Embed Size (px)
Citation preview

Technische Universität München
Lehrstuhl für Ökophysiologie der Pflanzen
Department für Ökologie
Bedeutung der Calcium - Versorgung für die Holzdifferenzierung
sowie die elektrische Signalleitung in der Pappel
Silke Lautner
Vollständiger Abdruck der von der Fakultät Wissenschaftszentrum Weihenstephan für
Ernährung, Landnutzung und Umwelt der Technischen Universität München zur Erlangung
des akademischen Grades eines Doktors der Naturwissenschaften (Dr. rer. nat.) genehmigten
Dissertation.
Vorsitzender: Univ.-Prof. Dr. Jörg Fromm
Prüfer der Dissertation:
1. Univ.-Prof. Dr. Rainer Matyssek
2. Univ.-Prof. Dr. Wolfgang Oßwald
Die Dissertation wurde am 28. April 2005 bei der Technischen Universität München
eingereicht und durch die Fakultät Wissenschaftszentrum Weihenstephan für Ernährung,
Landnutzung und Umwelt am 20. Juli 2005 angenommen.

I
Vorwort
Die vorliegende Arbeit entstand zwischen August 2002 und April 2005 am Lehrstuhl
Ökophysiologie der Pflanzen und am Fachgebiet Angewandte Holzbiologie der Fakultät
Wissenschaftszentrum Weihenstephan (WZW) an der Technischen Universität München. Das
Projekt wurde über ein Promotionsstipendium der TU München sowie über die Deutsche
Forschungsgemeinschaft im Rahmen der Pappelforschergruppe finanziert.
Mein besonderer Dank gilt meinem Doktorvater, Herrn Prof. Dr. Rainer Matyssek, und Herrn
Prof. Dr. Jörg Fromm für die Themenstellung, ihre Anregungen und ihre stete Unterstützung
bei der Durchführung der Versuche. Herrn Prof. Dr. Wolfgang Oßwald möchte ich für die
Übernahme des Zweitkorrektorats danken. Des weiteren möchte ich PD Dr. Thorsten Grams
für seine Unterstützung und Diskussionsbereitschaft danken, die mich über weite Teile der
Arbeit begleitet haben, und allen Mitarbeitern des Lehrstuhls für Ökophysiologie der Pflanzen
und der Fachgebiete Pathologie der Waldbäume und Angewandte Holzbiologie, insbesondere
Dr. F. Fleischmann, Dr. M. Arend, Herrn T. Feuerbach, Frau A. Vieler, Frau M. Rinas und
Herrn R. Rosin. Für die Hilfestellung bei der FTIR-Spektroskopie möchte ich mich beim
Chemielabor des Lehrstuhls für Holzkunde und Holztechnologie der TUM unter der Leitung
von Frau Dr. E. Windeisen herzlich bedanken.
Dank geht auch an meine Familie und meine Freunde, die mich stets unterstützten.

II
Inhaltsverzeichnis
Vorwort................................................................................................................................. I
Inhaltsverzeichnis................................................................................................................. II
Abkürzungsverzeichnis........................................................................................................ V
1 Einleitung........................................................................................................ 1
1.1 Die Bedeutung von Calcium für die Pflanze.................................................... 2
1.2 Charakteristika elektrischer Signale................................................................. 4
1.2.1 Aktionspotentiale.............................................................................................. 5
1.2.2 Variationspotentiale........................................................................................... 7
1.3 Grundlagen der Holzbildung............................................................................. 8
1.4 Zielsetzung.................................................................................................... 10
2 Material und Methoden................................................................................. 11
2.1 Pflanzenanzucht................................................................................................ 11
2.1.1 Anzucht von Populus tremula L. x Populus tremuloides Michx. ...………...11
2.1.1.1 Agarkultur……………………………………………………………………. 11
2.1.1.2 Hydrokultur...................................................................................................... 13
2.1.1.3 Anzucht der Pappeln unter unterschiedlicher Nährstoffversorgung………….13
2.1.2 Anzucht von Populus trichocarpa.................................................................... 16
2.2 Zuwachsmessungen.......................................................................................... 17
2.3 Anatomische Untersuchungen.......................................................................... 17
2.3.1 Anfertigung von Gewebeschnitten für lichtmikroskopische Untersuchungen..17
2.3.2 Mazeration.........................................................................................................18
2.3.3 Histometrische Auswertung..............................................................................19
2.3.4 Transmissionselektronenmikroskopie (TEM).................................................. 19
2.4 Chemische Analysen........................................................................................ 20
2.4.1 Fourier-Transform-Infrarot (FTIR) - Spektroskopie.........................................20
2.5 Elementanalysen................................................................................................22
2.5.1 Energiedispersive Röntgenanalyse (EDXA) am
Rasterelektronenmikroskop (REM)...................................................................22
2.6 Elektrophysiologische Untersuchungen............................................................23
2.6.1 Membranpotentialmessungen im Mesophyll................................................. 24

III
2.6.2 Membranpotentialmessungen im Phloem.........................................................24
2.7 Blattgaswechselmessungen...............................................................................26
2.8 Messungen der Chlorophyll-Fluoreszenz..........................................................28
3 Ergebnisse...................................................................................................... 30
3.1 Bedeutung der Calciumernährung für den Phänotyp der Pappel.................... 30
3.1.1 In Hydrokultur................................................................................................. 30
3.1.2 Im Freiland...................................................................................................... 33
3.2 Bedeutung der Calciumernährung für den Calciumhaushalt der Pappel........ 37
3.3 Bedeutung der Calciumernährung für die Anatomie der Pappel.................... 41
3.3.1 In Hydrokultur................................................................................................ 41
3.3.2 Im Freiland...................................................................................................... 45
3.4 Bedeutung der Calciumernährung für die Ultrastruktur
und die Zellwandchemie der Pappel................................................................ 47
3.4.1 Ultrastruktur des Cambiums und des Blattes................................................... 47
3.4.2 Chemie der Zellwand...................................................................................... 50
3.5 Elektrische Signalleitung der Pappel.............................................................. 55
3.5.1 Signalleitung an Populus trichocarpa............................................................. 55
3.5.1.1 Kältereizung mittels Eiswasser........................................................................ 56
3.5.1.2 Hitzereizung mittels offener Flamme............................................................... 57
3.5.1.3 Unterbrechung der elektrischen Signalleitung durch einen Kälteblock........... 60
3.5.1.4 Signalleitung an mit TEA+ behandelten Pappeln............................................ 61
3.5.2 Signalleitung an Calcium-mangelversorgten Pappeln
(P. tremula x P. tremuloides)……………………………………………… 62
3.6 Ionengehalte in geöffneten und geschlossenen Pappelstomata...................... 64
3.7 Der Einfluss der elektrischen Signalleitung auf den Blattgaswechsel der
Pappel.............................................................................................................. 67
3.7.1 Bei getopften Populus trichocarpa - Pflanzen................................................ 67
3.7.2 Bei unterschiedlicher Calciumernährung von
Populus tremula x Populus tremuloides…………………………………….. 71
3.8 Auswirkung der elektrischen Signalleitung auf die
Lichtreaktionen der Photosynthese der Pappel................................................ 74

IV
4 Diskussion...................................................................................................... 78
4.1 Bedeutung der Calciumernährung für die Holzdifferenzierung...................... 78
4.1.1 Ausblick auf die Bestandesebene.................................................................... 84
4.2 Signalleitung in der Pappel............................................................................. 85
4.2.1 Bedeutung elektrischer Signalleitung für den Baum....................................... 94
5 Schlussfolgerung............................................................................................. 96
6 Zusammenfassung.......................................................................................... 99
7 Literatur........................................................................................................ 102
8 Anhang.......................................................................................................... 116

V
Abkürzungsverzeichnis ABA Abszissinsäure
ALVPD Wasserdampfsättigungsdefizit des Blattes (Air to Leaf water Vapor
Pressure Deficit) [mbar / bar]
AP Aktionspotential
APW artificial pond water
BF projizierte Blattfläche [m²]
Ca2+ Calcium
[Ca2+]cyt freies Calcium im Cytoplasma
ce molare CO2-Konzentration vor der Küvette (≡ im Referenzgas); korrigiert
um Nullpunktverschiebung und Verzerrung des Infrarotgasanalysators (IRGA)
CO2 Kohlendioxid
∆C Differenz der CO2-Konzentration zwischen Referenzgas und Messgas;
korrigiert um Nullpunktverschiebung und Verzerrung des
Infrarotgasanalysators (IRGA)
DNP 2,4-Dinotrophenol
EDXA energie-dispersive Röntgenanalyse (Energy Dispersive X-ray Analysis)
ER endoplasmatisches Retikulum
FTIR Fourier - Transform - Infrarot - Spektroskopie
gH2O stomatäre Leitfähigkeit für Wasser
H2O Wasser
JCO2 Nettophotosyntheserate
JH2O Transpirationsrate [mmol m-2s-1]
KBr Kaliumbromid
KCl Kaliumchlorid
KOH Kalilauge
KSCN Kaliumthiocyanat
MES 2-Morpholinoethan-Sulfonsäure Monohydrat
n Stichprobenumfang
PAM Puls Amplituden-Modulation
PIN Proteinaseinhibitor-Gen
PPFD photosynthetisch aktive Photonenflussdichte
PS II Photosystem II

VI
REM Rasterelektronenmikroskop
Rubisco Ribulose-1,5-Biphosphat-Carboxylase/Oxygenase
svp Sättigungsdampfdruck (saturated vapor pressure) bei angegebener
Temperatur [mbar]
TEM Transmissionselektronenmikroskop
TL Blatttemperatur [°C]
TRIS Tris-(hydroxymethyl)-aminomethan
ue molarer Gasstrom am Küvetteneingang [µmol s-1]
VP Variationspotential
wa Molenbruch des Wasserdampfes in der Küvette
we molare Feuchtekonzentration vor der Küvette (≡ im Referenzgas)
wi Molenbruch des Wasserdampfes in den Interzellularen des Mesophylls
wo molare Feuchtekonzentration hinter der Küvette (≡ im Messgas)
Yield Electon Quantum Yield; Quantenausbeute am Photosystem II

Einleitung
1
1 Einleitung
Die mineralische Ernährung besitzt im pflanzlichen Metabolismus spezifische wie auch
essentielle Funktionen. Der heute weithin geläufige Begriff der essentiellen Nährelemente
wurde 1939 erstmals von Arnon und Stout geprägt. Nach ihrer Aussage lässt sich demnach
ein Nährelement als essentiell klassifizieren, wenn folgende Bedingungen erfüllt sind:
1. Die Pflanze kann in Abwesenheit dieses Elementes ihren Lebenszyklus nicht
abschließen.
2. Die Funktion dieses Elementes kann nicht durch ein anderes mineralisches Element
kompensiert werden.
3. Das Element muss in den Pflanzenmetabolismus direkt involviert oder für einen
definierten Stoffwechselvorgang (z.B. enzymatische Reaktion) unabdingbar sein.
Nach dieser doch sehr strikten Definition würden Mineralelemente, die toxische Effekte
kompensieren oder solche, welche in weniger spezifischen Funktionen, wie z.B.
Osmoseregulation, andere Mineralelemente ersetzen, nicht als essentiell eingestuft werden.
Sie würden lediglich als „vorteilsbringende“ Nährelemente eingestuft werden. Selbst heute ist
nicht endgültig geklärt, welche Elemente in der Pflanzenernährung ausschließlich als
essentiell zu klassifizieren sind; diese Problematik wird vor allem dann deutlich, wenn man
zwischen höheren und niederen Pflanzen unterscheidet. Essentielle Nährstoffe werden dabei,
je nach Bedarf für das pflanzliche Wachstum, generell in Makronährstoffe und
Mikronährstoffe unterteilt. Zu den Makronährstoffen höherer Pflanzen, wie den Bäumen,
gehören Stickstoff, Phosphor, Schwefel, Magnesium, Kalium und Calcium. Zu deren
Mikronährstoffen zählen Eisen, Mangan, Zink, Kupfer, Bor, Molybdän, Chlorid und Nickel;
als „vorteilsbringende“ Elemente werden bei höheren Pflanzen Natrium, Silizium und Kobalt
angesehen (Marschner, 1995). Bei dieser Art der Nährstoffklassifizierung wird jedoch nicht
deutlich, dass jedes Nährelement in der Pflanze verschiedenartige Funktionen ausüben kann,
welche wiederum häufig nur recht schwach bezüglich ihrer quantitativen oder
physiologischen Bedeutung miteinander korreliert sind. So kann ein mineralischer Nährstoff
dabei gleichzeitig als Grundbaustein einer organischen Struktur wie als Aktivator
enzymatischer Vorgänge, als Ladungsträger oder als Osmoseregulator fungieren (Marschner,
1995).

Einleitung
2
1.1 Die Bedeutung von Calcium für die Pflanze
Calcium gilt mit einem Ionenradius von 0,412 nm als ein relativ großes divalentes Kation; es
bildet zudem nur schwache Bindungen zu Wasser aus. Die Aufnahme von Calcium aus dem
Boden in den Apoplasten der Wurzel erfolgt passiv und der Transport dieses Nährelements
findet nahezu ausschließlich über den Transpirationsstrom in acropetaler Richtung statt, somit
kann dieses Ion als weitgehend Phloem-immobil angesehen werden.
Im Unterschied zu anderen Makronährelementen findet sich ein Grossteil des Calciums in der
Pflanze im apoplastischen Raum wieder. Dieses Phänomen ist hauptsächlich in der Fülle von
calciumbindenden Strukturen in der Zellwand begründet, aber auch in der beschränkten Ca-
Aufnahme des Protoplasten. Besonders in der Mittellamelle ist reichlich Calcium zu finden;
hier liegt es an den Carboxylgruppen der Pektine in gebundener, für die Pflanze nicht mehr
verfügbarer Form vor. Eine weitere Art der Calciumspeicherung in Pflanzen besteht in der
Bildung von Calciumoxalat-Kristallen. Die Anzahl selbiger steigt mit zunehmender Ca2+-
Versorgung der Pflanze an und findet in Angiospermen meist in den Vakuolen der Blattzellen
statt (Borchert, 1990). In Nadelbäumen jedoch wurden die Ca2+-Oxalat-Kristalle meist in den
Zellwänden oder den Interzellularräumen detektiert (Fink, 1991 a-c). In Pflanzenarten mit
einer geringen Calciumbindungskapazität an den Carboxylgruppen des Pektins der Zellwand
kann die Ausfällung von Ca2+-Oxalaten im Apoplasten, alternativ zur Vakuole des
Symplasten, somit als Ersatzmechanismus zur Bindung und Speicherung dieses Ions dienen
(Marschner, 1995). An der Pappel (P. tremula) wurden in der Rinde des Stammes neben
einzelnen Oxalatkristallen zudem sog. „Kristallkammerfasern“ beobachtet; letztere finden
sich in der Umgebung von Phloem-Fasern und sind in ihrer ganzen Länge mit Calcium-
Oxalat- Kristallen gefüllt (Trockenbrodt, 1995).
In voll entwickelten Geweben mit hoher Kationen-Austauschrate lassen sich Bereiche mit
hoher Ca2+-Konzentration deutlich von solchen mit niedriger Ca2+-Konzentration
unterscheiden (Abb. 1.1). Hohe Calcium-Konzentrationen finden sich im Apoplasten in den
Bereichen der Mittellamelle und an der Außenseite der Plasmamembran, im Symplasten in
der Vakuole und im endoplasmatischen Retikulum (ER), aber auch in den Chloroplasten. Im
Apoplasten ist Calcium teils fest, teils reversibel gebunden an den Außenseiten der
Membranen und in der Mittellamelle der Zellwände zu finden. Hier fallen dem Calcium
vornehmlich zwei Funktionen zu. Zum einen die Erhaltung der Membranstabilität, welche
einerseits durch eine Anlagerung des Ions an den Phosphatköpfen der membranbildenden

Einleitung
3
Phospholipiden, andererseits durch eine Brückenbildung zwischen Phospholipiden und
Proteinen an der Membranoberfläche geschieht und somit zu einer Stabilisierung der
Membranen führt. Zum anderen dient Calcium der Zellwandstabilisierung, indem es durch
Bindung an Pektinketten der Mittellamelle diese vernetzt und somit den Zellwänden Stabilität
gewährt. Unter reduzierter Ca2+-Konzentration in der Mittellamelle steigt dort die
Polygalakturonase-Aktivität, wodurch es zu der typischen Calciummangelerscheinung einer
Gewebedestabilisierung, bis hin zu einem Gewebekollaps kommen kann (Bussler, 1963).
Abb. 1.1: Schematische Darstellung der Ca2+-Verteilung (Ca2+ = ●) in benachbarten Zellen;
Schema in Anlehnung an Marschner, 1995.
Im Gegensatz zum Apoplasten ist die Konzentration von freiem Calcium im Cytoplasma
([Ca2+]cyt) äußerst gering; sie erreicht hier nur Werte zwischen 0,1 und 0,2 µM (Felle, 1988;
Evans et al., 1991; Hirschi, 2004). Die geringe Konzentration an freiem Ca2+ im Cytoplasma
wird einerseits durch eine generell geringe Ca2+-Permeabilität der Plasmamembran,
andererseits durch aktiven Export des Ions mittels Membrantransporter erreicht. Als solche
fungieren an der Plasmamembran und am ER Calcium-ATPasen (Ca2+/H+ Antiporter) (Kasai
und Muto, 1990; Jones et al., 1993), ebenso am Tonoplasten, wo sie zusätzlich durch PPiasen
energetisiert werden können. Auf diese Weise kann zwischen der Konzentration freien
Calciums im Cytosol und in der Vakuole ein bis zu 105-facher Konzentrationsunterschied
aufgebaut werden (Schumaker und Sze, 1990).
Calcium erfüllt im Cytoplasma vorrangig die Funktion eines sekundären Botenstoffes.
Veränderungen im [Ca2+]cyt -Gehalt treten während einer großen Bandbreite von biotischen
und abiotischen Einflüssen auf. Als biotische Stimuli wurden dabei vor allem die Hormone
Abszissinsäure und Gibberellinsäure untersucht, aber auch Elicitoren von Pilzen und

Einleitung
4
Nodulationsfaktoren (Marschner, 1995); abiotische Stimulatoren umfassen Licht, niedrige
und hohe Temperatur, Berührung sowie hyperosmotischen und oxidativen Stress (Sanders et
al., 1999; Knight, 2000; Anil und Rao, 2001; Knight und Knight, 2001; Rudd und Franklin-
Tong, 2001).
Die intrazellulären Funktionen des Calciums lassen sich dabei in drei Bereiche unterteilen.
Zunächst kann man recht allgemein postulieren, dass Calcium im Cytoplasma auf ionischen
Stress ausgleichend wirkt. Auch intrazellulär wirkt es membranstabilisierend und reduziert
durch diese Festigung der Membranen potentielle passive Ioneneinströme (White und
Broadley, 2003).
Des weiteren treten [Ca2+]cyt - Veränderungen häufig bei einer Enzymaktivierung in
Erscheinung. Allein in den Chloroplasten sind beispielsweise mehrere Ca-abhängige Proteine
(z.B. wasserspaltende Enzyme, Calmodulin-stimulierte NAD-Kinase, Thioredoxine,
Ferredoxine und Stromaenzyme des Calvin-Zyklus) angesiedelt (Plieth, 2005).
Schließlich kommt dem Calcium noch eine herausragende Bedeutung in der Aktivierung,
bzw. Blockierung von Ionenkanälen in Membranen zu. So konnte nachgewiesen werden, dass
bei Dunkelheit Ca2+ aus den Chloroplasten ins Cytoplasma strömt und dieser Ca2+-Einstrom
die auswärtsgerichteten K+-Kanäle der Plasmamembran aktiviert; dieser K+-Ausstrom hat
eine transiente Veränderung des Membranpotentials zur Folge (Plieth et al., 1998).
1.2 Charakteristika elektrischer Signale
Bereits seit über 100 Jahren ist das Phänomen elektrischer Signale in Pflanzen bekannt. Schon
Burdon-Sanderson (1873) und Charles Darwin (1875) konnten die Existenz der elektrischen
Signalleitung in insektivoren Pflanzen nachweisen. Dennoch fristete dieses Forschungsgebiet
über lange Jahre hinweg ein unbeachtetes Dasein. Darwin wandte sich in seiner weiteren
botanischen Forschung verstärkt der Circumnutation und der damit verbundenen chemischen
Signalleitung in Pflanzen zu (Darwin 1881). Aus bislang ungeklärten Gründen wurden diese
Arbeiten so in den Vordergrund gestellt, dass die Ergebnisse zur elektrischen Signalleitung
darüber in den Hintergrund gerieten und die Pflanzenphysiologie sich verstärkt auf die
chemische Signalleitung fokussierte. Aus diesem Trend entwickelte sich mit der Zeit die
Ansicht, dass elektrische Signale, später auch chemische Signale, in tierischen Systemen der
Reizübertragung dienen, wohingegen die Signalleitung im pflanzlichen System auf
chemischer Signaltransduktion beruhe. Erst Forschungsergebnisse von Bose (1924) und

Einleitung
5
später von Sibaoka (1966, 1969) und Pickard (1973) rückten die Existenz von elektrischen
Signalen in der Pflanzenphysiologie weltweit wieder in das Bewusstsein und in das Interesse
von Botanikern. Man begann, zwischen den verschiedenen Arten der Signaltransduktion, dem
Aktionspotential und dem Variationspotential zu unterscheiden.
1.2.1 Aktionspotentiale
Auch bei der elektrischen Signalleitung ist der [Ca2+]cyt -Wert von ausschlaggebender
Bedeutung bei der Entstehung von Aktionspotentialen entlang der Plasmamembran. Erst
durch einen vorübergehenden starken [Ca2+]cyt Anstieg von ~ 0,1µM auf ~1µM werden zuerst
Cl-- und anschließend K+- Kanäle geöffnet, welche zu der typischen De- bzw. Repolarisierung
der Plasmamembran während eines Aktionspotentials führen (Abb. 1.2). Ob der Anstieg von
[Ca2+]cyt durch Ca-Einstrom aus dem Apoplasten oder durch Ca2+-Einstrom aus
cytoplasmatischen Speicherorten genährt wird, ist noch nicht gänzlich geklärt. Entgegen der
herkömmlichen Meinung eines apoplastischen Einstroms sprechen allerdings erste
Beobachtungen, die das ER als Ca2+-Quelle für den [Ca2+]cyt -Anstieg sehen (Thiel et al.,
1997; Wacke et al., 2001; Plieth, 2005).

Einleitung
6
Abb. 1.2: Schematische Darstellung eines Aktionspotentials an der Plasmamembran von Chara, in
Anlehnung an Nultsch (2001). Das Ruhepotential beträgt hier -190 mV. Durch einen an der Plasmamembran auftreffenden Reiz (Pfeil) strömt Ca2+ in das Cytoplasma ein und bewirkt dadurch die
Öffnung auswärtsgerichteter Cl--Kanäle; dies führt zur Depolarisation der Plasmamembran bis hin zum Spitzenpotential (hier: +40 mV). Durch die Positivierung des Cytoplasmas werden
spannungsabhängige auswärtsgerichtete K+-Kanäle aktiviert, welche ihrerseits durch einen Ausstrom die Repolarisierung der Membran bis hin zu ihrem Ruhepotential bewirken.
Abszisse: Zeit in Sekunden; Ordinate: Membranpotential in mV.
Spannungsänderungen an den pflanzlichen Plasmamembranen gleichen in vielem den
Aktionspotentialen, wie sie schon seit langem in tierischen Systemen bekannt sind.
Aktionspotentiale werden durch an der Plasmamembran auftretende Reize, die ein
„Schwellenpotential“ überschreiten, ausgelöst; daraufhin erfolgt selbsttätig eine
Depolarisation bis hin zum Spitzenpotential, gefolgt von der Repolarisation zurück zum
Ruhepotential. Der Zustand, der bei der Überschreitung des Schwellenpotentials eintritt, wird
auch „Erregung“ genannt. Eine Erregung führt immer zu einem voll ausgebildeten
Aktionspotential, es gilt ein „Alles oder Nichts - Gesetz der Erregung“ (Dudel et al., 1996).
Aktionspotentiale können in Pflanzen durch Berührungs- oder Kältereize ausgelöst werden;
ihre Ausbreitungsgeschwindigkeit ist etwa um den Faktor 105 langsamer im Vergleich zu
tierischen Systemen (Burdon-Sanderson und Page, 1876; Sibaoka, 1991). In höheren Pflanzen
entwickeln Aktionspotentiale eine Ausbreitungsgeschwindigkeit zwischen 1 mm s-1 und
20 mm s-1; in sensitiven Pflanzen, wie z.B. der Mimose oder der Venusfliegenfalle, sogar bis
zu 200 mm s-1 (Sibaoka, 1969).

Einleitung
7
1.2.2 Variationspotentiale
Im Gegensatz zum Aktionspotential wird durch Verwundung (Flamme, Schnitt) von
pflanzlichem Gewebe ein sich langsam ausbreitendes Variationspotential mit irregulärer Form
ausgelöst (Pickard, 1973). Die Fortpflanzung eines Variationspotentials erfolgt nicht
selbsttätig, vielmehr handelt es sich um eine lokale elektrische Antwort auf eine chemische
Substanz, die im Xylem transportiert wird (Houwink, 1935; Sibaoka, 1966, 1969; Pickard,
1973). Diese chemischen Substanzen, als Reaktion auf die Verwundung hin gebildet,
gelangen an der Verwundungsstelle ins Xylem und werden durch hydraulische Ausbreitung
fortbewegt. Sie lösen in allen lebenden Zellen, die kontaktiert werden, elektrische
Spannungsänderungen an den Plasmamembranen aus; besonders ausgeprägt sind diese
Reaktionen in den Xylem-umgebenden Zellen. Ricca behauptete 1916 erstmals, dass das
Signal chemischer und nicht elektrischer oder hydraulischer Signatur ist, da es ihm gelang
über eine „Wasserfalle“ hinweg eine chemische Signalweiterleitung entlang des
Mimosensprosses nachzuweisen. Diese chemische Substanz wurde forthin „Ricca-Faktor“
genannt. Das Variationspotential ist somit in der Lage, auch über abgestorbenes
Sprossgewebe hinweg fortgeleitet zu werden. Malone (1994) überprüfte Riccas Theorie und
konnte ebenfalls an Mimosa bestätigen, dass eine hydraulische Ausbreitung von
Verwundungsstellen in acropetaler wie auch in basipetaler Richtung stattfindet.
Zusammenfassend lässt sich sagen, dass der Mechanismus eines Variationspotentials
folgendermaßen stattfindet: an der Verletzungsstelle gebildete Wundsubstanzen werden durch
hydraulische Ausbreitung über das Xylem im Pflanzenkörper fortgepflanzt. Ein
Variationspotential lässt sich daher durch folgende Eigenschaften charakterisieren: I.) es kann
totes Gewebe passieren; II.) sein Entstehen hängt vom vorherrschenden Wassergehalt der
Pflanze ab. Ein stark negatives Wasserpotential in den Xylemgefäßen führt zu einer schnellen
Ausbreitung; bei gesättigtem Wasserzustand ist die Gefäßspannung vernachlässigbar gering
und das Variationspotential kann nicht fortgepflanzt werden.
Im Gegensatz zum Variationspotential sollte ein Aktionspotential durch totes Gewebe in
seiner Ausbreitung geblockt werden; auch ist es nicht abhängig von hydraulischen Prozessen
im Xylem.

Einleitung
8
1.3 Grundlagen der Holzbildung
Neben Kalium gilt auch Calcium als sehr wichtiges Nährelement für die cambiale Aktivität in
Gehölzpflanzen (Eschrich und Blechschmidt-Schneider, 1992; Dünisch und Bauch, 1994a).
Beide Elemente sind für Vorgänge der Zellteilung und Zelldifferenzierung im cambialen
Meristem von besonderer Bedeutung. Calcium spielt dabei vor allem durch seine
versteifenden Eigenschaften an der dehnbaren Primärwand und der Mittellamelle im sich
differenzierenden Xylemgewebe eine wichtige Rolle (Brett und Waldron, 1996; Guglielmino
et al., 1997). Auch für die osmotische Regulation in der Vakuole ist es während der
Differenzierungsphase von herausragender Bedeutung.
Das Cambium ist ein laterales Meristem, das für die Bildung von sekundärem Xylem und
Phloem verantwortlich ist. In Bäumen wird so durch cambiale Zellteilung und
Zelldifferenzierung nach innen hin der Rohstoff Holz (sekundäres Xylem), nach außen hin
das sekundäre Phloem und die Rinde gebildet (Chaffey et al., 1997). Das vaskuläre Cambium
der Gehölze ist ein sekundäres Meristem, welches aus dem Procambium hervorgeht, das
seinerseits wiederum dem Apikalmeristem entspringt. Es besteht aus zwei verschiedenen
Zelltypen: aus den fusiformen Initialzellen, die sich anatomisch durch eine schmale und
langgestreckte äußere Form sowie durch eine starke Vakuolisierung von dem zweiten Zelltyp
unterscheiden, den Strahlinitialen (Raven, 1987). Diese sind in ihrer vertikalen Ausprägung
sehr kurz und stellen den Ausgangspunkt für die das Phloem wie auch das Xylem in radialer
Richtung durchziehende Strahlen dar. Das Strahlparenchym besteht aus lebenden Zellen, über
die eine Nährstofftranslokation zwischen Phloem und Xylem stattfinden kann (Mellerowitcz
et al., 2001).
Die cambiale Aktivität ermöglicht den Gehölzpflanzen durch ständig wiederkehrende Bildung
von Phloem und Xylem ihre mehrjährigen Lebenszyklen. Anatomisch unterteilt sich die sog.
cambiale Zone in das eigentliche Cambium, welches streng genommen aus nur einer
Zellschicht, den Initialzellen, besteht, und in dessen Derivate, den Phloem- und den
Xylemmutterzellen, welche wiederum aus den sich periklin teilenden Initialzellen
hervorgehen. Die Teilungsgeschwindigkeit der Xylemmutterzellen ist dabei bedeutend
schneller als die der Phloemmutterzellen. Aus Phloemmutterzellen gehen bei Angiospermen
wie der Pappel die Siebröhrenelemente (sieve elements/SEs) und die Geleitzellen (companion
cells/CCs) hervor, welche im ausdifferenzierten Phloemgewebe sog. SE/CC-Komplexe
bilden. Im Gegensatz zu den Xylemzellen schließen die Phloemzellen ihre Entwicklung nicht

Einleitung
9
mit der Autolyse des Protoplasten ab. Die Siebröhrenelemente behalten ein rudimentäres,
membranständiges Cytoplasma, ER, einige Mitochondrien, Plastiden und phloemspezifische
Proteine (van Bel et al., 2003); sie besitzen allerdings keinen Zellkern mehr. Die mit ihnen
durch zahlreiche Plasmodesmata verbundenen Geleitzellen hingegen bleiben im Besitz ihres
vollwertigen Cytoplasmas.
Aus den Xylemmutterzellen können sich bei Angiospermen Gefäße, Tracheiden, Parenchym-
und Faserzellen bilden. Der Holzaufbau der Pappel besteht in axialer Richtung nur aus
Gefäßen und Fasern, in radialer Richtung nur aus Parenchym. Die Ausdifferenzierung der aus
den Initialzellen hervorgehenden Xylemzellen umfasst drei Phasen: Zellstreckung,
Sekundärwandbildung mit Lignifizierung und Autolyse des Protoplasten (Abb. 1.3). Bei
einigen Baumarten kann es im Laufe des Lebenszyklus’ auch noch zu einer vierten Phase
kommen, der sog. Kernholzbildung (Plomion et al., 2001).
Abb. 1.3: Lichtmikroskopischer Querschnitt durch einen Pappelstamm im Sommerzustand. Von außen nach innen: Phloemgewebe (Ph); cambiale Zone (CZ), die sich aus fusiformen Initialen,
Strahlinitialen und deren Derivaten zusammensetzt; Expansion der Xylemzellen (Exp); Sekundärwandbildung (Sek), einhergehend mit Lignifizierung und abschließenden Zelltod. Des weiteren ist das radiale Strahlparenchym (S), entlang dessen sich häufig Gefäße (G) entwickeln,
deutlich zu erkennen.

Einleitung
10
Um mit einhergehender Umfangserweiterung des Stammes den cambialen Ring nicht
unterbrechen zu müssen sind die Initialzellen auch in der Lage, sich antiklin zu teilen und
somit durch in tangentialer Richtung neu eingefügte Initialzellen einen durchgehenden
cambialen Ring zu erhalten.
1.4 Zielsetzung
Ziel der vorliegenden Arbeit war es zum einen, die Bedeutung der Calciumversorgung für die
Entwicklung und das Wachstum der Pappel herauszuarbeiten. Ein besonderes Augenmerk
sollte in diesem Zusammenhang auf der Entwicklung des cambialen und des Xylemgewebes
liegen; besonders holzanatomische und holzchemische Veränderungen in Abhängigkeit der
Calcium-Ernährung galt es hierbei zu untersuchen. Die für diese Arbeit untersuchten
Pappelarten versprachen aufgrund ihrer Wüchsigkeit und ihrer relativ unkomplizierten
Anzucht, sei es in Hydrokultur oder im Stecklingsverfahren, das geeignete Pflanzenmaterial
zu sein.
Ein zweites Ziel dieser Arbeit lag darin, Charakteristika und Funktionen möglicher
elektrischer Signale an Bäumen zu untersuchen. Auch hier sollten die Untersuchungen in
einem Zusammenhang mit der bestehenden Calcium-Ernährung dargestellt werden. Bisher
durchgeführte Versuche auf diesem wissenschaftlichen Gebiet fanden bis auf wenige
Ausnahmen an niederen oder krautigen Pflanzen statt. Diese Arbeit sollte bisher gewonnene
Erkenntnisse der Signalleitung sowie deren Auswirkungen auf die Photosyntheserate nun
auch für Bäume, in diesem Fall die Pappel, verifizieren.

