Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/ijpara
International Journal for Parasitology 38 (2008) 1219–1237
Babesiosis: Recent insights into an ancient disease
K.-P. Hunfeld a,*, A. Hildebrandt b, J.S. Gray c
a Institute of Medical Microbiology & Infection Control, Hospital of the Johann Wolfgang Goethe-University Frankfurt am Main,
Department of Serology and Molecular Diagnostics, Paul-Ehrlich-Str. 40, D-60596 Frankfurt/Main, Germanyb Institute of Medical Microbiology, Friedrich-Schiller-University Jena, Semmelweis Str. 4, D-07743 Jena, Germany
c UCD School of Biology and Environmental Science, University College Dublin, Dublin 4, Ireland
Received 3 January 2008; received in revised form 29 February 2008; accepted 3 March 2008
Abstract
Ever since the discovery of parasitic inclusions in erythrocytes of cattle in Romania by Victor Babes at the end of the 19th century,newly recognised babesial pathogens continue to emerge around the world and the substantial public health impact of babesiosis on live-stock and man is ongoing. Babesia are transmitted by ixodid ticks and infection of the host causes a host-mediated pathology and eryth-rocyte lysis, resulting in anemia, hyperbilirubinuria, hemoglobinuria, and possibly organ failure. Recently obtained molecular data,particularly for the 18S rRNA gene, has contributed significantly to a better understanding of the sometimes puzzling phylogenetic sit-uation of the genus Babesia and new information has been added to help determine the taxonomic position of many species. Moreover, itseems that owing to higher medical awareness the number of reported cases in humans is rising steadily. Hitherto unknown zoonoticbabesias are now being reported from geographical areas where babesiosis was not known to occur and the growing numbers of immu-nocompromised individuals suggest that the frequency of cases will continue to rise. This review covers recent insights into human bab-esiosis with regard to phylogeny, diagnostics and treatment in order to provide new information on well known as well as recentlydiscovered parasites with zoonotic potential.� 2008 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Keywords: Babesia; Ticks; Zoonosis; Phylogeny; Human disease; Blood transfusion; IFAT; PCR; Diagnostics; Treatment; Prevention
1. Introduction
Tick-transmitted hemoparasites of the protozoan genusBabesia (phylum Apicomplexa) are the second most com-mon blood-borne parasites of mammals after the trypano-somes (Telford et al., 1993). It was Victor Babes who at theend of the 19th century first discovered microorganisms inerythrocytes of cattle in Romania and associated them withbovine hemoglobinuria or red water fever (Babes, 1888).Five years later, Smith and Kilbourne established Pyrosoma
– later renamed Babesia bigemina – as the causative agent ofTexas Cattle Fever (Smith and Kilbourne, 1893), a findingof historic significance because this piroplasm was thefirst recognised arthropod-borne pathogen of vertebrates
0020-7519/$34.00 � 2008 Australian Society for Parasitology Inc. Published b
doi:10.1016/j.ijpara.2008.03.001
* Corresponding author. Tel.: +49 69 6301 6441; fax: +49 69 6301 5767.E-mail address: [email protected] (K.-P. Hunfeld).
(Kjemtrup and Conrad, 2000). Since then, newly recognisedbabesia with zoonotic potential continue to emerge aroundthe world and the substantial economic impact of babesiosison livestock and companion animals especially in the tropicsand subtropics is ongoing (Collett, 2000; Kivaria et al.,2007). A fatal Babesia divergens infection in 1956 was the firstconfirmed case of human babesiosis (Skrabalo and Deanov-ic, 1957) and, ever since, babesiosis came into view as apotentially life threatening zoonotic infection in humans(Homer et al., 2000; Herwaldt et al., 2003). Although, severalbabesia species have been involved in human infectionsworldwide (Gorenflot et al., 1998), the major public healthburden on man lies in North America and is due to Babesia
microti, especially in the eastern parts of the US (Homeret al., 2000). In these classic areas of endemicity, babesiosisis on the rise and the number of cases appears to be increasingin some parts of the US relative to the number of Lyme
y Elsevier Ltd. All rights reserved.

1220 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
disease cases (Meldrum et al., 1992; Krause et al., 2003).Moreover, during the last decade, newly recognised babesiaparasites (Table 1) have been implicated in human diseaseand it seems that owing to higher medical awareness thenumber of reported cases is rising steadily (Hildebrandtet al., 2007). In addition, the occurrence of hitherto unknownzoonotic parasites is now reported from geographical areaswhere babesiosis was not known to occur and obviouslythe growing population of immunocompromised individualsis ever more involved (Hunfeld and Brade, 2004; Haselbarthet al., 2007; Hildebrandt et al., 2007; Karp and Auwaerter,2007). Most significantly, molecular analysis of the impli-cated pathogens suggests that the host-range of many babe-sia is less restricted than believed previously and also thathitherto unrecognised species can cause infections in a vari-ety of animal hosts and in humans (Zahler et al., 2000; Choet al., 2002; Herwaldt et al., 2003, 2004; Conrad et al.,2006; Kjemtrup et al., 2006; Haselbarth et al., 2007; Kimet al., 2007). Therefore, many past cases of human babesiosison both sides of the Atlantic that were attributed, based ontraditional methods, to classic species such as B. divergens
or B. microti, may indeed be due to species not yet knownto cause such infections in humans (Herwaldt et al., 2003;Gray, 2006; Hildebrandt et al., in press). This notion is fur-ther substantiated by the recent recognition of Babesia dun-
cani and B. divergens-like organisms as pathogens of medicalsignificance for humans in the US (Herwaldt et al., 1996;Beattie et al., 2002; Conrad et al., 2006). Moreover, con-firmed autochthonous B. microti infections have beenreported in Taiwan, Japan and Europe (Shih et al., 1997; Sai-to-Ito et al., 2000; Hildebrandt et al., 2007), and a new Euro-pean B. divergens-like organism (EU1), provisionally namedBabesia venatorum, has been discovered, which is probably aparasite of deer (Telford and Goethert, 2004; Bonnet et al.,2007). This parasite was involved in the first documentedcases of human babesiosis in Italy, Austria and Germany(Herwaldt et al., 2003; Haselbarth et al., 2007). Such newfindings now clearly challenge the dogma that human babe-siosis in North America is almost exclusively caused by B.
microti and that human babesiosis in Europe is solely dueto B. divergens infection in splenectomized individuals. Thisreview covers recent developments and important new infor-mation on well known and recently discovered babesias withzoonotic potential.
2. Classification and life cycle characteristics of Babesia spp.
Babesia are classified as apicomplexan parasites of thesuborder Piroplasmidea and family Babesiidae on the basisof their exclusive invasion of erythrocytes, multiplicationby budding rather than schizogony, and lack of hemozoin.The life cycles of the parasites are very similar (Fig. 1). Allspecies of babesia are naturally transmitted by the bite ofinfected ticks (almost all ixodids rather than argasids)and the main lifecycle difference amounts to the presenceof transovarial transmission in some species (Babesia spp.sensu stricto) and not in others (B. microti-like). During
the tick bite, sporozoites are injected into the host anddirectly infect red blood cells (Fig. 1). This phenomenonseparates Babesia spp. from Theileria spp., where sporozo-ites do not readily infect red blood cells but initially pene-trate a lymphocyte or macrophage in which developmentinto schizonts takes place (Uilenberg, 2006). In the host,babesia sporozoites develop into piroplasms inside theinfected erythrocyte resulting in two or sometimes fourdaughter cells that leave the host cell to infect other eryth-rocytes until the host dies or the immunity of the hostclears the parasites. The spleen with its lympho-reticular fil-ter function is essential in resisting primary infections ofBabesia spp. by specifically removing infected cells fromcirculation, probably through a combination of spleenmicrocirculation and stimulated phagocytic cell activity(de Vos et al., 1987; Gray and Weiss, 2008).
To date, more than 100 species have been identified,infecting many mammalian and some avian species (Grayand Weiss, 2008). Traditionally, babesias were mainlygrouped on the basis of their morphology, host/vectorspecificity, and susceptibility to drugs. Pragmatically, theyare divided into the small babesias (trophozoites of 1.0–2.5 lm; including Babesia gibsoni, B. microti, and Babesia
rodhaini and large babesias (2.5–5.0 lm; including Babesia
bovis, Babesia caballi, and Babesia canis). These morpho-logical classifications are generally consistent with thephylogenetic characterization based on nuclear small sub-unit-ribosomal RNA gene (18S rDNA) sequences, whichshows that the large and small babesias fall into two phylo-genetic clusters, with the small babesias being more relatedto Theileria spp. than the large (with the exception of B. div-
ergens, which appears small on blood smears [0.4–1.5 lm]but is genetically related to large babesias (Homer et al.,2000). Recently, molecular genetic analyses clarified thesomewhat confused phylogenetic situation, sometimesresulting in the emergence of new groups and 18S rDNAanalysis added new information to the taxonomic positionof many piroplasm species (Kjemtrup and Conrad, 2006).A careful study by Criado-Fornelio et al. (2003) recentlysuggested division of the piroplasms into five distinct clades:(i) B. microti group containing B. rodhaini, Babesia felis,Babesia leo, B. microti, and a B. microti-like canine isolate,(ii) western US Theileria-like group, containing Babesia con-
radae, (iii) Theileria-group, containing all Theileria speciesfrom bovines, (iv) a first group of ‘true’ Babesia spp. (sensustricto) including B. canis and B. gibsoni from caninestogether with B. divergens and Babesia odocoilei, and (v) asecond Babesia spp. sensu stricto group composed mainlyof Babesia spp. from ungulates: B. caballi, B. bigemina,B. ovis, B. bovis, and other Babesia spp. from cattle.
2.1. New developments in the phylogeny of B. microti and B.
microti-like organisms
Latest research suggests that B. microti is only distantlyrelated to Babesia species sensu stricto (B. bigemina,B. bovis, and B. divergens), that are best known as parasites

