Embed Size (px)
Citation preview

AZ MDMA HATÁSAI A VIGILANCIÁRA ÉS
NEURONKÁROSÍTÓ HATÁSÁNAK FARMAKOLÓGIAI
KÖVETKEZMÉNYEI
-Balogh Brigitta-
Témavezető: Dr. Bagdy György Programvezető: Dr. Nagy Zoltán
Semmelweis Egyetem, Általános Orvostudományi Kar
Farmakológiai és Farmakoterápiás Intézet
korábban:
Országos Pszichiátriai és Neurológiai Intézet Neuropszichofarmakológiai és Neurokémiai Laboratórium
SEMMELWEIS EGYETEM
SZENTÁGOTHAI JÁNOS IDEGTUDOMÁNYI DOKTORI ISKOLA
2008
A szigorlati bizottság tagjai: Dr. Török Tamás (elnök) Dr. Tímár Júlia Dr. Lévay György
Hivatalos bírálók: Dr. Hársing László Gábor Dr. Riba Pál

TARTALOMJEGYZÉK
A DOLGOZATBAN HASZNÁLT RÖVIDÍTÉSEK JEGYZÉKE 4
1 BEVEZETÉS 6 1.1 Történelmi áttekintés 6
1.2 A szerotonerg rendszer működésének áttekintése 8
1.2.1 A szerotonin (5-HT) szintézise és felszabadulása az idegsejtekből 8
1.2.2 Az 5-HT visszavétele és lebontása 9
1.2.3 Az 5-HT receptorai 10
1.2.4 A szerotonerg rendszer anatómiája 12
1.2.5 Az 5-HT szerepe az alvás szabályozásában 13
1.2.6 Az 5-HT szerepe a motoros aktivitás szabályozásában 14
1.3 A dopaminerg, a noradrenerg és a kolinerg rendszer szerepe az alvás
és a motoros aktivitás szabályozásában 16
1.4 Az MDMA akut hatásai állatokban 17
1.4.1 A monoaminok felszabadítása az agyban 18
1.4.2 Az acetilkolin felszabadítása az agyban 19
1.4.3 Az MDMA akut élettani és viselkedési hatásai állatokban 20
1.4.4 Aggregációs toxicitás 23
1.5 Az MDMA hosszútávú hatásai állatokban 23
1.5.1 A neurotoxikus hatás feltételezett mechanizmusai 23
1.5.2 Bizonyítékok a szerotonerg idegsejtek működésének tartós
csökkenésére patkányokban 25
1.5.3 A szerotonin-rendszer csökkent működésének következményei
patkányokban 27
1.6 Az állatkísérletekben használt és az ecstasyszedők által alkalmazott
dózisok összehasonlítása 30
1.7 A Dark Agouti patkánytörzs, mint az MDMA-t örökletesen lassan
metabolizáló emberek állatmodellje 30
1.8 Az MDMA (ecstasy) hatásai emberekben 31
1

1.8.1 Akut és szubakut hatások 31
1.8.2 Az ecstasy-szedés hosszú távú következményei 33
2 CÉLKITŰZÉSEK 37
3 MÓDSZEREK 38 3.1 Az állatok és tartásuk 38
3.2 Motoros aktivitás és alvás vizsgálatok 38
3.2.1 A kezelések 38
3.2.1.1 Kezelések az MDMA akut, szubakut és hosszútávú hatásainak
vizsgálatához 39
3.2.1.2 Kezelések a citalopram akut hatásainak vizsgálatához 40
3.2.2 A krónikus EEG és EMG elektródák műtéti beültetése 41
3.2.3 Az EEG, EMG és a motoros aktivitás mérése 42
3.2.4 Az alvásstádiumok meghatározása 43
3.2.5 A cosinor analízis módszer 44
3.2.6 Az adatok statisztikai elemzése 45
3.3 A [3H]-paroxetin kötés mérése autoradiográfiával 46
4 EREDMÉNYEK 48 4.1 Az MDMA akut hatásai: első 24 óra 48
4.2 Az MDMA hatásai a kezelést követő napokban 52
4.3 Az MDMA krónikus hatásai: 14. és 28. nap 58
4.4 Az MDMA hatásai a [3H]-paroxetin kötésre 65
4.5 Az MDMA akut hatásai az MDMA-val előkezelt állatokban 66
4.6 A citalopram akut hatásai az alvásra 72
4.7 A citalopram akut hatásai az alvásra MDMA-val előkezelt
állatokban 73
5 MEGBESZÉLÉS 81 5.1 Az MDMA akut hatásai: első 24 óra 81
5.2 Az MDMA hatásai a kezelést követő napokban 83
2

5.3 Az MDMA krónikus hatásai 84
5.4 Az MDMA akut hatásai az MDMA-val előkezelt állatokban 87
5.5 A citalopram akut hatásai az alvásra 88
5.6 A citalopram akut hatásai az alvásra MDMA-val előkezelt
állatokban 89
6 KÖVETKEZTETÉSEK 90
7 ÖSSZEFOGLALÁS 91
8 SUMMARY 93
9 IRODALOMJEGYZÉK 95
10 AZ ÉRTEKEZÉS TÉMÁJÁHOZ KÖZVETLENÜL
KAPCSOLÓDÓ KÖZLEMÉNYEK, KIVONATOK,
ELŐADÁSOK ÉS POSZTEREK 111
11 AZ ÉRTEKEZÉS TÉMÁJÁHOZ KÖZVETLENÜL NEM
KAPCSOLÓDÓ KÖZLEMÉNYEK, KIVONATOK,
ELŐADÁSOK ÉS POSZTEREK 115
12 KÖSZÖNETNYILVÁNÍTÁS 118
3

A DOLGOZATBAN HASZNÁLT RÖVIDÍTÉSEK
JEGYZÉKE
5-HIAA 5-hidroxi-indolecetsav
5-HT 5-hidroxi-triptamin, szerotonin
5-HTP 5-hidroxi-triptofán
AW active wakefulness, aktív ébrenlét
Ca2+ kalcium
CI konfidencia intervallum
CSF cerebrospinális folyadék
CYP2D6 debrisoquin-4-hidroxiláz
CYP450 citokróm-P-450
DAG diacil-glicerol
DAT dopamin transzporter
DHA dihidroxi-amfetamin
DHMA dihidroxi-metamfetamin
GABA γ-amino-vajsav
h óra
i.p. intraperitoneális
IP3 inozitol-trifoszfát
LSD lizergsav-dietilamid
MAO monoamin-oxidáz
MAO-A monoamin-oxidáz-A
MAO-B monoamin-oxidáz-B
m-CPP meta-klorofenilpiperazin
MDA 3,4-metiléndioxi-amfetamin
MDMA 3,4-metiléndioxi-metamfetamin, ecstasy
min perc
NAT noradrenalin transzporter
NMDA N-metil-D-aszpartát
NREM-alvás non-rapid eye movement, lassú hullámú alvás
4

NREM-1 felületes alvás, szendergés
NREM-2 mélyalvás
PET positron emission tomography, pozitron emissziós tomográfia
PKC protein-kináz C
PLC foszfolipáz C
PW passive wakefulness, passzív ébrenlét
REM-alvás rapid eye movement, paradox alvás
s másodperc
SERT szerotonin transzporter
SH-csoport szulfhidril-csoport
SPECT single photon emission computed tomography, egyfoton-emissziós
komputer tomográfia
SSRI selective serotonin reuptake inhibitor, szelektíven szerotonin-visszavételt
gátló
TPH triptofán-hidroxiláz
5

1 BEVEZETÉS
1.1 Történelmi áttekintés
Napjainkra az egyre szélesebb körben elterjedt, egyre fiatalabb korosztályt érintő
kábítószer-élvezet világméretű problémává vált. A klasszikus kábítószerek mellett
megjelentek a szintetikus drogok és ma az ecstasy a világon a leggyakrabban előforduló
szintetikus kábítószerek egyike (EU 2006).
Az ecstasytabletták hatóanyaga túlnyomó többségében a hallucinogén és
szimpatomimetikus hatásokkal egyaránt rendelkező amfetaminszármazék, az MDMA
(3,4-metilén-dioxi-metamfetamin, 1. ábra) (Parrott 2004; EU 2006). Az MDMA-t az
entaktogének csoportjába sorolják utalva ezzel sajátos hangulatmódosító hatásaira
(entaktogén = belsőleg megérintő hatás) (Green és mtsai 1995; Doyon 2001). Első
szintézisének időpontja és célja körül ma is viták vannak. A legvalószínűbb, hogy
Anton Kollisch, a német Merck Gyógyszergyár vegyésze szintetizálta először 1912-ben,
a vérzéscsillapító hidrasztinin előállítása közben, mint mellékterméket. Ezt követően
1914-ben kapta meg a szabadalmat a gyár az MDMA-nak (ekkor metil-zafrilaminnak
hívták), mint a hidrasztinin prekurzorának gyártására (Benzenhofer és Passie 2006;
Freudenmann és mtsai 2006). A következő évtizedekben az amerikai hadsereg néhány
kísérletétől eltekintve nem használták egészen addig, amíg az 1970-es években
Alexander Shulgin pszichofarmakológus Kaliforniában ismét előállította és humán
vizsgálatokat végzett vele (Shulgin 1986). Shulgin munkacsoportja elsőként írta le az
MDMA pszichés hatásait 1978-ban (Anderson és mtsai 1978), ezt követően a
pszichoterápiában járulékos szerként alkalmazták. Viszonylag enyhe és rövid hatású
szernek tartották, ami javítja az önelemző képességet, közvetlenebbé tesz észrevehető
változások, érzelmi kiszámíthatatlanság és ellenséges reakciók nélkül (amelyek pl. az
LSD-hez társulnak). Önkénteseken vizsgálva még 1986-ban is biztonságosnak tartották,
úgy vélték, használata nem fog visszaéléshez vezetni (Grinspoon és Bakalar 1986). Az
állatkísérletek azonban hosszútávú neurotoxikus hatásaira derítettek fényt (Schmidt és
mtsai 1986; Stone és mtsai 1986), ezért 1986 novemberében az FDA (Food and Drug
Administration, Amerikai Élelmiszer- és Gyógyszerhatóság) az illegális, gyógyászati
felhasználással nem rendelkező kábítószerek közé sorolta (Freudenmann és mtsai 2006).
6

A betiltás körül hatalmas viták voltak, számtalan újságcikk jelent meg róla, ami
sokakban felkeltette az érdeklődést az immár illegális kábítószer iránt (Parrott 2001).
Stimuláns és pszichoaktív hatásai, valamint könnyű alkalmazhatósága (per os jól
felszívódik) és egyszerű előállíthatósága is hozzájárultak ahhoz, hogy széles körben
elterjedt rekreációs droggá váljon (Doyon 2001; Parrott 2001). Ma már az ecstasy az
amfetaminal és a metamfetaminnal együtt – rögtön a kannabisz után - a világon a
második leggyakrabban használt kábítószerek csoportját alkotja. Fogyasztói manapság
főként fiatalok, az Európai Unió Kábítószerügyi Központjának 2006-os felmérése
szerint használatának életprevalenciája az Európai Unióban az összes felnőttet (15-64
évesek) figyelembe véve 2,6%, míg a fiatal felnőttek (15-34 évesek) körében ennek a
duplája, átlagosan 5,2% (EU 2006).
Az ecstasy népszerűségével párhuzamosan emelkedett a használatából eredő
halálesetek száma, sok esetben már néhány tabletta elfogyasztása végzetes kimenetelű
volt (Henry és mtsai 1992; Green és mtsai 1995). Úgy tűnik, a súlyos esetek növekvő
száma kapcsolatban áll megváltozott fogyasztásával is: az 1980-as évek közepéig főként
egyedül vagy kis társaságban használták, többnyire havonta egyszer és mindössze egy
tablettát fogyasztottak. Később divattá vált, hogy egyre gyakrabban használták és egyre
több tablettát vettek be alkalmanként (Parrott 2001). Ráadásul ma főként éjszakai
szórakozóhelyeken - diszkókban, rave partikon, klubokban – használják, ahol toxikus
hatását még tovább fokozzák a dehidratálódást elősegítő körülmények (tömeg, meleg,
intenzív mozgás) (Schifano 2000; Parrott 2001). A környezeti faktorok mellett a
tragikus kimenetelű esetek számát növeli az is, hogy az MDMA metabolizálásában fő
szerepet játszó CYP2D6 enzim polimorfizmusának köszönhetően fogyasztói különböző
mértékben érzékenyek toxikus hatásaira (Tucker és mtsai 1994). Az sem
elhanyagolandó, hogy bár az ecstasyként árusított tabletták többsége pszichoaktív
anyagként általában kizárólag MDMA-t vagy más, hasonló hatású
amfetaminszármazékot (metilén-dioxi-etamfetamint vagy metilén-dioxi-amfetamint)
tartalmaz (EU 2006), előfordulnak köztes termékek, más kábítószerek és pszichoaktív
anyagok is a tablettákban pl. LSD, amfetamin, metamfetamin, koffein, szelegilin,
efedrin, ketamin (Parrott 2004). További veszélyt jelent, hogy a tabletták MDMA-
tartalma nagyon változó és nem utolsósorban az ecstasyval együtt elfogyasztott alkohol
7

és más kábítószerek (pl. opiátok) is növelhetik toxicitását (Green és mtsai 1995; Parrott
2001).
MDMA amfetamin
1. ábra: A 3,4-metilén-dioxi-metamfetamin (MDMA) és az amfetamin szerkezeti
képlete
1.2 A szerotonerg rendszer működésének áttekintése
Számtalan vizsgálat bizonyítja, hogy az MDMA a szerotonerg rendszer tartós
károsodását okozza anélkül, hogy hasonlóan károsítaná a többi neurotranszmitter
rendszer idegsejtjeit (Green és mtsai 2003). Hatásmechanizmusa, valamint akut és
krónikus hatásainak értelmezése előtt ezért fontosnak tartom a szerotonerg rendszer
működésének rövid áttekintését.
1.2.1 A szerotonin (5-HT) szintézise és felszabadulása az idegsejtekből
A központi idegrenszerben lévő 5-HT (2. ábra) a szerotonerg neuronok
citoplazmájában szintetizálódik az esszenciális triptofán aminosavból. A szintézis
sebesség-meghatározó lépése a triptofán-hidroxiláz (TPH) által katalizált reakció,
melynek során 5-hidroxi-triptofán (5-HTP) keletkezik. A következő lépésben az 5-HTP
dekarboxiláció útján 5-HT-ná alakul, ami a preszinaptikus axonvégződésekben
található vezikulákba transzportálódik a vezikulák membránjához kötött vezikuláris
monoamin transzporterek (VMAT) segítségével (Rudnick és Clark 1993; Liu és
Edwards 1997). A VMAT-ek 12 transzmembrán doménnel rendelkező H+/ATP-áz
8

pumpák, melyek az 5-HT transzportjával párhuzamosan H+-t pumpálnak ki a
vezikulákból és a dopamin és a noradrenalin vezikuláris transzportját is biztosítják a
dopaminerg illetve noradrenerg neuronok axonvégződéseiben. A VMAT-eknek két
izoformája ismert: a VMAT1 és a VMAT2, a központi idegrendszerben szinte kizárólag
a VMAT2 fordul elő (Rudnick és Clark 1993; Liu és Edwards 1997).
A szerotonerg neuronok kisülésekor az axonvégződések vezikuláiban
raktározódó 5-HT exocitózissal a szinaptikus résbe ürül, ahol a posztszinaptikus
membrán 5-HT receptoraihoz kapcsolódva fejti ki hatásait (Azmitia 2000).
5-HT
2.ábra: A szerotonin (5-HT) szerkezeti képlete
1.2.2 Az 5-HT visszavétele és lebontása
A szinaptikus résbe ürült 5-HT egy része a preszinaptikus membránban lévő
szerotonin transzproteren (SERT) keresztül visszajut a szerotonerg neuronba (Azmitia
2000). A SERT egy 12 transzmembrán doménnel rendelkező fehérje, a Na+/Cl--függő
neurotranszmitter transzporter család tagja, ehhez a családhoz tartozik a dopamin
transzporter (DAT) és a noradrenalin transzporter (NAT) is. Az 5-HT transzportjához
szükséges energiát a Na+ és K+ kationok koncentráció- és feszültséggrádiensének
irányába történő kotranszportja biztosítja. Az 5-HT-nal együtt ezért az idegsejtbe jut egy
Na+ és egy Cl- is, miközben az intracelluláris térből egy K+ a szinaptikus résbe
transzportálódik (Rudnick és Clark 1993; Malpass és mtsai 1999).
9

Az axonvégződésekbe visszakerült 5-HT egy része újra a vezikulákba
transzportálódik, míg a többi a citoplazmában marad. A citoplazmában az 5-HT-t a
mitokondrium membránján elhelyezkedő MAO-B (monoamin oxidáz B) enzim bontja
(Azmitia 2000; Holschneider 2000). A MAO (monoamin oxidáz) enzimnek két
izoformája ismert: a MAO-A (monoamin oxidáz A) és a MAO-B, melyeknek eltérő a
szubsztrátspecificitása és a gátlószerek iránti érzékenysége (Holschneider 2000). A
szerotonerg neuronokban a MAO-B enzim található, amelynek az 5-HT iránti affinitása
viszonylag alacsony (jóval kisebb, mint a MAO-A enzimé), ezért az idegsejten belül az
5-HT bontása lassú (Westlund és mtsai 1988). A citoplazmában ennek köszönhetően
jelentős mennyiségű 5-HT marad, amit különböző vegyületek pl. MDMA, para-klór-
amfetamin, fenfluramin a SERT-en keresztül – depolarizációtól független módon – fel
tudnak szabadítani (Berger és mtsai 1992). A visszavételre nem került, extracelluláris
térbe jutott 5-HT-t a környezetben lévő különböző neuronok (pl. a dopaminerg neuron)
és asztrociták képesek felvenni és lebontani a bennük található, az 5-HT-t nagy
affinitással bontó MAO-A enzim segítségével (Azmitia 2000; Holschneider 2000).
.
1.2.3 Az 5-HT receptorai
Az 5-HT meglehetősen sokszínű hatásait pre- és posztszinaptikusan
elhelyezkedő receptorain keresztül fejti ki. Az utóbbi évtizedekben a farmakológia és a
molekuláris biológia fejlődésének köszönhetően ugrásszerűen megnőttek az ismereteink
az 5-HT receptorokról. Hoyer és munkatársai 1994-ben (Hoyer és mtsai 1994) 7 típusba
sorolták az 5-HT receptorokat farmakológiájuk, aminosavszekvenciájuk és
szignáltranszdukciós mechanizmusuk alapján (1. táblázat). Az elmúlt 20 évben
klónozásos vizsgálatokkal bizonyították, hogy legalább 14 altípust is
megkülönböztethetünk, amelyeket különböző gének kódolnak. Mára több altípusnak
különböző splice-variánsait is azonosították, s ezalapján kijelenthetjük, hogy legalább
30 különböző fehérjeszerkezetű 5-HT receptor létezik (Raymond és mtsai 2001; Oliver
von Bohlen und Halbach 2006). Az 5-HT3 receptorok ligand-függő ioncsatornák, míg a
többi 5-HT receptor a hét transzmembrán doménnel rendelkező receptorok családjába
tartozik. A hét tramszmembrán doménnel rendelkező receptorok szignáltranszdukciója
heterotrimer G-fehérjékhez kapcsolt. Az 5-HT1 receptorok a Gi/0 fehérje közvetítésével
10

gátolják az adenil-ciklázt. Az 5-HT2 receptorok a Gq fehérje serkentésén keresztül a
foszfolipáz C-t (PLC) aktiválják. A PLC által katalizált reakcióban inozitol-trifoszfát
(IP3) és diacil-glicerol (DAG) keletkezik. Az IP3 az intracelluláris Ca2+-koncentráció
fokozódását váltja ki, a DAG pedig a PKC fehérjecsaládot aktiválja. Az 5-HT4, 5-HT5A,
5-HT6 és 5-HT7 receptorok Gs fehérje közvetítésével aktiválják az adenil-ciklázt, amely
a protein kináz A stimulációjához vezet (Sanders-Bush 1998; Aghajanian 2002). Az 5-
HT5B-receptor szignáltranszdukciós mechanizmusa még nincs teljes részleteiben
megfejtve (Oliver von Bohlen und Halbach 2006).
1. táblázat: Az 5-HT receptorok csoportosítása (zárójelben a receptorok korábbi
elnevezései vannak feltüntetve; PLC: foszfolipáz C, IP3: inozitol-trifoszfát, DAG:
diacil-glicerol)
Az 5-HT receptorok típusai és altípusai
5-HT1A
5-HT1B (Dβ)
5-HT1D (Dα)
5-HT1E
5-HT1F
5-HT2A (2)
5-HT2B (2F)
5-HT2C (1C)
5-HT3 5-HT4 5-HT5A
5-HT5B
5-HT6 5-HT7
Szignáltranszdukciós mechanizmus
Gi/0-
fehérjén
keresztül
Adenil-
cikláz ↓
Gq-
fehérjén
keresztül
PLC ↑
IP3/DAG ↑
Ligand-függő
ioncsatorna
Na+-, K+-
csatorna ↑
Gs-
fehérjén
keresztül
Adenil-
cikláz ↑
5A: Gs-
fehérjén
keresztül
5B: ?
5A:
Adenil-
cikláz ↑
5B: ?
Gs-
fehérjén
keresztül
Adenil-
cikláz ↑
Gs-
fehérjén
keresztül
Adenil-
cikláz ↑
11

1.2.4 A szerotonerg rendszer anatómiája
A szerotonerg neuronok az agytörzsben, kevés kivételtől eltekintve a raphe
magvakban találhatók (Azmitia 2000; Hornung 2003). A raphe magvak a nyúltvelőtől a
középagyig terjednek ki és 9 régióra oszthatók: raphe pallidus (B1), raphe obscurus
(B2), raphe magnus (B3), a nyúltvelő központi szürkeállománya (B4), a hídi mediális
raphe (B5), a hídi dorzális raphe (B6), a középagyi dorzális raphe (B7), caudális lineáris
mag (B8) és a lemniscus mediális (B9) (Tohyama 1998). A caudalis (inferior) raphe
magvakból (B1-B4) kiinduló axonjaik a gerincvelőhöz futnak, ezek az axonok alkotják
a leszálló szerotonerg rendszert (Azmitia 2000; Hornung 2003). A rostralis (superior)
raphe magvakból (B5-B9) kiinduló pályák (felszálló szerotonerg rendszer) egyéb
agytörzsi struktúrákhoz, valamint a hipotalamuszhoz, a limbikus rendszerhez, a
hippocampushoz és az agykéreghez futnak, de kisebb-nagyobb sűrűségben az agyban
mindenütt kimutathatók (Tork 1990; Jacobs és Azmitia 1992; Azmitia 2000; Hornung
2003). A neurotranszmitter rendszerek közül a szerotoniné a legkiterjedtebb, rendkívül
szerteágazó, azonban igen különböző az egyes területek beidegzettségének mértéke
(Jacobs 2000). Egyes agyterületeken komplex, több különböző raphe régióból származó
szerotonerg beidegzésre is fény derült (Palkovits 1987; Takeuchi 1988; Murphy 1989;
Murphy és mtsai 1991).
Az 5-HT a szervezetben a központi idegrendszeren kívül a periférián is
előfordul: nagy mennyiségben megtalálható a gyomor-béltraktusban, vérlemezkékben,
hízósejtekben és a mellékvese velőállományában (Zifa és Fillion 1992). Széleskörű
előfordulásából következően fontos szerepet játszik számos élettani folyamat: többek
között a testhőmérséklet (Schwartz és mtsai 1995), az endokrin rendszer (Ruggiero és
mtsai 1999; Lowry 2002), a szexuális funkciók (Meston és Gorzalka 1992), a
táplálékfelvétel (Levine és mtsai 1987; Guy-Grand 1995), a fájdalom (Le Bars 1994), a
tanulás és a memória (McEntee és Crook 1991), a napszaki változások, az alvás (Portas
és mtsai 2000) és a motoros aktivitás (Jacobs és Azmitia 1992; Jacobs és Fornal 1997)
szabályozásában. A dolgozatom témájából adódóan az 5-HT-nak az alvás és a motoros
aktivitás szabályozásában betöltött szerepét fogom részletesen ismertetni a következő
fejezetekben.
12