Material und Methoden
11
2 Material und Methoden
2.1 Pflanzenanzucht
2.1.1 Anzucht von Populus tremula L. x Populus tremuloides Michx.
2.1.1.1 Agarkultur
Der Hybrid-Pappel-Klon Populus tremula L. x Populus tremuloides Michx. T89, welcher
freundlicherweise von Prof. Hedrich, Universität Würzburg, zur Verfügung gestellt wurde,
wurde über Kallusbildung mit Hilfe von Hormonen aus dem Sprossgewebe steril vermehrt.
Das erste Anzuchtstadium erfolgte in Petrischalen, dauerte etwa drei bis vier Wochen und
bewirkte eine Kallusbildung mit einsetzender Sprossbildung (in Medium I). Nach Umsetzen
der Sprosse in mit Medium II beschickte Magenta-Gefäße entwickelten sich die Sprosse für
weitere drei bis vier Wochen, bevor sie in Weckgläser mit Medium III überführt wurden, in
welchem sie für die Dauer von vier bis sechs Wochen Wurzeln ausbilden konnten (Abb. 2.1).
Abb. 2.1: Sterilkultur P. tremula L. x P. tremuloides Michx.;
Bewurzelung in Weckgläsern.
Die Sprossinduktion in den Medien I und II erfolgte im Licht/Dunkel-Wechsel von 8 h : 16 h
(25 W, 230 V OSRAM; TL70, F32T8/TL741, Philips) unter Temperaturen von 22°C während
der Licht- und 16°C während der Dunkel-Phase. Für die Spross und Wurzelbildung im
Medium III wurden die Pappeln unter 16 h Licht bei 22°C (TLD 58W/840 Super 80, Philips)
und 8 h Dunkelheit bei 17°C kultiviert.

Material und Methoden
12
Nährmedien für die Agarkulturen (pro 1 l)
Agarkultur Medium I: 4,4 g MS-Medium (Murashige and Skoog, Makro- und
Mikroelemente, Duchefa)
0,2 mg BAP (6-Benzylaminopurin, Sigma)
0,1 mg IBA (Indolbuttersäure, Duchefa)
0,01 mg TDZ (Thidiazuron, Duchefa)
20 g Sacharose
pH 5,8 (1 M KOH)
7g Agar (Roth, # 4508.1), autoklavieren
Agarkultur Medium II: 4,4 g MS-Medium
0,2 mg BAP
0,1 mg IBA
20 g Sacharose
pH 5,8 (1M KOH)
7g Agar, autoklavieren
Agarkultur Medium III: 4,4 g MS-Medium
10 g Sacharose
pH 5,8 (1 M KOH)
7 g Agar, autoklavieren
Die Anzucht der Pappeln in Agarkultur fand mit freundlicher Unterstützung der AG Hedrich,
Julius-von-Sachs Institut für Biowissenschaften, Universität Würzburg, statt.

Material und Methoden
13
2.1.1.2 Hydrokultur
Um die bewurzelten Pappeln der Sterilkultur in ein Raumklima zu überführen, wurden die
Pflanzen in mit Flüssigmedium gefüllte, abgedunkelte Kolbengläschen gegeben. Der
beblätterte Spross war dabei in der ersten Woche noch von einer kleinen Klarsichtbedeckung
umgeben, welche im Lauf der zweiten Woche Schritt für Schritt angehoben und schließlich
ganz entfernt wurde.
Hydrokultur Medium: 1,5 mM KH2PO4
2,0 mM KNO3
1,0 mM CaCl2
1,0 mM MgSO4
18 µM FeNaEDTA
8,1 µM H3BO3
1,5 µM MnCl2
pH 6,0
2.1.1.3 Anzucht der Pappeln unter unterschiedlicher Nährstoffversorgung
Um den Einfluss von Calcium auf verschiedene Prozesse der Holzbildung zu untersuchen,
wurden die aus dem Kallusgewebe gewonnenen Pappeln unter verschiedener
Calciumversorgung in Hydrokulturen angezogen. Dazu wurden sie in zehn Liter
Nährstofflösung fassende Curver-Boxen gegeben und über Tetratec®AP50 Luftpumpen (Tetra
Werke, 49324 Melle, Deutschland) mit Sauerstoff versorgt (Abb. 2.2). Um eine Veralgung
der Nährlösungen zu vermeiden und gleichzeitig den Pflanzen Stabilität zu gewährleisten
wurde die Lösungsoberfläche mit einer Styroporplatte abgedeckt.

Material und Methoden
14
Abb. 2.2: Anzucht von P. tremula x P. tremuloides unter verschiedenen
Ca2+-Regimen in Hydrokultur
Als Ausgangsnährlösung für die Hydrokulturen wurde sich an der Hoagland’schen
Nährlösung orientiert, die in ihrer Rezeptur 5 mM Ca2+ vorsieht (Hoagland and Arnon, 1950).
Diese Nährlösung wurde in zwei von insgesamt drei Versuchsreihen zum Vergleich auf
Calcium-Gehalte von 0 mM und 10 mM modifiziert (Tab 2.1), im dritten Durchlauf wurden
die Calciumgehalte der Nährlösungen von 5mM Ca2+ der Optimalversorgung auf 1 mM Ca2+
reduziert und auf 0,1 mM Ca2+ minimiert (Tab. 2.2). Zur Erhöhung des Calciumgehaltes der
Ausgangslösung wurde die Ca(NO3)2 -Konzentration erhöht, wobei die Mg(NO3)2-
Konzentration entsprechend erniedrigt wurde. Zur Reduktion von Calcium wurde der
Ca(NO3)2 -Anteil der Ausgangslösung durch entsprechende Anteile von Mg(NO3)2 ersetzt
(Tab. 2.1 und 2.2); der pH-Wert der Nährlösungen wurde auf 5,8 eingestellt. Durch
elementanalytische Untersuchungen konnte gezeigt werden, dass Magnesium nicht in der
Holzbildungszone akkumuliert wird und daher davon ausgegangen werden kann, dass es bei
der Holzbildung keine bedeutende Rolle einnimmt. Ein erhöhter Gehalt dieses Ions in den
modifizierten Nährlösungen hat somit auf Interpretationen zur calciumabhängigen
Holzbildung keinen Einfluss.
Die Nährlösungen wurden wöchentlich ausgetauscht.

Material und Methoden
15
Ca-Entzug
0 mM Calcium
Ca-normal
5 mM Calcium
Ca-Überfluss
10 mM Calcium
KNO3 5 mM 5 mM 5 mM
Ca(NO3)2 - 5 mM 10 mM
Mg(NO3)2 10 mM 5 mM -
MgSO4 2 mM 2 mM 2 mM
KH2PO4 1 mM 1 mM 1 mM
KCl 50 µM 50 µM 50 µM
Fe-EDTA 40 µM 40 µM 40 µM
H3BO3 25 µM 25 µM 25 µM
MnSO4 5 µM 5 µM 5 µM
ZnSO4 2 µM 2 µM 2 µM
CuSO4 0,5 µM 0,5 µM 0,5 µM
Tab. 2.1: Zusammensetzung der in ihrem Ca2+-Gehalt modifizierten Nährlösungen
der ersten beiden Versuchsreihen der Pappelanzucht in Hydrokultur
Ca-minimiert
0,1 mM Calcium
Ca-reduziert
1 mM Calcium
Ca-normal
5 mM Calcium
KNO3 5 mM 5 mM 5 mM
Ca(NO3)2 0,1 mM 1 mM 5 mM
Mg(NO3)2 4,9 mM 4 mM -
MgSO4 2 mM 2 mM 2 mM
KH2PO4 1 mM 1 mM 1 mM
KCl 50 µM 50 µM 50 µM
Fe-EDTA 40 µM 40 µM 40 µM
H3BO3 25 µM 25 µM 25 µM
MnSO4 5 µM 5 µM 5 µM
ZnSO4 2 µM 2 µM 2 µM
CuSO4 0,5 µM 0,5 µM 0,5 µM
Tab. 2.2: Zusammensetzung der in ihrem Ca2+-Gehalt modifizierten Nährlösungen
der dritten Versuchsreihe der Pappelanzucht in Hydrokultur

Material und Methoden
16
2.1.2 Anzucht von Populus trichocarpa
Stecklinge eines intraspezifischen Hybrids von Populus trichocarpa x Populus trichocarpa
(cv Trichobel) wurden vom Bayerischen Amt für forstliche Saat- und Pflanzenanzucht,
Teisendorf, bezogen. Dabei handelte es sich um einjährige, unverzweigte Apikaltriebe von
80 cm Länge mit einem Mitteldurchmesser von etwa 5 mm. Die Stecklinge wurden vor
Knospenaustrieb in ein möglichst nährstoffarmes Bodensubstrat gegeben, welches aus einem
Gemisch aus Quarzsand (2,0–2,5 mm Körnung) und einem handelsüblichen Weißmoostorf
(Europlant®, Fa. Euroflor GmbH, Raubersried, Deutschland) bestand. Dieses Gemisch wurde
vor dem Überführen der Stecklinge zweimal mit destilliertem Wasser durchgespült und in
Pflanztöpfe mit einem Fassungsvolumen von 25 Litern gegeben. In jeden Pflanztopf wurden
sieben Apikaltriebe je ca. 20 cm tief gesteckt (Abb. 2.3); die Töpfe wurden im Freiland
aufgestellt und stets von Verunreinigungen (Laub etc.) freigehalten.
Abb. 2.3: Im Freiland angezogene Populus trichocarpa - Pflanzen
während der Wachstumsperiode
Um den Einfluss von Ca2+ auf die Holzbildung der im Freiland gewachsenen Pappeln zu
bestimmen, wurden pro Nährstoffvariante zwei Töpfe regelmäßig mit den entsprechenden
Lösungen gegossen, so dass die gebildeten Wurzeln stets mit ausreichend Nährlösung
versorgt waren. Die Nährlösungen entsprachen den in Tab. 2.2 beschriebenen Lösungen einer
Optimalversorgung, einer Ca2+ reduzierten- und einer Ca2+ minimierten Variante. Für
anatomische Untersuchungen wurden die Pappeln in einem vierwöchigen Turnus beerntet.

Material und Methoden
17
2.2 Zuwachsmessungen
Um den Einfluss von Ca2+ auf die phänotypische Entwicklung der Pappeln über eine
Wachstumsperiode hinweg zu beobachten, wurden zu Beginn der Versuchsreihe fünf getopfte
Populus trichocarpa Stecklinge pro Variante ausgewählt; den Pappeln war allen eine Höhe
von 60 cm und ein Stammfußdurchmesser von ca. 5,8 mm gemeinsam. Sie wurden im
einwöchigen Turnus hinsichtlich ihres Blattflächenzuwachses, ihres Trieblängenzuwachses
und ihres Durchmesserzuwachses in 30 cm Höhe vermessen.
Da die jungen Triebe von P. trichocarpa Korkleisten aufweisen, wurde der Durchmesser
jeweils zweimal um 90° versetzt aufgenommen und der daraus resultierende Mittelwert als
Mitteldurchmesser gewählt. Bei den Messungen des Trieblängenzuwachses wurden der
zugewachsenen Länge des Terminaltriebes die der Seitentriebe zuaddiert. Um den
Blattflächenzuwachs innerhalb der verschiedenen Varianten zu bestimmen, wurde die
Blattanzahl der einzelnen Bäume wöchentlich aufgenommen. Die Blätter wurden am Ende
der Wachstumsperiode geerntet und den einzelnen Bäumen zugewiesen; aus dem Gewicht
und der Blattfläche einer Mischprobe aus zehn Blättern jeder Variante wurde der
Blattflächenzuwachs der einzelnen Messtermine rückwirkend berechnet.
2.3 Anatomische Untersuchungen
2.3.1 Anfertigung von Gewebeschnitten für lichtmikroskopische Untersuchungen
Für die Anfertigung von Gewebeschnitten für die Lichtmikroskopie wurden 1 cm lange
Proben in einer Fixierlösung mit 1% Formaldehyd, 1 mM EGTA, 50 mM Cacodylat-Puffer
und 5% Glutaraldehyd im Exicator infiltriert und für 2 Stunden bei Raumtemperatur fixiert.
Anschließend wurden die Proben mehrmals mit Cacodylat-Puffer gespült und in einer
ansteigenden Ethanolreihe (25% / 50% / 75% / 100%) entwässert. Die Proben wurden sodann
mit LR-White Acrylharz infiltriert und in Gelatinekapseln eingebettet. Die Polymerisation in
den Kapseln erfolgte für 24 Stunden bei 60°C in einem Trockenschrank.
Aus den eingebetteten Gewebeproben wurden mittels eines Mikrotoms (MT-X, RMC Inc.)
Semidünnschnitte von 1 µm Schnittdicke angefertigt, auf Objektträger übertragen und mit
Toluidin O angefärbt. Die solchermaßen präparierten Schnitte konnten nun im

Material und Methoden
18
Lichtmikroskop (Axiophot, Zeiss) betrachtet und mittels einer digitalen Camera (Axiocam,
Zeiss) fotographisch festgehalten werden.
2.3.2 Mazeration
Um die Länge der Libriformfasern bestimmen zu können, wurde der Zellverband des
Holzgewebes zunächst aufgelöst. Dazu wurde von ca. 1,5 cm langen Sprosstücken zunächst
die Rinde entfernt, sodann wurden mittels einer Rasierklinge Späne aus dem Holzkörper
abgeschabt, welche in eine Mazerationslösung gegeben wurden. Bei dieser Lösung handelt es
sich um das so genannte „Jeffrey’sche Gemisch“, welches aus 10%iger Salpetersäure (HNO3)
und Chromsäureanhydrid (CrO3) in einer 10%igen Lösung im Verhältnis 1:1
zusammengesetzt ist. Die Xylemspäne wurden in der Lösung in einem Wärmeschrank
(Memmert U30) für ca. 50 Minuten bei 60°C mazeriert. Anschließend wurden die weitgehend
aus dem Faserverband gelösten Libriformfasern durch einen Filter dekantiert und mit H2O
(bidest.) säurefrei gespült. Die Fasern konnten nun in einem Tropfen Glycerin eingeschlossen
auf dem Objektträger fixiert, im Lichtmikroskop (Axiophot, Zeiss) betrachtet und mit einer
digitalen Camera (Axiocam, Zeiss) fotografiert werden (Abb. 2.4).
Abb. 2.4: Für die Faserlängenmessung mazerierte Holzfasern

Material und Methoden
19
2.3.3 Histometrische Auswertung
Die histometrische Auswertung erfolgte an den digitalen Aufnahmen der Gewebeschnitte,
sowie an den Aufnahmen der mazerierten Fasern unter Verwendung der digitalen
Messfunktionen der Kamerasoftware Axio Vision 3.1 von Zeiss. Dabei wurden neben der
Länge der mazerierten Fasern auch der Holzzuwachs und die Gefäßgrößen anhand der
Stammquerschnitte gemessen.
2.3.4 Transmissionselektronenmikroskopie (TEM)
Sprossproben von 1 cm Länge wurden axial geviertelt und für zwei Stunden bei
Raumtemperatur in Fixierlösung gegeben (Lösung vgl. Kapitel 2.3.1). Nachdem sie mehrmals
in Cacodylat-Puffer gespült wurden, wurden sie über Nacht bei 4°C in einer Lösung mit 2%
Osmiumtetroxid in Puffer nachfixiert. Die Proben wurden sodann zweimal mit H2O (bidest.)
gespült und in 3 % Uranylacetat in 20% Ethanol für eine Stunde nachkontrastiert. Danach
erfolgte eine Entwässerung durch eine ansteigende Ethanolreihe (50% / 75% / 2x 100%) und
eine 1 : 1 Tränkung der Proben mit 100% Ethanol und einem Epoxydharz, dem Spurr’s
Gemisch (Spurr, 1969). Im Anschluss daran fand die Einbettung in reines Epoxydharz mit
einer zwölfstündigen Polymerisation bei 70°C statt. Für TEM-Untersuchungen wurden aus
den polymerisierten Proben an einem Ultramikrotom (Ultratome Nova, LKB)
Ultradünnschnitte von 80 - 100 nm angefertigt; diese wurden auf mit Formvar beschichtete
Kupfergrids (100 mesh) übertragen und mit alkalischer Bleicitratlösung (Reynolds, 1963) für
zehn Minuten unter CO2-Ausschluss bei Raumtemperatur nachkontrastiert. Die Beobachtung
der Ultradünnschnitte erfolgte am Transmissionselektronenmikroskop (EM 10 C, Zeiss) bei
einer Beschleunigungsspannung von 80 kV und 2.500 bis 100.000facher Vergrößerung. Für
die photographischen Aufnahmen wurde ein AGFA SCIENTIA EM Planfilm verwendet.

Material und Methoden
20
2.4 Chemische Analysen
2.4.1 Fourier-Transform-Infrarot (FTIR) - Spektroskopie
IR-spektroskopische Charakterisierungsmethoden haben seit der Entwicklung von
Routinegeräten in den 50er Jahren und besonders mit der Weiterentwicklung zur sog. Fourier-
Transformation eine große Bedeutung erlangt. Zu ihren Vorteilen gehören eine geringe
benötigte Substanzmenge und ein hoher Erkennungswert für bereits spektroskopierte und
entsprechend katalogisierte Substanzen. Aus den bisherigen FTIR-spektroskopischen
Untersuchungen haben sich bislang folgende Absorptionsbanden als für Holz charakteristisch
gezeigt (Faix 1992, Fengel und Ludwig 1991):
Wellenummer [cm-1] Funktionelle Gruppe Kommentar
3490 / 3330 O-H Bindungen
C-H Bindungen in CH3, CH2
2050 S=C=N Interner Standard
1740 C=O Bindungen (nicht-konjugiert) z.B. in Acetyl- Gruppen und Carboxylestern
1640
1652
C=O Bindungen (konjugiert)
Coniferyl-Aldehyd
Bereich oft unspezifisch, da hier adsorbiertes H2O bei 1635 überlagert, allerdings ist diese Bande bei Rinde signifikant
1600 / 1505 aromatische Schwingungen von Lignin
1515 phenolische Gruppen
1460 C-H Deformation z.B. in CH3, CH2
1425 C-H Deformation (und aromatische Schwingungen)
1380 C-H Bindung z.B. in CH3 der Acetyl-Gruppen, nicht in OCH3

Material und Methoden
21
Wellenummer [cm-1] Funktionelle Gruppe Kommentar
1330 C-O aus Aryl-Alkyl-Ether, z.B. Ar-O-CH3,
hier: S-Lignin
1240 C-O Bindung der Acetyl- Gruppen
in Verbindung mit 1740
1160 C-O-C antisym. Bindung
1110 asym. Ringbindung
1050 C-O Bindung, C-H Deformation
900 - 1300 Funktionelle Gruppen der Kohlenhydrate
898 C-H Deformation (Pyranring)
Tab. 2.3: Auswahl an spezifischen FTIR-Absorptionsbanden und den entsprechenden funktionellen Gruppen.
Die FTIR-Spektren wurden mit einem Gerät der Firma Biorad (FTS-40) erstellt. Die
Gewebeproben (Holz- und Rindenproben) wurden dafür nach der Kaliumbromid-Preßling-
Methode präpariert. Dazu wurde 1 mg einer zu untersuchenden Probe viermal für 1,5 Minuten
mit 300 mg Kaliumbromid (KBr) in einer Perkin Elmer Schwingmühle homogenisiert. Um
die Proben auf einen internen Standard zu eichen, wurde ihnen Kaliumthiocyanat (KSCN)
10%ig beigefügt; um eine Wasseranlagerung an das zugegebene KSCN zu vermeiden wurden
die Proben vorsichtig unter Vakuum getrocknet. Das Gemisch wurde anschließend zu einer
durchstrahlbaren Tablette gepresst. Pro Variante wurden auf diese Weise fünf Tabletten
zwischen 4000 und 450 cm-1 mit einer Auflösung von 4 cm-1 und 16 Scans IR-
spektroskopisch untersucht.

Material und Methoden
22
2.5 Elementanalysen
2.5.1 Energiedispersive Röntgenanalyse (EDXA) am Rasterelektronenmikroskop
(REM)
Das Prinzip der Röntgenmikroanalyse beruht auf der Interaktion zwischen den energiereichen
Elektronen des Strahls eines Elektronenmikroskops und den Atomen eines biologischen
Präparates. Energetisch niedrigere Elektronen der beschossenen Atome werden aus den
inneren Schalen geschleudert und durch Elektronen mit höherem Energieniveau aus den
äußeren Schalen ersetzt. Die dabei freiwerdende Energiedifferenz wird unter anderem als
Röntgenstrahlung emittiert. Da die Atomradien und die Anzahl der Elektronenschalen von der
unterschiedlichen Kernladung der jeweiligen Elemente abhängig sind, ist die freiwerdende
Röntgenstrahlung spezifisch für das betroffene Element. Die Röntgenstrahlung gibt aufgrund
kernnaher Emissionen jedoch keine Auskunft über den chemischen Status des betroffenen
Atoms.
Um EDX-Analysen an Pappel-Geweben durchzuführen, wurden frische Proben mit einer
Rasierklinge zugeschnitten, mittels in Flüssig-Stickstoff gekühlten Isopentans schockgefroren
und zur Trocknung für einen Tag an eine Gefriertrocknungsanlage (GT2-E, LYOVAC)
angeschlossen. Um die Kontrastierung der Proben im REM zu verbessern und gleichzeitig
eine elektrische Aufladung zu vermeiden wurden die gefriergetrockneten Proben in einem
Sputtergerät (K 575, EMITECH) für eine Minute und zehn Sekunden bei 90 mA mit Chrom
beschichtet. Die semiquantitative Ionenbestimmung der Gewebeproben erfolgte am
Rasterelektronenmikroskop (REM AMR 1200 B, Leitz), an welches eine energiedispersive
Röntgenanalysevorichtung (EDXA, KEVEX 4000) angeschlossen ist. Diese EDX-Anlage
ermöglicht eine Registrierung aller Elemente ab der Ordnungszahl 9 (Fluor).
Die Elementanalyse wurde unter einer Beschleunigungsspannung von 15 kV und bei
1000facher Vergrößerung der zu messenden Gewebeprobe durchgeführt. Nach einer
Analysendauer von 200 Sekunden wurde das resultierende Röntgenspektrum von einem
Analogschreiber aufgezeichnet; durch die Bestimmung des Verhältnisses der
elementspezifischen Röntgenstrahlung zum unspezifischen Hintergrund (peak:background
ratio) konnte eine relative Maßzahl für die Massenkonzentration des jeweiligen Ions ermittelt
werden (Abb 2.5).
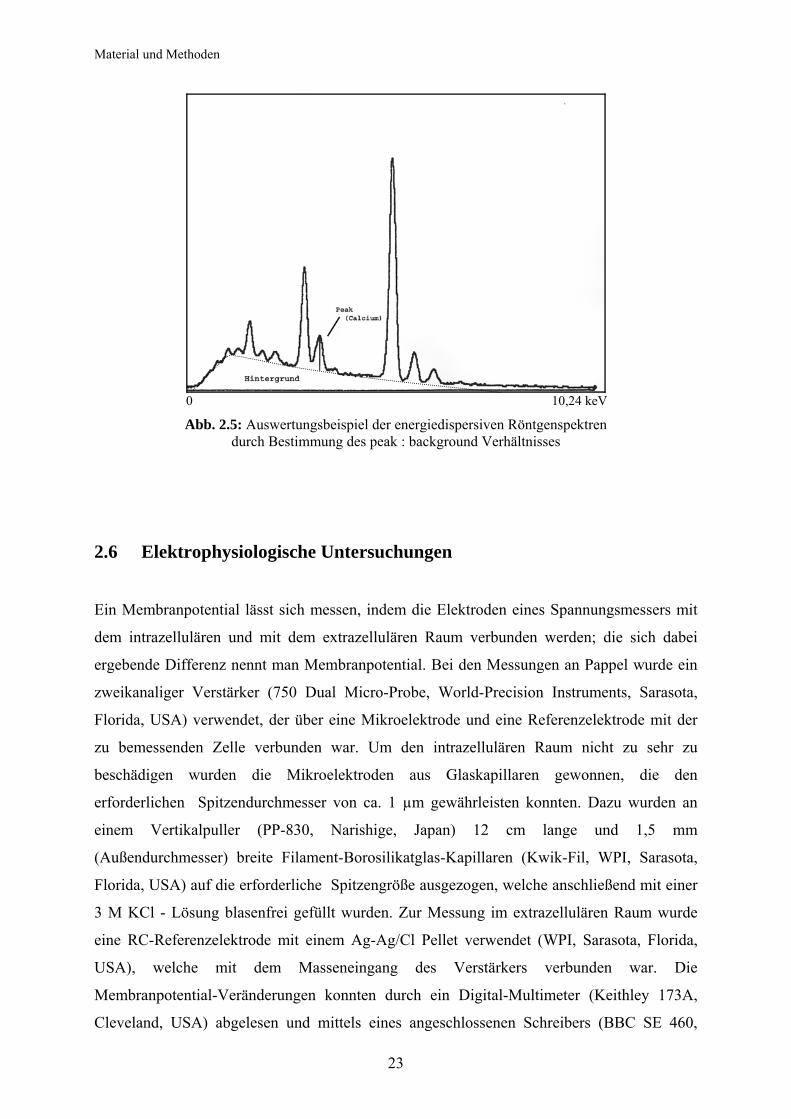
Material und Methoden
23
0 10,24 keV
Abb. 2.5: Auswertungsbeispiel der energiedispersiven Röntgenspektren durch Bestimmung des peak : background Verhältnisses
2.6 Elektrophysiologische Untersuchungen
Ein Membranpotential lässt sich messen, indem die Elektroden eines Spannungsmessers mit
dem intrazellulären und mit dem extrazellulären Raum verbunden werden; die sich dabei
ergebende Differenz nennt man Membranpotential. Bei den Messungen an Pappel wurde ein
zweikanaliger Verstärker (750 Dual Micro-Probe, World-Precision Instruments, Sarasota,
Florida, USA) verwendet, der über eine Mikroelektrode und eine Referenzelektrode mit der
zu bemessenden Zelle verbunden war. Um den intrazellulären Raum nicht zu sehr zu
beschädigen wurden die Mikroelektroden aus Glaskapillaren gewonnen, die den
erforderlichen Spitzendurchmesser von ca. 1 µm gewährleisten konnten. Dazu wurden an
einem Vertikalpuller (PP-830, Narishige, Japan) 12 cm lange und 1,5 mm
(Außendurchmesser) breite Filament-Borosilikatglas-Kapillaren (Kwik-Fil, WPI, Sarasota,
Florida, USA) auf die erforderliche Spitzengröße ausgezogen, welche anschließend mit einer
3 M KCl - Lösung blasenfrei gefüllt wurden. Zur Messung im extrazellulären Raum wurde
eine RC-Referenzelektrode mit einem Ag-Ag/Cl Pellet verwendet (WPI, Sarasota, Florida,
USA), welche mit dem Masseneingang des Verstärkers verbunden war. Die
Membranpotential-Veränderungen konnten durch ein Digital-Multimeter (Keithley 173A,
Cleveland, USA) abgelesen und mittels eines angeschlossenen Schreibers (BBC SE 460,

Material und Methoden
24
Schütt, Göttingen) aufgezeichnet werden. Da die Messungen nicht an isolierten Zellen
durchgeführt wurden, wurden die abgetrennten Triebspitzen mit ihrer Schnittstelle in
künstliches Teichwasser (artificial pond water: APW; nach Weisenseel et al., 1992) gestellt
und fixiert; die Referenzelektrode befand sich zur Bemessung des extrazellulären Raums in
der APW - Lösung.
Artificial pond water (APW): 1,0 mM NaCl
0,1 mM KCl
0,1 mM CaCl2
1,0 mM MES
pH mit TRIS auf 6,0 eingestellt
Während der Messungen befanden sich alle Geräte, mit Ausnahme des Verstärkers, des
Multimeters und des Schreibers unter einem Faraday’schen Käfig und waren somit von
äußeren elektrischen Einflüssen abgeschirmt.
2.6.1 Membranpotentialmessungen im Mesophyll
Vor Beginn der Membranpotentialmessungen im Mesophyll wurde die Mikroelektrode
zunächst zur Eichung im APW des extrazellulären Raums der Referenzelektrode auf den
Nullwert eingestellt. Danach wurde sie mit Hilfe eines Mikromanipulators durch die
Membran in das Cytoplasma der zu bemessenden Zelle eingeführt. Auf dem Digital-
Multimeter konnte nun das aktuelle Membranpotential abgelesen werden.
2.6.2 Membranpotentialmessungen im Phloem
Während sich Membranpotentialmessungen in peripheren Zellen von Pflanzenorganen durch
Einführung der Mikroelektrode mittels eines Mikromanipulators relativ einfach durchführen
lassen, sind Messungen an tiefer im Gewebe liegenden Zellen schwieriger durchzuführen.
Dies gilt insbesondere für Phloemzellen. Die hier durchgeführten
Membranpotentialmessungen an Phloemzellen basieren auf Messungen an abgetrennten
Rüsseln Phloemsaft-saugender Blattläuse. Diese Methode bot für die Messungen folgende
Vorteile:

Material und Methoden
25
- Das oft mühselige Suchen nach Siebröhren entfällt, da die meisten Blattläuse
ausschließlich im Phloem saugen (Evert et al., 1973)
- Das Gewebe wird nicht durch das Einführen einer Mikroelektrode verletzt und muß
nicht aufgetrennt werden.
- Das Zurückdrängen des Elektrolyten aufgrund eines hohen Zellturgors („backfiring“)
findet nicht statt, da der austretende Siebröhrensaft turgorlos ist.
Sobald man im Mikroskop eine Laus entdeckt, die dem Objektiv entweder seitlich oder
frontal gegenübersteht (Abb. 2.6 A) und zugleich Honigtau absondert, kann sie mit Hilfe
eines Laserimpulses (Neodym:Glas-Laser, Beck, Neu-Isenburg) von ihrem Rüssel abgetrennt
werden. Die Messelektrodenspitze wird nun mittels des Mikromanipulators mit dem aus dem
Rüssel austretenden Phloemexsudat kontaktiert (Abb. 2.6 B).
Abb. 2.6: Schematische Darstellung der Membranpotentialmessungen im Phloem des Blattleitbündels
unter Anwendung der Aphidentechnik. A: Skizzierte Darstellung einer im Phloem saugenden Blattlaus; zur besseren Darstellung wurde die Skizze um 180° gedreht. B: Photographische
Darstellung eines vom Laser abgetrennten Blattlausrüssels; der Rüssel befindet sich mit dem einen Ende noch im Phloem, die Mikroelektrode kontaktiert das andere Rüsselende mit dem austretenden
Phloemsaft.

Material und Methoden
26
2.7 Blattgaswechselmessungen
Die Messungen des Blattgaswechsels erfolgten an triebspitzennahen Pappelblättern, die ihr
Blattflächenwachstum im Wesentlichen abgeschlossen hatten.
Als Messgerät zur Erfassung der Photosyntheseleistung und der Transpiration wurde das
Kompakt-CO2/H2O-Porometer CQP 130 der Firma Walz in Effeltrich, Deutschland,
eingesetzt, welches sich aus folgenden Teilen zusammensetzt:
dem Porometermesskopf (inkl. Küvette) (Abb. 2.7),
dem externen Luftfeuchte- und Temperatursensor,
dem Infrarot-Gasanalysator (Binos 100) und
der Zentraleinheit CQP 130 mit mikroprozessorgesteuerter Datenerfassungseinheit.
Die Küvette besteht aus Plexiglas und ermöglicht so eine Beleuchtung des eingeschlossenen
Blattteils. Am Porometermesskopf misst ein Quantum Sensor (LI-190S, LiCor) die
einfallende photosynthetisch aktive Strahlung. Die Küvettentemperatur wird über einen
Pt100-Meßwiderstand und die Blatttemperatur über ein Thermoelement erfasst; mittels einer
kapazitiven Messzelle wird ebenfalls innerhalb der Küvette die relative Luftfeuchte erfasst.
Durch ein Peltieraggregat und einen Ventilator ist eine begrenzte Klimatisierung der
Messküvette möglich, so dass der eingeschlossene Blattteil bei hoher Lichtintensität keiner
Überhitzung ausgesetzt ist.
Abb 2.7: Pappelblatt, in die Küvette des Porometermeßkopfs
von CQP130 eingesetzt

Material und Methoden
27
Das Messprinzip des CQP 130 gibt vor, dass die dem System zugeführte Luft zunächst in ein
Puffergefäß gelangt, um kurzfristige Luftfeuchtigkeits- und CO2-Schwankungen
auszugleichen. Nach dem Puffergefäß teilt sich der Gasstrom in eine Referenzgasleitung und
in eine Messgasleitung. Das Referenzgas wird über ein Ausgleichsgefäß direkt, das Messgas
über die Küvette in den Gasanalysator geleitet, welcher dann im so genannten
Differenzmessverfahren die Veränderung der Gaszusammensetzung des Messgases relativ
zum Referenzgas bestimmt (Gaswegeplan: siehe Anhang). Zuerst wird die Veränderung des
Wasserdampfgehaltes in einem separaten Kanal gemessen. Danach wird in einem zweiten
Kanal der CO2-Gehalt analysiert. Die bis zu einem Volumenanteil von 1/1000000 (ppm)
genaue Gasanalyse basiert auf der Infrarotabsorption des Wasserdampfes, bzw. der CO2-
Moleküle.
Während der Messungen betrug die CO2-Konzentration der zugeführten Luft etwa 360 µl l-1
(Außenluft). Da die Messungen in einer Klimakammer stattfanden (York GmbH, Mannheim,
Deutschland) konnte von konstanten ca. 65% rel. Luftfeuchte, ca. 27°C und einer PPFD von
ca. 100 µmol m-2 s-1 ausgegangen werden. Mit Hilfe einer Kältefalle konnte die relative
Luftfeuchtigkeit in der Küvette der Umgebung angepasst werden. Die zeitliche Verzögerung
zwischen Veränderungen innerhalb der Küvette und deren Registrierung am Binos ist
abhängig von der Durchflussgeschwindigkeit des Messgases und wurde in den Berechnungen
berücksichtigt.
Die vom Porometer ermittelten Daten der Netto-Photosyntheserate (JCO2; µmol m-2 s-1) und
der stomatären Leitfähigkeit (g H2O; mmol m-2s-1) wurden über das Diagas Programm für CQP
130 folgendermaßen berechnet:
eOHee
CO cJCww
BFu
J **11
*22
0
−∆−−
=
ALVPDwJg OHOH
−−=
122
ue: molarer Gasstrom am Küvetteneingang [µmol s-1]
BF: projizierte Blattfläche [cm²]
we: molare Feuchtekonzentration vor der Küvette (≡ im Referenzgas)

Material und Methoden
28
w0: molare Feuchtekonzentration hinter der Küvette (≡ im Messgas)
∆C: Differenz der CO2-Konzentration zwischen Referenzgas und Messgas; korrigiert um
Nullpunktverschiebung und Verzerrung des Infrarotgasanalysators (IRGA)
ce: molare CO2-Konzentration vor der Küvette (≡ im Referenzgas); korrigiert um
Nullpunktverschiebung und Verzerrung des Infrarotgasanalysators (IRGA)
JH2O: Transpirationsrate [mmol m-2 s-1]
0
0
1*
2 www
BFu
J eeOH −
−=
ALVPD = wi-wa
wi : Molenbruch des Wasserdampfes in den Interzellularen des Mesophylls
pTsvpw L
i)(
=
svp: Sättigungsdampfdruck (saturated vapor pressure) bei angegebener
Temperatur [mbar]
TL: Blatttemperatur [°C]
wa: Molenbruch des Wasserdampfes in der Küvette
2.8 Messung der Chlorophyll-Fluoreszenz
Die Messungen der Chlorophyll-Fluoreszenz erfolgten, vergleichbar mit den
Gaswechselmessungen, an triebspitzennahen Blättern, die ihre Blattflächenentwicklung
bereits abgeschlossen hatten. Auch hier fanden die Messungen unter konstanten
Klimabedingungen von ca. 22°C, ca. 65% rel. Luftfeuchte und einer PPFD von
100 µmol m-2 s-1 statt.
Um die räumlichen und zeitlichen Veränderungen der Quantenausbeute der
Energieumwandlung im Photosystem II (PS II) visuell darzustellen, wurde das IMAGING-
PAM Chlorophyll Fluorometer der Firma Walz GmbH, Effeltrich, Deutschland verwendet.
Mit diesem System ist auf nichtinvasivem Weg möglich, die Quantenausbeute des PSII
mittels sättigender Lichtblitze (Genty et al., 1989; Schreiber et al., 1986) zu bestimmen. Als
gepulstes Messlicht sowie für die aktinische Ausleuchtung und für die sättigenden Lichtblitze
wird bei der Chlorophyll-Fluoreszenz-Messung mit der IMAGING-PAM blaues Licht des
Wellenbereichs 470 nm verwendet. Durch einen hochspezifischen Pulsverstärker ist

Material und Methoden
29
gewährleistet, dass nur das gepulste Fluoreszenzsignal weiterverarbeitet wird und die
aktinische Lichtintensität ohne Einflussnahme auf die Fluoreszenzmessung verändert werden
kann.
Das zu messende Blatt wurde in einen Probenhalter (IMAGE-USH, Walz GmbH, Effeltrich,
Deutschland) eingespannt (Messfläche 17 x 22 mm) und vor der Reizung für etwa zehn
Minuten an eine PPFD von 100 µmol m-2 s-1 adaptiert. Direkt vor den sättigenden Lichtblitzen
(alle 20 s) wurde jeweils die aktuelle Fluoreszenzausbeute, F, und am Ende der Lichtblitze die
maximale Fluoreszenzausbeute, F´m, erfasst. Hieraus wurde die Quantenausbeute des PSII,
∆F/F´m = (F´m - F) (Nomenklatur nach van Kooten und Snel, 1990), berechnet.
Abb. 2.8: IMAGING-PAM Versuchsaufbau zur Messung der
Chlorophyll-Fluoreszenz an mit Feuer gereizten Pappelblättern
Bereits während der laufenden Messungen können die oben genannten Parameter mittels einer
speziell entwickelten Windows Software (ImagingWin) ermittelt und sowohl in der
Darstellung der gemessenen Blattfläche, als auch in graphischer Form (Abb. 2.8) abgerufen
werden.