Table 1Important Babesia spp. with zoonotic potential including recently recognised defined parasites
Species Vector Vertebrate host Geographical occurrence Reported humancases (N)
Reportedmortality
Species StageLarge babesia
B. divergens and B. divergens-likeB. divergens s.s. Ixodes ricinusa, Ixodes ventalloi (?) Larvae, nymphs, adults Cattle, wild ruminants Europe >30 �42%B. venatorum (EU1) Ixodes ricinusa Larvae, nymphs, adults Deer Europe 3 0%MO1 and related parasites Ixodes dentatus (?) ? Cottontail rabbits USA 3 33%
B. ovis-likeKO1 Hemaphysalis longicornis (?) ? Sheep (?) Korea, Asia (?) 1 0%B. bovisb Boophilus spp., Ixodes spp.a,
Rhipicephalus bursa
Larvae Cattle, water buffalo,wild ruminants
Southern Europe, Africa,America, Asia, Australia
2 100%
B. canisb Rhipicephalus sanguineusa,Hemaphysalis leachi,Dermacentor reticularisa
Nymphs, adults dogs, Vulpes vulpes,wild canines
Europe, Asia, Africa,America, Australia
1 0%
Small babesia
B. microti & B. microti-likeB. microti complex Ixodes trianguliceps, Ixodes ricinusa,
Ixodes ovatusa, Ixodes scapularisa,Ixodes spinipalpis, Ixodes angustus,Ixodes muris
Nymphs, adults Rodents Europe, Asia, America >200 �5%
B. duncani & B. duncani-like ? ? ? USA 9 (?) 11%
a Known to regularly parasitize humans; ?: unknown; (?): questionable.b Unverified infections with B. canis and B. bovis have been reported (Gorenflot et al., 1998), though it is likely that B. bovis infections are in fact B. divergens.
K.-P
.H
un
feldet
al./In
terna
tion
al
Jo
urn
al
for
Pa
rasito
log
y3
8(
20
08
)1
21
9–
12
37
1221
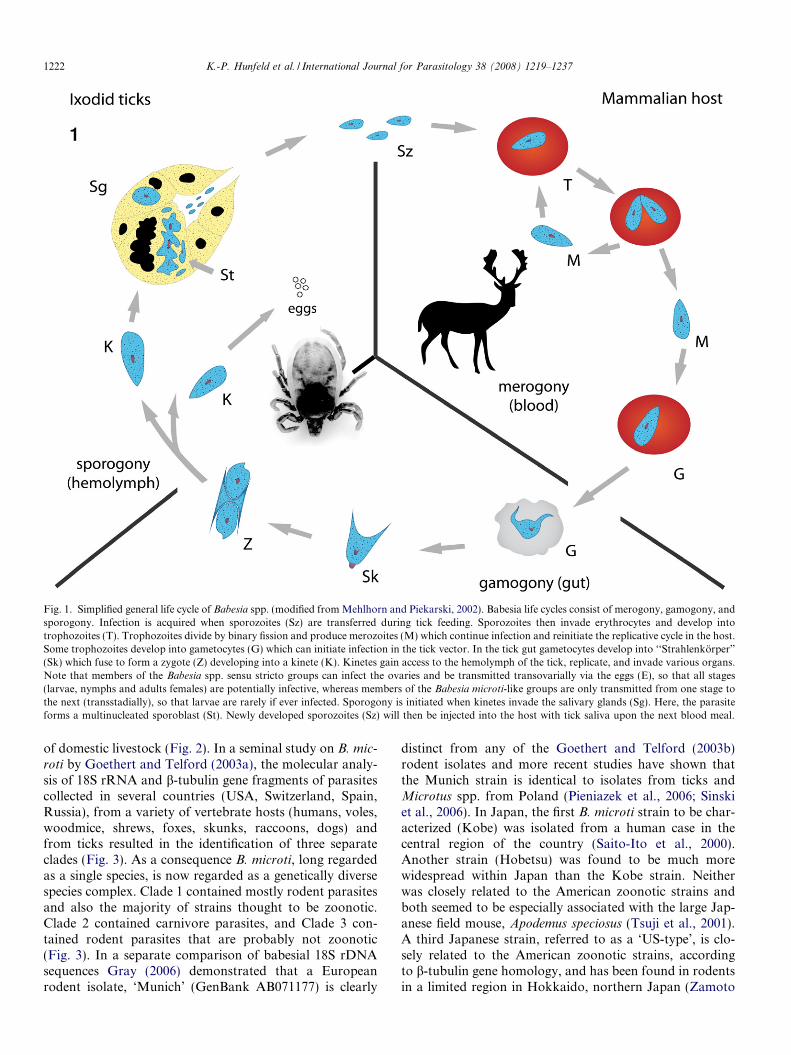
Fig. 1. Simplified general life cycle of Babesia spp. (modified from Mehlhorn and Piekarski, 2002). Babesia life cycles consist of merogony, gamogony, andsporogony. Infection is acquired when sporozoites (Sz) are transferred during tick feeding. Sporozoites then invade erythrocytes and develop intotrophozoites (T). Trophozoites divide by binary fission and produce merozoites (M) which continue infection and reinitiate the replicative cycle in the host.Some trophozoites develop into gametocytes (G) which can initiate infection in the tick vector. In the tick gut gametocytes develop into ‘‘Strahlenkorper”
(Sk) which fuse to form a zygote (Z) developing into a kinete (K). Kinetes gain access to the hemolymph of the tick, replicate, and invade various organs.Note that members of the Babesia spp. sensu stricto groups can infect the ovaries and be transmitted transovarially via the eggs (E), so that all stages(larvae, nymphs and adults females) are potentially infective, whereas members of the Babesia microti-like groups are only transmitted from one stage tothe next (transstadially), so that larvae are rarely if ever infected. Sporogony is initiated when kinetes invade the salivary glands (Sg). Here, the parasiteforms a multinucleated sporoblast (St). Newly developed sporozoites (Sz) will then be injected into the host with tick saliva upon the next blood meal.
1222 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
of domestic livestock (Fig. 2). In a seminal study on B. mic-
roti by Goethert and Telford (2003a), the molecular analy-sis of 18S rRNA and b-tubulin gene fragments of parasitescollected in several countries (USA, Switzerland, Spain,Russia), from a variety of vertebrate hosts (humans, voles,woodmice, shrews, foxes, skunks, raccoons, dogs) andfrom ticks resulted in the identification of three separateclades (Fig. 3). As a consequence B. microti, long regardedas a single species, is now regarded as a genetically diversespecies complex. Clade 1 contained mostly rodent parasitesand also the majority of strains thought to be zoonotic.Clade 2 contained carnivore parasites, and Clade 3 con-tained rodent parasites that are probably not zoonotic(Fig. 3). In a separate comparison of babesial 18S rDNAsequences Gray (2006) demonstrated that a Europeanrodent isolate, ‘Munich’ (GenBank AB071177) is clearly
distinct from any of the Goethert and Telford (2003b)rodent isolates and more recent studies have shown thatthe Munich strain is identical to isolates from ticks andMicrotus spp. from Poland (Pieniazek et al., 2006; Sinskiet al., 2006). In Japan, the first B. microti strain to be char-acterized (Kobe) was isolated from a human case in thecentral region of the country (Saito-Ito et al., 2000).Another strain (Hobetsu) was found to be much morewidespread within Japan than the Kobe strain. Neitherwas closely related to the American zoonotic strains andboth seemed to be especially associated with the large Jap-anese field mouse, Apodemus speciosus (Tsuji et al., 2001).A third Japanese strain, referred to as a ‘US-type’, is clo-sely related to the American zoonotic strains, accordingto b-tubulin gene homology, and has been found in rodentsin a limited region in Hokkaido, northern Japan (Zamoto

Fig. 2. Phylogenetic relationships of Babesia spp. using neighbour-joining analysis of the 18S rRNA gene (modified from Kjemtrup et al., 2006;reproduced from Gray and Weiss, 2008, with permission from Taylor and Francis Ltd).
Fig. 3. Phylogenetic relationships of Babesia microti strains using maximum-likelihood analysis (modified from Goethert and Telford, 2003a) of: (A) 18SrRNA gene; (B) b-tubulin gene (reproduced from Gray and Weiss, 2008, with permission of Taylor and Francis Ltd).
K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1223
et al., 2004a). This strain also occurs in South Korea, Vlad-ivostok in Russia, and Xinjiang in China, and appears toutilise a wider range of hosts, including insectivores, thanthe Hobetsu and Kobe strains (Zamoto et al., 2004b).The rather loosely constituted Clade 2 group of Goethertand Telford’s study (2003a) are all parasites of carnivores,
and the recently described B. microti-like parasite of dogsin Spain (GenBank AY534602, Theileria annae?) clearlybelongs to this group since it shows 100% 18S rDNAhomology to a Clade 2 parasite (GenBank AY144702)from a fox in Cape Cod, MA, USA (Fig. 3). The assigningof this Spanish dog parasite to the genus Theileria by

1224 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
Camacho et al. (2001) emphasises the lack of certainty inthe classification of B. microti-like parasites (Fig. 2).
At least 11 cases of human babesiosis have occurred inthe western USA caused by B. microti-like pathogens(Gray and Weiss, 2008). Recently, isolates from Washing-ton State (WA1) and from California (CA5) were charac-terized in detail and the parasite named as B. duncani
(Conrad et al., 2006). B. duncani shows only subtle mor-phological differences from B. microti, but analysis of the18S rDNA clearly demonstrated that it belongs to a sepa-rate phylogenetic group, together with isolates from dogsand wildlife, and was furthermore indistinguishable fromtwo other human isolates, CA6 and WA2-clone 1, thoughdistinguishable from the dog isolate B. conradae (Fig. 1).The manifestations of the B. duncani disease are very sim-ilar to those caused by B. microti but unlike B. microti, thereservoir hosts and vectors of B. duncani are unknown atpresent. In addition, uncharacterized B. microti-like babe-sias in human patients have been recorded in Brazil, China,Egypt, Mexico, South Africa and Taiwan (Gorenflot et al.,1998), and most recently in India (Marathe et al., 2005).
2.2. New developments in the phylogeny of Babesia sensustricto human pathogens
New phylogenetic information has also emerged forother recently recognised zoonotic Babesia spp. such asB. venatorum (EU1-3) in Europe (Herwaldt et al., 2003;Haselbarth et al., 2007) and B. divergens-like organisms(Gray, 2006) identified in the US based on 18S rDNAand ITS2 sequence analysis. Phylogenetic analysis of B.venatorum clearly demonstrates that it clusters togetherwith B. odocoilei, a parasite of deer from the US, and thesetwo organisms form a sister group with B. divergens
(Fig. 4). The clustering of these organisms was identical,regardless of which phylogenetic method was used (Her-waldt et al., 2003). B. divergens-like parasites isolated fromhumans in the US: MO1 from Missouri (Herwaldt et al.,1996); Washington State (Herwaldt et al., 2004); Kentucky(Beattie et al., 2002) are so close to B. divergens in terms of18S rDNA homology (Gray, 2006) they were sometimesreferred to as B. divergens. The Kentucky parasite appearsto be identical for the 18S rRNA gene to the Nantucketcottontail rabbit ‘B. divergens’ described by Goethert andTelford (2003b) and also to the Missouri parasite (MO1)(Holman et al., 2005; Gray, 2006). However, all bovine iso-lates of B. divergens, including resequenced isolates of theoriginal GenBank depositions, were found to be identicalfor this gene (Herwaldt et al., 2003; Slemenda et al., unpub-lished) so that parasites showing less than 100% homologyshould perhaps not be considered B. divergens. Indeed, arecent study by Holman (2006) on the cottontail rabbit iso-late, involving differential infectivity for cattle, parasite sizeand morphology in vitro, host erythrocyte specificityin vitro and ribosomal RNA gene sequences suggest thatB. divergens sensu stricto is not endemic to the US. At leasttwo European isolates from human cases that were identi-
fied at the time as B. divergens, also show less than 100%homology with the B. divergens 18S rDNA (Olmedaet al., 1997; Centeno-Lima et al., 2003) so that their iden-tity as B. divergens is questionable. This could mean thatthere are at least three B. divergens-like parasites with zoo-notic potential in Europe. Another human case occurredrecently in Korea (Kim et al., 2007) and this parasite, des-ignated KO1, was found to be most closely related to Babe-
sia spp of sheep that were described by Liu et al. (2007).This large babesia is clearly separate from all the otheragents of human babesiosis so far analysed, as shown inthe phylogenetic tree constructed by Kim et al. (2007).
3. Reservoir hosts and tick vectors
Babesia microti is now recognised as a diverse speciescomplex, parasitising a variety of hosts including rodents,insectivores and carnivores but the majority of the zoonoticstrains utilise microtine rodents as reservoir hosts (Goet-hert and Telford, 2003a). In the US the white-footed mousePeromyscus leucopus, is the main reservoir host and thevector is the human-biting Ixodes scapularis, the deer orblack-legged tick (Spielman, 1976). Other vectors of vari-ous strains of B. microti include Ixodes spinipalpis, Ixodes
angustus and Ixodes muris which transmit the parasites tovarious species of voles. However, these ticks do not bitehumans and the zoonotic potential of the strains of B. mic-
roti that they transmit is unknown. The vectors and reser-voirs of the main west coast US zoonotic babesia, B.
duncani, are not known.European strains of B. microti also parasitise a variety of
microtine rodents and two vectors are involved. Ixodes tri-
anguliceps is a specialised rodent tick that rarely if everbites man and is probably responsible for the transmissionof B. microti throughout Europe. Ixodes ricinus, which isclosely related to I. scapularis, and is well known to infesthumans, was identified as a vector of B. microti in Ger-many by Walter (1984), with the field vole, Microtus agres-
tis as the natural reservoir. B. microti transmission of aEuropean strain by I. ricinus using Mongolian gerbils inthe laboratory was confirmed by Gray et al. (2002) andthe parasite was detected in I. ricinus specimens collectedfrom vegetation in several countries (Gray and Weiss,2008). It seems likely, therefore, that this tick species isresponsible, for transmission to humans as evidenced byreports of positive serology (Hunfeld et al., 2002) and a sin-gle confirmed case (Hildebrandt et al., 2007). The only vec-tor implicated in transmission of B. microti in Japan, wherean autochthonous human case transmitted by blood trans-fusion was reported (Saito-Ito et al., 2000), is Ixodes ova-
tus, specimens of which were found in tick salivaryglands (Saito-Ito et al., 2004; Yano et al., 2005), althoughneither of the two types of B. microti that were detectedmatched the zoonotic (Kobe) strain. Microtine rodentsare the likely reservoir hosts.
The reservoir host for B. divergens sensu stricto, which isimplicated in most cases of human babesiosis in Europe, is