1.2.5 Az 5-HT szerepe az alvás szabályozásában
Az alvásszabályozásban részt vevő szerotonerg neuronok a dorsalis és medialis
raphe magvakban helyezkednek el, nyúlványaik a lateralis hypothalamuson futnak
keresztül és más, az alvásszabályozásban ugyancsak részt vevő (noradrenerg,
hisztaminerg, orexinerg) neuronok nyúlványaival együtt sűrűn behálózzák a cortexet
(Saper és mtsai 2001). Számos emlős állatfajban bizonyították, hogy a raphe magvak
szerotonerg neuronjainak lassú, tónusos tüzelési aktivitása van feltehetően belső,
tónusos pacemaker mechanizmusoknak köszönhetően (Aghajanian 2002). Ébrenlét alatt
a szerotonerg neuronok fokozott aktivitása figyelhető meg, NREM-alvás (non-rapid eye
movement, lassú hullámú alvás) alatt ehhez képest csökken az aktivitás, míg a REM-
alvás (rapid eye movement, paradox alvás) ideje alatt egyáltalán nem sülnek ki ezek az
idegsejtek (Portas és mtsai 2000; Pace-Schott 2002). Már az 1970-es években felmerült,
hogy a szerotonerg neuronok és a locus coeruleusban, valamint a peribrachialis magban
elhelyezkedő, nagyrészt noradrenerg neuronok gátolják a REM-alvást kiváltó, kolinerg
ún. REM-on neuronokat. Ez az oka annak, hogy a szervezet akkor kerül a REM-alvás
fázisába, amikor felfüggesztődik működésük (McCarley és Hobson 1975; Bódizs 2000).
Ma is vitatott, hogy az 5-HT mely receptorain keresztül, milyen módon vesz
részt az alvás-ébrenléti ciklus szabályozásában. A REM-alvás gátlásában úgy tűnik
legnagyobb szerepe az 5-HT1A receptoroknak van (Monti és Monti 2000; Filakovszky
és mtsai 2001; Monti és Jantos 2004). Emellett 5-HT2B (Popa és mtsai 2005) és 5-HT2C
(Martin és mtsai 1998) receptor agonistákról és 5-HT2A receptor antagonistákról
(Tortella és mtsai 1989; Monti és Jantos 2006) bizonyították, hogy csökkentik a REM-
alvás időtartamát. Meglepő módon viszont az 5-HT2B (Kantor és mtsai 2004; Popa és
mtsai 2005) és az 5-HT2C (Smith és mtsai 2002; Popa és mtsai 2005; Monti és Jantos
2006) receptorok antagonistái is gátolják a REM-alvást. Részt vesznek a REM-alvás
szabályozásában az 5-HT7 receptorok is (Thomas és mtsai 2003; Monti és Jantos 2006;
Bonaventure és mtsai 2007). Az ébrenlét és a NREM-alvás szabályozásában ugyancsak
több 5-HT-receptor játszik szerepet. Bizonyított, hogy az 5-HT1A (Monti és Jantos 1992;
Filakovszky és mtsai 2001), az 5-HT3 (Monti és mtsai 1999; Kantor és mtsai 2000) és
az 5-HT2C (Martin és mtsai 1998; Kantor és mtsai 2005) receptorok agonistái fokozzák
az ébrenlétet. Az 5-HT2A antagonisták és az 5-HT2B agonisták csökkentik az ébrenlét
13

idejét és növelik a NREM-alvás időtartamát (Popa és mtsai 2005), míg az 5-HT3 (Monti
és mtsai 1999; Kantor és mtsai 2000) és az 5-HT2A (Dugovic és mtsai 1989; Monti és
Jantos 1992; Kantor és mtsai 2000) receptorok agonistái csökkentik a NREM-alvás
időtartamát.
1.2.6 Az 5-HT szerepe a motoros aktivitás szabályozásában
Általános összefüggés van a tónusos motoros aktivitás, az éberség mértéke és a
szerotonerg neuronok aktivitása között, beleértve mind a superior, mind az inferior
magvakban elhelyezkedő idegsejtek működését (Jacobs és Fornal 1997; Frazer 1999;
Jacobs és Fornal 1999). A szerotonerg neuronok pacemaker aktivitását számos
neurotranszmitter, például a noradrenalin (NA) és maga az 5-HT is befolyásolja. A NA
α1-receptorokon keresztül fokozza a szerotonerg neuronok pacemaker aktivitását, míg
az 5-HT az 5-HT1A autoreceptorokon keresztül csökkenti azt, így korlátozva negatív
feedback mechanizmussal az idegsejtek aktivitását (Aghajanian 2002).
Az 5-HT és a motoros rendszer szoros kapcsolatának megfelelően az elsődleges
és másodlagos motoros területek nagyon sűrű szerotonerg beidegzéssel rendelkeznek:
patkányban a gerincvelő elülső szarvának, a trigeminális motoros magnak (a V. agyideg
magva), a faciális motoros magnak (az VII. agyideg magva), a substancia nigrának és a
globus pallidusnak igen sűrű a beidegzése (Jacobs 2000). Érdekesség, hogy a motoros
területek beidegzése nem homogén: a gerincvelőben sűrű beidegzéssel rendelkeznek az
axiális izmok, míg a disztálisak beidegzése ritka (Steinbusch 1981). Az agytörzsben
sűrű beidegzése van az áll, az arc és a nyak nagy izmaihoz vezető motoneuronoknak,
míg a szemmozgató izmok motoneuronjai viszonylag gyér beidegzéssel rendelkeznek
(Steinbusch 1981; Takeuchi és mtsai 1983). A finommozgásokat koordináló kisagyhoz
is kevés szerotonerg nyúlvány fut. A motoros területek különböző beidegzése sokat
elárul a szerotonerg rendszer szerepéről a motoros aktivitás szabályozásában. A
szerotonin-tartalmú idegsejtek fokozott működése figyelhető meg a nagyobb izmokat
igénybevevő, tónusos és repetitív mozgások esetén, például rágás, grooming (sztereotip
mosakodás), futás közben, míg a finommozgások alatt nem fokozódik aktivitásuk
(Jacobs 2000).
14

Jacobs és Fornal (1999) alkották meg azt a hipotézist, miszerint a szerotonin
rendszer fő funkciója a szervezet motoros működésének elősegítése (Jacobs és Fornal
1999). Másodlagos funkciói: az autonóm és a neuroendokrin rendszer szabályozása az
aktuális motoros működéstől függően, például részt vesz a vér oxigenizációjának, a
vázizmok vérellátásának, valamint az agy glükózellátásának fokozásában (Jacobs és
Fornal 1999; Jacobs 2000). Gátolja az ingerületátvitelt a legtöbb szenzoros idegpályán,
ez az oka annak, hogy amikor a szerotonin rendszer működése gátlódik figyelmi és
orientációs stimulusok következtében, megszűnik a motoros rendszer működésének
serkentése és felszabadulnak a szenzoros információkat közvetítő idegpályák a gátlás
alól (Frazer 1999; Jacobs és Fornal 1999; Jacobs 2000).
Több 5-HT receptorról is kimutatták, hogy részt vesz a motoros aktivitás
szabályozásában. Az 5-HT2C receptorok agonistái gátolják (Martin és mtsai 1998;
Bagdy és mtsai 2001), míg az 5-HT2B antagonista SB 215505 fokozza (Kantor és mtsai
2004) és a nem szubtípus szelektív 5-HT2 receptor antagonista ritanserin csökkenti a
motoros aktivitást (Kantor és mtsai 2002). Az 5-HT izgatja a gerincvelőben a
motoneuronokat 5-HT2A és 5-HT7 receptorok közvetítésével (Inoue és mtsai 2002;
Gilmore és Fedirchuk 2004; Liu és Jordan 2005; Jordan és mtsai 2008) és ezen
receptorok antagonistái ugyanott gátolják a motoros aktivitást (Jordan és Schmidt 2002;
Madriaga és mtsai 2004; Liu és Jordan 2005; Jordan és mtsai 2008). Az 5-HT1B
receptornak is fontos szerepe van a lokomotoros aktivitás szabályozásában (Sari 2004),
agonistái fokozzák a lokomotoros aktivitást (Green és mtsai 1984; Chaouloff és mtsai
1999).
A szerotonin rendszer működésének fokozása (például 5-HTP-nal vagy L-
triptofánnal) emlősállatokban szerotonin viselkedési szindrómát okoz, amelyet a
következők jellemeznek: megnövekedett lokomotoros aktivitás, tremor, ataxia, head-
weaving (fej fordítása egyik oldalról a másikra), hátsó végtag abductioja, piloerectio
(szőrszálak felborzolódása), nyálfolyás, székletürítés (Jacobs 2000; Green és mtsai
2003). Ugyanez történik az 5-HT felszabadulását fokozó indirekt agonisták, például az
MDMA alkalmazása esetén is (Geyer 1996).
15

1.3 A dopaminerg, a noradrenerg és a kolinerg rendszer szerepe az alvás és a
motoros aktivitás szabályozásában
Az MDMA, mint erről a következő fejezetben részletesen is írok nagymértékű,
akut 5-HT-felszabadító hatása mellett dopamint, noradrenalint és acetilkolint is
felszabadít az idegsejtekből. Ezeknek a neurotranszmittereknek is fontos szerepük van
az alvás illetve a motoros aktivitás szabályozásában, s ezért az MDMA – általam is
vizsgált – akut motoros aktivitás és alváshatásaiban.
A lokomotoros aktivitást fokozó környezeti ingerek növelik a dopamin
neurotranszmissziót és fordítva: a dopamin neurotranszmisszió növelésével a dopamin
agonisták fokozzák a lokomotoros aktivitást és az ébrenlétet emlősökben, míg az
antagonisták csökkentik (Brown és mtsai 2007; Sadock 2007). A dopaminerg rendszer
mesolimbicus pályája közvetíti a pszichostimulánsok lokomotoros aktivitást fokozó
hatását (Alcaro és mtsai 2007).
A locus coeruleusban, valamint a peribrachialis magban elhelyezkedő
noradrenerg neuronok kisülése ébrenlét alatt fokozott, a NREM-alvás ideje alatt
csökkent és a REM-alvás ideje alatt felfüggesztődik. Csakúgy, mint a szerotonerg
neuronoknak, az ébrenlét fenntartásában és a REM-alvás gátlásában van szerepük
(Bódizs 2000; Pace-Schott 2002). A locus coeruleus noradrenerg neuronjai különösen
aktívak stressz vagy nagyfokú éberség, figyelem esetén (Robbins 2000).
A hídban található pedunculopontinus és laterodorsalis tegmentális magvak
kolinerg neuronjai is aktívak az ébrenléti fázis alatt, aktivitásukkal hozzájárulnak az
ébrenlét fenntartásához (Saper és mtsai 2001; Pace-Schott 2002). Emellett a REM-alvás
létrejöttében és fenntartásában játszanak központi szerepet, ezek az ún. REM-on
neuronok; a REM-alvás ideje alatt különösen aktívak (Bódizs 2000; Pace-Schott 2002).
Ha kolinerg agonistát vagy acetil-kolinészteráz (az acetilkolint lebontó enzim) gátlót
juttatnak erre a területre, az REM-alvást vált ki kísérleti állatokban (Capece és mtsai
1997). Valószínűleg az agytörzsi retikuláris rendszer M2 receptorainak aktiválása a fő
mechanizmusa a REM-alvás kiváltásának (Baghdoyan és Lydic 1999). A szerotonerg
neuronok posztszinaptikus 5-HT1A receptorokon keresztül gátolják a REM-alvást
kiváltó kolinerg neuronokat (Monti és Monti 2000; Monti és Jantos 2004). A
noradrenerg neuronok REM-alvást gátló hatásában pedig úgy tűnik, mind az α1, mind az
16

α2 receptorok közvetítő szerepet játszanak (Williams és Reiner 1993; Ross és mtsai
1995).
A dopamin-felszabadulás nem változik olyan drámaian az alvás-ébrenléti ciklus
során, mint az 5-HT, a noradrenalin és az acetilkolin felszabadulása. REM-alvás
depriváció azonban növeli a dopamin-felszabadulást és a dopamin receptorok
érzékenységét (Pace-Schott 2002). Dopamin-reaptake gátlók és indirekt agonisták
növelik a REM-alvás időtartamát emberben (Nofzinger és mtsai 1995). A dopamin
fokozza a kortikális acetilkolin-felszabadulást, míg a REM-on neuronok fokozzák a
mesolimbikus pályán a dopamin-felszabadulást. Kölcsönös serkentés van tehát a
kolinerg és a dopaminerg rendszer között, aminek a REM-alvás fenntartásában és
mélyítésében lehet szerepe (Pace-Schott 2002).
1.4 Az MDMA akut hatásai állatokban
Az MDMA akut hatásai körülbelül 24 órán át tartanak (Schmidt és mtsai 1986;
Schmidt 1987; McKenna és Peroutka 1990), ezt követően hoszútávú, neurotoxikus
hatásai 12 hónapig, sőt tovább is fennmaradhatnak a kísérleti állatokban (Battaglia és
mtsai 1988; Scanzello és mtsai 1993). Akut viselkedési hatásainak egyedülálló
kombinációja annak köszönhető, hogy erőteljesen fokozza a dopamin, 5-HT,
noradrenalin és acetilkolin felszabadulását a preszinaptikus idegvégződésekből (Cole és
Sumnall 2003; Green és mtsai 2003; Gudelsky és Yamamoto 2007). Kisebb mértékben
az is hozzájárul hatásai sokszínűségéhez, hogy számos neurotranszmitter receptorhoz
kötődik: többek között az 5-HT2A-, α2-, M1-, H1- és M2-receptorokhoz (Cole és Sumnall
2003; Green és mtsai 2003).
Az MDMA királis molekula és sztereoizomérjei közül, az S (+) izomér
erőteljesebb monoamin-felszabadító hatással rendelkezik, hatásai jobban hasonlítanak
az amfetaminéra, míg az R (-) izomérre jellemzőbbek a meszkalin- és LSD-szerű
hatások (Kalant 2001; Baumann és mtsai 2007). Többnyire racém elegyét állítják elő és
használják az állatkísérletek során és az ecstasytabletták is ebben a formában
tartalmazzák az MDMA-t (Kalant 2001).
17

1.4.1 A monoaminok felszabadítása az agyban
Az SERT nagy affinitással köti az MDMA-t és aktív transzporttal, 5-HT-
MDMA csere mechanizmussal az idegsejtbe pumpálja. Az idegsejtbe jutott MDMA
fokozza az 5-HT felszabadulását exocitózis útján (Ca2+-függő felszabadulás) és Ca2+-tól
független mechanizmussal a SERT-en keresztül is (Crespi és mtsai 1997). A
transzporter intracelluláris részéhez kapcsolódva annak konformációváltozását idézi elő,
aminek hatására a transzport megfordul, és az 5-HT a szinaptikus térbe ürül (Levi és
Raiteri 1993; Rudnick és Clark 1993; Rothman és Baumann 2002). Az 5-HT kiürülését
tovább fokozza, hogy az MDMA a VMAT transzportját megfordítva a vezikulákból is
felszabadítja az 5-HT-t és így erőteljesen megnöveli a szinaptikus koncentrációt
(Rudnick és Clark 1993; Rothman és Baumann 2002). Az MDMA ezenkívül a MAO
enzimeket is gátolja, mind a MAO-A-t, mind a MAO-B-t, de az 5-HT-t nagyobb
affinitással bontó MAO-A-t mintegy tízszer erősebben (Leonardi és Azmitia 1994). A
kiáramlott 5-HT-t az idegsejtek nem tudják újraszintetizálni, mert az MDMA gátolja az
5-HT-szintézis sebességmeghatározó enzimének, a TPH enzimnek az aktivitását (Cole
és Sumnall 2003; Green és mtsai 2003).
Az MDMA által indukált noradrenalin-felszabadulás az 5-HT felszabadításához
hasonlóan történik (Baumann és mtsai 2007), a NAT (noradrenalin transzporter) is
jelentős, bár a SERT-nél kisebb affinitással köti az MDMA-t, ennek megfelelően a
noradrenalin-felszabadulás az 5-HT-énál kisebb mértékű MDMA hatására (Rothman és
Baumann 2002).
A DAT (dopamin transzporter) a NAT-nél is jóval kisebb affinitással köti az
MDMA-t (Rothman és Baumann 2002), ennek ellenére MDMA adását követően igen
erőteljes dopamin felszabadulás mérhető in vivo mikrodialízissel patkányokban (Nash
és Yamamoto 1992; Yamamoto és mtsai 1995; Colado és mtsai 1999). Megállapították
azt is, hogy a felszabadított dopamin mennyisége összefüggésben áll a felszabadított 5-
HT mennyiségével (Cole és Sumnall 2003). Ennek okát abban sejtik, hogy a
szerotonerg neuronokból felszabaduló 5-HT a striatumban és a substantia nigra-ban
elhelyezkedő GABA- (γ-amino-vajsav)-erg interneuronokat az 5-HT2A receptorok
közvetítésével gátolja, aminek hatására a dopaminerg neuronok felszabadulnak a
GABA-erg gátlás alól és kisülnek (több dopamin szintetizálódik és szabadul fel) (Ball
18

és Rebec 2005). Ugyanez történik a ventrális tegmentális áreaban is, ahol a GABA-erg
neuronok 5-HT2B/2C receptorainak aktiválása csökkent GABA-felszabadulással jár és
ennek hatására nő a dopamin-felszabadulás a nucleus accumbens-ben (Bankson és
Yamamoto 2004). Kimutatták azt is, hogy a NAT gátló desipramin is csökkenti az
MDMA-val kiváltott dopamin-felszabadulást a hippocampusban, amiből arra
következtettek, hogy a NAT által a noradrenerg neuronokba felvett MDMA fokozhatja
az ezekbe az idegsejtekbe bekerült dopamin felszabadulását (Shankaran és Gudelsky
1998).
Az akut monoaminfelszabadító hatás eredményeképpen a monoaminok
kiürülnek a neuronokból és míg a noradrenalin- és a dopaminraktárak néhány napon
belül újratöltődnek, az 5-HT tartós deplécióját okozza az MDMA (Green és mtsai
2003).
1.4.2 Az acetilkolin felszabadítása az agyban
Fischer és munkatársai (2000) mutatták ki először, hogy az MDMA patkány
stiatumában in vitro fokozza az acetilkolin-felszabadulást. Bizonyították azt is, hogy
H1-receptor antagonistákkal csökkenthető az MDMA-t követő acetilkolin-felszabadulás
(Fischer és mtsai 2000). Acquas és munkatársai (2001) ezután in vivo mikrodialízissel
bizonyították, hogy patkány striatumában és frontális cortexében is fokozódik az
acetilkolin felszabadulás MDMA hatására és ugyancsak az MDMA H1-receptorokhoz
kötődésének szerepét valószínűsítették a hatásban. Néhány évvel később ugyancsak in
vivo mikrodialízissel kimutatták, hogy az MDMA-t követő acetilkolin-felszabadulás a
prefrontális kéregben para-klór-fenilalanin (5-HT-szintézis gátló) és α-metil-para-tirozin
(dopamin-szintézis gátló), hatására csökken, viszont a dorzális hippocampusban az
acetilkolin-felszabadulást nem befolyásolják az említett gátlószerek (Nair és Gudelsky
2006). A prefrontális cortexben kimutatták, hogy D1-receptor antagonista és 5-HT4
receptor antagonista is csökkenti az acetilkolin-felszabadulást patkányokban (Nair és
Gudelsky 2005). A prefrontális cortexben ezalapján a szerotonerg és a dopaminerg
rendszerek szerepét feltételezhetjük az acetilkolin-felszabadítás mechanizmusában (Nair
és Gudelsky 2005; Nair és Gudelsky 2006; Gudelsky és Yamamoto 2007).
Elképzelhető, hogy az MDMA pszichomotoros stimuláns hatásainak egy részéért az
19

acetilkolin-felszabadulás felel (Cole és Sumnall 2003; Gudelsky és Yamamoto 2007),
mert számos pszichostimulánsról (pl.: kokain, amfetamin, metamfetamin) kimutatták,
hogy fokozza az acetilkolin-felszabadulást (Day és Fibiger 1992; Imperato és mtsai
1993; Taguchi és mtsai 1998; Arnold és mtsai 2001).
1.4.3 Az MDMA akut élettani és viselkedési hatásai állatokban
Az MDMA szobahőmérsékleten adva 1-2 oC-kal emeli a patkányok
testhőmérsékletét, 40-60 perccel adása után a legnagyobb a hőmérsékletváltozás
(Schmidt és mtsai 1990; Colado és mtsai 1993; O'Shea és mtsai 1998). A
testhőmérséklet-változás dózisfüggő (Dafters 1994; Dafters és Lynch 1998; Adori és
mtsai 2006) és nagyban befolyásolja a külső hőmérséklet is: magas hőmérsékleten adva
az MDMA erőteljesebben emeli a testhőmérsékletet, míg alacsony hőmérsékleten adva
általában csökkenti (Gordon és mtsai 1991; Dafters 1995; Dafters és Lynch 1998). A
hipertermiás válasz nagysága összefüggésben áll a monoamin-felszabadulás erősségével
(Green és mtsai 2004a). A testhőmérséklet emelkedésében úgy tűnik jelentős szerepe
van az 5-HT-felszabadulás hatására aktiválódott 5-HT2A/2C receptoroknak (Schmidt és
mtsai 1990; Green és mtsai 1995; O'Loinsigh és mtsai 2001; Herin és mtsai 2005). A
Dark Agouti patkányokkal történt vizsgálatok alapján viszont egyre inkább
valószínűsítik a D1-receptorok alapvető szerepét is a hipertermiás válaszban (Colado és
mtsai 1999; Mechan és mtsai 2002).
Más 5-HT-felszabadító drogokhoz és 5-HT-agonistákhoz (Levi 1994) hasonlóan
az MDMA is megemeli patkányokban a kortizon- és a prolaktin-szintet (Nash és mtsai
1988).
Gordon és munkatársai (1991) leírták, hogy az MDMA patkányokban arritmiát
okoz és növeli a szívfrekvenciájukat. Érösszehúzódást kiváltó és artériás vérnyomást
növelő hatása is ismert (Fitzgerald és Reid 1994; Vollenweider és mtsai 1998; Bexis és
Docherty 2006). A kardiovaszkuláris változások renin-felszabadulással járnak, aminek
hatására nő az angiotenzin-II-szint (Burns és mtsai 1996). Az angiotenzin-II
érösszehúzódást okoz és fokozza az aldoszteron felszabadulását a mellékvesekéregből,
mindezek a vérnyomás emelkedéséhez vezetnek (Fürst 2006). A kardiovaszkuláris
hatások úgy tűnik, nagyrészt az MDMA által indukált noradrenalin-felszabadulásnak
köszönhetőek (Fitzgerald és Reid 1994), de hozzájárulnak az MDMA α2- és 5-HT2-
20

