Embed Size (px)
Citation preview

Br
DR
a
ARRA
KHSDECRVM
1
urfificicef
ilpdabal
0d
Enzyme and Microbial Technology 47 (2010) 355–362
Contents lists available at ScienceDirect
Enzyme and Microbial Technology
journa l homepage: www.e lsev ier .com/ locate /emt
ehavior of different monocomponent endoglucanases on the accessibility andeactivity of dissolving-grade pulps for viscose process
avid Ibarra ∗, Viviana Köpcke, Monica Ekoyal Institute of Technology, Dept. of Fiber and Polymer Technology, Teknikringen 56, SE 10044 Stockholm, Sweden
r t i c l e i n f o
rticle history:eceived 24 May 2010eceived in revised form 29 July 2010ccepted 29 July 2010
eywords:ardwoodoftwood
a b s t r a c t
Three different commercial monocomponent endoglucanases, with and without a cellulose-bindingdomain (CBD) and differences in their glycosidic hydrolysis mechanisms, were compared with respect totheir ability to enhance the accessibility and reactivity of dissolving-grade pulps for viscose production.Hardwood (eucalyptus) and softwood (mixture of Norway spruce and Scots pine) commercial dried andnever-dried bleached sulfite dissolving pulps were used for this purpose. The effects of the enzymatictreatments on pulps were studied by reactivity, according to Fock’s method, and viscosity measurements,and recording of molecular weight distributions. Among the different assayed enzymes, endoglucanase
issolving-grade pulpndoglucanase (EG)ellulose-binding domain (CBD)eactivityiscosityolecular weight distribution (MWD)
with a CBD and an inverting hydrolysis mechanism was found to be the most effective in increasingthe reactivity of both pulps. Simultaneously, the viscosity decreased, being more marked for softwooddissolving pulp. A narrower molecular weight distribution, with a great reduction in the amount of long-chain cellulose molecules was observed in both pulps, being more pronounced for softwood dissolvingpulp. By contrast, endoglucanase without a CBD and a retaining hydrolysis mechanism showed a barleyenhancement of the studied properties. The effects of the different endoglucanase treatments were more
dried
pronounced when never-. Introduction
Dissolving-grade pulps are used as raw material in the man-facture of different cellulose-derived products, including viscoseayon, the first commercially manufactured regenerated celluloseber. In the viscose process, cellulose is treated with carbon disul-de (highly toxic solvent) in the presence of a base to produceellulose xanthate [1]. In contrast to paper-grade pulps, dissolv-ng pulps must contain a high content of cellulose (90–99%), lowontent of hemicelluloses (2–4%), and traces of residual lignin,xtractives and minerals. Low degree of polymerization and a uni-orm molecular weight distribution are also desired [2].
The processability of a dissolving pulp is often characterized byts reactivity towards solvents and reactants [2]. High reactivity cel-ulose could improve the homogeneity and quality of cellulose-endroducts and lower the demands of reactants, e.g. use of carbonisulfide in the viscose manufacture, reducing production costs
nd the environmental impact. However, increasing the accessi-ility and reactivity of cellulose is not a simple task. Cellulose hascompact fibrillar structure as a result of intra- and intermolecu-ar hydrogen bonds and hydrophobic interactions [3]. Only the less
∗ Corresponding author. Tel.: +46 8 790 8311; fax: +46 8 790 6166.E-mail address: [email protected] (D. Ibarra).
141-0229/$ – see front matter © 2010 Elsevier Inc. All rights reserved.oi:10.1016/j.enzmictec.2010.07.016
dissolving pulps were used.© 2010 Elsevier Inc. All rights reserved.
ordered cellulose located between and on the surface of the fibrilaggregates is accessible to chemicals [4]. In addition, the acces-sibility and reactivity of cellulose are influenced by the chemicalcomposition of the raw materials, the distribution, the structure,and morphology of the fibers [4]. Various treatments have beenassessed to increase the reactivity of the cellulose [4], including theuse of enzymes such as monocomponent endoglucanases [5–11].
Endo-1,4-�-glucanases (EC 3.2.1.4) are an essential compo-nent in the cellulose degradation, generating new ends randomlywithin the polysaccharide chain, which in turn are substrate for theexo-1,4-�-glucanases (cellobiohydrolases; EC 3.2.1.91) [12,13]. Theendoglucanases typically act on less ordered cellulose whereas cel-lobiohydrolases degrade preferentially crystalline cellulose-highlyordered-in a progressive manner [14]. In general, endoglucanaseshydrolyze the �-1,4 glycosidic bonds via two major hydrolysismechanisms, i.e. inversion and retention of the configuration of the�-glycosidic bond [15,16]. Inverting mechanism use a direct dis-placement of the carbohydrate leaving group, which consequentlyresults in the inversion of the anomeric configuration. Retainingmechanism use a double-displacement mechanism with retention
of the anomeric configuration. Both mechanisms use two carboxylresidues on opposite sites of the carbohydrate plane which act asa general acid and a general base (for inverting mechanism), or asa general acid/base and a nucleophile/leaving group (for retainingmechanism), respectively. In addition to hydrolysis mechanisms,
356 D. Ibarra et al. / Enzyme and Microbial Technology 47 (2010) 355–362
Table 1Characteristics of the three monocomponent endoglucanases used.
Enzyme Component Origin CBD Hydrolysis mechanism Activity (ECU/g)a
Novozyme 476 EG V (45) Humicola insolens Yes Inverting 5000Novozyme 613 EG I (7) Humicola insolens No Retaining 2500
ndo Ca
ttidbc
bdbiplp
dtmfinTiamtb
2
2
d(sincpPfafia
2
ofi
2
((dewdoe
Novozyme 51603 EG V (45) Humicola insolens
a The cellulolytic activity was determined by the manufacturer and expressed in Essignment of catalytic domain to corresponding structural family.
he cellulose hydrolysis efficiency depends largely on the adsorp-ion capacity of these cellulases to the substrate [17]. This capacitys determined by their modular structure, containing a catalyticomain responsible for the hydrolysis reaction and a cellulose-inding domain (CBD), which is a binding of the enzymes to theellulose, both connected by a flexible linker peptide [18].