Fig. 4. Phylogenetic tree calculated for the 18S rDNA sequences as available from GenBank for selected Babesia spp. and for the newly detected babesiaorganism (EU3). The tree was computed by using the Clustal method with weighted residue weight table (adopted from Haselbarth et al., 2007).
K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1225
cattle and the vector for this parasite is I. ricinus. However,B. divergens-like parasites have also been described fromdeer and from I. ricinus (Duh et al., 2001, 2005; Bonnetet al., 2007) and these parasites may have caused someinfections attributed to B. divergens sensu stricto. The stud-ies by Duh et al. (2001, 2005) and Bonnet et al. (2007) sug-gest strongly that the newly described B. venatorum, whichhas caused infections in Austria, Italy, and Germany (Her-waldt et al., 2003; Haselbarth et al., 2007) also have deer asa reservoir host and I. ricinus as a vector. The vector andreservoir host for the B. divergens-like parasite in a humancase from the Canary Isles are unknown (Olmeda et al.,1997). I. ricinus is not thought to be involved but a closerelative, Ixodes ventalloi, occurs there and may be thevector.
In the US, the reservoir hosts for the B. divergens-likeisolates from Missouri (MO1) and from Kentucky areprobably species of cottontail rabbits in view of the find-ings of Goethert and Telford (2003b) in Nantucket. Thevectors are unknown but it is likely that Ixodes dentatus
maintains the infection in the Nantucket rabbit population.Neither vector nor reservoir host are known for the B. div-ergens-like parasite isolated from a human case in Wash-ington State (Herwaldt et al., 2004), though interestingly,parasites from Texan black-tailed jackrabbits showed99.9% homology with this isolate (Yabsley et al., 2006).
4. Human disease due to Babesia spp
To date, seven distinct babesia parasites have beenfound to cause human babesiosis (Table 1): B. microti
and related organisms, B. divergens, B. bovis, B. canis,B. duncani, B. venatorum, and a novel type of Babesia sp.similar to ovine babesias provisionally named KO1 (CalvoDe Mora et al., 1985, Marsaudon et al., 1995; Gorenflotet al., 1998; Homer et al., 2000; Gray and Weiss, 2008;Hildebrandt et al., in press). Most patients infected withBabesia spp. sensu stricto share splenectomy as a risk fac-tor for acquiring the disease. In addition, for all babesiainfections advanced age and depressed cellular immunity
are associated with a higher risk of symptomatic infectionand more severe illness (Telford and Maguire, 2006). Thisis why, the rising number of HIV positive individuals andthe increasing population of immunocompromised individ-uals may serve to boost the number of human babesiosiscases (Kjemtrup and Conrad, 2000; Haselbarth et al.,2007; Hildebrandt et al., 2007; Karp and Auwaerter, 2007).
Humans acquire the disease through tick bites or trans-fusion of contaminated blood products (Homer et al.,2000). An exceptional way of infection which is rarelyobserved in humans is transplacental transmission (Kjemt-rup and Conrad, 2000). So far, only two confirmed humancases have been documented (Esernio-Jenssen et al., 1987;New et al., 1997). In case of maternal infection, treatmentcan successfully prevent infection of the foetus (Raucheret al., 1984).
The clinical features in patients with babesiosis varysubstantially from asymptomatic to life threatening,depending on the conditions of the patient and the parasiteinvolved. In general, patients of all ages including childrenare affected, but most present clinically in their 40s to 60s(Mylonakis, 2001; Hunfeld and Brade, 2004). For B. mic-roti children and adults are infected at similar frequencies,but the proportion of symptomatic infections in adults ishigher (Krause et al., 2003). Peak transmission occurs fromMay to September and incubation periods vary from five to33 days after a tick bite. However, most individuals do notremember a tick infestation (Homer et al., 2000; Hunfeldand Brade, 2004; Haselbarth et al., 2007; Hildebrandtet al., in press). In transfusion-transmitted cases or inimmunocompromised individuals, cases may arise at anytime of the year and incubation periods can be much longerdepending on the exact time point and means of transmis-sion (Haselbarth et al., 2007; Hildebrandt et al., in press).In immunocompetent individuals parasitemia can hardlybe detected. Patients may show up with, in part, non-spe-cific symptoms like fever, flu-like disease, headache, chills,sweats, and myalgia (Table 2). Clinical diagnosis of humanbabesiosis can be further complicated by long persistenceof subclinical infections (Krause et al., 1998) and in Europe

1226 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
and the US may underlie other tick-borne diseases (Hun-feld and Brade, 2004; Swanson et al., 2006), particularlyLyme borreliosis (Table 2). Symptoms, however, usuallyabate within a few weeks (Leiby, 2006). In HIV patientsfever may occur for more than 4 weeks and show high par-asitemias (>30%), and the parasite may persist despiteactive treatment (Kjemtrup and Conrad, 2000). Clinicalsymptoms in immunocompromised patients include highfever (up to 40 �C), high parasitemia (20–80%) diaphoresis,severe anemia, shortness of breath, weakness, and fatigue.Later, patients may develop more specific symptoms likejaundice, dark urine, CNS involvement, or complicationslike congestive heart failure, and respiratory distress syn-drome (Homer et al., 2000; Hatcher et al., 2001; Mylona-kis, 2001; Herwaldt et al., 2003; Hunfeld and Brade,2004; Hildebrandt et al., 2007). Babesia microti infectionsmay persist despite multiple courses of treatment andmay be associated with relapsing symptoms for more thana year in immunocompromised individuals as described ina recent case control study (Krause et al., 2008). A recentstudy on 139 patients with B. microti infection in NewYork State revealed that 25% required hospitalisationand 6.5% died. On average, a 12- to 14-day delay afterthe onset of symptoms was noted before initiation ofappropriate antibiotic treatment (White et al., 1998).
Infections with B. divergens generally present as fulmin-ant, life-threatening infections. To date, about 30 cases ofB. divergens infections have been reported in Europe pre-dominantly in asplenic patients (Homer et al., 2000; Zintlet al., 2003; Hildebrandt et al., in press). Symptoms appearrapidly within 1–3 weeks post infection with septic fever,hemoglobinuria, or jaundice due to severe hemolysis, andup to 42% of patients die (Homer et al., 2000; Leiby,
Table 2Clinical symptoms associated with babesiosis and/or Lyme borreliosis(modified from Homer et al., 2000)
Symptom % of patients exhibiting the indicated symptoms
Babesiosis(n = 10)
LD(n = 214)
Both infections(n = 26)
Fatigue 60 49 81Headache 60 42 77Erythema migrans
(EM)0 85 62
Fever 80 42 58Sweats 20 11 46Chills 50 23 42Myalgia 20 31 38Anorexia 10 14 31Arthralgia 50 36 27Nausea 10 5 23Neck stiffness 30 21 23Multiple EM 0 14 19Cough 20 10 15Sore throat 20 9 15Conjunctivitis 0 3 12Splenomegaly 10 0 8Vomiting 0 4 8Joint swelling 0 3 4
2006). In contrast to the findings in human B. divergens
infections, the first cases of B. venatorum babesiosis in Italyand Austria, which occurred in two asplenic men withHodgkin’s disease and large B-cell lymphoma, respectively,were different in that the disease manifestations rangedfrom mild to moderately severe and both patients werecured after successful chemotherapy with clindamycinand/or quinine (Herwaldt et al., 2003). Likewise, infectionswith the newly recognised B. divergens-like parasite thatoccurred in Washington State and a case of babesiosis inKorea due to a babesia parasite closely related to ovinebabesias and provisionally named KO1, took a morebenign clinical course and the patients recovered aftertreatment.
Similar to the Italian and Austrian cases, clinical symp-toms in the first German B. venatorum-infected patientincluded elevated body temperature, chills, anemia, weak-ness, fatigue, anorexia, and headache followed by jaundiceand dark urine from hemoglobinuria (Haselbarth et al.,2007; Hildebrandt et al., in press). There was significantdelay between the onset of symptoms and the initiationof specific chemotherapy. Such a delay can be fatal becausepatients can deteriorate due to congestive heart failure,intravascular coagulation, renal failure, and respiratorydistress syndrome if left untreated (Homer et al., 2000;Mylonakis, 2001). Therefore, such patients, most of whomare immunocompromised or asplenic must be regarded asmedical emergencies requiring immediate treatment toarrest hemolysis and to prevent renal failure (Gelfand,2000; Homer et al., 2000). The recently described Germancase due to B. venatorum, however, was unique in that thepatient remained seronegative for specific antibodies forseveral months and suffered from relapse after initial treat-ment. Moreover, re-treatment with atovaquone and azi-thromyin did not clear the parasite and low levelparasitemia persisted for several months despite mainte-nance therapy with atovaquone, possibly due to the previ-ous combined application of rituximab and prednisolonewhich have highly immunosuppressive effects (Haselbarthet al., 2007). Corticosteroids and depressed cellular immu-nity are associated with severe human babesiosis (Rosneret al., 1984; Meldrum et al., 1992; Telford and Maguire,2006). In such cases, monitoring of parasitemia by bloodsmear examination and PCR analysis, and clinical long-term follow-up is important (Haselbarth et al., 2007; Hilde-brandt et al., in press). Clinicians should be aware that inthese patients relapse and persistence of the parasite mayoccur despite treatment (Hildebrandt et al., in press;Krause et al., 2008).
4.1. Pathogenesis and immunobiology
Erythrocytic parasitism by babesia leads to anemia,hyperbilirubinuria, hemoglobinuria possibly followed bykidney failure, adult respiratory distress syndrome(ARDS), and central nervous system impairment (Hunfeldand Brade, 2004; Telford and Maguire, 2006). Upon

K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1227
examination, infected erythrocytes reveal membrane alter-ations (Sun et al., 1983; Krogstad et al., 1991) resulting,in animals, in reduced survival and enhanced erythrocyteremoval upon spleen passage (Commins et al., 1988). Thisfinding parallels the observation that asplenia is an impor-tant risk factor for severe babesiosis in humans and ani-mals (Hunfeld and Brade, 2004). However, people withintact spleens have died, and people without spleens haverecovered from severe babesiosis, even without specifictreatment (Rosner et al., 1984).
Most of what is known to date on babesia pathobiologycomes from in vitro experiments and animal studies(mainly in mice and cattle) on B. microti, B. bovis, and B.
divergens (Homer et al., 2000; Kjemtrup and Conrad,2000; Zintl et al., 2003). In many infections no isolates havebeen obtained and little information is available on thepathobiology of recently recognised babesia parasites suchas KO1, and B. venatorum (EU1). Humoral immunity,however, appears to be less important than cellular immu-nity in controlling babesiosis as demonstrated for examplein a mouse model of B. microti infection because B-celldeficient mice remain less susceptible to infection, whereasT-cell receptor-deficient mice are readily infected (Clawsonet al., 2002; Telford and Maguire, 2006). Immunologicalstudies on mice also indicate an important role of CD4+T cells in controlling parasitemia (Hemmer et al., 2000).This corresponds to the known difficulties of individualswith depressed cellular immunity to control persistent par-asitemia (Kjemtrup and Conrad, 2000; Telford and Magu-ire, 2006; Haselbarth et al., 2007; Hildebrandt et al., inpress). Similarly, depletion of host macrophages and natu-ral killer cells increases susceptibility to infection (Aguilar-Delfin et al., 2003). Animal studies with B. duncani showoverwhelming production of pro-inflammatory cytokinessuch as TNF-a and interferon-c suggesting that the patho-biology mainly results from the host response and not fromthe parasite itself. In humans, symptoms occur at parasite-mias of less than 1% and experiments on several Babesiaspp. suggest that an excessive host immune response is animportant pathogenetic cofactor for severe babesiosis,thereby resembling fulminate malaria due to Plasmodium
falciparum except for cerebral symptoms (Krause et al.,2007; Gray and Weiss, 2008).
Correspondingly, genetic disruption of the TNF-a path-way as well as CD4 and CD8 gene knockout in animalslowered the fatality risk whereas cd T-cell knockout miceand controls readily died after B. duncani infection (Hem-mer et al., 2000; Telford and Maguire, 2006). This suggestscontribution of CD8+ T cells to the pathobiology of B.
duncani infections (Hemmer et al., 2000; Telford andMaguire, 2006). Antibody-derived immunity is less impor-tant as passive transfer of immune sera to SCID and nudemice fails to protect such animals against challenge with B.
microti (Matsubara et al., 1993).Other authors, however, demonstrated that some degree
of immunity to B. microti and B. rodhaini can be transferredto cattle and mice with serum containing anti-babesia
antibodies although it could not prevent infection orultimate death (Mahoney, 1967; Abdalla et al., 1978).Whether such data hold true for human infection with newlyrecognised babesias requires further research. It is interest-ing to note though, that in a recently described B. venato-
rum-infection it took the patient 7 months to finallydevelop babesia-specific antibodies following prednisolonetreatment and rituximab induced depletion of CD20 posi-tive cells (Haselbarth et al., 2007). With the appearance ofthe antibodies the patient started clearing the parasitesand smears turned negative. Six weeks later, PCR turnednegative also. Although not for certain, the production ofanti-babesia antibodies obviously contributed to clearingthe parasite in this patient (Haselbarth et al., 2007). Thehypothesis of a significant role of the humoral immuneresponse for the successful control of babesia infections isfurther supported by the data presented in a recent casecontrol study on immunocompromised patients that alsoidentified rituximab as an important risk factor for persis-tent and relapsing babesiosis (Krause et al., 2008).
5. Laboratory diagnostics in human babesiosis
Besides a general lack of awareness, the major problemwith diagnosing babesia infections in humans is that conve-nient and well evaluated diagnostic tools designed for rapidand reliable detection of such pathogens are not yet readilyavailable to most routine diagnostic laboratories. Giemsa-stained slides are important in endemic areas where thediagnostic services are experienced. Outside endemic areas,however, Giemsa-stained blood smears are not regularlyperformed in patients with fever of unknown origin (Hilde-brandt et al., in press). Moreover, automated blood analy-sers cannot always reliably detect infected erythrocytes sothat diagnosis of babesiosis can be missed (Bruckneret al., 1985; Hildebrandt et al., in press). Laboratorytesting in apparent cases of human babesiosis may shownon-specific findings such as elevated transaminases,alkaline phosphatases, unconjugated bilirubin, and lacticdehydrogenase. Normochromia, normocytic anemia,thrombocytopenia, and, occasionally, leucopenia may alsobe observed, probably owing to a TNF-mediated immuneresponse similar to that seen in severe cases of malaria(Krause and Telford, 1999; Gelfand, 2000; Homer et al.,2000; Sharan and Krause, 2000; Mylonakis, 2001). A posi-tive Coombs test in combination with hemolytic anemiaand elevated procalcitonine levels is highly suspicious ofbabesiosis (Haselbarth et al., 2007; Hildebrandt et al., inpress) and should prompt further diagnostic tests.
5.1. Direct detection of parasites
Thus far, microbiological diagnosis of human babesiosisin general has been based on the detection of the parasitesin Giemsa-stained thin blood smears (Fig. 5). However,early in the infection the level of parasitemia in the non-immunocompromised host can be lower than 1% (Krause

