor i schen Quot ienten zum Ausdruck, fLir den in den beiden vor l iegenden t{eihen un te r gleichen Bedingungen und teilweise sogar fiir d ie gleichen Wti rmer e inmal der Wer t 0,75, das andere Mal 0,88 gefunden wurde.
Wie die Tabel le 7 und die graphi- sche Dars te l lung in Abb. 4 beweisen, ergeben sich zwischen der A tmung norma]er und CO-verg'ifteter Regen- wti rmer bet r~cht l iehe Unterschiede. Der Sauers tof fverbrauch normaler
86
%.
d0
Q
~. CO-g/Cimzez
\ \
2'0 /d ~'0 %
Abb, 4. Graphische Darstellung der er- halLenen Durchschnittswerte ffir die Ab- h/~ngigkeit normaler ( -) und CO-ver- gifteter WOrmer ( . . . . . ) yon der Sauer-
stoffkonzentration.
Wi i rmer zeigt in dem Bereich der Sauers tof fkonzent ra t ion zwischen 21% und 10% eine zwar geringe, aber doch eindeutige Abh~ngigkei t v o n d e r Sauers tof fkonzent ra t ion . Dieselbe is t jedoch un te r 10% Sauer- stoff wesentl ich st/~rker, was besonders deut l ich die graphische Dar-
Tabe l l e 7.
O2-Verbrauch normaler Wtirmer . . . Q-Verbrauch der CO-Wfirmer
02 vom l ib. transportiert in % . . . 02 vom Hb. transportiert . . . . . .
Luft
100 73
27 27
15 % 02
91 6O
34 31
10 % 02
86 55
36 31
5% o3
61 42
31 19
s tel lung erkennen l~Bt. Die Kohlens/~ureabscheidung ver ls zwischen 21% und 10% Sauerstoff nahezu vo l lkommen paral le l der Sauerstoffauf- nahme, wie der unver/~nderte R Q zeigt. Dagegen bedingt eine Sauer- s tof fspannung yon 5 % eine unve rkennbare Erh6hung dieses Quot ienten.

62 Friedrich Kriiger:
Bei den CO-vergifteten Wiirmern liegt der Sauerstoffverbrauch bei allen Spannungen unterhalb dem normaler Tiere, jedoch ~llt er in dem ganzen Bereich zwischen 21% und 5% nahezu gleichm~ig ab. Weiterhin ist fiir die CO-Tiere die ErhShung des respiratorischen Quotienten charakteristisch, die schon bei der Sauerstoflkonzentration der Luft einsetzt und sich mit sinkender Sauerstoffspannung steigert. Es konnte klargestellt werden, dal~ diese ErhShung des respiratorischen Quotienten nicht auf einer Sch~digung der Gewebeatmung beruht, sondern der Unterschied in der AtmungsgrSfle normaler und CO-ver- gifteter Wfirmer lediglich auf die Ausschaltung des H~moglobins zuriicl~- zufiihren ist.
Es ergibt sich hieraus zuns einmal die sehr bemerkenswerte Tatsache, da~ in dem untersuchten Temperaturbereich bei allen Sauer- stoffkonzentrationen - - also auch in der atmosph~rischen Luf t ' - - das Hi~moglobin an dem Transport des Sauerstoffs beteiligt ist. Wenn das ftir die w~hrend des Versuches in roller Ruhe befindlichen Wiirmer gilt, so sicher in erhShtem Mal~e fiir sich bewegende Wiirmer.