Ergebnisse
30
3 Ergebnisse
3.1 Bedeutung der Calciumernährung für den Phänotyp der Pappel
3.1.1 In Hydrokultur
Bereits erste Untersuchungen zum Einfluss der Calciumernährung auf die Holzbildung ließen
eindeutig erkennen, dass sich auch das phänotypische Erscheinungsbild der Pappel bei
Calciummangel verändert. Schon nach sechs Wochen Anzucht von P. tremula x P.
tremuloides in Hydrokultur unter konstanten Bedingungen ließen sich deutliche Unterschiede
in der Biomasseproduktion innerhalb der einzelnen Varianten erkennen (Abb.3.1 A).
B
■■ Wurzel ■ Spross
Abb. 3.1: A: P. tremula x P. tremuloides in Hydrokultur unter verschiedener Ca2+-Versorgung angezogen ; Variante1: 0mM Ca2+, Variante 2: 5mM Ca2+, Variante 3: 10mM Ca2+ .
B: Biomassewerte beziehen sich auf das Frischgewicht der Wurzel (Grob- und Feinwurzeln) und des Sprosses (Spross und Blätter); n=4
0
10
20
30
40
50
60
0 mM Ca 5 mM Ca 10 mM Ca
Bio
mas
se (
FG in
g)
A

Ergebnisse
31
Während die Pappeln, die unter Calciumabstinenz angezogen wurden, kaum Biomasse-
produktion erkennen ließen und bereits nach wenigen Wochen eingingen, erwiesen sich die
Zuwachsunterschiede zwischen der Ca2+-Optimalversorgung und der Ca2+-Überversorgung
als relativ gering (Abb. 3.1 B).
Um zu vermeiden, dass die Pappeln unter völliger Calciumabstinenz noch während der
laufenden Versuchsreihe absterben, wurde eine weitere Versuchsreihe mit leicht veränderten
Nährlösungen gestartet: die minimale Ca2+-Ernährung lag nun bei 0,1 mM Ca2+, eine
reduzierte Variante beinhaltete 1 mM Ca2+ in der Nährlösung, und die optimale Calcium-
Versorgung lag weiterhin bei den in der Hoagland’schen Nährlösung vorgegebenen 5 mM
Ca2+ (vgl. Tab. 2.2). Auch bei dieser Versuchsanordnung konnten schon nach kurzer Zeit
morphologische Unterschiede festgestellt werden. Hatten die Pappeln zu Beginn der
Versuchsreihe alle ein homogenes Erscheinungsbild (Abb. 3.2 A), so konnten bereits nach
zwei Wochen Wachstum in den Hydrokulturen Veränderungen in der Blattausbildung
festgestellt werden: die Blätter der Ca2+-Minimalversorgung zeigten eine deutlich geringere
Blattflächenausbildung und zudem ein Einrollen der Blattränder nach unten (Abb. 3.2 C);
zwischen den verschiedenen Varianten konnte zu diesem Zeitpunkt noch kein Unterschied im
Trieblängenwachstum festgestellt werden (Abb. 3.2 B). Nach einer sechswöchigen
Wachstumsperiode in den modifizierten Nährlösungen war jedoch ein deutlich reduziertes
Trieblängenwachstum bei der minimal Ca2+-versorgten Variante zu erkennen; zwischen der
optimal und der reduziert versorgten Variante hingegen ließ sich zu diesem Zeitpunkt nur ein
geringer Unterschied im Höhenwachstum beobachten (Abb. 3.2 D).
Abbildungen 3.2 A - D:
A
Abb. 3.2 A: P. tremula x P. tremuloides in verschiedenen Nährlösungen am Anfang der Versuchsreihe; homogenes Erscheinungsbild der Pappeln

Ergebnisse
32
B
Abb. 3.2 B: P. tremula x P. tremuloides nach zwei Wochen Wachstum in Hydrokultur;
kein Unterschied im Trieblängenwachstum, jedoch in der Blattausformung der Ca2+-minimierten Variante
C
Abb. 3.2 C: Ca2+-minimierte Pappeln nach zwei Wochen in Hydrokultur;
reduzierte Blattflächen und Wölbung der Blattränder nach unten

Ergebnisse
33
D
Abb. 3.2 D: P. tremula x P. tremuloides nach sechs Wochen Wachstum in Hydrokultur; deutlich reduziertes Trieblängenwachstum unter Ca2+-Minimierung (rechts), leichter Rückgang unter Ca2+-
Reduzierung (mitte) im Vergleich zur Ca2+-Normalversorgung (links). Gleichbleibende Abnormalitäten in der Blattausformung der Minimalvariante.
3.1.2 Im Freiland
Da die Anzucht von Pappeln in Hydrokultur in ihrer zeitlichen Ausdehnung begrenzt ist und
die klimatischen Bedingungen standardisiert sind, lässt sich bei diesem Versuchsaufbau keine
Aussage über eine jahreszeitliche Entwicklung unter verschiedenen Ernährungsvarianten
treffen. Um eine solche Fragestellung zu beantworten wurden Populus trichocarpa - Klone
vor dem Beginn der Vegetationsperiode als Stecklinge in ein nährstoffarmes Bodensubstrat
gebracht und im Freiland unter Ca2+-Minimierung, Ca2+-Reduzierung und Ca2+-
Normalversorgung angezogen. Alle Pflanzen hatten zu Versuchsbeginn ein homogenes
Äußeres. Die Bewurzelung der Stecklinge zum einen, aber auch der kühle und sonnenarme
Frühling 2004 könnten der Grund für einen relativ späten Blattaustrieb der Stecklinge im Juni
sein.
Die optischen Eindrücke (Abb. 3.6 A-C) eines mit steigender Ca2+-Versorgung in der
Nährlösung ebenfalls ansteigenden Zuwachses konnten durch diverse Messungen bestätigt
werden. Der Holzzuwachs, der sich im Zuwachs des Triebdurchmessers widerspiegelt, stieg
während der Vegetationsperiode in direkter Abhängigkeit zur steigenden Ca2+-Versorgung in
den Nährlösungen (Abb. 3.3) an. Die leichten Rückgänge in den Werten der reduzierten und
der optimierten Variante Anfang September sind nicht biologisch zu erklären, sondern
ergeben sich vielmehr aus der Tatsache der Korkleisten an der Rinde der Triebe, die eine
exakte Wiederholbarkeit der Durchmessermessungen erheblich erschwerten; der Mittelwert

Ergebnisse
34
aus zwei Messungen, mit welchem versucht wurde, diese Schwierigkeit zu überwinden,
konnte in diesem Fall die Abweichungen offensichtlich nicht ausreichend bereinigen.
Mitt
eldu
rchm
esse
r in
mm
Juli August September
0
2
4
6
8
10M
ittel
durc
hmes
ser i
n m
m
Juli August September
0
2
4
6
8
10
5 mM Ca1 mM Ca
0,1 mM Ca
5 mM Ca1 mM Ca5 mM Ca1 mM Ca
0,1 mM Ca Abb. 3.3: Durchmesserzuwachs von P. trichocarpa während einer
Vegetationsperiode unter verschiedener Ca2+-Versorgung
Auch bei dem Trieblängenwachstum konnte man zwischen den verschiedenen Ca2+-
versorgten Varianten Wachstumsunterschiede erkennen; ähnlich wie beim
Durchmesserzuwachs wurde auch beim Trieblängenzuwachs ein mit ansteigendem Ca2+-
Gehalt in den Nährlösungen ansteigendes Trieblängenwachstum deutlich. Während der
Zuwachs der Trieblängen bei der optimal mit Calcium versorgten Variante kontinuierlich
anstieg, konnten die reduzierte Variante mit 1 mM Ca2+ in der Nährlösung und die minimierte
Variante mit 0,1 mM Ca2+ in der Nährlösung eine deutliche Zuwachssteigerung erst Mitte
August aufweisen (Abb. 3.4).
Trie
blän
genw
achs
tum
in c
m
Juli August September
0
40
80
120
160
200
Trie
blän
genw
achs
tum
in c
m
Juli August September
0
40
80
120
160
200
5 mM Ca1 mM Ca
0,1 mM Ca
5 mM Ca1 mM Ca5 mM Ca1 mM Ca
0,1 mM Ca Abb. 3.4: Trieblängenzuwachs von P. trichocarpa während einer
Vegetationsperiode unter verschiedener Ca2+-Versorgung

Ergebnisse
35
Der Blattflächenzuwachs, welcher am Ende der Wachstumsperiode rückwirkend über eine
Mischprobe für die Anzahl der während der Vegetationsperiode gemessenen Blätter ermittelt
wurde, zeigte den deutlichsten Unterschied zwischen der Ca2+-Minimalvariante und den Ca2+-
optimal- und Ca2+-reduzierten Varianten; bei nur 0,1 mM Ca2+ in der Nährlösung ergab sich
ein sehr geringer Blattflächenzuwachs während der gesamten Vegetationsperiode. Einen
bedeutend höheren Blattflächenzuwachs konnten die Pappeln der Ca2+-reduzierten Variante
aufweisen, den höchsten jedoch die optimal mit Calcium versorgten Pflanzen. Zwischen den
beiden letztgenannten fand gegen Ende der Vegetationsperiode eine starke Annäherung der
Blattflächenausprägung statt (Abb. 3.5).
0
200
400
600
800
1000
1200
1400
Bla
ttflä
che
in c
m²
Juli August September
0
200
400
600
800
1000
1200
1400
Bla
ttflä
che
in c
m²
Juli August September
5 mM Ca1 mM Ca
0,1 mM Ca
5 mM Ca1 mM Ca5 mM Ca1 mM Ca
0,1 mM Ca Abb. 3.5: Blattflächenzuwachs von P. trichocarpa während einer
Vegetationsperiode unter verschiedener Ca2+-Versorgung
Die Standardabweichungen zu den Mittelwerten der oben abgebildeten Grafiken wurden aus
Gründen der Übersichtlichkeit nicht in die jeweilige Grafik integriert, sondern sind im
Anhang in Tabellenform ihren entsprechenden Werten zugeordnet.

Ergebnisse
36
Abbildungen 3.6 A - C:
A
Abb. 3.6 A: P. trichocarpa - Stecklinge im Juli
B
Abb. 3.6 B: P. trichocarpa - Stecklinge im August

Ergebnisse
37
C
Abb. 3.6 C: P. trichocarpa - Stecklinge im September
3.2 Bedeutung der Calciumernährung für den Calciumhaushalt der
Pappel
Der Einfluss der Calciumernährung auf die Calciumgehalte der unterschiedlichen
Pappelgewebetypen wurde mittels energiedispersiver Röntgenanalyse (EDXA) untersucht.
Die dazu durchgeführten Analysen erfolgten auf mikroskopischer Ebene in Kombination mit
der Rasterelektronenmikroskopie (REM) und ermöglichten so eine ortsgenaue Zuordnung von
einzelnen Messwerten zu den entsprechenden Zellen und Gewebebereichen. Zudem konnte
durch die EDX - Methode ein Überblick über die Elementzusammensetzung der
Gewebetypen gewonnen werden; davon ausgenommen waren jedoch Elemente unterhalb der
Ordnungszahl 9 (=Fluor), die durch die verwendete Methode nicht erfasst werden können.
Um die Auswirkungen der unterschiedlichen Calciumernährung auf den Ionenhaushalt der
Pappel zu untersuchen wurden verschiedene Gewebe der Organe Blatt (Leitbündel), Stamm
(Phloem, Cambium, Xylem-Differenzierungszone) und Wurzel (Zentralzylinder, Cortex) auf
ihre Ca2+-Konzentration in Abhängigkeit zur Nährstoffversorgung untersucht. Als

Ergebnisse
38
0
1
2
3
4
5
6
7
8
Xylem Phloem
rel.
Calc
ium
geha
lt
Pflanzenmaterial dienten aus Zellkultur gewonnene Populus tremula x Populus tremuloides,
welche unter Calciumabstinenz (0 mM Ca2+), unter normaler Calciumversorgung (5 mM
Ca2+) sowie unter Calciumüberversorgung (10 mM Ca2+) für sechs Wochen in Hydrokulturen
angezogen worden waren.
Bei der Röntgenanalyse der Blattleitbündel konnte zwischen dem Xylem- und dem
Phloemgewebe unterschieden werden; in beiden Geweben des Leitbündels wurde eine
parallel zur Nährstoffversorgung ansteigende Calciumkonzentration festgestellt (Abb. 3.7).
Abb. 3.7: Relativer Gehalt an Calciumionen im Leitbündel von Pappelblättern bei unterschiedlicher Ca-Konzentration in den Nährlösungen; Werte ermittelt durch
Peak:Background [P:B]-Verhältnisse von EDX-Analysen des jeweiligen Gewebes (n=5).
Der nahezu lineare Anstieg im Ca2+-Gehalt mit steigender Ca2+-Versorgung war im Phloem
sogar noch deutlicher ausprägt (P:B 5,0 bei 10 mM Ca2+) als im Xylem (P:B 2,9 bei 10 mM
Ca2+).
Der im Xylem und im Phloem des Blattleitbündels zu findende Calciumanstieg unter
verbesserter Ca2+-Versorgung ließ sich auch in den Transportgeweben des Stammes wieder
finden (Abb. 3.8). Auch hier konnte man im Phloem den deutlichsten Anstieg im P:B-
Verhältnis von ca. 0,4 über 1,0 auf 2,2 beobachten. Etwas verhaltener, aber immer noch
deutlich wirkte sich die in der Nährlösung erhöhte Ca2+-Konzentration auf die
Differenzierungszone des Xylems aus; die Calciumionenkonzentration unter völliger Ca2+-
Abstinenz in der Nährlösung hatte auch in der Differenzierungszone, ebenso wie in der
10 mM Ca 5 mM Ca 0 mM Ca

Ergebnisse
39
0
0,5
1
1,5
2
2,5
3
3,5
Phloem Cambium Differenzierungszone
rel.
Calc
ium
geha
lt
Cambiumzone, einen P:B-Wert von 0,4 , stieg unter optimaler Ca2+-Versorgung auf 0,8 ,
unter Ca2+-Überfluss jedoch nur weiter auf einen Wert von 1,0 an. Im Cambium des Stammes
zeigte sich auch ein im Vergleich zum Phloem mit zunehmender Ca2+-Versorgung der
Pflanzen geringerer Ca2+-Anstieg (Abb. 3.8).
Abb. 3.8: Relativer Gehalt an Calciumionen im Phloem, im Cambium und in der Differenzierungszone des Xylems im Stammbereich der Pappel bei unterschiedlicher Ca2+-Konzentration in den Nährlösungen; Werte ermittelt durch Peak:Background-Verhältnisse
von EDX-Analysen des jeweiligen Gewebes (n=5).
Anders als bei den Transportgeweben im Blattleitbündel und im Spross der Pappel konnte
man in der Wurzel keine lineare Kausalität der Ionenkonzentration zur Calciumversorgung
der Nährlösungen feststellen. Die Röntgenanalyse der untersuchten Feinwurzeln ergab hierbei
im Zentralzylinder keine signifikante Veränderung des Calciumgehaltes unter
unterschiedlicher Calciumernährung (Abb. 3.9). Sowohl unter Calciumabstinenz in der
Nährlösung, als auch unter optimaler Calciumversorgung wie unter einem
Calciumüberangebot beliefen sich die im Zentralzylinder gemessenen Ionenwerte für Calcium
zwischen 1,2 und 1,5.
10 mM Ca 5 mM Ca 0 mM Ca

Ergebnisse
40
00,5
11,5
2
2,53
3,54
Zentralzylinder Cortex
rel.
Calc
ium
geha
lt
Abb. 3.9: Relativer Gehalt an Calciumionen im Zentralzylinder und im Cortex der Feinwurzel von Pappeln bei unterschiedlicher Ca2+-Konzentration in den Nährlösungen; Werte ermittelt
durch Peak:Background-Verhältnisse von EDX-Analysen des jeweiligen Gewebes (n=5).
Im Wurzelcortex hingegen war ein deutlicher Anstieg der Calciumkonzentration zwischen der
Ca2+-Nullvariante mit ca. 0,5 hin zur Ca2+-Normalversorgung auf ca. 2,7 festzustellen (Abb.
3.9). Bei einem Überangebot in der Nährlösung änderte sich die Konzentration innerhalb des
Cortexgewebes im Vergleich zu der Optimalversorgung jedoch nicht mehr nennenswert.
10 mM Ca 5 mM Ca 0 mM Ca

Ergebnisse
41
3.3 Bedeutung der Calciumernährung für die Anatomie der Pappel
3.3.1 In Hydrokultur
Um einen möglichen Einfluss der Calciumernährung auf die Gewebeanatomie der Pappel zu
überprüfen, wurden verschiedene Gewebetypen von Populus tremula x Populus tremuloides,
welche für sechs Wochen unter verschiedenen Ca2+-Konzentrationen in den Nährlösungen in
Hydrokultur angezogen worden waren, lichtmikroskopisch analysiert und auf ihre
Unterschiede hin verglichen.
Wurzelgewebe:
Das Wurzelgewebe zeigte deutliche Unterschiede in Aufbau und Struktur zwischen der mit
0 mM Ca2+ versorgten Variante und der optimal mit 5 mM Ca2+ versorgten Variante (Abb.
3.10 A und B). Während sich das Cortexgewebe unter Optimalversorgung als ein fest
gebündeltes Gewebe mit nur geringen oder keinen Interzellularen zwischen den Cortexzellen
zeigte und einen klar begrenzten Zentralzylinder aufwies, konnte man in der unter
Calciumabstinenz angezogenen Variante eine sehr lockere Zellstruktur mit ausgeprägten
Interzellularen und einem kaum abgegrenzten Zentralzylinder beobachten. Diese
Unterschiede zeigten sich weitgehend unabhängig von der Art der Wurzelzone.
A B
Abb. 3.10: A: Wurzelquerschnitt einer Pappel aus Hydrokultur unter 0 mM Ca2+-Versorgung; typisch
ausgeprägte Interzellularen und kaum abgegrenzter Zentralzylinder. B: Wurzelquerschnitt einer Pappel aus Hydrokultur unter 5 mM Ca2+-Versorgung;
typische kompakte Gewebestruktur mit deutlich ausgeprägtem Zentralzylinder.

Ergebnisse
42
Blattgewebe:
Auch im Blattgewebe zeigten sich bei der Pappelanzucht ohne Calcium in der Nährlösung im
Vergleich zu Pappeln, die optimal mit Calcium versorgt waren, deutliche Unterschiede in der
Anatomie. Wieder konnten im Grundgewebe des Leitbündels zahlreiche deutlich ausgeprägte
Interzellularen und ein tendenziell lockerer Gewebeverband beobachtet werden (Abb. 3.11
A); die auch hier als Kontrolle dienende Optimalvariante hingegen zeigte wiederum einen
kompakten Zellverband im Grundgewebe des Leitbündels und dadurch kaum Interzellularen
(Abb. 3.11 B).
A B
Abb. 3.11: A: Blattquerschnitt einer Pappel aus Hydrokultur unter 0 mM Ca2+-Versorgung mit
ausgeprägten Interzellularen im Grundgewebe des Leitbündels. B: Blattquerschnitt einer Pappel aus Hydrokultur unter 5 mM Ca2+-Versorgung;
kompakte Gewebestruktur mit kaum Interzellularen im Grundgewebe des Leitbündels.
Cambiales Gewebe und seine Derivate:
Um den Einfluss der Calciumernährung auf das Cambium und seine Derivate zu untersuchen,
wurden die cambiale Zone sowie die Gefäße und Fasern des Xylems an den
Sprossquerschnitten der unter unterschiedlicher Ca2+-Ernährung angezogenen Pappeln
lichtmikroskopisch analysiert (Abb. 3.12, 3.13, 3.14). Dabei konnten deutliche Unterschiede
in der Ausprägung der cambialen Zone und der Zellexpansionszone des Xylems festgestellt
werden. Nach sechs Wochen Wachstum in Hydrokultur unter Calciumabstinenz war die
cambiale Zone dieser Pappeln sehr gering ausgeprägt; sie betrug nur etwa drei Zellreihen in
radialer Richtung (Abb. 3.12 A, Doppelpfeil). Eine Expansionszone des Xylems war in

Ergebnisse
43
diesem Zellverband nicht mehr deutlich abzugrenzen, vielmehr konnte im Anschluss an die
cambiale Zone eine rasch einsetzende Verholzung der Faserzellen durch die
Sekundärwandbildung beobachtet werden (Abb. 3.12 A).
Im Gegensatz zur Nullvariante zeigte die Kontrollvariante mit 5 mM Ca2+ in der Nährlösung
eine deutlich andere Ausprägung des cambialen Meristems und der Expansionszone; die
cambiale Zone umfasste hier etwa sieben bis acht Zellreihen in radialer Richtung (Abb. 3.12
B) und repräsentierte damit eine aktives Cambium, wie es in der Pappel im Frühjahr zu finden
ist. Die Expansionszone, in der die Gefäße und Fasern ihre Länge, ihren Durchmesser und ihr
Volumen festlegen, betrug in der Kontrollvariante etwa sieben im Anschluss an die cambiale
Zone radial aufeinander folgende Zellreihen (Abb. 3.12 B, lang gezogener Doppelpfeil), an
die sich zentripetal die Sekundärwandbildung anschloss.
A B
Abb. 3.12: Querschnitt durch Pappelstamm A: unter Ca2+-Abstinenz in der Nährlösung Reduktion der
cambialen Zone auf ca. drei Zellreihen in radialer Richtung (Doppelpfeil); cambiumnahe Sekundärwandbildung. B: normal mit Ca2+ versorgte Pappel mit ausgeprägter cambialer Zone
(kurzer Doppelpfeil) und ausgeprägter Zellexpansionszone (langer Doppelpfeil) mit anschließender Sekundärwandbildung.
Die lichtmikroskopischen Untersuchungen des neu gebildeten Holzgewebes ergaben neben
einem generellen Rückgang der Holzbildung unter Ca2+-Abstinenz (Abb. 3.13 C) auch eine
Veränderung der Gefäßquerschnittsflächen (Abb. 3.13 A und B). Die Gefäße im Xylem der
Ca2+-Nullvariante waren in direkter Nachbarschaft zu den gehäuft auftretenden Holzstrahlen
zu finden und wiesen eine durchschnittliche Gefäßgröße von nur etwa 400 µm² im
Querschnitt auf, wohingegen die durchschnittliche Gefäßquerschnittsfläche unter normaler
Ca2+-Versorgung der Pappeln mehr als das Doppelte betrug (Abb. 3.13 C).

Ergebnisse
44
A B
C
Abb. 3.13: Veränderungen der Gefäßquerschnittsflächen in Abhängigkeit des Ca2+-Gehalts in der Nährlösung. A: unter 0 mM Ca2+ in der Nährlösung; Gefäßlumina deutlich reduziert. B: unter optimaler Ca2+-Versorgung (5 mM Ca2+). C: graphische Darstellung der Unterschiede in der
Gefäßgröße (n=170) und im Holzzuwachs (n=4) unter Ca2+-Abstinenz und Ca2+-Optimalversorgung.
Calcium-abhängige Veränderungen im Xylem beschränkten sich nicht nur auf die
Gefäßbildung und ~Ausformung, sondern ließen sich auch an den Fasern beobachten.
Mazerierte Libriformfasern aus dem Xylem von Pappeln, welche für sechs Wochen unter
0,1 mM Ca2+, 1 mM Ca2+ oder 5 mM Ca2+ in Hydrokultur angezogen worden waren, ließen
eine deutliche Abhängigkeit zwischen der Ca2+-Versorgung in der Nährlösung und dem
Faserlängenwachstum erkennen (Abb. 3.14). Von durchschnittlichen 0,52 mm Faserlänge der
jungen Libriformfasern unter optimalen Nährstoffbedingungen konnte unter reduzierter Ca2+-
Ernährung ein Rückgang auf 0,47 mm Faserlänge und unter minimierter Ca2+-Ernährung auf
nur durchschnittliche 0,38 mm festgestellt werden. Damit ließ sich ein Einfluss von Calcium
auf die Faserlänge nachweisen, da mit abnehmender Ca2+-Versorgung in der Nährlösung auch
die Faserlänge abnahm.
0
200
400
600
800
1000
1200
1400
Gefäßgröße Holzzuwachs
Gef
äßgr
öße
in µ
m²
Hol
zzuw
achs
in µ
m
■ 0 m M Ca
■ 5 mM Ca

Ergebnisse
45
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
5 mM Ca 1 mM Ca 0,1 mM Ca
Fase
rläng
e in
mm
Abb. 3.14: Abhängigkeit der Faserlänge von der Ca2+-Ernährung. Oben: Abnahme der durchschnittlichen Faserlänge mit Abnahme des Ca2+-Gehalts in der Nährlösung (n=120);
Student’scher T-Test: P≤0,1. Unten: Mazerierte Libriformfasern unter dem Lichtmikroskop.
3.3.2 Im Freiland
Vergleichende Untersuchungen zur Gefäßentwicklung an P. trichocarpa - Stecklingen, die
unter den gleichen Nährstoffbedingungen im Freiland angezogen worden waren, ergaben
hinsichtlich der Gefäßgröße ein ähnliches Verteilungsbild. Nachdem zwei Pappeln pro
Variante am Ende der Wachstumsperiode auf die Gefäßquerschnittsflächen im neu gebildeten
Jahrring hin vermessen wurden, jeweils ca. 1050 Gefäße pro Variante, konnte man ebenfalls
eine Tendenz zur reduzierten Gefäßquerschnittsfläche bei Ca2+-Abnahme in der Nährlösung
entdecken (Abb. 3.15). So lag die Querschnittsflächenklasse mit der größten absoluten
Häufigkeit bei der minimal mit Ca2+ versorgten Variante bei 400 µm², wohingegen sie bei der
reduzierten Variante auf 1000 µm² anstieg. Bei der Normalvariante (5 mM Ca2+) lag die
größte absolute Häufigkeit ebenfalls bei 400 µm², jedoch erstreckten sich hier die
Querschnittsflächenklassen mit absoluten Häufigkeiten > 130 auf die Klassen 400 µm² bis
5 mM Ca 1 mM Ca 0,1 mM Ca
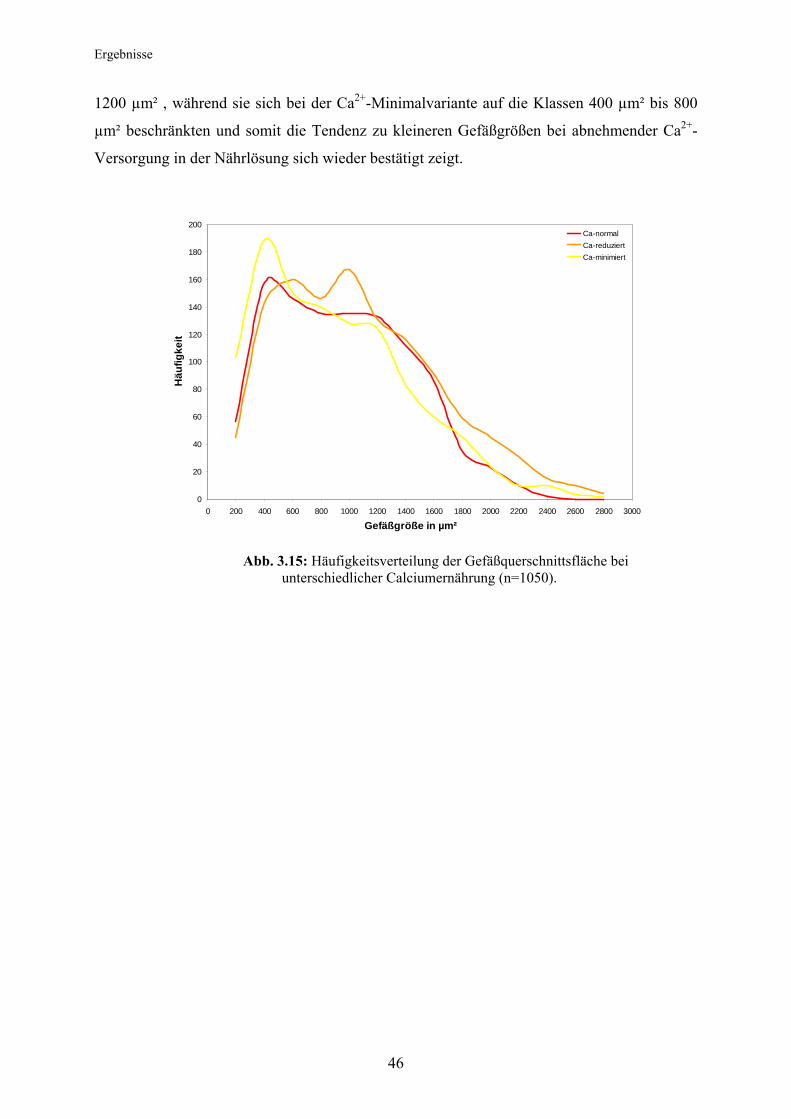
Ergebnisse
46
1200 µm² , während sie sich bei der Ca2+-Minimalvariante auf die Klassen 400 µm² bis 800
µm² beschränkten und somit die Tendenz zu kleineren Gefäßgrößen bei abnehmender Ca2+-
Versorgung in der Nährlösung sich wieder bestätigt zeigt.
0
20
40
60
80
100
120
140
160
180
200
0 200 400 600 800 1000 1200 1400 1600 1800 2000 2200 2400 2600 2800 3000
Gefäßgröße in µm²
Häu
figke
it
Ca-normalCa-reduziertCa-minimiert
Abb. 3.15: Häufigkeitsverteilung der Gefäßquerschnittsfläche bei
unterschiedlicher Calciumernährung (n=1050).

Ergebnisse
47
3.4 Bedeutung der Calciumernährung für die Ultrastruktur und die
Zellwandchemie der Pappel
3.4.1 Ultrastruktur des Cambiums und des Blattes
In Ergänzung zu den anatomischen Untersuchungen der Lichtmikroskopie wurden
ultrastrukturelle Untersuchungen am Transmissionselektronenmikroskop (TEM)
durchgeführt. Zwischen den unter Ca2+-Abstinenz in Hydrokultur angezogenen P. tremula x
P. tremuloides und der entsprechenden Kontrollvariante (5 mM Calcium) ergaben sich dabei
in der Ultrastruktur der cambialen Zellen sowie in den verschiedenen Geweben der Blätter
bemerkenswerte Unterschiede.
Wie die lichtmikroskopischen Untersuchungen bereits andeuteten, war unter optimaler Ca2+-
Versorgung (5 mM) die cambiale Zone mit etwa 7-8 Zellreihen in radialer Richtung stark
entwickelt. Zudem konnte unter elektronenmikroskopischer Betrachtung auch eine
ausgeprägte Vakuolisierung des cambialen Cytoplasmas festgestellt werden (Abb. 3.16 A und
B). Das cambiale Gewebe entsprach mit dieser Ultrastruktur der typischen Erscheinungsform
eines aktiven Cambiums der Pappel (Arend und Fromm, 2003).
Im Gegensatz dazu war unter Ca2+-Abstinenz in der Nährlösung nur eine geringe Ausprägung
der cambialen Zone in radialer Richtung festzustellen; der cytoplasmatische Anteil in diesen
Zellen war deutlich erhöht, die Vakuolen waren zahlreich, aber nur in geringer Größe
vertreten (Abb. 3.16 C und D). Mit diesem Erscheinungsbild entsprach das unter Ca2+-
Abstinenz gebildete cambiale Gewebe eher der typischen Ausprägung eines Pappelcambiums
während der Dormanz (Arend und Fromm, 2003).