1228 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
et al., 1998; Gelfand, 2000; Homer et al., 2000; Sharan andKrause, 2000; Mylonakis, 2001). In these patients, serialblood smears must be investigated. On the other hand, highlevel parasitemia ranging from 50–80% may be seen in sple-nectomized individuals (Kjemtrup and Conrad, 2000). InGiemsa-stained blood smears Babesia spp. appear asintra-erythrocyte ring forms or pyriform inclusions withlight blue cytoplasm (Fig. 5). Malaria is the most importantdifferential diagnosis as Plasmodium spp. may also showintraerythrocytic rings (Fig. 5). In malaria parasitic pig-ment (hemozoin) is usually detectable, but early parasiticstages may lack pigment and sometimes co-infections withboth agents cannot be ruled out (Bush et al., 1990). In suchcases, Babesia spp. may be easily misdiagnosed as Plasmo-dium spp. and vice versa (Fig. 5). This remains especiallytrue in malaria endemic regions or in travelers returningfrom areas where malaria is known to occur but wherebabesiosis has never been reported before.
In B. microti infections erythrocytes often show severalparasites that appear as rings, sometimes with incompleteclosures. Basket-shaped merozoites (Fig. 5) can be approx-imately 1.5–2.5 lm in size. Although B. microti is consid-ered a diverse species complex, parasites appearmonomorphic in human blood smears (Telford and Goet-hert, 2004). In B. divergens and also in B. capreoli, and B.
duncani infections, parasites are pleomorphic with accoleforms, and paired pyriform ring forms (Fig. 5). Merozoitescan be 1–3 lm in size. The morphology of intraerythrocyticB. divergens parasites (Fig. 5) is very similar to B. duncani,B. venatorum, and B. microti, except for the fact that in B.divergens and B. venatorum paired forms are typicallyfound at an obtuse or diverging angle usually in the centreof human erythrocytes (Telford et al., 1993; Gorenflotet al., 1998) (Fig. 5). Tetrad forms are more common inB. microti infections but are still relatively rare (Gray andWeiss, 2008; Hildebrandt et al., in press). Consequently,although helpful for diagnostic purposes, microscopy ofparasites in blood smears without additional molecularanalysis of the pathogen is not reliable for speciesidentification.
5.2. Detection of Babesia spp. by animal inoculation and
in vitro culture
Babesiosis can also be confirmed by directly inoculat-ing 1.0 ml ETDA whole blood into the peritoneum ofgolden hamsters, jirds, or mice (Krause and Telford,1999; Homer et al., 2000; Sharan and Krause, 2000).Blood smears of the inoculated animal usually becomepositive within 2–4 weeks. This is much too long to waitin acute human cases. Moreover, parasitemia in infectedanimals can be transient. Therefore, blood smears needto be monitored at least daily which is time-consuming.As such, animal inoculation is not useful in emergency sit-uations but still has its place in the diagnosis of crypticchronic babesia infections and especially for genomicstudies (Hildebrandt et al., in press). Similarly, bioassays
are not suitable for parasites that do not provoke parasi-temia in laboratory animals (Telford and Maguire, 2006).For example, jirds are highly susceptible to infection withB. divergens, but could not be infected by inoculation ofhuman blood containing B. venatorum in all three casesreported so far (Herwaldt et al., 2003; Haselbarth et al.,2007). In cases involving parasites where suitable animalhosts are not yet known, such Babesia spp. can be culti-vated directly from blood by in vitro culture using autol-ogous erythrocyte preparations (Malandrin et al., 2004;Bonnet et al., 2007). Such cultures then need to be mon-itored for parasitemia by examination of blood smears. Ifpositive, parasites can be adapted to culture in bloodfrom appropriate animal species after several passages(Chauvin et al., 2002). These laboratory methods, how-ever, are not readily available in most routine diagnosticlaboratories facing a first case of babesiosis.
5.3. Indirect detection of babesia infection by antibody
testing
IFAT for B. microti is the only serological test forhuman babesiosis that has so far been standardised toany extent (Homer et al., 2000; Kjemtrup and Conrad,2000; Hunfeld and Brade, 2004). Studies suggest B. microti-specific IFAT to be specific, sensitive, and reproduciblewith reported test specificities ranging from 90% to 100%(Krause et al., 1994; Gelfand, 2000; Homer et al., 2000;Hunfeld et al., 2002). Moreover, titres from 1:32 to 1:160are diagnostic and specific, with positive predictive valuesof 69–100% and negative predictive values of 96–99% (Kra-use et al., 1994). For IFAT based on B. divergens antigenand the more recently encountered parasites such dataare lacking. A B. duncani IFAT was used to screen poten-tially infected individuals (Kjemtrup and Conrad, 2000).However, a positive cutoff titer has not been established.
Specific IgG antibodies are found in patients with acuteand chronic babesia infections. The detection of IgM anti-bodies (>1:64) may indicate acute infection even in theabsence of demonstrable parasitemia but is less specificthan IgG antibody testing (Homer et al., 2000; Hunfeldet al., 2002). In immunocompromised patients with pro-longed prepatent periods a significant delay of seroconver-sion may occur (Telford and Maguire, 2006). Afterinfection titers persist from 13 months up to 6 years (Pers-ing et al., 1995).
Our knowledge of the serodiagnosis of human babesio-sis refers to B. microti-infected individuals mainly becauseobservations involving other Babesia spp. are based on casereports rather than on clinical studies (Hunfeld and Brade,2004). In acute cases, whether caused by B. divergens-likeparasites or B. microti in immunocompromised patients,IFAT are not reliable because seronegative results at dis-ease onset are common and may delay treatment (Persinget al., 1995; Haselbarth et al., 2007; Hildebrandt et al., inpress). Nevertheless, IFAT are important in demonstratingseroconversion to retrospectively confirm the diagnosis by
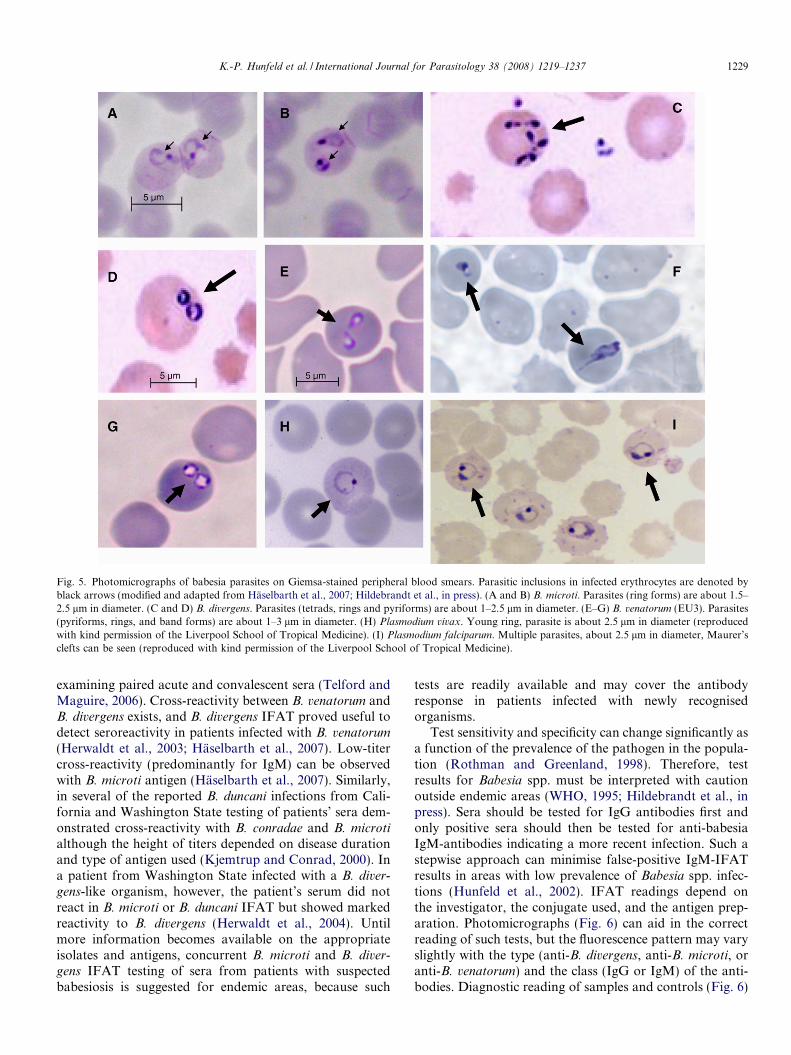
Fig. 5. Photomicrographs of babesia parasites on Giemsa-stained peripheral blood smears. Parasitic inclusions in infected erythrocytes are denoted byblack arrows (modified and adapted from Haselbarth et al., 2007; Hildebrandt et al., in press). (A and B) B. microti. Parasites (ring forms) are about 1.5–2.5 lm in diameter. (C and D) B. divergens. Parasites (tetrads, rings and pyriforms) are about 1–2.5 lm in diameter. (E–G) B. venatorum (EU3). Parasites(pyriforms, rings, and band forms) are about 1–3 lm in diameter. (H) Plasmodium vivax. Young ring, parasite is about 2.5 lm in diameter (reproducedwith kind permission of the Liverpool School of Tropical Medicine). (I) Plasmodium falciparum. Multiple parasites, about 2.5 lm in diameter, Maurer’sclefts can be seen (reproduced with kind permission of the Liverpool School of Tropical Medicine).
K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1229
examining paired acute and convalescent sera (Telford andMaguire, 2006). Cross-reactivity between B. venatorum andB. divergens exists, and B. divergens IFAT proved useful todetect seroreactivity in patients infected with B. venatorum
(Herwaldt et al., 2003; Haselbarth et al., 2007). Low-titercross-reactivity (predominantly for IgM) can be observedwith B. microti antigen (Haselbarth et al., 2007). Similarly,in several of the reported B. duncani infections from Cali-fornia and Washington State testing of patients’ sera dem-onstrated cross-reactivity with B. conradae and B. microtialthough the height of titers depended on disease durationand type of antigen used (Kjemtrup and Conrad, 2000). Ina patient from Washington State infected with a B. diver-
gens-like organism, however, the patient’s serum did notreact in B. microti or B. duncani IFAT but showed markedreactivity to B. divergens (Herwaldt et al., 2004). Untilmore information becomes available on the appropriateisolates and antigens, concurrent B. microti and B. diver-gens IFAT testing of sera from patients with suspectedbabesiosis is suggested for endemic areas, because such
tests are readily available and may cover the antibodyresponse in patients infected with newly recognisedorganisms.
Test sensitivity and specificity can change significantly asa function of the prevalence of the pathogen in the popula-tion (Rothman and Greenland, 1998). Therefore, testresults for Babesia spp. must be interpreted with cautionoutside endemic areas (WHO, 1995; Hildebrandt et al., inpress). Sera should be tested for IgG antibodies first andonly positive sera should then be tested for anti-babesiaIgM-antibodies indicating a more recent infection. Such astepwise approach can minimise false-positive IgM-IFATresults in areas with low prevalence of Babesia spp. infec-tions (Hunfeld et al., 2002). IFAT readings depend onthe investigator, the conjugate used, and the antigen prep-aration. Photomicrographs (Fig. 6) can aid in the correctreading of such tests, but the fluorescence pattern may varyslightly with the type (anti-B. divergens, anti-B. microti, oranti-B. venatorum) and the class (IgG or IgM) of the anti-bodies. Diagnostic reading of samples and controls (Fig. 6)