receptorokra kifejtett agonista hatásai is (Lyon és mtsai 1986; Battaglia és De Souza
1989; Lavelle és mtsai 1999).
Az agyban fiziológiás körülmények között szoros kapcsolat áll fenn a
vérkeringés és a glükózfelhasználás között (Kuschinsky 1991). Az MDMA akutan
számos agyterületen fokozza a glükózfelhasználást (Frederick és Paule 1997; Balogh és
mtsai 2004; Ando és mtsai 2006; Ferrington és mtsai 2006; Kirilly és mtsai 2006),
ugyanakkor ezek egy részében csökkenti a vérkeringést (Quate és mtsai 2004;
Ferrington és mtsai 2006). Az agyi vérkeringés csökkenésének feltehetően az az oka,
hogy a felszabaduló 5-HT érösszehúzódást vált ki az agyban 5-HT1B- (Craig és Martin
1993) és 5-HT2-es receptorokon keresztül (Chang és Owman 1987). A
glükózfelhasználás fokozódása és egyidejűleg a vérkeringés csökkenése, energiahiányos
állapotot hoz létre, ami hozzájárulhat a neurotoxicitás kialakulásához (Nixdorf és mtsai
2001).
Az MDMA akutan, dózisfüggő módon fokozza az állatok lokomotoros
aktivitását (Spanos és Yamamoto 1989; Callaway és Geyer 1992; Dafters 1994;
McNamara és mtsai 1995; Bengel és mtsai 1998). Érdekesség, hogy míg a kokain és az
amfetamin egyenletesen, minden irányba kiterjedően fokozzák a motoros aktivitást; az
MDMA hatására az állatok rotációs mozgást végeznek, körbe-körbe futnak a ketrecben
(Callaway és mtsai 1990; McCreary és mtsai 1999; Scearce-Levie és mtsai 1999). A
klasszikus, 5-HT-felszabadulást kiváltó hallucinogének hatására is hasonló
mozgássorozat látható az állatokon (Callaway és mtsai 1991; Cole és Sumnall 2003).
Megfigyelték azt is, hogy nagyobb dózis MDMA ismétlődő, sztereotíp
mozgássorozatokat vált ki, amelyek a szerotonin-szindróma elemeire emlékeztetnek
(Spanos és Yamamoto 1989). Úgy tűnik, hogy az 5-HT-felszabadulás és az 5-HT1B
receptorok aktivációja alapvetően fontos a motoros aktivitás-fokozódásban, mert
genetikailag módosított SERT-hiányos egerekben hiányzik (Bengel és mtsai 1998), míg
5-HT1B-hiányos egerekben jelentősen csökkent az MDMA-nak ez a hatása (Scearce-
Levie és mtsai 1999). Ráadásul az 5-HT1B receptor agonisták hasonló lokomotoros
hiperaktivitást váltanak ki, mint az MDMA (Rempel és mtsai 1993). A folyamat
összetettségére utal, hogy 5-HT2A receptor antagonista csökkenti az MDMA-t követő
lokomotoros aktivitás-fokozódást (Bankson és Cunningham 2002; Herin és mtsai 2005),
míg az 5-HT2C receptor antagonista potencírozza az MDMA ezen hatását (Bankson és
21

Cunningham 2002; Fletcher és mtsai 2006). Később bizonyították, hogy az 5-HT-
felszabadulás mellett a dopamin-felszabadulásnak is szerepe van az MDMA-t követő
lokomotoros aktivitás-fokozódásban, mert az dopamin receptor antagonistákkal
gátolható (Ball és mtsai 2003; Bubar és mtsai 2004). Nemrégiben knock-out egereken is
megvizsgálták a D1, a D2 és a D3 receptorok szerepét: a D1-hiányos egerek MDMA
hatására fokozott hiperaktivitást, míg a D2- és a D3-hiányos egerek csökkent
hiperaktivitást mutattak a vad típusú, MDMA-val kezeltekhez képest (Risbrough és
mtsai 2006). Ezt követően a noradrenalin-felszabadulás szerepét is bizonyították: az α1-
receptor gátló prazosin mind szisztémásan, mind lokálisan adagolva a prefrontális
kéregbe illetve a ventrális tegmentális áreába, gátolta a kis dózisú MDMA-által kiváltott
lokomotoros választ patkányokban (Selken és Nichols 2007).
Az MDMA-ról kimutatták, hogy akutan csökkenti az agresszivitást egerekben
(Navarro és Maldonado 1999; Maldonado és Navarro 2001) és patkányokban (Morley
és McGregor 2000; Kirilly és mtsai 2006). Az 5-HT1B receptor agonistái is antiagresszív
hatásúak (Bell és mtsai 1995; Fish és mtsai 1999; Clark és Neumaier 2001; Kirilly és
mtsai 2006), ezért ennek a receptortípusnak a közvetítő szerepét feltételezik a hatásban,
azonban valószínűnek tűnik a dopaminerg, a kolinerg, a GABA-erg és a glutamáterg
rendszerek résztvétele is (Sexton és mtsai 1999; Clark és Neumaier 2001). Az MDMA
akut anxiogén hatását is leírták különböző viselkedés-vizsgálatokban (pl.: szociális
interakciós teszt, „resident-intruder” teszt, megemelt keresztpalló teszt, „emergence”
teszt) (Bhattacharya SK 1998; Morley és McGregor 2000; Ando és mtsai 2006; Kirilly
és mtsai 2006). A szorongást fokozó hatásával lehet kapcsolatban, hogy több szorongás-
tesztben („open field” szorongás teszt, „resident-intruder” teszt, szociális interakciós
teszt) csökkentette a felfedező viselkedést, az ágaskodást és a lokomotoros aktivitást az
állatokban (Scearce-Levie és mtsai 1999; Ando és mtsai 2006; Kirilly és mtsai 2006).
Ezalapján úgy tűnik, hogy az MDMA hatására az állatok érzékenyebbekké válnak a
félelmet keltő szituációkra, s talán ez az oka, hogy fokozódik a szorongásuk, magával
vonzva a lokomotoros aktivitás csökkenését a szorongás-tesztekben (Cole és Sumnall
2003).
Az MDMA, főként nagy dózisban az állatok pusztulását okozhatja, többnyire a
testhőmérséklet-emelkedés és a szerotonin-szindróma együttes hatásaként. Nagyok az
egyéni érzékenységbeli különbségek, Sprague-Dawley és Dark Agouti patkányoknál is
22

leírtak az adott patkányfajra meghatározott LD50 dózisnál jóval kisebb dózisok esetén is
letalitást (Rochester és Kirchner 1999; Cole és Sumnall 2003).
1.4.4 Aggregációs toxicitás
Az aggregációs toxicitás fogalma először az 1940-es években merült fel az
amfetamin kutatása kapcsán, amikor észrevették, hogy amfetaminnal kezelt egereknél
súlyosabb toxicitás és jelentősebb viselkedési változások tapasztalhatók, ha a kísérlet
során egy ketrecben több egeret tartanak (Gunn és Gurd 1940; Chance 1947). A magas
hőmérséklet, az erős zaj és a folyadékhiány is növeli az amfetamin és a metamfetamin
toxicitását (Chance 1947; Morton és mtsai 2001). Később, az MDMA hatásait vizsgálva
kimutatták, hogy ha legalább 5 egeret tartottak egy ketrecben, az ötödére csökkent az
MDMA LD50 dózisa az egyedül tartott egerekéhez képest (Davis és Borne 1984). Az
egerekhez hasonlóan, embereknél is fokozhatják az ecstasy szedésének általános
körülményei a nem kívánt mellékhatások előfordulásának gyakoriságát és az előidézett
idegrendszeri károsodás mértékét (O'Shea és mtsai 1998; Green és mtsai 2003).
1.5 Az MDMA hosszútávú hatásai állatokban
Az MDMA fő célpontjai a szerotonerg neuronok, amelyekben tartós
neurotoxikus hatását mutatták ki számos állatfajban (Battaglia és mtsai 1987; Commins
és mtsai 1987; Schmidt 1987; Stone és mtsai 1987; McKenna és Peroutka 1990).
Egyedül az egerek tűnnek ellenállónak a szerotonerg idegsejteket károsító hatásával
szemben, viszont dopaminerg idegsejtjeik károsodnak (Colado és mtsai 2004), ennek
oka valószínűleg a többi fajtól jelentősen eltérő MDMA-metabolizmusuk (Escobedo és
mtsai 2005).
1.5.1 A neurotoxikus hatás feltételezett mechanizmusai
Az MDMA által okozott szerotonerg neuronkárosodás fő okát először a dopamin
központi idegrendszerbeli felhalmozódásában sejtették. Emellett szólt, hogy ha kísérleti
állatoknak L-DOPA-t, a dopamin prekurzorát adták MDMA kezelés előtt, még
23

súlyosabban károsodtak a szerotonerg idegsejtek (Levitt és mtsai 1982). További
bizonyítékul szolgált, hogy a dopamin-szintézis gátlása α-metil-p-tirozinnal (a tirozin-
hidroxiláz enzimet gátolja) vagy 6-hidroxi-dopaminnal (dopaminerg neurotoxin)
kivédte a neurotoxicitást (Stone és mtsai 1988; Schmidt és mtsai 1990).
Az elmélet szerint az MDMA kezelés után az extracelluláris térben felgyülemlett
dopamin eljut a kiürült szerotonerg idegsejtek SERT-eihez (Sprague és mtsai 1998). A
SERT csak kis affinitással köti a dopamint, ezért fiziológiás körülmények között csak
nagyon kevés dopamin jut be a szerotonin-tartalmú idegsejtekbe és a - MAO-B általi -
metabolizációja során felszabaduló szabadgyökök semlegesítődnek (Cadet és Brannock
1998). A kórosan felhalmozódott dopamin azonban jelentős mennyiségben bekerülhet a
szerotonerg neuronokba, ebben az esetben a nagy mennyiségben képződött
szabadgyököt az idegsejt nem tudja semlegesíteni és az károsítja (Levitt és mtsai 1982;
Cadet és Brannock 1998). A teória szerint ennek a következménye, hogy az 5-HT-nak
és metabolitjának, az 5-hidroxi-indolecetsav- (5-HIAA)-nak a szöveti koncentrációja,
valamint a SERT-ek denzitása tartósan csökken számos agyterületen (Schmidt és mtsai
1986; Stone és mtsai 1986; Schmidt 1987; Stone és mtsai 1987).
A dopaminteória mellett felvetődött az is, hogy az MDMA hosszútávú
neurotoxikus hatásait nem maga az MDMA, hanem metabolitjai okozzák, mert kísérleti
állatokban az MDMA intracerebrális injektálását követően nem jött létre szerotonerg
neurotoxicitás (Schmidt és Taylor 1988; Paris és Cunningham 1992; Esteban és mtsai
2001).
Az MDMA metabolizmusának első lépésében három metabolittá alakulhat: N-
demetilációval MDA-ná, demetilációval dihidroxi-metamfetaminná (DHMA) és gyűrűn
hidroxilálódva 2-hidroxi-4,5-metiléndioxi-metamfetaminná. Az MDA, az MDMA fő
metabolitja, ezt követően újra demetilálódik és dihidroxi-amfetamin (DHA) képződik
belőle. A DHA és a DHMA kinonnokká oxidálódnak, majd glutationnal konjugálódnak
(Hiramatsu és mtsai 1990; Tucker és mtsai 1994). A képződött glutation-
konjugátumokról feltételezik, hogy miután a vér-agy gáton átjutnak, majd bejutnak az
axonvégbe, reakcióba lépnek a TPH enzimeken található szulfhidril- (SH-)
csoportokkal, inaktiválják az enzimet, ami az 5-HT-szintézis gátlásához vezet és a
szerotonerg neuronok visszafordíthatatlan károsodását okozza (Rattray 1991; Tucker és
mtsai 1994; Capela és mtsai 2006). In vitro bizonyították súlyos, idegkárosító hatásaikat
24

(Jones és mtsai 2004; Capela és mtsai 2006). Az MDMA metabolizmusának bizonyos
lépései az agyban is végbemennek és az ezek során képződő szabadgyökök ugyancsak
károsíthatják a szerotonerg neuronok terminális szakaszait (Rattray 1991; Colado és
mtsai 1997; O'Shea és mtsai 1998; Lyles és Cadet 2003; Capela és mtsai 2006).
A sokáig a legvalószínűbbnek tűnt dopaminteória ellen szól és a
testhőmérséklet-emelkedés szerepére hívja fel a figyelmet Yuan és munkatársainak
(2002) kísérlete: ha az MDMA kezelés előtt rezerpint (kiüríti a monoaminraktárakat,
véd a neurotoxicitás ellen) vagy α-metil-p-tirozint adtak patkányoknak és a kísérlet alatt
a testhőmérsékletüket magasan tartották, az előkezelések nem védték ki az MDMA
neurotoxikus hatását. Emellett számtalan, a testhőmérsékletet csökkentő vegyületről
bizonyították, hogy kivédi az MDMA neuronkárosító hatását (Sprague és mtsai 1998;
Green és mtsai 2003) és in vitro bizonyították, hogy az MDMA-nak és metabolitjainak
neurotoxikus hatása hőmérsékletfüggő (Capela és mtsai 2006). Több vizsgálat is
kimutatott összefüggést a hipertermiás válasz nagysága és a neurotoxicitás mértéke
között MDMA-val kezelt állatokban (Broening és mtsai 1995; Malberg és Seiden 1998;
Sanchez és mtsai 2004). Vannak azonban olyan, a neurotoxicitást megakadályozó
vegyületek, amelyek nem csökkentik a testhőmérsékletet (Colado és mtsai 1997; Colado
és mtsai 1998; Sanchez és mtsai 2001). Az eddig rendelkezésünkre álló bizonyítékok
alapján valószínűnek tűnik, hogy a testhőmérséklet-emelkedés fokozza a
neurotoxicitást, de nem önmagában okozza azt (Green és mtsai 2004a).
1.5.2 Bizonyítékok a szerotonerg idegsejtek működésének tartós csökkenésére
patkányokban
Az MDMA kezelést követő extracelluláris 5-HT- és 5-HIAA-koncentrációk
tartós csökkenéséről tekintélyes számú cikkben olvashatunk (Battaglia és mtsai 1987;
Commins és mtsai 1987; Schmidt 1987; Stone és mtsai 1987; McKenna és Peroutka
1990; Nash és Yamamoto 1992; Colado és mtsai 1993; O'Shea és mtsai 1998). A
vizsgálatok során különböző patkánymodelleken, különböző dózisokat alkalmazva
számos agyterületen bizonyították a fenti paraméterek csökkenését és megállapítható,
hogy az egyes patkánymodellek eltérő érzékenységűek az MDMA neurotoxikus
hatására (Green és mtsai 2003). A legérzékenyebbek a Dark Agouti patkányok,
25

amelyekben 10-15 mg/kg MDMA legalább 30-50%-kal csökkentette a különböző
agyterületeken a szöveti 5-HT-koncentrációt (Colado és mtsai 1995; O'Shea és mtsai
1998), míg a Sprague-Dawley, a Hooded Lister és a Wistar patkányoknak legalább 20
mg/kg-os dózis többszöri adása volt szükséges ugyanolyan mértékű károsodás
létrejöttéhez (Colado és mtsai 1993; Aguirre és mtsai 1998; Shankaran és Gudelsky
1999). A beadást követően kb. 3-6 óra elteltével mérhető a legalacsonyabb szöveti 5-
HT-koncentráció, 24 óra elteltével a szintek nagyjából megegyeznek a kezelés előtti
értékekkel, majd egy héten belül újra csökkenés tapasztalható és az 5-HT-koncentráció
tartósan alacsony marad az agyszövetben (Stone és mtsai 1986; Schmidt 1987; Battaglia
és mtsai 1988). Hasonlóan alakul az 5-HIAA agyszöveti koncentrációja (Stone és mtsai
1986; Battaglia és mtsai 1988) és tartósan csökken a TPH aktivitása is (Stone és mtsai
1986; Schmidt és Taylor 1987).
A szerotonerg neuronok paroxetin- és citalopramkötő képessége is jelentősen
csökken patkányokban MDMA kezelés hatására (Battaglia és mtsai 1987; Sharkey és
mtsai 1991; Scanzello és mtsai 1993; Hewitt és Green 1994; Colado és mtsai 1995;
O'Shea és mtsai 1998; Ando és mtsai 2006; Ferrington és mtsai 2006). A paroxetin- és
citalopramkötő képesség csökkenésének oka, hogy csökken az ezeket kötő SERT-ek
mennyisége az idegsejtek nyúlványain, mutatva ezzel pusztulásukat (Schmidt 1987).
Ezen paraméterek mérése pontosabb képet ad az MDMA által okozott neurotoxicitás
mértékéről, mint az 5-HT és az 5-HIAA szöveti koncentrációinak mérése (Battaglia és
mtsai 1987; Green és mtsai 2003).
Az idegsejtek axonvégeinek károsodása láthatóvá tehető a SERT-ek
immunhisztokémiával történő kimutatásával (Ricaurte és mtsai 2000). O’Hearn és
munkatársai (1988) patkányokban vizsgálták a szerotonin-tartalmú idegsejteket ezzel a
módszerrel számos agyterületen. Napi 2*20 mg/kg-os MDMA-dózist alkalmaztak 4
egymást követő napon és megállapították, hogy két héttel a kezelést követően az axonok
alakja megváltozott, mennyiségük jelentősen csökkent a neocortexben, a striatumban és
a thalamusban. Az előzőeknél kisebb mértékű csökkenést találtak a hippocampus, a
septum és az amygdala területén (O'Hearn és mtsai 1988).
A prolin anterográd transzportjának kimutatásával is vizsgálható a szerotonerg
neurotoxicitás mértéke (Ricaurte és mtsai 2000; Green és mtsai 2003). A kísérlet során
előzetesen MDMA-val kezelt patkányok raphe magjaiba tríciummal jelölt prolint
26

injekcióznak, és ezzel a felszálló szerotonerg axonokat láthatóvá teszik. Az MDMA
jelentősen csökkenti a prolin anterográd transzportját (Callahan és mtsai 2001).
Úgy tűnik, a szerotonerg neuronok sejttestjei nem károsodnak, csak axonjaik,
mert immunhisztokémiai vizsgálat 2 héttel a kezelés után patkányokban a sejttestek
épségét igazolta (O'Hearn és mtsai 1988) és egy másik vizsgálatban a raphe magvak
kisülési aktivitásának mintázata nem tért el a kontrollokétól 2 héttel nagy dózisú
MDMA kezelést (napi 2*20 mg/kg 4 napig) követően (Gartside és mtsai 1996).
1.5.3 A szerotonin-rendszer csökkent működésének következményei patkányokban
In vivo mikrodialízis tanulmányok segítségével megvizsgálták a szerotonerg
rendszer csökkent működésének következményeit az MDMA neurotoxikus dózisával
előkezelt patkányokban:
• Az extracelluláris 5-HT-szint nem csökkent az előkezelt állatokban a
frontális cortexben illetve a striatumban (Series és mtsai 1994; Shankaran
és Gudelsky 1999), viszont az 5-HIAA-szint csökkent volt a frontális
cortexben 2 héttel a kezelés után (Series és mtsai 1994).
• 2 héttel az előkezelés után a dorzális raphe magvak elektromos ingerlése
kisebb 5-HT-felszabadulást okozott a frontális kéregben, mint nem
előkezelt állatokban (Gartside és mtsai 1996).
• 1 héttel az előkezelés után immobilizációs stressz hatására a
hippocampusban és a frontális kéregben kisebb 5-HT-felszabadulást
regisztráltak, mint nem előkezeltekben (Matuszewich és mtsai 2002).
• 1 héttel az előkezelés után a következő MDMA kezelés kisebb 5-HT-
felszabadulást okozott, mint nem előkezeltekben (Shankaran és Gudelsky
1999).
• Változásokat mutattak ki a fenfluraminnal kiváltott prolaktin- és kortizon-
szekréció mértékében, még egy év elteltével is volt kimutatható változás a
válaszban, annak ellenére, hogy ekkorra a szöveti 5-HT-szintek már
helyreálltak (Poland és mtsai 1997).
MDMA-val előkezelt patkányokban a hőszabályozás zavarát is megfigyelték:
magas hőmérsékleten az állatok testhőmérséklete gyorsabban emelkedett, mint a
27

kontrolloké és lassabban tért vissza a normális értékre, amikor visszahelyezték őket az
általuk megszokott hőmérsékleti viszonyok közé (Dafters és Lynch 1998; Mechan és
mtsai 2001). Ehhez hasonlóan, ha magas külső hőmérsékleten újabb MDMA kezelést
adtak patkányoknak, nagyobb testhőmérséklet-emelkedést figyeltek meg, mint nem
előkezeltekben és a testhőmérsékletük lassabban tért vissza az eredetire (Green és mtsai
2004b). Szobahőmérsékleten adva az újabb MDMA-kezelést ellentmondásos
eredményekhez vezetett: két korábbi tanulmányban kisebb (Shankaran és Gudelsky
1999), illetve nagyobb (Dafters 1995) hipertermiás választ figyeltek meg, míg az
előzőeknél kisebb mértékű idegkárosodást okozó tanulmányok szerint nem volt
különbség a hipertermiás válaszban (Beveridge és mtsai 2004; Green és mtsai 2004b;
Ferrington és mtsai 2006). Mindezek alapján úgy tűnik, az MDMA által okozott
neurotoxicitás a hőleadást károsítja leginkább (Green és mtsai 2004a).
Az MDMA-val kezelt állatok hosszútávú lokomotoros aktivitás-változásának
követésével kevés tanulmány foglalkozik és eredményeik is eléggé ellentmondásosak.
Wallace és munkatársai (2001) 4*10 mg/kg MDMA-val kezelt patkányok nappali és
éjszakai lokomotoros aktivitását monitorozták a kezelés utáni 7. naptól a 14. napig
folyamatosan és csökkentnek találták a kontrollokéhoz képest. Ezzel ellentétben
Marston és munkatársai (1999) 3 napig kezeltek patkányokat az első napon 2*10, a
másodikon 2*15, a harmadikon 2*20 mg/kg MDMA-val és nem találtak különbséget a
lokomotoros aktivitásban a kezelést követő 16 napon kontrollokhoz viszonyítva. A
különböző dózisok különböző mértékű neurotoxicitást okozva befolyásolhatták az
eredményeket.
A cirkadián ritmus pacemakere, a hipotalamuszban elhelyezkedő nucleus
suprachiasmaticus jelentős szerotonerg beidegzést kap a raphe magvakból és a
szerotonerg neuronok a laterális geniculate magon keresztül is befolyásolják a
működését (Meyer-Bernstein és Morin 1996). Ismert, hogy 5-HT agonisták segítségével
befolyásolható a nucleus suprachiasmaticus működése, átállítható a biológiai óra
(Cutrera és mtsai 1996; Morin 1999). Biello és munkatársai megvizsgálták, hogy az
MDMA szerotonerg neuronokat károsító hatása megváltoztatja-e hosszútávon a nucleus
suprachiasmaticusnak a 8-OH-DPAT-re (5-HT1A és 5-HT7 receptor agonista) adott
válaszát patkányokban (Biello és Dafters 2001; Dafters és Biello 2003). Az első
vizsgálatukban kimutatták, hogy a 8-OH-DPAT-tel kiváltott fáziseltolódás csökkent az
28