The mechanism of pulps activation by endoglucanases has noteen completely elucidated, although some hypotheses have beenescribed [6,13]. The attack on the less ordered cellulose regionsy the endoglucanase leads to a fiber wall swelling and thus an
ncrease in accessibility towards solvents and reactants. Anotherossibility is the degradation of cellulose II, less reactive than cel-
ulose I, by the endoglucanase, which might result in an increase ofulp reactivity.
In order to evaluate the role of catalytic and cellulose-bindingomains of endoglucanase enzymes on the cellulose activation,hree commercial monocomponent endoglucanases with different
odular structures were assayed on dissolving pulps. In addition,ber type (hardwood or softwood), and fiber history (dried orever-dried) were studied with respect to their activation mode.he effects of these enzymes on pulps were studied by reactiv-ty, according to Fock’s method [19], and viscosity measurements,nd recording molecular weight distribution by size exclusion chro-atography (SEC). Scanning electron microscopy (SEM) was used
o visualize morphological changes in the surface fibers producedy the endoglucanases action.
. Materials and methods
.1. Pulp samples
Commercial dried and never-dried totally chlorine-free (TCF) bleached sulfiteissolving-grade pulps from eucalyptus (Eucalyptus globulus), provided by SniaceSpain), and from a mixture of Norway spruce (Picea abies) and Scots pine (Pinusylvestris) (respectively 60%/40%), provided by Domsjö Fabriker AB (Sweden), werenvestigated. In general, these pulps presented a low kappa number (0.6–1), bright-ess higher 91% ISO, hemicelluloses content lower 2.4%, R18 higher 94.5%, and alphaellulose content higher 91%. The viscosity values were different, depending on theulp (460 and 560 mL/g for hardwood and softwood dissolving pulps, respectively).rior to the treatments, the sheets of dried pulp were maintained in deionized wateror 24 h, disintegrated in Lorentzen & Wettre equipment at 1.5% (w/v) consistencynd 30,000 revolutions, according to the ISO standard 5263-1:2004, and were finallyltrated. Never-dried pulp was stored as such in deep freeze (−20 ◦C) and defrostedt 4 ◦C for 24 h before the treatments.
.2. Enzymes
Three different commercial monocomponent endoglucanases (Table 1) werebtained from Novozymes (Bagsvaerd, Denmark). These enzymes are producedrom a genetically modified Aspergillus species, with an inserted gene from Humicolansolens, and differ either on the presence of a CBD or on the catalytic mechanism.
.3. Enzymatic treatments
Enzymatic treatments were carried out according to Engström et al. [7] on 10 gdry weight) of pulp at 3% (w/v) pulp consistency in phosphate buffer solution11 mM NaH2PO4 and 9 mM Na2HPO4), pH 7 (the optimal pH of the enzymes, as
escribed by the manufacturer Novozymes). For a homogeneous distribution, thenzymes were added to the buffer and then to the pulp. The enzymatic treatmentsere performed in plastic bags in a water bath at 50 ◦C, and 1 h, testing two differentosages (30 and 100 ECU/g dry weight pulp). The samples were periodically takenut of the bath and kneaded for 10–15 s to mix the sample. After treatment, thenzymes were denatured by filtration on a Büchner funnel and mixed with deion-No Inverting 5000
ellulase Units (ECU) per unit mass of material. The number in parentheses indicates
ized water at 90 ◦C. The treated pulps were placed in a 90 ◦C water bath for 30 minand subsequently filtered and washed with 1000 mL of deionized water. As a control,pulps were treated under identical conditions without enzymes.
2.4. Reactivity measurements
The reactivity of the treated pulps was analyzed according to a slightly modifiedversion of Fock’s method [19]. This test is a micro-scale process similar to the viscoseprocess, i.e. cellulose swelling in alkali, transformation to cellulose xanthate withCS2, which makes the cellulose polymer soluble, and removal of the –CS2 group indiluted sulfuric acid forming re-precipitated cellulose fibers [1]. Subsequently, theregenerated cellulose is oxidized and titrated [19]. Prior to Fock analysis, the treatedpulps were dried at 50 ◦C.
2.4.1. Step 1. Preparation of viscose from treated pulps and collection ofregenerated cellulose
0.5 g of pulp samples were weight in a 100 mL Erlenmeyer with a stopper. Fiftymilliliters of 9% (w/v) NaOH and 1.3 mL of CS2 were added, and the solutions werestirred with a magnetic stirrer (300 rpm) for 4 h at room temperature. The solu-tion was diluted to 100 g using deionized water and carefully shaken. The solutionwas then left for 2 h in order to allow any undissolved cellulose to settle. An aliquot(10 mL) from the upper clear solution was then transferred to another 100 mL Erlen-meyer flask and neutralized using 29% (w/v) H2SO4. The yellow solution turnedtransparent and was left overnight in a fume cupboard.
2.4.2. Step 2. Oxidation and titration of the regenerated celluloseThe regenerated cellulose samples were mixed with 20 mL of 68% (w/v) H2SO4
and stirred with a magnetic stirrer for 1 h. The milky solution was diluted to 50 mLwith deionized water. Ten milliliters of 1N K2Cr2O7 was added, and the solutionwas refluxed for 1 h to fully oxidize the regenerated cellulose and thereby clear thesolution. The solution was transferred to a 100 mL measuring flask and diluted withdeionized water. Forty milliliters of the solution was then transferred to a 250 mLbeaker containing 0.5 g of KI, stirred with a magnetic stirrer, and titrated with 0.1Nof Na2S2O3. When the brown solution started to change the color, 1.5 g of starch wasadded, and the solution turned blue-violet. The titration continued until all of the I2
was reduced and the solution turned pale blue. The volume of Na2S2O3 required ineach case was determined.
Reactivity measurements were carried out in triplicate and expressed as theregenerated cellulose yield (Eq. (1)):
X = (100)9.62a M(V1C1 − (V2C2100/40b)/6)4Y
(1)
where X is the reacted cellulose (%), Y is the weight of sample (g), M is the molecularmass of glucopyranosyl residue, C6H10O5 (162 g/mol), V1 is the volume of addedK2Cr2O7 (L), V2 is the volume of titrated Na2S2O3 (L), C1 is the concentration ofK2Cr2O7 (mol/L), C2 is the concentration of Na2S2O3 (mol/L), a is the first dilution to100 g and outtake of 10 mL (10.4 g) = 100/10.4 = 9.62, and b is the second dilution ofthe sample to 100 mL and outtake of 40 mL = 100/40.
