Embed Size (px)
Citation preview

bcr-abl-Induced Cell Lines Can Switch From Mast Cell to Erythroid or Myeloid Differentiation In Vitro
By Andrew G. Elefanty and Suzanne Cory
The chimericbcr-ablgene formed by the Philadelphia translo- cation is thought to initiate chronic myeloid leukemia. Engraft- ment of mice with bone marrow cells infected with a bcr-ab1 retrovirus has been shown to elicit multiple hematopoietic disorders, including a clonal but nontransplantable hyperpro- liferation of erythroid and/or mast cells. Culture of spleen and bone marrow cells from such mice usually yielded mast cell lines, even when erythroid disease dominated the pri- mary animal. The mast cells, which carried the same proviral insert as the primary disease, generally grew slowly and were neither transplantable nor clonogenic in agar until they had been cultured for several months. Unexpectedly, several bcr-ab/-induced lines switched in vitro from mast cell to
HRONIC MYELOID leukemia (CML) is a clonal C stem cell malignancy' typically distinguished by the Philadelphia chromosome (Ph')? a shortened chromosome 22 that results from a reciprocal 9;22 transl~cation.'.~ The translocation juxtaposes the N-terminal region of the bcr gene (on chromosome 22) and the body of the c-ab1 proto-oncogene (from chromosome 9), thereby creating a hybrid bcr-ab1 gene encoding a 210-Kd fusion polypeptide? The bcr-ab1 gene created by a variant form of the transloca- tion confined to Ph'-positive acute lymphoblastic leukemia (ALL) involves a smaller bcr moiety and encodes a polypep- tide of 190 Kd.6 The heightened tyrosine kinase activity of the bcr-ab1 proteins7 is believed to be the key feature inducing disease.
The chronic phase of CML, which can be regarded as a preleukemia, initially exhibits an excess of maturing my- eloid elements in the blood and hematopoietic organs. After 3 to 5 years the disease accelerates and eventually culminates in a fatal blast crisis, usually an acute myeloid or lymphoid leukemia, although erythroid, megakaryoblastic, and basophilic variants occur infrequently.'~~ The inability of chronic-phase CML cells to be maintained in culture has limited analysis of the events involved in progression to blast crisis. The existing CML cell lines, in fact, derive from patients in the terminal blast phase."
Early studies of the effect of expressing a bcr-ab1 cDNA" in hematopoietic cells established that immortalised cell lines were relieved of their previous growth factor require- ment and rendered tumorigeni~.'~"~ Low-level growth fac- tor production by one such line raised the possibility that ab1 oncogenes facilitate establishment of an autocrine l00p.'~ Most attempts to mimic CML, either in vitro by infecting long-term murine bone marrow cultures with a bcr-ab1 re t rovi r~s , '~~ '~ or in vivo through bcr-ab1 transgenic
yielded only lymphoid disease. A more promising approach has been to transplant lethally irradiated mice with bone marrow cells infected with a bcr-ab1 retrovirus. A high incidence of hematopoietic disease was o b s e ~ e d . ' ~ - ~ ' A few mice exhibited the granulocytosis expected of CML:0321 but most developed macrophage, erythroid, mast cell, or lymphoid tumors.
This report focuses on mast cell lines established from mice with erythroid and/or mast cell disease. Many were
megakaryocytic and/or erythroid character, and one became myeloid. The dramatic phenotypic shifts seem likely to involve changes occurring within progenitor cells maintain- ing the clone, rather than mutation of mature mast cells. The variant lines exhibited substantial spontaneous differentia- tion, despite being readily transplantable and therefore fully transformed. The production of hematopoietic growth fac- tors by the mast cell lines and their phenotypic variants may implicate an autocrine loop in their evolution. These novel bcr-ab/ cell lines should aid in the study of genetic events in the progression from chronic to acute leukemia and facilitate analysis of hematopoietic lineage commitment. o 1992 by The American Society of Hematology.
not malignant until they had been cultivated in vitro for several months. Surprisingly, several clonal bcr-abl-express- ing cell lines switched in vitro from mast cell to erythroid or myeloid differentiation. The implications of these findings for CML are discussed and hypotheses to explain the "lineage switching" proposed.
MATERIALS AND METHODS
Establishment of Cell Lines Bone marrow or spleen cell suspensions were seeded at 1 to 2 x
lo6 cells/mL in 10 mL Dulbecco's modified Eagle's medium containing 20% fetal calf serum and 0.05 mmol/L 2-mercaptoetha- no1 in 80 cm2 flasks (Nunc, Kamstrup, Denmark) and incubated at 37°C in a humidified atmosphere of 10% CO, in air. After 5 to 7 days, or sooner if cell proliferation was rapid, 5 to 10 mL of fresh medium was added. Thereafter, half medium changes were per- formed at least weekly. Cell lines that were 'slow' growing had doubling times of 2 to 7 days while 'fast' growing cell lines doubled every 1 to 2 days. Cell lines were frozen and stored in liquid nitrogen at regular intervals, especially after phenotypic 'switch- ing,' to circumvent the problem of dedifferentiation that occurred with continuous culture of these lines.
Analysis of DNA and RNA
Nucleic acid isolation and fractionation was as described previ- o ~ s l y . ~ The probes used for blot analysis included the following: IgE receptor, the 1,036-bp EcoRI fragment of the rat a subunit of the high-affinity IgE receptor (clone Edp 16)*3; myeloperoxidase,
From The Walter and Eliza Hall Institute of Medical Research, Post
Submitted July 23,1991; accepted October 16, 1991. Supported by the National Health and Medical Research Council of
Australia and the US National Cancer Institute (CA12421). A.G.E. is a National Health and Medical Research Council Postgraduate Scholar.
Address reprint requests to Suzanne Cory, PhD, The Walter and Eliza Hall Institute of Medical Research, Post Ofice Royal Melbourne Hospital, Victoria, 3050, Australia.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section I734 solely to indicate this fact.
office Royal Melbourne Hospital, victoria, Australia.
0 I992 by The American Society of Hematology. OOO6-4971/92/ 7905-0Oo9$3.OO/O
Blood, Vol 79, No 5 (March 1). 1992: pp 1271-1281 1271
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

1272 ELEFANTY AND CORY
the 1.9-kb Pst I fragment of the murine cDNA clone mMP06”; p53,0.4-kb and 0.87-kb Pst I fragments of the murine cDNA clone pp53-176=; and scl, the 1.9-kb EcoRI fragment of the murine cDNA, corresponding to nucleotides 406 to 2335.% The origin of other probes is described el~ewhere.’~~~’ Probes were labeled with [a-”PIdATP using a random hexamer priming kit (Bresatec Lim- ited, Adelaide, South Australia).
Hematopoietic Assays Conditioned medium collected from dense cell cultures was
concentrated 10-fold in an ultrafiltration cell (model 8050; Amicon, Danvers, MA) and filtered before use. Granulocye-macrophage colony-stimulating factor (GM-CSF) and interleukin-3 (IL-3) were assayed using FDC-PI cells (responsive to GM-CSF and IL-3) and 32D clone 23 cells (responsive to IL-3) as described previously.” Erythropoietin (Epo) was assayed on fetal liver cells using day-2 colony-forming unit-erythroid (CFU-E) as a read0ut.2~ Levels of CSF were converted to bone marrow units’” by comparison with purified recombinant GM-CSF and IL-3 of known activity. Colony- forming cells were assayed by culturing cell lines at a range of concentrations in triplicate 1 mL agar cultures, with and without addition of indicated growth factors for 7 to 14 days at 37°C as described previously.” Recombinant murine IL-3, GM-CSF, gran- ulocyte-CSF (G-CSF), and leukemia inhibitory factor (LIF) were generously provided by Dr D. Metcalf, and recombinant human Epo by Dr A.W. Burgess.
