Embed Size (px)
Citation preview

B. Control of Cell Number
1. Review of the mitotic cell cycle and cell death2. Regulation of cell number and quality during
mitosis 3. Regulation of cell type during mitosis

Numbers in a Cell Population
• Cell number is a combination of....
• Cell divisions – Cell deaths (necrotic + programmed)
• Necrosis is premature cell death– disease, injury, starvation, toxicity, excitotoxicity
• Programmed cell death is death by design– apoptosis, anoikis, cornification, autophagy
• Same for an organism, system, organ or tissue, and for single cell populations in an ecosystem

Figure 18-13 Molecular Biology of the Cell (© Garland Science 2008)
Programmed cell death is an essential component
It guarantees appropriate ennervation patterns

Figure Q18-1 Molecular Biology of the Cell (© Garland Science 2008)
Even gives us our fingers and toes!

Figure 17-51a Molecular Biology of the Cell (© Garland Science 2008)
Overview of the Mitotic Cell Cycle and Cell Division. Diploid cells duplicate the contents of their cytosol and nucleus prior to splitting to form two genetically exact daughter cells.

Figure 17-69 Molecular Biology of the Cell (© Garland Science 2008)
We’ve learned to both control it.........
A mutation in a signal molecule that limits muscle cell division has been bred in.

Fig. 11-19
2 µm
And fear it.........
A normal cell next to a tumor derived from uncontrolled cell divisions

Like everything else, the process of cell division has evolved over time

Fig. 12-11-1
Origin ofreplication
Two copiesof origin
E. coli cell Bacterialchromosome
Plasmamembrane
Cell wall
Prokaryotic Divisionis called binary fission.
Two copies of circular DNA, beginning at the origin of replication, actively move apart from each other.

Fig. 12-11-2
Origin ofreplication
Two copiesof origin
E. coli cell Bacterialchromosome
Plasmamembrane
Cell wall
Origin Origin

Fig. 12-11-4
Origin ofreplication
Two copiesof origin
E. coli cell Bacterialchromosome
Plasmamembrane
Cell wall
Origin Origin
When the daughter chromosomes reach the opposite poles, the cell separates into two daughter cells

Fig. 12-12
(a) Bacteria
Bacterialchromosome
Chromosomes
Microtubules
Intact nuclearenvelope
(b) Dinoflagellates
Kinetochoremicrotubule
Intact nuclearenvelope
(c) Diatoms and yeasts
Kinetochoremicrotubule
Fragments ofnuclear envelope
(d) Most eukaryotes
Since prokaryotes evolved before eukaryotes, mitosis probably evolved from binary fission.
Certain protists exhibit types of cell division that seem intermediate between binary fission and mitosis.

The complex mitotic cell cycle is used by the organism to control:
cell number
cell quality
cell type

Figure 15-8 Molecular Biology of the Cell (© Garland Science 2008)
TargetMechanisms
CellNumber
CellType
CellQuality

Figure 17-62 (part 1 of 3) Molecular Biology of the Cell (© Garland Science 2008)
We’ve even looked at one pathway already

Fig. 12-5
S(DNA synthesis)
MITOTIC(M) PHASE
Mito
sis
Cytokinesis
G1
G2

Interphase does the real work of mitosis
• Gap 1 (G1) is when the cell gets ready for DNA synthesis– Need DNA synthase complexes, repair enzymes, histones, etc..
• S-phase is when the cell synthesizes and edits chromosomes
– Synthesis and error editing are the primary objectives.
• Gap 2 (G2) is when the cell gets ready for cell division
– Need organelles, cytoskeletal proteins, molecular motors, metabolic enzymes, etc.
– Kinetochores are refined and finalized.

Fig. 12-UN1
Telophase andCytokinesis
Anaphase
Metaphase
Prometaphase
Prophase
MITOTIC (M) PHASE
Cytokinesis
Mitosis
SG1
G2

M-Phase
• Prophase: chromosomes condense with kinetichores, centrisomes move to poles, nuclear membrane disintegrates
• Metaphase: spindle fibers attach and push to midline
• Anaphase: kinetichores pull sister chromatids to poles
• Telophase: the reversal of prophase activities
• Cytokinesis: actin-based separation of cytosol into daughters

Figure 17-43a Molecular Biology of the Cell (© Garland Science 2008)
Sister chromatids are attached to each other at centromeres andeach is individually attached to a kinetochore from the opposite pole
metaphase

Figure 17-43b Molecular Biology of the Cell (© Garland Science 2008)
anaphase
Sister chromatids separate from each other, producing 92 individual chromosomes in humans, and are pulled by kinetichores to the poles.

