Embed Size (px)
Citation preview

ELSEVIER Journal of Experimental Marine Biology and Ecology,
204 (1996) 225-239
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
Behaviour and time allocation of the burrowing shrimp Callianassa subterranea (Decapoda, Thalassinidea)
E.J. Statnlmis”, T. Reede-Dekker, Y. van Etten, J.J. de Wiljes, J.J. Videler
Dept. of Marine Biology, University of Groningen, P.O. Box 14, 9750 AA, Haren, The Netherlands
Received 4 December 1995; revised 11 March 1996; accepted 28 March 1996
Abstract
The behaviour and allocation of time of the endobenthic shrimp Cullianassa subterranea from the central North Sea was studied in the laboratory. Animals were allowed to construct a two-dimensional burrow in large transparent sediment filled cuvettes tailored to their body width. The behaviour of the animals was found to consist of 12 behavioural states which were described in an ethogram. The allocation of time of animals in cuvettes was recorded to the nearest 0.1 s during 1 or 2 h observation periods randomly distributed over the day. All records were statistically tested for their validity with respect to visibility of the animals studied. The behavioural states were grouped into six classes using a type of Markov-chain sequence analysis: Wandering, Resting, Burrowing, Surveying, Grooming and Ventilating. An average time budget of 10 animals based on 98 valid 1 h protocols shows that C. subterranea spends 27.4?2.2% (SEM) of the time Burrowing. This is a little more than 40% of the time the animal is active. Wandering and Grooming each take about half that amount, with 12.6?0.9% and 14.92 1.4% respectively. Ventilating takes 8.6+1.5% and only little time is spent Surveying the burrow (2.6?0.4%). The animal is Resting during 33.9+2.4% of the time. Circumstantial evidence indicates that feeding is part of the burrowing behaviour. No major periodicity in the behaviour of C. subterranea was
encountered.
Keywords: Thalassinidea; Callianassa subterranea; Behaviour; Burrowing; Time budget
1. Introduction
The behaviour of endobenthic thalassinid shrimps is difficult to study because of their hidden way of life in burrows in the sea bed. That is probably the reason why studies on thalassinid behaviour concentrated mainly on burrow ventilation and sediment excava-
*Corresponding author. Tel.: ( + 31-50) 363-2078; fax: ( + 31-50) 363-5205; e-mail: [email protected]
0022.0981/96/$15.00 0 1996 Elsevier Science B.V All rights reserved
PII SOO22-098 1(96)02587-7

226 E.J. Starnhuis rt al. I J. E.xp. Mrrr. Bid. Ecol. 204 (1996) 2252.39
tion rates measured outside the burrows, and on burrow architecture. Species belonging to the predominantly filter feeding family Upogebiae are estimated to spend up to half their time on burrow ventilation for feeding purposes (e.g., Dworschak, 1987; Scott et al., 1988). Sediment excavation rates are usually low after the initial establishment of the burrows (Ott et al., 1976; Suchanek, 1985). In contrast, most species of the family Callianassidae appear to be continuous excavators. Daily sediment turnover estimates range from 10 g dry weight (DW) for the temperate Cul/ianassa subterruneu to 3.4 kg DW per m2 for tropical Culliunussu-species (Rowden and Jones, 1993). Callianassidae only ventilate their burrow every now and then for respiratory purposes. Occasionally, excavated sediment is removed from the burrow by pumping it to the bottom surface (Suchanek, 1985; Witbaard and Duineveld, 1989). Most Callianassidae are established as or assumed to be deposit feeders, constantly mining for food, and thereby continuously changing their burrow systems (Suchanek, 1985; Griffis and Suchanek, 1991). Some species of the Upogebiae, the Callianassidae and the Jaxeae are assumed to have a mixture of feeding strategies: they may filter the ventilation flow for food, as well
as feed on deposit, graze on the micro-flora gardened on plant debris accumulated in the burrow, or scavenge on organic remains. This mixed feeding strategy results in relatively simple burrow architectures and intermediate sediment excavation rates (Ott et al., 1976; Griffis and Suchanek, 1991; Nickel1 and Atkinson, 1995).
Information on the actual behaviour of burrowing thalassinid shrimps is scarce and descriptions can be found scattered throughout the literature (e.g., MacGinitie, 1930, 1934; Lutze, 1938; Pearse, 1945; MacGinitie and MacGinitie, 1968; Ott et al., 1976; Dworschak, 1981, 1983, 1987; Scott et al., 1988; Witbaard and Duineveld, 1989; Nickel1 and Atkinson, 1995). In most studies detailed information on the total behaviour and allocation of time is lacking, which makes it difficult to causally link feeding strategies, burrow architectures and general ecology (Grifhs and Suchanek, 1991). This gap needs to be filled up, at least for a number of species belonging to the different guilds in feeding strategy and burrow architecture (Suchanek, 1985; Griffis and Suchanek, 199 1: Nickel1 and Atkinson, 1995).
Descriptions of the burrow architecture of the temperate callianassid Culliunussa
suhterruneu (Montagu, 1808) by several authors are mainly based on resin casting. Essentially, the burrow consists of a few vertical shafts descending from funnel-shaped openings or openings on top of a mound at the sea floor. The shafts lead down to spherical chambers at various depths. These chambers are interconnected by a lattice of horizontal and oblique tunnels (Witbaard and Duineveld, 1989; Atkinson and Nash, 1990, Rowden and Jones, 1995; Nickel1 and Atkinson, 1995). This architecture points at deposit feeding as the most plausible feeding mechanism (Griffis and Suchanek, 1991: Nickel1 and Atkinson, 1995). Support for this assumption is given by measurements indicating relatively high excavation rates (Rowden and Jones, 1993) and by single observations of sediment processing (Witbaard and Duineveld, 1989: Nickel1 and Atkinson, 1995). Witbaard and Duineveld (1989) do not exclude gardening as a possible feeding strategy, and Nickel1 and Atkinson ( 1995) assume C. subt~rruneu to be capable of filter feeding.