Detection of Cell Su~ace Antigens Cells were stained with the monoclonal antibodies (MoAbs)
M1/70 (aMac-l),SZ RB6-8C5 (‘8C5’/aGr-l),” F4/80,” and TER- 119”; each had been directly conjugated to fluorescein isothiocy- anate (FITC) or phycoerythrin (PE) (Southern Biotechnology Associates, Birmingham, AL), or biotinylated and revealed by an FITC- or PE-streptavidin second stage. All staining was performed in the presence of excess unlabeled anti-Fcy receptor antibody 2.4G2% to reduce nonspecific binding of antibodies, and cells were analyzed on a FACScan (Becton Dickinson, Sunnyvale, CA).
Cytochemical Stains
May-Grunwald-Giemsa, Lux01 Fast Blue, Astra Blue, and stain- ing for a-naphthyl acetate esterase, myeloperoxidase, and acetylcho- linesterase were performed on cytocentrifuge preparations or fixed agar colonies according to standard protocol^.^' Detection of the pseudoperoxidase activity of hemoglobin in agar colonies or cell suspensions was with benzidine HCl” or 2,7-diaminofl~orene.’~
RESULTS
Erythroid and mixed erythroid/mast cell hyperplasia was a frequent cause of morbidity in mice we reconstituted with bcr-abl virus-infected bone marrow cellsI9 (Elefanty and Cory, manuscript submitted). To further characterize this syndrome, we sought to establish cell lines by culturing the bone marrow and spleen cells of 19 mice in the absence of added growth factors. Twelve of the 15 lines that were derived (Table 1) were mast cell in origin; three grew rapidly from the outset but the majority grew very slowly, doubling every 2 to 7 days. The proviral insertion pattern shown by Southern blot analysis established that all but one of the cell lines were clonal and, in 9 of 10 cases, the pattern obtained from the cell line was identical to that obtained from the tissue of origin. Because six of the seven mast cell
lines in this analysis derived from tissue infiltrated predom- inantly by erythroid cells, both populations probably de- scended from a common infected prec~rsor.’~
Six mast cell lines were maintained continuously in vitro for several months. Remarkably, they proved to be pheno- typically unstable (Table 2), most evolving towards eryth- roid/megakaryocytic differentiation. After cloning in agar, the phenotypic variants from all six lines were shown by proviral insertion analysis to derive from the same infected precursor as the initial mast cell line” (this report and unpublished results). The history of one erythroid/ megakaryocytic line (96.4) will be described in detail, as will that of an exceptional line (34.6) that became myeloid. The evolution of the bcr-ab1 lines towards a fully malignant phenotype will also be considered.
The 96.4 Line: A Switch From Mast Cell to Erythroid and Megakaryocytic Differentiation
Clonal phenotypic variants. Figure 1A summarizes the evolution of the 96.4 mast cell line, which was obtained from the spleen of a BALB/c mouse with erythroid, mast cell, and macrophage disease (Elefanty and Cory, manu- script submitted). The slow-growing line was independent of the exogenous IL-3 required by normal mast cells, but may have been provided with growth factors by the stromal component present during the first few weeks of culture. After about 3 months, a rapidly growing subline of mast cells (designated 96.43Mst) arose in one of several cultures grown in parallel. The cells in another flask of the slow- growing line (designated 96.42Mst) underwent a dramatic change after another 2 months. Within 1 week, the culture became pleomorphic (compare Fig 1B with 1C and D) and cytospin preparations (Fig 1E and F) showed that the mast cells had been replaced by maturing erythroid cells and megakaryocytes that stained with acetylcholinesterase (not shown). Most cells (70%) in this subline (96.42EMeg) contained hemoglobin, as judged by staining with benzidine or diaminofluorene and over 50% reacted strongly with the erythroid-specific MoAb TER-119.35 After 2 months, one 96.42EMeg culture yielded a subline (96.42Ery) that re- tained erythroid character but exhibited few mature mega- karyocytes.
Evidence that the mast cell, erythroid/megakaryocytic, and erythroid subclones all carried the same proviral insert as the original splenic tissue and hence derived from a single infected precursor was provided by Southern blot analysis for provirus-host DNA junction fragments in EcoRI digests (Fig 2A) and Hind111 digests (not shown). RNA analysis (Fig 2B) established that the bcr-ab1 gene was expressed and confirmed the lineage assignments of the cell lines. Expression of the a chain of the high-affinity IgE receptor’’ and c-kit, the receptor for mast cell growth factor (MGF)? was detectable only in the mast cell lines (lanes 1 and 2), while p globin and Epo receptor expression was found only in sublines containing erythroid cells (lanes 3 and 4). Two genes encoding transcription factors impli- cated in erythroid differentiation, GATA-1 and scl/tal, were expressed in the mast, erythroid, and erythroid/
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

bcr-ab/ MAST CELLS SWITCH DIFFERENTIATION 1273
Table 1. Derivation of Cell Lines From Primary bcr-ab/ Tumors
Primary Disease
Latency Clonal Mouse Strain (wk)* Pathologyt Transplantation* Cell Llne5 Relationship11
33.3 C57BL/6 46 Ery, Mst + ** Erythroid [f] I 55.3 BALB/c 22 Ery, Mst ND Erythroid [fl I 52.10 BALB/c 30 Ery, Mst +tt Erythroid [f] I 34.4 C57BL/6 49 Mst + Mast [fl I 45.14 DBA/2 16 Ery, Mst + Mast [fl I 99.4 BALB/c 24 Ery + ** Mast [fl D 32.1 C57BL/6 8 Ery, Mst - Mast [SI ND 33.1 C57BL/6 7 Ery Mast [SI ND 33.2 C57BL/6 7 Ery, Tn Mast [SI ND
- Mast [SI ND 34.6 C57BLl6 33 Ery 34.8 C57BLl6 22 Tll Mast [SI I 45.1 1 DBA/2 3 ErY -tt Mast [SI I 96.4 BALBIc 11 Ery, Mst +# Mast [SI I 52.5 BALB/c 11 Ery, Mst Mast [SI I 59.17 BALB/c 14 Ery, Mst ND Mast [SI I
-
-
-
-
Abbreviation: ND, not done. *Time elapsed (in weeks) between transplantation of irradiated mouse with bcr-ab/ virus-infected bone marrow cells and autopsy. tHistopathology of primarybcr-ab/ mice that exhibited erythroid (Ery) and mast cell (Mst) diseaselg (Elefanty and Cory, manuscript submitted). Two
of these mice also had a lymphoid (T) tumor component and all had a variable amount of macrophage tumor that, at least in some cases, derived from the same infected progenitor as the erythroid/mast cell t ~ m o r . ’ ~
STransplantability of the primary tumor was usually assessed by injecting lo6 spleen cells intraperitoneally into 2 to 6 sublethally irradiated (4.5 Gy) or nonirradiated (two daggers) recipients, except for those (two asterisks) that were tested as cell lines within 25 days of initiating in vitro culture. Recipients were monitored for tumors for 12 months; +,transplantable; -, not transplantable.
§Cell type in primary line derived by culturing spleen or bone marrow cells of sick mice. [fl or [SI indicates fast or slow growth. IlRetroviral integration pattern of cell line determined by Southern blotting (see Fig 2A) to be identical (I) or different (D) to that of tissue of origin or,
for mouse 34.8, of tumor deposits elsewhere in the primary mouse. ND, not done due to insufficient material; these four animals had oligoclonal tumors at other sites.
llA clonal T-lymphoid thymoma was present that derived from the same progenitor (mouse 33.2) or a different progenitor (mouse 34.8) to the macrophage or mast cell disea~e.’~
#Four of six recipients developed tumors following a long latency (6 months).