Ergebnisse
48
A B
C D
Abb. 3.16: Einfluss der Calciumernährung auf die Ultrastruktur cambialer Zellen. A. Ausgeprägte cambiale Zone unter optimaler Ca2+-Versorgung.
B. Unter optimaler Ca2+-Versorgung nimmt die Vakuole nahezu den gesamten Zellraum ein; das Cytoplasma bildet nur einen schmalen Saum an der Innenseite der Zellwände.
C. Reduzierte cambiale Zone unter Ca2+-Abstinenz in der Nährlösung. D. Unter Ca2+-Abstinenz nimmt das Cytoplasma nahezu den gesamten Zellraum ein und beinhaltet
viele kleine Vakuolen. Abkürzungen: CZ - cambiale Zone; Ph - Phloem; V - Vakuole; X - Xylem

Ergebnisse
49
Nicht nur in der Ultrastruktur der cambialen Zellen konnten unter Ca2+-Abstinenz
Veränderungen wahrgenommen werden, sondern auch in der Ultrastruktur der Blattgewebe.
In den primären Leitbündeln der Blätter konnten zwar in der Ausprägung des Phloems und
des Xylems keine signifikanten Veränderungen festgestellt werden, jedoch konnten unter
Ca2+-Abstinenz keine Calcium-Oxalat-Kristalle im Grundgewebe der Blattadern beobachtet
werden, wie sie im Gegensatz dazu in den optimal mit Calcium versorgten Blättern im
Grundgewebe der Blattadern häufig zu finden waren (Abb. 3.17).
A B
Abb. 3.17: Einfluss der Calciumernährung auf die Ultrastruktur der Blattadern. A. Querschnitt durch Phloem und Grundgewebe einer optimal mit Calcium (5 mM) versorgten
Blattader; deutliche Ansammlung von Ca2+-Oxalat-Kristallen im Grundgewebe (Pfeile). B. Querschnitt durch Phloem und Grundgewebe einer Blattader unter Ca2+-Abstinenz; im
Grundgewebe sind keine Ca2+-Oxalat-Kristalle festzustellen. Abkürzungen: GG - Grundgewebe der Blattadern; Ph - Phloem der Blattadern.

Ergebnisse
50
Weitere Veränderungen im Blattgewebe ließen sich unter Ca2+-Abstinenz an den
Mesophyllzellen beobachten. Einlagerungen von Stärkekörnern in den Chloroplasten, wie sie
unter optimaler Nährstoffversorgung gehäuft auftraten, waren in den Chloroplasten der unter
Ca2+-Abstinenz gebildeten Mesophyllzellen nicht, bzw. nur sehr selten, zu beobachten (Abb.
3.18).
A B
Abb. 3.18: Einfluss der Calciumernährung auf die Ultrastruktur der Mesophyllzellen. A. Typisches Bild der Mesophyllzellen unter optimaler Nährstoffversorgung; in den Chloroplasten
sind häufig Einlagerungen von Stärkekörnern (S) zu beobachten (Pfeile). B. Typisches Bild der Mesophyllzellen unter Ca2+-Abstinenz in der Nährlösung; in den Chloroplasten
sind keine Stärkeeinlagerungen festzustellen.
3.4.2 Chemie der Zellwand
Um einen möglichen Einfluss der Calciumversorgung auf die chemische Zusammensetzung
der Zellwände zu überprüfen, wurden die unter verschiedenen Ca2+-Regimen in Hydrokultur
angezogenen Pappeln (P. tremula x P. tremuloides) mittels FTIR-Spektroskopie untersucht.
Dabei wurde zwischen Proben des Xylemgewebes und der Rinde unterschieden; aus
technischen Gründen konnten Strahl- und Markzellen jedoch nicht aus den Holzproben
separiert werden, ebenso wie die Rindenproben auch das Phloem und Cambium umfassten.

Ergebnisse
51
Die Absorptionsbanden der Proben (n=5) wurden auf der Grundlage der Absorptionsintensität
von Kaliumthiocyanat, welches mit der Wellennummer 2050 cm-1 als interner Standard
fungierte, berechnet (Nakano und Miyazaki 2003, Wiberley et al. 1957, Fraser 1959).
Bei einer Betrachtung der Absorptionsspektren von Xylemproben konnte festgestellt werden,
dass es durchaus eine Abhängigkeit in der chemischen Zusammensetzung der Holzzellwand
von der Calciumversorgung über die Nährlösung gibt. Für den Spektrenbereich zwischen
4000 und 450 cm-1 konnte unter Calciumabstinenz in der Nährlösung im Vergleich zu der
optimal mit Ca2+ versorgten Variante ein Rückgang in der Absorption nahezu aller
funktioneller Gruppen des Xylems beobachtet werden (Abb. 3.19).
Abb. 3.19: Vergleichende Darstellung zweier FTIR-Spektren von Holzproben (n=5) im Wellenzahl-Bereich 4000 cm-1 bis 450 cm-1. Unter Calciumabstinenz in der Nährlösung (schwarze Linie) ist im
Vergleich zur Ca-Optimalversorgung (grüne Linie) ein allgemeiner Rückgang der Zellwandkomponenten zu beobachten. Interner Standard bei 2050 cm-1.
Um die Unterschiede in den funktionellen Gruppen der Holzzellwände besser ersichtlich zu
machen, ist in Abbildung 3.20 ein Ausschnitt (Wellenzahl-Bereich 1800 cm-1 bis 800 cm-1)
der obigen Darstellung gegeben.

Ergebnisse
52
Abb. 3.20: Vergleichende Darstellung zweier FTIR-Spektren von Holzproben (n=5) im Wellenzahl-Bereich 1800 cm-1 bis 800 cm-1. Unter Ca-Abstinenz deutliche Reduktion der Methoxy-Gruppen des
S-Lignins bei Wellenzahl 1330 cm-1 sowie verschiedener Acetylgruppen bei Wellenzahlen 1240 cm-1, 1380 cm-1 und 1740 cm-1.
Die Wellenzahl 1330 cm-1 repräsentiert eine S-Lignin - spezifische Methoxygruppe; der
Rückgang in dieser Bande deutet somit einen Rückgang des S-Ligningehalts in
Holzzellwänden unter Calciumabstinenz in der Nährlösung an. Des weiteren konnten unter
Ca2+-Abstinenz Rückgänge in C-O Doppelbindungen (1740 cm-1), in C-O Bindungen
(1240 cm-1) und in C-H Bindungen (1380 cm-1) von Acetylgruppen festgestellt werden.
Nachdem die Unterschiede im chemischen Aufbau der Holzzellwand zwischen den optimal
mit Ca2+ versorgten und den unter Ca2+-Abstinenz angezogenen Pappeln deutlich
hervortraten, sollte in einem weiteren Schritt untersucht werden, ob es für einen Rückgang der
beobachteten Absorption einen Schwellenwert bei der Calciumernährung gibt. Aus diesem
Grund wurden Holzproben aus minimal mit Ca2+ (0,1 mM) versorgten Pappeln denen aus
Calciumabstinenz gegenübergestellt (Abb. 3.21).
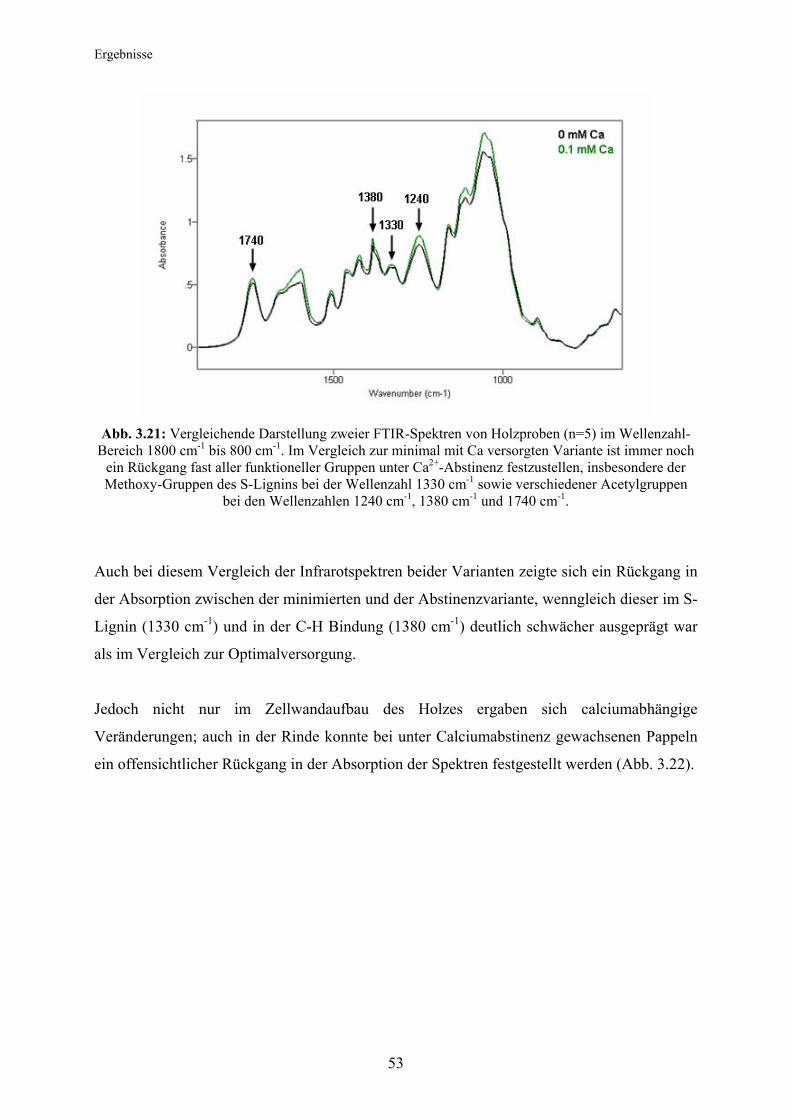
Ergebnisse
53
Abb. 3.21: Vergleichende Darstellung zweier FTIR-Spektren von Holzproben (n=5) im Wellenzahl-
Bereich 1800 cm-1 bis 800 cm-1. Im Vergleich zur minimal mit Ca versorgten Variante ist immer noch ein Rückgang fast aller funktioneller Gruppen unter Ca2+-Abstinenz festzustellen, insbesondere der Methoxy-Gruppen des S-Lignins bei der Wellenzahl 1330 cm-1 sowie verschiedener Acetylgruppen
bei den Wellenzahlen 1240 cm-1, 1380 cm-1 und 1740 cm-1.
Auch bei diesem Vergleich der Infrarotspektren beider Varianten zeigte sich ein Rückgang in
der Absorption zwischen der minimierten und der Abstinenzvariante, wenngleich dieser im S-
Lignin (1330 cm-1) und in der C-H Bindung (1380 cm-1) deutlich schwächer ausgeprägt war
als im Vergleich zur Optimalversorgung.
Jedoch nicht nur im Zellwandaufbau des Holzes ergaben sich calciumabhängige
Veränderungen; auch in der Rinde konnte bei unter Calciumabstinenz gewachsenen Pappeln
ein offensichtlicher Rückgang in der Absorption der Spektren festgestellt werden (Abb. 3.22).

Ergebnisse
54
Abb. 3.22: Vergleichende Darstellung zweier FTIR-Spektren von Rindenproben (n=5) im Wellenzahl-
Bereich 4000 cm-1 bis 450 cm-1. Unter Calciumabstinenz in der Nährlösung (punktierte Linie) ist im Vergleich zur optimal mit Calcium versorgten Variante deutlich ein allgemeiner Rückgang der
Absorption bei fast allen funktionellen Gruppen zu beobachten. Interner Standard bei 2050 cm-1.
Bei den vergleichenden Untersuchungen der Rindenspektren wurde eine signifikante
Reduktion bei der Wellenzahl 1640 cm-1 deutlich. Diese Bande repräsentiert in IR-Spektren
von Zellwänden eine C=O Doppelbindung im Lignin, wie sie beispielsweise in Aryl-Ketonen
vorzufinden ist. In direkter Nachbarschaft zu dieser Wellenzahl absorbiert Coniferylaldehyd
(1652 cm-1), welches somit ebenfalls als unter Ca2+-Abstinenz reduziert gelten kann.

Ergebnisse
55
3.5 Elektrische Signalleitung der Pappel
3.5.1 Signalleitung an Populus trichocarpa
Um die elektrische Signalleitung an der Pappel zu untersuchen, wurden junge Triebe von im
Freiland wachsenden Bäumen (Populus trichocarpa) geerntet und sogleich unter einem
Faraday’schen Käfig mit der Schnittstelle in ein Gefäß mit artificial pondwater (APW)
gestellt. Die für die Messungen geernteten oberen Triebbereiche waren etwa 20 cm lang und
mit mindestens sechs Blättern belaubt, an deren Blattunterseite reichlich für die
Aphidentechnik notwendige Blattläuse zu finden waren. Das oberste Messblatt war stets in
seiner Blattfläche ausgewachsen. Für die Messungen der Signalleitung wurde folgender
Versuchsaufbau eingehalten (Abb. 3.23):
Abb. 3.23: Experimenteller Versuchsaufbau zur Messung der elektrischen Potentiale in der Pappel. Hitzereizung erfolgte durch Stimulation mit einer Flamme an der Spitze von Blatt 1, sowie an der
Blattbasis von Blatt 4. Kältereizung erfolgte durch Stimulation mit Eiswasser an der Blattspitze von Blatt1, sowie an einem tiefer gelegenen Sprossabschnitt. Um die Reizleitung bei Distanzreizen zu
unterbrechen wurde am Spross ein mit Eiswasser gefüllter Trichter (Kälteblock) angebracht.

Ergebnisse
56
Bei Messungen im Mesophyll wurde die Spitze der Mikroelektrode in das Mesophyllgewebe
eingeführt. Das Ruhepotential der Mesophyllzellen lag stets bei den für diesen Gewebetyp
charakteristischen Werten von -80 mV bis -100 mV. Bei Messungen im Phloem wurde die
Mikroelektrodenspitze mit dem aus einem abgetrennten Lausrüssel austretenden Phloemsaft
kontaktiert; das Ruhepotential der Phloemzellen lag dabei stets bei Werten zwischen -116 mV
und -165 mV, was ebenfalls die für das Phloem charakteristische Größenordnung
widerspiegelt (Fromm 1991; Fromm und Bauer 1994).
3.5.1.1 Kältereizung mittels Eiswasser
Wie schon aus vielfachen Versuchen an verschiedenen Pflanzen bekannt ist, kann die
elektrische Erregbarkeit von Pflanzen generell durch Reizung mit Eiswasser überprüft
werden. Am Beispiel der Pappel konnte nun bei einer Stimulation der Blattspitze 1 mit
Eiswasser eine basipetale Signalleitung festgestellt werden (Abb. 3.24). Beide
Mikroelektroden verzeichneten auf die Reizung hin ein elektrisches Signal mit einer
hyperpolarisierenden Amplitude von etwa 25 mV in den Siebröhren des Phloems. Die
Signalleitungsgeschwindigkeit betrug bei diesen Untersuchungen 4 bis 8 mm s-1.
Abb. 3.24: Reizung der Blattspitze von Blatt 1 mit Eiswasser; basipetale Signalleitung
in den Siebröhren mit einer Geschwindigkeit von 4 - 8 mm s-1.
Im Gegensatz dazu konnten bei einer Reizung mit Eiswasser an einem tiefer gelegenen
Sprossabschnitt Aktionspotentiale gemessen werden, für die eine Depolarisation der
Zellmembran charakteristisch sind (Abb. 3.25). Diese bewegten sich über die
Plasmamembranen der Siebröhren mit einer ähnlichen Ausbreitungsgeschwindigkeit acropetal
fort und wiesen dabei Amplituden zwischen 12 und 20 mV auf.

Ergebnisse
57
Abb. 3.25: Reizung am Spross mit Eiswasser; acropetale Signalleitung
in den Siebröhren mit einer Geschwindigkeit von 4 - 8 mm s-1.
Auch hier war drei bis vier Minuten nach der Reizinduktion das Ruhepotential in den
Membranen wiederhergestellt.
3.5.1.2 Hitzereizung mittels offener Flamme
Die Reizungen mit Feuer wurden mittels der offenen Flamme eines Gasfeuerzeugs
durchgeführt; dabei wurde die Flamme für ca. 3 Sekunden so nah an das Gewebe geführt, bis
die von ihr ausgehende Wärme die Zellstruktur sichtlich schädigte (Abb. 3.26).
Abb. 3.26: Beispielhafte Darstellung einer Membranpotentialmessung im Mesophyll bei Feuerreizung an der Blattspitze; Schädigung des Gewebes durch Hitze.

Ergebnisse
58
Bei einem Hitzereiz an der Blattspitze von Blatt 1 konnte sowohl an den beiden
Phloemmesspunkten, als auch im Mesophyll des gereizten Blattes ein basipetal fortlaufendes
elektrisches Signal festgestellt werden. Das Membranpotential hyperpolarisierte mit einer
Amplitude von etwa 25 mV, wobei sich das Signal mit einer Geschwindigkeit von etwa 1 bis
2 mm s-1 fortpflanzte (Abb. 3.27).
Abb. 3.27: Feuerreiz an der Blattspitze; basipetale Signaltransduktion eines hyperpolarisierenden elektrischen Signals im Phloem und im Mesophyll.

Ergebnisse
59
Bei einem Hitzereiz an der Blattbasis von Blatt 4 konnte sowohl an den beiden
Phloemmesspunkten, als auch im Mesophyll ein elektrisches Signal mit einer unregelmäßigen
Erscheinungsform und einer Amplitude von über 50 mV bei Depolarisation des
Membranpotentials aufgezeichnet werden (Abb. 3.28).
Abb. 3.28: Feuerreiz an der Blattbasis von Blatt 4; acropetale Signaltransduktion
eines depolarisierenden elektrischen Signals im Phloem wie im Mesophyll.
Die Leitungsgeschwindigkeit der Signale betrug auch hier 1 bis 2 mm s-1 und deckt sich somit
mit den basipetal gemessenen Leitungsgeschwindigkeiten nach Feuerreizung.
Bemerkenswert an den obigen Messungen ist unter anderem sicherlich, dass die sich basipetal
fortpflanzenden elektrischen Signale die Membranspannung hyperpolarisieren, wohingegen
sich acropetal fortpflanzende Signale die Plasmamembranen depolarisieren; diese Erkenntnis
gilt unabhängig von der Art der Reizung (Kälte- wie Hitzereiz).

Ergebnisse
60
3.5.1.3 Unterbrechung der elektrischen Signalleitung durch einen Kälteblock
Wie bereits an Mais gezeigt werden konnte, kann die Ausbreitung elektrischer Signale durch
das Anlegen eines Kälteblocks unterbunden werden (Koziolek et al., 2005). Um nach diesem
Prinzip die elektrische Signalleitung bei der Pappel zu unterbrechen wurde am Spross der
Versuchspflanzen oberhalb des zu reizenden Blattes ein Trichter befestigt, welcher mit
Eiswasser gefüllt wurde; somit konnte der jeweilige Sprossabschnitt auf konstante + 4°C
gekühlt werden (Abb. 3.29).
Abb. 3.29: Beispielhafte Darstellung des Versuchsaufbaus bei
Distanzsignalmessungen im Phloem mit angelegtem Kälteblock.
Nachdem die Blattbasis von Blatt 4 mit Feuer gereizt wurde, ergaben die Messungen im
oberhalb des Kälteblocks befindlichen Phloem nur eine leichte Membran-Depolarisierung an
beiden Messpositionen (Abb. 3.30).

Ergebnisse
61
Abb. 3.30: Acropetale Signalleitung nach Feuerreiz;
Messungen oberhalb eines Kälteblocks.
Zusätzlich zu diesen Versuchen an abgetrennten Trieben von P. trichocarpa wurden die oben
beschriebenen Messungen auch an intakten P. tremula x P. tremuloides durchgeführt; diese
unter optimaler Nährstoffversorgung in Hydrokultur angezogenen Pappeln wurden samt ihres
intakten Wurzelwerks in APW gestellt und wiesen bei der Signalleitung die gleichen
Resultate auf wie die oben gezeigten Triebspitzen aus P. trichocarpa.
3.5.1.4 Signalleitung an mit TEA+ behandelten Pappeln
Aufgrund der vom klassischen Signalleitungsmodell (vorübergehende Depolarisation des
Membranpotentials) abweichenden Hyperpolarisierung, wie sie bei basipetal fortlaufenden
elektrischen Signalen in der Pappel auftritt (s.o.), wurden P. trichocarpa - Triebe vor der
Feuerreizung für 24 Stunden mit Tetraethylamoniumchlorid (TEA+) behandelt; es wurde
dafür in 1 mM-Konzentration dem APW beigefügt. TEA+ ist ein Kaliumkanal-spezifischer
Blocker, der bei einem elektrischen Signal den K+-Ausstrom aus dem Cytoplasma unterbindet
(Thiel et al., 1996). Somit sollte getestet werden, ob die Hyperpolarisierung des
Membranpotentials durch einen K+-Ausstrom hervorgerufen wird.
Die mit TEA+ behandelten Pappeln zeigten nach einer basipetal gerichteten Signalinduktion
durch einen Feuerreiz an der Blattspitze 1 keine ausgeprägte Veränderung des
Membranpotentials im Phloemgewebe an den beiden Messpositionen Elektrode A und
Elektrode B (Abb. 3.31).

Ergebnisse
62
Abb. 3.31: Basipetale Signalleitung durch Feuerreizung an der Blattspitze 1 nach TEA+-Behandlung
des Sprosses; lediglich an der Elektrode A ist als Reaktion auf das elektrische Signal eine sehr schwache Hyperpolarisierung im Membranpotential festzustellen.
Mit dieser Versuchsreihe konnte auch für ein hyperpolarisierendes Membranpotential die
Abhängigkeit des elektrischen Signals von der Kaliumkanalaktivität belegt werden.
3.5.2 Signalleitung an Calcium-mangelversorgten Pappeln (P. tremula x P. tremuloides)
Über die Ionenabhängigkeit der pflanzlichen Erregbarkeit sind aus der Literatur zahlreiche
Arbeiten bekannt, z. B. bei Characeen (Tazawa et al. 1987) und der Korbweide (Fromm and
Spanswick, 1993).Um weitere Informationen über die am Aufbau der elektrischen Signale
beteiligten Ionenflüsse bei der Pappel zu bekommen wurden unter Ca2+-Mangel (0,1 mM) in
Hydrokultur angezogene Pappeln hinsichtlich elektrischer Erregbarkeit überprüft.
Der Versuchsaufbau war dabei deckungsgleich mit dem oben beschriebenen, nur dass hier
keine abgetrennten Triebspitzen in APW gegeben wurden, sondern die ganze Pflanze mit
intaktem Wurzelsystem.
Nach einer Stimulation durch einen Hitzereiz an der Blattspitze von Blatt 1 konnte an keiner
der beiden im Phloem messenden Elektroden ein elektrisches Signal gemessen werden (Abb.
3.32).

Ergebnisse
63
Abb. 3.32: Basipetale Signalleitung nach Feuerreiz an der Blattspitze von Blatt 1; die gemessenen
Pappeln waren für sechs Wochen unter Calcium-Mangel in Hydrokultur angezogen worden.
Nach einer Feuerreizung an der Blattbasis von Blatt 4 waren im Gegensatz dazu jedoch
Signale, wenngleich auch mit wesentlich geringer Amplitude, im Phloem festzustellen (Abb.
3.33).
Abb. 3.33: Acropetale Signalleitung nach Feuerreiz an der Blattbasis von Blatt 4; die bemessenen
Pappeln waren für sechs Wochen unter Calcium-Mangel in Hydrokultur angezogen worden.
Diese Resultate sind ein Hinweis darauf, dass Calciumflüsse am Aufbau der elektrischen
Signale an der Pappel beteiligt sind.

Ergebnisse
64
3.6 Ionengehalte in geöffneten und geschlossenen Pappelstomata
Untersuchungen über die Öffnungsgeschwindigkeit von Stomata der Pappel im Vergleich zu
anderen, krautigen Pflanzen (Tabak, Ackerschmalwand und Saubohne) haben gezeigt, dass
die Pappel sehr schnell mit ihrem Spaltöffnungsapparat auf veränderte Umweltbedingungen
reagieren kann. Beispielsweise konnte gezeigt werden, dass die stomatäre
Öffnungsgeschwindigkeit unter CO2 - freier Luft bis zu fünf mal schneller ablief als bei
Nicotiana tabacum, und sich auch die Schließgeschwindigkeit mit durchschnittlichen
2,3 µmol m-2 sec-2 als noch deutlich schneller erwies als bei Tabak (1,4 µmol m-2 sec-2)
(Langer et al., 2004). P. tremula x P. tremuloides erwies sich somit im Vergleich zu anderen
höheren Pflanzen als sehr reaktionsstark in der stomatären Apparatur. Nach unseren
Erkenntnissen begründet sich die rasche Gaswechselreaktion dieses Pappelklons zum einen
auf die hohe stomatäre Dichte der Blattoberfläche, zum anderen auf das optimale Verhältnis
zwischen der Schließzellenoberfläche und ihrem Volumen (Abb. 3.34).
A B
Abb. 3.34: Schließzellenanatomie von P. tremula x P. tremuloides. A Rasterelektronische Aufnahme der Blattunterseite; hohe stomatäre Dichte.
B Transmissionselektronenmikroskopische Aufnahme eines Querschnitts durch einen Schließzellenapparat.
Die Anzahl der Schließzellen pro mm² beträgt bei dem P. tremula x P. tremuloides Klon T89
auf der Blattunterseite etwa 190 und ist damit in etwa gleich mit der Schließzellenanzahl pro
mm² von Arabidopsis thaliana ; auch in der Größe des Schließzellenapparates ist diese Pappel
mit 267 µm² durchaus mit Schließzellen der Ackerschmalwand, welche etwa 390 µm²

Ergebnisse
65
messen, vergleichbar (Langer et al., 2004). Dennoch beträgt die Öffnungsgeschwindigkeit bei
Pappel das etwa 2,5-fache von Arabidopsis. Um den Mechanismus der
Schließzellenbewegung bei Pappel näher zu untersuchen und Ursachen für die schnelle
Öffnungsgeschwindigkeit zu erforschen, wurden die Ionengehalte der Pappelschließzellen im
geöffneten sowie im geschlossenen Zustand mittels EDXA untersucht. Die Proben der
geöffneten Stomata wurden von Blättern während der Lichtphase aus CO2-freier Luft
geerntet; die Proben der geschlossenen Stomata wurden, zwei Stunden nachdem sie mit
100 µM ABA besprüht worden waren, von Pappeln während der Dunkelphase geerntet (Abb.
3.35).
Abb. 3.35: EDX-Analyse an Pappelschließzellen. Die abgebildeten Spektren spiegeln die relative
Ionenkonzentration in den geöffneten, bzw. geschlossenen Schließzellen wieder. Cr-Peaks resultieren aus der Chrombesputterung der Proben.
Die elementspezifischen Röntgenspektren für Kalium, Chlorid, Calcium und Phosphor
wurden für geöffnete und für geschlossenen Stomata bestimmt (n=5); dabei zeigten sich in

Ergebnisse
66
den Kalium- sowie in den Chloridgehalten der Schließzellen zwischen den beiden
physiologischen Zuständen deutliche Unterschiede (Abb. 3.36).
Abb. 3.36: Relative Kalium- und Chloridgehalte der Stomata im geöffneten und im geschlossenen
Zustand, berechnet aus den Peak-Werten von je fünf Spektren. Im geschlossenen Zustand senkte sich die Chloridkonzentration auf 70%, die Kaliumkonzentration auf 60% des Gehaltes im geöffneten
Zustand.
Beide Ionengehalte lagen in geschlossenen Stomata mit deutlich niedrigerer Konzentration
vor als in geöffneten Stomata. Der Kaliumgehalt in den offenen Schließzellen belichteter
Blätter betrug demnach die 1,6-fache Konzentration dessen der geschlossenen Schließzellen;
der Chloridgehalt in den Stomata betrug im geöffneten Zustand das 1,4-fache des
Chloridgehaltes bei Dunkelheit. Diese Unterschiede in den Ionenkonzentrationen zwischen
beiden Zuständen zeigen, dass K+ und Cl- eine große osmotische Bedeutung bei der
Schließzellenbewegung haben. Die schnelle Öffnungsgeschwindigkeit der Pappel-
schließzellen lässt sich durch die elektrischen Eigenschaften der K+-Aufnahmekanäle
erklären. Mikroelektrodenmessungen in Schließzellen von intakten Blättern zeigten, dass die
K+-Einwärtsströme eine hohe Stromdichte von 4,4 pA µm-2 Schließzellenoberfläche hatten.
Diese Stromdichte sowie die Zahl der aktiven K+-Kanäle ist wesentlich höher als in
Schließzellen anderer Pflanzenarten (Langer et al., 2004).

Ergebnisse
67
3.7 Der Einfluß der elektrischen Signalleitung auf den Blattgaswechsel
der Pappel: stomatäre Leitfähigkeit für Wasserdampf und
Dunkelreaktion der Photosynthese
3.7.1 Bei getopften Populus trichocarpa - Pflanzen
Um die Auswirkungen der elektrischen Signale auf den Blattgaswechsel der Pappel zu
untersuchen wurden triebspitzennahe, bereits vollständig ausdifferenzierte Blätter getopfter
Populus trichocarpa in einer Klimakammer gemessen. Auch hier wurde wieder zwischen
basipetal gerichteten Kurzstreckensignalen innerhalb desselben Blattes und zwischen
acropetalen, sich über den Spross hinweg fortpflanzenden Distanzsignalen unterschieden. Die
im Folgenden dargestellten Messungen stehen repräsentativ für alle im Anhang aufgeführten.
Bei einer Verwundung des Blattgewebes durch einen Hitzereiz an der Messblattspitze mit
einer Distanz von 3 cm zur Gaswechselküvette (Abb. 3.37 A) konnte 30 Sekunden nach
Stimulation ein starker Abfall der CO2 - Aufnahmerate (JCO2) festgestellt werden (Abb. 3.37
B); innerhalb von 200 Sekunden fiel JCO2 von etwa 1,6 µmol m-2 s-1 auf Werte um -0,4 µmol
m-2 s-1, bevor sie sich nach etwa einer Minute wieder in Richtung auf ihren Ausgangszustand
hin erholte. Die ursprünglichen Werte konnten im Laufe der Messung jedoch nicht wieder
erreicht werden. Weiteren Beobachtungen zufolge stellten sich diese jedoch nach etwa 30
Minuten wieder ein.
Im Gegensatz zu der CO2 - Aufnahmerate konnte für die stomatäre Leitfähigkeit für
Wasserdampf (g H2O) nach Stimulation der Blattspitze durch einen Hitzereiz keine signifikante
Änderung in den Gaswechselwerten festgestellt werden. Auch nach der Reizung bewegten
sich hier die Werte zwischen 110 und 125 mmol m-2 s-1 (Abb. 3.37 B).

Ergebnisse
68
A
B
Abb. 3.37: Einfluss von Kurzstreckensignalen aus Hitzereizung auf den Gaswechsel des gereizten Blattes. A: Versuchsaufbau; basipetale Signalleitung durch Hitzereiz an der Messblattspitze zum
Zeitpunkt 0. B: Gaswechselmessung; 30 s nach Stimulation deutlicher Abfall von JCO2.
Bei einer acropetalen Signalleitung über den Spross konnten ähnliche Reaktionen im
Gaswechsel des triebspitzennahen Messblattes festgehalten werden. Nach Verwundung der
Blattbasis des unteren Blattes in über 10 cm Entfernung durch einen Hitzereiz (Abb. 3.38 A)
konnte 140 Sekunden nach der Stimulation am oberen Messblatt ein starker Abfall der CO2 -
Aufnahmerate verzeichnet werden. Auch hier bewegte sich JCO2 gegen Null und verharrte für
etwa zwei Minuten auf diesem Tiefpunkt, bevor sich der Wert wieder langsam seinem
ursprünglichem Niveau von 1,4 µmol m-2 s-1 näherte (Abb. 3.38 B). Ähnlich wie bei der
Kurzstreckensignalleitung wurde auch hier im Gegensatz zur CO2 - Aufnahmerate keine
signifikante Änderung der stomatären Leitfähigkeit festgestellt; die Werteskala für gH2O
bewegte sich auch während dieses Versuchs unverändert zwischen 110 und 125 mmol m-2 s-1.

Ergebnisse
69
A
B
Abb. 3.38 : Einfluss von Distanzsignalen aus Hitzereizung auf den Gaswechsel des >10 cm entfernten Messblatts. A: Versuchsaufbau; acropetale Signalleitung durch Hitzereiz an der Blattbasis eines
unteren Blattes zum Zeitpunkt 0. B: Gaswechselmessung; 140 s nach Stimulation deutlicher Abfall von JCO2 des Messblatts.

Ergebnisse
70
Die interne CO2-Konzentration stieg während des Rückgangs in der CO2-Aufnahme sowohl
bei acropetal, wie auch bei basipetal gerichteter Signalleitung um circa 25 ppm
antiproportional zu JCO2 an (Abb. 3.39).
A B
Abb. 3.39: Interne CO2-Konzentration (ci) bei Kurzstreckensignalen (A) sowie bei Distanzsignalen (B) aus Hitzereizung. Werte resultieren aus den gleichen Messungen wie
Abb. 3.37 B, bzw. Abb. 3.38 B.
Um die Abhängigkeit des rapiden Abfalls von JCO2 von der elektrischen Signalleitung nach
Verwundung durch einen Hitzereiz zu überprüfen, wurde am Sprossabschnitt zwischen dem
zu verletzenden Blatt und dem Messblatt eine Kälteblock angelegt; hierfür wurde ein mit
Eiswasser gefüllter Trichter an dem Sprossabschnitt befestigt und dieser somit auf konstante
+ 4°C gekühlt (Abb. 3.40 A). Bei einer Hitzereizung der Blattbasis des unteren Blattes wurde
dadurch die elektrische Signalleitung in acropetale Richtung reduziert (Abb. 3.30); die CO2 -
Aufnahmerate sowie die stomatäre Leitfähigkeit für Wasserdampf des Messblattes zeigten
daraufhin keine Veränderung (Abb. 3.40 B). Damit konnte der kausale Zusammenhang des
Abfalls von JCO2 mit der elektrischen Signalleitung eindeutig in Verbindung gebracht werden.