This study is dedicated to recording and describing the behaviour and allocation of time of Culliunussu subterrunea. in an attempt to relate burrow architecture with

E.J. Stamhuis et al. I J. Exp. Mar. Bid. Ed. 204 (1996) 225-239 227
burrowing activities. Detailed knowledge of C. subterranea’s behaviour will also provide insight in its foraging habits, and in the functions of different parts of the
burrow.
2. Methods
2.1. Collection and housing of experimental animals
Callianassa subterranea individuals were collected from several locations in the central North Sea, around 53”45’ N and 4”30’ E. Sampling trips were made in October 1988, May 1989, October 1989, June 1990, October 1991 and September 1992 with the Dutch Research Vessels Aurelia and Pelagia. The sampling trips were made in spring and in autumn, before the summer thermocline settled or after it had disappeared to avoid a temperature shock when the animals were lifted up from 40 m depth to the surface. The animals were sampled using a boxcorer which penetrated the sediment to a depth of about 0.5 m. The shrimps were washed out of the cores and single, undamaged individuals with an estimated rostrum-to-telson length of 20 to 45 mm were placed in plastic 0.5 1 jars filled with a 3-4 cm layer of sediment. Each jar was covered with 2 mm mesh size netting to prevent escape and submerged in a container which was continuously flushed with oxygenated seawater. During car transport to the laboratory, the water-filled containers with the jars were kept cool by covering them with insulating
blankets. In the laboratory, the jars were kept overnight in aquaria with artificial seawater. The
water was kept at a temperature of 12 “C and at a salinity of about 30%0, values close to those at the sampling sites. The next day each animal was removed into a 5 or 10 1 plastic (PE) container largely filled with sediment originating from the sampling sites. These containers were stored submerged in the sea water aquaria in a 12 “C dark climatized room. Usually, the animals started digging immediately and disappeared into the sediment within a few h. Only a small mound with a hole in the top was left as visible proof of their presence.
Animals were not individually fed, but the aquaria were enriched regularly by adding a mixture of ground dried algae and dried zooplankton to the water. Most of this organic material settled on the sediment, the remaining part enriched the water until it was removed by the filter system. This storage technique allowed us to keep animals alive in the laboratory for over 2 years, which is about their estimated life span (Rowden and Jones, 1994).
2.2. Experimental setup
To enable observation of Callianassa subterranea’s behaviour, single animals were introduced into narrow, sediment filled perspex cuvettes, 30 cm high X 30 cm long. The widths of the cuvettes were tailored to the body widths of the animals, ranging between 6 and 10 mm for shrimps with rostrom-telson lengths of 25 to 35 mm. These cuvettes were stored vertically, submerged in the seawater aquaria. Before the animals were

228 E.J. Stumhuis et ~1. I J. Elp. Mur. Bid. Ed. 204 (1996) 22-C2.79
introduced, the sediment filled cuvettes were kept undisturbed for several weeks to allow the sediment to bed down properly. Most animals started digging immediately after introduction, and established a (2-dimensional) burrow within 3 weeks. Due to the limited width of the cuvettes, the shafts, tunnels and chambers of the burrows were usually of the same diameter as the cuvette width, enabling observation of the behaviour of the animals in virtually the whole burrow when positioning the cuvettes against the front glass of the aquarium. The inside of the back panels of the cuvettes was roughened to prevent sliding of sediment or the collapse of burrows.
Cullianussu suhterrunecz individuals were observed starting at least 3 weeks after introducing them into the cuvette, using two macro video cameras displaying the animal four to eight times magnihed on a TV-screen. Both cameras could be moved horizontally and vertically, which enabled the animal to be followed throughout the burrow. One camera monitored the front side of the cuvette, the other one monitored the back side of the cuvette through a set of mirrors. Red light (A > 600 nm) was used for illumination because it did not provoke any visible reaction from the animals. In total 30 animals in cuvettes were studied. In 14 of these the behaviour was recorded explicitly.
2.3. Recording hehaviour
Preliminary observations of the behaviour of Culliunu.s.su suhterraneu resulted in an ethogram consisting of 12 exclusive behavioural states (Altmann, 1974). Each of these was assigned to one key of an electronic event recorder, capable of storing start and end of 2500 events at 0.1 s precision. Another key on the recorder was used to indicate when the animal was concealed from observation due to sediment covering the glass of the tunnel wall in some parts of the burrow. After each observation period, the recorded data were stored on a persona1 computer.
Observations were usually made in periods of 1 or 2 h, randomly distributed over the 24 h day. The duration of the observation periods always exceeded the average duration of single bouts or characteristic sequences of behavioural states, and was short enough to prevent introduction of errors due to fatigue of the observer (Altmann, 1974). Long-term periodicity was studied by recording five uninterrupted observation periods of 8, 24 (twice), 34 and 72 h. During these periods at least four observers, trained in advance to eliminate subjectivity in interpretation of behaviour, worked in shifts of 2 h.
