Embed Size (px)
Citation preview

SHORT COMMUNICATION
Bacterial communities in sediments of a drinkingwater reservoir
Wen Cheng & Jingxu Zhang & Zhao Wang & Min Wang &
Shuguang Xie
Received: 27 March 2013 /Accepted: 1 August 2013# Springer-Verlag Berlin Heidelberg and the University of Milan 2013
Abstract Sediment microbial communities play crucial rolesin biogeochemical cycles of aquatic systems. Knowledge ofmicrobial community structure could aid in our understandingof these roles. In this study, the sediment bacterial communitystructure of a drinking water reservoir in China was investi-gated. A large difference in major bacterial groups was ob-served at different sampling sites. Proteobacteria was thelargest bacterial phylum, with a marked shift in the propor-tions of its major subdivisions. Microorganisms within phy-lum Proteobacteria might play important roles in variousbiogeochemical processes.
Keywords Reservoir . Sediment . Proteobacteria . Microbialcommunity . Freshwater
Freshwater sediment harbors high densities of viable microor-ganisms which are major components of aquatic ecosystems(Borrel et al. 2012). Sediment microbial communities playcrucial roles in decomposition of organic matter and in biogeo-chemical cycling of elements such as nitrogen, phosphorus,
sulfur, and iron (Martins et al. 2011). Themicrobial communitystructure of lake sediment can be affected by various factorssuch as total phosphorus (Song et al. 2012), and pollution(Haller et al. 2011). Moreover, the microbial composition oflake sediment shows seasonal and spatial variations (Song et al.2012). However, the links between environmental conditionsand aquatic microbial ecology remain poorly understood(Lymperopoulou et al. 2012). Reservoirs are artificial lakes,and usually experience shorter retention times andmore intensewater level fluctuations than natural lakes, so different physical,chemical, and biological processes can occur in these kinds ofaquatic systems (Lymperopoulou et al. 2012). However, fewstudies have been performed to investigate the composition andvariability of the bacterial community of reservoir sediment(Qu et al. 2008; Roske et al. 2008, 2012).
In China, there is no information available on the sedimentmicrobial community of drinking water reservoirs, althoughthere have been a few reports on sediment microbial commu-nities of shallow and eutrophic freshwater lakes (Song et al.2012) and eutrophic and heavily polluted reservoirs (Qu et al.2008). The Tangpu Reservoir, located in the rural region ofShaoxing City, is a typical temperate drinking water reservoir.The Tangpu Reservoir covers 13.64 km2 and its catchmentarea is about 460 km2. The main objective of the current studywas to investigate the sediment bacterial community structureof the Tangpu Reservoir. This work could add some newinsights into the structure of freshwater sedimentary microbialcommunities.
In this study, sediment samples in triplicate were collectedusing a grab sampler at four different sites (Fig. S1): A(29°50’32”N, 120°46’ 58”E, 21.8 m deep), B(29°49’57.97”N, 120°45’ 51”E, 19.2 m deep), C(29°49’13”N, 120°45’ 38”E, 15.5 m deep), and D(29°50’14.68”N, 120°46’ 23.35”E, 19.7 m deep). Samplingsites C and B were in the inflow area of the main basin, whilesampling site Awas in the outflow area. Sampling site D was inthe center of the main basin. Sediment samples were sliced into
Electronic supplementary material The online version of this article(doi:10.1007/s13213-013-0712-z) contains supplementary material, whichis available to authorized users.
W. Cheng :M. WangState Key Laboratory of Ecohydraulic Engineering in Shaanxi,Xi’an University of Technology, Xi’an 710048, China
J. Zhang : Z. Wang : S. XieCollege of Environmental Sciences and Engineering, The KeyLaboratory of Water and Sediment Sciences (Ministry of Education),Peking University, Beijing 100871, China
S. Xie (*)College of Environmental Sciences and Engineering,Peking University, Beijing 100871, Chinae-mail: [email protected]
Ann MicrobiolDOI 10.1007/s13213-013-0712-z
layers. The layer at 0–5 cm sediment depth was homogenizedand subsampled for further molecular analysis. Organic andphosphorus contents of the sediments ranged between 12.6and 26.8 g kg−1, and 0.29 and 0.59 g kg−1, respectively.Ammonium, nitrate and total nitrogen contents of the sedimentswere 121.7–167.2mg kg−1, 4.4–6.7mg kg−1, and 1.1–2.8 g kg−1,respectively (Table S1).