1230 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
should start with a magnification of 400� and, in case ofany doubt, be confirmed by checking the fluorescence pat-tern with a magnification of 1000� with oil (Fig. 6). This isdone to exclude rare non-specific fluorescence of babesialantigen which mainly involves the apex of the parasite (Hil-debrandt et al., in press). False positive results have beendescribed in Plasmodium spp. and Toxoplasma gondii infec-tions, and in lupus erythematosus or rheumatoid arthritis(Homer et al., 2000; Hildebrandt et al., in press).
ELISA and immunoblot can be used to confirm IFATresults and to identify babesia-positive carriers. ELISAdetection of anti-Babesia spp. antibodies is less subjectivethan that of IFAT and can be automated. A peptide-basedELISA has recently been described using secreted antigensof babesia parasites (Homer et al., 2003) but, comparedwith IFAT, greater amounts of antigen are required andtest specificity depends on optimised blocking conditionsand purification of the antigens (Zintl et al., 2003). More-over, tests such as ELISAs and immunoblot are not yet suf-ficiently standardised for routine application in diagnosticlaboratories (Ryan et al., 2001; Hunfeld and Brade, 2004;Gray and Weiss, 2008).
5.4. New tools for a more rapid molecular biological
diagnosis of Babesia spp. infection
The development of sensitive and specific multiplexPCR assays may be an important future improvement inthe laboratory diagnosis of human disease following singleor multiple infection with tick-borne pathogens (Hilde-brandt et al., in press). Detection may then be attemptedby use of a single diagnostic test covering agents such asBabesia spp., Borrelia spp., Anaplasma spp., and Rickettsia
spp. (Swanson et al., 2006). This seems attractive becausepatients suffering from Lyme disease, human babesiosis,human anaplasmosis, or most other tick-borne diseasescan all present with relatively non-specific influenza-like ill-nesses complicating any clinical differential diagnostic con-siderations (Swanson et al., 2006). With the development ofmore sensitive molecular techniques for diagnosing fastidi-ous organisms like Babesia spp. and Anaplasma spp., PCRis increasingly relied on to detect such rare pathogens inpatients with suspected infection but low parasite loadsand negative blood smears (Hunfeld and Brade, 2004;Swanson et al., 2006; Hildebrandt et al., in press).
In patients with suspected babesiosis, PCR results forBabesia spp. can be available within one day and as fewas three parasites in 100 ll blood can yield a positive result(Persing et al., 1992; Armstrong et al., 1998). Studies haveshown that PCR targeting the 18S rDNA (Fig. 6) is moresensitive than and equally specific as blood smear evalua-tion or bioassay in the detection of acute Babesia spp.infections (Homer et al., 2000). For purposes of qualitycontrol, the 12S rDNA of ticks or the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene can be used ashousekeeping genes to control for high quality of DNAextraction from ticks or human blood, respectively (Barber
et al., 2005; Casati et al., 2006). As mentioned above, suchnew rapid and sensitive PCR methods are also attractive incases of human babesiosis where the effectiveness of anti-microbial therapy has to be monitored. Detection of babe-sia DNA indicates a parasitemia and persistent DNAdetection clearly points to ongoing infection (Krauseet al., 1998; Homer et al., 2000; Haselbarth et al., 2007).Nevertheless, PCR suffers from limitations such as the dif-ficulties to distinguish between nucleic acids from viableand non-viable organisms and, therefore, reversion toPCR negativity may significantly lag behind a clinicalresponse to antimicrobial therapy (Persing, 1993). In thiscontext, the major advantage of detecting RNA insteadof DNA targets is a more direct correlation of rRNAand specific gene transcript quantities with growth of theorganism. This is why a more sensitive indicator of thera-peutic response in the near future may be to quantitativelymonitor for RNA target sequences such as rRNA ratherthen specific DNA because of their higher turnover (Hilde-brandt et al., in press). Whether PCR indeed can be usefulin monitoring the success of specific treatment in patientswith Babesia spp. infection and prolonged persistent para-sitemia, however, awaits further evidence from clinicalstudies (Haselbarth et al., 2007). For purposes of epidemi-ology and phylogeny, PCR technology and sequence anal-yses of the amplicons proved powerful in more exactspecies identification. Because a high degree of 18S rDNAsequence identity exists between many Babesia spp., thecomplete 18S rRNA gene (about 1700 bp) should alwaysbe analysed especially in newly recognised organisms(Gray, 2006; Hildebrandt et al., in press).
6. Treatment and outcome of babesiosis
Although babesiosis is a common disease in domesticanimals and many drugs have been developed for treat-ment, none have been adopted for routine use in humans,primarily because of licensing problems. Current knowl-edge on clinical course and treatment of human babesiosisis mainly derived from clinical data on B. microti and B.
divergens-infected patients. Several drugs have been testedin vitro against a variety of babesia species, but resultsfor pyrimethamine, tetracyclines, primaquine, and pen-tamidines have varied. Moreover, their use in humans atappropriate dosage is associated with significant side effectsand treatment failure. These drugs, therefore, have notbeen applied consistently in the clinical context (Pudneyand Gray, 1997; Homer et al., 2000; Gray and Weiss,2008). The anti-malaria drug chloroquine is regarded asineffective against Babesia spp. (Homer et al., 2000; Zintlet al., 2003). This remains true also for antimalarials suchas mefloquine and artemisinin at least in the animal model(Wittner et al., 1996; Marley et al., 1997). Imidocarb ishighly effective in vitro and has been used successfully totreat two Irish patients infected with B. divergens (Zintlet al., 2003) but has not been approved for general use inhumans (Mylonakis, 2001; Vial and Gorenflot, 2006). Since

Fig. 6. Direct and indirect diagnostic methods for detection of babesial infections (modified and adapted from Haselbarth et al., 2007; Hildebrandt et al.,in press). (A) Photomicrograph of positive B. divergens-IFAT, 400�. (B) Display detail 1000�, oil (zoomed). (C) Negative IFAT control. (D) Babesia spp.-specific PCR designed for amplification of a highly specific region of the 18S rRNA gene (Armstrong et al., 1998). Molecular weight of the fragment(437 bp) amplified by use of PIRO-A and PIRO-B primers in bp appears on the left together with the 123-bp marker. P – positive control, N – negativecontrol, S – patient sample, I – inhibition control, M – 123-bp marker.
K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1231
treatment of human babesiosis with a combination of qui-nine and clindamycin (Table 3) for seven to 10 days isapplied, disease outcome has dramatically improved(Rowin et al., 1982; Wittner et al., 1982). Quinine can beexchanged for quinidine and administered intravenouslyalong with clindamycin, if necessary (Vial and Gorenflot,2006). The mortality rate for clinically apparent B. microti
infections is �5% and most infections resolve on their own(Meldrum et al., 1992). Chemotherapy, thus, is indicatedonly in moderately to severely ill cases with such infections.Individuals with B. divergens infection, however, are usu-ally regarded as medical emergencies and require immedi-ate treatment to arrest hemolysis and prevent renalfailure. Many severe B. divergens infections in the pastended fatally with general organ failure 4–7 days afterthe manifestation of hemoglobinuria, and outcome datain severely ill individuals with asplenia suggest a mortalityrate of 42% (Gorenflot et al., 1998; Homer et al., 2000;Kjemtrup and Conrad, 2000; Zintl et al., 2003; Leiby,
2006). The in vivo effectiveness of quinine, however, hasalways been questioned and the drug related toxicity of thisdrug is significant (Brasseur et al., 1996). It is noteworthythat in recent years, a more favourable outcome wasincreasingly reported in B. divergens-infected patients withsevere complications even though they were not treatedwith a full course of quinine and clindamycin, mainlybecause of quinine side effects (Denes et al., 1999; Corpeletet al., 2005). These findings underscore the impact ofimproved adjunctive measures provided by modern inten-sive care medicine, including exchange transfusion whichis usually reserved for extremely ill individuals, i.e., thosewith massive hemolysis, asplenia, immunosuppression,and parasitemia of more than 10% (Mylonakis, 2001; Zintlet al., 2003), but has been recommended for all emergencycases involving B. divergens (Gorenflot et al., 1987).
This measure is particularly helpful because in humansbabesia have no exo-erythrocytic stages. Therefore, theremoval of parasitised erythrocytes is curative. In addition,

1232 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
anemia is corrected and toxic and harmful metabolites areremoved. Clindamycin monotherapy has been proposedwhen accompanied by such adjunctive measures (Brasseuret al., 1996; Denes et al., 1999; Corpelet et al., 2005). It isnotable, however, that in an experimental B. divergens/ger-bil model, atovaquone alone proved more effective than thebest animal antibabesial, imidocarb dipropionate, (Pudneyand Gray, 1997) and perhaps this drug should also be con-sidered for emergency treatment of B. divergens-like infec-tions (Table 3).
For treatment of B. microti infections, animal studiesshowed that azithromycin in combination with quinine(Weiss et al., 1993), azithromycin with atovaquone (Witt-ner et al., 1996), and atovaquone with clindamycin (Grayand Pudney, 1999), were all effective. More recently, ran-domized trials in humans infected with B. microti showedthat atovaquone plus azithromycin therapy was as effectiveas the standard quinine/clindamycin combination andthere were fewer side effects (15% versus 72%) (Krauseet al., 2000). In view of the low risk of side effects associ-ated with atovaquone/azothromycin and the possibility ofparasite persistence and occasional recrudescence, it hasbeen argued that all patients diagnosed with B. microti
infection be treated with this drug combination. In severecases similar adjunctive measures to those used for B. div-
ergens infections may be necessary. In HIV patients and inotherwise immunocompromised individuals substantiallyhigher dosage regimens and longer treatment schedulesmay be required for clearing the infection (Kjemtrup andConrad, 2000; Krause et al., 2008). Little is known so farabout the exact in vitro susceptibilities to potential antiba-besia drugs of the newly recognised Babesia spp. such as B.
duncani, B. venatorum or the B. divergens-like organismsfrom the US, though the cottontail rabbit babesia thoughtto be responsible for cases in Kentucky and Missouri has
Table 3Commonly used effective drugs for the treatment of human babesiosis (accordin2006)
Drug (generic name) Regular single dose (Adults, 70 kg)
Adults
Quinine 650 mgClindamycin 600 mgAzithromycin 500 mg/1st day, 250 mg theraftera
Atovaquone 750 mgDoxycycline 200 mg
Drug (generic name) Regular single dose/kg
Children
Quinine 8 mgc
Clindamycin 7–10 mgd
Azithromycinb 10 mg/1st day, 5 mg/day thereafterAtovaquone 20 mg/dayf
a In immunocompromised patients higher doses (600–1000 mg/day) may beb In immunocompromised patients higher dose may be required.c Maximum: 650 mg per dose.d Maximum: 600 mg per dose.e Maximum: 250 mg per dose.f Maximum: 750 mg per dose.
now been adapted to an in vitro system (Spencer et al.,2006). Having reviewed the currently available informationon the antimicrobial susceptibility data and in vivo treat-ment outcome available from in vitro susceptibility studies,case reports, and clinical investigations (Homer et al., 2000;Herwaldt et al., 2003; Vial and Gorenflot, 2006; Haselbarthet al., 2007) published so far, it appears that there is noconvincing scientific evidence for any clinically relevant dif-ferences in the susceptibilities of pathogenic Babesia spp.with regard to the therapeutic agents commonly used totreat human disease, though it is known that in animalmodels B. microti-like infections are more difficult to treatthan those caused by Babesia species s.s., such as B. diver-
gens (Gray and Pudney, 1999). The clindamycin and qui-nine combination appears to be the regimen of choice forhuman cases, despite problems with side effects and therequirement for aggressive adjunctive procedures in rapidlyfulminating infections. Most of the recent cases of humanbabesiosis caused by previously unknown babesias haveresponded well to this drug combination (Quick et al.,1993; Persing et al., 1995; Saito-Ito et al., 2000; Kjemtrupet al., 2002; Herwaldt et al., 2003, 2004; Conrad et al.,2006; Wormser et al., 2006; Haselbarth et al., 2007; Hilde-brandt et al., 2007; Kim et al., 2007). However, problemswith speed of response to therapy and also parasite persis-tence remain (Krause et al., 1998; Haselbarth et al., 2007),emphasising the importance of closely monitoring thecourse of parasitemia, the necessity for long-term follow-up in such patients, and the need for further antibabesialdrug research.
7. Babesia spp. and blood products
In recent years, transfusion medicine massively focusedon HIV, hepatitis B, and hepatitis C virus, whereas patho-
g to Telford and Maguire, 2006; Vial and Gorenflot, 2006; Wormser et al.,
Application Dosage regimen
p.o. 3 times dailyp.o., i.v. 3 times dailyp.o. Once dailyp.o. Twice dailyp.o. Once daily
Application Dosage regimen
p.o. 3 times dailyp.o., i.v. 3 times daily
e p.o. Once dailyp.o. Twice daily
required.