2.4. Record validation
A data set of 55 one h records of five animals was used to test if the percentage of time animals were hidden had a significant effect on the relative percentage of time per behavioural state. In 15 out of the 55 records, the animals were visible 100% of the time, in the remaining 40 records less than 100%. First the records were labelled according to their percentages of time visible. Then the percentages of time per behavioural state per record were compensated for the time not-visible, assuming the same behaviour during hidden periods as during visible periods. Each of the 12 behavioural states was tested for positive correlation with the percentages of time hidden. This resulted in significantly influenced, and not significantly influenced states. Subsequently, the 40 records with

E.J. Stamhuis et al. I .I. Exp. Mar. Bid. Ed. 204 (1996) 225-239 229
visibility less than 100% were sorted according to their time visible, from high to low, and divided into groups of five records. The percentages of time per record spent to significantly influenced states, averaged per group were tested against the average of the same states of the 15 records with 100% visibility (Student’s r-test after root-arcsine transformation, (Y = 0.05). The tests resulted in a critical visibility percentage below which the recorded times per state were significantly affected by the time hidden. Only protocols with a visibility percentage above the critical value were used to analyze the behaviour of Callianassa subterranea.
2.5. Sequence analysis
The occurrence of typical sequences in the behaviour of Callianassa subterranea was analyzed by performing a type of Markov-chain analysis (Martin and Bateson, 1986) on a data set of 98 one h records of ten animals: the frequencies of duplets, triplets, quadruplets, quintuplets and sextuplets of successive behavioural states in the protocols were tested against the estimated chance of occurrence if it would be determined by purely stochastic processes. We used x2 probability-density distributions, spreading stochastically around f when having f degrees of freedom, to find levels of significant discrimination. When f equals the estimated frequency f,,, a defined sequence can occur stochastically, the 95% confidence interval can be found following normal procedures, setting the critical frequency fCri,. When the sequence occurs more often than the critical frequency, it is assumed to occur significantly more often than can be expected according to stochastics. All behavioural states were classified to functional behavioural units (classes) based on the sequence analysis. To improve the signal-to-noise ratio, sequences occurring in less than 10% of the records, as well as sub-units of higher order sequences were left out.
2.4. Frequencies and time allocation
The data of 98 validated one h records recorded of ten different animals were also processed to listings of recorded behavioural states grouped to the behavioural classes with their onset and end, and tables of frequencies, bout-lengths, interval-lengths and allocation of time per class per h. From these data averages were calculated and a time budget was composed to quantify the behaviour of Callianassa subterranea of average size (25-35 mm rostrum-telson length) under laboratory conditions.
2.7. Periodic@ analysis
Small-scale as well as large-scale periodicity (Naylor, 1988) was tested for by performing Fourier analysis (Martin and Bateson, 1986) on the behavioural data transformed to percentages of time active per period. This yielded a spectrum of amplitudes for all the possible periodicities between 2 X the period length (15 min to 1 h) and the total time of the protocol tested (e.g., 36 h). When the amplitude of an individual periodicity surpassed the average amplitude plus 2 S.D. (95% confidence interval) in all protocols, it was assumed to be a significant periodicity.

3. Results
-3.1. The ethograrn
The behaviour of single Calliana.ssa subterranea in narrow sediment filled cuvettes under laboratory conditions can be subdivided into 12 behavioural states, as listed in Table 1.
A few of the states summarized in Table 1 need some additional description. The Ventilation movements produce a flow through an inhalant shaft with an average velocity in the order of magnitude of a few mm per sec. Ventilation renews the water inside the burrow. The pleopod beat frequency during Pumping was estimated between 3 and 5 Hz. Pumping is always performed in one particular spherical chamber connected to the opening on top of a mound on the sea bed by a thin twining shaft. Sediment removed in this way is first transported to the spherical chamber until the chamber is tilled up about half. Then this sediment is stirred with large amplitude stirring movements. Subsequently, the animal performs a quick turn and protrudes its abdomen backwards in the chamber, after which it starts pumping fiercely. The water pumped in
Table I Behavioural stiltes of C~rlliunassa suhfrrrwwrr m burrows in narrow sediment tilled cuvettes under laboratory
conditions
Sitting
Walking
Turning
Stirring
Lifting
Carrying
Dropping
Tampmg
Bulldozering
Ventilating
Pumping
No visible motion, apart from antenna1 movements.
Walking through the burrow using all but the first pair of pereiopods. without carrying
sediment.
Changing the direction of locomotton by I X0” by performing a somersault followed by a roll
around the body axis, always performed in one of the spherical chambers.
Loosening the sediment roatro-ventrally of the cephalothorax using stirring movements of the
third pair of maxillipeds and the second pair of pereiopods. Sometimes the first pair of
pereiopods are also mvolved including the large cheliped.
Lifting a portton of loosened sediment with the third pair of maxillipeds and the first and
second pair of pereiopods.
Walking through the burrow while holding a portion of sediment with the third pair of
maxillipeds and the first pair of pereiopods.
Dropping the portion of sediment that was hfted and transported, most of the time on the
sediment surface, or at another part of the burrow.
Pressing or tamping at the sediment using the second and the third pair of pereiopod\,
thereby consolidating it or mcorporating it into the surroundin g sediment, usually the tunnel
wall.
Walking while pushing sediment m front of the cephalothorax using the first and the second
pair of pereiopods and the third pair of maxillipedh.
Relatively \low metachronous movements of the pairs of pleopods of the third. fourth and
fifth abdominal segments, with an average beat frequency of about I Hr.
Fierce ventilation movements of the pleopods of the third, fourth and fifth abdominal
segments. causing a sediment eruption from the shaft opening at the top of a mound.
Brushing various parts of the cephalothorax. the abdomen and all of the extremities, to clean
these parts from sediment, using the third maxillipeds and the setal brushes at the distal ends
of the third, fourth and fifth pair of pereiopods.