Ergebnisse
71
A
B
Abb. 3.40: Auswirkung eines Kälteblocks auf den Gaswechsel bei acropetaler Signalleitung nach Reizinduktion durch Verwundung eines Blatts. A: Versuchsaufbau; Feuerreizung an der Blattbasis
des unteren Blattes. Das elektrische Signal wird durch einen Kälteblock am Spross gehemmt. B: Gaswechselmessung; keine signifikanten Veränderungen in JCO2 nach Stimulation
des unteren Blattes zum Zeitpunkt 0.
3.7.2 Bei unterschiedlicher Calciumernährung von Populus tremula x Populus
tremuloides
Um die Erkenntnisse der Auswirkungen der elektrischen Signalleitung auf den Gaswechsel
um den Einfluss der Calciumernährung zu erweitern, wurden unter verschiedenen Ca2+-
Regimen in Hydrokultur angezogene P. tremula x P. tremuloides untersucht. Bei den beiden
untersuchten Varianten handelte es sich um eine Calcium-Minimalversorgung (0,1 mM Ca2+)
und um eine Calcium-Normalversorgung (5 mM Ca2+). Da die Blätter bei diesen in
Hydrokultur angezogenen Pappeln unter Calciummangel sehr klein waren, musste auf eine
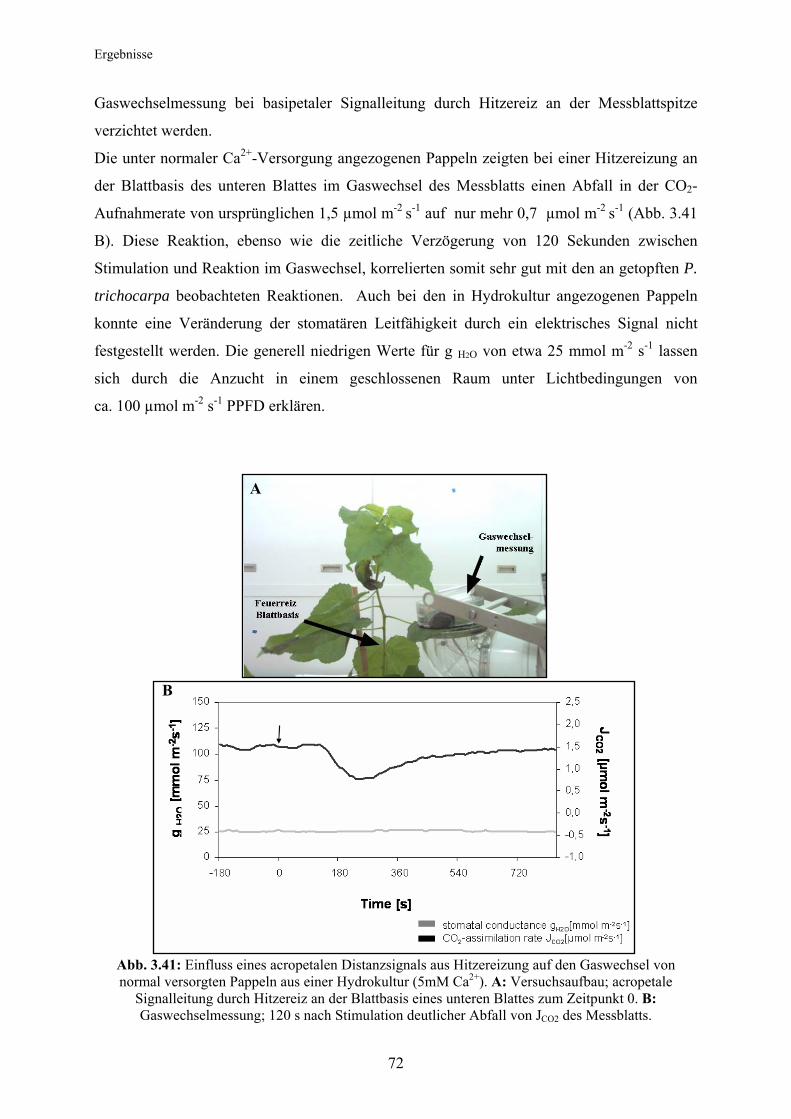
Ergebnisse
72
Gaswechselmessung bei basipetaler Signalleitung durch Hitzereiz an der Messblattspitze
verzichtet werden.
Die unter normaler Ca2+-Versorgung angezogenen Pappeln zeigten bei einer Hitzereizung an
der Blattbasis des unteren Blattes im Gaswechsel des Messblatts einen Abfall in der CO2-
Aufnahmerate von ursprünglichen 1,5 µmol m-2 s-1 auf nur mehr 0,7 µmol m-2 s-1 (Abb. 3.41
B). Diese Reaktion, ebenso wie die zeitliche Verzögerung von 120 Sekunden zwischen
Stimulation und Reaktion im Gaswechsel, korrelierten somit sehr gut mit den an getopften P.
trichocarpa beobachteten Reaktionen. Auch bei den in Hydrokultur angezogenen Pappeln
konnte eine Veränderung der stomatären Leitfähigkeit durch ein elektrisches Signal nicht
festgestellt werden. Die generell niedrigen Werte für g H2O von etwa 25 mmol m-2 s-1 lassen
sich durch die Anzucht in einem geschlossenen Raum unter Lichtbedingungen von
ca. 100 µmol m-2 s-1 PPFD erklären.
A
B
Abb. 3.41: Einfluss eines acropetalen Distanzsignals aus Hitzereizung auf den Gaswechsel von normal versorgten Pappeln aus einer Hydrokultur (5mM Ca2+). A: Versuchsaufbau; acropetale
Signalleitung durch Hitzereiz an der Blattbasis eines unteren Blattes zum Zeitpunkt 0. B: Gaswechselmessung; 120 s nach Stimulation deutlicher Abfall von JCO2 des Messblatts.

Ergebnisse
73
Im Gegensatz zu der obigen Reaktion zeigten Hydrokulturpflanzen, welche unter einer
minimalem Ca2+-Versorgung angezogen wurden, auf eine acropetales Distanzsignal über den
Spross hinweg keine Veränderungen im Blattgaswechsel (Abb. 3.42 B). Sowohl die CO2-
Aufnahmerate, als auch die stomatäre Leitfähigkeit zeigten nach der Stimulation zum
Zeitpunkt 0 keine signifikanten Änderungen und verblieben weiterhin in ihren
Wertebereichen von etwa 1,7 µmol m-2 s-1 (JCO2), bzw. 40 mmol m-2 s-1 (g H2O).
A
B
Abb. 3.42: Einfluss eines acropetalen Distanzsignals aus Hitzereizung auf den Gaswechsel von
minimal mit Calcium versorgten Pappeln aus einer Hydrokultur (0,1mM Ca2+). A: Versuchsaufbau; acropetale Signalleitung durch Hitzereiz an der Blattbasis eines unteren Blattes.
B: Gaswechselmessung; keine signifikante Änderung von JCO2 und g H2O nach Stimulation durch einen Hitzereiz an der Blattbasis eines unteren Blattes zum Zeitpunkt 0.

Ergebnisse
74
3.8 Auswirkung der elektrischen Signalleitung auf die Lichtreaktionen
der Photosynthese der Pappel
Der Einfluss von elektrischen Signalen auf verschiedene Prozesse der Pflanzenphysiologie ist
seit ihrer Entdeckung ein Forschungsbereich, der weltweit schon viele Botaniker faszinierte.
Jedoch erst in jüngster Vergangenheit gelang es anhand von Mimosa pudica einen
Zusammenhang zwischen elektrischen Signalen und der Photosynthese aufzudecken
(Koziolek et al., 2004).
Um mögliche Auswirkungen der elektrischen Signale auf die Lichtreaktionen der
Photosynthese der Pappel zu überprüfen, wurde ein Chlorophyll Fluorometer der Firma Walz
(Heinz Walz GmbH, Effeltrich, Deutschland), das Imaging-PAM, verwendet. Mit Hilfe dieses
Gerätes konnte die Quantenausbeute der Energieumwandlung des Photosystems II räumlich
wie zeitlich aufgelöst werden.
Die noninvasiven Messungen wurden an triebspitzennahen, ausdifferenzierten Blättern
intakter Pflanzen (Populus trichocarpa) durchgeführt, die in ihrer Position mit der Skizze
3.23 in Kapitel 3.5.1 vergleichbar waren.
Mittels einer Flamme wurde die Blattspitze des Messblattes (Blatt 1) gereizt. Die Distanz zur
gemessenen Blattfläche in der Messblattmitte betrug ca. 3 cm. Dort wurde 80 Sekunden nach
der Stimulation an der Blattspitze eine einsetzende Abnahme in der photosynthetischen
Quantenausbeute des PSII deutlich, welche sich innerhalb von 160 Sekunden von den
ursprünglichen Werten um 0,6 , in der Falschfarbendarstellung blau angezeigt, auf Werte um
0,2 , in der Falschfarbendarstellung gelb angezeigt, substanziell reduzierte (Abb. 3.43). Die
photosynthetische Reaktion auf das sich basipetal fortsetzende elektrische Signal zeigt somit
eine deutliche Verzögerung zu dem eigentlichen elektrischen Signal, welches an der
Elektrode A bereits zwei Sekunden, an der Elektrode B 60 Sekunden nach der Stimulation der
Blattspitze aufgezeichnet werden konnte. Nachdem die Quantenausbeute ihr Minimum nach
etwa 300 Sekunden erreicht hatte, begann sie wieder in Richtung auf ihren Ausgangswert
anzusteigen.

Ergebnisse
75
Abb. 3.43: Räumliche und zeitliche Veränderungen in der photosynthetischen Quantenausbeute des PSII; ∆F/F´m. Die Bildausschnitte (22 x 17 mm) zeigen die Blattmitte des Messblattes 1. Stimulation der Messblattspitze durch Hitzereiz zum Zeitpunkt 0s; Distanz 3 cm. Erste Veränderungen in ∆F/F´m
nach 80 Sekunden.
Die Abbildung 3.44 zeigt im Gegensatz dazu die photosynthetischen Reaktionen im
Pappelblatt auf ein Distanzsignal. Nachdem die Blattbasis von Blatt 4 mit einem Hitzereiz
stimuliert wurde, war 240 Sekunden später in der Blattmitte von Blatt 1 ein Rückgang der
photosynthetischen Quantenausbeute festzustellen. Dabei wurde beobachtet, dass die
Blattadern von Blatt 1 zuerst auf das Signal reagierten, die Intercostalfelder zeitlich verzögert
(Abb. 3.45 B), was darauf schließen lässt, dass sich das acropetal fortpflanzende elektrische
Signal über die Blattadern ins Mesophyll ausbreitet; dort konnte nach 360 Sekunden eine
Reduktion der Quantenausbeute von ca. 0,6 auf ca. 0,2 festgestellt werden. Auch bei diesem
Distanzsignal konnte eine deutliche Verzögerung der Reaktion in der Fluoreszenz im
Vergleich zum elektrischen Signal, welches bereits 80 Sekunden nach Stimulation an der
Elektrode A aufgezeichnet werden konnte (vgl. Kapitel 3.5.1.2, Abb. 3.28), gemessen werden.
Die Reaktion in der Fluoreszenz erreichte auf der einen Blattseite von Blatt 1 ihr Maximum
etwa 420 Sekunden nach der Stimulation (Abb. 3.44, offener Pfeil), bevor sie sich wieder
ihrem Ursprungswert annäherte.
Getrennt durch die Mittelrippe erreichte die gegenüberliegende Blattseite ihren minimalen
Wert in der photosynthetischen Quantenausbeute erst nach etwa 600 Sekunden, welcher dann
ebenfalls bei 0,1 bis 0,2 lag (Abb. 3.44, geschlossener Pfeil).

Ergebnisse
76
Abb. 3.44: Räumliche und zeitliche Veränderungen in der photosynthetischen Quantenausbeute des PSII; ∆F/F´m. Die Bildausschnitte (22 x 17 mm) zeigen die Blattmitte von Blatt 1. Stimulation der Blattbasis von Blatt 4 durch Hitzereiz zum Zeitpunkt 0 s; Distanz 11 cm. Erste Veränderungen in
∆F/F´m nach 240 Sekunden.
Bei näherer Betrachtung des basipetal laufenden Kurzstreckensignals und des acropetal
laufenden Distanzsignals wird hinsichtlich der zeitlichen Reaktion in der Fluoreszenz ein
weiterer Unterschied deutlich. Wie in Abbildung 3.45 A ersichtlich ist, löst das durch
Hitzereiz an der Messblattspitze induzierte elektrische Signal eine simultane Reaktion in der

Ergebnisse
77
Fluoreszenz, sowohl in der Blattnervatur, als auch in den Intercostalfeldern, aus; in beiden
Bereichen des Blattes wird in 4 cm Entfernung zum verwundeten Blattgewebe nach 160
Sekunden ein gleichzeitig einsetzender Rückgang der photosynthetischen Quantenausbeute
gemessen. Ein anderes Bild jedoch lässt sich bei genauerer Betrachtung des acropetal
laufenden Distanzsignals erkennen. Hier war nach Reizung der Blattbasis von Blatt 4 in der
Blattmitte von Messblatt 1 deutlich zu erkennen, dass die Reaktion in der Fluoreszenz zuerst
in den Blattadern erfolgte, bevor sie auf die benachbarten Intercostalfelder überging (Abb.
3.45 B, vergrößerter Ausschnitt). Sie trat dort mit einer ca. 60-sekündigen Verzögerung auf.
Abb. 3.45: Kinetik der Veränderungen in der photosynthetischen Quantenausbeute des PSII, ∆F/F´m,
gemessen mittels Fluoreszenz Imaging-PAM in der Blattmitte von Messblatt 1. Die Graphen repräsentieren in ihren Farben die Flächen von Adern und angrenzenden Intercostalfeldern, wie sie in
den unteren Darstellungen abgebildet sind. A Hitzereiz an der Messblattspitze zum Zeitpunkt 0. B Hitzereiz an der Blattbasis von Blatt 4 zum Zeitpunkt 0. Die Verzögerung der Reaktion zwischen Blattadern und Intercostalfeldern ist hier durch eine Vergrößerung der relevanten Zeitpunkte in der
Grafik hervorgehoben.
Der Unterschied in der Fluoreszenzreaktion auf das Distanzsignal und auf das im Messblatt
induzierte elektrische Signal zeigt damit eindeutig, dass bei einer Distanzsignalleitung die
elektrischen Signale über die Blattadern in die Intercostalfelder weitergeleitet werden,
während bei den basipetalen Kurzstreckensignalen die durch die Verwundung
hervorgerufenen Signale sowohl über das Mesophyll, als auch über die Blattadern mit der
selben Geschwindigkeit weitergeleitet zu werden scheinen.

Diskussion
78
4 Diskussion
4.1 Bedeutung der Calciumernährung für die Holzdifferenzierung
Obwohl die Bedeutung von Bäumen als Lieferanten eines der weltweit wichtigsten Rohstoffe,
dem Holz, immer stärker ins öffentliche Bewusstsein rückt, so war dennoch die cytologische
Erforschung des Cambiums in seiner Funktion als holzbildendes Meristem über lange Zeit
hinweg relativ unpopulär. Erst seit einigen Jahren, einhergehend mit technischen Fortschritten
in der Molekularbiologie, wird man sich bewusst, dass eine mögliche Modifizierung des
Holzwachstums auch von immensen wirtschaftlichen Vorteilen sein könnte. Pionierarbeiten
auf diesem Gebiet anhand der Modellbaumart Populus betrafen Untersuchungen zur
Ligninreduzierung sowie dessen leichtere Extrahierbarkeit. Lignine sind Zellwand-Polymere,
die in höheren Pflanzen für mechanische Festigkeiten, für den Wassertransport und für
Pathogenabwehr essentiell sind. Die Entfernung dieses Polymers aus den Holzfasern stellt den
entscheidenden Schritt bei der Zellstoff- und Papierherstellung dar. Wegen seiner
Wasserunlöslichkeit und der starken Vernetzung in den Fasern ist man bei der kostspieligen
Delignifizierung zudem auf den Einsatz von umweltschädigenden Chemikalien angewiesen.
Wissenschaftliche Untersuchungen stützten sich fortan vorwiegend auf genetische
Modifikationen der Ligninbiosynthese. Als problematisch stellt sich dabei nicht nur die
Ausbringung der genetisch veränderten Pflanzen unter ökologischen Gesichtspunkten, vor
allem Aspekte der Fertilität der Bäume betreffend, dar, sondern auch die eigentliche
Modifizierung an dem komplexen Polymer Lignin (Campbell und Sederoff, 1996; Lewis,
1999; Boerjan et al., 2003). Einhergehend mit einer immer tieferen Erkenntnis über die
Ligninbiosynthese wurde offenbar, um welch sensiblen und verzweigt arrangierten Vorgang
es sich dabei handelt. Heute ist man sich darüber einig, dass eine Vielzahl von Enzymen und
Substraten in enger Zusammenarbeit mit Regulatorproteinen nötig ist, um in einem
spezifischen Pflanzengewebe eines bestimmten Entwicklungsstadiums den erforderlichen
Lignintyp bereitzustellen (Ranocha et al., 2000).
Die in dieser Arbeit dargestellten Ergebnisse beruhen nicht auf genetischen Veränderungen
des untersuchten Organismus’, jedoch zeigen auch diese Pappeln Veränderungen in ihrem
Ligningehalt in den Holzzellwänden. So konnte durch FTIR-Spektroskopie gezeigt werden,
dass sich bei stark reduzierter Calcium-Ernährung eine deutliche Reduktion der Syringyl-
Einheiten ergab; da in Pappeln das G/S - Verhältnis recht stabil ist (Hu et al., 1999) kann
daraus geschlossen werden, dass sich die gesamte Ligninkonzentration in diesem Holzgewebe

Diskussion
79
im Vergleich zur Calcium-Optimalversorgung verändert. Die exakte Einflussnahme von
Calcium auf die Ligninbiosynthese ist bislang noch nicht vollständig bekannt, jedoch stimmen
die in der vorliegenden Arbeit gezeigten Ergebnisse mit Beobachtungen von Eklund und
Eliasson überein, die für die Ligninbiosynthese in jungen Bäumen eine notwendige
Versorgung mit Calcium feststellen konnten (Eklund und Eliasson, 1990). Eine
Erklärungsmöglichkeit für das Phänomen der Ligninreduktion unter Calciummangel gibt sich
auch aus Untersuchungen an apoplastischen Peroxidasen, welche über eine Ca2+-Pektat-
Bindungsstelle verfügen (Carpin et al., 2001). Nachdem die Mittellamelle und die
Zellwandecken die ersten Bereiche einer einsetzenden Lignifizierung darstellen und eben
diese Bereiche im Normalfall über eine hohe Ca2+-Pektat-Konzentration verfügen, ließe sich
daraus ableiten, dass diese Ca2+-Pektat - gebundenen Peroxidasen eine nicht zu
unterschätzende Auswirkung auf die räumliche Verteilung der Lignindeposition ausüben.
Änderungen in der apoplastischen Ca2+-Konzentration könnten damit Änderungen in der
Peroxidaselokalisation und dadurch in der Ligninsynthese nach sich ziehen (Carpin et al.,
2001).
Eine weitere Calciumabhängigkeit der Ligninbiosynthese wurde bereits in den 80er Jahren
von Westermark (1982) untersucht. In ihren Arbeiten hat sie in vitro einen möglichen
Oxidationsmechanismus für Coniferyl-Alkohol durch ein Superoxid-Radikal erforscht, eine
Reaktion, die bei einer apoplastischen Calcium-Konzentration von ≥ 50 mM gefördert wurde.
Mit abnehmender Ca2+-Versorgung ging auch eine Abnahme in der Reaktionsintensität
einher; unter Calcium-Abstinenz konnten nurmehr etwa 60% des Coniferyl-Alkohols mit
Superoxid reagieren. Durch diese Untersuchungen ergab sich auch eine zusätzliche
Erklärungsmöglichkeit für die erhöhten Calcium-Konzentrationen in den Wandbereichen
noch unlignifizierter Zellschichten verglichen mit bereits lignifizierten Zellwandschichten
(Wardrop, 1976); diese wären somit für die Bildung von Radikalen aus Coniferylalkohol
förderlich. Die im Rahmen dieser Arbeit vorliegenden Ergebnisse weisen ebenfalls auf eine
mit Rückgang der Ca2+-Versorgung rückgängige Lignin-Biosynthese hin; insofern lassen sich
durchaus Parallelen zu Westermarks Untersuchungen ziehen.
Neben calciumabhängigen Veränderungen des Ligningehaltes in den Zellwänden konnten die
FTIR - Untersuchungen auch noch Veränderungen anderer funktioneller Gruppen darlegen.
Besonders bemerkenswert erscheint hier ein unter Calcium-Mangel genereller Rückgang der
Absorptionsbanden in dem Wellenbereich 1300 cm-1 bis 900 cm-1, welcher für funktionelle
Gruppen der Carbohydrate steht (Williams und Fleming, 1980). Diese Wellenbereiche
gewinnen an Bedeutung, wenn man sie Ergebnissen einer Arbeitsgruppe gegenüberstellt, die

Diskussion
80
FTIR - Spektren an Zellsuspensionen der Karotte gemessen haben (McCann et al., 1993). In
dieser Arbeit gelang es den Autoren, gewichtige Veränderungen in der
Zellwandzusammensetzung während des Streckungswachstums der untersuchten Zellen
festzustellen. Demnach enthielten die Zellwände noch nicht gestreckter, runder Zellen
deutlich mehr Proteine, Ester und Phenole pro gemessenen Zellwandausschnitt als die
Zellwände bereits gestreckter Zellen, welche ihrerseits einen signifikant höheren Anteil an
Carbohydraten enthielten. In der vorliegenden Arbeit haben Untersuchungen zur
Xylemfaserlänge der Pappel in Abhängigkeit der Calciumernährung ergeben, dass mit
abnehmender Calcium-Versorgung ein Rückgang in der Faserlänge einhergeht. Ebenso
konnte mittels FTIR - Spektroskopie unter Calcium-Optimalversorgung ein im Vergleich zur
Ca2+-Abstinenz stark erhöhter Anteil an Carbohydraten festgestellt werden. Diese Korrelation
zwischen optimaler Calcium-Versorgung, dem Carbohydratanteil der Zellwand und der
Faserlänge ist demnach in einigen Punkten mit den Ergebnissen von McCann et al. an
Karottenzellen übereinstimmend und bekräftigt somit die Aussagen zum calciumabhängigen
Streckungswachstum der Faserzellen.
Dennoch darf bei allen Übereinstimmungen nicht außer Acht gelassen werden, aus welchem
Meristem die Faserzellen der Pappel entspringen und wieweit sie sich überhaupt longitudinal
entwickeln können. So liegt der erste große Unterschied in der Ausgangsform der Faserzellen
von Angiospermen; diese ist nach der Differenzierung aus den cambialen Mutterzellen bereits
lang gestreckt. Im Vergleich zu den oben genannten Karottenzellen tritt auch noch ein sog.
intrusives Spitzenwachstum auf. Dieses Längenwachstum der Libriformfasern kann wiederum
in verschiedenen Laubbaumarten unterschiedlich intensiv ausgeprägt sein (Tab. 4.1).

Diskussion
81
Baumart
Durchschnittslänge cambiale Initialen
(mm)
Durchschnittslänge Libriformfasern
(mm)
Acer rubrum 0,49 0,84
Alnus incana 0,6 1,89
Betula populifolia 0,94 1,31
Fraxinus americana 0,29 0,96
Populus sp. 0,49 0,9
Quercus alba 0,53 1,0
Robinia pseudoacacia 0,17 0,87
Tab. 4.1: Durchschnittliche Länge cambialer Initialzellen und Xylemfasern
ausgewählter Laubbaumarten; nach Bailey, 1920.
Neben einem baumartenspezifischen Faserlängenwachstum ist zudem noch eine
altersabhängige Veränderung in den jeweiligen Gewebeexpansionen festzustellen. Demnach
nimmt sowohl die Länge der cambialen Initialzellen, als auch die Länge der aus ihnen
hervorgehenden Xylemfaserzellen im juvenilen Holz (≤ 30 Jahre) mit steigendem Alter des
Baumes zu. Dieses Phänomen gilt für Gymnospermen genauso wie für Angiospermen, kann
jedoch wieder baumartenabhängig unterschiedlich stark ausgeprägt sein (Bailey und Shepard,
1915; Shimaji, 1950).
Neben der Zunahme der Faserlänge mit zunehmendem Baumalter lassen sich auch saisonale
Veränderungen des intrusiven Spitzenwachstums der Libriformfasern feststellen. Bissett und
Dadswell fanden bereits in den 50er Jahren heraus, dass in allen 28 von ihnen untersuchten
Laubbaumarten innerhalb eines Jahrrings deutliche Schwankungen der Faserlänge existieren
(Bissett und Dadswell, 1950): zwischen den ersten Fasern des Frühholzes und den
spätgebildeten des Spätholzes findet ein kontinuierlicher Anstieg in der Faserlänge statt; an
einer Spätholzgrenze zum nächsten Jahrring hin konnte ein steiler Abfall in der Faserlänge
gemessen werden.
Zieht man die altersabhängigen sowie die intrasaisonalen Abhängigkeiten des
Spitzenwachstums in Betracht, so lassen sich auch die relativ kurzen Faserlängen der in dieser
Arbeit vorgestellten mazerierten Pappelfasern erklären; die Proben stammten von einjährigen

Diskussion
82
Pappeln aus der Frühholzbildung und betrugen zwischen 0,35 mm und 0,6 mm. Trotz einer
möglichen biologischen Schwankung kann die calciumabhängige Reduzierung im
Spitzenwachstum aufgrund der hohen Stichprobenanzahl (n = 120) dennoch als signifikant
angesehen werden.
Im Gegensatz zu den Xylemfasern findet bei den Gefäßen der Angiospermen kaum ein
Spitzenwachstum statt; sie behalten demnach meist die Länge ihres Ursprungsmeristems, der
cambialen Initialzellen bei (Larson, 1994). Durch ihre perforierten stumpfen Enden können
Gefäße axial miteinander vernetzt sein und so Verbindungen bis zu mehreren Metern Länge
(z.B. Ringporer) bilden. Aufgrund ihrer Funktion des Wassertransports ist die radiale
Ausprägung der Gefäße von großer Bedeutung für den Baum. Eine Vergrößerung des
Gefäßdurchmessers vergrößert die Effizienz des Wassertransportes infolge eines rückläufigen
Wasserwiderstandes deutlich, vermindert jedoch gleichzeitig den Selbstschutz des Systems
vor z.B. Folgen von Kavitationen (Aloni, 1987). In der vorliegenden Arbeit kann bei dem
Gefäßweitenwachstum ein calciumabhängiger Trend von Gefäßen mit großem Durchmesser
bei Optimalversorgung hin zu Gefäßen mit reduziertem Durchmesser bei Minimalversorgung
festgestellt werden. Diese Beobachtung ließe sich erklären, wenn man die Auswirkungen
einer ebenfalls unter Calcium-Mangel rückläufigen Blattflächenentwicklung in Betracht zieht.
Wie sowohl Biomasseuntersuchungen der in Hydrokultur angezogenen Pappeln, als auch
Blattflächenmessungen im Freiland angezogener Pappeln zweifelsfrei zeigen konnten, sank
die Biomasseproduktion und damit auch die Blattflächenbildung mit abnehmender Calcium-
Versorgung im Nährmedium, bzw. Bodensubstrat. Parallel zur reduzierten Blattfläche sinkt
damit auch die Transpirationsrate der einzelnen Probebäume, womit auch die Notwendigkeit
für einen maximalen Gefäßdurchmesser nicht mehr gegeben ist.
Eine weitere Möglichkeit für das unter Calcium-Mangel reduzierte Gefäßweitenwachstum
könnte in einer allgemeinen Stressreaktion der Pappel auf Nährstoffmangel liegen. In der
vorliegenden Dissertation konnte anhand von ultrastrukturellen Untersuchungen gezeigt
werden, dass die cambiale Zone unter Calcium-Mangel in ihrer radialen Ausprägung
erheblich rückläufig ist; anstelle der ca. acht Zellreihen der Kontrolle war die cambiale Zone
auf etwa drei Zellreihen beschränkt. Das Cytoplasma wies auch nicht die typische starke
Vakuolisierung eines aktiven Cambiums auf, sondern zeigte ein sehr dichtes Cytoplasma mit
reduzierten Vakuolen. Mit diesem Erscheinungsbild ähnelten sie sehr einem typischen
angiospermen Cambium während der Winterruhe (Farrar und Evert, 1996; Krabel, 2000;

Diskussion
83
Arend und Fromm, 2003). Da Calciumstress auf das cambiale Meristem einen ebenso
gearteten Einfluss auszuüben scheint, könnte die beobachtete Gefäßflächenreduzierung auch
auf ein nahezu inaktives Cambium zurückzuführen sein. Einen weiteren, diese Theorie
unterstützenden Hinweis liefern die elektronenmikroskopischen Beobachtungen der
Chloroplasten. Auch hier ergaben sich im Vergleich zu der Kontrolle deutliche Unterschiede
in der Ultrastruktur. In der Kontrolle deutlich sichtbare Stärkeeinlagerungen, wie sie bereits
vielfach als chloroplastentypisch und ~notwendig charakterisiert wurden, fehlen in den unter
Calcium-Mangel gewachsenen Pappelblättern nahezu vollständig. Dieser Mangel an Stärke in
den Chloroplasten könnte ebenfalls auf eine Reaktion der Pappel auf Calciumstress hindeuten,
die in ihrer Ausprägung der Vorbereitung zur Seneszenz sehr ähnelt. Die zur Vorbereitung auf
den Laubfall typischen Vorgänge im Blatt beinhalten unter anderem eine Translokation
mobilisierbarer Nährelemente in den Stamm, in dessen Zug auch Stärke abgebaut wird (Mohr
und Schopfer, 1992). Ein Abbau von Chlorophyll, wie er ebenfalls bei der Blattalterung
auftritt, konnte bei den durchgeführten Versuchen jedoch nicht festgestellt werden. Wie in der
Arbeit gezeigt werden konnte, wurden in vergleichenden Gaswechselmessungen unter
Calcium-Mangel keine Unterschiede in der Photosyntheseleistung gemessen.
Als dritter Aspekt für die Tendenz eines rückläufigen Gefäßdurchmessers unter Calcium-
Mangel wäre die mit Calcium-Mangel einhergehende Reduktion der Phloembe- und
~entladung in Betracht zu ziehen. Wie mikroautoradiographische Untersuchungen an 14C-
markierten Photoassimilaten bereits zeigen konnten, fand bei unter Calcium-Abstinenz
angezogenen Pappeln eine drastisch reduzierte Phloembeladung in den Blättern und keine
Entladung von Photoassimilaten aus dem Phloem des Stamms in das Cambium statt (Schulte-
Baukloh und Fromm, 1993). Da dieser Effekt bei vergleichenden Untersuchungen unter
Kalium-, bzw. Magnesium-Mangel nicht gezeigt werden konnte, kann davon ausgegangen
werden, dass die Phloementladung im Stamm möglicherweise calciumspezifisch reguliert
wird. Da die cambialen Zellen unter Calcium-Mangel demnach nicht mehr mit Kohlenstoff
versorgt werden, könnte die Tendenz zu den geringer ausgeprägten Gefäßdurchmessern auf
den C-Mangel zurückgeführt werden. Ein weiteres Argument, das für eine Funktion des
Calciums bei der Phloementladung spricht sind die in dieser Dissertation dargestellten
Calciumgehalte in den verschiedenen Geweben des Stamms. Wie mittels energiedispersiver
Röntgenanalyse gezeigt werden konnte, steigt der Gehalt an Calcium im Phloem, im
Cambium und in der Differenzierungszone des Xylems mit steigender Calcium-Versorgung in
der Nährlösung ebenfalls an. In allen drei bemessenen Geweben bleibt die Ionenkonzentration

Diskussion
84
pro Ernährungsvariante dabei etwa gleich stark ausgeprägt, lediglich bei einem Überangebot
von Calcium steigt die Konzentration im Phloem überproportional an, was wiederum auf eine
phloemspezifische Funktion des Calciums hinweist. Da Calcium relativ Phloem-immobil ist,
muss es sich dabei um eine apoplastische Verlagerung über den Transpirationsstrom handeln.
Eine ähnliche Situation mit erhöhten Calcium-Vorkommen im Phloem lässt sich auch
während der „Prä-Aktivierungs-Phase“ des Cambiums im Frühjahr feststellen. Wie Follet-
Gueye et al. anhand von Untersuchungen an der Buche mittels sekundärer Ionen-
Massenspektronomie (SIMS) zeigen konnten, sind die Calciumgehalte insbesondere im
Phloem zu dieser Zeit außerordentlich erhöht (Follet-Gueye et al., 1998). Da diese
Beobachtungen jedoch vor Einsetzen des Transpirationsstroms gemacht wurden, wird das
benötigte Calcium in diesem Fall entweder aus in der Rinde lokalisierten Oxalatkristallen
oder durch basipetale Diffusion aus den calciumreichen Apikalmeristemen geliefert,
einhergehend mit der ebenfalls basipetal verlaufenden auxingesteuerten cambialen
Reaktivierung (Lachaud und Bonnemain, 1982).
4.1.1 Ausblick auf die Bestandesebene
Die Effektivität, mit der ein Baum auf die unterschiedlichsten auf ihn einwirkenden Kräfte
reagieren kann, ist die im Bestandesalltag unserer Waldbäume eigentlich relevante Größe. In
diesem Zusammenhang kann Calcium-Mangel die mechanischen Fähigkeiten von Holz
negativ beeinflussen; dies kann sich in einer reduzierten Zugfestigkeit, einer reduzierten
Bruchresistent sowie eines reduzierten Elastizitätsmoduls äußern (Brady, 1969; Panshin und
DeZeeuw, 1980; McLaughlin und Wimmer, 1999).
Über die rein mechanischen Auswirkungen auf den Rohstoff hinaus kann sich Calcium-
Mangel natürlich auch auf das Stützgewebe des biologischen Systems der Pflanze auswirken.
Das kann zum einen eine Lockerung des Zellverbandes nach sich ziehen, wie es bereits 1929
von Albrecht und Davis am Spross der Sojabohne beobachtet wurde und wie es in der
vorliegenden Arbeit auch an elektronenmikroskopischen Aufnahmen für Wurzel- und
Blattgewebe bestätigt werden konnte. Zum anderen kann Calcium-Mangel auch, wie an der
Kiefer gezeigt werden konnte, zu einer erhöhten Brüchigkeit des Stamms führen (Davies,
1949).
Eine geringe Calcium-Verfügbarkeit in stark sauren Böden kann als mögliche Konsequenz
einen Rückgang in der Lignifizierung des Holzgewebes zur Folge haben (Eklund und