Table 2. Phenotypic Shifts of bcr-ab/ Mast Cell Lines
Cell Line*
96.4Mst 34.6Mst
34.4Mst 45.14Mst 52.5Mst 99.4Mst
Culture Period (mo)t
5 3
12 6
1 II 1.511
6 6
Secondary Lineage*
Erythroid/megakaryocytic Erythroid/mast cell Erythroid§ Myeloid§ Erythroid/megakaryocytic Erythroid/megakaryocyticT Erythroid/megakaryocytic# Erythroid/mast cell
*Six other mast cell lines did not demonstrate a phenotypic shift in
tTotal period in culture (in months) before the observed shift. Sln all cases, agar cloned cell lines of the secondary lineage harbored
identical proviral integrations to the primary cell line (Figs 2 and 5, Elefanty et aI.,”and data not shown).
vitro, but they were only cultured for 1 to 4 months.
§These lines evolved from the erythroid/mast cell line. IlAlthough mast cells dominated the primary cultures from these
mice, some erythroblasts persisted from the outset. BPreviously described,” but the megakaryocytic component was not
initially appreciated. #Rare clones obtained after infection of the 52.5Mst line with a
retrovirus expressing a mutated p53 cDNA; whether p53 played any role is obscure.
megakaryocytic variants, consistent with previous observa- t i o n ~ . ~ ’ . ~ ~
Four of five 96.4 mast cell sublines expressed GM-CSF RNA (eg, Fig 2B, lane 2) and, although no IL-3 (or MGF) RNA was detectable, more sensitive bioassays (see Materials and Methods) showed that the three clones tested secreted low levels of IL-3 (1 to 5 U/mL) as well as GM-CSF (5 to 900 U/mL). All three erythroid/megakaryocytic and both erythroid subclones tested secreted GM-CSF (5 to 100 U/mL). A low level of Epo bioactivity (0.05 U/mL) was detected in the single erythroid clone tested, but not in an erythroid/megakaryo- cytic clone.
The range of differentiation stages detectable in the 96.42EMeg and 96.42Ery lines was impressive. Both produced macroscopic colonies of erythroblasts in the spleen of lethally irradiated syngeneic mice 12 to 14 days after injection. Spleen colony formation, a property displayed by some early stem cells (CFU-S), was not obtained with the other bcr-ab1 erythroid cell line (33.3Ery) tested. When cultured in semi-solid agar, the 96.42 erythroid lines yielded diverse colony types (Table 3). The EMeg line produced large compact colonies contain- ing blast cells (some colonies were white, others red due to a large erythroid component), as well as small single-
Factor production.
Differentiation capacity of the 96.4 variants.
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

1274 ELEFANTY AND CORY
MOUSE 96.4 96.42EMeg c erythroid, mast &
tumor \
- macrophage spleen (SIOW)
culture
Fig 1. Differentiation of 96.4 cell lines. (A) In vitro history (see text). The interval between stages (boxed) is indicated in months. (B through D) Appear- ance in culture of (B) 96.42Mst (8 months) (C) 96.42EMeg
* (5 months) (original magnifica- tion of both x 125). and (0 ) mega- karyocyte in 96.42EMeg culture (original magnification x400). (E through G) Cytocentrifuge prepa- rations stained with May-Grun- wald-Giemsa of (E) 96.4Mst showing granulated mast cells (5 weeks) and IF) 96.42EMeg showina meaakarvocvte and iY
lineage or mixed colonies of maturing erythroid cells and megakaryocytes (Fig 3). In contrast, the Ery subline yielded fewcr megakaryocytic colonies and many more overtly hemoglobinized colonies (Table 3). Only the white compact colonies retained a high proliferative potential upon replat- ing (data not shown). Addition to the agar cultures of IL-3 or Epo (Table 3) or GM-CSF (not shown) affected neither the plating efficiency nor the distribution, size, or morphol- ogy of colonies.
The 96.4 line did not readily switch its differentiation pattem. Despite the phenotypic shift just described, the 96.4 primary mast cell line was not intrinsically unstable in vitro. After being maintained for 8 months in liquid culture, 96.42Mst cells could be cloned in agar, but only on onc occasion over the next 5 months were erythroid colonies detected amongst the mast cell colonies. These were established as EMeg lines that displayed the same proviral insert as all other 96.4 sublines (Fig 2A, lane 5), but the liquid culture from which they derived was not overtaken by erythroid cells, perhaps because the mast cells themselves now grew more rapidly or because the second EMegvariant was not as competitive as the earlier one.
Because the demethylating agent 5-azacytidine has been
" "- , , . .
erythroid differentiation with ma- turing normoblasts and an eryth- rocyte (arrowed) (5 months) (original magnification x500). (G) Peripheral blood film from an erythroleukemic recipient of 96.42EMeg (5 months) (original magnification x500). Note the maturity of the circulating eryth- roblasts. The time indicated in parentheses in this and all subse- quent figures refers to the total period in culture at the time of analysis.
shown to induce a lymphoid to macrophage switch in an Abelson-transformed ccll line,M we attempted to induce a mast to erythroid/megakaryoqtic switch by incubating three 96.42 Mst clones with up to 6 pmol/L 5-azacytidine for 1 to 5 days. As only one treated culture acquired a small number ( - 12%) of benzidine-staining cells, we infer that the genetic change(s) required to permit erythroid/ megakaryocytic differentiation of the 96.42Mst progenitor cell is not readily induced by demethylation.
The 34.6 Line: A Switch From Mast Cell to Erythroid and Myeloid Differentiation
The 34.6 mast cell line (34.6Mst) was derived from the near-normal bone marrow of a C57BL/6 mouse with a modest hcr-ahl-induced erythroid and macrophage tumor burden." As outlined in Fig 4A, prolonged culture of this line produced both an erythroid and a myeloid variant. Once again, Southern blotting established that the sub- cloned phenotypic variants and the primary mast cell line derived from a single infected precursor (Fig 5A).
After 3 to 4 months in culture, the cells became smaller, generally exhibiting a morphology intermediate between that of an erythroid and
Evolution of the erythroid variant.