Durch einen Vergleich der Atmungsgr61~e normaler und vergifteter Tiere kSnnen wir uns daher ein Bild machen yon dem Umfang, in dem das H~moglobin einerseits und der im Blutplasma physikalisch gelSste Sauerstoff andererseits an der Gesamtatmung beteiligt sind. Wir nehmen dabei an, dal3 die fiir die AtmungsgrSBe der CO-Tiere gefundenen Werte etwa dem bei den verschiedenen Sauerstoffkonzentrationen in physikalisch gelSster Form transportierten Sauerstoff entsprechen. Da die ErhShung des respiratorischen Quotienten anzeigt, dab bei ihnen Sauerstoff- mangel in den Geweben auftritt, daft man vermuten, da]3 - - soweit m5glich - - aller im Blut vorhandene Sauerstoff verbraucht wird, was bei normalen Tieren nicht vorausgesetzt zu werden braucht. Es wiirden also diese Zahlen die HSchstwerte darstellen, die durch den physikalisch ge]Ssten Sauerstoff gedeckt werden kSnnen. Rechnet man diesen Anteil um auf den Gesamtsauerstoffverbrauch, so ergibt er sich in Luft zu 73%, bei 15% 02 zu 66%, bei 10% 02 zu 65% und bei 5% 02 zu 69%. Wie man siehf, ergeben sich nicht sehr erhebliche Unterschiede fiir die verschiedenen Sauerstoffspannungen, so da]~ man a!!gemein etwa sagen kann, dal~ 2/3 des verbrauchten Sauerstoffs in physikalisch gelSstem Zustande den Geweben zugefiihrt wird.
Bei diesem Ergebnis ist weniger iiberraschend der Umfa~g, in dem die physikalische LSsung zur Befriedigung des Sauerstoffbediirfnisse~ herangezogen wird, als die Tatsache, da~ der Anteil des physikalisch gelSsten Sauerstoffs in einem so weiten Konzentrationsbereich nahezu unver~ndert bleibt.
Der Anteil des vom Hs transportiertel~ Sauerstoffs ergibt sich danach zu etwa 27% bzw. 34% bzw. 36% und 31%. Er ent~pricht also etwa dem restlichen Drittel des Sauerstoffverbr~uches, Ein etwas

Beitrgge zur Physiologie des H~moglobins wirbelloser Tiere. I. 63
anderes Bild erhalten wir, wenn wir nicht den relativen, sondern den absoluten Anteil des vom Hs transportierten Sauerstoffs bestimmen, indem wir ffir die verschiedenen Sauerstoffkonzentrationen die Differenz aus der Atmung normaler und CO-vergifteter Tiere bilden. Ffir a~mosph~risehe Luft betr~gt diese Differenz etwa 27, ffir 15% O9 und 10% Oz etwa 31, ffir 5% O2 etwa 19.
Wenn wir die als nicht vollkommen eindeutig anzusehenden Aus- gangswerte berfieksichtigen, kSnnen wir den Anteil des vom Hs globin transportierten Sauertoffs zwischen den Konzentrationen 21% und 10% mit gleiehm~Big etwa 30 annehmen. Wir dfirfen diesen Wert mit einiger Wahrseheinlichkeit ~ls den maximalen Wert annehmen. Um so mehr erstaunt uns dann, dab bei 5 % 02 der vom H~moglobin trans- portierte Sauerstoff nur 2/3 dieses Wertes betrs ahem, da ja unter diesen Bedingungen die durchweg erhShten respiratorischen Quotienten einen Sauerstoffmangel in den Geweben verraten, was eher eine erhShte Ausnutzung des hs ndenen Sauerstoffes erwarten lassen wfirde. Verst~ndlieh wird uns diese Erscheinung nut, wenn wir annehmen, dab bei 5% Sauerstoff keine vollstAndige S/~ttigung der Kapazit~t des Hs unter dan gegebenen Umstgnden erreicht wird. Die Ladungsspannung des Regenwurmh~moglobins scheint also unter den Versuehsbedingungen erst oberhalb 5%, wahrscheinlieh etwa bei 10% Sauerstoff zu liegen.
Mit dam Nachweis, dab das H~moglobin auch bei hSherer Sauerstoff- konzentration in den AtmungsprozeB eingreift und daft bei 5% Sauer- stoffgehalt die Ladungsspannung des HAmoglobins nicht erreicht ist, wird die Anschauung hinf~llig, dab es ]ediglich zur Ausnutzung niederer Sauerstoffspannungen dient. Ebensowenig aber beschrgnkt sieh das Hgmoglobin auf das Eingreifen nach Zeiten yon Sauerstoffmangel zur Besehleunigung der Erholungsatmung, wie es HAR~ISCH (z. B. 1937) vermutet, denn es liegt kein Anlaft vor, anzunehmen, daft die in ihren Schalen in Luft gehaltenen Wfirmer irgendwie unter respiratoriseh ungiinstigen Bedingungen lebten und ebensowenig spricht die Gestaltung des respiratorisehen Quotienten oder der Verlauf der ganzen Atmung ffir eine solche Annahme. Wir stehen daher vor der Notwendigkeit, nach neuen Gesichtspunkten fiir die physiologisehe Bedeutung des Hgmoglobins Umschau zu halten. Wenn wir das tun wollen, so ergibt sieh die Notwendigkeit, uns fiber die Art des Eingreifens des H~moglobins Klarheit zu verschaffen.