Diskussion
85
Eliasson, 1990). Dieser Rückgang wäre aufgrund der ausgeprägten Bindungsmöglichkeiten
entlang des Transpirationsstroms im apoplastischen Raum vor allem in den Kronenbereichen
älterer Bestände vorstellbar, wenn man davon ausgeht, dass nur noch ein Bruchteil der im
Wurzelraum aufgenommenen Ionen den Kronenraum erreicht. Gerade bei
Nadelholzbeständen wäre im Kronenraum eine ausreichende Calciumversorgung jedoch
unbedingt nötig, da Lignin dort eine stützende Funktion bei der Astbildung innehat
(Druckholzbildung) und somit stark zur Stabilität des Baumes beiträgt (McLaughlin und
Wimmer, 1999). Im Stammbereich könnte Calcium-Mangel zu einem weniger stabilen und
damit brüchigen Xylem führen und damit zu einer höheren Anfälligkeit, mechanischen
Belastungen wie Wind oder Schnee nicht mehr standhalten zu können (McLaughlin und
Wimmer, 1999).
4.2 Signalleitung in der Pappel
Die Entwicklung molekularbiologischer Techniken ermöglichte eine über die deskriptive
Ebene hinausgehende Erforschung der elektrischen Signalleitung, insbesondere ihrer Rolle
bei inter- sowie in intrazellulären Vorgängen. Besonders die Entwicklung von Calcium-
spezifischen Farbstoffen hat es erlaubt, räumliche und zeitliche Abläufe bei der Signalleitung
aufzudecken (Webb et al., 1996). Dadurch wurde der Weg geebnet, den Prozess der
Signalentstehung, der Signalausbreitung und den pflanzenphysiologischen Reaktionen darauf
zu entschlüsseln (Trewavas und Malho, 1997; McAnish und Hetherington, 1998; Snedden
und Fromm, 1998; Plieth et al., 1998). Anfang der 80er Jahre des letzten Jahrhunderts gelang
es beispielsweise Davies und Schuster (1981) an der Erbse, Auswirkungen von elektrischen
Signalen auf die Genexpression nachzuweisen. 1992 veröffentlichten Wildon et al. einen
wegweisenden Artikel, der die Aktivierung von Proteinaseinhibitor (PIN) - Genen durch
Hitzereiz - induzierte elektrische Signale beschrieb.
Wenn man die Reaktion einer Pflanze auf diverse Umwelteinflüsse betrachtet kann man
erkennen, dass auch unter ökophysiologischen Gesichtspunkten elektrischen Signalen eine
Schlüsselrolle zukommt. Ihr Aktionsradius erstreckt sich dabei von einzelnen Zellen bis hin
zu verschiedenen Organen der Pflanze (Volkov und Mwesigwa, 2001). So konnte
beispielsweise anhand von Untersuchungen an der Sojabohne festgestellt werden, dass 2,4-
Dinitrophenol (DNP), welches in seinen Auswirkungen auf Mensch und Umwelt als durchaus

Diskussion
86
bedenklich eingestuft wird, in der Pflanze verstärkt Aktionspotentiale hervorruft und
gleichzeitig einen Rückgang im Variationspotential bewirkt (Mwesigwa et al., 2000).
Gleichermaßen wurde für Pentachlorphenol, ein weiteres Umweltgift, das die oxidative
Phosphorylierung entkoppelt, eine Induktion von Aktionspotentialen ebenfalls an der
Sojabohne nachgewiesen (Volkov et al., 2000).
Es sind sicherlich noch nicht annähernd alle einzelnen Auswirkungen von elektrischen
Signalen auf die unterschiedlichen Aspekte der Pflanzenphysiologie erforscht. So konnten
beispielsweise erst in jüngerer Vergangenheit Forschungsergebnisse zur Signalleitung an der
sensitiven Pflanze Mimosa pudica eine Einflussnahme der elektrischen Signale auf die
Photosyntheseleistung aufzeigen (Koziolek et al., 2004).
Die hier vorgestellte Arbeit bestätigt diese Erkenntnisse nun auch für die Gattung Populus;
darüber hinaus unterscheidet sie noch zwischen verschiedenen Arten und Positionen der
Signalinduktion und den dadurch hervorgerufenen Reaktionen in der Photosyntheseleistung.
Dabei wurde zwischen Veränderungen in der Lichtreaktion der Photosynthese, welche mittels
zweidimensionaler Bildanalyse der Chlorophyllfluoreszenz gemessen wurde, und
Veränderungen in der Dunkelreaktion der Photosynthese, welche durch Schwankungen von
JCO2 im Blattgaswechsel detektiert wurden, unterschieden.
Die elektrischen Signale, die in der vorliegenden Arbeit dargestellt sind, wurden überwiegend
mittels der sog. Aphidentechnik in Phloemzellen gemessen. Dieser Zelltyp stimmt in vielen
grundlegenden Eigenschaften mit Nervenzellen des tierischen Systems überein, wie z. B. der
elektrischen Erregbarkeit der Zellmembranen, durch die elektrische Signale von Zelle zu
Zelle weitergegeben werden können. Wenngleich es in tierischen Nervenzellen auch
Natrium- und Kaliumströme sind, die für die Entstehung eines Aktionspotentials
verantwortlich sind, so basieren die Mechanismen der Erregbarkeit in pflanzlichen
Zellemembranen auf einem Calciumeinstrom und auf einem Chlorid- und Kaliumausstrom
(Beilby und Coster, 1979; Kikuyama und Tazawa, 1983; Lunevsky et al., 1983; Tsutsui et al.,
1986; Fromm und Spanswick, 1993).
In der Pappel konnte nun bei den Veränderungen des Membranpotentials gezeigt werden, dass
die Art der Spannungsänderung des fortlaufenden Signals von der Richtung abhängig ist, in
der es sich fortbewegt. Sowohl bei Hitze- als auch bei Kältereiz zeigten basipetal laufende
Signale durch negative Spannungsänderungen an der Plasmamembran eine Hyperpolarisation

Diskussion
87
der Membranpotentiale, wohingegen sich acropetal fortpflanzende Signale durch eine
kurzzeitige Depolarisierung der Membran charakterisieren ließen.
Der Ionenmechanismus, der hinter einer Depolarisation der Membran steht, basiert auf einem
Calcium-Einstrom in das Cytoplasma, welcher einen Chlorid-Ausstrom in den Apoplasten
induziert, dem wiederum ein Kalium-Ausstrom folgt (Beilby und Coster, 1979; Kikuyama
und Tazawa, 1983; Kikuyama, 1987; Lunevsky et al., 1983; Tsutsui et al., 1986; Fromm und
Spanswick, 1993). Weniger genau definiert ist jedoch die Entstehung der Hyperpolarisation
der Zellmembran. Um die bei der basipetalen Signalleitung beobachtete Hyperpolarisation auf
ihre Kalium-Kanal - Abhängigkeit zu überprüfen, wurden daher die Kalium-Kanäle in den
Pappeltrieben mit Tetraethylammoniumchlorid (TEA+), einem K+-Kanal - spezifischen
Hemmstoff, geblockt. Da TEA+ in den meisten Fällen apoplastisch appliziert wird, hemmt
dieser Wirkstoff in den meisten Untersuchungen ausschließlich die einwärtsgerichteten
Kalium-Kanäle (z.B. Langer et al., 2002). Untersuchungen, bei denen TEA+ intrazellular
injiziert wurde konnten jedoch belegen, dass es im intrazellularem Raum auch
auswärtsgerichtete Kalium-Kanäle blockt (Wong und Adler, 1986; Ochi und Nishiye, 1974);
TEA+ wirkt demnach generell auf alle K+-Kanäle, die von der jeweiligen Position
transmembran gerichtet sind. In der vorliegenden Arbeit wurden abgetrennte Pappeltriebe für
einen Tag in eine mit TEA+ versetzte Nährlösung (APW + TEA+) gegeben. Der Hemmstoff
konnte sich somit nicht nur über den Transpirationsstrom im apoplastischen Raum
ausbreiten, sondern auch über die Schnittstelle in den intrazellularen Raum des
Phloemgewebes eindringen. Auf diese Weise konnten die K+-Kanäle auf beiden Seiten der
Membran geblockt werden. Ein sich durch einen Hitzereiz an der Blattspitze basipetal
ausbreitendes elektrisches Signal konnte nach einer TEA+-Behandlung nicht mehr detektiert
werden (Abb. 3.31). Dieses Ergebnis deutet darauf hin, dass auch bei einer Hyperpolarisation
der Plasmamembran das sich ausbreitende Signal über einen K+- Efflux aus den Phloemzellen
verursacht wird, insbesondere wenn weitere Arbeiten zum Mechanismus der
Hyperpolarisation herangezogen werden. So konnten Schönknecht et al. an der einzelligen
Grünalge Eremosphaera viridis eine vorübergehende Hyperpolarisation feststellen, nachdem
die photosynthetische Elektronenaufnahme durch kurzzeitiges Abdunkeln unterbrochen
wurde (Schönknecht et al., 1998). Mittels ionenspezifischer Farbstoffe gelang es den Autoren
nachzuweisen, dass die Hyperpolarisation durch einen intrazellulären Calcium-Anstieg
induziert wurde, welcher seinerseits einen K+-Efflux bewirkte. Dass dieser
Hyperpolarisationsmechanismus im Pflanzenreich jedoch nicht der einzige ist, konnten
jüngste Untersuchungen an einem Phytoplankton zeigen, bei welchem durch einen Chlorid-

Diskussion
88
Einstrom eine Hyperpolarisation ausgelöst wird (Taylor und Brownlee, 2003). An höheren
Pflanzen wurde bislang jedoch der Calcium-Kalium -Mechanismus bestätigt.
Die vorliegenden Ergebnisse an unter Calcium-Minimalversorgung angezogenen Pappeln
zeigen bei einer Hitzereizung der Blattspitze, dass die Erregbarkeit der Phloemzellen bei
einem basipetal laufenden elektrischen Signal gänzlich unterdrückt wurde. Daraus lässt sich
zum einen schließen, dass bei einer basipetalen Signalleitung Calcium-Kanäle involviert sind;
ein Schluss, der die bisher bekannte Funktion von Calcium-Kanälen und -Transportern als
Angelpunkt für ein vielseitiges Netzwerk intrazellularer Signalverarbeitung unterstützen
würde (Trewavas und Malho, 1997; Trewavas, 2000). Zum anderen lässt sich unter
Berücksichtigung der TEA+ geblockten K+-Kanäle schließen, dass unter optimaler
Nährstoffversorgung der Pappel ein K+-Efflux für das Entstehen der Hyperpolarisation
entlang der Phloemzellen verantwortlich ist. Dieser verläuft mit hoher Wahrscheinlichkeit
auch in Bäumen nach dem aus Eremosphaera bekannten Prinzip der Calcium-Induktion
(Schönknecht et al., 1998).
Die vorliegenden Ergebnisse zur Distanzsignalleitung erlauben eine weitere Unterscheidung
zwischen basipetal und acropetal gerichteten Signalen. Die Phloemmessungen an unter
Calcium-Mangel angezogenen Pappeln haben gezeigt, dass unter identischen Bedingungen
das in acropetaler Richtung laufende Signal trotz erheblichen Calcium-Mangels noch leicht
ausgeprägt war, wohingegen in basipetaler Richtung keine Amplitude mehr festgestellt
werden konnte (Abb. 3.32). Diese Beobachtung deutet auf eine Präferenz der Signalleitung
hin, die ihre Ursprünge in der Funktion der Informationsweiterleitung vom Wurzelsystem /
Stamm zum Kronenraum haben könnte. Anhand der Wasserpflanze Azolla konnte ein
ähnliches Phänomen festgestellt werden (Overall und Gunning, 1982); elektrophysiologische
Messungen in den Wurzeln haben auch hier gezeigt, dass über die Plasmodesmata die
elektrische Kopplung zwischen zwei Zellen in acropetaler Richtung deutlich ausgeprägter war
als in basipetaler. Die Signalweiterleitung im Phloem der Pappel findet durch die in diesem
Gewebetyp häufig auftretenden Siebporen von Siebelement zu Siebelement statt. Da die recht
großen Siebporen (bis zu 1 µm Durchmesser) an den Querwänden der Siebelemente bei der
Phloemdifferenzierung aus Plasmodesmata hervorgehen, scheinen sie bei der elektrischen
Signalleitung eine entscheidende Rolle zu spielen.
In dieser Arbeit wurden an der Pappel elektrische Signale durch zweierlei Reize induziert:
zum einen durch Kältereizung und zum anderen durch Hitzereizung. Die durch Kälte
induzierten, acropetal fortgeleiteten Signale wiesen in ihrer Geschwindigkeit ebenso wie in

Diskussion
89
ihrer Amplitude die für pflanzliche Aktionspotentiale typischen Merkmale auf und stimmen
in diesen Punkten mit bereits bekannten Reaktionen in Mais (Fromm und Fei, 1998) und in
Weide (Fromm und Spanswick, 1993) überein. Generell gilt, dass Aktionspotentiale sich mit
einer konstanten Geschwindigkeit fortbewegen. Bisherige Erkenntnisse lassen darauf
schließen, dass sie in ihrer Ausbreitung von spannungsabhängigen Ionenkanälen abhängig
sind und sich entlang der Plasmamembran durch Plasmodesmata lebender Zellen oder durch
Siebporen der Siebröhren fortpflanzen. Im Gegensatz dazu lösen Reizungen durch
Gewebeverwundung, insbesondere durch Hitzereizung, sich langsamer ausbreitende und
unregelmäßig verlaufende Variationspotentiale aus (Sibaoka, 1966; Sibaoka, 1969; Pickard,
1973). Sie galten bislang nicht als sich selbständig fortpflanzende Signale, sondern vielmehr
als lokale elektrische Reaktionen, die auf einen chemischen Reiz erfolgen, welcher wiederum
am Ort der Verletzung entsteht und sich hydraulisch über das Xylem fortpflanzt. Tinz-
Füchtmeier und Gradmann führten zur hydraulischen Signalleitung mittels Elektrometer und
Interferometer Messungen an der Mimose durch, welche dieser Theorie der hydraulischen
Signalleitung nach einer Hitzereizung jedoch entgegenstehen (Tinz-Füchtmeier und
Gradmann, 1990).
Grundsätzlich ist es durchaus möglich, dass hydraulische Signale bei der Erregbarkeit der
Pappel eine Rolle spielen. Nachdem die Messungen zu den hier vorgestellten Daten jedoch im
Phloem stattfanden und es dort gelang, die durch einen Hitzereiz induzierten Signale zu
messen, stellte sich die Frage, ob Druck- oder chemische Veränderungen im Xylem
Ionenkanäle des Phloems aktivieren können, und zwar in dem Ausmaß, dass daraufhin in
diesem Gewebe die Variationspotentiale hervorgerufen werden könnten. Zur Klärung dieser
Frage wurden Kontrollmessungen durchgeführt, die deutlich zeigen, dass durch das Anlegen
eines Kälteblocks am Spross sowohl die charakteristische Signalleitung unterbrochen ist, als
auch dass die Reaktion im Gaswechsel unterbleibt. Es erwies sich als ausschlaggebend, das
elektrische Signal von einem hydraulischen mittels eines Kälteblocks unterscheiden zu
können. Da auf das Anbringen eines Kälteblock hin keine Veränderung in der CO2-
Austauschrate oder in der stomatären Leitfähigkeit im Gaswechsel festzustellen war, konnte
das Distanzsignal eindeutig als elektrisches bestätigt werden, da es den gekühlten
Sprossabschnitt offensichtlich nicht passieren konnte. Für ein hydraulisches Signal würde ein
Kälteblock kein Hindernis darstellen, da die Transduktion über die toten Zellen des Xylems
hinweg erfolgen würde.

Diskussion
90
Somit konnte bei den an Pappel durchgeführten Messungen ausgeschlossen werden, dass bei
der Signalweiterleitung in acropetaler Richtung hydraulische Signale einen Einfluss auf den
Gaswechsel haben. Zudem gelten hydraulische Signale als stark abhängig vom
Bewässerungsstatus der Messpflanze. Dabei gilt, dass bei einem wassergesättigten Zustand
des Sprosses, wie er bei den in dieser Arbeit untersuchten Pappeln vorlag, die
Xylemspannung vernachlässigbar gering ist und damit hydraulische Signale sich kaum
ausbreiten könnten. Diese Punkte sprechen dafür, dass die hier gemessenen Wund-Signale im
Pappel-Phloem sich eigenständig fortbewegen und nicht von sich im Xylem fortpflanzenden
hydraulischen Signalen beeinflusst sind.
Es ist nicht auszuschließen, dass auch chemische Signale einen Einfluss beispielsweise auf die
Photosynthese haben können. Die Transportgeschwindigkeit für chemische Signale in
Pflanzen liegt jedoch bei Werten zwischen 50 und 100 cm h-1 (Canny, 1975) und erweist sich
damit als bei weitem zu langsam, um an den Reaktionen der vorliegenden Messungen
beteiligt zu sein.
Im Hinblick auf eine Beeinflussung der photosynthetischen Primärprozesse durch elektrische
Signale konnte die nicht-invasive Bildanalyse der Chlorophyllfluoreszenz zeigen, dass
innerhalb eines Blattes Kurzstreckensignale, welche durch einen Hitzereiz an der Blattspitze
hervorgerufen wurden und sich somit basipetal ausbreiteten, in den Blattadern und in den
Intercostalfeldern einen simultanen, deutlichen Rückgang der Quantenausbeute im
Photosystem II hervorrufen. Im Gegensatz dazu konnte bei Distanzsignalen, an einem unteren
Blatt hervorgerufen und sich somit acropetal zum Messblatt fortpflanzend, eine eindeutige
zeitliche Verzögerung im Rückgang der Quantenausbeute des PSII zwischen sekundären
Blattadern und den sich direkt anschließenden Intercostalfeldern beobachtet werden. Diese
Tatsache lässt darauf schließen, dass sich das Distanzsignal über das Phloem des Stammes
und der Blattadern, welche zuerst reagieren, in das Mesophyllgewebe des Blattes ausbreitet.
Diese Beobachtung unterstützt die mittels Aphidentechnik gemessene Signaltransduktion, die
ebenfalls eine Ausbreitung über das Phloem charakterisierte.
Bei der photosynthetischen Reaktion auf ein elektrisches Signal ist zu vermuten, dass dabei
Veränderungen der Ionenströme innerhalb der Chloroplasten eine beachtliche Rolle spielen
könnten. Für weiterführende Forschungen könnte es also als erfolgversprechend gelten, wenn
ein Hauptaugenmerk auf die Ionentranslokation in den Mesophyllzellen und ihren
Chloroplasten gelegt werden würde. Hinweise auf eine veränderte Enzymtätigkeit als
Reaktion auf signalverursachende Änderungen der Ionenkonzentrationen in der Zellwand, in

Diskussion
91
den Plasmodesmata sowie im Cytoplasma liegen bereits vor (Davies, 1987). Bulychev et al.
(1986, 1987) berichteten bereits in den 80er Jahren von Änderungen der
Chlorophyllfluoreszenz in intakten Chloroplasten als Reaktion auf ein verändertes
Membranpotential. Aus diesen Gründen scheint es wahrscheinlich zu sein, dass der schnellen
Reaktion in der CO2-Aufnahmerate und nachfolgend in der Chlorophyllfluoreszenz in den
Pappelblättern lokale Schwankungen der Ionenkonzentration zugrunde liegen.
Für die Reduktion der Quantenausbeute am PS II wären dabei jedoch Ionenkonzentrations-
änderungen über die Thylakoidmembran hinweg entscheidend. Zieht man dabei die
Membranständigkeit der Chloroplasten in Betracht, so scheint es durchaus möglich zu sein,
dass so deutliche Spannungsänderungen an der Plasmamembran, wie sie bei einem
elektrischen Signal auftreten können, auf die direkt benachbarten Chloroplasten und auch auf
deren Thylakoidmembranen übergreifen könnten.
Anhand von Messungen an isolierten Thylakoidmembranen von Spinat gelang es bereits, eine
direkte Calcium-Abhängigkeit der Photolyse im Photosystem II nachzuweisen (Vrettos et al.,
2001); die Autoren legten dabei die dringend notwendige Präsenz von Calcium für diesen
Vorgang dar. Durch Störungen des intrastromatären Calcium-Gehalts aufgrund von
eintreffenden und auf die Chloroplasten- und Thylakoidmembran überspringenden
elektrischen Signalen, wäre es denkbar, dass die O2 - Bildung aufgrund lokaler Calcium-
Flüsse kurzfristig unterbrochen ist. Dadurch würden der Elektronentransportkette des PS II
die bei der Photolyse freiwerdenden Elektronen fehlen, wodurch die Vorgänge in den
Primärprozessen der Photosynthese gehemmt werden würden. Im intakten System der
Pappelblätter muss dieser Möglichkeit jedoch eine untergeordnete Rolle zugewiesen werden,
da trotz einer Unterbrechung der Photolyse die Elektronen weiterhin auf das PS II eintreffen
würden und in dem Fall, dass sie nicht abtransportiert werden könnten, am Chlorophyll
Schäden anrichten würden. In den vorliegenden Ergebnissen konnte jedoch keine Schädigung
des Chlorophylls beobachtet werden, da es sich um reversible Reaktionen am PS II handelte.
Die ersten Veränderungen in der CO2-Aufnahmerate, und damit in der Dunkelreaktion der
Photosynthese, erfolgten bei einer Verwundung des Blattgewebes an der 3 bis 4 cm zur
Meßküvette entfernten Blattspitze bereits 30 s nach Feuerreizung. Als bemerkenswertes
Phänomen ist im Gaswechsel zu beobachten, dass die stomatäre Leitfähigkeit sich trotz des
transienten Rückgangs in JCO2 nicht verändert. Entgegen der an Mimose beobachteten
Reaktion, dass auf ein elektrisches Signal hin die stomatäre Leitfähigkeit einhergehend mit
dem Abfall in JCO2 rückläufig war (Koziolek et al., 2004), blieb sie in der Pappel trotz

Diskussion
92
ebenfalls rückläufigen JCO2 in etwa auf ihrem Ursprungsniveau. Der Grund, weshalb trotz
steigendem ci-Werte die Stomata nicht schlossen, konnte im Rahmen dieser Arbeit nicht
untersucht werden. Aufgrund der generell hohen Öffnungs- und Schließgeschwindigkeit der
Pappel im Allgemeinen und des verwendeten Klons T89 im besonderen (Langer et al., 2003)
kann eine Trägheit des Schließzellenapparates als Ursache jedoch ausgeschlossen werden.
Nach Nobel (1991) sind die in wässriger Lösung vorkommenden Konzentrationen von CO2
temperaturabhängig und pH-sensitiv. Die Temperaturabhängigkeit bestimmt dabei vor allem
die Unterschiede der CO2-Konzentration zwischen der wässrigen Lösung und der
angrenzenden Gasphase, die Gleichgewichtskonzentration zwischen Bicarbonat und gelöstem
CO2 ist hingegen vorwiegend durch den pH-Wert beeinflusst. In der wässrigen Lösung
reagiert CO2 mit OH- und bildet so Bicarbonat, welches wiederum durch das Anhängen eines
Protons einen Kohlensäure-Komplex bilden kann. Die Umwandlung von CO2 zu H2CO3 wird
durch die Aktivität des Enzyms Carboanhydrase deutlich beschleunigt; da in diesem Prozess
Protonen involviert sind, gilt der pH-Wert als bedeutender Faktor der Katalysation (Nobel,
1991). Da die Gleichgewichtskonzentration für Kohlensäure nur etwa 1/400 des gelösten
Kohlendioxids entspricht, liegt bei diesem Vorgang der Schwerpunkt auf der Umwandlung
von CO2 zu Bicarbonat. Der Anstieg des ci-Wertes könnte nun aus Veränderungen des pH-Wertes im Cytoplasma
resultieren. Bei einem durch Hitze induziertem Signal, das wie bereits mehrfach erwähnt, in
seiner Repolarisation sehr träge ist und dadurch ein zeitlich lang anhaltendes Spitzenpotential
vorherrscht, welches seinerseits wiederum mit einem ansteigenden [Ca2+]cyt - Gehalt
einhergeht, könnte sich der intrazellulare pH-Wert deutlich ändern. Bush berichtete bereits
1995 von einer Vielzahl abiotischer und biotischer Stimuli, welche Veränderungen des
cytoplasmatischen Calciumgehaltes bewirkten und dadurch eine Veränderung des
cytoplasmatischen pH-Wertes nach sich zogen (Bush, 1995). Zieht man nun in Betracht, dass
die Aktivität des Enzyms Carboanhydrase stark pH-abhängig ist, könnte man daraus
schlussfolgern, dass die Enzymaktivität durch einen veränderten [Ca2+]cyt - Wert gehemmt
wird. Eine Hemmung der Carboanhydrase hätte zur Folge, dass an den Chloroplasten keine
Rückumwandlung von Bicarbonat in CO2 stattfinden kann. Geht man nun davon aus, dass die
Aufnahmefähigkeit von Bicarbonat im Cytoplasma begrenzt ist, würde sich demnach die
CO2-Konzentration in der Atemhöhle gleichfalls „zurückstauen“, was den Rückgang von JCO2
und damit auch den parallel dazu erhöhten ci-Wert erklären würde.

Diskussion
93
Der Rückgang der Quantenausbeute im PSII als Reaktion auf elektrische Signale ist bereits an
Mimose (Koziolek et al., 2004) und als Reaktion auf diverse abiotische Reize, wie z.B. das
Anlegen eines elektrischen Stroms, an Tomate (Herde et al., 1999) beobachtet worden; bei
den Untersuchungen an Tomate wurde dieser Rückgang jedoch erst nach fünf Stunden
gemessen. Dies steht im deutlichen Widerspruch zu den hier vorliegenden Beobachtungen
eines transienten Rückgangs im PS II, wie auch im JCO2 der Pappelblätter. Auch konnten die
Autoren bei Untersuchungen an der Tomate keinen zeitlichen Unterschied zwischen
Veränderungen im JCO2 und PS II feststellen; die Reaktionen, die mittels
Gaswechselmessungen ebenfalls einen Rückgang in JCO2 zeigten, wurden als zeitlich
übereinstimmend mit den Reaktionen im PS II erkannt.
In den hier dargestellten Ergebnissen an der Pappel wiesen die Photosynthesereaktionen
jedoch in ihrer Kinetik bemerkenswerte zeitliche Differenzen zwischen der Lichtreaktion und
der Dunkelreaktion auf. Im Vergleich zwischen den ersten auftretenden Reaktionen an JCO2
im Gaswechsel und denen der Chlorophyllfluoreszenz ist eindeutig zu beobachten, dass der
Rückgang in JCO2 um mindestens 120 Sekunden schneller einsetzte als der Rückgang der
Quantenausbeute im PS II. Diese Beobachtung konnte sowohl für basipetal gerichtete
Kurzstreckensignale, als auch für acropetal gerichtete Distanzsignale gezeigt werden. Beide
Systeme haben jedoch gemeinsam, dass sie nur für kurze Zeit in ihrem jeweiligen Minimum
verweilen, bevor sie sich wieder in Richtung auf ihren Ausgangswert erholen.
Die genauen Mechanismen, die bei dem Rückgang der Quantenausbeute im PS II über ein
elektrisches Signal ausgelöst werden, müssen erst noch aufgedeckt werden. Wenn man den
oben beschrittenen Gedankengang der Carboanhydrase-Blockierung weiterverfolgt, kommt
man zu der Schlussfolgerung, dass für das Enzym Rubisco im Stroma der Chloroplasten kein
CO2 mehr als Substrat zur Verfügung steht. Daraus folgt, dass im Calvin-Zyklus kein ADP
mehr frei wird. Durch ein Fehlen von ADP wiederum kann an der Thylakoidmembran der
sich ständig aufbauende Protonengradient nicht mehr abgebaut werden, es kommt zu einem
Anstieg des Protonengradienten. Als Reaktion auf einen daraus resultierenden niedrigen pH-
Wert im Thylakoidinnenraum tritt eine Veränderung im Xanthophyllzyklus auf (Schäfer und
Schmid, 1993).
Biotische und abiotische Stressfaktoren wie sehr starke Strahlungsintensität, aber auch
Verwundung und Stromfluss (Herde et al., 1999; Bergantino et al., 2003) konnten bereits als
ausschlaggebende Faktoren charakterisiert werden, die verschiedene Schutzmechanismen an
dem PS II hervorrufen. Ein solcher Schutzmechanismus ist der Xanthophyllzyklus, in
welchem daraufhin durch das Enzym Violaxanthin-Oxidase vermehrt Zeaxanthin über die

Diskussion
94
Zwischenstufe Antheraxanthin auf Kosten von Violaxanthin gebildet wird. Dieser
Mechanismus kann innerhalb weniger Minuten (ab 1,5 Minuten, nach Schäfer und Schmid,
1993) in Kraft treten, ist reversibel und einhergehend mit einem Anstieg des
nichtphotochemischen Quenchings der Chlorophyllfluoreszenz (Bergantino et al., 2003;
Schäfer und Schmid, 1993). Indirekt proportional zum nichtphotochemischen Quenching
verhält sich das photochemische Quenching, welches in den vorliegenden
Fluoreszenzmessungen an Pappelblättern entsprechend rückläufig war. Es resultiert ein
Rückgang der Quantenausbeute am PS II. Neben der zeitlichen Übereinstimmung dieser
Reaktion mit den gemessenen Daten und der beobachteten Reversibilität, sprechen für diese
Reaktion auf elektrische Signale auch Ergebnisse an Tomate, welche auf Verwundung einen
Anstieg von Zeaxanthin auf Kosten von Violaxanthin bestätigen konnten (Herde et al., 1999).
Die Autoren weisen in diesem Zusammenhang darauf hin, dass das Zeaxanthin bei
Stressreaktionen mit Lipiden der Thylakoidmembran interagieren könnte und auf selbige
gleichsam stabilisieren und schützen wirken könnte.
4.2.1 Bedeutung elektrischer Signalleitung für den Baum
Bei der Frage nach den Ursprüngen der pflanzlichen Signalleitung weiß man heute, dass man
die Evolution sehr weit zurückverfolgen muss; bereits Bakterien, die als Vorläufer der
Pflanzen wie auch der Tiere gelten, zeigen auf bestimmte Reize hin präzise elektrische
Aktivitäten entlang ihrer Oberfläche ( Szmelcman und Adler, 1976). An der einzelligen
Grünalge Acetabularia konnten Aktionspotentiale gemessen werden, die vermutlich
osmotische Funktionen haben (Mummert und Gradmann, 1976). Bei den höheren Pflanzen
haben sich im Laufe der Evolution hochspezialisierte und hochsensitive Systeme entwickelt,
die selbst heute noch erstaunen. Eines der berühmtesten dieser Art ist wohl die
Klappbewegung der Venusfliegenfalle. Der lange Weg vom relativ einfachen System des
Bakteriums zu den komplexen höherer Pflanzen war stets geprägt von der bestmöglichen
Anpassung an den Lebensraum. Die ersten Pflanzen benutzten elektrische Signale unter
anderem als Antrieb für Schwimmbewegungen im Wasser und, wie bei Dinoflagellaten, zur
Biolumineszenz. Seit die Pflanzen das Land eroberten nutzen sie ihre latente Erregbarkeit auf
vielfältige Art und Weise, ihre Immobilität zu kompensieren, beispielsweise durch das Öffnen
und Schließen der Stomata oder durch Blattbewegungen, wie bei der Mimose zu sehen.

Diskussion
95
Durch ihr ausgeprägtes vertikales Wachstum mussten Bäume verschiedene Mechanismen
entwickeln, um ihren Wasser- und Nährstoffhaushalt an ihr Wuchsverhalten anzupassen. Die
Differenzierung des Xylem- und des Phloemgewebes aus den cambialen Initialzellen löste
dieses Problem meisterlich. Wie die Ergebnisse dieser Arbeit verdeutlichen, nutzen Bäume
das Phloemgewebe bei weitem nicht nur zum Assimilattransport; vielmehr ermöglicht ihnen
auch dieser spezielle Gewebetyp mit seinen langgestreckten, lebenden Zellen und seinen
typischen Siebporen in den Siebelementen, mittels elektrischer Signale Informationen
innerhalb kürzester Zeit über verschiedene Pflanzenorgane zu übermitteln. Die spezifischen
Reaktionen auf diese „Warnsignale“ sind bei Bäumen noch nicht ausschöpfend geklärt. Im
Rahmen der vorliegenden Dissertation konnten als baumphysiologische Antwort
Veränderungen in verschiedenen Prozessen der Photosynthese dargestellt werden.
Vergleichbare Untersuchungen an krautigen Pflanzen, wie beispielsweise die Proteinase-
Inhibitor (PIN) - Genaktivierung (Wildon et al., 1992; Rhodes et al., 1996; beide an Tomate),
die auf Phloem - weitergeleiteten elektrischen Signalen basiert, bieten jedoch eine reiche
Inspirationsquelle für weitergehende Untersuchungen der Reaktionen der Bäume auf
elektrische Distanzsignale.