2.5. Determination of viscosity
The viscosity of the treated pulps, defined as the intrinsic viscosity of a sample ofcellulose dissolved in a diluted solution of cupri-ethylenendiamene, was calculatedaccording to SCAN-CM 15:99. Viscosity measurements were carried out in duplicate.
2.6. Size exclusion chromatography
Prior to characterization on a SEC system, the treated pulps were dissolved[20]. Fifteen milligrams of treated pulp samples were activated for 1 h in 15 mLof deionized water at 4 ◦C. The excess water was removed, and the pulps weresolvent-exchanged once with methanol and then three times with DMAc (N,N-dimethylacetamide) with an intermediate equilibration period of 30 min. Then,1.9 mL of 8% (w/v) LiCl/DMAc was added, and the samples were left at 4 ◦C for 5
days with mild magnetic stirring. Finally, the samples were diluted to 0.5% (w/v)LiCl by the addition of 27.4 mL of DMAc. The dissolved treated pulps were filteredthrough a 0.45 �m PTFE (poly(tetrafluorethylene)) filter before chromatographiccharacterization.Samples were analyzed on a SEC system consisting of a DGU-20A3 degasser(Shimadzu), a LC-20AD liquid chromatograph (Shimadzu), a CT-20A column oven
robial Technology 47 (2010) 355–362 357
(rsoo(1mucei((t
2
wst
3
epotWcueo
3
dshrocccIoDoxrcli
sd1eh8ercAlfi
D. Ibarra et al. / Enzyme and Mic
Shimadzu) equipped with a Rheodyne 7725i fixed-loop (100 �L) and a RID-10Aefractive index detector (Shimadzu). The injection volume was 100 �L, and theeparations were performed at 80 ◦C with 0.5% (w/v) LiCl/DMAc at a flow ratef 0.5 mL/min on four Mixed-A 20 �m columns (7.5 mm × 300 mm, Polymer Lab-ratories) connected in series and preceded by a Mixed-A 20 �m guard column7.5 mm × 50 mm, Polymer Laboratories). Pullulan standards 800 K, 400 K, 200 K,10 K, 50 K, 22 K, 12 K, 6 K, 1.3 K and 320 Da (Fluka) were used to calibrate theolecular weight. The linear coefficient of determination (r2) between the molec-
lar weight of the standards and the elution time was 0.996. Data acquisition andalculation were carried out with LC Solution software (Shimadzu). A numericalvaluation from the molecular weight distribution (MWD) of samples was then stud-ed, determining the number-average (Mn) and weight-average molecular weightMw) [21]. The number-average (DPn) and weight-average degree of polymerizationDPw) were calculated by dividing the respective molecular weight values by 162,he molecular weight of glucopyranosyl residue.
.7. Scanning electron microscopy
For micro-structural analysis of the treated pulps, the samples were analyzedith a Hitachi s-4300 field emission scanning electron microscope (FE-SEM). The
pecimens were fixed on a metal stub with carbon paint and coated with a 3 nmhick gold layer using an Agar HR Sputter coater.
. Results and discussion
Monocomponent endoglucanases are considered to be the mostfficient enzymes in the activation of dissolving and paper-gradeulps from different raw materials [6–11]. Better understandingf the action of their individual domains in the cellulose activa-ion could proceed towards its applicability in the viscose process.
ith this purpose, the present paper studies the behavior of threeommercial monocomponent endoglucanases with different mod-lar structures (Table 1) on dissolving-grade pulps, as well as theirffect on the cellulose activation according to fiber type (hardwoodr softwood) and fiber history (dried or never-dried).
.1. Effect of endoglucanase treatment on reactivity
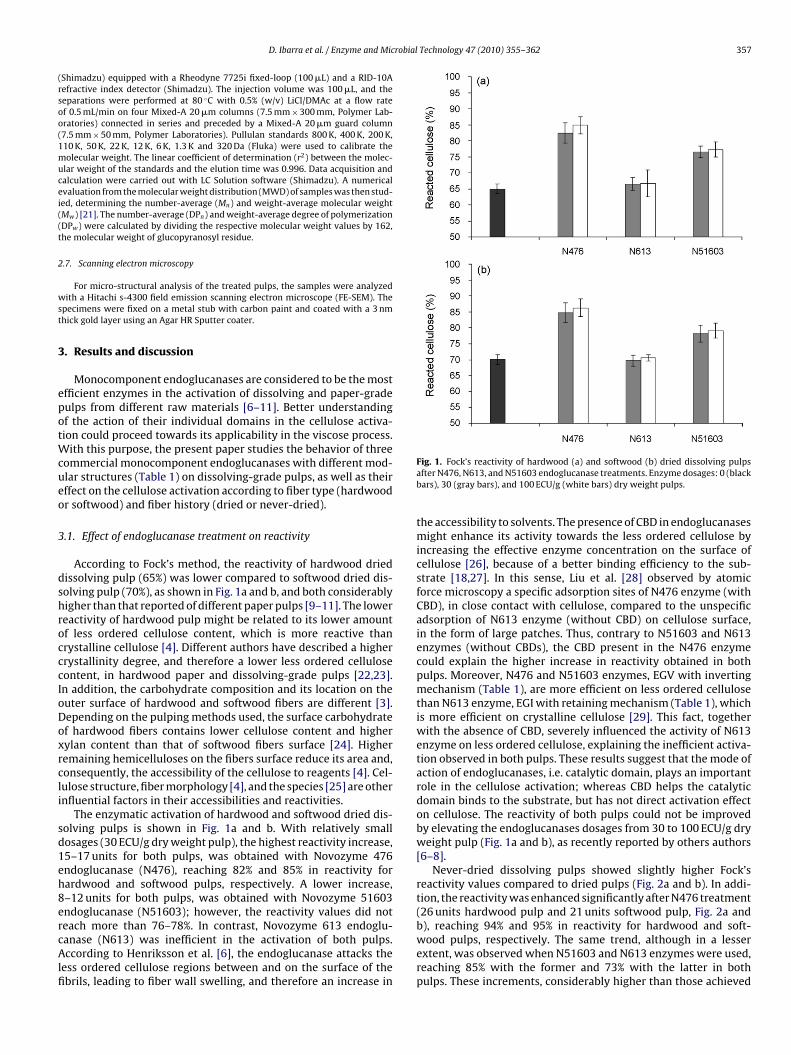
According to Fock’s method, the reactivity of hardwood driedissolving pulp (65%) was lower compared to softwood dried dis-olving pulp (70%), as shown in Fig. 1a and b, and both considerablyigher than that reported of different paper pulps [9–11]. The lowereactivity of hardwood pulp might be related to its lower amountf less ordered cellulose content, which is more reactive thanrystalline cellulose [4]. Different authors have described a higherrystallinity degree, and therefore a lower less ordered celluloseontent, in hardwood paper and dissolving-grade pulps [22,23].n addition, the carbohydrate composition and its location on theuter surface of hardwood and softwood fibers are different [3].epending on the pulping methods used, the surface carbohydratef hardwood fibers contains lower cellulose content and higherylan content than that of softwood fibers surface [24]. Higheremaining hemicelluloses on the fibers surface reduce its area and,onsequently, the accessibility of the cellulose to reagents [4]. Cel-ulose structure, fiber morphology [4], and the species [25] are othernfluential factors in their accessibilities and reactivities.