In diesem Zusammenhang ist zun~chst die Frage zu stellen, worauf die Konzentrationsabh~ngigkei$ des Sauerstoffverbrauches zurfick- zuffihren ist, die wir sowohl bei normalen als auch vergifteten Wiirmern fanden. Man kSrmte im Sinne yon HE~zE (1910) an den EinfluB be- schrgnkter Sauerstoffzufuhr denken. Hiergegen spricht aber meines Ermessens der in weitem Bereieh konstant bleibende respiratorische

64 Friedrich Kriiger:
Quotient, der in manchen F/fllen auch bei 5% Sauerstoff noeh seine normale GrSl~e beibeh/tlt. Ein Beispiel fiir die Beschr/~nkung der Atmung durch die Sauerstoffzufuhr bieten die CO-vergifteten Wiirmer, bei denen wir dann aueh die erwartete Abh/s des respiratorisehen Quotienten yon der Sauerstoffspannung fanden, wenngleich ebenfalls hierbei die Konzentrationsabh/~ngigkeit der Atmung eine Rolle spielt.
Es liegt hier allerdings eine gewisse Schwierigkeit vor. Da die physi- kalische LSslichkeit des Sauerstoffes parallel zu dessen Partiardruck verl/~uft, miiBte man annehmen, dab sich in Luft 4real so viel Sauerstoff im Blur 15st als in dem Gasgemiseh mit 5% Sauerstoff. Wiirde der Sauerstoff als einziger begrenzender Faktor auftreten, so diirfte die Atmung in 5% Sauerstoff bei den CO-vergifteten Wiirmern nur 1/4 der Atmung in Luft betvagen, ein Wert, der in Wirklichkeit in keinem Fal]e erreicht wird. Wenn wir auch die hier zugrunde liegenden Faktoren nieht kennen, so miissen wir doch allgemein Ieststellen, dal~ die Gewebe des Wurmes die niederen Sauerstoffkonzentrationen besser ausnfitzen als die h6heren. Wenn man davon sprieht, dab ein respirato- rischer Farbstoff dazu dient, ,,niedere Sauerstoffkonzentrationen aus- zunutzen", so mug man sieh dariiber klar sein, dab ein soleher im wesentliehen nur die Menge des transportierten Sauerstoffs giinstig beeinflussen kann, dagegen nur in beschr/s Umfang - - durch den B o h r e f f e k t - - dem Gewebe den Sauerstoff unter hSherer Spannung anzubieten vermag, als er in der Umgebung vorhanden ist.
Die Menge des vom Blur transportierten Sauerstoffs ist aul]er yon der Anwesenheit respiratorischer Farbstoffe noch abh/~ngig v o n d e r Menge der Blutfliissigkeit, ferner der Umlaufsgeschwindigkeit und Geschwindigkeit der Sauerstoffaufnahme in den Atmungsorganen - - hier der KSrperoberfl/~ehe. ~ber die beiden letzteren Faktoren vermag ieh wenig zu sagen. Z~hlungen der Herzfrequenz bei versehiedenen Sauerstoffspannungen ergaben keinen Anhaltspunkt fiir deren Anpassung an ver/~nderte Atmungsbedingungen. Die yon Ko~oPacXI sehon be- schriebene Streckung der Wiirmer bei niederen Sauerstoffkonzen- trationen habe ieh auch h/~ufiger beobachtet, dureh sie wiirde die fiir den Sauerstoffzutritt zur Verfiigung stehende Fl~che vergrSBert werden, aber naeh meinen Beobachtungen wird die Streckung regelm~tBig nach einiger Zeit riiekg/~ngig gemacht, so dab ihr wohl keine besondere Be- deutung ftir die Regulation des Sauerstoffverbrauches zukommt.