MDMA-val 6-10 nappal, illetve 20 héttel korábban előkezelt patkányok koronális
hypothalamus szeleteiben in vitro. Az eredmény arra utal, hogy a patkányok nucleus
suprachiasmaticusában az 5-HT1A és/vagy az 5-HT7 receptorok mennyisége és/vagy
eloszlása a neuronkárosodás következtében megváltozott (Biello és Dafters 2001).
Később bizonyították, hogy a nucleus suprachiasmaticus működésének károsodása
akkor is létrejön, ha kivédik az MDMA akut hipertermiás hatását (Dafters és Biello
2003).
Már egy dózis MDMA kezelés hosszútávú szorongás-fokozó hatását is
kimutatták különféle szorongás-tesztekkel („emergence”, „open field”, megemelt
keresztpalló (elevated plus maze), szociális interakciós teszt) 1-3 hónappal a kezelést
követően Sprague-Dawley és Wistar patkányokban (Morley és mtsai 2001; Fone és
mtsai 2002; Gurtman és mtsai 2002; Walker és mtsai 2007). Előfordult, hogy a szöveti
5-HT-koncentrációk mérésével nem tudtak kimutatni idegkárosodást, a szorongás mégis
fokozódott (Fone és mtsai 2002). Érdekes azonban, hogy a Dark Agouti patkányokban,
amelyeknél MDMA kezelés nélkül az „open field” és a megemelt keresztpalló
tesztekkel nagyobb mértékű szorongás mérhető, mint a Wistar és Sprague-Dawley
patkányoknál; az MDMA csökkentette a szorongást (Morley és mtsai 2001; Mechan és
mtsai 2002; Mechan és mtsai 2002). Megmagyarázhatja az ellentmondást, hogy a
szerotonin-rendszer működésének csökkenése maga után vonhatja a szorongás
fokozódását és csökkenését is (Griebel 1995; Green és McGregor 2002).
Meglepő módon számos vizsgálat, ami az MDMA-val kezelt állatok tanulási és
memória károsodását vizsgálta, nem talált rendellenességre utaló jeleket (Slikker és
mtsai 1989; Ricaurte és mtsai 1993; Robinson és mtsai 1993; Seiden és mtsai 1993;
McNamara és mtsai 1995; Byrne és mtsai 2000). Kivételt képez két munkacsoport
(Marston és mtsai 1999; McGregor és mtsai 2003) eredménye, akik az MDMA-val
kezelt állatok rövid távú memóriájának károsodását bizonyították. Emellett Piper és
munkatársai a figyelem károsodását mutatták ki patkányokban, amelyeket fiatalkorban
kezeltek többszöri MDMA dózissal (Piper és Meyer 2004; Piper és mtsai 2005). A sok
negatív eredmény oka lehet, hogy a szerotonerg neurotoxicitást követő kompenzációs
mechanizmusok elégségesek ahhoz, hogy a kialakult szerotonin-hiány ellenére is
fenntartsák a normál fiziológiás működést vagy az állatoknál használt tanulási és
29

memóriatesztek nem elég érzékenyek a létrejött tanulási és memória problémák
kimutatására (Baumann és mtsai 2007).
1.6 Az állatkísérletekben használt és az ecstasyszedők által alkalmazott dózisok
összehasonlítása
Nehéz az MDMA állatkísérletekben kimutatott hatásai alapján egyértelmű
következtetéseket levonni arra vonatkozólag, hogy mit okozhat pontosan az
ecstasyszedőkben. A kisebb emlősöknek gyorsabb a szívritmusuk, a vérkeringésük és
általában gyorsabban eliminálódnak belőlük a szervezetükbe juttatott vegyületek, mint
nagyobb állatokból (Green és mtsai 2003; Baumann és mtsai 2007). Az anatómiai,
élettani és biokémiai ismeretek alapján felállítottak egy képletet, amellyel a különböző
állatfajokban alkalmazott dózisok összehasonlíthatók (White és Seymour 2005).
Ezalapján a Dark Agouti patkányokban neurotoxikusnak talált 10-15 mg/kg-os dózis
egy 70 kg súlyú ember esetében 140-190 mg-os dózisnak felel meg (Green és mtsai
2003). Az ecstasy tabletták MDMA-tartalma igen változó, az átlagos hatóanyagtartalom
az utóbbi évek felmérései szerint 80-100 mg között van. Az ecstasyfogyasztók dózisai
igen különbözőek, van, aki csak negyed vagy fél tablettát fogyaszt egyszerre, vannak
azonban, akik 5-10 tablettát is bevesznek rövid időn belül. A többségük egyszerre 1-2
tablettát fogyaszt, tehát 2 tablettát bevéve már eléri a képlet alapján számított dózist
(Schifano 2004). Klinikai vizsgálatokban azonban kimutatták, hogy ha egyszerre nagy
dózist vagy több kisebb dózist vesznek be a fogyasztók egymás után, a vártnál jóval
magasabb vérszintet érhet el az MDMA, tehát a kinetikája nem lineáris (de la Torre és
mtsai 2000). A szokatlanul magas vérszint és szöveti koncentráció súlyos
következményekkel járhat (Parrott 2002; Schifano 2004).
1.7 A Dark Agouti patkánytörzs, mint az MDMA-t örökletesen lassan metabolizáló
emberek állatmodellje
Az emberi szervezetben az MDMA lebontásának sebességmeghatározó enzime a
nagy genetikai polimorfizmust mutató debrisoquin-4-hidroxiláz (CYP2D6) (Tucker és
mtsai 1994). Az európaiak 5-10%-ában a CYP2D6 enzim kódolásáért felelős gén
30

mutációjának köszönhetően a CYP2D6 enzim működése deficiens, vagyis az MDMA-t
csak lassabban vagy elégtelenül képesek lebontani, ezért az MDMA igen magas
vérkoncentrációt érhet el bennük és különösen nagy veszélyt jelentenek számukra az
MDMA akut hatásai (de la Torre és Farre 2004; de la Torre és mtsai 2005).
A Dark Agouti patkánytörzsben - főként a nőstény, de kisebb mértékben a hím
állatokban is - az MDMA metabolizálásáért elsődlegesen felelős CYP2D1 enzim
működése deficiens, ezért ezzel az állatmodellel jól vizsgálhatók a lassú
metabolizálókban lezajló MDMA-hatások (Colado és mtsai 1995; Malpass és mtsai
1999). A Dark Agouti patkányok érzékenysége jelentősen nagyobb az MDMA toxikus
hatásaira, mint a Sprague Dawley-ké: Dark Agouti patkányokban magas hőmérsékleten
10 mg/kg-os i.p. dózist követően leírtak letalitást, míg ugyanolyan körülmények között
vizsgált Sprague Dawley patkányoknak ez a dózis nem okozta a pusztulását (Malpass és
mtsai 1999).
1.8 Az MDMA (ecstasy) hatásai emberekben
1.8.1 Akut és szubakut hatások
Az ecstasyfogyasztók elmondásuk alapján mintegy 20 perccel a bevétel után
eufóriát, másokhoz való közelséget, nyitottságot, empátiát éreznek, beszédesebbekké
válnak, fokozódik éberségük, energikusabbak, nő az önbizalmuk és nyitottabbak az új
ötletek iránt (Morgan 2000; Parrott 2001; Fürst 2006). Citalopram (szelektíven
szerotonin-visszavételt gátló, SSRI), ketanserin (5-HT2A antagonista) és haloperidol (D2
antagonista) segítségével MDMA-val kezelt önkénteseken megállapították, hogy a
pozitív hangulati hatásokért és az entaktogén jellegért főként az 5-HT-felszabadulás
felel, míg a stimuláns hatások nagyrészt a dopamin-felszabadulásnak köszönhetők
(Liechti és Vollenweider 2001). A pozitív hatások mellett a felhasználók harmada-
negyede tapasztal akut, nemkívánatos mellékhatásokat: pl. egyensúlyzavart, szájzárt,
csökkent étvágyat, koncentrációs zavarokat, látászavarokat, izomfájdalmat,
nyugtalanságot, erős szívdobogást, remegést, lehangoltságot vagy gyengeségérzést
(McCann és mtsai 1996; Burgess és mtsai 2000; Parrott 2002).
31

A fogyasztók túlnyomó többségének megemeli a testhőmérsékletét, ami
különösen veszélyes, ha intenzív testmozgás, tánc és magas külső hőmérséklet is társul
mellé, amelyek jellemzően kapcsolódnak az ecstasy fogyasztásához (Parrott 2001;
Parrott 2002). A túlzott testhőmérséklet-emelkedésnek életveszélyes szövődményei
lehetnek: rhabdomyolysis (az izmokat alkotó fehérjék szétesnek és a vérbe kerülnek),
disszeminált intravaszkuláris koaguláció (a vérlemezkék kicsapódnak az érpályában és
vérrögöket képezve elzárják az ereket, majd vérzések lépnek fel), akut veseelégtelenség,
májelégtelenség, szívmegállás (Henry és mtsai 1992; Kalant 2001; Green és mtsai 2003;
Schifano 2004). A legtöbb fogyasztó verejtékezésről és szomjúságérzésről panaszkodik
ecstasy fogyasztását követően. Sokan emiatt egyszerre túl sok folyadékot fogyasztanak,
aminek következménye hyponatraemia lehet és súlyos esetben a fogyasztó halálát
okozhatja (Henry és mtsai 1992; Parrott 2002). Leírtak halálos neurológiai
szövődményeket is ecstasy-szedést követően: agyburok alatti vérzést, agyinfarktust,
agyi vénás trombózist, amelyek eredhetnek az ecstasy akut hipertóniát, illetve
dehidrációt okozó hatásaiból (McCann és mtsai 1996; Milroy és mtsai 1996). A legtöbb
emberben létrehoz egy enyhe szerotonin-szindrómát és annak jellegzetes tünetei közül
is tapasztalnak néhányat: viselkedéses hiperaktivitás, zavartság, izgatottság, láz,
hiperreflexia, tachycardia, reszketés, izomgörcsök, tremor, szemtekerezgés, szájzár,
fogcsikorgatás (Parrott és Lasky 1998; Parrott 2002). A pozitív tünetekhez gyakran
tolerancia fejlődik ki, míg a nemkívánatos hatások erősödnek a gyakori használattal
(Parrott 2001; Parrott 2002; Fürst 2006).
Az akut hatások a bevételtől számítva általában 6 órán belül lecsengenek és 24-
48 órán belül jelentkeznek a szubakut hatások, amelyek akár egy hétig is tarthatnak
(Morgan 2000; Green és mtsai 2003). Jellemző, hogy ekkor többnyire az akut
hatásokkal ellentétesen kimerültségről, rossz hangulatról, depresszióról, fokozott
ingerlékenységről számolnak be a fogyasztók (Parrott és Lasky 1998; Topp és mtsai
1999; Huxster és mtsai 2006). Ezekhez sok esetben a koncentrálóképesség csökkenése,
fejfájás, álmatlanság, étvágytalanság, illetve izomfájdalom társul (Topp és mtsai 1999;
Huxster és mtsai 2006).
32

1.8.2 Az ecstasy-szedés hosszútávú következményei
Az ecstasy szerotonerg idegsejteket károsító hatását jóval nehezebb emberekben
vizsgálni, mint állatokban. A rendelkezésünkre álló vizsgáló módszerekkel mért
változások viszont összhangban állnak az állatkísérletek eredményeivel (Morgan 2000;
Green és mtsai 2003; McCann és Ricaurte 2007).
Állatokban a szöveti 5-HT- és 5-HIAA-koncentráció tartós csökkenését
bizonyították, mérésükre emberben csak post mortem van lehetőség. Kish és
munkatársai (2000) egy, a halála előtt rendszeresen ecstasy-t szedő férfi striatumában
post mortem a fenti paraméterek 50-80%-os csökkenését mérték.
A cerebrospinális folyadék (CSF) 5-HIAA-koncentrációja élő emberben is
mérhető és mérésének eredményeiből következtetéseket vonhatunk le a szerotonerg
neuronok működésére vonatkozóan (Morgan 2000). Peroutka (Peroutka 1987) nem
talált különbséget a CSF 5-HIAA-koncentrációjában szórakozási droghasználókat, akik
átlagosan 18 ecstasy tablettát fogyasztottak el életük során, kontrollokhoz hasonlítva.
Több vizsgálat viszont az említett paraméter csökkenését mérte ecstasyt
rendszeresebben szedőket (átlagosan mintegy 50-200 ecstasy tablettát fogyasztottak)
más kábítószereket fogyasztókkal összehasonlítva (Ricaurte és mtsai 1990; McCann és
mtsai 1994; Bolla és mtsai 1998; McCann és mtsai 1999).
Mint az állatkísérleteknél is írtam erről, a különböző 5-HT-agonisták kortizol- és
prolaktinszint növelő hatása is jelezheti a szerotonin-rendszer működési
rendellenességeit (Morgan 2000). L-triptofán használatával nem tudtak különbséget
kimutatni ecstasyszedők és más kábítószereket használó kontrollok, illetve kábítószert
egyaltalán nem szedő kontrollok között (Price és mtsai 1989; McCann és mtsai 1994),
D-fenfluramin és m-CPP (meta-klorofenilpiperazin) segítségével viszont több
kutatócsoport is csökkent prolaktin- illetve kortizolszint növekedést írt le (Gerra és
mtsai 1998; McCann és mtsai 1999; Gerra és mtsai 2000; Verkes és mtsai 2001).
Közvetlenebb bizonyítékokkal szolgálnak a modern képalkotó eljárások, a
pozitron emissziós tomográfia (PET)- és az egyfoton-emissziós komputer tomográfia
(SPECT)-tanulmányok eredményei. Ezen eljárások során olyan radioaktív izotóppal
jelölt ligandot juttatnak a szervezetbe, amely a SERT-hez vagy valamelyik 5-HT
receptorhoz nagy szelektivitással kötődik, így mérhető a SERT illetve az adott 5-HT
33

receptor denzitása a különböző agyterületeken (Morgan 2000). A SERT-ek
denzitásának csökkenését mutatták ki PET-val és SPECT-val is ecstasyt rendszeresen
fogyasztókban (McCann és mtsai 1998; Semple és mtsai 1999; Reneman és mtsai 2001;
Buchert és mtsai 2003; Buchert és mtsai 2004; McCann és mtsai 2005). SPECT-val
kimutatták a corticalis 5-HT2A receptorok sűrűségének változását ecstasy-
fogyasztókban: ha néhány hetes absztinenciát követően mérték a receptorsűrűséget, a
corticalis 5-HT2A receptorok csökkenését tapasztalták (Reneman és mtsai 2000), míg, ha
hosszabb absztinenciát követően mérték, az occipitális cortexben ezen receptorok
sűrűségének növekedését figyelték meg (Reneman és mtsai 2000; Reneman és mtsai
2002). A fokozott receptorsűrűség hátterében valószínűleg az 5-HT-depléció
kompenzálására irányuló törekvések állnak (Reneman és mtsai 2002).
Ecstasy-t fogyasztókon végzett vizsgálatok számos esetben számolnak be arról,
hogy a krónikus pszichiátriai megbetegedésekre jellemző tünetek gyakrabban fordulnak
elő az ecstasysokban, mint az azt nem fogyasztóknál. Leírták a pánik rohamok,
alvászavarok, rossz hangulat, szorongásfokozódás, hallucinációk, paranoid képzetek, a
testsúly és a személyiség megváltozása, bulímia, szomatizáció, kényszeresség,
ellenségesség és a flashback (az akut hangulati hatások, hallucinációk újra átélése a
drog fogyasztása nélkül) gyakori előfordulását (Green és mtsai 1995; Morgan 2000;
Schifano 2000). Súlyos pszichiátriai kórképekről is pl. depresszióról, pánikzavarról,
étkezési zavarokról, kognitív zavarokról, impulzuskontroll-zavarokról, szociális
fóbiáról, paranoiáról, deperszonalizációról, pszichotikus dekompenzációról is gyakran
beszámolnak. A vizsgálatok alapján összefüggés van az elfogyasztott ecstasytabletták
száma és a tünetek gyakorisága között (Schifano és mtsai 1998).
A kialakult alvászavarok és cirkadián ritmusbeli változások önmagukban is
tanulási és memóriakárosodáshoz, hangulatzavarokhoz, az impulzivitás és az endokrin
működés megváltozásához vezethetnek az ecstasy-fogyasztókban (Parrott 2001;
McCann és Ricaurte 2007). Kevés, az ecstasyfogyasztók alvásának minőségi és
mennyiségi változásait tanulmányozó vizsgálat történt napjainkig. Allen és munkatársai
(1993) a NREM-alvás 2. fázisának csökkenését és ennek köszönhetően a teljes alvásidő
csökkenését állapították meg olyan szórakozási kábítószer-élvezőkön, akik életük során
legalább 25 alkalommal vettek be ecstasyt, de a vizsgálatot megelőzően már
abbahagyták a szedését. Később ugyanez a kutatócsoport hosszabb alvásidőt mért
34

ecstasyfogyasztókat vizsgálva. Enyhén csökkent ugyan a NREM-alvás 2. fázisának
időtartama, de a csökkenés nem volt szignifikáns, továbbá a 3. és a 4. fázis
szignifikánsan hosszabb volt az ecstasy-fogyasztókban, ez növelte meg a teljes alvásidőt
(McCann és mtsai 2000). Megvizsgálták az alvásstruktúrát m-CPP adását követően is:
az m-CPP növelte a NREM-alvás 4. fázisának hosszát az ecstasy-fogyasztókban,
viszont csökkentette a kontrollokban (Ricaurte és McCann 2001). Nemrégiben ugyanez
a kutatócsoport azt figyelte meg, hogy az ecstasy-fogyasztók NREM-alvásának 2. fázisa
rövidebb, az 1. fázisa viszont hosszabb időtartamú, mint a kontrolloké és a teljes
alvásidő csökkenését trend jelezte. Megvizsgálták az α-metil-para-tirozin hatását is a két
csoporton: a kontrollok alváslatenciáját (az alvás kezdetéig eltelt idő) jobban
megnövelte, mint az ecstasyhasználókét, míg a REM-latenciát (az alvás kezdetétől a
REM-fázis kezdetéig eltelt idő) növelte az ecstasysokon és csökkentette a kontrollokon.
Negatív korrelációt figyeltek meg a 2. fázis időtartama és az elfogyasztott ecstasy
tabletták száma, valamint az ecstasyhasználat hossza között, és a két utóbbi paraméter
pozitívan korrelált az 1. fázis időtartamával (McCann és mtsai 2007). Az
alvástanulmányok eredményei tehát kissé ellentmondásosak, de abban megegyeznek,
hogy az ecstasyhasználat változásokat okoz az alvásstruktúrában és a korrelációk az
ecstasyhasználat hosszával és az elfogyasztott tabletták számával megerősítik az
MDMA lehetséges szerepét a változásokban (McCann és Ricaurte 2007).
Kuypers és munkatársai (2007) 14 ecstasy-fogyasztót vizsgáltak, akiknek egy
része 125 mg MDMA-t, másik része placebo-t kapott és az éjszakai alvásmegvonás alatt
különböző tanulási teszteken vettek részt. Az MDMA csökkentette az álmosság érzését,
viszont a megosztott figyelmet vizsgáló tesztekben azok, akik MDMA-t kaptak
rosszabb eredményeket értek el (Kuypers és mtsai 2007). Más vizsgálatok is a
kontrollokénál gyakoribb memória- és gondolkodásbeli zavarokról írnak
ecstasyfogyasztókban (Morgan 2000; Parrott 2001).
A rendelkezésünkre álló, a szerotonerg rendszer károsodását bizonyító, illetve
arra utaló preklinikai és klinikai bizonyítékok alapján feltételezhetjük, hogy a
psziciátriai és pszichológiai zavarok oka az MDMA szerotonerg neuronkárosító hatása.
Támogatja ezt az is, hogy az alacsony CSF-beli 5-HIAA-koncentrációval rendelkező
személyekről már évtizedekkel ezelőtt bizonyították, hogy impulzívabbak,
agresszívebbek, ellenségeskedőbbek, míg az átmeneti 5-HT-aktivitás csökkenéséről,
35

hogy egészségesekben rossz hangulatot eredményez (Brown és mtsai 1979; Young és
mtsai 1985; Smith és mtsai 1987; Coccaro 1989; Linnoila és mtsai 1993). Egyértelmű
következtetések azonban nem vonhatók le az ecstasy fogyasztása és az említett
pszichiátriai betegségek, pszichiátriai és pszichológiai tünetek megjelenése között, mert
a kábítószerfogyasztók között eleve nagyobb arányban vannak azok, akik valamilyen
pszichiátriai megbetegedéssel vagy arra való hajlammal rendelkeznek (Morgan 2000;
McCann és Ricaurte 2007). Másrészt, a humán vizsgálatoknál sosem zárhatók ki teljes
biztonsággal az ecstasy mellett fogyasztott más illegális kábítószerek és az
alkoholfogyasztás következményei, ezért különösen fontosak az MDMA tartós
pszichiátriai és neuropszichológiai következményeit vizsgáló állatkísérletek eredményei
is (Parrott 2001; McCann és Ricaurte 2007).
36