The enzymatic activation of hardwood and softwood dried dis-olving pulps is shown in Fig. 1a and b. With relatively smallosages (30 ECU/g dry weight pulp), the highest reactivity increase,5–17 units for both pulps, was obtained with Novozyme 476ndoglucanase (N476), reaching 82% and 85% in reactivity forardwood and softwood pulps, respectively. A lower increase,–12 units for both pulps, was obtained with Novozyme 51603ndoglucanase (N51603); however, the reactivity values did not
each more than 76–78%. In contrast, Novozyme 613 endoglu-anase (N613) was inefficient in the activation of both pulps.ccording to Henriksson et al. [6], the endoglucanase attacks theess ordered cellulose regions between and on the surface of thebrils, leading to fiber wall swelling, and therefore an increase in
Fig. 1. Fock’s reactivity of hardwood (a) and softwood (b) dried dissolving pulpsafter N476, N613, and N51603 endoglucanase treatments. Enzyme dosages: 0 (blackbars), 30 (gray bars), and 100 ECU/g (white bars) dry weight pulps.
the accessibility to solvents. The presence of CBD in endoglucanasesmight enhance its activity towards the less ordered cellulose byincreasing the effective enzyme concentration on the surface ofcellulose [26], because of a better binding efficiency to the sub-strate [18,27]. In this sense, Liu et al. [28] observed by atomicforce microscopy a specific adsorption sites of N476 enzyme (withCBD), in close contact with cellulose, compared to the unspecificadsorption of N613 enzyme (without CBD) on cellulose surface,in the form of large patches. Thus, contrary to N51603 and N613enzymes (without CBDs), the CBD present in the N476 enzymecould explain the higher increase in reactivity obtained in bothpulps. Moreover, N476 and N51603 enzymes, EGV with invertingmechanism (Table 1), are more efficient on less ordered cellulosethan N613 enzyme, EGI with retaining mechanism (Table 1), whichis more efficient on crystalline cellulose [29]. This fact, togetherwith the absence of CBD, severely influenced the activity of N613enzyme on less ordered cellulose, explaining the inefficient activa-tion observed in both pulps. These results suggest that the mode ofaction of endoglucanases, i.e. catalytic domain, plays an importantrole in the cellulose activation; whereas CBD helps the catalyticdomain binds to the substrate, but has not direct activation effecton cellulose. The reactivity of both pulps could not be improvedby elevating the endoglucanases dosages from 30 to 100 ECU/g dryweight pulp (Fig. 1a and b), as recently reported by others authors[6–8].
Never-dried dissolving pulps showed slightly higher Fock’sreactivity values compared to dried pulps (Fig. 2a and b). In addi-tion, the reactivity was enhanced significantly after N476 treatment(26 units hardwood pulp and 21 units softwood pulp, Fig. 2a andb), reaching 94% and 95% in reactivity for hardwood and soft-
wood pulps, respectively. The same trend, although in a lesserextent, was observed when N51603 and N613 enzymes were used,reaching 85% with the former and 73% with the latter in bothpulps. These increments, considerably higher than those achieved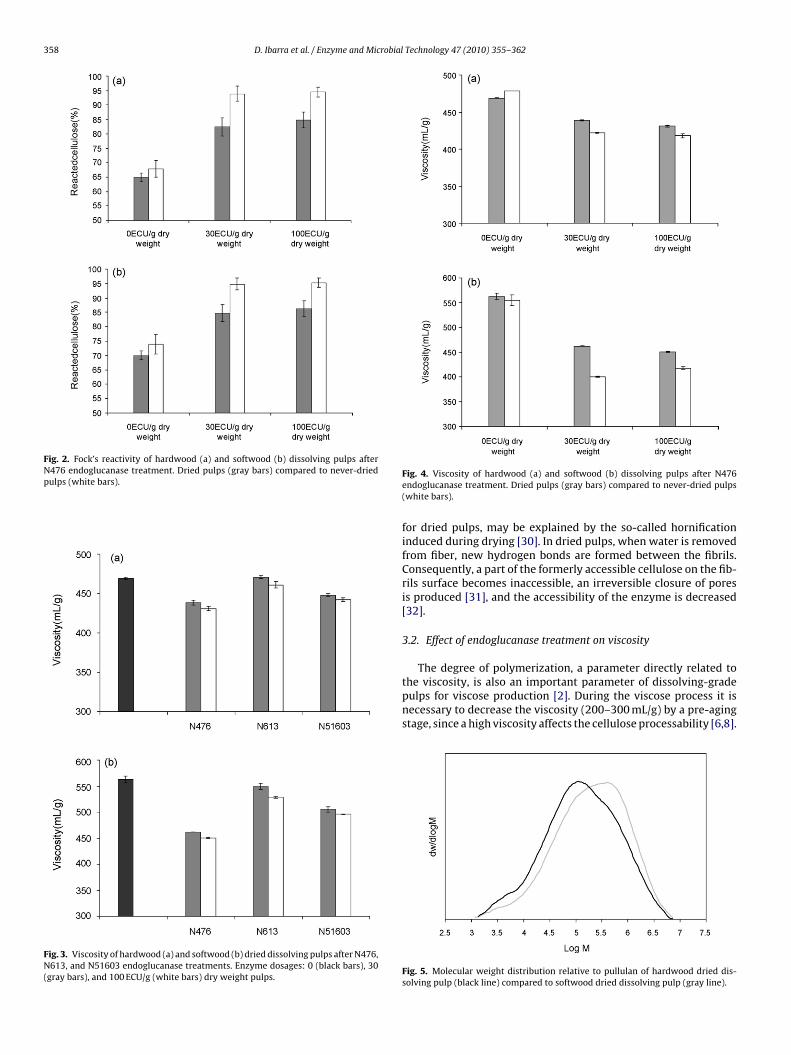
358 D. Ibarra et al. / Enzyme and Microbial Technology 47 (2010) 355–362
Fig. 2. Fock’s reactivity of hardwood (a) and softwood (b) dissolving pulps afterN476 endoglucanase treatment. Dried pulps (gray bars) compared to never-driedpulps (white bars).