E.J. Stamhuis et al. I J. Exp. Mar. Bid. Ed. 204 (1996) 225-239 231
the twining shaft starts flowing slowly until the shaft is opened up by flushing away a plug of sediment remaining from the previous pumping bout. Finally a water/sediment mixture erupts from the opening at the sediment surface until the animal stops pumping. The sediment then beds down around the opening forming a smooth mound, and closing off the opening of the shaft again. Often during Grooming a typical posture is observed, the animal supporting itself with the first pair of pereiopods (mainly the large cheliped) sometimes assisted by the second pair of pereiopods, and the uropods + telson. This leaves the other pereiopods free to clean one another, the pleopods, the cephalothorax, the abdomen and the gill cavities on the lateral sides of the cephalothorax. Cleaning the mouthparts by rubbing them along each other, is also recorded as grooming.
Fig. 1 shows schematic drawings representing the behavioural states of C. subterrunea projected on a simplified typical burrow. The thin lines indicate the parts of the burrow where a behavioural state mainly takes place. Lifting and Walking are not depicted, because these would not be different from Carrying. Lifting is performed at the excavation site, and Walking is observed throughout the burrow, but mostly in the lattice of tunnels.
3.2. Record validation
The behavioural states which show a significant positive correlation with the percentage of time the animals were hidden are: Walking, Carrying, Dropping, Stirring
Fig. 1. Schematic drawings of the behavioural states of Calliunassa subterranea projected on a simplified burrow.

232 E.J. Sramhuis et al. I J. Exp. Mar. Bid. Ed. 204 (1996) 22_5-239
Table 2
Correlation coefficients of linear regressions between percentage of time spent to a behavioural state and
percentage of time not visible per I h protocol, from data of a set of 55 protocols (* = significant at 95%
confidence level)
Behavioural component Correlation coefficient Si.uuficance
Walk
Carry
Drop
Stir
Turn
Pump
Lift
Tamp
Groom
Bulldozer
Sit
Ventilate
r= 0.612
r= 0.338
r= 0.321
r = 0.302
r= 0.271
r= 0.154
r = o.oso
I = ~ 0.035
r = ~ 0.072
I = ~ 0.082 I = ~ 0.181
r = ~ 0.325
P <0.001*
P < 0.02*
P < 0.02*
P < 0.05*
P < o.os*
P > 0.2
P > 0.2
P > 0.2
P > 0.2
P > 0.2 P>O.l
P < 0.02*
and Turning (Table 2). Behavioural records with a relatively high percentage of time hidden apparently overestimate mobile states and underestimate non-mobile states (e.g., Sitting and Ventilating). When testing the total percentage of time spent on overesti- mated states from records with 100% visibility (15 one-hour records) against the ones with less than 100% visibility (40 one-hour records) divided into groups, a distinct level of significant deviation was found (see Table 3). All classes with visibilities above 90% of the time showed no significant deviation (in all cases P > 0.20), the classes with visibilities below 90% of the time deviated slightly or significantly from the records with 100% visibility (P 5 0.056).
Only records of animals with a visibility percentage higher than 90% were used for further analysis of the behaviour of Cullianasssa subterruneu. In total 273 h of protocol were recorded from a total of 14 different C. subterrunea individuals. In 129 h out of
Table 3
Total percentage of time (?SEM) spent to over-estimated states ordered according to visibility percentage. all
protocols with visibility percentage less than 100% were divided into groups of five protocols, the groups were
tested against the 100% visibility group (Student’s t-test after root-arcsine transformation, (Y = 0.05, H,, = no
difference, ** = P < 0.05, * = P = 0.05).
Visibility range (%time) Overestimated states (%time) Test result + significance (** / *)
100 (n = 15)
99.0-97.4
97.3-96.2
9.5.9-94.0
93.9-92.8
92.7-90. I 89. I pX6.4
85.9~83.8
83.0-54. I
21.1 (4.X)
30.3 (6.5)
30.2 (6.1)
17.7 (6.3) 26.4 (3.6)
27.X (4.8)
36.5 (4.2)
33.5 (4.4)
48.1 (6.6)
P = 0.202
P = 0.244
P = 0.830
P = 0.326 P = 0.309
P = 0.023**
P = 0.056* P = o.ooi**

E.J. Stamhuis et al. I .I. Exp. Mar. Biol. Ecol. 204 (1996) 225-239 233
these 273, recorded from 13 animals, the observed animal was visible for more than 90% of the time.
3.3. Frequency analysis and time allocation
The Markov-chain sequence analysis resulted in 100 sequences of 2, 3, 4, 5 or 6 states which occurred significantly more often than could be explained stochastically. After improving the signal-to-noise ratio, 18 sequences remained as listed in Table 4, ordered from high to low Markov-order.
The 12 behavioural states were grouped into six classes, based on the sequence analysis. This resulted in three classes composed of two or more states, and three classes consisting of only one state. The classes are described below with reference to relevant sequences in Table 4.
WANDERING = sequences of Walking and Turning, without carrying sediment (Table 4: 17 and 18).