Schlussfolgerung
96
5 Schlussfolgerung
Für die pflanzliche Entwicklung ist eine ausreichende Versorgung mit Calcium dringend
notwendig. Die dargelegten Ergebnisse konnten zeigen, dass sowohl der apoplastische, als
auch der symplastische Calciumgehalt für verschiedene Prozesse in der Pappel limitierend
sein kann. Die Größenordnungen der notwendigen Versorgung gehen dabei allerdings weit
auseinander. Wenn bei der apoplastischen Calciumversorgung in der vorliegenden Arbeit 5
mM Ca2+ als Optimalversorgung angesehen werden, betragen die Konzentrationen für freies
Ca2+ im Cytoplasma nur etwa 0,1 bis 0,2 µM. Bereits dieser Gradient deutet auf eine
grundsätzlich verschiedene Funktion des Nährelements in den verschiedenen Zellbereichen
hin. Wie in dieser Studie gezeigt werden konnte, kommen dem Calcium im apoplastischen
Raum diverse Funktionen zu:
- Es stabilisiert die Mittellamellen und übt somit auf den gesamten Zellverband der
jeweiligen Gewebe eine stützende Funktion aus
- Es nimmt Einfluss auf die Ligninbiosynthese; ob es dabei im Zellwandbereich oder an
der Außenseite der Plasmamembranen verfügbar sein muss, konnte jedoch nicht mit
Sicherheit prognostiziert werden.
- Bei dem intrusiven Spitzenwachstum von Fasern wirkt eine verbesserte
Calciumversorgung wachstumsfördernd.
Auch im Cytoplasma der cambialen Zellen hat die Konzentration an freiem Calcium einen
Einfluss auf die Strukturbildung des sekundären Xylems. Wie die vorliegenden Ergebnisse
zeigen, kann man unter minimierter Calciumversorgung einen allgemeinen Rückgang der
Teilungsrate der Xylemmutterzellen erkennen; der Holzzuwachs sinkt somit rapide. Auch die
unter reduzierter Calcium-Versorgung tendenzielle Rückläufigkeit im Gefäßdurchmesser
könnte auf einen reduzierten [Ca2+]cyt -Wert zurückführbar sein.
Die deutlichsten physiologischen Veränderungen auf einen Rückgang des [Ca2+]cyt -Werts
ließen sich jedoch bei der Signalweiterleitung in den Siebröhren des Phloems feststellen.
Gerade die Untersuchungen an Calcium-verarmten Pappeln ließen für die Signalweiterleitung
den Schluss zu, dass die Richtung, in die sich das Signal fortpflanzt, von entscheidender
Bedeutung für die elektrische Leitfähigkeit der Phloemzellen ist. Die Fähigkeit, elektrische

Schlussfolgerung
97
Signale in ihrer Ausbreitung zu unterstützen könnte durch sich schnell öffnende bzw.
schließende Plasmodesmata sowie Siebporen reguliert werden. Wo Calcium als limitierender
Faktor ins Spiel kommt, scheint die Informationsweiterleitung eines elektrischen Signals nur
noch acropetal zu funktionieren. In Anbetracht der strukturellen und der funktionellen
Übereinstimmungen zwischen Siebporen und Plasmodesmata ist zu vermuten, dass die
elektrische Signalleitung entlang des Phloems für Langstreckensignale optimiert wurde,
wobei in anderen Gewebetypen die elektrische Signalleitung zur Kommunikation über kurze
Strecken von Zelle zu Zelle über Plasmodesmata abläuft (Abb. 5.1).
Abb. 5.1: Schematische Darstellung der elektrischen Signalleitung über kurze und weite Distanzen im
Baum am Beispiel eines typischen Aktionspotentials. Ein Reiz wie z.B. Eiswasser (Stern) induziert einen Ca2+-Einstrom in eine Mesophyllzelle des Blattes (MC). Das Membranpotential depolarisiert
über den Schwellenwert hinaus, so dass sich zunächst Cl--Auswärtskanäle öffnen (Depolarisation) und anschließend K+-Auswärtskanäle (Repolarisation). Die Ausbreitung des induzierten Aktionspotentials erfolgt von Zelle zu Zelle über Plasmodesmata (P) im Rahmen einer Kurzstreckenleitung. Erreicht das
Signal das Phloem, kann es über die Geleitzellen (CC) und deren Plasmodesmata in eine Siebröhre (SE) gelangen und dort bidirektional über weite Distanzen fortgepflanzt werden.
Aufgrund des reich verzweigten Leitbündelsystems ist davon auszugehen, dass viele Reize, wie z. B. Hitze direkt vom Phloem aufgenommen werden und weitergeleitet werden.
Weitere Abkürzung: PA, Parenchymzelle

Schlussfolgerung
98
Im Rahmen dieser Arbeit zeigte sich, dass die Signalweiterleitung über das Phloem
schließlich im Mesophyllgewebe des Blattes endet. Die mit leichter zeitlicher Verzögerung
gemessenen transienten Rückgänge in der CO2-Aufnahmerate der Dunkelreaktion und der
Quantenausbeute des Photosystems II lassen darauf schließen, dass die über das Phloem
transportierte Information des Distanzsignals einen Schutzmechanismus des photosynthetisch
aktiven Gewebes in den Blattorganen der Pappel hervorrief. Der Anstieg des
nichtphotochemischen Quenchings, einhergehend mit dem Abfall des photosynthetischen
Quenchings deutet auf eine Veränderung in dem Xanthophyllzyklus als Schutzreaktion hin.

Zusammenfassung
99
6 Zusammenfassung
In der vorliegenden Arbeit wurde die Bedeutung der Calcium-Versorgung für die
Holzdifferenzierung und die elektrische Signalleitung an der Pappel untersucht. Dafür
wurden zwei verschiedene Pappelklone, Populus tremula x Populus tremuloides T89 und ein
intraspezifischer Hybrid aus Populus trichocarpa x Populus trichocarpa cv Trichobel, in
Hydrokultur bzw. in Bodensubstrat unter verschiedener Calciumernährung angezogen.
Vergleichende Untersuchungen an unterschiedlich mit Calcium versorgten Pappeln (0 mM,
0,1 mM, 1 mM, 5 mM und 10 mM Calcium) haben gezeigt, dass die Calciumernährung
sowohl die Struktur, als auch die Chemie des gebildeten Holzes erheblich beeinflusst:
- Die cambiale Zone war unter Calcium-Mangel in ihrer radialen Ausprägung deutlich
reduziert; im Vergleich zur Optimalversorgung reduzierte sich die cambiale Zone um
etwa die Hälfte auf drei bis vier Zellreihen in radialer Richtung. Die Ausprägung der
Zellexpansionszone war ebenfalls deutlich rückläufig. Die cambialen Zellen selbst
zeigten unter Calciummangel eine deutliche Limitierung in der Ausprägung ihrer
großen Zentralvakuole; im Gegensatz zur Optimalversorgung konnte unter Calcium-
Mangel ein dichtes Cytoplasma mit vielen kleinen Vakuolen beobachtet werden.
- Die aus den cambialen Xylemmutterzellen sich ausdifferenzierenden Libriformfasern
zeigten einhergehend mit einer reduzierten Calciumversorgung einen signifikanten
Rückgang in der Faserlänge. Chemische Analysen der Holzzellwände konnten zudem
darlegen, dass unter minimierter Calciumversorgung der Anteil an S-Lignin rückläufig
ist; es kann somit von einer Reduktion der Ligninsynthese unter Calcium-Mangel
ausgegangen werden. Bei der Ausprägung der Gefäßquerschnittsflächen wurde unter
abnehmender Calciumversorgung eine Tendenz zu kleineren Durchmessern
deutlich.
- Elementanalysen zeigten eine Zunahme von Calcium im Phloem, Cambium und
Xylem des Stammes mit ansteigender Calcium-Versorgung. Diese Beobachtung zeigte
sich besonders deutlich im Phloem.

Zusammenfassung
100
- Bei saisonalen Untersuchungen im Verlauf einer Wachstumsperiode wurde deutlich,
dass mit abnehmender Calciumversorgung ein Rückgang im Trieblängenwachstum,
im Blattflächenzuwachs sowie im Holzzuwachs einhergeht. Demnach wird die
Biomasseproduktion mit abnehmender Calciumversorgung gehemmt.
Die Untersuchungen zur elektrischen Signalleitung sollten zeigen, ob calciumabhängige
Signale über weite Distanzen im Baum fortgeleitet werden, welches Gewebe dem
Ferntransport dient und ob elektrische Signale einen Einfluss auf die Photosynthese ausüben.
Die Durchführung der elektrophysiologischen Untersuchungen an der Pappel erfolgte mit der
Mikroelektrodentechnik im Mesophyll oder mittels der Aphidentechnik im Phloem. Durch die
Aphidentechnik konnte gezeigt werden, dass die gemessenen elektrischen
Spannungsänderungen über die Plasmamembran der Siebröhren über weite Distanzen
fortgeleitet werden. Bei den Untersuchungen wurde zwischen basipetaler und acropetaler
Signalleitung unterschieden. Dabei wiesen die Reaktionen auf die beiden Reiztypen Eiswasser
und Verwundung durch Hitzereizung sowohl Gemeinsamkeiten als auch signifikante
Unterschiede auf. Die wesentlichen Resultate lassen sich wie folgt zusammenfassen:
- Im Phloem konnten als Reaktion auf Kältereizung mittels Eiswasser in acropetaler
Richtung typische Aktionspotentiale, auf Verwundung von Blattgewebe mittels
Hitzreizung hingegen Variationspotential - ähnliche Signale aufgezeichnet
werden. Dabei unterschied sich die acropetal gerichtete Signalleitung mit einer
Depolarisation der Plasmamembran in den Siebröhren deutlich von der basipetal
gerichteten Signalleitung, die an den Plasmamembranen eine Hyperpolarisation
auslöste. Diese Reaktionen konnten auch für das Mesophyllgewebe bestätigt
werden.
- Unter Calcium-Mangel konnte eine Signalweiterleitung lediglich bei acropetal
gerichteten Distanzsignalen festgestellt werden; die Amplitude der Signale fiel unter
Calcium-Mangel zudem deutlich geringer aus als unter Optimalversorgung. Eine
basipetal gerichtete Signalweiterleitung konnte unter Calcium-Mangel nicht detektiert
werden. Dieser Umstand deutet auf eine Selektivität der Signaltransduktion im
Phloemgewebe der Pappel hin.
Durch Applikation von TEA+, einem Kaliumkanal-Blocker, konnten
hyperpolarisierende Signale eliminiert werden. Aus dieser Feststellung ließ sich

Zusammenfassung
101
folgern, dass ein Kalium-Ausstrom die Hyperpolarisation des Membranpotentials
verursacht.
- Als physiologische Reaktion auf die elektrische Signalleitung konnte in der
Dunkelreaktion der Photosynthese ein transienter Rückgang in der CO2-Aufnahme
festgestellt werden. Diese Beobachtung konnte bei basipetal gleichsam wie bei
acropetal gerichteter Signalweiterleitung in den Blättern gemacht werden. Eine
parallel dazu erfolgende Veränderung in der stomatären Leitfähigkeit konnte jedoch
nicht festgestellt werden. Durch Anlage eines Kälteblocks ließ sich weiterhin zeigen,
dass das elektrische, im Phloem fortgeleitete Signal hierdurch geblockt wird und
anschließend keine Veränderung in der CO2-Aufnahmerate stattfindet. Somit konnte
deutlich gezeigt werden, dass die vorübergehende Reduktion der CO2-Aufnahmerate
durch das elektrische Signal ausgelöst wird.
- Einige Sekunden nach Eintreffen des elektrischen Signals im Blatt zeigte sich
zunächst die Reaktion im Gaswechsel. Anschließend konnte an den Blättern ein
transienter Rückgang der Quantenausbeute des Photosystems II beobachtet
werden. Dieser ging einher mit einem dazu indirekt proportionalen Anstieg des
nichtphotochemischen Quenchings und lässt damit auf eine Veränderung im
Xanthophyllzyklus als Reaktion auf ein elektrisches Signal schließen. Die
Signalweiterleitung konnte bei elektrischen Distanzsignalen in acropetaler Richtung
deutlich als von den Blattadern ins Mesophyllgewebe der Intercostalfelder übergehend
gezeigt werden. Im Gegensatz dazu konnte bei basipetaler Signalleitung innerhalb
eines Blattes gezeigt werden, dass die Quantenausbeute des PS II in Blattadern und
Intercostalfeldern gleichzeitig absinkt, was auf eine simultane Signalleitung über
Plasmodesmata des Mesophylls und des Phloems innerhalb eines Blattes hindeutet.
Anhand der vorliegenden Ergebnisse konnte gezeigt werden, dass das Kation Calcium in
Angiospermen mehrere wichtige Funktionen erfüllt. Zum einen ist es in Bäumen in zahlreiche
Prozesse wie z.B. der Gewebestabilisierung, der Zellwandbildung und der
Faserdifferenzierung involviert, zum anderen spielt es auch eine wichtige Rolle bei der
elektrischen Informationsweiterleitung innerhalb von Blattorganen sowie über weite
Distanzen zwischen verschiedenen Blättern.

Literatur
102
7 Literatur
Albrecht, W.A., Davis, D.E. (1929) Physiological importance of calcium in legume
inoculation. Botanical Gazette 88: 310-321
Aloni, R. (1987) Differentiation of vascular tissues. Annual Review of Plant Physiology 38:
179-204
Anil, V.S., Rao, K.S. (2001) Calcium-mediated signal transduction in plants: A perspective on
the role of Ca2+ and CDPKs during early plant development. Journal of Plant
Physiology 158: 1237-1256
Arend, M., Fromm, J. (2003) Ultrastructural changes in cambial cell derivates during xylem
differentiation in poplar. Plant Biology 5: 255-264
Arnon, D.I., Stout, P.R. (1939) The essentiality of certain elements in minute quantity for
plants with special reference to copper. Plant Physiology 14: 371-375
Bailey, I.W. (1920) The cambium and its derivative tissues. II. Size variations of cambial
initials in gymnosperms and angiosperms. American Journal of Botany 7: 355-367
Bailey, I.W., Shepard, H.B. (1915) Sanio´s laws for the variation in size of coniferous
tracheids. Botanical Gazette 60: 66-71
Beilby, M.J., Coster, H.G.L. (1979) The action potential in Chara corallina. II. Two
activation-inactivation transients in voltage clamps of plasmalemma. Australian
Journal of Plant Physiology 6: 329-335
van Bel; A., Gaupels, F., Will, T., Kogel, K.-H. (2003) Phloem-mediated remote control by
long-distance signals. In: Progress in Botany 65. Springer Verlag. pp.372-392
Bergantino, E., Segalla, A., Brunetta, A., Teardo, E., Rigoni, F., Giacometti, G.M.,
Szabò, I. (2003) Light- and pH-dependent structural changes in the PsbS subunit of
photosystem II. Proceedings of the National Academy of Sciences 100: 15265-15370

Literatur
103
Bissett, I.J.W., Dadswell, H.E. (1950) The variation in cell length within one growth ring of
certain angiosperms and gymnosperms. Australian Forestry 14: 17-29
Boerjan, W., Ralph, J., Baucher, M. (2003) Lignin biosynthesis. Annual Review of Plant
Biology 54: 519-546
Borchert, R. (1990) Ca2+ as developmental signal in the formation of Ca-oxalate crystal
spacing patterns during leaf development in Carya ovata. Plant 182: 339-347
Bose, J.C. (1924) The physiology of photosynthesis. Longman, Green and Co (eds.), London,
UK
Brady, S.L. (1969) The effect of mineral nutrients on the anatomy of Larix occidentalis
seedlings. Dissertation. Pennsylvania State University, USA
Brett, C., Waldron, K. (1996) Physiology and biochemistry of plant cell walls. 2. Auflage.
Chapman & Hall. London. 255 pp.
Bulychev, A.A., Niyazova, M.M., Rubin, A.B. (1987) Fluorescence changes of chloroplasts
caused by the shifts of membrane-potential and their dependence on the redox state of
the acceptor of photosystem-II. Biologicheskie Membrany 4: 262-269
Bulychev, A.A., Niyazova, M.M., Turovetsky, V.B. (1986) Electro-induced changes of
chlorophyll fluorescence in individual intact chloroplasts. Biochimica and Biophysica
Acta 850: 218-225
Burdon-Sanderson, J. (1873) Note to the electrical phenomena which accompany irritation of
the leaf of Dionaea muscipula. Proceedings of the Royal Society (London) 21: 495-
496
Burdon-Sanderson, J., Page, J.F.M. (1876) On the mechanical effects and on the electrical
disturbance consequent on excitation of the leaf of Dionaea muscipula. Proceedings of
the Royal Society, London 25: 411-434

Literatur
104
Bush, D.S. (1995) Calcium regulation in plant cells and its role in signaling. Annual Review
of Plant Physiology and Plant Molecular Biology 46: 95-122
Bussler, W. (1963) Die Entwicklung von Calcium-Mangelsymptomen. Zeitschrift für
Pflanzenernährung, Düngung und Bodenkunde 100: 53-58
Campbell, M.M., Sederoff, R.R. (1996) Variation in lignin content and composition. Plant
Physiology 11: 3-13
Canny, M.J.P. (1975) Mass transfer. In: Zimmermann,H.M., Milburn, J.A. (eds.)
Enzyclopedia of plant physiology. Springer, Berlin, Germany, S. 139-153
Carpin, S., Crèvecoeur, M., de Meyer, M., Simon, P., Greppin, H., Penel, C. (2001)
Identification of a Ca2+-pectate binding site on an apoplastic peroxidase. Plant Cell
13:511 20
Chaffey, N., Barnett, J., Barlow, P. (1997) Endomembranes, cytoskeleton and cell walls:
aspects of the ultrastructure of the vascular cambium of taproots of Aesculus
hippocastanum L. (Hippocastanacées). International Journal of Plant Sciences 158:
97-109
Darwin, C. (1875) Insectivorous plants. Murray, J. (ed.), London, UK
Darwin, C. (1881) The power of movement in plants. Murray, J. (ed.), London, UK
Davies, E. (1987) Actionpotentials as multifunctional signals in plants - a unifying hypothesis
to explain apparently disparate wound responses. Plant, Cell & Environment 10: 623-
631
Davies, E., Schuster, A.M. (1981) Intercellular communication in plants: evidence for a
rapidly-generated, bidirectionally-transmitted wound signal. Proceedings of the
National Academy of Sciences, USA 78: 2422-2426

Literatur
105
Davies, D.E. (1949) Some effects of calcium deficiency on the anatomy of Pinus taeda.
American Journal of Botany 86: 276-282
Dudel, J., Menzel, R., Schmidt, R. (1996) Neurowissenschaft - vom Molekül zur Kognition.
Springer-Verlag, 559 pp.
Dünisch, O., Bauch, J. (1994a) Influence of mineral elements on wood formation of old
growth spruce (Picea abies [L.] Karst.). Holzforschung 48: 5-14
Eklund,L., Eliasson, L. (1990) Effects of calcium ion concentration on cell wall synthesis.
Journal of Experimental Botany 41: 863-867
Eschrich, W., Blechschmidt-Schneider, S. (1992) Extrinsic and intrinsic influences on tree
growth - anatomical analysis of spruce branches. Trees 6: 179-185
Evans, D.E., Briars, S.-A., Williams, L.E. (1991) Active calcium transport by plant cell
membranes. Journal of Experimental Botany 42: 285-303
Evert, R.F., Eschrich, W., Eichhorn, S.E., Limbach, S.T. (1973) Observations on penetration
of barley leaves by the aphid Rhopalosiphum maidis (Fitch.). Protoplasma 77: 95-110
Faix, O. (1992) Fourier Transform Infrared Spectroscopy. In: Methods in Lignin Chemistry.
Lin S Y, Dence C W (Eds.). pp. 83-109
Felle, H. (1988) Cytoplasmic free calcium in Riccia fluitans L. and Zea mais L.: Interactions
of Ca2+ and pH? Planta 176: 248-255
Fengel, D., Ludwig, M. (1991) Möglichkeiten und Grenzen der FTIR-Spektroskopie bei der
Charakterisierung von Cellulose. Papier 45: 45-51
Fink, S. (1991a) Comparative microscopical studies on the patterns of calcium oxalate
distribution in the needles of various conifer species. Botanica Acta 102: 306-315

Literatur
106
Fink, S. (1991b) The micromorphological distribution of bound calcium in needles of Norway
spruce (Picea abies (L.) Karst.) New Phytologist 119: 33-40
Fink, S. (1991c) Unusual patterns in the distribution of calcium oxalate in spruce needles and
their possible relationships to the impact of pollutants. New Phytologist 119: 41-51
Follet-Gueye, M.-L., Verdus, M.C., Demarty, M., Thellier, M., Ripoll, C. (1998) Cambium
pre-activation in beech correlates with a strong temporary increase of calcium in
cambium and phloem but not in xylem cells. Cell Calcium 24: 205-211
Fraser R. T. (1959) Sodium Azide as an Internal Standard for Quantitative Infrared Analysis.
Analytical Chemistry 31: 1602-1603
Fromm, J., Fei, H. (1998) Electrical signalling and gas exchange in maize plants of drying
soil. Plant Science 132: 203-213
Fromm J (1991) Control of phloem unloading by action potentials in Mimosa. Physiologia
Plantarum 83: 529-533
Fromm, J., Bauer, T. (1994) Action potentials in maize sieve tubes change phloem
translocation. Journal of Experimental Botany 45: 463-469
Fromm J, Spanswick R (1993) Characteristics of action potentials in willow (Salix viminalis
L.). Journal of Experimental Botany 44:1119-1125
Genty, B., Briantais, J.-M., Baker, N.R. (1989) The relationship between the quantum yield of
photosynthetic electron transport and quenching of chlorophyll fluorescence.
Biochimica et Biophysica Acta 990: 87-92
Guglielmino, N., Liberman, M., Jauneau, A., Vian, B., Catesson, A.M., Goldberg, R. (1997)
Pectin immunolocalization and calcium visualization in differentiating derivatives
from poplar cambium. Protoplasma 199: 151-160

Literatur
107
Herde, O., Pena-Cortes, H., Fuss, H., Willmitzer, L., Fisahn, J. (1999) Effects of mechanical
wounding, current application, and heat treatment on chlorophyll fluorescence and
pigment position in tomato plants. Physiologia Plantarum 105: 179-184
Hirschi, K.D. (2004) The calcium conundrum. Both versatile nutrient and specific signal.
Plant Physiology 136: 2438-2442
Hoagland, D.R., Arnon, D.I. (1950) The water culture method for growing plants without soil.
Calif. Agric. Exp. Stn. Bull. 347: 1-32
Houwink, A.L. (1935) The conduction of excitation in Mimosa pudica. Recueil des Travaux
Botaniques Néerlandaise 32: 51-91
Hu, W.-J., Harding, S.A., Lung, J., Popko, J.L., Ralph, J. (1999). Repression of lignin
biosynthesis promotes cellulose accumulation and growth in transgenic trees. Nature
Biotechnology 17:808 12
Jones, R.L., Gilroy, S., Hillmer, S. (1993) The role of calcium in the hormonal regulation of
enzyme synthesis and secretion in barley aleurone. Journal of Experimental Botany
44: 207-212
Kasai, M., Muto, S. (1990) Ca2+ pump and Ca2+/H+ antiporter in plasma membrane vesicles
isolated by aqueous two phase partitioning from corn leaves. Journal of Membrane
Biology 114: 133-142
Kikuyama, M. (1987) Ion efflux during a single action potential of Nitella axilliformis in
medium lacking Ca2+. Plant and Cell Physiology 28: 179-186
Kikuyama, M., Tazawa, M. (1983) Transient increase of intracellular Ca2+ during excitation
of tonoplast-free Chara cells. Protoplasma 117: 62-67
Knight, H. (2000) Calcium signalling during abiotic stress in plants. International Review of
Cytology 195: 269-324

Literatur
108
Knight, H., Knight, M.R. (2001) Abiotic stress signalling pathways: Specificity and cross-
talk. Trends in Plant Science 6: 262-267
van Kooten, O., Snel, J.F.H. (1990) The use of chlorophyll fluorescence nomenclature in
plant stress physiology. Photosynthesis Research 25: 147-150
Koziolek, C., Grams, T.E.E., Knobloch, P. Koch, M. Matyssek, R., Fromm, J. (2005) Electric
and hydraulic signals trigger stomatal movement upon re-irrigation in Zea mays L.
Plant & Cell Physiology , eingereicht zur Publikation
Koziolek, C., Grams, T.E.E., Schreiber, U., Matyssek, R., Fromm, J. (2004) Transient
knockout of photosynthesis mediated by electrical signals. New Phytologist 161: 715-
722
Krabel, D. (2000) Influence of sucrose on cambial activity. In: Cell and Molecular Biology of
Wood Formation. Hsg. Savidge, R.A., Barnett, J.R. und Napier, R.. BIOS Scientific
Publishers Ltd, Oxford, 113-125 pp.
Lachaud, S., Bonnemain, J.I. (1982) Xylogénése chez les dicotylédones arborescentes. III.
Transport de láuxine et activité cambiale dans les jeunes tiges de hêtre. Canadian
Journal of Botany 60 : 869-876
Langer, K., Ache, P., Geiger, D., Stinzing, A., Arend, M., Wind, C., Regan, S., Fromm, J.,
Hedrich, R. (2002) Poplar potassium transporters capable of controlling K+
homeostasis and K+-dependent xylogenesis. Plant Journal 32: 997-1009
Langer, K., Levchenko, V., Fromm, J., Geiger, D., Steinmeyer, R., Lautner, S., Ache, P.,
Hedrich, R. (2003) The poplar K+ channel KPT1 is associated with K+ uptake during
stomatal opening and bud development. Plant Journal 37: 828-838
Larson, P.R. (1994) The Vascular Cambium. Development and Structure. Springer Verlag.
Series in Wood Science. 725 pp.

Literatur
109
Lewis, N.G. (1997) A 20th century roller coaster ride: a short account of lignification. Current
Opinion in Plant Biology 2: 153-162
Lunevsky, V.Z., Zherelovam O.M., Vostrikov, I.Y., Berestovsky, G.N. (1983) Excitation of
Characeae cell membranes as a result of activation of calcium and chloride channels.
Journal of Membrane Biology 72: 43-58
Malone, M. (1994) Wound-induced hydraulic signals and stimulus transmission in Mimosa
pudica L. New Phytology 128: 49-56
Mansson, P., Samuelsson, B. (1981) Quantitative determination of O-acetyl and other O-
acetyl groups in cellulosic material. Svensk Papperstidning 84: R15-R24
Marschner, H. (1995) Mineral nutrition of higher plants. 2 Auflage. Academic Press. San
Diego. 889 pp.
McAnish , M.R., Hetherington, A.M. (1998) Encoding specificity in Ca2+ signalling systems.
Perspectives 3: 22-26
McCann, M.C., Stacey, N.J., Wilson, R., Roberts, K. (1993) Orientation of macromolecules in
the walls of elongating carrot cells. Journal of Cell Science 106: 1347-1356
McLaughlin, S.B., Wimmer, R. (1999) Calcium physiology and terrestrial ecosystem
processes. Tansley Review No. 104. New Phytologist 142: 373-417
Mellerowitcz, E.J., Baucher, M., Sundberg, B., Boerjan, W. (2001) Unravelling cell wall
formation in the woody dicot stem. Plant Molecular Biology 47: 239-274
Mohr, H., Schopfer, P. (1992) Pflanzenphysiologie. 4. Auflage. Springer Verlag, Berlin. 619
pp.
Morikawa, H., Hayashi, R., Senda, M. (1978) Infrared analysis of pea stem cell walls and
oriented structure of matrix polysaccharides in them. Plant and Cell Physiology 19:
1151-1159

Literatur
110
Mummert, H., Gradmann, D. (1976) Voltage dependent potassium fluxes and the significance
of action potentials in Acetabularia. Biochimica et Biophysica Acta 443 :443-450
Mwesigwa, J., Collins, D.J., Volkov, A.G. (2000) Electrochemical signalling in green plants:
effects of 2,4-dinitophenol on variation and action potentials in soybean.
Bioelectrochemistry 51: 201-205
Nakano T, Miyazaki J (2003) Surface fractal dimensionality and hygroscopicity for heated
wood. Holz als Roh- und Werkstoff 57: 289-294
Nobel, P.S. (1991) Physiochemical and Environmental Plant Physiology. Academic Press,
San Diego, USA
Nultsch, W. (2001) Allgemeine Botanik. 11. Auflage. Thieme Verlag, Stuttgart. 599 pp.
Ochi, R., Nishiye, H. (1974) Effect of intracellular tetraethylammonium ion on action
potential in the guinea-pig´s myocardium. European Journal of Physiology 348: 305-
316
Overall, R.L., Gunning, B.E.S. (1982) Intercellular communication in Azolla roots. I.
Electrical coupling. Protoplasma 111: 151-160
Panshin, A.J., DeZeeuw, C. (1980) Textbook of wood technology. 4. Ausgabe. McGraw-Hill
New York
Pickard, E. (1973) Action potentials in higher plants. Botanical Review 39: 172-201
Plieth, C., Sattelmacher, B., Hansen, U.-P. (1998) Light-induced Ca2+ -transients in green
Plants I: Methodological aspects. Planta 207: 42-51
Plieth, C., Sattelmacher, B., Hansen, U.-P. (1998) Light induced cytosolic calcium transients
in green plant cells II: The effect on the K+-channel as studied by kinetic analysis in
Chara corallina. Planta 207: 52-59

Literatur
111
Plieth, C. (2005) Calcium: Just another regulator in the machinery of life? Annals of Botany -
Botanical Briefings (im Druck)
Plomion, C., Leprovost, G., Stokes, A. (2001) Wood formation in trees. Plant Physiology 127:
1513-1523
Ranocha, P., Goffner, D., Boudet, A.M. (2000) Plant laccases: are they involved in
lignification? In: Cell and Molecular Biology of Wood Formation. Hsg. Savidge, R.A.,
Barnett, J.R. und Napier, R.. BIOS Scientific Publishers Ltd, Oxford, 397-410 pp.
Raven, J.A. (1987) The role of vacuoles. New Phytology 106: 357-422
Reynolds, E.S. (1963) The use of lead citrate at high pH as an electronopaque stain in electron
microscopy. Journal of Cell Biology 17: 208-212
Rhodes, J., Thain, J.F., Wildon, D.C. (1996) The pathway for systemic electrical signal
transduction in the wounded tomato plant. Planta 200: 50-57
Ricca, U., (1916) Soluzione d´un problema di fisiologia. La propagazione di stimulo nelle
Mimosa. Nuovo Giornale Botanico Italiano 23: 51-170
Rudd, J.J., Franklin-Tong, V.E. (2001) Unravelling response-specificity in Ca2+ signalling
pathways in plant cells. New Phytologist 151: 7-33
Sanders, D., Brownlee, C., Harper, J.F. (1999) Communicating with calcium. Plant Cell 11:
691-706
Schäfer, C., Schmid, V. (1993) Pflanzen im Lichtstreß. Biologie in unserer Zeit 23: 55-62.
Schönknecht, G., Bauer, C.S., Simonis, W. (1998) Light-dependent signal transduction and
transient changes in cytosolic Ca2+ in a unicellular green alga. Journal of Experimental
Botany 49: 1-11

Literatur
112
Schreiber, U., Schliwa, U., Bilger, W. (1986) Continuous recording of photochemical and
non-photochemical chlorophyll fluorescence quenching with a new type of modulation
fluorometer. Photosynthesis Research 10: 51-62
Schulte-Baukloh, C., Fromm, J. (1993) The effect of calcium starvation on assimilate
partitioning and mineral distribution of the phloem. Journal of Experimental Botany
44: 1703-1707
Schumaker, K.S., Sze, H. (1990) Solubilization and reconstitution of the oat root vacuolar
H+/Ca2+ exchanger. Plant Physiology 92:340-345
Shimaji, K. (1950) On the relation between fiber length of some dicotyledonous woods and
the tension wrought on the cambium. Journal of Japanese Forest Society 32: 371-372
Sibaoka, T. (1991) Rapid plant movements triggered by action potentials. Botanical Magazine
Tokyo 104: 73-95)
Sibaoka, T. (1969) Physiology of rapid movements in higher plants. Annual Review of Plant
Physiology 20: 165-184
Sibaoka, T. (1966) Action potentials in plant organs. Symposium of the Society for
Experimental Biology 20: 49-73
Snedden, W.A., Fromm, H. (1998) Calmodulin, calmodulin-related proteins and plant
responses to the environment. Trends in Plant Science 3: 299-304
Spurr, A.E. (1969) A low-viscosity epoxy resin embedding medium for electron microscopy.
Journal of Ultrastructural Research 26: 31-41
Szmelcman, S., Adler, J. (1976) Change in membrane potential during bacterial chemotaxis.
Proceedings of the National Academy of Sciences, USA 73: 4387-4391

Literatur
113
Taylor, A.R., Brownlee, C. (2003) A novel Cl- inward-rectifying current in the plasma
membrane of the calcifying marine phytoplankton Coccolithus pelagicus. Plant
Physiology 131: 1391-1400
Tazawa M, Shimmen T, Mimura T (1987) Membrane control in the Characeae. Annual
Review of Plant Physiology 38: 95-117
Thiel, G., Brüdern, A., Gradmann, D. (1996) Small inward rectifying K+ channels in
coleoptiles: Inhibition by external Ca2+ and function in cell elongation. Journal of
Membrane Biology 149: 9-20
Thiel, G., Homann, U., Plieth, C. (1997) Ion channel activity during the action potential in
Chara: a new insight with new techniques. Journal of Experimental Botany 48: 609-
622
Tinz-Füchtmeier, A., Gradmann, D. (1990) Laser-interferometric re-examination of rapid
conductance of excitation in Mimosa pudica. Journal of Experimental Botany 41: 15-
19
Trewavas, A.J. (2000) Signal perception and transduction. In: Buchnan, B.B., Gruissem, W.,
Jones, R.L. (eds.) Biochemistry and Molecular Biology of Plants. American Society of
Plant Physiologists, Rockville, Maryland, USA, S. 930-987
Trewavas, A.J., Malho, R. (1997) Signal perception and transduction: the origin of the
phenotype. Plant Cell 9: 1181-1195
Trockenbrodt, M. (1995) Calcium oxalate crystals in the bark of Quercus robur, Ulmus
glabra, Populus tremula and Betula pendula. Annals of Botany 75: 281-284
Tsutsui, I., Ohkawa, T., Nagai, R., Kishimoto, U. (1986) Inhibition of Cl- channel activation
in Chara corallina membrane by lanthanum ion. Plant and Cell Physiology 27: 1197-
1200