Fig. 3. Viscosity of hardwood (a) and softwood (b) dried dissolving pulps after N476,N613, and N51603 endoglucanase treatments. Enzyme dosages: 0 (black bars), 30(gray bars), and 100 ECU/g (white bars) dry weight pulps.
Fig. 4. Viscosity of hardwood (a) and softwood (b) dissolving pulps after N476endoglucanase treatment. Dried pulps (gray bars) compared to never-dried pulps(white bars).
for dried pulps, may be explained by the so-called hornificationinduced during drying [30]. In dried pulps, when water is removedfrom fiber, new hydrogen bonds are formed between the fibrils.Consequently, a part of the formerly accessible cellulose on the fib-rils surface becomes inaccessible, an irreversible closure of poresis produced [31], and the accessibility of the enzyme is decreased[32].
3.2. Effect of endoglucanase treatment on viscosity
The degree of polymerization, a parameter directly related tothe viscosity, is also an important parameter of dissolving-gradepulps for viscose production [2]. During the viscose process it isnecessary to decrease the viscosity (200–300 mL/g) by a pre-agingstage, since a high viscosity affects the cellulose processability [6,8].
Fig. 5. Molecular weight distribution relative to pullulan of hardwood dried dis-solving pulp (black line) compared to softwood dried dissolving pulp (gray line).
D. Ibarra et al. / Enzyme and Microbial Technology 47 (2010) 355–362 359
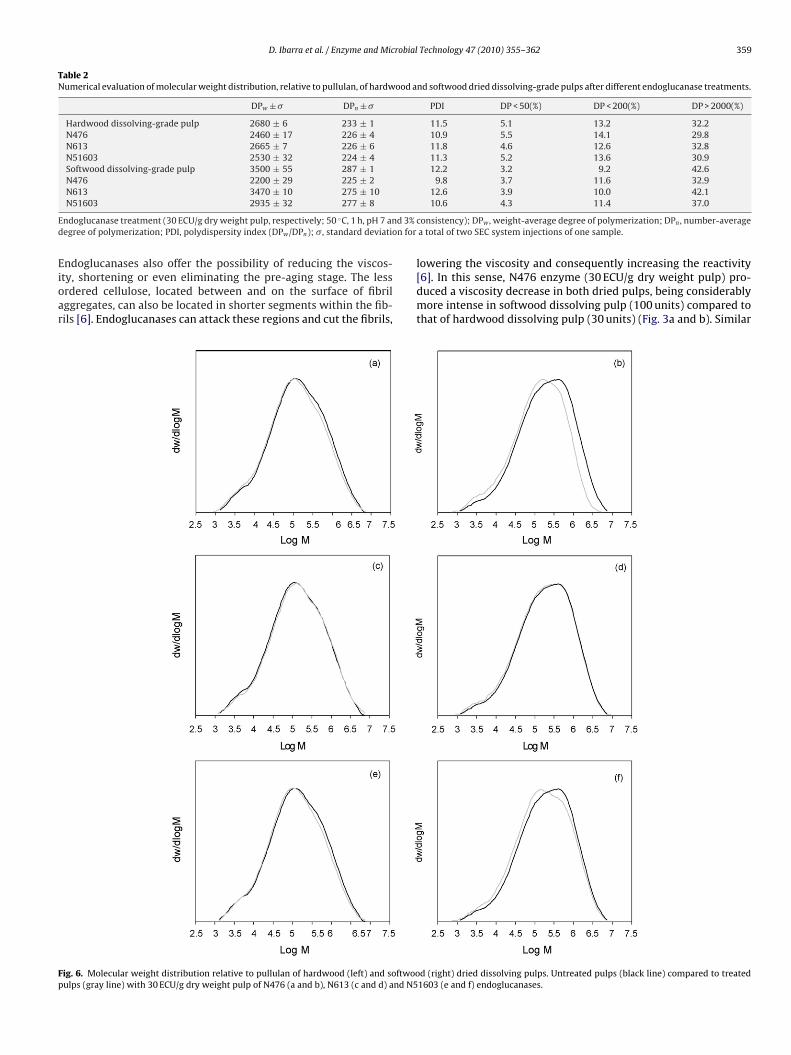
Table 2Numerical evaluation of molecular weight distribution, relative to pullulan, of hardwood and softwood dried dissolving-grade pulps after different endoglucanase treatments.