2 CÉLKITŰZÉSEK
Tekintélyes és szerteágazó szakirodalommal rendelkezünk az MDMA
neurotoxikus hatásaival kapcsolatban, ennek ellenére kevés az irodalomban fellelhető,
az MDMA rövid- és hosszútávú alvás-ébrenléti hatásaival foglalkozó vizsgálat. Nem
rendelkezünk információval arról, hogy miben különböznek az MDMA-val előkezelt
állatokon az újabb MDMA dózis és az SSRI citalopram akut alvás-ébrenléti hatásai
valamint, hogy befolyásolja-e hosszútávon az állatok napszaki ritmusát az MDMA
kezelés.
Az MDMA rövid- és hosszútávú káros hatásai különösen veszélyesek és
kiszámíthatatlanok a lassú metabolizálókban, akiknek állatmodelljei a Dark Agouti
patkányok. Munkám során ezen az állatmodellen tanulmányoztam az MDMA akut,
szubakut és hosszútávú motoros aktivitás- és alváshatásait a kábítószer-élvezők által
fogyasztott dózisnak megfelelő MDMA-dózist alkalmazva. Kísérleteimmel a
következőkre kerestem a választ:
1) Milyen akut változásokat idéz elő az MDMA 15 mg/kg-os dózisa a lassú
metabolizáló Dark Agouti patkányok motoros aktivitásában és alvásparamétereiben?
2) Milyen szubakut és hosszútávú változások figyelhetők meg a kezelés hatására
ezekben a paraméterekben?
3) Miben térnek el az általunk alkalmazott MDMA dózis akut hatásai az azonos dózisú
MDMA-val 3 héttel korábban előkezelt állatokon?
4) Megváltoztatja-e a 3 héttel korábban alkalmazott MDMA előkezelés a 2,5 mg/kg
dózisú citalopram kezelés akut alváshatásait?
37

3 MÓDSZEREK
Az állatkísérleteket a helyi etikai bizottságok engedélyével, a hazai (az 1998.
december 31-én az állatkísérletekről kiadott kormányrendelet) és a nemzetközi (az
Európai Közösség Tanácsának 1986. november 24-én kiadott 86/609/EEC számú
direktívája és a Nemzeti Egészségügyi Intézet által kiadott „A laboratóriumi állatok
tartásának alapelvei” című, 1985-ben átdolgozott, 85-23. számú kiadvány) előírásoknak
megfelelően végeztük.
3.1 Az állatok és tartásuk
A vizsgálatokat hím Dark Agouti (Harlan, Olac Ltd, Egyesült Királyság)
patkányokon végeztük. Az állatok a laborba érkezésükkor 4-5 hetesek voltak, súlyuk
50- 80 g között volt. A szabványos tápot (CRLT AM, Charles River, Magyarország) és
a vizet szabadon fogyaszthatták, almukat a kísérlet menetétől függően 2-3 naponta
cseréltük és legalább 24 óra telt el az alomcsere és a kezelések között. A helyiségek
állandó hőmérsékletét (21±1 oC) és 12 órás világos/sötét periodicitását (a világítás
kezdete: 09:00) biztosítottuk, a relatív páratartalom 40-50 % között volt.
3.2 Motoros aktivitás és alvás vizsgálatok
3.2.1 A kezelések
A kezeléseket 7 napos szoktatási periódus előzte meg, amelynek során az
állatokat minden nap a világos fázis első 5 percében fiziológiás sóoldattal kezeltük
intraperitoneálisan. A szoktatási periódusra azért volt szükség, hogy az állatok
hozzászokhassanak a kezeléseket végző személy átmeneti jelenlétéhez, a kézbevételhez
és a kezelésekhez.
38

3.2.1.1 Kezelések az MDMA akut, szubakut és hosszútávú hatásainak vizsgálatához
Az állatokat (összesen 66-ot) véletlenszerűen 4 csoportra osztottuk (2. táblázat).
A kezelésekhez használt (±)MDMA-t (>99,5%-os tisztaságú, Sanofi-Synthelabo-
Chinoin, Magyarország) fiziológiás sóoldatban oldottuk fel és 15 mg/ml-es töménységű
oldatot készítettünk. A vizsgálat -21. napján (3 héttel az MDMA akut hatásainak
tanulmányozása előtt) az MDMA+Veh (n=12) és az MDMA+MDMA (n=12)
csoportba sorolt állatoknak 15 mg/kg MDMA-t adtunk intraperitoneálisan. A Veh+Veh
(n=21) és a Veh+MDMA (n=21) csoportba tartozó állatokba pedig fiziológiás sóoldatot
(0,9%-os NaCl-oldat) juttatunk 1 ml/kg-os térfogatban, szintén intraperitoneálisan. A
vizsgálat 1. napján (az akut kezelés napja) a Veh+MDMA és az MDMA+MDMA
csoport tagjai 15 mg/kg MDMA-t kaptak, míg a Veh+Veh és az MDMA+Veh csoportba
tartozó állatok fiziológiás sóoldatot kaptak intraperitoneálisan. Az állatokat minden
esetben a világos periódus kezdetének első 5 percében kezeltük.
EEG, EMG és motoros aktivitás felvételeket készítettünk a Veh+Veh és a
Veh+MDMA csoportban lévő állatokról az 1., a 3., az 5., a 14. és a 28. napon, míg az
MDMA+Veh és az MDMA+MDMA csoportban lévőkről az 1. napon (2. táblázat). A
vizsgálat előtti napon (0. nap) minden állatról felvételt készítettünk és ellenőriztük az
EEG- és az EMG-jel minőségét. Azokat az állatokat, amelyeknél a jel nem volt
megfelelő minőségű, kizártuk a vizsgálatból. A vizsgálat alatt is figyelemmel kísértük a
jelminőséget és indokolt esetben kizártuk az állatot a további tanulmányozás alól. Az
állatok a vizsgálat 1. napját követően minden nap (azokon a napokon is, amikor nem
készült felvétel) kaptak egy fiziológiás sóoldat injekciót a világos periódus kezdetének
első 5 percében. Erre azért volt szükség, hogy kivédjük az injekció által okozott
esetleges viselkedésbeli különbségeket, valamint, hogy az egyes felvételeket a
korábbiakhoz tudjuk hasonlítani.
39

2. táblázat: A kezelések menete és a csoportok elnevezése az MDMA
hatásainak vizsgálata során
A csoport neve -21. nap 1. nap 3.nap 5. nap 14. nap 28. nap
Veh + Veh fiz. só fiz. só fiz. só fiz. só fiz. só fiz. só
Veh + MDMA fiz. só MDMA fiz. só fiz. só fiz. só fiz. só
MDMA + Veh MDMA fiz. só
MDMA + MDMA MDMA MDMA
MDMA: 15 mg/kg MDMA i.p., fiz. só: 1 ml/kg 0.9% NaCl (fiziológiás sóoldat) i.p.
3.2.1.2 Kezelések a citalopram akut hatásainak vizsgálatához
Az állatokat (összesen 61-et) véletlenszerűen 4 csoportra osztottuk (3. táblázat).
Három héttel a citalopram akut hatásainak vizsgálata előtt (-21. nap) az MDMA+Veh
(n=15) valamint az MDMA+Citalopram (n=15) csoportokba tartozó állatoknak 15
mg/kg MDMA-t adtunk intraperitoneálisan, míg a Veh+Veh (n=16) és a
Veh+Citalopram (n=15) csoportokba sorolt állatoknak fiziológiás sóoldatot adtunk 1
ml/kg-os térfogatban, intraperitoneálisan. Az akut citalopram kezelés napján (1. nap) az
MDMA+Citalopram és a Veh+Citalopram csoport tagjai 2,5 mg/kg citalopram
(Citalopram hydrobromide, Tocris Bioscience, Biotrend, Köln, Németország) kezelést,
míg az MDMA+Veh és a Veh+Veh csoportokba tartozó állatok fiziológiás sóoldatot
kaptak intraperitoneálisan. A kezeléseket a világos periódus első 5 percében végeztük és
a vizsgálat napján (1. nap) a következő fejezetekben részletezett módon - 24 órás EEG,
EMG és motoros aktivitás felvételeket készítettünk az állatokról. Felvételt készítettünk
a kezelés előtti napon (0. nap) is minden állatról, hogy ellenőrizhessük az EEG- és
EMG-jel megfelelő minőségét. Ha a 0. napon vagy a vizsgálat napján a jel nem volt
megfelelő minőségű, csakúgy, mint az MDMA hatásainak vizsgálata során, kizártuk az
állatot a vizsgálatból.
40

3. táblázat: A kezelések menete és a csoportok elnevezése a citalopram hatásainak
vizsgálata során
A csoport neve -21. nap 1. nap
Veh+Veh fiz. só fiz. só
Veh+Citalopram fiz. só citalopram
MDMA+Veh MDMA fiz. só
MDMA+Citalopram MDMA citalopram
MDMA: 15 mg/kg MDMA i.p., fiz. só: 1 ml/kg 0.9% NaCl (fiziológiás sóoldat) i.p.,
citalopram: 2,5 mg/kg citalopram i.p.
3.2.2 A krónikus EEG és EMG elektródák műtéti beültetése
Az MDMA- illetve fiziológiás sóoldat előkezelések után kb. egy héttel történt az
EEG és EMG regisztrálásához használt krónikus elektródák műtéti beültetése. A
műtéteket altatás (2% halotán oxigénben, Fluotec 3) mellett, sztereotaxikus készülék
(Kopf) segítségével végeztük. Az EEG-hez használt, rozsdamentes acélból készült,
csavar-elektródákat epiduralisan a bal frontális cortex (a bregmához viszonyítva L: 2,0
mm és A: 2,0 mm) valamint a bal parietális cortex (a lambdához viszonyítva L: 2,0 mm
és A: 2,0 mm) felett rögzítettük. A föld elektródát a kisagy felett helyeztük el. Az EMG-
hez használt, rozsdamentes acélból készült, szilikon-gumi bevonatú, spirál-elektródák
(Subcutaneous Electrode Wire, Plastics One Inc., Roanoke, Egyesült Államok, 550 mm
hosszú, 1,2 mm átmérőjű a szilikon-gumival) megtisztított végét két öltéssel a hátsó
nyaki izmokba varrtuk. A rugalmas szilikon-gumi bevonatú spirál-elektródák lehetővé
tették az állatok szabad mozgását. Az EEG és EMG elektródák szabad végeit egy
kisméretű csatlakozóba vezettük, amelyet rögzítő-csavarok és speciális cement
(cranioplastic cement, Plastics One Inc., Roanoke, Egyesült Államok) segítségével az
állat koponyacsontjához rögzítettünk.
A műtétet követően az állatokat egyesével 3500 mm * 3500 mm * 4050 mm-es
üvegketrecekbe helyeztük, majd 8 nap elteltével kábelre csatlakoztattuk (3. ábra) és így
további 7 napot töltöttek el a vizsgálat előtt.
41

3.2.3 Az EEG, EMG és a motoros aktivitás mérése
Az EEG és EMG jeleket elvezető kábelre mágnest rögzítettünk, a kábel a
mágnes szintjén áthaladt egy elektromos tekercsen, majd a ketrec fölött elhelyezett
forgócsatlakozóban végződött. A forgócsatlakozó lehetővé tette az állatok szabad
mozgását, s tevékenységük közben mozgásba hozták a fejükhöz csatlakoztatott kábelt a
rajta lévő mágnessel, ami áramot indukált a tekercsben (Svensson és mtsai 1987; Kantor
és mtsai 2002). Az így kapott jeleket, mint a motoros aktivitás mutatóit az EEG és EMG
jelekkel együtt, megfelelő alsó és felső szűrést követően (0,53 – 30 Hz, 6 dB/ oktáv),
differenciál erősítő segítségével felerősítettük (EEG * 5000, EMG * 20000 és motoros
aktivitás * 5000). Az analóg jeleket csatornánkénti 64 Hz-es mintavételezéssel
digitalizáltuk, a számítógép képernyőjén megjelenítettük és merevlemezén tároltuk a
későbbi feldolgozásokhoz.
A motoros aktivitás megállapításához a felerősített jelsorozatokat 4 másodperces
szakaszokra osztottuk. Az eredeti jelet, ami - az állat mozgásának köszönhetően - a
görbén mind pozitív, mind negatív irányban megjelenhet, átalakítottuk egy csak a
pozitív tartományban megjelenő jellé, az eredeti abszolút értékévé. Emellett
meghatároztunk egy zajszintet, ami alatt a jelet nem tekintettük az állat mozgásának és a
zajszint feletti jelet tartalmazó 4 másodperces szakaszokat összegeztük, s az így kapott
idő adta a motoros aktivitás idejét (Svensson és mtsai 1987; Kantor és mtsai 2002).
42

mágnes
forgó-csatla-kozó
Elektro-mos tekercs
3. ábra: Az állatok elhelyezése a ketrecekben: az EEG-, EMG- és motoros aktivitás
tanulmányozásához a megműtött állatokat egyesével üvegketrecekbe helyeztük és
kábelre csatlakoztattuk. A vizsgálathoz használt elektromos tekercs, mágnes és
forgócsatlakozó helyét a nyilak mutatják.
3.2.4 Az alvásstádiumok meghatározása
A citalopram akut alváshatásait a szakirodalomra támaszkodva az első néhány
órában vártuk, ezért az első 4 óra időtartamait, a minél pontosabb kiértékelés érdekében
vizuálisan elemeztük és a következő szempontok alapján határoztuk meg az egyes
alvásfázisokat:
• Ébrenlét: alacsony amplitúdójú (max. 60-80 µV), gyors frekvenciájú (alfa: 8-13
és béta: 14-30 Hz) EEG, fokozott EMG és motoros aktivitás (aktív ébrenlét,
AW) illetve a motoros aktivitás hiánya (passzív ébrenlét, PW).
• A szendergés és felületes alvás (NREM-1): átmeneti szakasz az ébrenlét és a
mélyalvás között, amelynek EEG-jét a lassú hullámokkal (delta: 0,75-6 Hz) és
43

alvási orsókkal (6-15 Hz) tarkított ébrenlétszerű háttértevékenység, valamint
csökkent izomtónus és motoros aktivitás jellemez.
• A mélyalvás (NREM-2): nagy amplitúdójú, kis frekvenciájú (max 350-400 µV;
delta: 0,5-6 Hz) EEG, amely csökkent izomtónussal és a motoros aktivitás
hiányával párosul.
• A paradox alvás (REM): alacsony, közel egyforma amlitúdójú, gyors théta
hullámok az EEG-n, az izomtónus és a motoros aktivitás teljes hiánya mellett,
amelyet időnként 1-2 másodperces izomrángások szakíthatnak meg (Gottesmann
1992; Filakovszky és mtsai 2001).
Vizuálisan állapítottuk meg, ugyancsak a pontosság érdekében, az MDMA akut,
szubakut és hosszútávú hatásainak vizsgálatánál az alváslatencia hosszát is: az
alváslatencia a világos fázis kezdetétől az alvás kezdetéig eltelt időt jelenti.
A citalopram akut hatásainak vizsgálatánál (az első 4 óra kivételével) és az
MDMA akut, szubakut és hosszútávú hatásainak vizsgálatánál az éberségi szintek
megállapítása alvás-elemző program (Sleepsign for Animal, Kissei Comtec America,
Inc., Egyesült Államok) segítségével történt. A vizuális értékelés kapcsán a fent leírtak
szerint egy algoritmust építettünk fel. Az algoritmus alapján a program 4 másodperces
szakaszokra osztotta az EEG-, EMG- és motoros aktivitás görbéket, majd minden
szakaszra vonatkozóan megállapította az éberségi szinteket. Az MDMA hatásainak
vizsgálata során a motoros aktivitás értékelését különösen fontosnak tartottuk, így azt
külön értékeltük, ezért az ébrenlétet nem osztottuk fel AW-re és PW-re, hanem azokat
összesítve értékeltük a külön megadott motoros aktivitás érték mellett.
Az alvás-elemző programhoz felépített algoritmus pontosságát ellenőriztük
azzal, hogy 8 állat 4*30 perces görbéit (0:00-0:30, 9:00-9:30, 13:30-14:00 és 22:30-
23:00 között) a programmal és vizuálisan is elemeztünk és az eredményeket
összehasonlítottuk. A programmal kapott eredmények szorosan korreláltak a vizuális
vizsgálat eredményeivel.
3.2.5 A cosinor analízis módszer
A ciklikus élettani működéseket (amelyek közé a cirkadián ritmus is tartozik)
matematikailag szinuszoid függvényekkel lehet jellemezni. A különböző időpontokban
44

mért értékekből a periódikus regresszió segítségével állíthatjuk fel a biológiai ritmust
jellemző szinuszoid függvényt. A periódikus regresszió számítására vizsgálatainkban a
cosinor analízis módszerét használtuk. A szinuszoid hullámokat mesor értékükkel
(átlagszintjükkel), amplitúdójukkal (a legnagyobb eltérés az átlagszinttől, a hullám
maximumánál mérhető), és akrofázisukkal (az az időpont, amelynél a hullám eléri a
maximumát) jellemeztük (Nelson és mtsai 1979).
4. ábra: A biológiai ritmust jellemző szinuszoid hullám jellemzése az amplitúdó, a
mesor és az akrofázis segítségével
3.2.6 Az adatok statisztikai elemzése
Az MDMA akut, szubakut és hosszútávú hatásainak vizsgálatához az adatokat
többszempontos variancia analízissel (MANOVA) dolgoztuk fel, a két fő tényező: a
kezelés (nem ismételt mintavételezéses, fiziológiás sóoldat (Veh) és MDMA kezelés,
illetve első MDMA kezelés és második MDMA kezelés) és az idő (ismételt
mintavételezéses, 1-23 óra). A post-hoc összehasonlításhoz a Tukey honest significant
difference (Tukey HSD) tesztet (STATISTICA 6.0, Statsoft, Tulsa, Egyesült Államok)
használtuk. A cosinor analízist (az amplitúdó, akrofázis, mesor értékek és ezek
±konfidecia határainak számítását) az El Temps, 1998 program segítségével végeztük
(Cambras és mtsai 2000).
A citalopram akut alváshatásainak vizsgálatához az adatokat variancia
analízissel (MANOVA) dolgoztuk fel, a három fő tényező: az előkezelés (fiziológiás
45

sóoldat (Veh) vagy MDMA), a kezelés (fiziológiás sóoldat (Veh) vagy citalopram) és az
idő (ismételt mintavételezéses, 1-23 óra) volt. Faktoriális variancia analízist (ANOVA)
végeztünk, a két fő tényező, az előkezelés és a kezelés hatásának vizsgálatára. A post-
hoc összehasonlításhoz a Tukey honest significant difference (Tukey HSD) tesztet
(STATISTICA 6.0, Statsoft, Tulsa, Egyesült Államok) használtuk. A cosinor analízist
(az amplitúdó, akrofázis, mesor értékek és ezek ±konfidecia határainak számítását) a
Time Series Analysis, Serial Cosinor 6.2-Lab View (Expert Soft Technologie, Esvres,
Franciaország) program segítségével végeztük.
3.3 A [3H]-paroxetin kötés mérése autoradiográfiával
A [3H]-paroxetin kötés autoradiográfiás mérésével a különböző agyterületeken a
SERT-hez kötött, radioaktív izotóppal (tríciummal) jelölt ligand (paroxetin)
mennyiségét mutattuk ki. Célunk az volt, hogy igazoljuk az alkalmazott MDMA-dózis
neuronkárosító hatását az occipitális cortexben valamint, hogy megvizsgáljuk,
károsodtak-e a szerotonerg neuronok nyúlványai a substantia nigra pars reticulata, pars
compacta és a striatum területén.
A patkányok egyik csoportját 15 mg/kg MDMA-val, másik csoportját
fiziológiás sóoldattal kezeltük. 21 nappal később mindkét állatcsoportot gyors hatású
intravénás barbiturát injekcióval elaltattuk, majd az agyukat kivettük, lefagyasztottuk és
cryostat segítségével 2 metszetsorozatot készítettünk. A zselatinnal borított
tárgylemezekre felvitt metszeteket -70 oC-on tároltuk az autoradiográfiás feldolgozásig.
A feldolgozás során a metszeteket először kétszer 5 percre puffer-oldatba
helyeztük (előinkubáció), hogy növeljük a receptor-kötés specificitását, majd az egyik
metszetsorozatot tríciummal jelölt paroxetint - telítő, 0,25 nM-os koncentrációban -
tartalmazó puffer-oldattal jégen, 2 órán keresztül inkubáltuk. A nem-specifikusan
kötődött ligandok mennyiségének megállapítása érdekében a másik metszetsorozatot a
0,25 nM paroxetin mellett citalopramot is – 4 µM-os koncentrációban - tartalmazó
puffer-oldattal inkubáltuk. A citalopram leszorítja a paroxetint az 5-HT-felvevő
helyekről, így csak a nem-specifikusan kötődött paroxetin ligandok maradnak kötve ez
utóbbi metszetsorozatban. Az egyes inkubáló edényekből 100 µl-es mintákat vettünk és
10 ml szcintillációs folyadék hozzáadásával, szcintillációs számlálóval ellenőriztük az
46

oldatban a radioaktív ligand koncentrációját. Az inkubáció befejeztével 3-szor 5 percig
puffer-oldatba, majd a puffer sók eltávolítása és az autoradiográfiás műtermékek
kivédése céljából, desztillált vízbe helyeztük a metszeteket. Ezt követte a metszetek
szárítása, melynek során vákum-kapcsolt Pasteur-pipettával távolítottuk el a
vízcseppeket és forró levegővel szárítottuk a metszeteket. Mindezek után kartonpapírra
ragasztottuk és fényvédő kazettába helyeztük őket az ismert koncentrációjú tríciumot
tartalmazó standardokkal és a trícium-érzékeny filmmel együtt. Végül a film gyári
előírásainak megfelelően 6 hétig tároltuk a metszeteket, majd a standardok segítségével
kiszámítottuk a létrejött paroxetin-kötés mennyiségét mindkét metszetsorozatban és a
két metszetsorozat értékeinek különbsége adta a specifikusan kötött paroxetin-kötés
mennyiségét. Az adatok statisztikai elemzését Student’s t-teszt segítségével végeztük.
47

4 EREDMÉNYEK
4.1 Az MDMA akut hatásai: első 24 óra
Az MDMA a kezelés napján erőteljesen fokozta az állatok motoros aktivitását
(5. ábra) és gátolta az alvást (6. ábra). A Veh+MDMA csoportba tartozó állatok motoros
aktivitása szignifikánsan megemelkedett a 2., 3., 4., 5. és 6. órában a Veh+Veh csoport
értékeihez képest. A lokomotoros aktivitás növekedésével párhuzamosan nőtt az
ébrenlét időtartama és csökkent az NREM-1, az NREM-2 és a REM fázisok időtartama
az első 5-11 órában az MDMA-val kezelt (Veh+MDMA csoport) állatoknál. A REM-
alvás gátlása jóval tovább tartott, mint az NREM-1 és az NREM-2 fázisoké. A hatás
lecsengése után, a sötét (aktív) fázis alatt a motoros aktivitás időtartamok csaknem a
fázis teljes ideje alatt szignifikánsan kisebbek ennél az állatcsoportnál, mint az MDMA-
val nem kezelt, Veh+Veh csoportba tartozó állatoknál.
A motoros aktivitás és az alvás-ébrenléti fázisok időtartamainak cosinor
analízisével kapott amplitúdó, akrofázis és mesor értékei is többnyire szignifikáns
különbségeket mutattak a kezelés napján (4. táblázat). Az amplitúdók szignifikánsan
megemelkedtek az MDMA-val kezelt állatoknál (Veh+MDMA csoport) az ébrenlét és
az NREM-2 fázis esetén, ami az óránként mért időtartamok egymáshoz viszonyított
nagyobb különbségeire utal (lásd 3.2.6 fejezet). A mesorok (az óránként mért
időtartamok értékeinek átlagára utal) és az akrofázisok (az az időpont, amelynél a mért
paraméterek cirkadián ritmusát jellemző (periódikus regresszióval illesztett) szinuszoid
hullám eléri a maximumát) szignifikáns eltéréseit figyeltük meg a Veh+MDMA és a
Veh+Veh csoportok értékeit összehasonlítva majdnem minden (kivéve az NREM-1)
paraméter esetén. A hatások megmutatkoznak a két csoport akrofázisértékeinek
különbségeiben is (11. táblázat).
Az alváslatencia 7±3-ról 392±28 min-re emelkedett az MDMA hatására [kezelés
interakció: F(1,20)=187,031, p<0,001] az 1. napon (7. ábra).
48

5. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok motoros aktivitására
és ébrenléti idejére az első 24 órában. Az állatok 3 héttel korábban fiziológiás sóoldat
előkezelést, majd a vizsgálat napján MDMA-t (jobb oldal, Veh+MDMA csoport) vagy
fiziológiás sóoldatot (veh, bal oldal, Veh+Veh csoport) kaptak a világos fázis
kezdetekor. Az ANOVA (2 fő tényező: kezelés, idő) kezelés-idő interakciók (F, df és p
értékek) az ábrákon láthatók és csillag (*) jelöli a Tukey honest post-hoc teszt alapján
megállapított szignifikáns (p<0,05) eltérést a Veh+Veh csoporttól.
49

6. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok felszínes alvás
(NREM-1), mélyalvás (NREM-2) és paradox alvás (REM) fázisokban töltött
idejére az első 24 órában. Az állatok 3 héttel korábban fiziológiás sóoldat előkezelést,
majd a vizsgálat napján MDMA-t (jobb oldal, Veh+MDMA csoport) vagy fiziológiás
sóoldatot (veh, bal oldal, Veh+Veh csoport) kaptak a világos fázis kezdetekor. Az
ANOVA (2 fő tényező: kezelés, idő) kezelés-idő interakciók (F, df és p értékek) az
ábrákon láthatók és csillag (*) jelöli a Tukey honest post-hoc teszt alapján megállapított
szignifikáns (p<0,05) eltérést a Veh+Veh csoporttól.
50

4. táblázat: Az MDMA (15 mg/kg) kezelés akut hatásainak vizsgálata cosinor
analízissel. Az állatok 3 héttel korábban fiziológiás sóoldat előkezelést, majd a
vizsgálat 1. napján MDMA (Veh+MDMA csoport) vagy fiziológiás sóoldat (Veh+Veh
csoport) kezelést kaptak és cosinor analízissel kiszámítottuk az első 24 órában mért
motoros aktivitás, ébrenlét, felszínes alvás (NREM-1), mélyalvás (NREM-2) és paradox
alvás (REM) időtartamok amplitúdó (min/h), akrofázis (h) és mesor (min) értékeit. Az
értékek átlagai és a konfidencia intervallumok (CI) láthatók a táblázatban. Csillag (*)
jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05) eltérést
a Veh+Veh csoporttól.
Motoros aktivitás Ébrenlét Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
1 Átlag CI Átlag CI 1 Átlag CI Átlag CI Ampl. 5,3 3,7-6,9 8,0 5,1-10,9 Ampl. 9,4 7,0-11,9 18,4* 12,8-23,9 Akr. 16,9 15,7-18,1 2,3* 0,9-3,8 Akr. 17,1 16,1-18,1 2,4* 1,2-3,6 Mesor 8,9 8,0-9,8 7,2* 5,6-8,8 Mesor 22,1 20,7-23,5 32,2* 29,0-35,3
NREM-1 NREM-2 Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
1 Átlag CI Átlag CI 1 Átlag CI Átlag CI Ampl. 1,7 0,8-2,5 4,4 2,5-6,4 Ampl. 5,4 3,1-7,7 13,1* 9,1-17,1 Akr. 3,8 1,6-6,0 16,0 14,2-17,9 Akr. 4,7 2,9-6,4 13,4* 12,2-14,5 Mesor 11,1 10,6-11,6 10,2 9,1-11,3 Mesor 21,6 20,3-22,9 15,6* 13,4-17,8
REM Nap Veh+Veh Veh+MDMA
1 Átlag CI Átlag CI Ampl. 2,6 1,9-3,2 2,3 1,8-2,8 Akr. 6,8 5,8-7,8 17,3* 16,4-18,2 Mesor 4,8 4,5-5,2 2,0* 1,7-2,3
51

7. ábra: Az MDMA (15 mg/kg) kezelés hatása az állatok alváslatenciájára. Az
állatok 3 héttel korábban fiziológiás sóoldat előkezelést, majd a vizsgálat 1. napján
MDMA (Veh+MDMA csoport) vagy fiziológiás sóoldat (Veh+Veh csoport) kezelést, s
ezt követően minden nap fiziológiás sóoldatot kaptak a világos fázis kezdetekor. Az
ábrán csillag (*) jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns
(p<0,05) eltérést a Veh+Veh csoporttól.
4.2 Az MDMA hatásai a kezelést követő napokban
Szignifikáns kezelés-idő interakciókat kaptunk az MDMA-val kezelt állatoknak
(Veh+MDMA csoport) a kezelés utáni 3. napon mért motoros aktivitás, ébrenlét (8.
ábra) és NREM-2 (9. ábra) fázisokban töltött időtartamait összehasonlítva a Veh+Veh
csoport értékeivel. Az 5. napon is szignifikánsak voltak a kezelés-idő interakciók a
motoros aktivitás (10. ábra) és a REM (11. ábra) időtartamok esetében a két csoport
eredményeit összehasonlítva. A motoros aktivitás csökkenése a 3. napon a mesor
értékének csökkenésében is megmutatkozott (5. táblázat) a Veh+MDMA csoportnál. Az
5. napon az MDMA kezelésnek köszönhetően ennél az állatcsoportnál az amplitúdók
szignifikánsan lecsökkentek a motoros aktivitás, az ébrenlét és a REM esetén a
Veh+Veh csoport értékeihez képest és a mesor értékek is szignifikánsan különböztek
ennél a 3 paraméternél a 2 csoport eredményeit összehasonlítva (6. táblázat). Ezenkívül
a REM időtartama drámaian lecsökkent ezen a napon a Veh+MDMA csoport állatainál
52

[szignifikáns kezelés hatás: F(1,19)=12,002, p<0,01], míg az ébrenléti idő
szignifikánsan nőtt [szignifikáns kezelés hatás: F(1,19)=12,558, p<0,01].
8. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok motoros aktivitására
és ébrenléti idejére a kezelés utáni 3. napon. Az állatok 3 héttel korábban fiziológiás
sóoldat előkezelést, majd a vizsgálat 1. napján 15 mg/kg MDMA-t (jobb oldal,
Veh+MDMA csoport) vagy fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt
követően minden nap fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az
ANOVA (2 fő tényező: kezelés, idő) kezelés-idő interakciók (F, df és p értékek) az
ábrákon láthatók és csillag (*) jelöli a Tukey honest post-hoc teszt alapján megállapított
szignifikáns (p<0,05) eltérést a Veh+Veh csoporttól.
53

9. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok felszínes alvás
(NREM-1), mélyalvás (NREM-2) és paradox alvás (REM) fázisokban töltött
idejére a kezelés utáni 3. napon. Az állatok 3 héttel korábban fiziológiás sóoldat
előkezelést, majd a vizsgálat 1. napján MDMA-t (jobb oldal, Veh+MDMA csoport)
vagy fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt követően minden nap
fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő tényező:
kezelés, idő) kezelés-idő interakciók (F, df és p értékek) láthatók az ábrákon.
54

5. táblázat: Az MDMA (15 mg/kg) kezelés szubakut hatásainak vizsgálata cosinor
analízissel a kezelés utáni 3. napon. Az állatok 3 héttel korábban fiziológiás sóoldat
előkezelést, majd a vizsgálat 1. napján MDMA (Veh+MDMA csoport) vagy fiziológiás
sóoldat (Veh+Veh csoport) kezelést, s ezt követően minden nap fiziológiás sóoldatot
kaptak és cosinor analízissel kiszámítottuk a kezelés utáni 3. napon mért motoros
aktivitás, ébrenlét, felszínes alvás (NREM-1), mélyalvás (NREM-2) és paradox alvás
(REM) időtartamok amplitúdó (min/h), akrofázis (h) és mesor (min) értékeit. Az
értékek átlagai és a konfidencia intervallumok (CI) láthatók a táblázatban. Csillag (*)
jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05) eltérést
a Veh+Veh csoporttól.
Motoros aktivitás Ébrenlét Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
3 Átlag CI Átlag CI 3 Átlag CI Átlag CI Ampl. 4,6 3,4-5,9 2,5 1,0-4,0 Ampl. 8,9 5,7-12,1 6,1 2,4-9,8 Akr. 16,5 15,5-17,6 19,5 16,9-22,0 Akr. 16,4 14,9-17,9 19,2 16,6-21,6 Mesor 7,4 6,7-8,1 5,9* 5,0-6,7 Mesor 20,5 18,7-22,4 19,7 17,6-21,8
NREM-1 NREM-2 Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
3 Átlag CI Átlag CI 3 Átlag CI Átlag CI Ampl. 1,4 0,4-2,5 0,3 22,4-2,2 Ampl. 5,4 2,8-8,0 4,6 1,4-7,7 Akr. 3,9 0,8-7,1 23,7 20,4-3,0 Akr. 4,0 2,0-6,0 6,7 3,7-9,6 Mesor 12,8 12,2-13,4 11,9 11,4-12,4 Mesor 21,6 20,1-23,1 22,5 20,7-24,3
REM Nap Veh+Veh Veh+MDMA
3 Átlag CI Átlag CI Ampl. 2,5 1,7-3,3 1,7 0,9-2,5 Akr. 6,2 4,9-7,5 9,1 7,2-10,9 Mesor 5,0 4,5-5,4 5,5 5,1-6,0
55

10. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok motoros aktivitására
és ébrenléti idejére a kezelés utáni 5. napon. Az állatok 3 héttel korábban fiziológiás
sóoldat előkezelést, majd a vizsgálat 1. napján MDMA-t (jobb oldal, Veh+MDMA
csoport) vagy fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt követően minden
nap fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő
tényező: kezelés, idő) kezelés-idő interakciók (F, df és p értékek) láthatók az ábrákon.
56

11. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok felszínes alvás
(NREM-1), mélyalvás (NREM-2) és paradox alvás (REM) fázisokban töltött
idejére a kezelés utáni 5. napon. Az állatok 3 héttel korábban fiziológiás sóoldat
előkezelést, majd a vizsgálat napján MDMA-t (jobb oldal, Veh+MDMA csoport) vagy
fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt követően minden nap
fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő tényező:
kezelés, idő) kezelés-idő interakciók (F, df és p értékek) az ábrákon láthatók és csillag
(*) jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05)
eltérést a Veh+Veh csoporttól.
57

6. táblázat: Az MDMA (15 mg/kg) kezelés szubakut hatásainak vizsgálata cosinor
analízissel a kezelés utáni 5. napon. Az állatok 3 héttel korábban fiziológiás sóoldat
előkezelést, majd a vizsgálat 1. napján MDMA (Veh+MDMA csoport) vagy fiziológiás
sóoldat (Veh+Veh csoport) kezelést, s ezt követően minden nap fiziológiás sóoldatot
kaptak és cosinor analízissel kiszámítottuk a kezelés utáni 5. napon mért motoros
aktivitás, ébrenlét, felszínes alvás (NREM-1), mélyalvás (NREM-2) és paradox alvás
(REM) időtartamok amplitúdó (min/h), akrofázis (h) és mesor (min) értékeit. Az
értékek átlagai és a konfidencia intervallumok (CI) láthatók a táblázatban. Csillag (*)
jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05) eltérést
a Veh+Veh csoporttól.
Motoros aktivitás Ébrenlét Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
5 Átlag CI Átlag CI 5 Átlag CI Átlag CI Ampl. 5,5 4,1-7,0 1,9* 0,2-3,5 Ampl. 10,5 7,7-13,4 4,2* 1,1-7,2 Akr. 17,5 16,4-18,6 19,4 15,2-23,3 Akr. 17,0 16,0-18,1 18,2 14,9-21,4 Mesor 8,4 7,6-9,2 6,5* 5,6-7,4 Mesor 23,8 22,2-25,4 31,7* 30,0-33,4
NREM-1 NREM-2 Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
5 Átlag CI Átlag CI 5 Átlag CI Átlag CI Ampl. 2,3 1,1-3,5 0,0 22,6-1,4 Ampl. 5,1 2,9-7,3 3,7 0,8-6,6 Akr. 3,8 1,6-6,1 15,7 6,0-1,3 Akr. 4,6 2,8-6,4 6,0 2,5-9,4 Mesor 11,2 10,5-11,9 7,4 7,1-7,6 Mesor 19,4 18,1-20,6 19,6 18,0-21,3
REM Nap Veh+Veh Veh+MDMA
5 Átlag CI Átlag CI Ampl. 2,4 1,7-3,2 0,5* 0,0-0,9 Akr. 6,9 5,7-8,2 7,1 2,2-11,8 Mesor 4,1 3,7-4,6 1,2* 1,0-1,5
4.3 Az MDMA krónikus hatásai: 14. és 28. nap
Az MDMA-val kezelt állatoknál (Veh+MDMA csoport) a kezelést követő 14.
napon szignifikáns kezelés-idő interakciókat kaptunk a motoros aktivitás (12. ábra) és a
REM (13. ábra) fázis időtartamát összehasonlítva a Veh+Veh csoportba tartozó állatok
értékeivel. A 28. napon is szignifikánsak voltak a kezelés-idő interakciók a motoros
58

aktivitás, az ébrenlét (14. ábra) és az NREM-2 (15. ábra) fázisok esetén a két csoport
értékeit összehasonlítva. A Veh+MDMA csoportnál a paramétereknek – a Veh+Veh
csoporténál - nagyobb ingadozásai láthatók a görbéken. A motoros aktivitás átmenetileg
fokozódott a sötét fázis kezdete előtt 4-5 órával a 14. napon (12. ábra) az MDMA-val
kezelt állatoknál, s ehhez társult az NREM-2 és a REM (13. ábra) időtartamának
csökkenése. A 28. napon az időtartamok értékeinek ingadozásai később, a passzív és az
aktív fázis végén láthatók a motoros aktivitás, az ébrenlét (14. ábra), a NREM-2 és a
REM (15. ábra) fázisok esetén. A motoros aktivitás általános fokozódását a mesor
értékének növekedése (7. táblázat) és szignifikáns kezelés hatás mutatja a kezelés utáni
14. napon a Veh+Veh csoporthoz képest (12. ábra, [F(1,19)=7,065, p<0,05]). Ezen a
napon a post-hoc tesztek a motoros aktivitás szignifikáns növekedését mutatták a sötét
fázis kezdetekor (12 és 13 óránál, 12. ábra), míg a 28. napon hasonló, de szignifikanciát
el nem érő változások láthatók az MDMA-val kezelteknél (14. ábra).
59

12. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok motoros aktivitására
és ébrenléti idejére a kezelés utáni 14. napon. Az állatok 3 héttel korábban fiziológiás
sóoldat előkezelést, majd a vizsgálat 1. napján MDMA-t (jobb oldal, Veh+MDMA
csoport) vagy fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt követően minden
nap fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő
tényező: kezelés, idő) kezelés-idő interakciók (F, df és p értékek) láthatók az ábrákon és
csillag (*) jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns
(p<0,05) eltérést a Veh+Veh csoporttól.
60

13. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok felszínes alvás
(NREM-1), mélyalvás (NREM-2) és paradox alvás (REM) fázisokban töltött
idejére a kezelés utáni 14. napon. Az állatok 3 héttel korábban fiziológiás sóoldat
előkezelést, majd a vizsgálat 1. napján MDMA-t (jobb oldal, Veh+MDMA csoport)
vagy fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt követően minden nap
fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő tényező:
kezelés, idő) kezelés-idő interakciók (F, df és p értékek) láthatók az ábrákon.
61

7. táblázat: Az MDMA (15 mg/kg) kezelés hosszútávú hatásainak vizsgálata
cosinor analízissel a kezelés utáni 14. napon. Az állatok 3 héttel korábban fiziológiás
sóoldat előkezelést, majd a vizsgálat 1. napján MDMA (Veh+MDMA csoport) vagy
fiziológiás sóoldat (Veh+Veh csoport) kezelést, s ezt követően minden nap fiziológiás
sóoldatot kaptak és cosinor analízissel kiszámítottuk a kezelés utáni 14. napon mért
motoros aktivitás, ébrenlét, felszínes alvás (NREM-1), mélyalvás (NREM-2) és paradox
alvás (REM) időtartamok amplitúdó (min/h), akrofázis (h) és mesor (min) értékeit. Az
értékek átlagai és a konfidencia intervallumok (CI) láthatók a táblázatban. Csillag (*)
jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05) eltérést
a Veh+Veh csoporttól.
Motoros aktivitás Ébrenlét Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
14 Átlag CI Átlag CI 14 Átlag CI Átlag CI Ampl. 4,9 3,5-6,3 5,8 2,3-9,4 Ampl. 10,7 6,7-14,8 9,7 4,5-14,8 Akr. 16,9 15,8-18,1 15,7 13,2-18,3 Akr. 18,1 16,6-19,6 16,5 14,3-18,8 Mesor 7,6 6,8-8,4 14,9* 12,9-16,9 Mesor 23,4 21,1-25,7 23,2 20,2-26,1
NREM-1 NREM-2 Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
14 Átlag CI Átlag CI 14 Átlag CI Átlag CI Ampl. 1,9 0,8-2,9 1,3 0,1-2,4 Ampl. 6,2 2,3-10,1 6,8 2,5-11,2 Akr. 4,3 1,9-6,7 22,9 18,5-27,3 Akr. 5,8 3,1-8,4 4,9 2,1-7,6 Mesor 10,0 9,4-10,6 8,9 8,3-9,5 Mesor 22,0 19,8-24,2 22,8 20,3-25,3
REM Nap Veh+Veh Veh+MDMA
14 Átlag CI Átlag CI Ampl. 2,9 2,1-3,7 2,6 1,1-4,1 Akr. 7,3 6,2-8,3 5,3 2,8-7,7 Mesor 5,4 4,9-5,8 4,8 3,9-5,6
62

14. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok motoros aktivitására
és ébrenléti idejére a kezelés utáni 28. napon. Az állatok 3 héttel korábban fiziológiás
sóoldat előkezelést, majd a vizsgálat 1. napján MDMA-t (jobb oldal, Veh+MDMA
csoport) vagy fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt követően minden
nap fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő
tényező: kezelés, idő) kezelés-idő interakciók (F, df és p értékek) az ábrákon láthatók.
63

15. ábra: Az MDMA (15 mg/kg) kezelés hatásai a patkányok felszínes alvás
(NREM-1), mélyalvás (NREM-2) és paradox alvás (REM) fázisokban töltött
idejére a kezelés utáni 28. napon. Az állatok 3 héttel korábban fiziológiás sóoldat
előkezelést, majd a vizsgálat 1. napján MDMA-t (jobb oldal, Veh+MDMA csoport)
vagy fiziológiás sóoldatot (bal oldal, Veh+Veh csoport), s ezt követően minden nap
fiziológiás sóoldatot (veh) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő tényező:
kezelés, idő) kezelés-idő interakciók (F, df és p értékek) láthatók az ábrákon.
64

8. táblázat: Az MDMA (15 mg/kg) kezelés hosszútávú hatásainak vizsgálata
cosinor analízissel a kezelés utáni 28. napon. Az állatok 3 héttel korábban fiziológiás
sóoldat előkezelést, majd a vizsgálat 1. napján MDMA (Veh+MDMA csoport) vagy
fiziológiás sóoldat (Veh+Veh csoport) kezelést, s ezt követően minden nap fiziológiás
sóoldatot kaptak és cosinor analízissel kiszámítottuk a kezelés utáni 28. napon mért
motoros aktivitás, ébrenlét, felszínes alvás (NREM-1), mélyalvás (NREM-2) és paradox
alvás (REM) időtartamok amplitúdó (min/h), akrofázis (h) és mesor (min) értékeit. Az
értékek átlagai és a konfidencia intervallumok (CI) láthatók a táblázatban. Csillag (*)
jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05) eltérést
a Veh+Veh csoporttól.
Motoros aktivitás Ébrenlét Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
28 Átlag CI Átlag CI 28 Átlag CI Átlag CI Ampl. 5,6 4,2-7,0 7,9 4,7-11,1 Ampl. 8,1 4,6-11,6 12,7 7,4-18,0 Akr. 17,1 16,1-18,1 16,6 15,0-18,2 Akr. 16,6 14,9-18,4 16,6 14,9-18,3 Mesor 8,3 7,5-9,1 12,6* 10,8-14,4 Mesor 20,9 18,8-22,9 22,7 19,7-25,7
NREM-1 NREM-2 Nap Veh+Veh Veh+MDMA Nap Veh+Veh Veh+MDMA
28 Átlag CI Átlag CI 28 Átlag CI Átlag CI Ampl. 0,8 23,0-2,6 2,9 1,5-4,3 Ampl. 5,4 2,2-8,7 6,7 2,5-10,9 Akr. 3,0 0,1-5,9 5,1 3,0-7,1 Akr. 4,1 1,6-6,7 4,2 1,5-6,9 Mesor 9,6 9,0-10,2 10,2 9,4-11,0 Mesor 23,6 21,7-25,4 21,4 19,1-23,8
REM Nap Veh+Veh Veh+MDMA
28 Átlag CI Átlag CI Ampl. 2,2 1,0-3,3 2,7 1,4-3,9 Akr. 6,4 4,3-8,6 5,3 3,4-7,1 Mesor 5,7 5,0-6,3 5,0 4,3-5,7
4.4 Az MDMA hatásai a [3H]-paroxetin-kötésre
A [3H]-paroxetin-kötést a 3 héttel korábban fiziológiás sóoldattal illetve
MDMA-val kezelt állatok különböző agyterületein mértük (9. táblázat). Nem találtunk
szignifikáns különbséget a substantia nigra pars compacta (–13%), a pars reticulata
65