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom
bcr-ab/ MAST CELLS SWITCH DIFFERENTIATION
B
1275
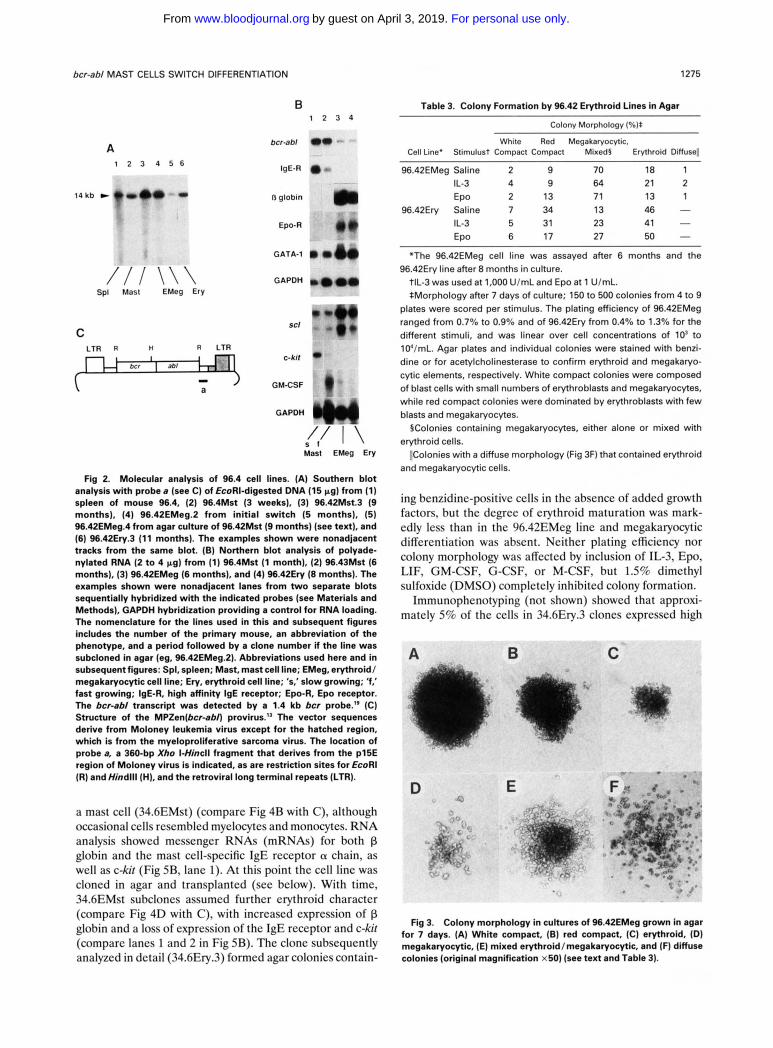
Table 3. Colony Formation by 96.42 Erythroid Lines in Agar
A 1 2 3 4 5 6
/ / I \ \ \ SpI Mast EMeg Ery
C
bcr-abl
IgE-R
13 globln
Epo-R
GATA-1
GAPDH
SCl
LTR R R LTR C-klt
GYCSF
GAPDH
1 2 3 4
S f / T I \ Mast EMeg Ery
Fig 2. Molecular analysis of 96.4 cell lines. (A) Southem blot analysis with probe 8 (see C) of EcoRI-digested DNA (15 pg) from (1) spleen of mouse 96.4, (2) 96.4Mst (3 weeks), (3) 96.42Mst.3 (9 months), (4) 96.42EMeg.2 from initial switch (5 months), (5) 96.42EMeg.4 from agar culture of 96.42Mst (9 months) (see text), and (6) 96.42Ery.3 (11 months). The examples shown were nonadjacent tracks from the same blot. (B) Northern blot analysis of polyade- nylated RNA (2 to 4 pg) from (1) 96.4Mst (1 month), (2) 96.43Mst (6 months), (3) 96.42EMeg (6 months), and (4) 96.42Ery (8 months). The examples shown were nonadjacent lanes from two separate blots sequentially hybridized with the indicated probes (see Materials and Methods), GAPDH hybridization providing a control for RNA loading. The nomenclature for the lines used in this and subsequent figures includes the number of the primary mouse, an abbreviation of the phenotype, and a period followed by a clone number if the line was subcloned in agar (eg, 96.42EMeg.2). Abbreviations used here and in subsequent figures: Spl, spleen; Mast, mast cell line; EMeg, erythroid/ megakaryocytic cell line; Ery, erythroid cell line; 's,'slow growing; 'f,' fast growing; IgE-R, high affinity IgE receptor; Epo-R, Epo receptor. The bcr-ab/ transcript was detected by a 1.4 kb bcr probe.19 (C) Structure of the MPZen(bCr-8bn provirus." The vector sequences derive from Moloney leukemia virus except for the hatched region, which is from the myeloproliferative sarcoma virus. The location of probe a, a 360-bp Xho I-Hincll fragment that derives from the p15E region of Moloney virus is indicated, as are restriction sites for EcoRl (R) andHindlll (HI, and the retroviral long terminal repeats (LTR).
a mast cell (34.6EMst) (compare Fig 4B with C), although occasional cells resembled myelocytes and monocytes. RNA analysis showed messenger RNAs (mRNAs) for both p globin and the mast cell-specific IgE receptor a chain, as well as c-kit (Fig 5B, lane 1). At this point the cell line was cloned in agar and transplanted (see below). With time, 34.6EMst subclones assumed further erythroid character (compare Fig 4D with C), with increased expression of p globin and a loss of expression of the IgE receptor and c-kit (compare lanes 1 and 2 in Fig 5B). Thc clone subsequently analyzed in detail (34.6Ery.3) formed agar colonies contain-
Colony Morphology (%):
White Red Megakaryocytic, Cell Line' Stimulust Compact Compact Mixed5 Erythroid Diffuse11
96.42EMeg Saline 2 9 70 18 1 IL-3 4 9 64 21 2 EPO 2 13 71 13 1
96.42Ery Saline 7 34 13 4 6 - IL-3 5 31 23 41 -
27 50 - 6 17 EPO
"The 96.42EMeg cell line was assayed after 6 months and the
tlL-3 was used at 1,000 UlmL and Epo at 1 UlmL. *Morphology after 7 days of culture; 150 to 500 colonies from 4 to 9
plates were scored per stimulus. The plating efficiency of 96.42EMeg ranged from 0.7% to 0.9% and of 96.42Ery from 0.4% to 1.3% for the different stimuli, and was linear over cell concentrations of lo' to 1O'lmL. Agar plates and individual colonies were stained with benzi- dine or for acetylcholinesterase to confirm erythroid and megakaryo- cytic elements, respectively. White compact colonies were composed of blast cells with small numbers of erythroblasts and megakaryocytes, while red compact colonies were dominated by erythroblasts with few blasts and megakaryocytes.
§Colonies containing megakaryocytes, either alone or mixed with erythroid cells.
I!Colonies with a diffuse morphology (Fig 3F) that contained erythroid and megakaryocytic cells.
96.42Ery line after 8 months in culture.
ing benzidine-positive cells in the absence of added growth factors, but the degree of erythroid maturation was mark- edly less than in the 96.42EMeg line and megakaryocytic differentiation was absent. Ncither plating efficiency nor colony morphology was affected by inclusion of IL-3, Epo, LIF, GM-CSF, G-CSF, or M-CSF, but 1.5% dimethyl sulfoxide (DMSO) completely inhibited colony formation.
Immunophcnotyping (not shown) showed that approxi- mately 5% of the cells in 34.6Ery.3 clones expressed high
Fig 3. Colony morphology in cultures of 96.42EMeg grown in agar for 7 days. (A) White compact, (B) red compact, (C) erythroid, (Dl megakaryocytic, (E) mixed erythroid/megakaryocytic, and (F) diffuse colonies (original magnification x50) (see text and Table 3).
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

ELEFANTY AND CORY 1276
cloned 34.6Ery + A
MOUSE 34.6 erythroid & macrophage BM 3 mo
tumor
34.6MSt 34.6EMst
34.6Myl + culture
Fig 4. Differentiation of 34.6 cell lines. (A) In vitro history (see text). (B through D) Cytocentri- fuga preparations stained with May-Griinwald-Giemsa (MGG) of (B) 34.6Mst (2 months), (C) 34.6EMst (3.5 months), and (D) 34.6Ery (12 months) (original magnification x 500). Appearance in culture of (E) 34.6Ery (12 months) and (F) 34.6Myl (14 months) (original magnification x 125). Cytocentrifuge prepara- tions of 34.6Myl stained (G) with MGG, (H) for myeloperoxidase, ( I ) with MGG after 2 days of induction with 1.5% DMSO, and (J) with MGG after 24 hours of treatment with 16 nmol/L tet- radecanoyl phorbol acetate and overnight incubation with latex beads (0.8 pm; Difco, Detroit, MI) (original magnifications: G, H, 1, x500; J, x800).
levels of TER-119. Curiously, most cells expressed very high levels of F4/80, an antigen not seen in the 96.42EMeg line and previously thought to be macrophage al- though other features suggestive of macrophage diffcrcnti- ation were not present. Attempts to differentiate 34.6Ery.3 cells with 1.5% DMSO in liquid culture showed marked toxicity, but the surviving cells acquired some cytoplasmic granules suggestive of reversion to a mast cell phenotype.