BURROWING = sequences of sediment interaction and transport activities (Stirring, Lifting, Carrying, Dropping, Tamping, Bulldozering, Pumping), including Walking- bouts and short (a few s) Sitting-bouts when appearing in between two other burrowing activities (Table 4: 1, 2, 3, 4, 10, 11, 12, 13). Callianassa subterranea frequently transports sediment directly to the sediment surface. The animal approaches the shaft opening slowly, and stops just before appearing at the surface. After a few s of antenna1 movements the animal emerges partly to dump the sediment at the rim of the
Table 4
Average frequencies per h (?SEM) of duplets, triplets, quadruplets, quintuplets and sextuplets of successive
behavioural states of Calliunassa subterranea which were found to occur more often than expected according
to stochastics (x2 frequency-density distribution test, P < 0.05). F,” ,,o~ = averaged over records with > 10%
occurrence, F,_,, = averaged over all records
Sequence
1. Sit-Walk-Stir-Lift-Carry-Drop
2. Walk-Stir-Lift-Carry-Drop-Tamp
3. Stir-Lift-Carry-Drop-Tamp-Walk
4. Sit-Stir-Sit-Stir-Sit-Stir
5. Sit-Vent-Sit-Vent-Sit-Vent
6. Sit-Walk-Sit-Walk
7. Sit-Groom-Sit
8. Walk-Tamp-Walk
9. Tamp-Walk-Tamp
10. Walk-Stir-Walk
I 1. Tamp-Walk-Stir
12. Tamp-Walk-Sit
13. CarrySit-Carry
14. Groom-Sit-Groom 1.5. Walk-Groom-Walk
16. Vent-Walk
17. Walk-Turn
18. Turn-Walk
2.5 (0.3) 5.3 (0.9)
6.0 (1.1)
2.5 (0.5)
9.7 (2.9)
6.2 (0.5)
4.8 (0.7)
3.8 (1.1)
4.0 (1.3)
3.9 (0.4)
5.4 (0.8)
4.0 (0.6)
3.6 (0.7)
4.7 (0.8) 4.9 (0.7)
3.8 (0.3)
3.8 (0.4)
4.0 (0.4)
0.7 (0.2)
2.7 (0.7)
2.5 (0.7)
0.5 (0.2)
1.3 (1.1)
4.5 (0.4)
1.2 (0.4)
0.4 (0.4)
0.5 (0.4)
1.4 (0.2)
1.7 (0.4)
0.9 (0.3)
0.4 (0.2)
0.5 (0.3) 0.6 (0.2)
0.4 (0.1)
1.6 (0.3)
1.5 (0.2)

234 E.J. Stamhuis et al. I J. Exp. Mur. Bid. Ed. 204 (1996) 22-T-2.39
shaft-opening, sometimes bulldozering it away from the opening. Then it retreats backwards into the burrow until it arrives at a turning chamber. There it turns and goes back to the excavation location for a next run. This procedure of removing sediment is often observed during burrow construction or periods of reconstruction. The almost continuous supply of sediment during these periods causes growing mounds around the burrow openings. Sometimes sediment is used to partly till up rarely used old parts of the burrow. This sediment is collected again later and dumped at the surface. Special chambers connected with a thin vertical shaft to the top of a mound are also used as dumping sites. The sediment in these chambers is removed from the burrow by pumping it to the surface.
SURVEYING = sequences of Walking and Tamping, sometimes combined with Stirring (Table 4: 8, 9, 11 and 12). These combinations of walking and tamping are often observed when an animal is walking through its burrow meanwhile scanning the walls of the burrow with its second pair of antennae, extending almost one body-length in front of the animal. Burrow imperfections are repaired immediately by tamping, sometimes followed by smoothing the tunnel-lining with sediment collected near the place of damage.
GROOMING = Grooming. RESTING = Sitting, when not embedded in between Burrowing states. VENTILATING = Ventilating. No separate class FEEDING was distinguished, although sequences showing combi-
nations of Stirring with Sitting or Walking (see Table 4: 4, 10) indicated sediment processing for feeding purposes. Table 4 also shows that behavioural states were often interrupted by short periods of Resting (4, 5, 6, 7, 1.3, 14) or sometimes Walking (15, 16).
The functional description of Cullianassu~subterraneu’s behaviour resulting from the sequence analysis, was used to analyze the frequencies and durations of the behavioural classes and of the intervals between those classes (see Table 5). Finally a map of the allocation of time was composed (Fig. 2).
On average, Wandering is displayed most frequently, but due to very short bout lengths it does not occupy a major part of the time budget. Most of the time is spent Resting (large number of short bouts), Burrowing (small number of long bouts), and Grooming. Surveying takes little time, on average being displayed only once per h for a short period. The shrimp ventilates its burrow a few times per h. The hatch-patterns in
Table 5
Average frequencies per h, bout-length and interval-length (-tSEM) of behavioural classes averaged from 98
one hour behavioural records of ten Callianu.s.w .suhtrrruneu
Frequency Bout-length (s) Interval-length (s)
Wander
Rest
Burrow
Survey
Groom
Ventilate
26.8 (I .7) 17( I) I02 (22)
21.5 (1.2) 57 (14) 106 (14)
6.0 (0.4) I65 (12) 266 (35)
1.2 (0.2) 77 ( 4) 541 (58)
II.4 (1.3) 47( 3) I93 (48)
4.1 (0.6) 77 (II) 252 (44)

E.J. Stamhuis et al. I J. Exp. Mar. Biol. Ecol. 204 (1996) 225-239 235
40 r
35 -
30 -
E 25-
i! .- : : 20
Q 15- e a”
10 -
5-
E3 Walk TlNll
UIIUI Tamp hmzl Stir 0 Lift 0 Cany E33 Drop B Bulldozer m Pump r?zz3 Rest [LzLzi3 Groom eZa Ventilate
1 Wander Rest Burrow Survey Groom Ventilate
Fig. 2. General time budget of Callianassa subterranea under lab conditions in percentages of time per
behavioural class (?SEM) averaged from 98 one hour behavioural records of ten animals; hatch patterns refer
to the percentages of time per state.