(–2%) és a striatum (–15%) területén, az occipitális cortexben viszont jelentős volt az
eltérés (–81%) a két csoport értékei között.
9. táblázat: A [3H]-paroxetin kötés mértékének vizsgálata fiziológiás sóoldattal
(Veh) és MDMA-val (15 mg/kg) 3 héttel korábban kezelt állatokon. Az adatok az
átlagos kötés denzitás értékeket mutatják (fmol/mg szövet)±SEM. Csillag (*) jelöli a
Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05) eltérést a
fiziológiás sóoldattal kezelt csoporttól. SNR: substantia nigra pars reticulata, SNC:
substantia nigra pars compacta.
Agyterület Veh (n=6) MDMA (n=5) Lézió
SNR 229±40 225±20 –2%
SNC 274±33 237±30 –13%
Striatum 57±11 49±6 –15%
Occipitális cortex 77±20 14±7* –81%
4.5 Az MDMA akut hatásai az MDMA-val előkezelt állatokban
Az MDMA-val 3 héttel korábban előkezelt állatok motoros aktivitását és
éberségét fokozta, míg a különböző alvásfázisokban töltött időt csökkentette az újabb
15 mg/kg dózisú MDMA kezelés (MDMA+MDMA csoport, 16. ábra). A mért
paraméterek időtartamait cosinor analízissel elemezve az akrofázisok és a mesorok
szignifikáns változását figyeltük meg majdnem minden (kivéve az NREM-1 fázist, 10.
táblázat) paraméternél az MDMA+Veh csoport értékeihez képest. Az amplitúdók
közötti eltérések nem érték el a szignifikancia szintjét (10. táblázat) a két csoport
értékeit összehasonlítva.
Az MDMA-val előkezelt állatok (MDMA+MDMA csoport) akut MDMA
kezelésre adott válaszát a Veh+MDMA csoportéval összehasonlítva a következőket
figyeltük meg: az akut MDMA kezelésnek rövidebb hatása volt a motoros aktivitásra,
66

az NREM-1-re, az NREM-2-re és a REM-re (5-6., 16-17. ábra) az MDMA-val előkezelt
állatoknál, mint azt a Veh+MDMA csoportnál láthattuk (lásd 4.1 fejezet). Szignifikáns
előkezelés-idő interakciókat figyeltünk meg a motoros aktivitás- [F(22,330)=1,924,
p<0,01] és a REM-eredményeknél [F(22,330)=1,665, p<0,05] a két csoport értékei
között, míg az ébrenléti [F(22,330)=0,675, p=0,863], valamint az NREM-1
[F(22,330)=0,779, p=0,751] és az NREM-2 fázisban töltött idők [F(22,330)=0,818,
p=0,704] esetén az előkezelés-idő interakciók nem voltak szignifikánsak (5-6., 16-17.
ábra). Az időtartamokat cosinor analízissel vizsgálva is megfigyeltünk néhány
különbséget: a REM időtartamok mesor értéke nagyobb az MDMA+MDMA csoportba,
mint a Veh+MDMA csoportba tartozó állatoknál (10. táblázat). Az MDMA kezelés által
okozott akrofázis-változás is jelentősen eltért a két csoport értékei között a NREM-1
fázis időtartama esetén (11. táblázat).
Az akut MDMA kezelés 8±3 min-ről 301±19 min-re emelte az MDMA-val
előkezelt (MDMA+MDMA) állatok átlagos alváslatenciáját (18. ábra) a kezelés napján
[kezelés hatás: F(1,16)=326,761, p<0,001]. Ugyanakkor a 301±19 min alváslatencia
érték a Veh+MDMA csoport átlagos alváslatenciájánál szignifikánsan kisebb
[előkezelés hatás: F(1,15)=4,992, p<0,05].
67

16. ábra: Az MDMA (15 mg/kg) kezelés hatásai az MDMA-val (15 mg/kg) előkezelt
patkányok motoros aktivitására és ébrenléti idejére az első 24 órában. Az állatok 3
héttel korábban MDMA előkezelést, majd a vizsgálat napján MDMA-t (jobb oldal,
MDMA+MDMA csoport) vagy fiziológiás sóoldatot (veh, bal oldal, MDMA+Veh
csoport) kaptak a világos fázis kezdetekor. Az ANOVA (2 fő tényező: kezelés, idő)
kezelés-idő interakciók (F, df és p értékek) az ábrákon láthatók és csillag (*) jelöli a
Tukey honest post-hoc teszt alapján megállapított szignifikáns (p<0,05) eltérést az
MDMA+Veh csoporttól.
68

17. ábra: Az MDMA (15 mg/kg) kezelés hatásai az MDMA-val (15 mg/kg) előkezelt
patkányok felszínes alvás (NREM-1), mélyalvás (NREM-2) és paradox alvás
(REM) fázisokban töltött idejére az első 24 órában. Az állatok 3 héttel korábban
MDMA előkezelést, majd a vizsgálat napján MDMA-t (jobb oldal, MDMA+MDMA
csoport) vagy fiziológiás sóoldatot (veh, bal oldal, MDMA+Veh csoport) kaptak a
világos fázis kezdetekor. Az ANOVA (2 fő tényező: kezelés, idő) kezelés-idő
interakciók (F, df és p értékek) az ábrákon láthatók és a Tukey honest post-hoc teszt
alapján megállapított szignifikáns (p<0,05) eltérést csillag (*) jelöli az MDMA+Veh
csoporttól és kettős kereszt (#) jelöli a Veh+MDMA csoporttól.
69

18. ábra: Az MDMA (15 mg/kg) kezelés hatása az MDMA-val (15 mg/kg) előkezelt
állatok alvás latenciájára. Az állatok 3 héttel korábban fiziológiás sóoldat (Veh+Veh
és Veh+MDMA (Veh+MD) csoport, bal oldal) vagy MDMA (MDMA+Veh (MD+Veh)
és MDMA+MDMA (MD+MD) csoport, jobb oldal) előkezelést majd a vizsgálat napján
MDMA (Veh+MDMA és MDMA+MDMA csoport) vagy fiziológiás sóoldat (Veh+Veh
és MDMA+Veh csoport) kezelést kaptak a világos fázis kezdetekor. Csillag (*) jelöli a
Veh+MDMA csoport értékeinek a Tukey honest post-hoc teszt alapján megállapított
szignifikáns (p<0,05) eltérését a Veh+Veh csoporttól és az MDMA+MDMA csoport
értékeinek eltérését az MDMA+Veh csoporttól. Kettős kereszt (#) jelöli az
MDMA+MDMA csoport értékeinek a Tukey honest post-hoc teszt alapján megállapított
szignifikáns (p<0,05) eltérését a Veh+MDMA csoporttól.
70

10. táblázat: Az MDMA (15 mg/kg) kezelés akut hatásainak vizsgálata cosinor
analízissel az MDMA-val (15 mg/kg) előkezelt állatokon. Az állatok 3 héttel
korábban MDMA előkezelést, majd a vizsgálat napján MDMA (MDMA+MDMA
csoport) vagy fiziológiás sóoldat (MDMA+Veh csoport) kezelést kaptak és cosinor
analízissel kiszámítottuk az első 24 órában mért motoros aktivitás, ébrenlét, felszínes
alvás (NREM-1), mélyalvás (NREM-2) és paradox alvás (REM) időtartamok
amplitúdó (min/h), akrofázis (h) és mesor (min) értékeit. Az értékek átlagai és a
konfidencia intervallumok (CI) láthatók a táblázatban. A Tukey honest post-hoc teszt
alapján megállapított szignifikáns (p<0,05) eltérést csillag (*) jelöli az MDMA+Veh
csoporttól és kettős kereszt (#) jelöli a fiziológiás sóoldattal előkezelt Veh+MDMA
csoporttól.
Motoros aktivitás Ébrenlét Nap MDMA+Veh MDMA+MDMA Nap MDMA+Veh MDMA+MDMA
1 Átlag CI Átlag CI 1 Átlag CI Átlag CI Ampl. 7,6 4,2-11,0 7,3 2,8-11,8 Ampl. 11,3 6,3-16,3 17,8 9,8-25,9 Akr. 16,1 14,3-17,9 0,9* 22,5-3,4 Akr. 16,6 14,8-18,4 1,9* 0,1-3,7 Mesor 16,2 14,2-18,1 8,5* 6,0-11,0 Mesor 21,9 19,0-24,7 28,6* 24,1-33,1
NREM-1 NREM-2 Nap MDMA+Veh MDMA+MDMA Nap MDMA+Veh MDMA+MDMA
1 Átlag CI Átlag CI 1 Átlag CI Átlag CI Ampl. 0,9 22,2-3,6 5,1 2,5-7,7 Ampl. 7,8 3,7-11,9 10,8 4,7-17,0 Akr. 23,7 16,1-7,3 14,1 12,0-16,2 Akr. 4,6 2,4-6,8 13* 10,8-15,3 Mesor 9,7 9,0-10,4 12,6 11,1-14,0 Mesor 22,4 20,1-24,8 15,5* 12,1-18,9
REM Nap MDMA+Veh MDMA+MDMA
1 Átlag CI Átlag CI Ampl. 2,9 1,5-4,3 2,8 1,6-4,1 Akr. 5,71 3,7-7,7 16.8* 15,0-18,7 Mesor 5,5 4,7-6,3 3,1*# 2,4-3,8
71

11. táblázat: A motoros aktivitás, ébrenlét, NREM-1, NREM-2 és REM
időtartamok akrofázis értékeinek különbségei MDMA (15 mg/kg) vagy fiziológiás
sóoldat kezelést követően a kezelés napján (1. nap). Az állatok MDMA
(MDMA+MDMA és MDMA+Veh csoport) vagy fiziológiás sóoldat (Veh+Veh és
Veh+MDMA csoport) előkezelést kaptak 3 héttel az akut kezelés előtt. Csillag (*) jelöli
a szignifikáns (p<0,05) eltérést az MDMA+Veh csoporttól.
Motoros aktivitás Nap Veh+Veh Veh+MDMA Különbség MDMA+Veh MDMA+MDMA Különbség Akrofázis (h) Akrofázis (h)
1 16,9 2,3* 14,6 16,1 0,9* 15,2 Ébrenlét
Nap Veh+Veh Veh+MDMA Különbség MDMA+Veh MDMA+MDMA Különbség Akrofázis (h) Akrofázis (h)
1 17,1 2,4* 14,7 16,6 1,9* 14,7 NREM-1
Nap Veh+Veh Veh+MDMA Különbség MDMA+Veh MDMA+MDMA Különbség Akrofázis (h) Akrofázis (h)
1 3,8 16,0 -12,2 23,7 14,1 9,6 NREM-2
Nap Veh+Veh Veh+MDMA Különbség MDMA+Veh MDMA+MDMA Különbség Akrofázis (h) Akrofázis (h)
1 4,7 13,4* -8,7 4,6 13* -8,4 REM
Nap Veh+Veh Veh+MDMA Különbség MDMA+Veh MDMA+MDMA Különbség Akrofázis (h) Akrofázis (h)
1 6,8 17,3* -10,5 5,7 16,8* -11,1
4.6 A citalopram akut hatásai az alvásra
A citalopram gátolta a REM-alvást az MDMA előkezelést nem kapott állatokban
(Veh+Citalopram csoport, 19-20. ábra): 73%-kal csökkent a REM-alvás időtartama az
első 2 órában (20. ábra) és 55%-kal az ezt követő 2 órában (3. és 4. óra, 21. ábra). A
REM-alvás 24 órás görbéjén is jól látható, hogy a citalopramnak markáns hatása volt a
REM-alvás időtartamokra az első 4 órában (19. ábra). A citalopram emellett jelentősen
fokozta az ébrenlétet: az első 3 órában a Veh+Citalopram csoport PW fázis időtartama
198%-kal nagyobb volt, mint a Veh+Veh csoporté (22. ábra). Az AW fázis időtartama is
72

emelkedett kis mértékben a citaloprammal kezelt állatoknál, de a változás nem érte el a
szignifikancia szintjét (23. ábra). A NREM-1 és a NREM-2 fázisok esetében sem
figyeltünk meg szignifikáns változást a Veh+Veh és a Veh+Citalopram csoport értékeit
összehasonlítva.
Cosinor analízissel a REM időtartamok mesorjának szignifikáns csökkenését
mutattuk ki a citaloprammal kezelt állatoknál (Veh+Citalopram csoport, 12. táblázat), az
amplitúdó és akrofázis értékek azonban nem tértek el szignifikánsan a Veh+Veh csoport
értékeitől. Nem találtunk szignifikáns különbségeket a többi alvásstádium értékeinek
cosinor analízisével sem a két csoport eredményei között.
4.7 A citalopram akut hatásai az alvásra MDMA-val előkezelt állatokban
A citalopram csökkentette a REM-alvás időtartamát az MDMA-val előkezelt
állatoknál is (MDMA+Citalopram csoport, 19-21. ábra). A csökkenés mértéke az első 2
órában pontosan ugyanakkora volt, mint a nem előkezelt állatoknál (73%, 20. ábra). A
következő 2 órában azonban jelentős volt a különbség: míg a Veh+Citalopram csoport
REM időtartamai 55%-kal kisebbek voltak a 3. és a 4. órában a Veh+Veh csoporténál,
addig az MDMA+Citalopram csoport eredményei nem különböztek szignifikánsan az
MDMA+Veh csoportétól (-16%, 21. ábra). A PW időtartamának növekedése sem volt
szignifikáns az előkezelt állatoknál a kezelést követő első 3 órában (8%, 22. ábra), míg a
nem előkezelt állatoknál ugyanezekben az órákban vizsgálva jelentős volt az emelkedés
(198%, 22. ábra). A citalopram kezelés nagyon szoros trendet okozott a REM fázis
kezelés-idő interakciójában (F22,572=1,560, p=0,0504, 19. ábra) a kezelést követő 24 órára
számítva, míg a többi alvásstádium időtartamában nem volt szignifikáns változás az
MDMA+Citalopram és az MDMA+Veh csoport értékei között a 24 órás
összehasonlításban.
Cosinor analízissel elemezve az időtartamokat megfigyeltük, hogy az
MDMA+Citalopram csoport REM időtartamának mesorja szignifikánsan kisebb, mint az
MDMA+Veh csoporté, azonban a különbség jóval kisebb, mint a Veh+Citalopram
csoportot a Veh+Veh csoporthoz hasonlítva látható (12. táblázat). Az akrofázisok és az
amplitúdók között nem volt szignifikáns különbség (12. táblázat) és a többi alvásstádium
73

időtartamainak cosinor analízisével sem figyeltünk meg szignifikáns eltérést az
MDMA+Citalopram és az MDMA+Veh csoport eredményei között.
Az MDMA-val előkezelt és nem előkezelt állatok adatait összehasonlítva a
következőket figyeltük meg: az MDMA+Veh csoport REM-alvás időtartama a világos
fázis első 2 órájában 88%-kal több volt, mint a Veh+Veh csoportba tartozó állatoké (19-
20. ábra, F1,27=5,284, p=0,029), viszont az első 2 óra után már nem volt különbség a két
csoport eredményei között (19. ábra). Ezenkívül a 3. órában az MDMA+Citalopram
csoportba tartozó állatok szignifikánsan több időt töltöttek REM-alvásban, mint a
Veh+Citalopram csoport (19. ábra) és a 3. és 4. óra REM időtartamait elemezve is
szignifikáns különbséget találtunk a két utóbbi csoport értékeit összehasonlítva (21.
ábra). Az AW és a PW adatait elemezve nem találtunk szignifikáns előkezelés hatásokat
(22-23. ábra).
74

19. ábra: A citalopram (2,5 mg/kg) kezelés hatásai a 3 héttel korábban előkezelt
patkányok REM (rapid eye movement, paradox alvás) fázisban töltött idejére az
első 24 órában. Az állatok 3 héttel korábban fiziológiás sóoldat (Veh+Veh és
Veh+Citalopram csoport, felső sor) vagy MDMA (15 mg/kg, MDMA+Veh és
MDMA+Citalopram csoport, alsó sor) előkezelést kaptak, majd a vizsgálat napján
citalopram (cit, Veh+Citalopram és MDMA+Citalopram csoport) vagy fiziológiás
sóoldat (veh, Veh+Veh és MDMA+Veh csoport) kezelést kaptak a világos fázis
kezdetén. Csillag (*) jelöli a Tukey honest post-hoc teszt alapján megállapított
szignifikáns (p<0,05) eltéréseit a Veh+Citalopram csoport értékeinek a Veh+Veh csoport
értékeitől és az MDMA+Citalopram csoport értékeinek az MDMA + Veh csoportétól.
Kettős kereszt (#) jelöli a Tukey honest post-hoc teszt alapján megállapított szignifikáns
(p<0,05) eltéréseit az MDMA+Veh csoport értékeinek a Veh+Veh csoport értékeitől és
az MDMA+Citalopram csoport értékeinek a Veh+Citalopram csoportétól. Az ANOVA
(3 fő tényező: előkezelés, kezelés, idő) kezelés-idő interakció értékek (F, df és p érték) az
ábrán fel vannak tüntetve.
75

20. ábra: A citalopram kezelés (2.5 mg/kg) hatásai a 3 héttel korábban előkezelt
patkányok REM (rapid eye movement, paradox alvás) fázisban töltött idejére a
kezelést követő első 2 órában. Az állatok fiziológiás sóoldat (Veh+Veh és
Veh+Citalopram (Veh+Cit) csoport) vagy MDMA (MDMA+Veh (MD+Veh) és
MDMA+Citalopram (MD+Cit)) előkezelést kaptak 3 héttel a citalopram kezelés előtt.
Csillag (*) jelöli a Veh+Citalopram csoport értékeinek szignifikáns (p<0,05) eltéréseit a
Veh+Veh csoporthoz képest és az MDMA+Citalopram csoport értékeinek szignifikáns
(p<0,05) eltéréseit az MDMA+Veh csoporthoz képest. Kettős kereszt (#) jelöli az
MDMA+Veh csoport értékeinek szignifikáns (p<0,05) eltéréseit a Veh + Veh
csoporthoz képest.
76

21. ábra: A citalopram kezelés (2.5 mg/kg) hatásai a 3 héttel korábban előkezelt
patkányok REM (rapid eye movement, paradox alvás) fázisban töltött idejére a
kezelést követő 3. + 4. órában. Az állatok fiziológiás sóoldat (Veh+Veh és
Veh+Citalopram (Veh+Cit) csoport) vagy MDMA (MDMA+Veh (MD+Veh) és
MDMA+Citalopram (MD+Cit)) előkezelést kaptak 3 héttel a citalopram kezelés előtt.
Csillag (*) jelöli a Veh+Citalopram csoport értékeinek szignifikáns (p<0,05) eltéréseit a
Veh+Veh csoporthoz képest. Kettős kereszt (#) jelöli az MDMA+Citalopram csoport
értékeinek szignifikáns (p<0,05) eltéréseit a Veh+Citalopram csoporthoz képest.
77

22. ábra: A citalopram (2,5 mg/kg) kezelés hatásai az MDMA-val előkezelt
patkányok passzív ébrenléti (PW) fázisban töltött idejére a kezelést követő első 3
órában. Az állatok 3 héttel korábban fiziológiás sóoldat (Veh+Veh és Veh+Citalopram
(Veh+Cit) csoport, bal oldal) vagy MDMA (15 mg/kg, MDMA+Veh (MD+Veh) és
MDMA+Citalopram (MD+Cit) csoport, jobb oldal) előkezelést kaptak, majd a vizsgálat
napján citalopram (Veh+Citalopram és MDMA+Citalopram csoport) vagy fiziológiás
sóoldat (Veh+Veh és MDMA+Veh csoport) kezelést a világos fázis kezdetén. Csillag
(*) jelöli a Veh+Citalopram csoport értékeinek szignifikáns (p<0,05) eltéréseit a
Veh+Veh csoporthoz képest.
78

23. ábra: A citalopram (2,5 mg/kg) kezelés hatásai az MDMA-val előkezelt
patkányok aktív ébrenléti (AW) fázisban töltött idejére a kezelést követő első 3
órában. Az állatok 3 héttel korábban fiziológiás sóoldat (Veh+Veh és Veh+Citalopram
(Veh+Cit) csoport, bal oldal) vagy MDMA (15 mg/kg, MDMA+Veh (MD+Veh) és
MDMA+Citalopram (MD+Cit) csoport, jobb oldal) előkezelést kaptak, majd a vizsgálat
napján citalopram (Veh+Citalopram és MDMA+Citalopram csoport) vagy fiziológiás
sóoldat (Veh+Veh és MDMA+Veh csoport) kezelést a világos fázis kezdetén.
79

12. táblázat: A citalopram (2,5 mg/kg) kezelés akut hatásainak vizsgálata a
paradox alvás (REM) időtartamok cosinor analízisével. Az állatok 3 héttel korábban
MDMA (15 mg/kg, MDMA+Veh és MDMA+Citalopram csoport) vagy fiziológiás
sóoldat (Veh+Veh és Veh+Citalopram csoport) előkezelést, majd a vizsgálat napján
citalopram (Veh+Citalopram és MDMA+Citalopram) vagy fiziológiás sóoldat
(Veh+Veh és MDMA+Veh csoport) kezelést kaptak és cosinor analízissel kiszámítottuk
az első 24 órában mért REM időtartamok amplitúdó (min/h), akrofázis (h) és mesor
(min) értékeit. Az értékek átlagai és a konfidencia intervallumok (CI) láthatók a
táblázatban. Csillag (*) jelöli a Tukey honest post-hoc teszt alapján megállapított
szignifikáns (p<0,05) eltérését a Veh+Citalopram csoport értékeinek a Veh+Veh
csoporttól és az MDMA+Citalopram csoport értékeinek szignifikáns (p<0,05) eltérését
az MDMA+Veh csoporttól.
REM kezelés
előkezelés veh citalopram amplitúdó: 3,5 amplitúdó: 2,1
CI: 2,8 – 4,2 CI: 1,2-3,1 akrofázis: 8,0 akrofázis: 7,4CI: 7,2 – 8,7 CI: 6,0 – 8,8 mesor: 5,3 mesor: 3,2 *
veh
CI: 4,9 – 5,8 CI: 2,7 – 3,7 amplitúdó: 2,6 amplitúdó: 2,1
CI: 2,0 – 3,2 CI: 1,4 – 2,8 akrofázis: 6,3 akrofázis: 5,7CI: 5,3 – 7,3 CI: 4,3 – 7,0 mesor: 4,8 mesor: 3,7 *
MDMA
CI: 4,4 – 5,3 CI: 3,2 – 4,2
80