After 2 months in vitro, the mast cell culture generated from the spleen of a mouse transplanted with 34.6EMst cells was monopolized by maturing myeloid cells that grew in tight clumps (Fig 4F), unlike the crythroid/mast ccll or its erythroid derivative (Fig 4E). The line was cloned in agar and designated
A shft to myeloid differentiation.
cytic cells (Fig 4G) that stained strongly for myeloperoxi- dase (Fig 4H). Depending on the clone, 5% to 80% of the cells were cosinophils and their precursors, because Lux01 Fast Blue showed small numbers of emerald green cytoplas- mic granules. Less than 5% of the cells were phagocytic adherent macrophages but - 30% expressed a-naphthyl acetate esterase. RNA analysis showed myeloperoxidase transcripts in 34.6Myl but no detectable mRNA for p globin or the IgE receptor (Fig 5B, lane 3).
34.6MyI cells formed granulocyte-macrophage colonies in agar in the absence of added growth factors. Neither plating efficiency nor colony morphology was affected by the inclusion of IL-3, GM-CSF, G-CSF, M-CSF, Epo, or LIF in the assays (not shown). However, analysis of
34.6Myl. All 12 clones analyzed were dominated by granulo- conditioned medium showed endogenous production of
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

bcr-ab/ MAST CELLS SWITCH DIFFERENTIATION 1277
A 1 2 3
6.5kb ww-w
/ I \ Mast E h My1
1 2 3 4 5 6
bcr-abl 11) @a (c
13 globin
C-kit
MPO
GM-CSF
GAPDH
IgE-R
GAPDH
111 - -
a
Ilo - * // - + + +
EMast Ery Myl +DMSO
Fig 5. Molecular analysis of 34.8 cell liner. (A) Southem blot analysis with probea (see Fig 2C) of EcoRI-digested DNA (15 pg)from (1) 34.6Mst (2.5 months), (2) 34.6Ery.3 (16 months), and (3) 34.6Myl.l (16 months). (B) Northem blot analysis of polyadenylated RNA (2 to 4 pg) from (1) 34.6EMst (3 months), (2) 34.6Ery.3 (13 months), (3) 34.6Myl.l (15 months), and (4 through 6) 34.6Myl.l incubated with 1.5% DMSO for 1,3, or 5 days (16 months). The examples shown were nonadjacent lanes from two separate blots sequentially hybridized with the indicated probes (see Materials and Methods), GAPDH hybridization providing a control for RNA loading. Abbreviations as in Fig 2 and Myl, myeloid cell line; EMst, erythroid/mast cell line; MPO, myeloperoxidase.
GM-CSF (up to 1,400 U/mL) and IL-3 (up to 100 U/mL). In contrast, minimal levels of GM-CSF were detected in the 34.6 mast and erythroid phenotypic variants.
Granulocytic differentiation of 34.6Myl cells was en- hanced by DMSO and sodium butyrate (Fig 41), and 16 nmol/L tetradecanoyl phorbol acetate increased the num- ber of adherent phagocytic macrophages in some clones (Fig 45). DMSO increased Mac-1 expression and most cells (>90%) acquired F4/80, while a subpopulation (5% to 15%) also expressed the 8C5 mature granulocytic marker (data not shown). Granulocytic differentiation was associ- ated with downregulation of myeloperoxidase, as assessed by RNA analysis (Fig SB, lanes 3 through 6) and cytochem- ical staining (not shown), and also with considerable GM-CSF mRNA induction (Fig 5B).
Evolution of the Transformed Phenotype The slowly growing mast cell lines were not malignant
(Table 4) and, with one exception, were derived from animals in which the primary disease, although lethal, was not transplantable (Table 1). Furthermore, although at 18
weeks the still slowly growing 96.42Mst culture contained transplantable cells, the tumor latency was very long (37 weeks) and only after 29 weeks in culture were clonogenic cells detectable in agar. These observations suggest that although bcr-ab1 expression confers a growth advantage in vivo and in vitro, further somatic mutation(s) must accrue to achieve complete transformation. Those cell lines (eryth- roid or mast) that were transplantable from the outset (Table 4), and that derived from transplantable primary tumors (Table l), had presumably already acquired second- ary mutation(s) in vivo.
Because the tumor suppressor gene p53 is mutated in many CML patients in blast crisis:d7 we investigated p53 expression in bcr-ab1 lines that had become malignant. Diminished expression of p53 may have contributed to the malignancy of the 33.3Ery line, because p53 RNA levels were much lower in this line, especially after prolonged culture (Fig 6C, lanes 13 and 14), than in other erythroid lines (Fig 6, lanes 3, 4, 6, 12, 15, and 16). Levels were also low in the myeloid derivative of 34.6 (Fig 6B) and in three of the four mast cell lines examined (Fig 6, lanes 1,2, and 10). Sequence analysis will be required to fully assess the role of p53, because oncogenic changes affecting p53 function frequently involve point mutations" and thus may not be detectable by Northern analysis.
The ets-related transcription factor PU.1, expressed in macrophage and B-lymphoid cells,@' corresponds to a gene previously designated Spi-1 or Sfpi-1 that is silent in normal erythroid cells but frequently activated by proviral insertion in Friend erythroleukemia.m Sfpi-1 expression was readily detectable in several of the erythroid lines (Fig 6, lanes 6, 12,13, and 15) and may therefore have played a role in their neoplastic progression. Other bcr-ab1 erythroid lines, how- ever, lacked Sfii-1 expression (Fig 6, lanes 3,4, and 16).
Differentiation Capacity of the Cell Lines In vivo Histologic analysis of animals transplanted with the
established cell lines permitted assessment of their differen- tiation capacity in vivo (Table 4). Mice injected with 96.42EMeg cells developed erythroleukemia with greatly elevated numbers of circulating erythroblasts (Fig 1G). Although both 96.42Mst and 96.43Mst cells produced disseminated mast cell tumors, all four animals examined also contained areas of erythroid tumor in the spleen. This finding may indicate that the cells had a wider differentia- tion potential in vivo than in vitro, or that the frequency of 'lineage switch' mutations was relatively high in vivo. Within the 34.6 series, the 34.6Myl.l clonc produced disseminated myelomonocytic tumors, while all six recipi- ents of 34.6EMst cells succumbed to various combinations of erythroid, mast cell, and macrophage tumors. Unexpect- edly, the tumors arising after injcction of the relatively undifferentiated erythroid clone 34.6Ery.3 were mast cells, implying that its differentiation potential had also been modified by environmental signals or by mutation.