Fig. 2 show that Wandering mainly consists of Walking (12.1+0.8%). During Burrow- ing most time is spent Tamping (13.5?2.1), but also Stirring (5.920.7) and Carrying (5.8+0.7), whereas Lifting (0.8t0.3), Dropping (0.3+0.1), Turning (0.62O.l), Bull- dozering (0.4?0.2) and Pumping (0.04-+0.02) take little time. Surveying mainly consists of Tamping (1.6kO.3) and less Walking (l.OtO.l).
3.4. Periodicity analysis
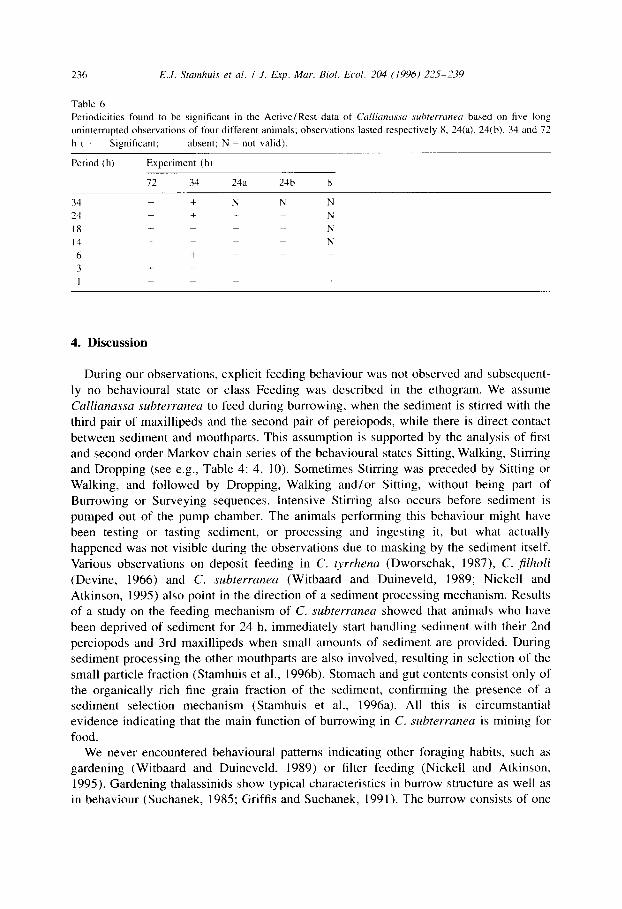
The Fourier analysis tests only for a restricted number of possible periods. In total we tested for 99 different periods between 72 h and 30 min based on the Active/Rest data of the four long term observations. Analysis of variance showed some significant periodicities in individual Cullianassa subterranea (see Table 6), but none of these periods was consistent for all or most of the animals.
3.5. The average behavioural pattern
On average, Callianassa subterranea is Burrowing every 7 min for about 3 min followed by Wandering (17 s), Resting (1 min) or sometimes Surveying (1.3 min), or combinations of these. Every 5 min the animal interrupts this pattern for a period of Grooming (46 s), and Ventilation of the burrow is performed every 14.5 min lasting about 1.3 min. This pattern is performed constantly, and no major significant periodicity was found. More than 40% of the time that an animal is active ( = not resting) is spent Burrowing, during which the animal is probably foraging as well. Pumping sediment to the sea floor surface is performed usually once, but sometimes up to a few times per day.
236 E.J. Starnhuis et al. 1 J. Ekp. MOT. Bid. Ecol. 204 (1996) 22.5-2.39
Table 6
Periodicities found to be significant in the Active/Rest data of Cdliuruuw .suhterranetr based on five long
uninterrupted observations of four different animals; observations lasted respectively 8, 24(a), 24(b), 34 and 72
h ( + = Significant; ~ = absent; N = not valid).
Period (h) Experiment (h)
72 34 24a 24b 8
34 + N N N
24 + t N 18 + N
14 + N
6 + 3 + ._
I _ +
4. Discussion
During our observations, explicit feeding behaviour was not observed and subsequent- ly no behavioural state or class Feeding was described in the ethogram. We assume Culliunassa suhterranea to feed during burrowing, when the sediment is stirred with the third pair of maxillipeds and the second pair of pereiopods, while there is direct contact between sediment and mouthparts. This assumption is supported by the analysis of first and second order Markov chain series of the behavioural states Sitting, Walking, Stirring and Dropping (see e.g., Table 4: 4, 10). Sometimes Stirring was preceded by Sitting or Walking, and followed by Dropping, Walking and/or Sitting, without being part of Burrowing or Surveying sequences. Intensive Stirring also occurs before sediment is pumped out of the pump chamber. The animals performing this behaviour might have been testing or tasting sediment, or processing and ingesting it, but what actually happened was not visible during the observations due to masking by the sediment itself. Various observations on deposit feeding in C. tyrrhena (Dworschak, 1987) C. jilholi
(Devine, 1966) and C. subterrunea (Witbaard and Duineveld, 1989; Nickel1 and Atkinson, 1995) also point in the direction of a sediment processing mechanism. Results of a study on the feeding mechanism of C. subterranea showed that animals who have been deprived of sediment for 24 h, immediately start handling sediment with their 2nd pereiopods and 3rd maxillipeds when small amounts of sediment are provided. During sediment processing the other mouthparts are also involved, resulting in selection of the small particle fraction (Stamhuis et al., 1996b). Stomach and gut contents consist only of the organically rich fine grain fraction of the sediment, confirming the presence of a sediment selection mechanism (Stamhuis et al., 1996a). All this is circumstantial evidence indicating that the main function of burrowing in C. subterrunea is mining for food.