Figure 17-37 Molecular Biology of the Cell (© Garland Science 2008)
dynein motors

Figure 17-40 Molecular Biology of the Cell (© Garland Science 2008)

Figure 17-49a Molecular Biology of the Cell (© Garland Science 2008)
Cytokinesis results from the assembly of an actin-myosin ring that gets smaller and smaller as mysoin pulls actin along actin. Ring assembly is microtubule-dependent and myosin light chain kinase must be phosphorylated to begin.

The cleavage furrow

Cell Division in Plants
In some plants growth continues over the life of the organism

Regulation of the Cell Cycle
• There are four major ‘checkpoints’ that monitor sensor systems and trigger molecular switches to get to next stage– The G1/S checkpoint – to enter the cycle or not– The S-checkpoint – to synthesize DNA or not– The G2/M checkpoint – to divide the cell or not– The M-checkpoint – to shift from metaphase to anaphase
• The sensor systems are focused outside the cell as well as inside the cell to make sure conditions are appropriate
• When ‘off’, the molecular switches halt progression, when ‘on’ they biochemically start the next stage

Fig. 12-14
SG1
M checkpoint
G2M
Controlsystem
G1 checkpoint
G2 checkpoint
S checkpoint
The book claims 3 and describes 4, so we’ll go with 4

• For many cells, the G1 checkpoint seems to be the most important one
• If a cell receives a go-ahead signal at the G1 checkpoint, it will usually complete the S, G2, and M phases and divide
• If the cell does not receive the go-ahead signal, it will exit the cycle, switching into a nondividing state called the G0 phase
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Fig. 12-15
G1
G0
G1 checkpoint
(a) Cell receives a go-ahead signal
G1
(b) Cell does not receive a go-ahead signal

Stop and Go Signals at the G1 Checkpoint
• In multicellular organisms, the classic required external signals are growth factors, cytokines and hormones
• Most cells also exhibit anchorage dependence, in which they must be attached in order to divide
• Most cells also exhibit density-dependent inhibition, in which crowded cells stop dividing
• Many single cells also monitor external nutrient availability and will not divide if it is inadequate
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Figure 17-15 Molecular Biology of the Cell (© Garland Science 2008)
The Cell Cycle Switches at the First Three Checkpoints are Molecules Called Cyclins and Cyclin-Dependent Kinases
Cyclins are expressed and degraded, when present the cell moves forward
1. G1 and G1/S cyclins regulate the G1 checkpoint
2. S cyclins regulate S checkpoint
3. M cyclins regulate G2/M checkpoint
Cyclin-dependent kinases are always present and are activated by the binding of their appropriate cyclin

Cyclin isdegraded
Cdk
MPF
M
SG2
checkpoint
Cyclin
Cyclin accumulation
1. Cdk is always there, 2. if conditions are OK cyclin is expressed,3. the combination is an active kinase, and 4. activity is lost with cyclin

What does cyclin-cdk phosphorylate?
• G1-cdk– Rb protein: loses inhibition of E2F– E2F then activates transcription of S-cyclin
• M-cdk– histones: chromosome condensation– lamins: nuclear membrane fragmentation– myosin: purse-string cytokinesis

Figure 17-62 (part 1 of 3) Molecular Biology of the Cell (© Garland Science 2008)
So, back to our growth factor example.....
What does immediate early gene expression cause?