Literatur
114
Volkov, A.G., Mwesigwa, J. (2001) Electrochemistry of soybean: effects of uncouplers,
pollutants and pesticides. Journal of Electroanalytical Chemistry 496: 153-157
Volkov, A.G., Collins, D.J., Mwesigwa, J. (2000) Plant electrophysiology: pentachlorophenol
induces fast action potentials in soybean. Plant Science 153:185-190
Vrettos, J.S., Stone, D.A., Brudvig, G.W. (2001) Quantifying the ion selectivity of the Ca2+
site in photosystem II: evidence for direct involvement of Ca2+ in O2 formation.
Biochemistry 40: 7937-7945
Wacke, M., Thiel, G. (2001) Electrically triggered all-or-non Ca2+-liberation during action
potential in the giant alga Chara. Journal of General Physiology 118: 11-21
Wardrop, A.B. (1976) Lignification of the plant cell wall. Applied Polymeric Symposium 28:
1041-1063
Weisenseel, M.H., Becker, H.F., Ehlgötz, J.G. (1992) Growth, gravitropism and endogenous
ion currents of cress roots (Lepidium sativum L.). Plant Physiology 100: 16-25
Westermark, U. (1982) Calcium promoted phenolic coupling by superoxide radical - a
possible lignification reaction in wood. Wood Science and Technology 16: 71-78
White, P.J., Broadley, M.R. (2003) Calcium in plants. Annals of Botany 92: 487-511
Wiberley S. E., Sprague J. W., Campbell J. E. (1957) Quantitative infrared analysis of solids
in potassium bromide using an internal standard. Analytical Chemistry 29: 210-213
Wildon, D.C., Thain, J.F. Minchin P.E.H., Gubb, I.R., Reilly, A.J., Skipper, Y.D., Doherty,
H.M., O´Donnell, P.J., Bowles, D.J. (1992) Electrical Signalling and systemic
proteinase inhibitor induction in the wounded plant. Nature 360: 62-65
Williams, D.H., Fleming, I. (1980) Infrared spectroscopy. In: Spectroscopic Methods in
Organic Chemistry, 3. Auflage. McGraw Hill, New York. pp 35-73

Literatur
115
Wong, B.S., Adler, M. (1986) Tetraethylammonium blockade of calcium-activated potassium
channels in clonal anterior pituitary cells. European Journal of Physiology 407: 279-
284

Anhang
116
8 Anhang
8.1 Gaswegeplan der Porometers CQP 130:
Abb. 8.1: Gaswegeplan des Porometers CQP 130 zur Messung des Blattgaswechsels an Pappel.
8.2 Reaktion im Gaswechsel auf Hitzereiz an der Blattspitze (Populus trichocarpa) A)

Anhang
117
B)
C)

Anhang
118
D)
E)
Abb. 8.2: A - E Repräsentative Reaktionen der Netto-Photosyntheserate (JCO2) und der stomatären Leitfähigkeit (gH2O) in Pappelblättern auf einen Hitzereiz an der Blattspitze des Messblattes. Der
Zeitpunkt der Reizung ist durch Pfeile gekennzeichnet.
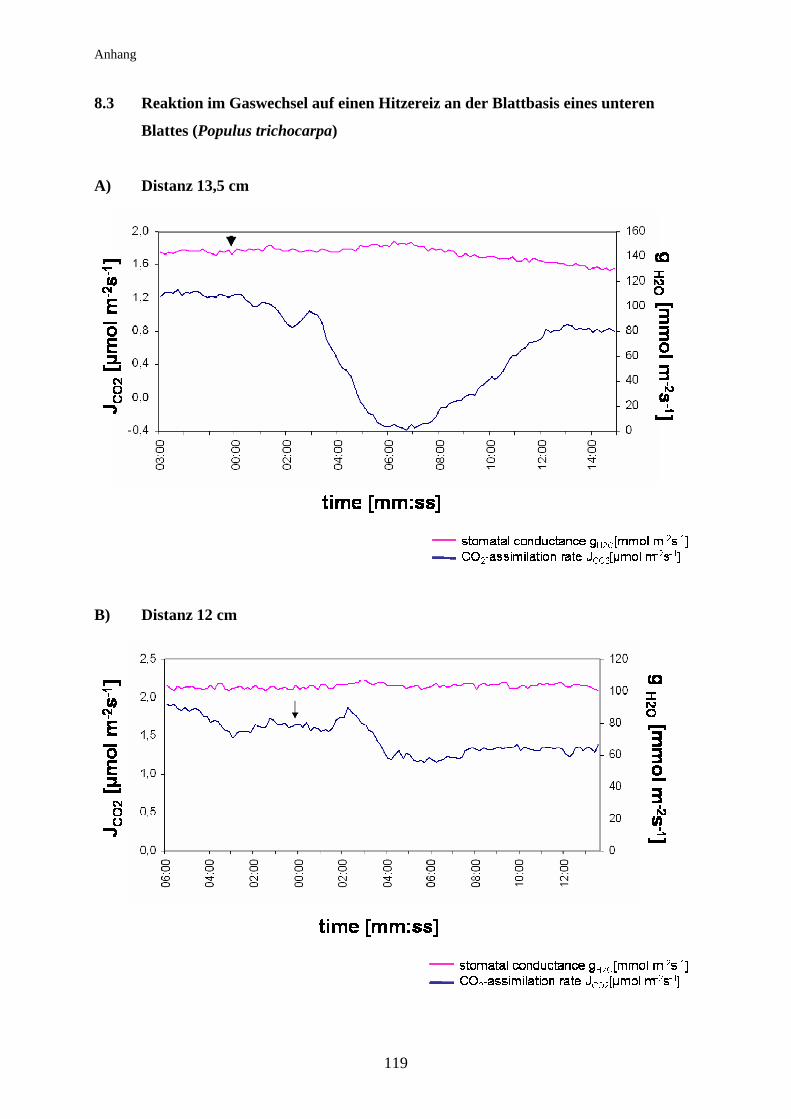
Anhang
119
8.3 Reaktion im Gaswechsel auf einen Hitzereiz an der Blattbasis eines unteren
Blattes (Populus trichocarpa)
A) Distanz 13,5 cm
B) Distanz 12 cm

Anhang
120
C) Distanz 9 cm
Abb. 8.3: A - C Repräsentative Reaktionen der Netto-Photosyntheserate (JCO2) und der stomatären Leitfähigkeit (gH2O) in Pappelblättern auf einen Hitzereiz an der Blattbasis eines unteren Blattes hin.
Der Zeitpunkt der Reizung ist durch Pfeile gekennzeichnet.

Anhang
121
8.4 Reaktion im Gaswechsel auf einen Hitzereiz an der Blattbasis eines unteren
Blattes bei unter Calcium-Mangel angezogenen Pappeln
(Populus tremula x Populus tremuloides)
A) Distanz 7,5 cm
B) Distanz 5 cm

Anhang
122
C) Distanz 10 cm
D) Distanz 5,5cm

Anhang
123
E) Distanz 6,5 cm
Abb. 8.4: A - E Repräsentative Reaktionen der Netto-Photosyntheserate (JCO2) und der stomatären Leitfähigkeit (gH2O) in Pappelblättern auf einen Hitzereiz an der Blattbasis eines unteren Blattes hin.
Die bemessenen Pappeln wurden in Hydrokultur unter minimaler Calciumversorgung (0,1 mM) angezogen. Der Zeitpunkt der Reizung ist durch Pfeile gekennzeichnet.

Anhang
124
8.5 Reaktionen in der Quantenausbeute des Photosystems II auf einen Hitzereiz an
der Blattspitze des Messblattes (Populus trichocarpa)
A)
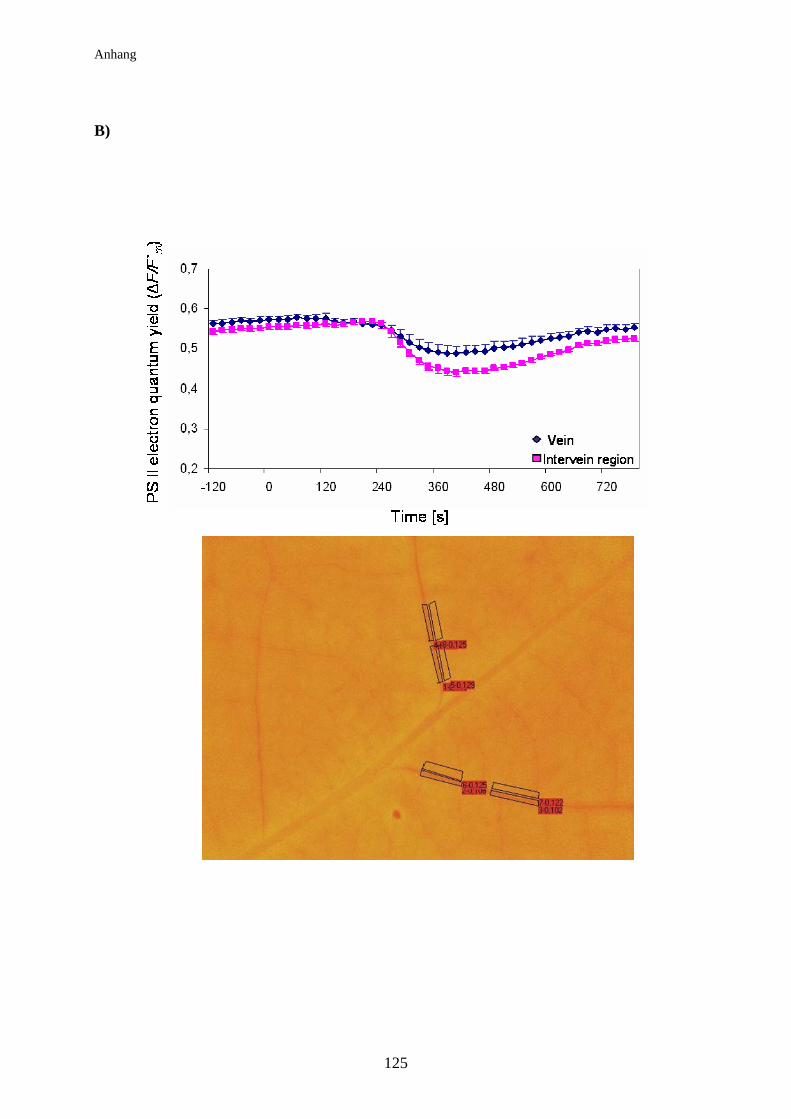
Anhang
125
B)

Anhang
126
C)

Anhang
127
D)
Abb. 8.5: A - D Repräsentative Reaktionen der Quantenausbeute des PS II auf eine Hitzereizung an der Messblattspitze zum Zeitpunkt 0. Distanz jeweils ca. 4 cm. Gleichzeitiger Abfall im Yield-Wert der Adern und der direkt angrenzenden Intercostalfelder auf das basipetal gerichtete Signal hin. Die
unteren Bilder zeigen die Messflächen der oben dargestellten Graphen.
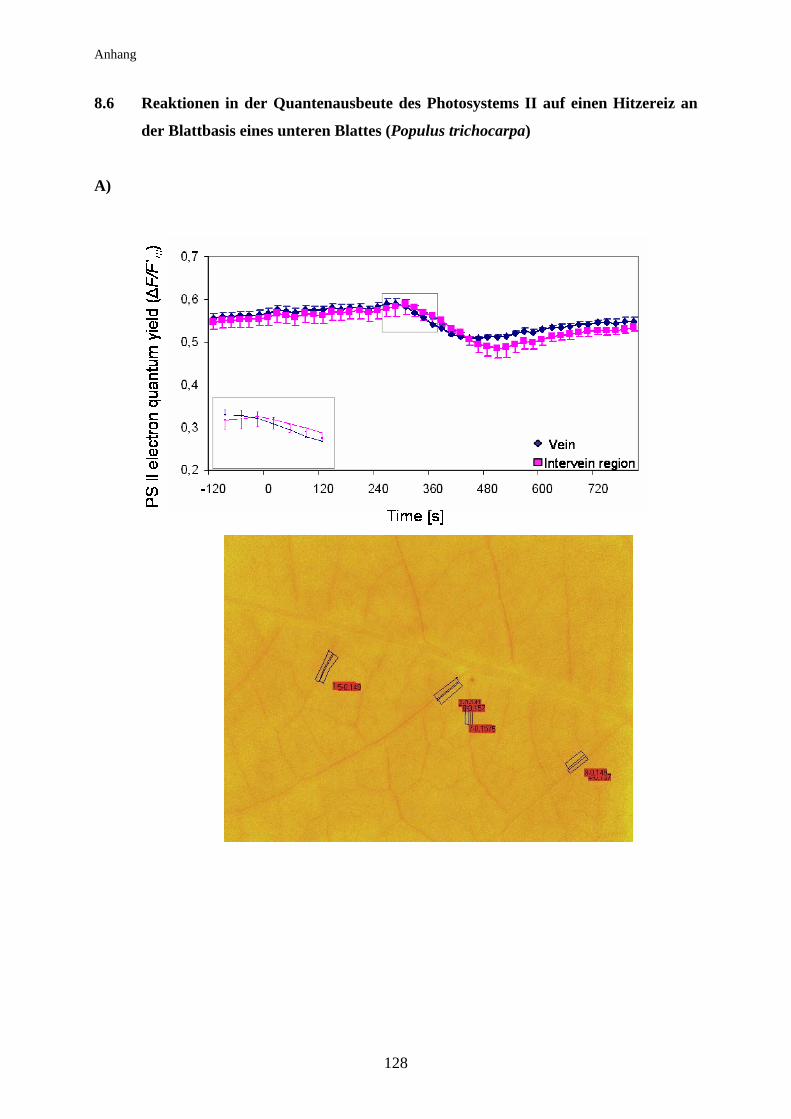
Anhang
128
8.6 Reaktionen in der Quantenausbeute des Photosystems II auf einen Hitzereiz an
der Blattbasis eines unteren Blattes (Populus trichocarpa)
A)

Anhang
129
B)
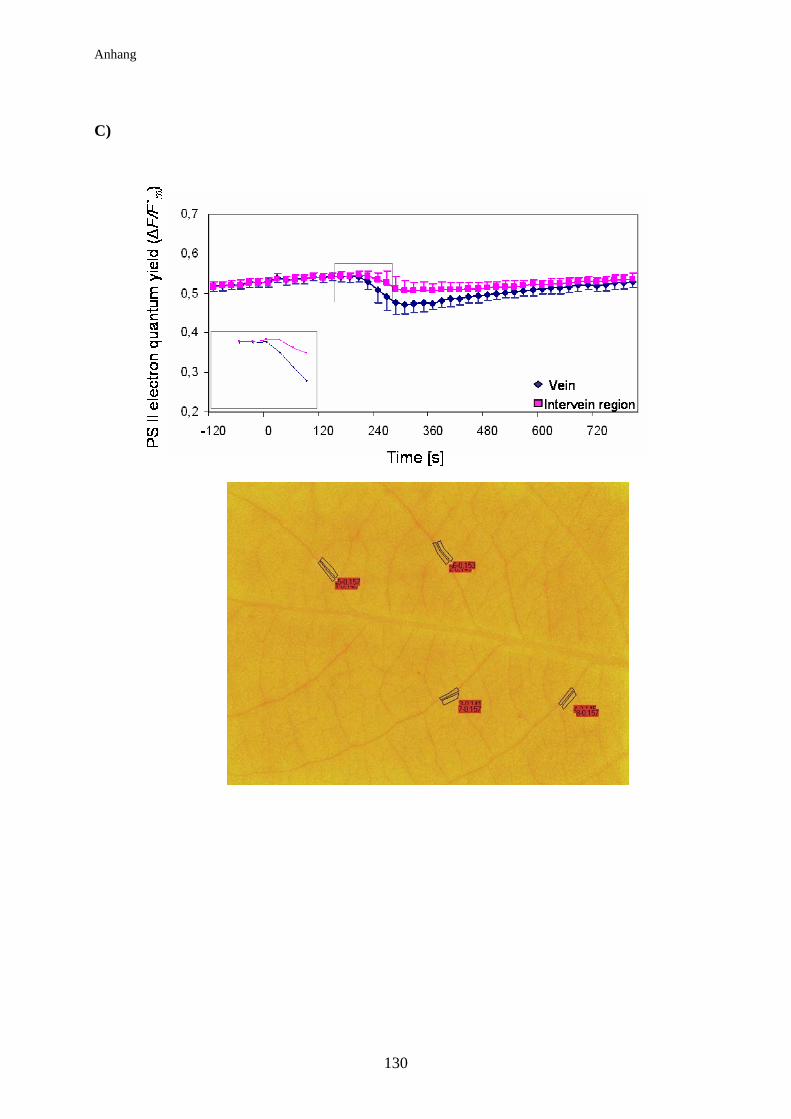
Anhang
130
C)
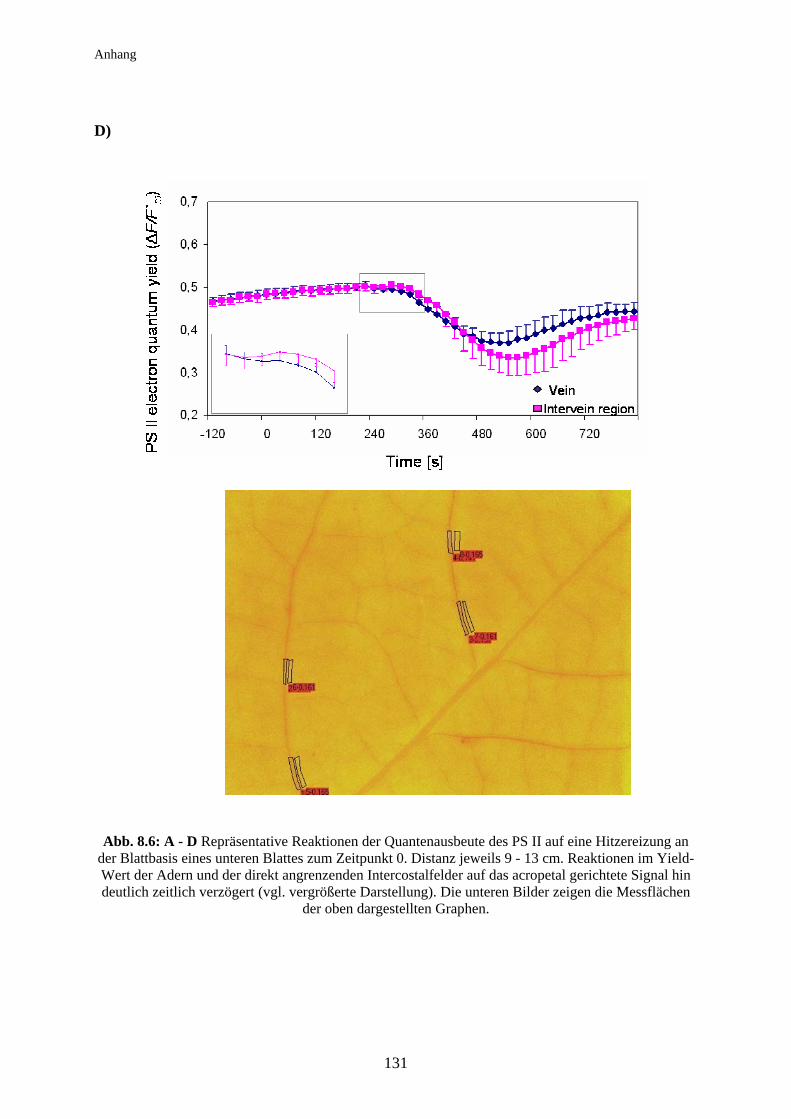
Anhang
131
D)
Abb. 8.6: A - D Repräsentative Reaktionen der Quantenausbeute des PS II auf eine Hitzereizung an der Blattbasis eines unteren Blattes zum Zeitpunkt 0. Distanz jeweils 9 - 13 cm. Reaktionen im Yield-Wert der Adern und der direkt angrenzenden Intercostalfelder auf das acropetal gerichtete Signal hin deutlich zeitlich verzögert (vgl. vergrößerte Darstellung). Die unteren Bilder zeigen die Messflächen
der oben dargestellten Graphen.
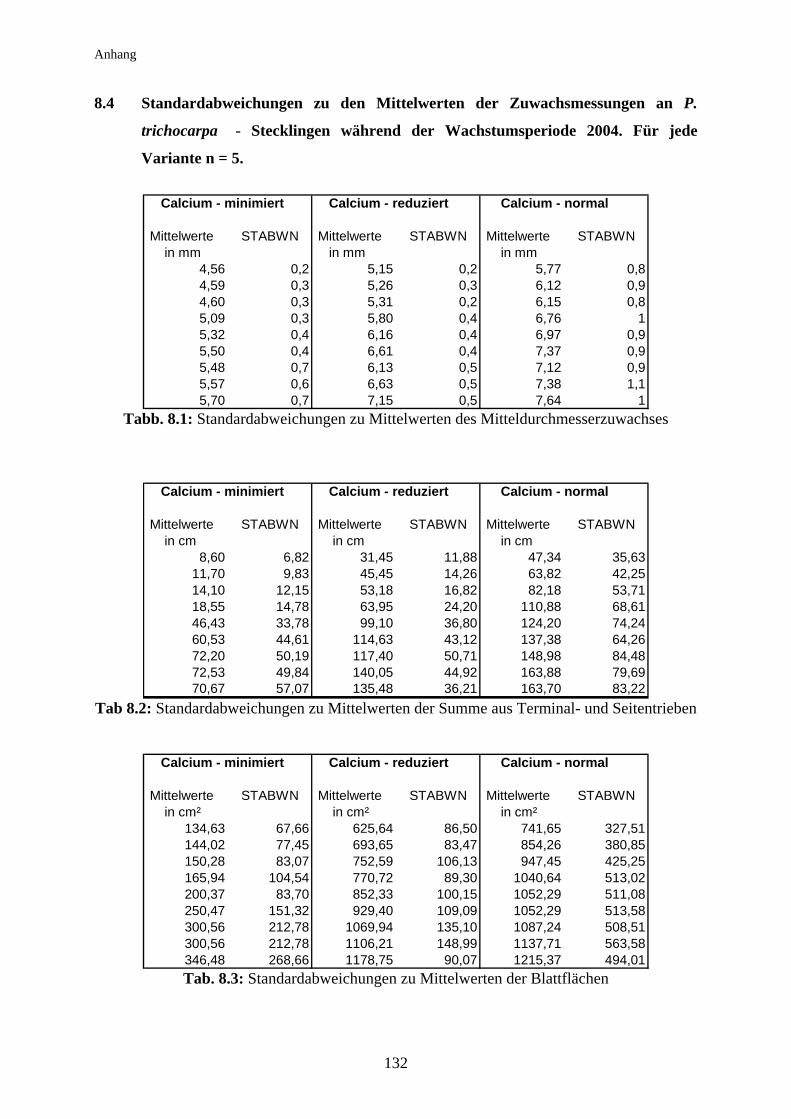
Anhang
132
8.4 Standardabweichungen zu den Mittelwerten der Zuwachsmessungen an P.
trichocarpa - Stecklingen während der Wachstumsperiode 2004. Für jede
Variante n = 5.
Calcium - minimiert Calcium - reduziert Calcium - normal
Mittelwerte STABWN Mittelwerte STABWN Mittelwerte STABWN in mm in mm in mm
4,56 0,2 5,15 0,2 5,77 0,84,59 0,3 5,26 0,3 6,12 0,94,60 0,3 5,31 0,2 6,15 0,85,09 0,3 5,80 0,4 6,76 15,32 0,4 6,16 0,4 6,97 0,95,50 0,4 6,61 0,4 7,37 0,95,48 0,7 6,13 0,5 7,12 0,95,57 0,6 6,63 0,5 7,38 1,15,70 0,7 7,15 0,5 7,64 1
Tabb. 8.1: Standardabweichungen zu Mittelwerten des Mitteldurchmesserzuwachses
Calcium - minimiert Calcium - reduziert Calcium - normal
Mittelwerte STABWN Mittelwerte STABWN Mittelwerte STABWN in cm in cm in cm
8,60 6,82 31,45 11,88 47,34 35,6311,70 9,83 45,45 14,26 63,82 42,2514,10 12,15 53,18 16,82 82,18 53,7118,55 14,78 63,95 24,20 110,88 68,6146,43 33,78 99,10 36,80 124,20 74,2460,53 44,61 114,63 43,12 137,38 64,2672,20 50,19 117,40 50,71 148,98 84,4872,53 49,84 140,05 44,92 163,88 79,6970,67 57,07 135,48 36,21 163,70 83,22
Tab 8.2: Standardabweichungen zu Mittelwerten der Summe aus Terminal- und Seitentrieben
Calcium - minimiert Calcium - reduziert Calcium - normal
Mittelwerte STABWN Mittelwerte STABWN Mittelwerte STABWN in cm² in cm² in cm²
134,63 67,66 625,64 86,50 741,65 327,51144,02 77,45 693,65 83,47 854,26 380,85150,28 83,07 752,59 106,13 947,45 425,25165,94 104,54 770,72 89,30 1040,64 513,02200,37 83,70 852,33 100,15 1052,29 511,08250,47 151,32 929,40 109,09 1052,29 513,58300,56 212,78 1069,94 135,10 1087,24 508,51300,56 212,78 1106,21 148,99 1137,71 563,58346,48 268,66 1178,75 90,07 1215,37 494,01 Tab. 8.3: Standardabweichungen zu Mittelwerten der Blattflächen

Anhang
133
8.8 Abbildungsverzeichnis
Abb. 1.1: Schematische Darstellung der Ca2+-Verteilung in benachbarten Zellen........... 3
Abb. 1.2: Schematische Darstellung eines Aktionspotentials an der Plasmamembran
von Chara............................................................................................................... 6
Abb. 1.3: Lichtmikroskopischer Querschnitt durch einen Pappelstamm
im Sommerzustand.................................................................................................. 9
Abb. 2.1: Sterilkultur P. tremula L. x P. tremuloides Michx............................................ 11
Abb. 2.2: Anzucht von P. tremula x P.tremuloides unter verschiedenen
Ca-Regimes in Hydrokultur..................................................................................... 14
Abb. 2.3: Im Freiland angezogene Populus trichocarpa-Pflanzen..................................... 16
Abb. 2.4: Für die Faserlängenmessung mazerierte Holzfasern........................................... 18
Abb. 2.5: Auswertungsbeispiel der energiedispersiven Röntgenspektren
durch Bestimmung des peak : background Verhältnisses........................................ 23
Abb. 2.6: Schematische Darstellung der Membranpotentialmessungen im
Phloem des Blattleitbündels unter Anwendung der Aphidentechnik....................... 25
Abb. 2.7: Pappelblatt, in die Küvette des Porometermeßkopfs eingespannt...................... 26
Abb. 2.8: IMAGING-PAM Versuchsaufbau............................................................................. 29
Abb. 3.1: P. tremula x P. tremuloides in Hydrokultur unter verschiedener
Ca2+-Versorgung angezogen….................................................................................. 30
Abb. 3.2 A-D: P. tremula x tremuloides zu Beginn, nach zwei und
nach sechs Wochen Wachstum in Hydrokultur......................................................... 31
Abb. 3.3: Durchmesserzuwachs von P. trichocarpa während einer
Vegetationsperiode unter verschiedener Ca2+-Versorgung....................................... 34
Abb. 3.4: Trieblängenzuwachs von P. trichocarpa während einer
Vegetationsperiode unter verschiedener Ca2+-Versorgung....................................... 34
Abb. 3.5: Blattflächenzuwachs von P. trichocarpa während einer
Vegetationsperiode unter verschiedener Ca2+-Versorgung....................................... 35
Abb. 3.6 A-C: P. trichocarpa - Stecklinge im Juli, August und September........................ 36
Abb. 3.7: Relativer Gehalt an Calciumionen im Leitbündel von Pappelblättern................. 38
Abb. 3.8: Relativer Gehalt an Calciumionen im Phloem, im Cambium
und in der Differenzierungszone des Xylems im Stammbereich der Pappel........... 39

Anhang
134
Abb. 3.9: Relativer Gehalt an Calciumionen im Zentralzylinder
und im Cortex der Feinwurzel................................................................................... 40
Abb. 3.10: Wurzelquerschnitt unter 0 mM und unter 5 mM Ca2+....................................... 41
Abb. 3.11: Blattquerschnitt unter 0 mM und unter 5 mM Ca2+........................................... 42
Abb. 3.12: Querschnitt durch Pappelstamm unter 0 mM und unter 5 mM Ca2+................. 43
Abb. 3.13: Veränderungen der Gefäßquerschnittsflächen in Abhängigkeit
des Ca2+-Gehalts....................................................................................................... 44
Abb. 3.14: Abhängigkeit der Faserlänge von der Ca2+-Ernährung...................................... 45
Abb. 3.15: Häufigkeitsverteilung der Gefäßquerschnittsfläche........................................... 46
Abb. 3.16: Einfluss der Calciumernährung auf die Ultrastruktur cambialer Zellen............. 48
Abb. 3.17: Einfluss der Calciumernährung auf die Ultrastruktur der Blattadern................. 49
Abb. 3.18: Einfluss der Calciumernährung auf die Ultrastruktur der Mesophyllzellen....... 50
Abb. 3.19: Vergleichende Darstellung zweier FTIR-Spektren
von Holzproben im Wellenbereich 4000 cm-1 bis 450 cm-1...................................... 51
Abb. 3.20: Vergleichende Darstellung zweier FTIR-Spektren
von Holzproben im Wellenbereich 1800 cm-1 bis 800 cm-1....................................... 52
Abb. 3.21: Vergleichende Darstellung zweier FTIR-Spektren
von Holzproben im Wellenbereich 1800 cm-1 bis 800 cm-1...................................... 53
Abb. 3.22: Vergleichende Darstellung zweier FTIR-Spektren
von Rindenproben im Wellenbereich 4000 cm-1 bis 450 cm-1.................................... 54
Abb. 3.23: Experimenteller Versuchsaufbau zur Messung der elektrischen Potentiale......... 55
Abb. 3.24: Reizung der Blattspitze von mit Eiswasser.......................................................... 56
Abb. 3.25: Reizung am Spross mit Eiswasser........................................................................ 57
Abb. 3.26: Beispielhafte Darstellung einer Membranpotentialmessung im Mesophyll........ 57
Abb. 3.27: Feuerreiz an der Blattspitze.................................................................................. 58
Abb. 3.28: Feuerreiz an der Blattbasis von Blatt 4................................................................ 59
Abb. 3.29: Beispielhafte Darstellung des Versuchsaufbaus bei
Distanzsignalmessungen im Phloem mit angelegtem Kälteblock.............................. 60
Abb. 3.30: Acropetale Signalleitung nach Feuerreiz mit Kälteblock.................................... 61
Abb. 3.31: Basipetale Signalleitung durch Feuerreizung nach TEA+-Behandlung............. 62
Abb. 3.32: Basipetale Signalleitung nach Feuerreiz an Ca2+-mangelversorgten Pappeln..... 63
Abb. 3.33: Acropetale Signalleitung nach Feuerreiz an Ca2+-mangelversorgten Pappeln..... 63
Abb. 3.34: Schließzellenanatomie von P. tremula x P. tremuloides...................................... 64
Abb. 3.35: EDX-Analyse an Pappelschließzellen.................................................................. 65

Anhang
135
Abb. 3.36: Relative Kalium- und Chloridgehalte der Stomata im
geöffneten und im geschlossenen Zustand............................................................... 66
Abb. 3.37: Einfluss von basipetal gerichteten Kurzstreckensignalen aus Hitzereizung
auf den Blattgaswechsel............................................................................................ 68
Abb. 3.38 : Einfluss von acropetal gerichteten Distanzsignalen aus Hitzereizung
auf den Blattgaswechsel.............................................................................................. 69
Abb. 3.39: Interne CO2-Konzentration bei Kurzstreckensignalen sowie bei
Distanzsignalen aus Hitzereizung............................................................................... 70
Abb. 3.40: Auswirkung eines Kälteblocks auf den Gaswechsel bei acropetal
gerichteten Distanzsignalen aus Hitzereizung............................................................. 71
Abb. 3.41: Einfluss eines acropetal gerichteten Distanzsignals aus Hitzereizung auf
den Blattgaswechsel von normal versorgten Pappeln aus einer Hydrokultur ............ 72
Abb. 3.42: Einfluss eines acropetale gerichteten Distanzsignals aus Hitzereizung auf
den Blattgaswechsel von minimal mit Calcium versorgten Pappeln aus
einer Hydrokultur........................................................................................................ 73
Abb. 3.43: Veränderungen in der photosynthetischen Quantenausbeute
des PSII auf basipetale Kurzstreckensignale aus Hitzereizung.................................. 75
Abb. 3.44: Veränderungen in der photosynthetischen Quantenausbeute
des PSII auf acropetale Distanzsignale aus Hitzereizung........................................... 76
Abb. 3.45: Kinetik der Veränderungen in der photosynthetischen Quantenausbeute
des PSII zwischen Blattadern und Intercostalfeldern................................................ 77
Abb. 5.1: Schematische Darstellung der elektrischen Signalleitung im Baum................... 97
8.9 Tabellenverzeichnis
Tab. 2.1: Zusammensetzung der in ihrem Ca2+-Gehalt modifizierten Nährlösungen
der ersten beiden Versuchsreihen............................................................................... 15
Tab. 2.2: Zusammensetzung der in ihrem Ca2+-Gehalt modifizierten Nährlösungen
der dritten Versuchsreihe............................................................................................ 15
Tab. 2.3: Auswahl an spezifischen FTIR-Absorptionsbanden und den
entsprechenden funktionellen Gruppen...................................................................... 21
Tab. 4.1: Durchschnittliche Länge cambialer Initialzellen und Xylemfasern
ausgewählter Laubbaumarten...................................................................................... 81

Anhang
136
8.10 Veröffentlichungen
Fromm, J., Rockel, B., Lautner, S., Windeisen, E., Wanner, G. (2003) Lignin distribution in
wood cell walls determined by TEM and backscattered SEM techniques. Journal of Structural
Biology 143: 77-84
Langer, K., Levchenko, V., Fromm, J., Geiger, D., Steinmeyer, R., Lautner, S., Ache, P.,
Hedrich, R. (2004) The poplar K+ channel KPT1 is associated with K+ uptake during stomatal
opening and bud development. The Plant Journal 37: 828-838
Lautner, S., Grams, T.E.E., Matyssek, R., Fromm, J. (2005) Characteristics of electrical
signals in poplar and responses in photosynthesis. Plant Physiology 138: 2200-2209
Fromm, J., Lautner, S. (2005) Characteristics and functions of phloem-transmitted electrical
signals in higher plants. In: Communication in Plants - Neuronal Aspects of Plant Life. F.
Baluska, S. Mancuso, D. Volkmann (Eds.). Springer-Verlag, Heidelberg, im Druck





