DPw ± � DPn ± � PDI DP < 50(%) DP < 200(%) DP > 2000(%)
Hardwood dissolving-grade pulp 2680 ± 6 233 ± 1 11.5 5.1 13.2 32.2N476 2460 ± 17 226 ± 4 10.9 5.5 14.1 29.8N613 2665 ± 7 226 ± 6 11.8 4.6 12.6 32.8N51603 2530 ± 32 224 ± 4 11.3 5.2 13.6 30.9Softwood dissolving-grade pulp 3500 ± 55 287 ± 1 12.2 3.2 9.2 42.6N476 2200 ± 29 225 ± 2 9.8 3.7 11.6 32.9N613 3470 ± 10 275 ± 10 12.6 3.9 10.0 42.1
E d 3%d n for
Eioar
Fp
N51603 2935 ± 32 277 ± 8
ndoglucanase treatment (30 ECU/g dry weight pulp, respectively; 50 ◦C, 1 h, pH 7 anegree of polymerization; PDI, polydispersity index (DPw/DPn); �, standard deviatio
ndoglucanases also offer the possibility of reducing the viscos-ty, shortening or even eliminating the pre-aging stage. The lessrdered cellulose, located between and on the surface of fibrilggregates, can also be located in shorter segments within the fib-ils [6]. Endoglucanases can attack these regions and cut the fibrils,
ig. 6. Molecular weight distribution relative to pullulan of hardwood (left) and softwooulps (gray line) with 30 ECU/g dry weight pulp of N476 (a and b), N613 (c and d) and N5
10.6 4.3 11.4 37.0
consistency); DPw , weight-average degree of polymerization; DPn , number-averagea total of two SEC system injections of one sample.
lowering the viscosity and consequently increasing the reactivity[6]. In this sense, N476 enzyme (30 ECU/g dry weight pulp) pro-duced a viscosity decrease in both dried pulps, being considerablymore intense in softwood dissolving pulp (100 units) compared tothat of hardwood dissolving pulp (30 units) (Fig. 3a and b). Similar
d (right) dried dissolving pulps. Untreated pulps (black line) compared to treated1603 (e and f) endoglucanases.

360 D. Ibarra et al. / Enzyme and Microbial Technology 47 (2010) 355–362
F pulps.w
blptdls
dpwddEidapn
dseiTcvSekifpaioCe
ig. 7. SEM micrographs of hardwood (left) and softwood (right) dried dissolvingeight pulp of N476 endoglucanases.
ehavior has been recently described [22]. The lower hemicellu-oses content in the softwood fibers surface [24] enable an easierenetration of the endoglucanase to cellulose as well as a bet-er adsorption onto the fibers [32], explaining the higher viscosityecrease produced by N476 enzyme. In addition, this softwood cel-
ulose, with higher less ordered cellulose content [22,23] is moreusceptible to endoglucanase attack.
In spite of the greater viscosity decrease observed in softwoodissolving pulp, Fock’s reactivity improvement was similar for bothulps (15–17 units with 30 ECU/g dry weight pulp). Similar effectas described by Kvarnlöf et al. [8], who observed a viscosityecrease without reactivity increment after treatment of softwoodissolving pulp with multicomponent cellulases. In the same way,ngström et al. [7] compared the reactivity of softwood dissolv-ng pulp after acid hydrolysis and endoglucanase treatment. It wasemonstrated that, at a given viscosity, the dissolving pulp hadlower reactivity compared to endoglucanase treated dissolving
ulp. Therefore, the activation phenomenon seems to be relatedot only to viscosity decrement.
Compared with N476 enzyme, the decrease of viscosity in bothissolving pulps with N51603 enzyme was less marked (57 unitsoftwood pulp and 21 units hardwood pulp, Fig. 3a and b). Thisffect on viscosity is consistent with previously results concern-ng the role of CBD in the activation of cellulose by endoglucanases.he absence of CBD in N51603 enzyme reduces its effectiveness inutting fibrils, i.e. on less ordered cellulose, resulting in a loweriscosity decrease and consequently a lower reactivity increase.imilar behavior was described with the intact Trichoderma reeseindoglucanases and their core proteins (without CBD) on softwoodraft pulp [33], resulting in somewhat lower pulp viscosity with thentact enzymes. Finally, a barely viscosity decrease was observedor both pulps when N613 enzyme was used (Fig. 3a and b), com-ared to values obtained with N476 and N51603 enzymes. The
bsence of CBD and its glycosidic hydrolysis mechanism, retain-ng the anomeric configuration, explain the low efficiency on lessrdered cellulose. Thus, as previously described for reactivity, theBD has no catalytic activity by itself; it supports the action ofndoglucanase enzyme by helping the catalytic domain bind to theUntreated pulps (a and b) compared to treated pulps (c and d) with 30 ECU/g dry
cellulose. A scarcely viscosity decrease was obtained by increasingthe enzymes dosages from 30 to 100 ECU/g dry weight pulp (Fig. 3aand b), as previously described [6–8].
The reduction of viscosity was more pronounced when never-dried dissolving pulps were treated with N476 enzyme (154 unitssoftwood pulp and 57 units hardwood pulp, Fig. 4a and b), in agree-ment with previous studies [34]. These results support the higherreactivity levels described above for never-dried pulps, demon-strating the role of the hornification phenomenon in the loweraccessibility of enzymes. The same effect, although in a lesserextent, was observed with N51603 and N613 enzymes (data notshown).
3.3. Size exclusion chromatography
Pulp source with a uniform and narrow MWD is of primaryimportance in order to obtain a homogeneous product [2]. Asobserved in Fig. 5, both pulps showed a single peak of cellulose,with a lower molecular mass for hardwood dissolving pulp. How-ever, in spite of their uniform MWDs, a broad cellulose distributionwas showed for both dissolving pulps, which can affect the cel-lulose processability and properties of the cellulose-end products,such as the viscose strength [2].