Was die Blutmenge des Regenwurmes anbetrifft, so ist sie, wie die Versuche mit den CO-Tieren erwiesen, aueh in Verbindung mit anderen regulierenden Faktoren nieht in der Lage, den vollen Sauerstoffbedarf der Gewebe zu decken und bedarf der Erg/inzung durch das H/~mo- globin. Die Eigentfimliehkeit des H/~moglobins ist, dab es oberhalb einer bestimmten Konzentration - - in dem vorliegenden Falle etwa yon 10% Sauerstoff ab - - eine gleiehbleibende Menge Sauerstoff trans.

Beitr~ge zur Physiologie des H~moglobins wirbelloser Tiere. I. 65
portiert. W~re die Blutmenge des Regenwurmes yon vorneherein so groin, dab sie aueh bei niederen Sauerstoffkonzentrationen den gesamten Bedarf decken kSnnte, so wiirde das Sauerstoffangebot bei steigender Sauerstoffkonzentration wesentlieh starker anwaehsen, als das dureh die Mitbeteiligung des oberhalb einer gewissen Grenze gesi~ttigten H~moglobins der Fall ist. Bei der Konzentrationsabhiingigkeit der Atmung des Regenwurmes wiirde das ein st~rkeres Anwachsen des Sauerstoffverbrauehes bedeuten als sie durch die M]tbeteiligung des Hi~moglobins gegeben ist. Wie der einfaehe Vergleich der normalen und CO-vergifteten Tiere in dem K0nzentrationsbereich zwischen 10% und 21% Sauerstoffgehalt - - in welehem Bereiche das H~moglobin votl wirksam ist - - zeigt, bedingt das H~moglobin verminderte Kofizen- trationsabh~ngigkeit durch Ausgleichung des Sauerstoffangebotes.
Trotzdem mSehte ieh nicht annehmen, da[t sieh hiermit die Bedeutung des Hi~moglobins erschSpft. Eine solche Funktion kSnnte ein anderer respiratorischer Farbstoff - - z. B. das H~tmoeyanin - - in i~hnlicher Weise iibernehmen, wi~hrend ja gerade das Auftreten der roten, eisen- haltigen Farbstoffe bei sehr vielen, unter respiratorisch ungfinstigen Bedingungen lebenden Tieren das besondere Problem darstellt. Es kann daher mit den vorliegenden Versuchen noch nicht als gelSst betrachtet werden. Die Versuehe wurden unter Bedingungen durehgefiihrt, die sieh erheblich yon den unter normalen Umsts gegebenen entfernen. Vor allem ist die verhi~ltnism~i~ig sehr hohe Temperatur zu beanstanden, unter der alle bisherigen Untersuehungen ausgeffihrt wurden. Nur wenn gelegentlieh der Regenwurm die Erde verls wird er ihnen ausgesetzt sein, dann aber nie mit niederen Sauerstoffkonzentrationen in Bertihrung kommen. Solehen wird der Regenwurm im allgemeinen nut in der Erde begegnen, in der niedere Temperaturen vorherrschen. Es gilt also zu- n~ehst die Atmung bei der im Boden im Mittel herrsehenden Temperatur von etwa 10 ~ zu untersuchen. Wi~hrend diese Temperaturbedingung verh~ltnismi~l~ig leieht zu erfiillen ist, wi~re au~erdem als schwieriger zu untersuchender Faktor der Einflu]~ der Kohlenss zu prtifen, ein Faktor, der aber gerade unter respiratorisch ungiinstigen Bedingungen ftir den Regenwurm erhShte Bedeutung hat.
Zusammenfassung der Ergebnisse.
1. Unter den angew~ndten Versuchsbedingungen - - kohlenss ffeie Gasgemische yon etwa 200 C - - erweist sich die Atmung der Regen- wiirmer im Bereiche der Sauerstoffkonzentration zwischen 21% und 5% als spannungsabhi~ngig, jedoch ist die Reaktion der Wiirmer auf Konzentrationss sehr unterschiedlich und ver~ndert sich auch beim gleichen Wurm. Im Durchschnitt fi~llt die Atmung zwischen 21% und 10% nur langsam ab, unterhalb 10% dagegen wesentlieh steiler (Abb. 4).