DISCUSSION
The bcr-abl-expressing mast cell lines characterized here were obtained from mice that had developed erythroid and/or mast cell hyperplasia after lethal irradiation and
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

1278 ELEFANTY AND CORY
Table 4. Clonogenicity and Transplantability of bcr-abl Cell Lines
Transplanted Tumors:
Culture Period Clonogenicity Latency Cell Line (wk)' (%)t Incidence5 lwk)ll Pathologfl
96.4 lines - - 96.4Mst Is] 5 ND 014
96.42Mst Is] 18 213 37 Mst(Ery) 23.29 < 0.002
33 15 96.43Mst [fl 14 ND 313 8 Mst(Ery) 96.42EMeg 24 1 213 11 Ery#
313 (IV) 4 EW 34.6 lines
34.6EMst 15 616 8 Mst, Ery, Mac** 17 10
34.6Ery.3 51 313 7 Mst 56 15
34.6Myl.l 50 45 66 1 /3 44 My1
2/3 (IV) 8 My1 Other lines
- - 33.1Mst Is] 8 ND 013 52.5Mst Is] 5 0/3 - -
8.26 <0.01 42 20
7 5 99.4Mst [q 4 313 3 Mst
33.3Ery 2 1 313 3 EryWst)
Abbreviation: ND, not done. 'Time of continuous culture (in weeks) before transplantation or plating in agar; Is] and (q indicate slow and fast growing mast cell lines. t u p to 10'cells (5 x 10'for 96.42Mst cells) plated in triplicate 1 mL agar cultures were scored for colonyformation at day 7 and 14, and the cloning
efficiency expressed as a percentage of the cells plated. No colonies were observed for 96.42Mst before 33 weeks of culture or for 52.5Mst before 42 weeks of culture. Equivalent results were obtained for mast cell lines cultured with or without IL-3 (1,000 UlmL).
SCeils (lo8) were transplanted intraperitoneally into sublethally (4.5 Gy) irradiated recipients, or nonirradiated mice (boldface), and monitored for up to 12 months. Additional animals were inoculated intravenously (IV) where indicated.
§Number of mice developing tumors versus total number injected. IlMean tumor latency (in weeks) posttransplantation. THistopathology of secondary tumors: Mst, mast cell; Ery, erythroblast; Mac, macrophage; Myl, myeloid. Minor tumor comDonents are indicated
#One of two recipients also had mast cell tumor around the common bile duct. **Previously described."
in parentheses (see text).
A 96.4 lines
1 2 3 4
P53
SfpC1
GAPDH m m w
S 't/ I \ Mast EMeg Ery
B 34.6 lines
5 6 7 8 9
I I + + + EMast Ery Myl+~hlSO
C
10 11 12 13 14 15 16
-e/-- ----- / / I \ I \ \
Mast EMeg Ery EMeg Ery
Fig 6. Northern blot analysis of cell lines harboringbcr-ab/ pro- virus. Polyadenylated RNA sam- ples (2 t o 4 pg) were sequentially hybridized with the indicated probes (see Materials and Meth- ods). Samples for (A) are as for Fig 28. In (E), (5) 34.6EMst (3 months), (6) 34.6Ery.3 (13 months), and (7 through 9) 34.6Myl.l incubated with 1.5% DMSO for 1, 3, or 5 days (16 months). In (C), (10) 52.5Mst (3 months), (11) 99.4Mst (5 weeks), (12) 45.14EMeg.2 (14 months), (13) 33.3Ery (1 week), (14) 33.3Ery.2 (11 months), (15) 34.4EMeg (7 months), and (16) 55.3Ery (2 weeks).
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

bcr-ab/ MAST CELLS SWITCH DIFFERENTIATION 1279
reconstitution with hcr-ah1 virus-infected bone marrow" (Elcfanty and Cory, manuscript submitted). The virus- infected cell responsible for the syndrome appears to have been a progenitor cell with capacity for both mast cell and erythroid differentiation, as judged by clonal analysis of affected tissues and derived cell lines (Elcfanty et all" and this report). The hcr-ah1 mast cell lines grew in the absence of exogenous IL-3, unlike normal mast cells, but initially may have required factors produced by associated stromal cells, eg, the recently identified MGF or Steel factor. They produced significant levels of GM-CSF and IL-3. When first established, most lines were neither transplantable nor clonogenic in soft agar, but transplantable cells emerged after prolonged culture, prcsumably due to additional (undefined) oncogenic mutations. Analogous factor-indc- pendent mast cell lines produced by Abelson virus infection of multilineage or blast cell colonies"." or 18-day fetal liver cells" also secreted multiple growth factors"." and initially required stromal cell support,"." but became fully malig- nant much more rapidly than the hcr-ah1 lines (2 to 3 months after virus infcction v 6 to 20 months). This finding probably reflects differences in the transforming potency of v-uhl and p210 hcr-ah1 proteins.
The striking feature of the hcr-ah1 cell lines was their propensity to spontaneously change hematopoietic lineage on prolonged culture, a property never previously docu- mented for mast cell lines. Several assumed an erythroid or erythroid/megakaryocytic character, but one became my- eloid (Table 2). In each case, the variant line contained the same proviral insert as the parental mast cell line (Fig 2A and SA and data not shown).
Several of the variant sublines exhibited a marked capac- ity to continuously differentiate, despite being highly malig- nant. The 96.4EMeg line and its subclones, for example, yielded mature erythrocytes and megakaryocytes in the absence of added growth factors (Fig 1). Because normal erythroid differentiation requires Epo and/or IL-3, and Epo was detected in the conditioned medium of one hcr-ah1 erythroid clone, an autocrine loop may have figured in their evolution. Intriguingly, spontaneous hemoglobinization is a feature of some erythroid progenitors in the chronic phase of CML.'6 A surprisingly high proportion (5 of 13) of the cell lines established from CML blast crisis patients (eg, K562) exhibit some erythroid and/or megakaryocytic char- acter, but induction by chemical agents is usually required to promote differentiation."' Equivalent lines have not been isolated with Abelson virus; differentiating erythroid colo- nies isolated from infected fctal liver cells could not be established in culture."
The differentiation capacity of the hcr-ah1 myeloid line (34.6Myl) strongly resembled that of the well-studied pro- myelocytic line HL-60 derived from a patient with acute myeloid leukemia.'" Both differentiate spontaneously along neutrophil, eosinophil, and macrophage lines, the degree of differentiation being enhanced by a range of chemical inducers. 34.6Myl cells secrete GM-CSF and IL-3 and an 'autocrinc activity' has been observed in H L d k o n d i - tioned medium.5N None of the existing CML lines shows such a degree of myeloid differentiation and no equivalent
lines have been isolated with Abelson virus. However, one clone of the IL-3-dcpcndcnt line 32D, which was initially characterized as a mast cell line,'" has the capacity to terminally differentiate into granulocytes if IL-3 is replaced by G-CSF."'
The nature of the cells involved in the 'lineage switches' is an intriguing, although unresolved, issue. Figure 7 depicts our current working hypothesis It seems unlikely that mature mast cells are capable of the phenotypic shifts observed, so we presume that the affected cells were precursors present at low concentration in the cultures. These cells may have been committed, unipotential mast cell progenitors or oligopotential cells unable to express their full potential under the growth conditions provided. (The observation that certain lines exhibited a wider differentiation potential in vivo than in vitro may favor the latter hypothesis.) In either case, phenotypic switching in vitro prcsumably resulted from somatic mutation(s) that either provided necessary growth factor(s) or induced key transcriptional programming gene(s). Our failure to mimic such mutation by treatment with 5-azacytidinc suggests that the mechanism of the lineage switch is more complex than activation of a single gene by demethylation.
For 96.42, the switch from mast to erythroid/megakaryo- cytic differentiation was accompanied by transformation, raising the possibility that the same gene(s) orchestrated both events. Many of the genes encoding nuclear oncopro- tcins (which might cooperate with cytoplasmic hcr-ahl) are known to be transcription factors and might well influence the differentiation program. However, transformation and phenotypic change were not always linked. The myeloid switch in the 34.6 line, for example, occurred in a cell that was already tumorigenic and clonable in agar.