Total genomic DNA of each sediment sample wasextracted using the UltraClean DNA extraction kit (MobioLaboratories, USA) according to the manufacturer’s protocol.The same amount of DNA from triplicate samples was pooledtogether for clone library analysis. Bacterial 16S rRNA geneswere amplified using primers 27 F (5’-GAGTTTGATCMTGGCTCAG-3’) and 1492R (5’-GGTTACCTTGTTACGACTT-3’) according to the literature (Wan et al. 2012;Zhang et al. 2012). The amplicons were cloned into apGEM-T-easy Vector (Takara, Japan). The clones containingcorrect sizes were sequenced. The taxonomic identities ofbacterial sequences were assigned using the Classifier pro-gram of the Ribosomal Database Project (RDP)-II at confi-dence level of 80 % (Wang et al. 2007). The bacterial se-quences obtained in this study were submitted to GenBank,under accession numbers KC559913–KC559960 for SampleA, KC559961–KC560010 for Sample B, KC520581–KC520626 for Sample C, and KC520627–KC520674 forSample D.
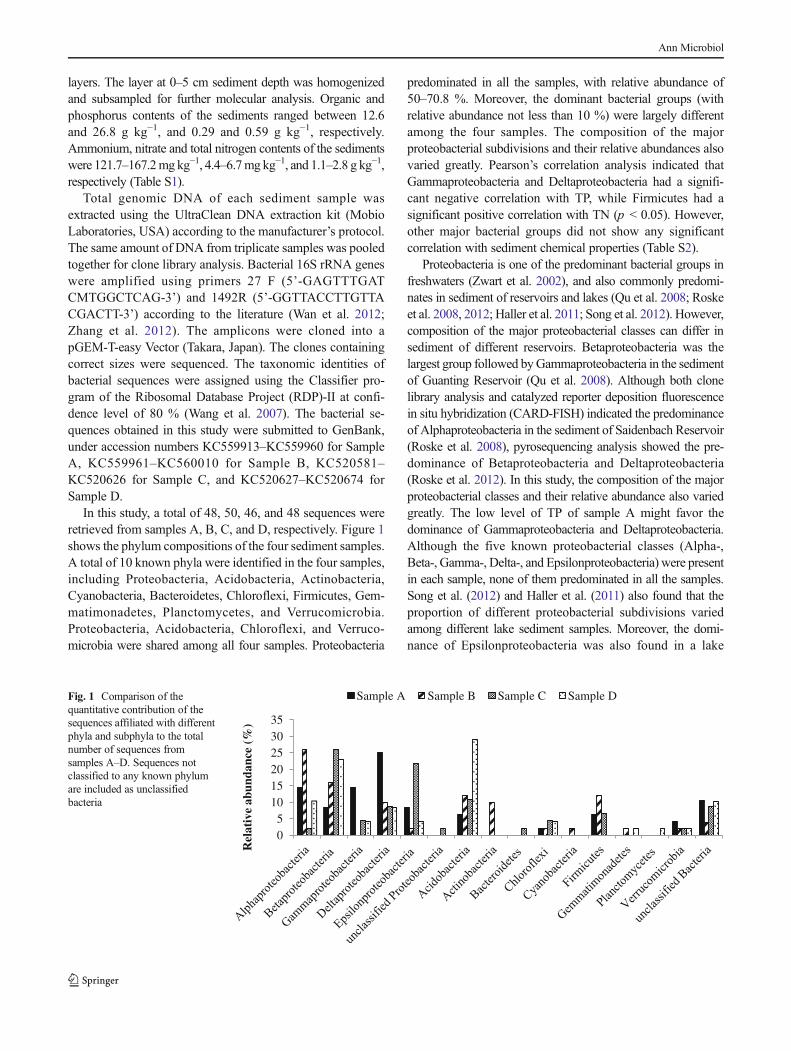
In this study, a total of 48, 50, 46, and 48 sequences wereretrieved from samples A, B, C, and D, respectively. Figure 1shows the phylum compositions of the four sediment samples.A total of 10 known phyla were identified in the four samples,including Proteobacteria, Acidobacteria, Actinobacteria,Cyanobacteria, Bacteroidetes, Chloroflexi, Firmicutes, Gem-matimonadetes, Planctomycetes, and Verrucomicrobia.Proteobacteria, Acidobacteria, Chloroflexi, and Verruco-microbia were shared among all four samples. Proteobacteria
predominated in all the samples, with relative abundance of50–70.8 %. Moreover, the dominant bacterial groups (withrelative abundance not less than 10 %) were largely differentamong the four samples. The composition of the majorproteobacterial subdivisions and their relative abundances alsovaried greatly. Pearson’s correlation analysis indicated thatGammaproteobacteria and Deltaproteobacteria had a signifi-cant negative correlation with TP, while Firmicutes had asignificant positive correlation with TN (p < 0.05). However,other major bacterial groups did not show any significantcorrelation with sediment chemical properties (Table S2).