Z. f. ve rg l . Phys io log i e . Bd , 28. 5

66 Friedrieh Krtiger: Physiologie des H/s wirbelloser Tiere. I.
2. Die Kohlens/iureabgabe verl/iuft zwischen 21% und 10% Sauer- stoff parallel zur Sauerstoffaufnahme, so da$ der respiratorisehe Quotient konstant bleibt. Bei 5% Sauerstoff beobachtet man eine relative Zu- nahme der Kohlens~ureabgabe, so da$ der respiratorisehe Quotient ira allgemeinen erhShte Werte erh/flt.
3. Die Atmung CO-vergifteter Wtirmer ist stets niedriger als die normaler Tiere. Die Druckabhi~ngigkeit zeigt in dem untersuchten Bereieh einen gleichmi~$igeren Verlauf als bei den normalen Tieren. Da insgesamt bei ihnen der Atmungsabfall geringer ist, n~hern sieh die ffir die Durehsehnittswerte erhaltenen Zahlen bei niederen Sauerstoff- konzentrationen.
4. Der respiratorisehe Quotient der CO-vergifteten Wiirmer ist gegentiber dem normaler Wiirmer stets erhSht, und zwar steigt er mit sinkender Sauerstoffspannung an. Die ErhShung des Quotienten ist eine Folge der mangelhaften Sauerstoffversorgung der Gewebe dutch den Ausfall des Hi~moglobins.
5. Die bei allen Sauerstoffkonzentrationen verminderte Atmung zeigt an, dab das H/~moglobin aueh bei allen Sauerstoffkonzentrationen am Sauerstofftransport beteiligt ist. Der Anteil des vom Hi~moglobin be- fSrderten Sauerstoffs betr/~gt bei allen untersuehten Konzentrationen gleiehmi~l~ig etwa 1/s des gesamten Sauerstoffverbrauehes. Die uner- wartete Tatsaehe, da$ unter den gew/~hlten Versuchsbedingungen der Anteil des vom Hi~moglobin transportierten Sauerstoffs keine ErhShung bei niederen Sauerstoffspannungen erf/ihrt, kann auf unvollsti~ndige Ladung des H~moglobins bei 5 % Sauerstoffgehalt zuriickgefiihrt werden.
Schriftenverzeichnis. DOLK, H. E. u. F. VAI~ DER PAAUW: Die Leistungen des H~moglobins beim
Regenwurm. Z. vergl. Physiol. 10 (1929). - - GIESCHE~, A. : Beitrgge zur Atmungs- physiologie des Regenwurms. Zool. J . Physiol. 48 (1931). - - HAnNISC~, O.: Primi~re und sekundi~re Oxybiose wirbelloser Tiere. Verh. dtsch, zool. Ges. 1987. - - I-IE~ZE, M.: Ober den EinfluB des Sauerstoffdruckes auf den G~swechsel einiger Meerestiere. Bioehem. Z. 26 (1910). - - JORDAN, H.: Allgemeine vergleiehende Physiologie der Tiere. Berlin u. Leipzig 1929. - - JOrDAn, H. u. B. Sc~wA~z: Einfache Apparate zur Gasanalyse und 1V[ikrorespirometrie in best immten Gas- gemischen und fiber die Bedeutung des Iti~moglobins beim Regenwurm. Pflfigers Arch. 185 (1920). - - KO~OPACKI, M. M.: lJber den AtmungsprozeB bei l~egen- wiirmern. Anz. Akad. Wiss. Krakau 1907. - - KRtiOER, F.: Ein neuer Mikro- respirationsapparat zur fort laufenden manometrischen Bestimmung des respi- ratorisehen Quotienten. Z. vergl. Physiol. 21 (1934). - - Neue Versuche fiber die Bedeutung des Hgmoglobins im Blute des l~egenwurmes. Verh. dtsch, zool. Ges. 1938. - - T~oMAs, J. B. : t ibe r die Atmung beim Regenwurm. Z. vergl. Physiol. 22 (1935). - - Die Atmungsregulierung yon LV~BRICVS. Kon. Akad. Wetenseh. Amsterd. 88 (1935).





