The evolution of the hcr-ahl-cxpressing cell lines pro-
Progenitor cells
mutation/ \ mutation
Mast cell differentiation
Erythroid/megakaryocytic Myeloid differentiation differentiation
Fig 7. Differentiation pathways in bcr-ab/ mast cell lines and their phenotypic variants. This model suggests that mutationfs) in mast cell progenitors 'switch' differentiation either to erythroid and megakaryocytic or to myeloid lineages. The mast cell precursors are cells with self-renewal potential clonally descended from the bcr-ab/ virus-infected oligopotential progenitor that caused disease in the primary mice. These cells may be unipotential or they may retain oligopotentiality but be unable to express it in vitro.
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

1280 ELEFANTY AND CORY
vides clues to the function of bcr-ab1 and its role in CML. Expression of bcr-ab1 by a hematopoietic progenitor and its progeny provides only a modest proliferative advantage and relieves certain growth factor requirements but probably not ail. Whether growth factor production, particularly of IL-3,I3 is crucial to the action of bcr-ab1 (and v-abl) or is merely an epiphenomenon remains an important unre- solved issue. The unexpected phenotypic plasticity of the cell lines suggests that bcr-ab1 expression may facilitate and/or complement the (postulated) somatic mutations in lineage-determining genes. Finally, our observation that even fully transformed lines can retain substantial capacity for maturation suggests that the chronic phase of CML may accumulate mutations that increase malignancy without significantly curtailing differentiation. Thus, more events
may be needed to progress to blast crisis than hitherto suspected. The lines characterized here may facilitate identification not only of genes able to cooperate with bcr-ab1 in leukemogenesis but also those instrumental in determining lineage commitment during normal hematopoi- esis.
ACKNOWLEDGMENT
We thank Drs J.M. Adams, A.W. Harris, D. Metcalf, and N.M. Gough for helpful discussions and critically reviewing the manu- script, Dr T. Kina for providing the TER-119 MoAb, Dr J. Visvader for the scl probe, Dr G. Rovera for the myeloperoxidase probe, Dr J.P. Kinet for the IgE receptor probe, Dr G.R. Johnson for performing the Epo bioassay, and Dr A. Strasser for providing reagents and helpful advice for flow cytometry.
REFERENCES
1. Fialkow PJ, Jacobson RJ, Papayannopoulou T Chronic myelocytic leukemia: Clonal origin in a stem cell common to the granulocyte, erythrocyte, platelet and monocyteImacrophage. Am J Med 63:125,1977
2. Nowell PC, Hungerford D A A minute chromosome in human chronic granulocytic leukemia. Science 132:1497,1960
3. Rowley JD: A new consistent chromosomal abnormality in chronic myelogenous leukaemia identified by quinacrine fluores- cence and Giemsa staining. Nature 243:290,1973
4. de Klein A, Geurts van Kessel A, Grosveld G, Bartram CR, Hagemeijer A, Bootsma D, Spurr NK, Heisterkamp N, Groffen J, Stephenson JR: A cellular oncogene is translocated to the Philadel- phia chromosome in chronic myelocytic leukaemia. Nature 300: 765,1982
5. Daley GQ, Ben-Neriah Y: Implicating the bcrlabl gene in the pathogenesis of Philadelphia chromosome-positive human leuke- mia. Adv Cancer Res (in press)
6. Kurzrock R, Gutterman JU, Talpaz M: The molecular genet- ics of Philadelphia chromosome-positive leukemias. N Engl J Med 319:990,1988
7. Konopka JB, Witte O N Activation of the ab1 oncogene in murine and human leukaemias. Biochim Biophys Acta 823:1, 1985
8. Champlin RE, Golde DW: Chronic myelogenous leukemia: Recent advances. Blood 65:1039,1985
9. Kyle RA, Pease G L Basophilic leukemia. Arch Intern Med 118:205,1966
10. Keating A: Ph positive CML cell lines, in Goldman JM (ed): Baillitre's Clinical Haematology International Practice and Re- search, vol 1, no. 4. London, UK, Baillitre Tindall, 1987, p 1021
11. Mes-Masson A-M, McLaughlin J, Daley GQ, Paskind M, Witte ON: Overlapping cDNA clones define the complete coding region for the P210c-"h' gene product associated with chronic myelogenous leukemia cells containing the Philadelphia chromo- some. Proc Natl Acad Sci USA 83:9768,1986
12. Daley GQ, Baltimore D: Transformation of an interleukin 3-dependent hematopoietic cell line by the chronic myelogenous leukemia-specific P2lCP""' protein. Proc Natl Acad Sci USA 85:9312,1988
13. Hariharan IK, Adams JM, Cory S: bcr-ab1 oncogene renders myeloid cell line factor independent: Potential autocrine mecha- nism in chronic myeloid leukemia. Oncogene Res 3:387,1988
14. Laneuville P, Heisterkamp N, Groffen J: Expression of the chronic myelogenous leukemia-associated p2lObcrlabl oncopro- tein in a murine IL-3 dependent myeloid cell line. Oncogene 6:275, 1991
15. McLaughlin J, Chianese E, Witte O N In vitro transforma-
tion of immature hematopoietic cells by the P210 BCRIABL oncogene product of the Philadelphia chromosome. Proc Natl Acad Sci USA 84:6558,1987
16. Young JC, Witte O N Selective transformation of primitive lymphoid cells by the BCRIABL oncogene expressed in long-term lymphoid or myeloid cultures. Mol Cell Biol8:4079, 1988
17. Hariharan IK, Harris AW, Crawford M, Abud H, Webb E, Cory S, Adams JM: A bcr-v-ab1 oncogene induces lymphomas in transgenic mice. Mol Cell Biol9:2798,1989
18. Heisterkamp N, Jenster G, ten Hoeve J, Zovich D, Patten- gale PK, Groffen J: Acute leukaemia in bcrlabl transgenic mice. Nature 344:251,1990
19. Elefanty AG, Hariharan IK, Cory S: bcr-abl, the hallmark of chronic myeloid leukaemia in man, induces multiple haemopoietic neoplasms in mice. EMBO J 9:1069,1990
20. Daley GQ, Van Etten RA, Baltimore D: Induction of chronic myelogenous leukemia in mice by the P210b"'"" gene of the Philadelphia chromosome. Science 247:824, 1990
21. Kelliher MA, McLaughlin J, Witte ON, Rosenberg N: In- duction of a chronic myelogenous leukemia-like syndrome in mice with v-ab1 and BCRIABL. Proc Natl Acad Sci USA 87:6649,1990
22. Bowtell DDL, Johnson GR, Kelso A, Cory S: Expression of genes transferred to haemopoietic stem cells by recombinant retroviruses. Mol Biol Med 4229,1987
23. Kinet J-P, Metzger H, Hakimi J, Kochan J: A cDNA presumptively coding for the a subunit of the receptor with high affinity for immunoglobulin E. Biochemistry 26:4605,1987
24. Venturelli D, Shirsat N, Gemperlein I, Bittenbender S, Hudson S, Rovera G: Nucleotide sequence of cDNA for murine myeloperoxidase. Nucleic Acids Res 17:5852,1989
25. Zakut-Houri R, Oren M, Bienz B, Lavie V, Hazum S, Givol D: A single gene and a pseudogene for the cellular tumour antigen p53. Nature 306594,1983