K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1233
gens such as babesia that represent a more scattered butongoing blood safety risk continue to ‘‘fly under the radar”
(Leiby, 2006). Characteristics of Babesia spp. clearly favorsuccessful transmission via blood products: First, once theyare introduced into a mammalian host they infect and rep-licate in red blood cells. Infected blood or residual erythro-cytes in platelet units, therefore, represent suitable vehiclesfor transmission to transfusion recipients (Homer et al.,2000; Leiby, 2006). Second, infected hosts may suffer fromlong-lasting subclinical parasitemia that is hard to detectand infected donors may be at risk for transmitting babesiafor extended periods of time (Krause et al., 1998). Lastly,based on data obtained from the analysis of transfusion-transmitted infections, parasites can survive for 21–35 daysunder standard blood storage conditions (Mintz et al.,1991; Eberhard et al., 1995; Leiby and Gill, 2004). So far,about 60 cases of transfusion-transmitted babesiosis havebeen reported in the literature (Leiby, 2006). Transfusion-transmitted cases involved blood recipients ranging in agefrom neonate to 79 years and most of the cases were attrib-uted to B. microti (Herwaldt et al., 1997; Kjemtrup et al.,2002; Pantanowitz et al., 2002; Della-Giustina et al.,2005; Leiby, 2006; Hildebrandt et al., 2007). Observedincubation periods for these cases ranged from 1 to 9weeks. The exact number of cases, however, is difficult toassess because there are no mandatory reporting systemsand frequently, cases of new infections are not publishedbecause they are not considered novel or noteworthy(Leiby, 2006). Few reliable estimates of transmission riskare currently available with regard to transfusion-acquiredbabesiosis. In two recent studies from endemic areas inConnecticut the risk of acquiring B. microti from a unitof red blood cells was reported to range between 1 in 601and 1 in 1800 and the risk after transfusion of a unit ofplatelets was 0 in 371 patients investigated (Gerber et al.,1994; Leiby, 2006). Increasing evidence suggests that thegeographical range of distribution of Babesia spp. appearsto be greater than initially thought and on a worldwidescale progressively more transfusion-transmitted cases arereported from areas where such infections were not knownto occur before (Herwaldt et al., 1997; Saito-Ito et al.,2000; Kjemtrup et al., 2002; Leiby, 2006; Hildebrandtet al., 2007). Moreover, newly recognised organisms suchas B. duncani in the US and B. microti-like parasites inCanada, Japan, and Germany are increasingly implicatedin transfusion-transmitted cases, particularly, in highly sus-ceptible patients who are immunocompromised, asplenic,and have received large amounts of blood (Saito-Itoet al., 2000; Kain et al., 2001; Leiby, 2006; Hildebrandtet al., 2007, in press). Clearly, the successful detectionand characterization of such blood-derived parasitesdepends on medical awareness and the availability ofsophisticated diagnostic methods such as PCR technology,serological tests, and culture techniques (Saito-Ito et al.,2000; Kain et al., 2001; Hildebrandt et al., 2007, in press).As such the currently reported distribution and frequencyof transfusion-acquired cases of babesiosis is probably
not representative and may instead be regarded as the tipof the iceberg when assessing the true dimension of theproblem (Hildebrandt et al., in press).
8. Prevention and vaccines
With increasing outdoor activities more people areexposed to ticks. However, I. ricinus nymphs can be easilyoverlooked and most infested persons do not remember atick bite (Leiby, 2006; Hildebrandt et al., in press). Unfor-tunately, antibiotic prophylaxis for human babesiosis hasnot been established and live vaccines are available for ani-mal babesiosis only (de Waal and Combrink, 2006). Thereis a need for effective recombinant vaccines against apicom-plexan pathogens including Babesia, Plasmodium, or Thei-leria spp. but development has proved unsuccessful so far(de Waal and Combrink, 2006). Therefore, prevention ofbabesia infection mainly relies on wearing appropriateclothing, application of repellents (DEET, permethrin),and the prompt removal of any ticks (Gray and Weiss,2008). With regard to prevention of transfusion-acquiredbabesiosis no licensed techniques are currently availableto prevent or reduce transmission of Babesia spp. (Zavizionet al., 2004; Leiby, 2006). PCR- or antibody screening ofdonors is not yet established because molecular and sero-logical diagnostics are not readily available. Moreover, reg-ular blood donor screening is not cost-effective even in theknown geographical areas of high endemicity in the US(Leiby, 2006). In summary, protective and preventive mea-sures against zoonotic babesia, to date, are limited andmust focus on clinical awareness in susceptible populationsafter tick exposure or after transfusion of blood products(Hunfeld and Brade, 2004; Leiby, 2006; Hildebrandtet al., in press) to readily diagnose and treat babesiainfections.
9. Conclusions
The diversity of Babesia spp. and the ubiquity of tickspoint to a high potential for Babesia spp. to further emergeas zoonotic pathogens on a worldwide scale. Concordantly,several new babesia parasites have recently been recognisedand cases of human babesiosis have been increasinglyreported in geographical areas where the presence of Babe-
sia spp. in enzootic cycles was obvious for decades butwhere the risk for humans of acquiring Babesia spp. eitherfrom ticks or from human blood products was not knownbefore (Hildebrandt et al., in press). Most importantly,human babesiosis may become progressively more impor-tant for the steadily increasing population of highly suscep-tible immunocompromised individuals and should beregularly considered in the differential diagnosis of infec-tion or fever of unknown origin, especially, in splenectom-ized patients and in individuals with recent transfusion ofblood products. The need for better diagnostics is obviousand laboratories require broad access to reliable tests suchas IFAT and PCR for timely diagnosis in suspected cases.

1234 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
Better molecular detection and strain typing of parasites isalso necessary to clarify the epidemiology of zoonoticBabesia spp. and whether their virulence or enhancedtransmissibility is potentially strain-dependent. Furtherstudies are therefore urgently needed to better characterizethe distribution and medical relevance of these pathogensin many parts of the world.
References
Abdalla, H.S., Hussein, H.S., Kreier, J.P., 1978. Babesia rodhaini: passiveprotection of mice with immune serum. Tropenmed. Parasitol. 29,295–306.
Aguilar-Delfin, I., Wettstein, P.J., Persing, D.H., 2003. Resistance to acutebabesiosis is associated with interleukin-12- and gamma interferon-mediated responses and requires macrophages and natural killer cells.Infect. Immun. 71, 2002–2008.
Armstrong, P.M., Katavolos, P., Caporale, D.A., Smith, R.P., Spielman,A., Telford 3rd, S.R., 1998. Diversity of Babesia infecting deer ticks(Ixodes dammini). Am. J. Trop. Med. Hyg. 58, 739–742.
Babes, V., 1888. Sur l’hemoglobinurie bacterienne du boeuf. Compt.Rend. Acad. Sci. Ser. III Sci. Vie. 107, 692–694.
Barber, R.D., Harmer, D.W., Coleman, R.A., Clark, B.J., 2005. GAPDHas a housekeeping gene: analysis of GAPDH mRNA expression in apanel of 72 human tissues. Physiol. Genomics 21, 389–395.
Beattie, J.F., Michelson, M.L., Holman, P.J., 2002. Acute babesiosiscaused by Babesia divergens in a resident of Kentucky. N. Engl. J.Med. 347, 697–698.
Bonnet, S., Jouglin, M., L’Hostis, M., Chauvin, A., 2007. Babesia sp. EU1from roe deer and transmission within Ixodes ricinus. Emerg. Infect.Dis. 13, 1208–1210.
Brasseur, P., Lecoublet, S., Kapel, N., Favennec, L., Ballet, J.J., 1996.Quinine in the treatment of Babesia divergens infections in humans.Eur. J. Clin. Microbiol. Infect. Dis. 15, 840–841.
Bruckner, D.A., Garcia, L.S., Shimizu, R.Y., Goldstein, E.J., Murray,P.M., Lazar, G.S., 1985. Babesiosis: problems in diagnosis usingautoanalyzers. Am. J. Clin. Pathol. 83, 520–521.
Bush, J.B., Isaacson, M., Mohamed, A.S., Potgieter, F.T., de Waal, D.T.,1990. Human babesiosis – a preliminary report of 2 suspected cases inSouth Africa. S. Afr. Med. J. 78, 699.
Calvo De Mora, A., Garcia Castellano, J.M., Herrera, C., Jimenez-Alonso, J., 1985. Babesiosis humana: aportacion de un caso deevolucion fatal. Medicina Clinica 85, 515–516.
Camacho, A.T., Pallas, E., Gestal, J.J., Guitian, F.J., Olmeda, A.S.,Goethert, H.K., Telford, S.R., 2001. Infection of dogs in north-westSpain with a Babesia microti-like agent. Vet. Rec. 149, 552–555.
Casati, S., Sager, H., Gern, L., Piffaretti, J.C., 2006. Presence ofpotentially pathogenic Babesia sp. for human in Ixodes ricinus inSwitzerland. Ann. Agric. Environ. Med. 13, 65–70.
Centeno-Lima, S., do Rosario, V., Parreira, R., Maia, A.J., Freudenthal,A.M., Nijhof, A.M., Jongejan, F., 2003. A fatal case of humanbabesiosis in Portugal: molecular and phylogenetic analysis. Trop.Med. Int. Health 8, 760–764.
Chauvin, A., Valentin, A., Malandrin, L., L’Hostis, M., 2002. Sheep as anew experimental host for Babesia divergens. Vet. Res. 33, 429–433.
Cho, S.H., Kim, T.S., Lee, H.W., Tsuji, M., Ishihara, C., Kim, J.T., Wee,S.H., Lee, C.G., 2002. Identification of newly isolated Babesia
parasites from cattle in Korea by using the Bo-RBC-SCID mice.Korean J. Parasitol. 40, 33–40.
Clawson, M.L., Paciorkowski, N., Rajan, T.V., La Vake, C., Pope, C., LaVake, M., Wikel, S.K., Krause, P.J., Radolf, J.D., 2002. Cellularimmunity, but not gamma interferon, is essential for resolution ofBabesia microti infection in BALB/c mice. Infect. Immun. 70, 5304–5306.
Collett, M.G., 2000. Survey of canine babesiosis in South Africa. J. S. Afr.Vet. Assoc. 71, 180–186.
Commins, M.A., Goodger, B.V., Waltisbuhl, D.J., Wright, I.G., 1988.Babesia bovis: studies of parameters influencing microvascular stasis ofinfected erythrocytes. Res. Vet. Sci. 44, 226–228.
Conrad, P.A., Kjemtrup, A.M., Carreno, R.A., Thomford, J., Wain-wright, K., Eberhard, M., Quick, R., Telford 3rd, S.R., Herwaldt,B.L., 2006. Description of Babesia duncani n.sp. (Apicomplexa:Babesiidae) from humans and its differentiation from other piroplasms.Int. J. Parasitol. 36, 779–789.
Corpelet, C., Vacher, P., Coudore, F., Laurichesse, H., Conort, N.,Souweine, B., 2005. Role of quinine in life-threatening Babesia
divergens infection successfully treated with clindamycin. Eur. J. Clin.Microbiol. Infect. Dis. 24, 74–75.
Criado-Fornelio, A., Martinez-Marcos, A., Buling-Sarana, A., Barba-Carretero, J.C., 2003. Molecular studies on Babesia, Theileria andHepatozoon in southern Europe. Part II. Phylogenetic analysis andevolutionary history. Vet. Parasitol. 114, 173–194.
de Waal, D.T., Combrink, M.P., 2006. Live vaccines against bovinebabesiosis. Vet. Parasitol. 138, 88–96.
Della-Giustina, D., Laird Jr, T.W., Smith, T., 2005. Transfusion-acquiredbabesiosis in a nonendemic area. Mil. Med. 170, 295–296.
Denes, E., Rogez, J.P., Darde, M.L., Weinbreck, P., 1999. Management ofBabesia divergens babesiosis without a complete course of quininetreatment. Eur. J. Clin. Microbiol. Infect. Dis. 18, 672–673.
de Vos, A.J., Dalgliesh, R.J., Callow, L.L., 1987. Babesia. In: Soulsby,E.J.L. (Ed.), Immune Responses in Parasitic Infections, vol. III.Protozoa. CRC Press, Boca Raton, FL, pp. 183–222.
Duh, D., Petrovec, M., Avsic-Zupanc, T., 2001. Diversity of Babesia
Infecting European sheep ticks (Ixodes ricinus). J. Clin. Microbiol. 39,3395–3397.
Duh, D., Petrovec, M., Avsic-Zupanc, T., 2005. Molecular characteriza-tion of human pathogen Babesia EU1 in Ixodes ricinus ticks fromSlovenia. J. Parasitol. 91, 463–465.
Eberhard, M.L., Walker, E.M., Steurer, F.J., 1995. Survival and infec-tivity of Babesia in blood maintained at 25 C and 2–4 C. J. Parasitol.81, 790–792.
Esernio-Jenssen, D., Scimeca, P.G., Benach, J.L., Tenenbaum, M.J., 1987.Transplacental/perinatal babesiosis. J. Pediatr. 110, 570–572.
Gelfand, J.A., 2000. Babesia. In: Mandell, G.L., Douglas, R.G., Bennett,J.E., Dolin, R. (Eds.), Mandell, Douglas, and Bennett’s Principles andPractice of Infectious Diseases, fifth ed. Churchill Livingstone, NewYork, pp. 2899–2902.
Gerber, M.A., Shapiro, E.D., Krause, P.J., Cable, R.G., Badon, S.J.,Ryan, R.W., 1994. The risk of acquiring Lyme disease or babesiosisfrom a blood transfusion. J. Infect. Dis. 170, 231–234.
Goethert, H.K., Telford 3rd, S.R., 2003a. What is Babesia microti?.Parasitology 127 301–309.
Goethert, H.K., Telford 3rd, S.R., 2003b. Enzootic transmission ofBabesia divergens among cottontail rabbits on Nantucket Island,Massachusetts. Am. J. Trop. Med. Hyg. 69, 455–460.
Gorenflot, A., Bazin, C., Ambroise-Thomas, P., 1987. Human babesiosis.Treatment of severe forms. Presse Med. 16, 1099.
Gorenflot, A., Moubri, K., Precigout, E., Carcy, B., Schetters, T.P., 1998.Human babesiosis. Ann. Trop. Med. Parasitol. 92, 489–501.
Gray, J.S., Pudney, M., 1999. Activity of atovaquone against Babesia
microti in the Mongolian gerbil, Merionesunguiculatus. J. Parasitol. 85,723–728.
Gray, J., von Stedingk, L.V., Gurtelschmid, M., Granstrom, M., 2002.Transmission studies of Babesia microti in Ixodes ricinus ticks andgerbils. J. Clin. Microbiol. 40, 1259–1263.
Gray, J.S., 2006. Identity of the causal agents of human babesiosis inEurope. Int. J. Med. Microbiol. 296 (40), 131–136.
Gray, J.S., Weiss, L.M., 2008. Babesia microti. In: Khan, N. (Ed.),Emerging Protozoan Pathogens. Taylor and Francis, Abingdon, UK,pp. 303–349.
Haselbarth, K., Tenter, A.M., Brade, V., Krieger, G., Hunfeld, K.P.,2007. First case of human babesiosis in Germany – Clinical presen-tation and molecular characterization of the pathogen. Int. J. Med.Microbiol. 297, 197–204.