Proteobacteria is one of the predominant bacterial groups infreshwaters (Zwart et al. 2002), and also commonly predomi-nates in sediment of reservoirs and lakes (Qu et al. 2008; Roskeet al. 2008, 2012; Haller et al. 2011; Song et al. 2012). However,composition of the major proteobacterial classes can differ insediment of different reservoirs. Betaproteobacteria was thelargest group followed byGammaproteobacteria in the sedimentof Guanting Reservoir (Qu et al. 2008). Although both clonelibrary analysis and catalyzed reporter deposition fluorescencein situ hybridization (CARD-FISH) indicated the predominanceof Alphaproteobacteria in the sediment of Saidenbach Reservoir(Roske et al. 2008), pyrosequencing analysis showed the pre-dominance of Betaproteobacteria and Deltaproteobacteria(Roske et al. 2012). In this study, the composition of the majorproteobacterial classes and their relative abundance also variedgreatly. The low level of TP of sample A might favor thedominance of Gammaproteobacteria and Deltaproteobacteria.Although the five known proteobacterial classes (Alpha-,Beta-, Gamma-, Delta-, and Epsilonproteobacteria) were presentin each sample, none of them predominated in all the samples.Song et al. (2012) and Haller et al. (2011) also found that theproportion of different proteobacterial subdivisions variedamong different lake sediment samples. Moreover, the domi-nance of Epsilonproteobacteria was also found in a lake
05
101520253035
Rel
ativ
e ab
unda
nce
(%)
Sample A Sample B Sample C Sample DFig. 1 Comparison of thequantitative contribution of thesequences affiliated with differentphyla and subphyla to the totalnumber of sequences fromsamples A–D. Sequences notclassified to any known phylumare included as unclassifiedbacteria
Ann Microbiol

sediment sample (Song et al. 2012). However, to the authors’knowledge, this was the first report on the dominance ofEpsilonproteobacteria in reservoir sediment.
Acidobacteria, Actinobacteria, and Firmicutes have usuallybeen found to be minor components of sediment bacterialcommunities (Qu et al. 2008; Roske et al. 2008, 2012; Halleret al. 2011). Song et al. (2012) reported the dominance ofAcidobacteria and Firmicutes in sediment of Lake Dongping.However, to the authors’ knowledge, the dominance ofAcidobacteria, Actinobacteria, or Firmicutes in reservoir sedi-ment has not been reported in previous studies. In this study,interestingly, Actinobacteria and Firmicutes were the dominantbacterial groups in sample B, while Acidobacteria were abun-dant in samples B, C, and D. The high TN level of sample Bmight favor the growth of Firmicutes.
Nearly 40–60 % of the sequences from each sample couldbe classified at genus level (data not shown). Table 1 showsthe 27 known proteobacterial genera detected in the foursamples. Most of these genera had only a single sequence.Only genus Sulfuricurvum was found in all four samples, and34 genera were present only in a single sample. Therefore, atthe genus level of taxonomic classifications, the differences inthe bacterial community structure were more evident.
Microorganisms from alphaproteobacterial genera (Bradyrhi-zobium , Hyphomicrobium , Sphingomonas and Novosphing-obium) can degrade a variety of environmental organic pollutants(Liao et al. 2012, 2013).Hydrogenophaga (Betaproteobacteria ),Comamonas (Betaproteobacteria), and Rhodanobacter(Gammaproteobacteria) are also known for biodegrading manykinds of environmental chemicals (Kanaly et al. 2002; Lamboand Patel 2007; Ruzicka et al. 2011). Steroidobacter(Gammaproteobacteria) has been linked to anaerobic degrada-tion of testosterone (Fahrbach et al. 2010). Moreover, membersof generaMethylocaldum (Gammaproteobacteria) andMethylo-bacter (Gammaproteobacteria) are known as methanotrophs andcan assimilate methane carbon (Kalyuzhnaya et al. 1998;Medvedkova et al. 2009). Therefore, these proteobacterial mi-croorganisms might play active roles in biodegradation of organ-ic pollutants and carbon cycling in the Tangpu Reservoir.