26. Begley CG, Visvader J, Green AR, Aplan PD, Metcalf D, Kirsch IR, Gough NM: Molecular cloning and chromosomal localization of the murine homolog of the human helix-loop-helix gene SCL. Proc Natl Acad Sci USA 88:869,1991
27. Cory S, Maekawa T, McNeall J, Metcalf D: Murine eryth- roid cell lines derived with c-myc retroviruses respond to leukemia- inhibitory factor, erythropoietin, and interleukin 3. Cell Growth Diff 2165,1991
28. Metcalf D: Molecular control of granulocyte and macro- phage production. Prog Clin Biol Res 191:323, 1985
29. Johnson GR, Barker DC: Erythroid progenitor cells and stimulating factors during murine embryonic and fetal develop- ment. Exp Hematol13:200,1985
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

bcr-ab/ MAST CELLS SWITCH DIFFERENTIATION 1281
30. Metcalf D: Multi-CSF-dependent colony formation by cells of a murine hemopoietic cell line: Specificity and action of multi-CSF. Blood 65:357,1985
31. Metcalf D: Clonal Culture of Hemopoietic Cells: Techniques and Applications. Amsterdam, The Netherlands, Elsevier, 1984
32. Springer T, Galfrk G, Secher DS, Milstein C: Mac-1: A macrophage differentiation antigen identified by monoclonal anti- body. Eur J Immunol9:301,1979
33. Holmes KL, Langdon WY, Fredrickson TN, Coffman RL, Hoffman PM, Hartley JW, Morse HC 111: Analysis of neoplasms induced by Cas-Br-M MuLV tumor extracts. J Immunol 137:679, 1986
34. Austyn JM, Gordon S: F4/80, a monoclonal antibody di- rected specifically against the mouse macrophage. Eur J Immunol 11:805,1981
35. Ikuta K, Kina T, MacNeil I, Uchida N, Peault B, Chien Y, Weissman IL: A developmental switch in thymic lymphocyte maturation potential occurs at the level of hematopoietic stem cells. Cell 629363,1990
36. Unkeless J C Characterization of a monoclonal antibody directed against mouse macrophage and lymphocyte Fc receptors. J Exp Med 150:580,1979
37. Cooper MC, Levy J, Cantor LN, Marks PA, Rifkind R A The effect of erythropoietin on colonial growth of erythroid precursor cells in vitro. Proc Natl Acad Sci USA 71:1677,1974
38. Kaiho S-I, Mizuno K Sensitive assay systems for detection of hemoglobin with 2,7-diaminofluorene: Histochemistry and colo- rimetry for erythrodifferentiation. Anal Biochem 149:117,1985
39. Witte ON: Steel locus defines new multipotent growth factor. Cell 63:5,1990
40. Romeo P-H, Prandini M-H, Joulin V, Mignotte V, Prenant M, Vainchenker W, Marguerie G, Uzan G: Megakaryocytic and erythrocytic lineages share specific transcription factors. Nature 344:447,1990
41. Martin DIK, Zon LI, Mutter G, Orkin SH: Expression of an erythroid transcription factor in megakaryocytic and mast cell lineages. Nature 344444,1990
42. Visvader J, Begley CG, Adams JM: Differential expression of the L E , SCL and E24 helix-loop-helix genes within the hemopoietic system. Oncogene 6187,1991
43. Green AR, Salvaris E, Begley CG: Erythroid expression of the 'helix-loop-helix' gene, SCL. Oncogene 6:475,1991
44. Boyd AW, Schrader JW: Derivation of macrophage-like lines from the pre-B lymphoma ABLS 8.1 using 5-azacytidine. Nature 297:691,1982
45. Ahuja H, Bar-Eli M, Advani SH, Benchimol S, Cline MJ: Alterations in the p53 gene and the clonal evolution of the blast crisis of chronic myelocytic leukemia. Proc Natl Acad Sci USA 86:6783, 1989
46. Kelman Z, Prokocimer M, Peller S, Kahn Y, Rechavi G, Manor Y, Cohen A, Rotter V Rearrangements in the p53 gene in Philadelphia chromosome positive chronic myelogenous leukemia. Blood 74:2318,1989
47. Mabhal R, Shtalrid M, Talpaz M, Kantarjian H, Smith L, Beran M, Cork A, Trujillo J, Gutterman J, Deisseroth A Rearrange- ment and expression of p53 in the chronic phase and blast crisis of chronic myelogenous leukemia. Blood 75:180,1990
48. Lane DP, Benchimol S: p53: Oncogene or anti-oncogene? Genes Dev 4:1,1990
49. Klemsz MJ, McKercher SR, Celada A, Van Beveren C, Maki RA: The macrophage and B cell-specific transcription factor PU.1 is related to the ets oncogene. Cell 61:113,1990
50. Moreau-Gachelin F, Ray D, Mattei M-G, Tambourin P, Tavitian A: The putative oncogene Spi-1: Murine chromosomal localization and transcriptional activation in murine acute erythro- leukemias. Oncogene 4:1449,1989
51. Wong PMC, Humphries RK, Chen TR, Eaves CJ: Evidence for a multistep pathogenesis in the generation of tumorigenic cell lines from hemopoietic colonies exposed to Abelson virus in vitro. Exp Hematol15:280,1987
52. Wong PMC, Chung S-W, Raefsky E, Eaves CJ, Nienhuis AW: Blast colonies containing hemopoietic progenitor cells can give rise to Abelson virus (A-MuLV)-transformed cell lines. Exp Hematol 16:5,1988
53. Pierce JH, Di Fiore PP, Aaronson SA, Potter M, Pumphrey J, Scott A, Ihle J N Neoplastic transformation of mast cells by Abelson-MuLV Abrogation of IL-3 dependence by a nonauto- crine mechanism. Cell 41:685,1985
54. Chung SW, Wong PMC, Shen-Ong G, Ruscetti S, Ishizaka T, Eaves CJ: Production of granulocyte-macrophage colony- stimulating factor by Abelson virus-induced tumorigenic mast cell lines. Blood 68:1074, 1986
55. Humphries RK, Abraham S, Krystal G, Landsdorp P, Lemoine F, Eaves CJ: Activation of multiple hemopoietic growth factor genes in Abelson virus-transformed myeloid cells. Exp Hematol16:774,1988
56. Eaves AC, Henkelman DH, Eaves CJ: Abnormal erythropoi- esis in the myeloproliferative disorders: An analysis of underlying cellular and humoral mechanisms. Exp Hematol8:235,1980 (suppl
57. Waneck GL, Rosenberg N Abelson leukemia virus induces lymphoid and erythroid colonies in infected fetal liver cultures. Cell 26:79, 1981
58. Collins SJ: The HL-60 promyelocytic leukemia cell line: Proliferation, differentiation, and cellular oncogene expression. Blood 70:1233,1987
59. Greenberger JS, Sakakeeny MA, Humphries RK, Eaves CJ, Bckner RJ: Demonstration of permanent factor-dependent multipo- tential (erythroid/neutrophil/basophil) hematopoietic progenitor cell lines. Proc Natl Acad Sci USA 80:2931, 1983
60. Valtieri M, Tweardy DJ, Caracciolo D, Johnson K, Mavilio F, Altmann S, Santoli D, Rovera G: Cytokine-dependent granulo- cytic differentiation. Regulation of proliferative and differentiative responses in a murine progenitor cell line. J Immunol 138:3829, 1987
8)
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom

1992 79: 1271-1281
AG Elefanty and S Cory myeloid differentiation in vitrobcr-abl-Induced cell lines can switch from mast cell to erythroid or
http://www.bloodjournal.org/content/79/5/1271.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on April 3, 2019. by guest www.bloodjournal.orgFrom





