A numerical evaluation from their MWDs was determined(Table 2). The Mw relative to pullulan, expressed as DPw, wasconsiderably lower for hardwood dissolving pulp (DPw 2680), asdepicted in Fig. 5. This value is in agreement with its lower viscos-ity (460 mL/g), its fiber length, shorter for hardwood fibers [3], andits lower amount of long-chain cellulose molecules (DP > 2000 of32.2%). Moreover, the polydispersity index (PDI), a parameter thatdetermines the width of the MWD, confirmed the broader distribu-tion observed for both dissolving pulps, as indicated by their highPDI values (11.5 and 12.2 for hardwood and softwood pulps, respec-
tively), due to the high amount of long-chain cellulose molecules(DP > 2000 of 32.2% for hardwood pulp and of 42.6% for softwoodpulp).Changes in the MWD of both pulps were observed after N476treatment (30 ECU/g dry weight pulp). In hardwood dissolving

robial
pmodam((pdNonttfi
dMu
afeiefval(T
3
atfiuowedfafidoe
4
cpite(AdCnc
[
[
[
[
[
D. Ibarra et al. / Enzyme and Mic
ulp the cellulose peak shifted barely towards the low molecularass region (Fig. 6a), as is also evident by the small decrement
f DPw (8% drop) (Table 2), in agreement with its low viscosityecrease (Fig. 3a). In contrast, softwood dissolving pulp showedgreater shift of the cellulose peak towards the low molecularass region (Fig. 6b) with a more pronounced reduction of its DPw
37% drop) (Table 2), in agreement with its high viscosity decreaseFig. 3b). Similar behavior has been described in both dissolvingulps [5,7,8]. In addition, lower PDI values were obtained for bothissolving pulps, displaying the narrowing of their MWDs after476 treatment. It was supported by the reduction of the amountf long-chain cellulose molecules (DP > 2000) (Table 2), being sig-ificantly less marked in hardwood dissolving pulp (7% drop) thanhat of softwood dissolving pulp (23% drop). These results provehe higher accessibility of endoglucanase enzymes to the softwoodbers.
On the other hand, N613 treatment (30 ECU/g dry weight pulp)id modify neither hardwood (Fig. 6c) nor softwood pulps (Fig. 6d)WDs, in accordance with the scarcely reduction of their DPw val-
es (Table 2), and viscosity decrease (Fig. 3).Finally, N51603 treatment (30 ECU/g dry weight pulp) also
ltered the MWDs of both dissolving pulps, being less pronouncedor hardwood pulp (Fig. 6e) than softwood pulp (Fig. 6f). Nev-rtheless, the distribution profiles of both pulps were modifiedn a lesser extent in comparison to MWDs obtained with N476nzyme, as supported the lower reduction of their DPw (6% dropor hardwood and 16% drop for softwood, Table 2), and their loweriscosity decreases (Fig. 3). A lower narrowing of their MWDs waslso noticed, as showed their PDI values (Table 2), observing aower reduction of the amount of long-chain cellulose moleculesDP > 2000) (4% drop for hardwood and 13% drop for softwood,able 2).
.4. Scanning electron microscopy
In order to attain a better understanding of the endoglucanasection on enhanced accessibility, the fiber surface morphology ofhe pulps was studied by scanning electron microscopy (SEM). Theber surface of the untreated pulps appeared relatively rough andneven, showing a fibrillation from the surface in certain regionsf the fibers (Fig. 7a and b). After N476 treatment (30 ECU/g dryeight pulp for 1 h), the major part of the fibrilar material was
liminated, leaving a smoother and cleaner surface (Fig. 7c and). Consequently, this modified surface, i.e. with an increased sur-ace area, let to a greater accessibility to reagents and thereforen enhancement of cellulose reactivity. The polishing or cleaningbers surface by cellulase enzymes has been already described onifferent wood and non-wood pulps [35,36]. Even Liu et al. [28]bserved by atomic force microscopy the same effect when N476nzyme was applied on bleached reed kraft pulp.
. Conclusion
With the aim to proceed towards the application of endoglu-anase treatment in the activation of dissolving pulps for viscoserocess different monocomponent endoglucanases have been
nvestigated. The results obtained in this study reveal substan-ial differences on the cellulose activation as a function of thendoglucanases used (with different modular structures), fiber typehardwood or softwood), and fiber history (dried or never-dried).
s observed, the mode of action of endoglucanases, i.e. catalyticomain, plays an important role in the cellulose activation; whereasBD helps the catalytic domain binds to the substrate, but hasot direct activation effect on cellulose. In this sense, endoglu-anase with a CBD and an inverting hydrolysis mechanism (N476)[
[
[
Technology 47 (2010) 355–362 361
was found to be the most effective in increasing the reactiv-ity of both pulps. Simultaneously, the viscosity decreased, beinglower for softwood dissolving pulp. A narrower molecular weightdistribution with a great reduction in the amount of long-chaincellulose molecules was also observed. Endoglucanase with invert-ing hydrolysis mechanism and without a CBD (N51603) affectedthe cellulose reactivity, viscosity and molecular weight distribu-tion in a lesser extent. By contrast, endoglucanase with retaininghydrolysis mechanism and without a CBD (N613) showed an inef-ficient action on the studied properties. These properties weresubstantially enhanced in all cases when the different endoglu-canase treatments were applied on never-dried dissolving pulps.
In summary, low charges of certain monocomponent endoglu-canases, preferably with cellulose-binding domain and invertingglycosidic hydrolysis mechanism, could be efficiently used prior toviscose production to enhance the reactivity and reduce the viscos-ity of both hardwood and softwood never-dried dissolving-gradepulps. Accordingly, the demand of carbon disulfide could be low-ered and the pre-aging step shortened or even eliminated, reducingthe environmental impact and production costs.
Acknowledgements
Dr. D.I. acknowledges a grant from the Jacob WallenbergResearch Foundation (Sweden). V.K. acknowledges a financial sup-port from Stiftelsen Nils och Dorthi Troëdssons forksningsfond(Sweden). Sniace (Spain) and Domsjö (Sweden) are acknowledgedfor providing the pulp samples. Novozymes (Denmark) is acknowl-edged for supplying the enzymes.
References
[1] Treiber E, Nevell TP, Zeronian S. Formation of fibers from cellulose solutions.In: Cellulose chemistry and its applications. Chichester: H. Ellis Horwood Ltd.;1985. p. 456–57.
[2] Sixta H. Handbook of pulp. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA;2006. p. 1022–67.
[3] Fengel D, Wegener G. Wood: chemistry, ultrastructure, reactions. Berlin: DeGruyter; 1984.
[4] Krässig HA. Cellulose—structure, accessibility and reactivity. Yverdon: Yverdon,Gordon and Breach Science Publishers; 1993.
[5] Rahkamo L, Siika-aho M, Vehlainen M, Dolk M, Viikari L, Nousiainen P, et al.Modification of hardwood dissolving pulp with purified Trichoderma reeseicellulases. Cellulose 1996;3:153–63.
[6] Henriksson G, Christiernin M, Agnemo R. Monocomponent endoglucanasetreatment increases the reactivity of softwood sulphite dissolving pulp. Journalof Industrial Microbiology and Biotechnology 2005;32:211–4.