Figure 17-62 (part 2 of 3) Molecular Biology of the Cell (© Garland Science 2008)
Immediate early gene, Myc, expression activates the expression of G1 cyclin

M Checkpoint Regulation
• If any kinetochores are not attached to spindle microtubules during metaphase they send a molecular signal that delays cycle progression
• The signal activates the anaphase-promoting complex, or cyclosome, known as APC/C
• APC/C is a protease that destroys:– cyclins: stopping cell cycle– securin: which drives sister chromatid separation

Apoptosis During Mitosis Controls Cell Quality
• Every cell cycle in multicellular organisms are molecularly hardwired with apoptosis as a potential outcome if things aren’t just right
• The primary cause is imperfect DNA copying
• This is one of our primary defenses against erroneous or uncontrolled proliferation

Characteristics: 1. Cessation of DNA repair mechanisms2. Cell shrinkage3. Nuclear membrane blebbing 4. DNA fragmentation5. Death

Figure 17-63 (part 1 of 2) Molecular Biology of the Cell (© Garland Science 2008)
Can result from bad copies during DNA replication
p%#Mdm2 will cause p53 degradation

Figure 17-63 (part 2 of 2) Molecular Biology of the Cell (© Garland Science 2008)
p21 has been shown to block cyclin/cdk activity at G1-S, S and G2-M

The Roles of p21 and PCNA
• The apoptotic clock:
– PCNA = the proliferating cell nuclear antigen
– If [p21] rises to 5X [PCNA] apoptosis will happen
– p21 will reach binding affinity for PCNA
– The dimer will shut off DNA repair mechanisms

Figure 18-11a Molecular Biology of the Cell (© Garland Science 2008)
When apoptosis is inactive, Bcl2 is bound to APAF-1 and the dimer acts to block assemblage of mitochondrial membrane
channel proteins, BH123 (Bax and Bac) ,

The loss of DNA repair mechanisms:
1. Activates BH3-only proteins
2. Signal Bax translocation to the mitochondria

Figure 18-11b Molecular Biology of the Cell (© Garland Science 2008)
apoptosisinducing factor
Bax/Bac dimers
BH3-only inactivates Bcl2/APAF-1 inhibition and along with Bax dimerization with Bac forms active channels

Figure 18-8 Molecular Biology of the Cell (© Garland Science 2008)
Release of cytochrome C from the mitochondria outcompetes Bcl2 for APAf-1 and activates assembly of the apoptosome
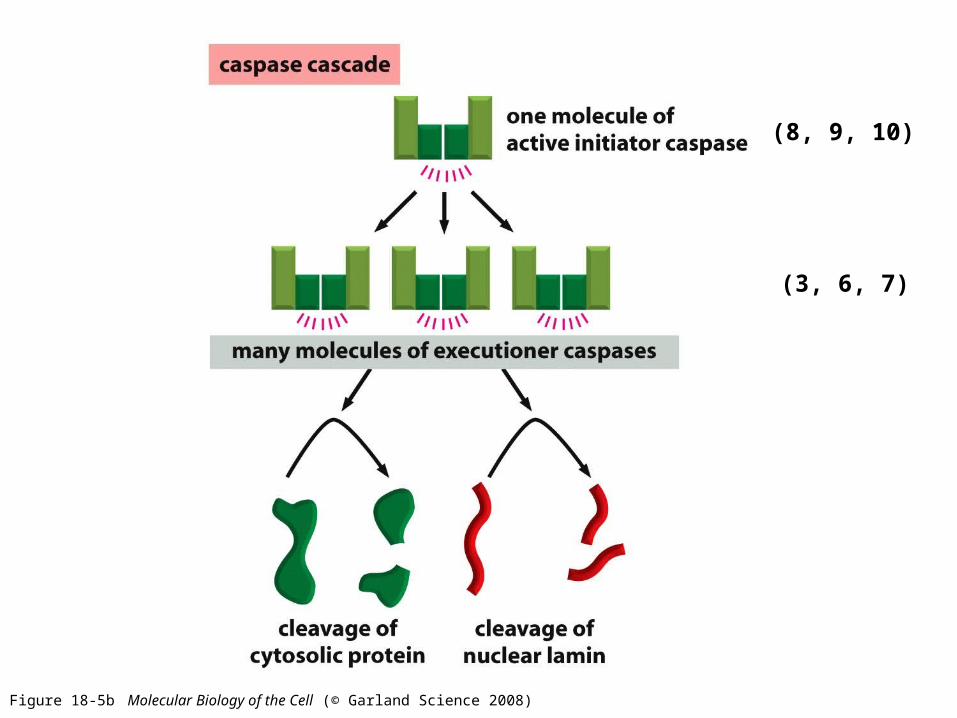
Figure 18-5b Molecular Biology of the Cell (© Garland Science 2008)
(8, 9, 10)
(3, 6, 7)