Members of genera Sulfuricella (Betaproteobacteria),Thiovirga (Gammaproteobacteria), and Sulfuricurvum(Epsilonproteobacteria) have been linked to sulfur oxidation(Kodama and Watanabe 2004; Ito et al. 2005; Kojima andFukui 2010). The dominance of Sulfuricurvum species as wellas the presence of Sulfuricella and Thiovirga species sug-gested a strong sulfur oxidation process in reservoir sediment.Acinetobacter (Gammaproteobacteria) is one of commonlydetected phosphorus-accumulating microorganisms in sediment(Martins et al. 2011). The presence of Acinetobacter speciesmight be involved in phosphorus cycling. Moreover, microor-ganisms from genera Anaeromyxobacter (Deltaproteobacteria)and Geobacter (Deltaproteobacteria) are known as Fe(III) re-ducers (Haaijer et al. 2008; Chao et al. 2010). This two
deltaproteobacterial species might be involved in Fe(III)reduction.
In conclusion, major bacterial groups showed a large varia-tion at different sampling sites. Proteobacteria was the largestphylum and might be involved in various biogeochemicalprocesses in sediment. However, further workwill be necessaryin order to elucidate the composition of sediment microbialcommunities and their ecological function in drinking waterreservoirs.
Acknowledgments This work was financially supported by NationalNatural Science Foundation of China (No. 51076130).
Table 1 Distribution of the sequences affiliated with the identifiedproteobacterial genera in samples A–D
Phylogenetic affiliation Sample A Sample B Sample C Sample D
Alphaproteobacteria
Bradyrhizobium 1 —a — —
Roseomonas 1 1 — —
Hyphomicrobium — 1 — —
Rhodoplanes — 1 — 1
Phenylobacterium — 1 — —
Novosphingobium — 1 — —
Sphingomonas — 1 — —
Defluviicoccus — 1 — —
Betaproteobacteria
Sulfuricella — 1 — —
Herminiimonas — 1 — —
Massilia — — 2 —
Hydrogenophaga — — 2 —
Comamonas — — 1 —
Leeia — — 3 —
Vogesella — — 1 —
Gammaproteobacteria
Rhizobacter 1 — — —
Rhodanobacter 1 — — —
Methylocaldum 1 — — —
Acinetobacter 1 — — —
Steroidobacter 1 — — 1
Methylobacter — — 1 —
Thiovirga — — — 1
Deltaproteobacteria
Anaeromyxobacter 1 1 — —
Smithella 2 — — 1
Geobacter 2 — 1 —
Syntrophorhabdus — — 1 —
Epsilonproteobacteria
Sulfuricurvum 4 1 10 2
Total 16 11 22 6
a— not detected.
Ann Microbiol

References
Borrel G, Lehours AC, Crouzet O, Jezequel D, Rockne K, Kulczak A,Duffaud E, Joblin K, Fonty G (2012) Stratification of Archaea in thedeep sediments of a freshwater Meromictic Lake: vertical shift frommethanogenic to uncultured archaeal lineages. Plos ONE 7(8)doi:10.1371/journal.pone.0043346
Chao TC, Kalinowski J, Nyalwidhe J, Hansmeier N (2010) Comprehen-sive proteome profiling of the Fe(III)-reducing myxobacteriumAnaeromyxobacter dehalogenans 2CP-C during growth with fuma-rate and ferric citrate. Proteomics 10(8):1673–1684
Fahrbach M, Krauss M, Preiss A, Kohler HPE, Hollendera J (2010)Anaerobic testosterone degradation in Steroidobacter denitrificans–identification of transformation products. Environ Pollut 158(8):-2572–2581
Haaijer SCM, Harhangi HR, Meijerink BB, Strous M, Pol A, SmoldersAJP, Verwegen K, Jetten MSM, den Camp HJMO (2008) Bacteriaassociated with iron seeps in a sulfur-rich, neutral pH, freshwaterecosystem. ISME J 2(12):1231–1242
Haller L, Tonolla M, Zopfi J, Peduzzi R, Wildi W, Pote J (2011) Compo-sition of bacterial and archaeal communities in freshwater sedimentswith different contamination levels (Lake Geneva, Switzerland).Water Res 45(3):1213–1228
Ito T, Sugita K, Yumoto I, Nodasaka Y, Okabe S (2005) Thiovirgasulfuroxydans gen. nov., sp nov., a chemolithoautotrophic sulfur-oxidizing bacterium isolated from a microaerobic waste-water bio-film. Int J Syst Evol Microbiol 55:1059–1064