We never encountered behavioural patterns indicating other foraging habits, such as gardening (Witbaard and Duineveld, 1989) or tilter feeding (Nickel1 and Atkinson, 1995). Gardening thalassinids show typical characteristics in burrow structure as well as in behaviour (Suchanek, 1985; Griffis and Suchanek, 1991). The burrow consists of one

E.J. Stamhuis et al. I J. Exp. Mar. Bid. Ed. 204 (1996) 225-239 237
or a few vertical shafts running down to partitioned storage chambers. The gardening
shrimps collect and transport organic debris to these chambers where micro-organisms are cultured and harvested. The architecture of the burrows of Callianassa subterranea
is quite different, and organic debris is neither transported to special chambers nor foraged upon. Therefore we exclude gardening as a probable trophic mode.
Nickel1 and Atkinson (1995) observed how Caflianassa subterranea’s setal structures on the first and second pereiopods caught suspended particles when sediment was pumped out. They describe how the shrimps use the third maxillipeds to remove the particles from the setae, and ingest these. The behaviour described is interpreted as filter feeding. We have several reasons to rate this behaviour as Grooming. Our observations suggest that during pumping these fine particles accidentally adhere to the long setae on the second pereiopods, as well as to fringes of shorter setae on other appendages. At the end of a pumping bout, the animals often quickly groom the first pairs of pereiopods and the antennae using the third maxillipeds. Sometimes, more thorough grooming, involving other appendages, occurs as well. The animal might indeed ingest removed particulate material, but it is difficult to actually see that happening. Even on high (20 X ) magnification video recordings from the rostra1 and the ventral side, it was difficult to see if sediment actually was ingested (Stamhuis et al., 1996b).
The pereiopodal setal fringes of C. subterranea differ from those of obligate filter feeding thalassinids such as most Upogebiae. Filter feeding Upogebiae have large and densely implanted plumose setal structures on their first and second pereiopods forming a basket to filter the pleopod-generated flow through the burrow for food particles (Scott et al., 1988; personal observations on U. detaura). The basket-shaped filter covers the whole diameter of the tube during active filter feeding preventing flow from evading the filter. C. subterranea’s setal fringes do not have the right size to cover the whole diameter of the tube, and the density of the setae is low. Most of the flow would pass around the setae, making the filter inefficient (Vogel, 1981), and filter feeding relatively expensive (Riisgaard and Larsen, 1995). Furthermore, pumping in C. subterranea does not last longer than 10 s and ventilating not longer than 1.5 min. This is long enough to flush out sediment or to renew the burrow water, but does not seem to be sufficient to filter food from the overlaying water. These arguments make filter feeding an unlikely trophic mode in C. subterranea. The setal fringes on the first pereiopods are used for Lifting and Carrying sediment during Burrowing.
We observed three ways of removing sediment from excavation sites, in Callianassa subterranea. In young burrows and in burrows under heavy reconstruction, most sediment is directly transported to the sea floor surface and dumped there. In established burrows, most sediment is transported to the pump chamber and flushed to the sea floor once or a few times per day. Sometimes, but not regularly, excavated sediment is dumped in an old tunnel.
Sequences of behavioural states including Pumping were not encountered in the results of the sequence analyses, because the applied method did not allow detection of rarely occurring sequences. One of these rare sequences included Pumping, and these showed a stereotypical pattern: sediment was Dropped in the pump chamber after a number of Burrowing bouts. Subsequently, a number of Sitting-Stirring duplets were performed followed by a Walking-Turning-Walking triplet as the animal walked to

238 E.J. Stumhuis et al. I J. Exp. Mar. Btol. Ed. 204 (1996) 22_7-239
another spherical chamber, turned, and returned backwards to the pump chamber, where it started Pumping.
No courtship or other explicit mating behaviour was observed during this study, presumably because the animals were always kept in separate containers. Although these containers were open at the top, only rarely an animal escaped from its container moving to another one. When two or more animals were brought together in an experimental setup, they immediately started to demolish one another. This was also observed by Rowden and Jones (1994), who suggest this behaviour to be functional in the regulation of population density. We believe that this behaviour might be induced by stress as a result of handling.
Sex determination required rather intensive handling, risking damage to the animals. Nevertheless we sexed six individuals in total, four males and two females. The behavioural patterns did not differ between the sexes. Variations in time allocation among or between females and males did not differ from variations between individuals regardless of their sex.
The behavioural study presented here covered a period of several years. In this period the behaviour of more than 30 animals was studied under constant laboratory conditions and in 14 of these the behaviour was explicitly recorded. Callianassa suhterranea’s
behaviour appeared to be very stereotypical during the whole experimental period and we never needed to change or adapt the ethogram. Also the time budgets of the different animals or groups of animals were very much the same and often statistically indistinguishable. We therefore assume our results to reflect closely the natural behaviour of C. subtrrranea at the Oyster Grounds.
Acknowledgments
We would like to thank the crews of the Research Vessels Aurelia and Pelagia for their assistance during the sampling trips. J.H. Koop, P.J. Geerlink, M. Staal, M. Hesseling, H. Zemmelink, F. Zegwaard, M. Renckema, P. Agterberg, M. Palmen and H. Lippert are gratefully acknowledged for their shifts in the long-period observations. W. Beukema, E. Paap, R. Vol and H. v.d. Hengelaar designed and assembled the event recorders.