K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1235
Hatcher, J.C., Greenberg, P.D., Antique, J., Jimenez-Lucho, V.E., 2001.Severe babesiosis in Long Island: review of 34 cases and theircomplications. Clin. Infect. Dis. 32, 1117–1125.
Hemmer, R.M., Ferrick, D.A., Conrad, P.A., 2000. Role of T cells andcytokines in fatal and resolving experimental babesiosis: protection inTNFRp55�/� mice infected with the human Babesia WA1 parasite. J.Parasitol. 86, 736–742.
Herwaldt, B., Persing, D.H., Precigout, E.A., Goff, W.L., Mathiesen,D.A., Taylor, P.W., Eberhard, M.L., Gorenflot, A.F., 1996. A fatalcase of babesiosis in Missouri: identification of another piroplasm thatinfects humans. Ann. Intern. Med. 124, 643–650.
Herwaldt, B.L., Kjemtrup, A.M., Conrad, P.A., Barnes, R.C., Wilson,M., McCarthy, M.G., Sayers, M.H., Eberhard, M.L., 1997. Transfu-sion-transmitted babesiosis in Washington State: first reported casecaused by a WA1-type parasite. J. Infect. Dis. 175, 1259–1262.
Herwaldt, B.L., Caccio, S., Gherlinzoni, F., Aspock, H., Slemenda, S.B.,Piccaluga, P., Martinelli, G., Edelhofer, R., Hollenstein, U., Poletti,G., Pampiglione, S., Loschenberger, K., Tura, S., Pieniazek, N.J.,2003. Molecular characterization of a non-Babesia divergens organismcausing zoonotic babesiosis in Europe. Emerg. Infect. Dis. 9, 942–948.
Herwaldt, B.L., de Bruyn, G., Pieniazek, N.J., Homer, M., Lofy, K.H.,Slemenda, S.B., Fritsche, T.R., Persing, D.H., Limaye, A.P., 2004.Babesia divergens-like infection, Washington State. Emerg. Infect. Dis.10, 622–629.
Hildebrandt, A., Hunfeld, K.P., Baier, M., Krumbholz, A., Sachse, S.,Lorenzen, T., Kiehntopf, M., Fricke, H.J., Straube, E., 2007. Firstconfirmed autochthonous case of human Babesia microti infection inEurope. Eur. J. Clin. Microbiol. Infect. Dis. 26, 595–601.
Hildebrandt, A., Tenter, A.M., Straube, E., Hunfeld, K.P., in press.Human babesiosis in Germany: just overlooked or truly new? Int. J.Med. Microbiol. doi:10.1016/j.ijmm.2007.11.001.
Holman, P.J., Spencer, A.M., Drolesky, R.E., Goethert, H.K., Telford Jr,S.R., 2005. In vitro cultivation of a zoonotic Babesia sp. isolated fromeastern cottontail rabbits (Sylvilagus floridanus) on Nantucket islandassachusetts. J. Clin. Microbiol. 43, 3995–4001.
Holman, P.J., 2006. Phylogenetic and biologic evidence that Babesia
divergens is not endemic in the United States. Ann. N.Y. Acad. Sci.1081, 518–525.
Homer, M.J., Aguilar-Delfin, I., Telford 3rd, S.R., Krause, P.J., Persing,D.H., 2000. Babesiosis. Clin. Microbiol. Rev. 13, 451–469.
Homer, M.J., Lodes, M.J., Reynolds, L.D., Zhang, Y., Douglass, J.F.,McNeill, P.D., Houghton, R.L., Persing, D.H., 2003. Identificationand characterization of putative secreted antigens from Babesia
microti. J. Clin. Microbiol. 41, 723–729.Hunfeld, K.P., Lambert, A., Kampen, H., Albert, S., Epe, C., Brade, V.,
Tenter, A.M., 2002. Seroprevalence of Babesia infections in humansexposed to ticks in midwestern Germany. J. Clin. Microbiol. 40, 2431–2436.
Hunfeld, K.P., Brade, V., 2004. Zoonotic Babesia: possibly emergingpathogens to be considered for tick-infested humans in CentralEurope. Int. J. Med. Microbiol. 293 (37), 93–103.
Kain, K.C., Jassoum, S.B., Fong, I.W., Hannach, B., 2001. Transfusion-transmitted babesiosis in Ontario: first reported case in Canada.CMAJ 164, 1721–1723.
Karp, C.L., Auwaerter, P.G., 2007. Coinfection with HIV and tropicalinfectious diseases. II. Helminthic, fungal, bacterial, and viral patho-gens. Clin. Infect. Dis. 45, 1214–1220.
Kim, J.Y., Cho, S.H., Joo, H.N., Tsuji, M., Cho, S.R., Park, I.J., Chung,G.T., Ju, J.W., Cheun, H.I., Lee, H.W., Lee, Y.H., Kim, T.S., 2007.First case of human babesiosis in Korea: detection and characteriza-tion of a novel type of Babesia sp. (KO1) similar to ovine Babesia. J.Clin. Microbiol. 45, 2084–2087.
Kivaria, F.M., Ruheta, M.R., Mkonyi, P.A., Malamsha, P.C., 2007.Epidemiological aspects and economic impact of bovine theileriosis(East Coast fever) and its control: a preliminary assessment withspecial reference to Kibaha district, Tanzania. Vet. J. 173, 384–390.
Kjemtrup, A.M., Conrad, P.A., 2000. Human babesiosis: an emergingtick-borne disease. Int. J. Parasitol. 30, 1323–1337.
Kjemtrup, A.M., Lee, B., Fritz, C.L., Evans, C., Chervenak, M., Conrad,P.A., 2002. Investigation of transfusion transmission of a WA1-typebabesial parasite to a premature infant in California. Transfusion 42,1482–1487.
Kjemtrup, A.M., Conrad, P.A., 2006. A review of the small caninepiroplasms from California: Babesia conradae in the literature. Vet.Parasitol. 138, 112–117.
Kjemtrup, A.M., Wainwright, K., Miller, M., Penzhorn, B.L., Carreno,R.A., 2006. Babesia conradae, sp. Nov., a small canine Babesia
identified in California. Vet. Parasitol. 138, 103–111.Krause, P.J., Telford 3rd, S.R., Ryan, R., Conrad, P.A., Wilson, M.,
Thomford, J.W., Spielman, A., 1994. Diagnosis of babesiosis: evalu-ation of a serologic test for the detection of Babesia microti antibody.J. Infect. Dis. 169, 923–926.
Krause, P.J., Spielman, A., Telford 3rd, S.R., Sikand, V.K., McKay, K.,Christianson, D., Pollack, R.J., Brassard, P., Magera, J., Ryan, R.,Persing, D.H., 1998. Persistent parasitemia after acute babesiosis. N.Engl. J. Med. 339, 160–165.
Krause, P.J., Telford, S.R., 1999. Babesiosis. In: Gilles, H.M. (Ed.),Protozoal diseases. Arnold, London, pp. 236–248.
Krause, P.J., Lepore, T., Sikand, V.K., Gadbaw Jr, J., Burke, G., Telford3rd, S.R., Brassard, P., Pearl, D., Azlanzadeh, J., Christianson, D.,McGrath, D., Spielman, A., 2000. Atovaquone and azithromycin forthe treatment of babesiosis. N. Engl. J. Med. 343, 1454–1458.
Krause, P.J., McKay, K., Gadbaw, J., Christianson, D., Closter, L.,Lepore, T., Telford 3rd, S.R., Sikand, V., Ryan, R., Persing, D.,Radolf, J.D., Spielman, A., 2003. Increasing health burden of humanbabesiosis in endemic sites. Am. J. Trop. Med. Hyg. 68, 431–436.
Krause, P.J., Daily, J., Sam, R., Telford 3rd, S.R., Vannier, E., Lantos, P.,Spielman, A., 2007. Shared features in the pathobiology of babesiosisand malaria. Trends Parasitol. 23, 605–610.
Krause, P.J., Gewurz, B.E., Hill, D., Marty, F.M., Vannier, E., Foppa,I.M., Furman, R.R., Neuhaus, E., Skowron, G., Gupta, S., McCalla,C., Pesanti, E.L., Young, M., Heiman, D., Hsue, G., Gelfand, J.A.,Wormser, G.P., Dickason, J., Bia, F.J., Hartman, B., Telford 3rd,S.R., Christianson, D., Dardick, K., Coleman, M., Girotto, J.E.,Spielman, A., 2008. Persistent and relapsing babesiosis in immuno-compromised patients. Clin. Infect. Dis. 46, 370–376.
Krogstad, D.J., Sutera, S.P., Boylan, C.W., Gluzman, I.Y., Qian, Z.F.,Rao, P.R., 1991. Intraerythrocytic parasites and red cell deformability:Plasmodium berghei and Babesia microti. Blood Cells 17, 209–221,discussion 222–207.
Leiby, D.A., Gill, J.E., 2004. Transfusion-transmitted tick-borne infec-tions: a cornucopia of threats. Transfus. Med. Rev. 18, 293–306.
Leiby, D.A., 2006. Babesiosis and blood transfusion: flying under theradar. Vox Sang. 90, 157–165.
Liu, A.H., Yin, H., Guan, G.Q., Schnittger, L., Liu, Z.J., Ma, M.L.,Dang, Z.S., Liu, J.L., Ren, Q.Y., Bai, Q., Ahmed, J.S., Luo, J.X.,2007. At least two genetically distinct large Babesia species infective tosheep and goats in China. Vet. Parasitol. 147, 246–251.
Mahoney, D.F., 1967. Bovine babesiosis: the passive immunization ofcalves against Babesia argentina with special reference to the role ofcomplement fixing antibodies. Exp. Parasitol. 20, 119–124.
Malandrin, L., L’Hostis, M., Chauvin, A., 2004. Isolation of Babesia
divergens from carrier cattle blood using in vitro culture. Vet. Res. 35,131–139.
Marathe, A., Tripathi, J., Handa, V., Date, V., 2005. Human babesiosis –a case report. Indian J. Med. Microbiol. 23, 267–269.
Marley, S.E., Eberhard, M.L., Steurer, F.J., Ellis, W.L., McGreevy, P.B.,Ruebush 2nd, T.K., 1997. Evaluation of selected antiprotozoal drugsin the Babesia microti-hamster model. Antimicrob. Agents Chemother.41, 91–94.
Marsaudon, E., Camenen, J., Testou, D., Bourree, P., Samson, P.,Luneau, F., 1995. Babesia canis human babesiosis causing a 40-dayanuria. Ann. Med. Interne (Paris) 146, 451–452.
Matsubara, J., Koura, M., Kamiyama, T., 1993. Infection of immuno-deficient mice with a mouse-adapted substrain of the gray strain ofBabesia microti. J. Parasitol. 79, 783–786.