KalyuzhnayaMG, Khmelenina VN, Starostina NG, Baranova SV, SuzinaNE, TrotsenkoYA (1998) Newmoderately halophilic methanotrophof the genus Methylobacter. Microbiology 67(4):438–444
Kanaly RA, Harayama S, Watanabe K (2002) Rhodanobacter sp strainBPC1 in a benzo[a]pyrene-mineralizing bacterial consortium. ApplEnviron Microbiol 68(12):5826–5833
Kodama Y, Watanabe K (2004) Sulfuricurvum kujiense gen. nov., sp nov., afacultatively anaerobic, chemolithoautotrophic, sulfur-oxidizing bacteri-um isolated from an underground crude-oil storage cavity. Int J SystEvol Microbiol 54:2297–2300
Kojima H, Fukui M (2010) Sulfuricella denitrificans gen. nov., sp nov., asulfur-oxidizing autotroph isolated from a freshwater lake. Int J SystEvol Microbiol 60:2862–2866
Lambo AJ, Patel TR (2007) Biodegradation of polychlorinated biphenylsin Aroclor 1232 and production of metabolites from 2,4,4'-trichlorobiphenyl at low temperature by psychrotolerant Hydro-genophaga sp strain IA3-A. J Appl Microbiol 102(5):1318–1329
Liao XB, Chen C, Chang CH, Wang Z, Zhang XJ, Xie SG (2012)Heterogeneity of microbial community structures inside the up-
flow biological activated carbon (BAC) filters for the treatment ofdrinking water. Biotechnol Bioprocess Eng 17(4):881–886
Liao XB, Chen C, Wang Z, Wan R, Chang CH, Zhang X, Xie S (2013)Changes of biomass and bacterial communities in biological acti-vated carbon filters for drinking water treatment. Process Biochem48(2):312–316
Lymperopoulou DS, Kormas KA, Karagouni AD (2012) Variability ofprokaryotic community structure in a drinking water reservoir(Marathonas, Greece). Microbes Environ 27(1):1–8
Martins G, Terada A, Ribeiro DC, Corral AM, Brito AG, Smets BF,Nogueira R (2011) Structure and activity of lacustrine sedimentbacteria involved in nutrient and iron cycles. FEMS Microbiol Ecol77(3):666–679
Medvedkova KA, Khmelenina VN, Suzina NE, Trotsenko YA (2009)Antioxidant systems of moderately thermophilic methanotrophsMethylocaldum szegediense andMethylococcus capsulatus. Micro-biology 78(6):670–677
Qu JH, Yuan HL, Wang et, Li C, Huang HZ (2008) Bacterial diversity insediments of the eutrophic Guanting Reservoir, China, estimated byanalyses of 16S rDNA sequence. Biodivers Conserv 17(7):1667–1683
Roske K, Roske I, Uhlmann D (2008) Characterization of the bacterialpopulation and chemistry in the bottom sediment of a laterallysubdivided drinking water reservoir system. Limnologica 38(3–4):367–377
Roske K, Sachse R, Scheerer C, Roske I (2012) Microbial diversity andcomposition of the sediment in the drinking water reservoirSaidenbach (Saxonia, Germany). Syst Appl Microbiol 35(1):35–44
Ruzicka J, Dvorackova M, Koutny M (2011) Trichloroethylene degrada-tion by two phenol utilizing strains of Comamonas testosterone .Asian J Chem 23(10):4255–4259
Song H, Li Z, Du B, Wang G, Ding Y (2012) Bacterial communities insediments of the shallowLakeDongping in China. J Appl Microbiol112(1):79–89
Wang Q, Garrity GM, Tiedje JM, Cole JR (2007) Naïve Bayesianclassifier for rapid assignment of rRNA sequences into the newbacterial taxonomy. Appl Environ Microbiol 73(16):5261–5267
Wan R, Zhang SY, Xie SG (2012) Microbial community changes inaquifer sediment microcosm for anaerobic anthracene biodegrada-tion under methanogenic condition. J Environ Sci 24(8):1498–1503
Zhang SY, Wang QF, Xie SG (2012) Molecular characterization ofphenanthrene-degrading methanogenic communities in leachate-contaminated aquifer sediment. Int J Environ Sci Technol9(4):705–712
Zwart G, Crump BC, AgterveldMPKV, Hagen F, Han SK (2002) Typicalfreshwater bacteria: an analysis of available 16S rRNA gene se-quences from plankton of lakes and rivers. Aquat Microb Ecol28(2):141–155
Ann Microbiol





