References
Altmann, .I., 1974. Observational study of behaviour: sampling methods. Bchaviour. Vol. 49, pp. 227-267.
Atkinson and Nash, 1990. J. Exp. Mar. Bid. Em/., Vol. 52, pp. 25-35.
Devine, C.E., 1966. Ecology of Callkmussu .filholi Milne Edwards I X78 (Crustacea: Thalassinidea). Trams. K. Ser. N. z. zoo/., Vol. 8, pp. 93-l IO.
Dworschak, P.C., 1981. The pumping rates of the burrowing shrimp Upogehiu pu.silla (Petagna) (Decapoda:
Thalassinidea).
Dworschak, P.C., 1983. The biology of Upo~&iu pusilla (Petagna) (Decapoda: Thalassinidea) I. The Burrows.
P.S.Z.N.lr Marinr Ecology, Vol. 4, pp. 19-43.
Dworschak, P.C., 1987. Feeding behaviour of Upo&icr pusillu and Cdlianussu tyduwr (Crustacea,
Decapoda, Thalassinidea). Inv. Pesy., Vol. 5 I (suppl. I ). pp. 421-429.

E.J. Stamhuis et al. I J. Exp. Mar. Biol. Ecol. 204 (1996) 225-239 239
Griffis, R.B. and T.H. Suchanek, 1991. A model of burrow architecture and trophic modes in thalassinidean
shrimp (Decapoda: Thalassinidea). Mar. Ecol. Progr. Ser., Vol. 79, pp. 171-183.
Lutze, J., 1938. Ueber Systematik, Entwicklung und Oekologie von Callianassa. Helg. Wiss. Meeresunters., Vol. 1, pp. 162-190.
MacGinitie, G.E., 1930. The natural history of the mud shrimp Upogebia pugettensis (Dana). Ann. Mag. Nat. Hi.vt., Vol. 6, pp. 36-44.
MacGinitie, G.E., 1934. The natural history of Callianassa californiensis Dana. Am. Midl. Nat., Vol. 15, pp.
166-177.
MacGinitie, G.E. and N. MacGinitie, 1968. Natural History of Marine Animals. McGraw-Hill, New York.
Martin, P. and P. Bateson, 1986. Measuring Behaviour, an introductory guide. Cambridge Univ. Press,
Cambridge.
Montagu, G., 1808. Description of several marine animals found on the south coast of Devonshire. Trans. Linn. Sot. Land., (ZooI.),Vol. 9, pp. 81-114.
Naylor, E., 1988. Rhythmic behaviour of decapod crustaceans. Symp. Zool. Sot. Lond.,Vol. 59, pp. 177-199.
Nickel], L.A. and R.J.A. Atkinson, 1995. Functional morphology of burrows and trophic modes of three
thalassinidean shrimp species, and a new approach to the classification of thalassinidean burrow
morphology. Mar. Ecol. Progr. Ser., Vol. 128, pp. 181-197.
Ott, J.A., B. Fuchs, R. Fuchs and A. Malasek, 1976. Observations on the biology of Callianassu stebbingi Borrodaille and Upogebia littoralis Risso and their effect upon the sediment. Senckenberg. Marit., Vol. 8,
pp. 61-79.
Pearse, AS., 1945. Ecology of Upogebia afinis (Say). Ecology, Vol. 26, pp. 303-305. Riisgaard, H.U. and P.S. Larsen, 1995. Filter-feeding in marine macro-invertebrates: pump characteristics,
modelling and energy costs. Biol. Rev., Vol. 70, pp. 67-106.
Rowden, A.A. and M.B. Jones, 1993. Critical evaluation of sediment turnover estimates for Callianassidae
(Decapoda: Thalassinidea). J. Exp. Mar. Biol. Ecol., Vol. 173, pp. 265-272.
Rowden, A.A. and M.B. Jones, 1994. A contribution to the biology of the burrowing mud shrimp Callianassa subterranea (Decapoda: Thalassinidea). J. Mar. Biol. Ass. U.K., Vol. 74, pp. 623-635.
Rowden, A.A. and M.B. Jones, 1995. The burrow structure of the mud shrimp Callianassa subterranea (Decapoda: Thalassinidea) from the North Sea. .I. Nut. Hist., Vol. 29, pp. 1155-l 165.
Scott, P.J.B., H.M. Reiswig and B.M. Marcotte, 1988. Ecology, functional morphology, behaviour and feeding
in coral- and sponge-boring species of Upogebia (Crustacea: Decapoda: Thalassinidea). Can. J. Zool., Vol.
66, pp. 483-495. Suchanek, T., 1985. Thalassinid shrimp burrows: ecological significance of species specific architecture. Proc.
5th lnt. Coral Reqf Congr. at Tahiti, Vol. 5, pp. 205-210.
Stamhuis, E.J., J.J. Videler and P.A.W.J. de Wilde, 1996a. Optimal foraging in the thalassinid shrimp
Cullianassa suhterranea: Improving food quality. by selection on grain size. in prep.
Stamhuis, E.J., B. Dauwe and J.J. Videler, (1996b). Morphology and function of the feeding appendages in the
deposit feeding thalassinid shrimp Callianassa subterranea. in prep.
Vogel, S. 1981. Life in moving fluids: The physical biology of flow. Princeton Univ. Press, Princeton, NJ.
Witbaard, R. and C.G.A. Duineveld, 1989. Some aspects of the biology and ecology of the burrowing shrimp
Culliunassa subterranea (Montagu) (Thalassinidea) from the southern North Sea. Sarsia, Vol. 74, pp.
209-219.





