1236 K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237
Mehlhorn, H., Piekarski, G., 2002. Grundriß der Parasitenkunde, 6threvised edition. Spektrum Akademischer Verlag GmbH, Heidelberg,Berlin, Germany, pp. 38–39.
Meldrum, S.C., Birkhead, G.S., White, D.J., Benach, J.L., Morse, D.L.,1992. Human babesiosis in New York State: an epidemiologicaldescription of 136 cases. Clin. Infect. Dis. 15, 1019–1023.
Mintz, E.D., Anderson, J.F., Cable, R.G., Hadler, J.L., 1991. Transfu-sion-transmitted babesiosis: a case report from a new endemic area.Transfusion 31, 365–368.
Mylonakis, E., 2001. When to suspect and how to monitor babesiosis.Am. Fam. Physician 63, 1969–1974.
New, D.L., Quinn, J.B., Qureshi, M.Z., Sigler, S.J., 1997. Verticallytransmitted babesiosis. J. Pediatr. 131, 163–164.
Olmeda, A.S., Armstrong, P.M., Rosenthal, B.M., Valladares, B., delCastillo, A., de Armas, F., Miguelez, M., Gonzalez, A., RodriguezRodriguez, J.A., Spielman, A., Telford 3rd, S.R., 1997. A subtropicalcase of human babesiosis. Acta Trop. 67, 229–234.
Pantanowitz, L., Aufranc 3rd, S., Monahan-Earley, R., Dvorak, A.,Telford 3rd, S.R., 2002. Transfusion medicine illustrated. Morphologichallmarks of Babesia. Transfusion 42, 1389.
Persing, D.H., Mathiesen, D., Marshall, W.F., Telford, S.R., Spielman,A., Thomford, J.W., Conrad, P.A., 1992. Detection of Babesia microti
by polymerase chain reaction. J. Clin. Microbiol. 30, 2097–2103.Persing, D.H., 1993. Diagnostic molecular microbiology. Current chal-
lenges and future directions. Diagn. Microbiol. Infect. Dis. 16, 159–163.
Persing, D.H., Herwaldt, B.L., Glaser, C., Lane, R.S., Thomford, J.W.,Mathiesen, D., Krause, P.J., Phillip, D.F., Conrad, P.A., 1995.Infection with a Babesia-like organism in northern California. N.Engl. J. Med. 332, 298–303.
Pieniazek, N., Sawczuk, M., Skotarczak, B., 2006. Molecular identifica-tion of Babesia parasites isolated from Ixodes ricinus ticks collected innorthwestern Poland. J. Parasitol. 92, 32–35.
Pudney, M., Gray, J.S., 1997. Therapeutic efficacy of atovaquone againstthe bovine intraerythrocytic parasite, Babesia divergens. J. Parasitol.83, 307–310.
Quick, R.E., Herwaldt, B.L., Thomford, J.W., Garnett, M.E., Eberhard,M.L., Wilson, M., Spach, D.H., Dickerson, J.W., Telford 3rd, S.R.,Steingart, K.R., Pollock, R., Persing, D.H., Kobayashi, J.M., Juranek,D.D., Conrad, P.A., 1993. Babesiosis in Washington State: a newspecies of Babesia? Ann. Intern. Med. 119, 284–290.
Raucher, H.S., Jaffin, H., Glass, J.L., 1984. Babesiosis in pregnancy.Obstet. Gynecol. 63, 7S–9S.
Rosner, F., Zarrabi, M.H., Benach, J.L., Habicht, G.S., 1984. Babesiosisin splenectomized adults. Review of 22 reported cases. Am. J. Med. 76,696–701.
Rothman, K.L., Greenland, S. (Eds.), 1998. Modern Epidemiology,second ed. Lippincott-Raven, Philadelphia, PA.
Rowin, K.S., Tanowitz, H.B., Wittner, M., 1982. Therapy of experimentalbabesiosis. Ann. Intern. Med. 97, 556–558.
Ryan, R., Krause, P.J., Radolf, J., Freeman, K., Spielman, A., Lenz, R.,Levin, A., 2001. Diagnosis of babesiosis using an immunoblotserologic test. Clin. Diagn. Lab. Immunol. 8, 1177–1180.
Saito-Ito, A., Tsuji, M., Wei, Q., He, S., Matsui, T., Kohsaki, M., Arai, S.,Kamiyama, T., Hioki, K., Ishihara, C., 2000. Transfusion-acquired,autochthonous human babesiosis in Japan: isolation of Babesia
microti-like parasites with hu-RBC-SCID mice. J. Clin. Microbiol.38, 4511–4516.
Saito-Ito, A., Yano, Y., Dantrakool, A., Hashimoto, T., Takada, N.,2004. Survey of rodents and ticks in human babesiosis emergence areain Japan: first detection of Babesia microti-like parasites in Ixodes
ovatus. J. Clin. Microbiol. 42, 2268–2270.Sharan, K.P., Krause, P.J., 2000. Babesiosis. In: Cunha, B.A. (Ed.), Tick-
Borne Infectious Diseases: Diagnosis and Management. MarcelDecker Inc., New York, pp. 111–120.
Shih, C.M., Liu, L.P., Chung, W.C., Ong, S.J., Wang, C.C., 1997. Humanbabesiosis in Taiwan: asymptomatic infection with a Babesia microti-like organism in a Taiwanese woman. J. Clin. Microbiol. 35, 450–454.
Sinski, E., Bajer, A., Welc, R., Pawelczyk, A., Ogrzewalska, M., Behnke,J.M., 2006. Babesia microti: prevalence in wild rodents and Ixodes
ricinus ticks from the Mazury Lakes District of North-Eastern Poland.Int. J. Med. Microbiol. 296 (40), 137–143.
Skrabalo, Z., Deanovic, Z., 1957. Piroplasmosis in man; report of a case.Doc. Med. Geogr. Trop. 9, 11–16.
Smith, T., Kilbourne, E.L., 1893. Investigation into the nature, causation,and prevention of southern cattle fever. US Dept. Agr. Bur. Anim.Indust. Bull. 1.
Spencer, A.M., Goethert, H.K., Telford 3rd, S.R., Holman, P.J., 2006. Invitro host erythrocyte specificity and differential morphology ofBabesia divergens and a zoonotic Babesia sp. from eastern cottontailrabbits (Sylvilagus floridanus). J. Parasitol. 92, 333–340.
Spielman, A., 1976. Human babesiosis on Nantucket Island: transmissionby nymphal Ixodes ticks. Am. J. Trop. Med. Hyg. 25, 784–787.
Sun, T., Tenenbaum, M.J., Greenspan, J., Teichberg, S., Wang, R.T.,Degnan, T., Kaplan, M.H., 1983. Morphologic and clinical observa-tions in human infection with Babesia microti. J. Infect. Dis. 148, 239–248.
Swanson, S.J., Neitzel, D., Reed, K.D., Belongia, E.A., 2006. Coinfectionsacquired from Ixodes ticks. Clin. Microbiol. Rev. 19, 708–727.
Telford 3rd, S.R., Gorenflot, A., Brasseur, P., Spielman, A., 1993.Babesial infection in human and wildlife. In: Kreier, J.P. (Ed.),Parasitic Protozoa, vol. 5. Academic Press, San Diego, CA, pp. 1–47.
Telford 3rd, S.R., Goethert, H.K., 2004. Emerging tick-borne infections:rediscovered and better characterized, or truly ‘new’? Parasitology 129,301–327, Suppl.
Telford 3rd, S.R., Maguire, J.H., 2006. Babesiosis. In: Guerrant, R.L.,Walker, D.H., Weller, P.F. (Eds.), Tropical Infectious Diseases:Principles, Pathogens, and Practice. Churchill Livingstone, pp. 1063–1071.
Tsuji, M., Wei, Q., Zamoto, A., Morita, C., Arai, S., Shiota, T.,Fujimagari, M., Itagaki, A., Fujita, H., Ishihara, C., 2001. Humanbabesiosis in Japan: epizootiologic survey of rodent reservoir andisolation of new type of Babesia microti-like parasite. J. Clin.Microbiol. 39, 4316–4322.
Uilenberg, G., 2006. Babesia – A historical overview. Vet. Parasitol. 138,3–10.
Vial, H.J., Gorenflot, A., 2006. Chemotherapy against babesiosis. Vet.Parasitol. 138, 147–160.
Walter, G., 1984. [Transmission and course of parasitemia of Babesia
microti (Hannover I strain) in the bank vole (Clethrionomys glareolus)and field vole (Microtus agrestis)]. Acta Trop. 41, 259–264.
Weiss, L.M., Wittner, M., Wasserman, S., Oz, H.S., Retsema, J.,Tanowitz, H.B., 1993. Efficacy of azithromycin for treating Babesia
microti infection in the hamster model. J. Infect. Dis. 168, 1289–1292.White, D.J., Talarico, J., Chang, H.G., Birkhead, G.S., Heimberger, T.,
Morse, D.L., 1998. Human babesiosis in New York State: review of139 hospitalized cases and analysis of prognostic factors. Arch. Intern.Med. 158, 2149–2154.
WHO, 1995. World Health Organisation Veterinary Public Health Unit:WHO Workshop on Lyme Borreliosis Diagnosis and Surveillance.Document No. WHO/CDS/VPH/95.141. World Health Organization,Geneva, Switzerland.
Wittner, M., Rowin, K.S., Tanowitz, H.B., Hobbs, J.F., Saltzman, S.,Wenz, B., Hirsch, R., Chisholm, E., Healy, G.R., 1982. Successfulchemotherapy of transfusion babesiosis. Ann. Intern. Med. 96, 601–604.
Wittner, M., Lederman, J., Tanowitz, H.B., Rosenbaum, G.S., Weiss,L.M., 1996. Atovaquone in the treatment of Babesia microti infectionsin hamsters. Am. J. Trop. Med. Hyg. 55, 219–222.
Wormser, G.P., Dattwyler, R.J., Shapiro, E.D., Halperin, J.J., Steere,A.C., Klempner, M.S., Krause, P.J., Bakken, J.S., Strle, F., Stanek,G., Bockenstedt, L., Fish, D., Dumler, J.S., Nadelman, R.B., 2006.The clinical assessment, treatment, and prevention of Lyme disease,human granulocytic anaplasmosis, and babesiosis: clinical practiceguidelines by the Infectious Diseases Society of America. Clin. Infect.Dis. 43, 1089–1134.

K.-P. Hunfeld et al. / International Journal for Parasitology 38 (2008) 1219–1237 1237
Yabsley, M.J., Romines, J., Nettles, V.F., 2006. Detection of Babesia andAnaplasma species in rabbits from Texas and Georgia, USA. VectorBorne Zoonotic Dis. 6, 7–13.
Yano, Y., Saito-Ito, A., Anchalee, D., Takada, N., 2005. Japanese Babesia
microti cytologically detected in salivary glands of naturally infectedtick Ixodes ovatus. Microbiol. Immunol. 49, 891–897.
Zahler, M., Rinder, H., Schein, E., Gothe, R., 2000. Detection of a newpathogenic Babesia microti-like species in dogs. Vet. Parasitol. 89,241–248.
Zamoto, A., Tsuji, M., Kawabuchi, T., Wei, Q., Asakawa, M., Ishihara,C., 2004a. U.S.-type Babesia microti isolated from small wild mammalsin Eastern Hokkaido. Jpn. J. Vet. Med. Sci. 66, 919–926.
Zamoto, A., Tsuji, M., Wei, Q., Cho, S.H., Shin, E.H., Kim, T.S.,Leonova, G.N., Hagiwara, K., Asakawa, M., Kariwa, H., Takashima,I., Ishihara, C., 2004b. Epizootiologic survey for Babesia microti amongsmall wild mammals in northeastern Eurasia and a geographic diversityin the beta-tubulin gene sequences. J. Vet. Med. Sci. 66, 785–792.
Zavizion, B., Pereira, M., de Melo Jorge, M., Serebryanik, D., Mather,T.N., Chapman, J., Miller, N.J., Alford, B., Bzik, D.J., Purmal, A.,2004. Inactivation of protozoan parasites in red blood cells usingINACTINE PEN110 chemistry. Transfusion 44, 731–738.
Zintl, A., Mulcahy, G., Skerrett, H.E., Taylor, S.M., Gray, J.S., 2003.Babesia divergens, a bovine blood parasite of veterinary and zoonoticimportance. Clin. Microbiol. Rev. 16, 622–636.





