[7] Engström A-C, Ek M, Henriksson G. Improved accessibility and reactivity ofdissolving pulp for the viscose process: pretreatment with monocomponentendoglucanase. Biomacromolecules 2006;7:2027–31.
[8] Kvarnlöf N, Germgård U, Jönsson L, Söderlund CA. Enzymatic treatment toincrease the reactivity of a dissolving pulp for viscose production. Appita Jour-nal 2006;59:242–6.
[9] Köpcke V, Ibarra D, Ek M. Increasing accessibility and reactivity of paper-gradepulp by enzymatic treatment for use as dissolving pulp. Nordic Pulp and PaperResearch Journal 2008;4:363–8.
10] Ibarra D, Köpcke V, Ek M. Exploring enzymatic treatments for the production ofdissolving grade pulp from different wood and non-wood paper grade pulps.Holzforschung 2009;63:721–30.
11] Ibarra D, Köpcke V, Larsson PT, Jääskeläinen A-S, Ek M. Combination of alkalineand enzymatic treatments as a process for upgrading sisal paper-grade pulp todissolving-grade pulp. Bioresource Technology 2010;101:7416–23.
12] Tomme P, Warren RAJ, Gilkes NR. Cellulose hydrolysis by bacteria and fungi.Advances in Microbial Physiology 1995;37:1–81.
13] Rabinovich MS, Melnik MS, Bolobova AV. The structure and mechanism ofaction of cellulose enzymes. Biochemistry 2002;67:850–71.
14] Henrissat B, Driguez H, Viet C, Schüelin M. Synergism of cellulases fromTrichoderma reesei in the degradation of cellulose. Nature Biotechnology1985;3:722–6.
15] Koshland DE. Stereochemistry and mechanism of enzymatic reactions. Biolog-ical Reviews 1953;28:416–36.
16] Davies G, Henrissat B. Structures and mechanism of glycosyl hydrolases. Struc-ture 1995;3:853–9.
17] Mansfield SD, Mooney C, Saddler JN. Substrate and enzyme characteristics thatlimit cellulose hydrolysis. Biotechnology Progress 1999;15:804–16.

3 robial
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
62 D. Ibarra et al. / Enzyme and Mic
18] Linder M, Teeri TT. The roles and function of cellulose-binding domains. Journalof Biotechnology 1997;57:15–28.
19] Fock W. A modified method for determining the reactivity of viscose-gradedissolving pulp. Papier 1959;13:92–5.
20] Sjöholm E, Gustafsson K, Berthold F, Colmsjö A. Influence of the carbohydratecomposition on the molecular weight distribution of kraft pulps. CarbohydratePolymers 2000;41:1–7.
21] Berggren R, Molin U, Berthold F, Lennholm H, Lindström M. Alkaline degra-dation of birch and spruce: influence of degradation conditions on molecularmass distributions and fibre strength. Carbohydrate Polymers 2003;51:255–64.
22] Cao Y, Tan H. Study on crystal structures of enzyme-hydrolyzed cellulosic mate-rials by X-ray diffraction. Enzyme and Microbial Technology 2005;36:314–7.
23] Popescu C-M, Singurel G, Popescu M-C, Vasile C, Argyropoulos DS, WillförS. Vibrational spectroscopy and X-ray diffraction methods to establishthe differences between hardwood and softwood. Carbohydrate Polymers2009;77:851–7.
24] Suurnäkki A, Heijnesson A, Buchert J, Tenkanen M, Viikari L, Westermark U.Chemical characterization of the surface layers of unbleached pine and birchkraft pulp fibres. Journal Pulp and Paper Science 1996;22:43–7.
25] Ye D, Farriol D. Improving accessibility and reactivity of celluloses of annual
plants for the synthesis of methylcellulose. Cellulose 2005;12:507–15.26] Ståhlberg J, Johansson G, Pettersson G. A new model for enzymatic hydrol-ysis of cellulose based on the two-domain structure of cellobiohydrolase I.Biotechnology 1991;9:286–90.
27] Gilkes NR, Henrissat B, Kilbum DG, Miller Jr KC, Warren RA J. Domains inmicrobial beta-1,4-glycanases: sequence conservation, function, and enzyme
[
[
Technology 47 (2010) 355–362
families. Microbiology and Molecular Biology Reviews 1991;55(2):303–15.
28] Liu H, Fu S, Zhu JY, Li H, Zhan H. Visualization of enzymatic hydrolysisof cellulose using AFM phase imaging. Enzyme and Microbial Technology2009;45:274–9.
29] Schülein M. Enzymatic properties of cellulases from Humicola insolens. Journalof Biotechnology 1997;57:71–81.
30] Jayme G. Mikro-Quellungsmessungen an Zellstoffen. Wochenblatt Paper Fabric1994;6:187–94.
31] Stone JE, Scallan AM. The effect of component removal upon the porous struc-ture of a cell wall of wood. Part III. A comparison between the sulphite and kraftprocesses. Pulp and Paper Canada Magazine 1968;6:69–74.
32] Gerber PJ, Joyce TW, Heitmann JA. Adsorption of a Trichoderma reesiendoglucanase and cellobiohydrolase onto bleached Kraft fibres. Cellulose1997;4:255–68.
33] Suurnäkki A, Tenkanen M, Siika-Aho M, Niku-Paavola M-L, Viikari L, BuchertJ. Trichoderma reesi cellulases and their core domains in the hydrolysis andmodification of chemical pulp. Cellulose 2000;7:189–209.
34] García O, Torres AL, Colom JF, Pastor FIJ, Díaz P, Vidal T. Effect of cellulase-assisted refining on the properties of dried and never-dried eucalyptus pulp.Cellulose 2002;9:115–25.
35] Mansfield SD, de Jong Ed, Stephens RS, Saddler JN. Physical characteriza-tion of enzymatically modified kraft pulp fibers. Journal of Biotechnology1997;57:205–16.
36] Buschle-Diller G, Zeronian SH, Pan N, Yoo MY. Enzymatic hydrolysis of cot-ton, linen, ramie, and viscose rayon fabrics. Textile Research Journal 1994;64:270–9.





