Characteristics: 1. Cessation of DNA repair mechanisms2. Cell shrinkage3. Nuclear membrane blebbing 4. DNA fragmentation5. Death

Figure 18-6 Molecular Biology of the Cell (© Garland Science 2008)
Extrinsic apoptotic cascade also uses the common caspase cascade

3. Regulation of cell type during mitosis
• Review – Symmetric vs. Asymmetric Cell Division– Stem Cell and Embryonic Cell Divisions
• Regulation– Control in the Cytosol– Control in the Nucleus


Common Cell Divisions Produce Daughters Like the Parent to Replenish the Population
Common during growth and the repair of damaged tissues

The Stem Cell Concept
• Asymmetric division of stem cells produces one new stem cell and one differentiated daughter– Many stem cells in the embryo and adult
• In some organs: frequent replenishing divisions– gut, epidermis, bone marrow– example: billions of blood cells are destroyed by the spleen
every hour
• In others, they only divide in response to stress or the need to repair the organ– heart, prostate

Stem Cell Mitosis
Each division produces daughter cell(s) unlike the parent cell:
- The first two are asymmetric and produce one stem cell
- The last one produces two like daughters unlike parent
HSC

Stem Cell Types: Embryonic Stem Cells
The Inner Cell Mass
Produces all cells of the embryo

Stem Cell Types: Adult Stem Cells
• Committed stem cells with limited potential
– hematopietic stem cells - hair stem cells
– mesenchymal stem cells - melanocyte stem cells
– epidermal stem cells - muscle stem cells
– neural stem cells - tooth stem cells
– gut stem cells - germline stem cells
– mammary stem cells

The hair shaft is composed of keratinocytes, lubricated by sebaceous secretions, with melanin for color. - “Bulge” is the stem cell niche for hair basal cells, sebocytes and melanocytes.
- The first two arise from a common stem cell but melanocytes arise from a committed stem cell.

Hematopoietic stem cells in the bone marrow are the kings of differentiation choices
Progenitor cells will produce two daughters like each other but not like the parent, with the choice being determined by the current, local signaling combination

Control of Assymetrical Division in the Cytosol
• G2 is the key to differentiation events
• The daughter cells can be built differently– DNA is the same but....– RNA’s can be different– Proteins can be different– Cytoskeleton and organelles can even be different
• The parent cell must focus its placement of these components on either side of the furrow

Isolation of transcription factors across midline

Fig. 18-15a
Two differentcytoplasmicdeterminants
Unfertilized egg cell
Sperm
Fertilization
Zygote
Mitoticcell division
Two-celledembryoshave twodifferentcell types
NucleusThe unfertilized egg is the queen of cytoplasmic isolation of cell fate determinants
All stem cells do it effectively

We use the embryonic process of cleavage to disperse the careful distribution of cytoplasmic determinants laid down in the egg.

The G-phases of somatic mitosis allow for cytoplasmic growth so that the daughter cells are equal in size to the parent cell.
In cleavage we want to use the egg cytoplasmic material so we just skip the G-phases all together.
It makes the cell cycle go very fast! - Frogs can make 37,000 cells in 43 hours. - Fruit flies can make 50,000 in 12 hours (10 min!)

They don’t even bother to make plasmamembranes until later!

Control of asymmetric division in the nucleus

Two DNA methyltransferases are important in modifying DNA

Development of pluripotency in the inner cell mass depends on methylation pattern
Both parent gametes have methylation patterns that must be removed for all genes to be available to the developing organism.

One of the big hurdles to somatic cell nuclear transfer cloning was overcoming the adult methylation pattern
It took hundreds of failed attempts before the successful cloning of Dolly from adult mammary epithelium.

Each step of differentiation of a given cell type depends on changing methylation patterns
These genes maximize effectiveness when coding for transcription factors, SNuRPs, signaling receptors, etc.





