5 MEGBESZÉLÉS
Az előző fejezetekben tárgyalt kísérleteinkkel arra kerestük a választ, hogy
milyen változásokat okoz az MDMA egyszeri dózisa a DA patkányok szerotonerg
rendszerének működésében. A DA patkányok az MDMA-t lassabban metabolizáló
emberek állatmodelljei. Vizsgálatukkal ezért jobb következtetéseket vonhatunk le arra
vonatkozólag, hogy az ecstasy fogyasztása milyen potenciális veszélyeket rejt magában
az MDMA-t lassabban metabolizáló embercsoportra nézve. Tanulmányoztuk, hogy az
MDMA egyszeri dózisa hogyan befolyásolja a DA patkányok motoros aktivitásának és
alvás-ébrenléti paramétereinek cirkadián ritmusát a kezelést követő 28 napon.
Párhuzamosan kezelt csoportokban a szerotonin transzporterek (3H)-paroxetin-kötését
elemeztük a motoros agyterületeken és az occipitális cortexben, hogy igazoljuk, hogy az
általunk alkalmazott dózis tartósan károsította a szerotonerg neuronok nyúlványait az
állatokban. Megvizsgáltuk, hogy egy újabb MDMA kezelés akut motoros aktivitás és
alvás-ébrenléti hatásai milyen változásokat mutatnak az MDMA-val 3 héttel korábban
előkezelt állatokban. Emellett a citalopram (SSRI) akut alváshatásait is
összehasonlítottuk MDMA-val előkezelt állatokon és nem előkezelteken. Eredményeink
igazolták azt a hipotézist, mely szerint az MDMA által okozott szerotonerg
idegkárosodás változásokat okoz az állatok motoros aktivitásában és alvás-ébrenléti
ciklusában. A továbbiakban részletesen áttekintem az általunk megfigyelt, a szerotonerg
rendszer működésének károsodására utaló változásokat.
5.1 Az MDMA kezelés akut hatásai: első 24 óra
Az MDMA akutan fokozza a monoaminok (legnagyobb mértékben az 5-HT)
felszabadulását számos agyi régióban és akut lokomotoros aktivitás-növekedést okoz
kísérleti állatokban (Spanos és Yamamoto 1989; Callaway és Geyer 1992; Dafters
1994; McNamara és mtsai 1995; Bengel és mtsai 1998). A megfigyelt, dózisfüggő
lokomotoros aktivitás-fokozódásban bizonyították az 5-HT-felszabadulás (Callaway és
mtsai 1990; Rempel és mtsai 1993; Bengel és mtsai 1998; Scearce-Levie és mtsai 1999;
Bankson és Cunningham 2002; Herin és mtsai 2005), a dopamin-felszabadulás (Ball és
mtsai 2003; Bubar és mtsai 2004; Risbrough és mtsai 2006) és a noradrenalin-
81

felszabadulás (Selken és Nichols 2007) szerepét is. Az MDMA kezelést követő
lokomotoros válasz (vizsgálatunkban: Veh+MDMA csoport, 5. ábra) tehát feltehetően
komplex neurokémiai folyamatok, hatások, ellenhatások összegződése (Green és mtsai
2003).
Az MDMA fokozza az acetilkolin felszabadulását is (Fischer és mtsai 2000;
Acquas és mtsai 2001; Nair és Gudelsky 2005; Nair és Gudelsky 2006). A hídban
található pedunculopontinus és laterodorsalis tegmentális magok kolinerg neuronjai
kisülési aktivitásukkal fokozzák az ébrenlétet és különösen aktívak a REM-alvás ideje
alatt. A locus coeruleusban, valamint a peribrachialis magban elhelyezkedő noradrenerg
neuronok és a raphe magvak szerotonerg neuronjai az ébrenlét fenntartásában játszanak
szerepet, emellett gátolják a REM-alvást kiváltó kolinerg neuronokat (Bódizs 2000;
Saper és mtsai 2001; Pace-Schott 2002). Az ébrenléti idők jelentős növekedését
figyeltük meg (Veh+MDMA csoport, 5. ábra), amiben a monoamin-felszabadulás (és
ennek hatására a motoros aktivitás és az ébrenlét fokozása) mellett az acetilkolin-
felszabadulás szerepe is feltételezhető. A REM-alvás időtartamok jelentősen csökkentek
az első 11 órában az MDMA hatására (Veh+MDMA csoport, 6. ábra). Az acetilkolin-
felszabadulás úgy tűnik, ezt nem befolyásolta, ami az első 6-7 órában azzal indokolható,
hogy az állatok nem aludtak el (az ébrenléti idők és az alvás latencia jelentős
növekedése, Veh+MDMA csoport, 7. ábra). A későbbiekben, a 8-11. órában a
monoamin-felszabadulás REM-alvást gátló hatásának túlsúlya mutatkozik meg, mert
ezekben az órákban az MDMA csak a REM-alvást gátolta, a NREM-1-et és a NREM-2-
t nem (Veh+MDMA csoport, 6. ábra). Más, az MDMA akut alváshatásait vizsgáló
preklinikai vagy klinikai vizsgálatot nem közöltek még le napjainkig, a magyarázat
tehát későbbi vizsgálatok tárgya lehet.
10-12 órával az MDMA kezelés után kezdődtek az akut hatásokkal ellentétes,
szubakut hatások (csökkent motoros aktivitás és hosszabb alvásidők, Veh+MDMA
csoport, 5-6. ábra). Az ecstasy-fogyasztók is az akut hatások lecsengését követően,
azokkal ellentétes, szubakut hatásokról: pl. kimerültségről, depresszióról számolnak be
(Parrott és Lasky 1998; Topp és mtsai 1999; Parrott 2002; Green és mtsai 2003; Huxster
és mtsai 2006). A szubakut hatások megjelenésének oka feltehetően a monoamin-
(elsősorban az 5-HT-) -raktárak kiürülése (Parrott 2001; Parrott 2002), habár a
tünetekkel párhuzamosan a klinikai tanulmányok nem vizsgálták a monoamin-szintekre
82

utaló markereket. Az általunk alkalmazott dózis esetén patkányokban az MDMA
kezelést követően különböző agyterületeket érintő, már az első 12 órán belül kialakuló
és hosszantartó szöveti 5-HT-szint csökkenésről számos cikkben olvashatunk, míg ez a
dózis nem okoz tartós dopamin-, noradrenalin- vagy acetilkolin-szint csökkenést az
agyban (Green és mtsai 2003). 10-20 mg/kg MDMA-t követően beszámolnak a DAT-ek
működésének átmeneti, 24 órán belül helyreálló csökkenéséről patkány striatumában
(Metzger és mtsai 1998), a szakirodalom alapján azonban nincs kellő információnk
arról, hogy mikorra állnak helyre 15 mg/kg MDMA adását követően patkányokban a
noradrenalin- és az acetilkolin-szintek. A kísérleteinkben alkalmazottnál jóval nagyobb
dózis esetén a noradrenalin-szint tartós csökkenését is leírták patkányokban
(Mayerhofer és mtsai 2001). Az általunk megfigyelt szubakut tünetek kialakulásában
tehát az 5-HT-szint csökkenésének hatása mellett valószínűsíthetjük a dopamin-szint
csökkenését is az első 24 órában, de nem zárhatjuk ki a noradrenalin- és az acetilkolin-
szintek átmeneti csökkenésének hatását sem.
5.2 Az MDMA hatásai a kezelést követő napokban
A kezelés utáni 3. napon a motoros aktivitás általános csökkenését (mesor
csökkenése, 5. táblázat) tapasztaltuk főként a sötét (aktív) fázis alatt (Veh+MDMA
csoport, 8. ábra). Tímár és munkatársai (2003) ugyancsak a kezelés utáni 3. napon a
spontán lokomotoros aktivitás csökkenését figyelték meg Wistar patkányokban 4*10
mg/kg (+) MDMA kezelést követően. Az általuk alkalmazott dózis hosszú távon
csökkenti az 5-HT-szintet a striatumban (Wallace és mtsai 2001), tehát feltételezhető,
hogy a tartós 5-HT-depléció eredményezte a lokomotoros aktivitás csökkenését (Timar
és mtsai 2003). Vizsgálatunkban mi jóval kisebb dózist és (±)MDMA-t (a (+) izomér
erőteljesebb monaminfelszabadító hatással rendelkezik, mint a racém) adtunk az
állatoknak és a lokomotoros aktivitás csökkenését az állatok régóta megszokott
környezetében tapasztaltuk (míg az említett tanulmányban azt új környezetben figyelték
meg). 10-15 mg/kg (±)MDMA is csökkenti az 5-HT-szintet, az 5-HIAA-szintet és a
[3H]-paroxetin-kötés mértékét az MDMA-t a Wistar-nál lassabban metabolizáló DA
patkányokban számos agyterületen (Colado és mtsai 1995; O'Shea és mtsai 1998). Az
MDMA által felszabadított többi neurotranszmitter szintje (az általunk alkalmazott
83

dózis esetén) nem csökken tartósan (Green és mtsai 2003), tehát a motoros aktivitás
csökkenésének a 3. napon elsősorban az 5-HT tartós depléciója lehet az oka.
Még az 5. napon is tart a motoros aktivitás csökkenése, ugyanakkor
szignifikánsan nőtt az ébrenlét (a passzív ébrenlét növekedése miatt) és csökkent a
REM-alvás időtartama (Veh+MDMA csoport, 10-11. ábra). Ismert, hogy egy dózis
MDMA (10 mg/kg) patkányokban 2 fázisban szabadítja fel az 5-HT-t, az első, akut 5-
HT-felszabadítást követi egy második, kisebb mértékű 5-HT-felszabadítás kb. egy héttel
a kezelés után (Schmidt 1987). Feltételezzük, hogy az 5. napon megfigyelt változások
mögött ez a második, az elsőnél kisebb mértékű 5-HT-felszabadulás, az 5-HT
„kiszivárgása” a károsodott axonokból, állhat. Ugyanezen a napon az MDMA-val kezelt
állatok (Veh+MDMA csoport) cirkadián ritmusa jelentősen eltért a kontrollokétól
(Veh+Veh csoport): a motoros aktivitás, az ébrenlét és a REM-alvás időtartamok
amplitúdói csökkentek (6. táblázat), az időtartamok minimum-maximum különbségei
kisebbek (10-11. ábra). Az 1. napon ennek az ellenkezőjét tapasztaltuk (nagy minimum-
maximum különbségek, amplitúdók növekedése: ébrenlét, NREM-2, 5-6. ábra, 4.
táblázat). Felmerül a kérdés, hogy mi lehet az ellentétes irányú cirkadián ritmusbeli
változás oka, amely még a kismértékű 5-HT-felszabadulás ellenére is jelentkezett.
Feltételezéseinket az MDMA krónikus hatásainak megbeszélésén belül tárgyalom.
5.3 Az MDMA krónikus hatásai
Az MDMA-val kezelt állatoknál a motoros aktivitás időtartamokat elemezve
szignifikáns kezelés-hatást és a mesor értékének növekedését mutattuk ki a kezelés
utáni 14. és 28. napon, amelyek feltehetően a fázisváltás környékén megfigyelhető
nagyarányú motoros aktivitás-fokozódás eredményei (4.3 fejezet, Veh+MDMA csoport,
7-8. táblázat). Wallace és munkatársai (2001) 4*10 mg/kg MDMA-val kezelt patkányok
motoros aktivitásának csökkenését figyelték meg a kezelés utáni 7. nap és 14. nap
között. Marston és munkatársai (1999), akik 3 napig kezeltek patkányokat az első napon
2*10, a másodikon 2*15, a harmadikon 2*20 mg/kg MDMA-val, nem találtak
különbséget a lokomotoros aktivitásban a kezelést követő 16 napon kontrollokhoz
viszonyítva. Az egymástól eltérő eredmények a különböző dózisokból adódóan a
különböző mértékű neurotoxicitás következményei lehetnek. Vizsgálatunkban a
84

fázisváltás környékén megfigyelt nagyarányú motoros aktivitás-fokozódás, az
időtartamok értékeinek a kontrollokénál (Veh+Veh csoport) nagyobb ingadozásai
(Veh+MDMA csoport, főként a sötét fázis kezdete körül, 12-15. ábra) a motoros
aktivitás cirkadián ritmusbeli zavarára utalnak. Az MDMA kezelés hosszútávon
megváltoztatja a cirkadián ritmus pacemakerének, a nucleus suprachiasmaticusnak a
működését patkányokban és hörcsögökben is (Biello és Dafters 2001; Colbron és mtsai
2002; Dafters és Biello 2003; Gardani és mtsai 2005). MDMA-val 6-10 nappal, illetve
20 héttel korábban előkezelt patkányok koronális hypothalamus szeleteiben in vitro
kimutatták, hogy a 8-OH-DPAT-tel (5-HT1A/7 agonista) kiváltott fáziseltolódás csökkent
a kontrollokéhoz képest (Biello és Dafters 2001). Hörcsögöket vizsgálva azt
tapasztalták, hogy a 8-OH-DPAT-kezelés és a 15 percig alkalmazott fény a
kontrollokétól kisebb fáziseltolódást okozott az MDMA-val kezelt hörcsögök cirkadián
ritmusában 3-5 héttel a kezelés után (Colbron és mtsai 2002). Az MDMA tehát
megváltoztatta a cirkadián ritmus pacemakerének működését, amelyet a fény stimulusra
és a 8-OH-DPAT-re adott, a kontrollokétól különböző válaszok jeleznek (Colbron és
mtsai 2002). Az eredmények arra utalnak, hogy a patkányok nucleus
suprachiasmaticusában az 5-HT1A és/vagy az 5-HT7 receptorok mennyisége és/vagy
eloszlása a neuronkárosodás következtében megváltozott (Biello és Dafters 2001;
Colbron és mtsai 2002). Megvizsgálták MDMA-val 2 héttel korábban előkezelt
hörcsögök motoros aktivitásának cirkadián ritmusbeli változását triazolam
(benzodiazepin) hatására is és tanulmányozták, hogy ha a fény-sötét ciklust
mesterségesen eltolják, mennyi idő alatt alkalmazkodnak az új ciklushoz.
Ugyanezekben az állatokban megvizsgálták a szerotonerg neuronok nyúlványainak
denzitását is immunhisztokémiával. A triazolam a kontrollokénál kisebb fáziseltolódást
okozott az MDMA-val kezelt állatokban, míg az új fény-sötét ciklushoz lassabban
alkalmazkodtak ezek az állatok. A kezelt állatok nucleus suprachiasmaticusában
csökkent a szerotonerg neuronok denzitása, amiből arra következtettek, hogy a
neuronkárosodás okozta a kontrollokétól eltérő válaszokat (Gardani és mtsai 2005).
Vizsgálatunkban a [3H]-paroxetin-kötés mértékének csökkenését mutattuk ki az
occipitális cortexben 3 héttel korábban MDMA-val előkezelt állatokban (9. táblázat),
ami az általunk alkalmazott dózis neurotoxikus hatását mutatja. A dolgozatomban
bemutatott kísérletsorozatokban nem vizsgáltuk a szerotonerg neuronok károsodására
85

utaló markereket a nucleus suprachiasmaticusban. Munkacsoportunk egy későbbi
vizsgálatában viszont kimutatta az MDMA, az itt leírtakkal azonos dózisának (15
mg/kg) neuronkárosító hatását DA patkányok nucleus suprachiasmaticusában: a
szerotonerg neuronok nyúlványainak a sűrűsége csökkent 7 és 21 nappal a kezelést
követően (Kirilly és mtsai megjelenés alatt). Az általunk tapasztalt cirkadián ritmusbeli
változások (az 5. napon több vizsgált paraméter cirkadián ritmusának amplitúdója
csökkent és a paraméterek ingadozásai láthatók még a 14. és a 28. napon is, 6. táblázat,
12-15. ábra) mögött tehát valószínűsíthető, hogy a szerotonerg neuronok nyúlványainak
károsodása áll.
Az 5-HT gátolja a REM-alvást kiváltó kolinerg, ún. REM-on neuronokat (Monti
és Monti 2000), tehát nem kizárt, hogy a neuronkárosodás következtében csökkent 5-
HT-neurotranszmisszió csökkenése okozta a REM-alvás növekedését a világos
(nyugalmi) fázis első óráiban 3 héttel a kezelés után (19-21. ábra). Genetikailag
módosított SERT-hiányos egerekben ugyancsak a REM-alvás időtartamának
növekedését figyelték meg (Alexésre és mtsai 2006).
Eredményeink bizonyítják a cirkadián- és az alvásparaméterek kóros eltéréseit
14, 21 és 28 nappal a kezelés után kísérleti állatokban. Ecstasy-fogyasztókban több, az
éjszakai alvást tanulmányozó EEG-vizsgálat is alvásstruktúra-változásokról, illetve m-
CPP (5-HT agonista) hatására különböző alvásparaméter-változásokról számol be
(Allen és mtsai 1993; McCann és mtsai 2000; Ricaurte és McCann 2001; McCann és
mtsai 2007). A humán vizsgálatoknál mindig felmerül, hogy a változásokat nem
okozhatták-e az ecstasy mellett fogyasztott más kábítószerek, illetve nem, már az
ecstasy-fogyasztás előtt is meglévő különbségekről van-e szó. Az ecstasy, illetve az
MDMA szerepére utal azonban, hogy az említett vizsgálatokban megfigyelt
alvásstruktúra-változások mértéke korrelált az ecstasyhasználat hosszával és az
elfogyasztott tabletták számával, illetve, hogy ugyanezekben az emberekben m-CPP-vel
a szerotonerg rendszer működésbeli változása is kimutatható volt (McCann és mtsai
2007). A szerotonerg rendszer károsodása tehát úgy tűnik, emberekben is
alvásstruktúra-változásokhoz vezet (McCann és Ricaurte 2007).
86

5.4 Az MDMA akut hatásai az MDMA-val előkezelt állatokban
Az MDMA előkezelést követő akut MDMA kezelés hatásainak vizsgálata
különösen jelentős, hiszen az előkezelt állatokon tapasztalt akut hatáskülönbségek
mutatják a szerotonerg rendszer tartós károsodásának funkcionális következményeit.
Vizsgálatunkban az MDMA kisebb változásokat okozott az MDMA-val előkezelt
állatok (MDMA+MDMA csoport) motoros aktivitás- és alvásparamétereiben, mint a
nem előkezelteknél (Veh+MDMA csoport) láthattuk: a motoros aktivitás és az ébrenlét
fokozása, valamint a NREM-1, a NREM-2 és a REM időtartamok csökkenése rövidebb
ideig tartott (16-17. ábra. 10. táblázat) és az amplitúdók nem változtak meg (4. táblázat).
Az alvás latencia kisebb (18. ábra) és a REM mesor értéke nagyobb (4., 10. táblázat) az
MDMA+MDMA csoport állatainál, mint a Veh+MDMA csoportnál. A REM mesor
értékeinek különbsége adódhat abból, hogy az előkezelt állatokon az MDMA rövidebb
ideig gátolta a REM-alvást. Másrészt nem kizárt, hogy ehhez az előkezelt állatokban az
MDMA krónikus, a REM-alvás időtartamát növelő hatása is hozzájárult (5.3 fejezet,
19-21. ábra).
Más vizsgálatok is alátámasztják az MDMA akut hatásainak csökkenését
MDMA-val előkezelt állatokon. Egy héttel nagy dózisú MDMA előkezelés (4*10
mg/kg) után a következő MDMA (7,5 mg/kg) kezelés kisebb 5-HT-felszabadulást
okozott az előkezelt állatokban, mint nem előkezeltekben, míg a dopamin-
felszabadulásban nem volt különbség. Ugyanezekben az állatokban kisebb
testhőmérséklet-változást és kisebb mértékű szerotonin szindrómát váltott ki az újabb
MDMA kezelés és csökkent volt az 5-HT-szint a striatumban (Shankaran és Gudelsky
1999). Mi jóval kisebb dózist alkalmaztunk az előkezelés során és talán ez az ok, hogy a
párhuzamosan kezelt állatokban nem volt különbség a striatumban a [3H]-paroxetin-
kötés mértékében (9. táblázat). Az akut motoros aktivitás és alvás-ébrenléti hatások
azonban csökkentek, ami a szerotonerg rendszer működésének károsodására utal.
Poland és munkatársai (1997) is arról számoltak be, hogy bár az 5-HT-szintekben már
nem volt kimutatható változás, a szerotonerg rendszer működészavara kimutatható volt
(a fenfluraminnal kiváltott prolaktin- és kortizonszekréciós válasz csökkent) még egy
évvel az MDMA kezelés után is.
87

Az előbb leírtakkal ellentétben megfigyeltek hatásfokozódást is az újabb
MDMA kezelés hatására. Ha magas külső hőmérsékleten újabb MDMA kezelést adtak
patkányoknak, nagyobb testhőmérséklet-emelkedést figyeltek meg, mint nem
előkezeltekben és a testhőmérsékletük lassabban tért vissza az eredetire (Green és mtsai
2004b). Dafters (1995) szobahőmérsékleten is nagyobb hipertermiás választ figyelt
meg. Emellett több tanulmány szerint nő az akut MDMA kezelést követő lokomotoros
aktivitás-fokozódás az MDMA többszöri dózisával előkezelt patkányokban (Spanos és
Yamamoto 1989; Kalivas és mtsai 1998; Modi és mtsai 2006; Aberg és mtsai 2007). A
magyarázat talán abban rejlik, hogy az MDMA által kiváltott akut hipertermia és
hiperaktivitás mögött több monoaminerg rendszer interakciói állnak, ezért a szerotonerg
rendszer hosszútávú és különböző mértékű károsodása különbözőképpen
befolyásolhatja ezeket.
5.5 A citalopram akut hatásai az alvásra
A citalopram kezelést követően a REM-alvás időtartamának csökkenését
tapasztaltuk (19-21. ábra). A hatás jó néhány órán keresztül fennállt, mert a REM 24
órára számított mesor értéke (ami az óránként mért értékek átlagának csökkenésére utal)
is szignifikánsan csökkent volt. Más kutatócsoportok is beszámoltak a citalopram REM-
alvást gátló hatásáról, melynek oka feltehetően az, hogy nő a szinaptikus 5-HT-
koncentráció a szerotonin-visszavételének gátlása következtében (Neckelmann és mtsai
1996; Monaca és mtsai 2003; Wilson és mtsai 2004; Ivarsson és mtsai 2005;
Bonaventure és mtsai 2007).
Vizsgálatunkban a citalopram kezelés fokozta a passzív ébrenlétet (22. ábra). A
szakirodalomban néhány klinikai vizsgálat ugyancsak az ébren töltött idő növekedéséről
és a teljes alvási idő csökkenéséről számol be SSRI-kezelés hatására (Sharpley és mtsai
1996; Feige és mtsai 2002). Az ok feltehetően – csakúgy, mint a REM-alvás gátlásánál -
a szerotonin-visszavétel gátlásának a következménye: a szinaptikus 5-HT-koncentráció
növekedése. Valószínűleg a szinaptikus 5-HT-szint gyors növekedésének köszönhető,
hogy a REM-alvást gátló hatás és az ébrenlét fokozása azonnal létrejön az SSRI beadása
után, míg a hangulati hatások – amelyeket feltehetően csak az 5-HT receptorok
88

lassabban kialakuló adaptációja hoz létre - csak jóval később jelentkeznek (Wilson és
mtsai 2004).
5.6 A citalopram akut hatásai az alvásra MDMA-val előkezelt állatokban
Bonaventure és munkatársai (2007) azt tapasztalták, hogy az 5-HT7 agonista SB-
269970 fokozta a citalopram hatására létrejött extracelluláris 5-HT-koncentráció-
növekedést és ezzel együtt a REM-alvást gátló hatást. Vizsgálatunkban a citalopram
rövidebb időre változatta meg a REM-alvás időtartamát az MDMA-val előkezelt
állatokon, mint a nem előkezelteken (19-21. ábra) és nem fokozta a passzív ébrenlétet
az előkezelt állatokon (22. ábra). Feltételezhetjük, hogy mindez annak köszönhető, hogy
az axonkárosodás következtében a citalopram kevésbé növelte meg a szinaptikus 5-HT-
koncentrációt az MDMA-val előkezelt állatokban.
Dolgozatomban az MDMA tartós, szerotonerg idegkárosító hatásának
funkcionális következményeit vizsgáltam az MDMA-t örökletesen lassabban
metabolizáló DA patkányokban. Eredményeink felhívják a figyelmet arra, hogy
akár egyetlen, viszonylag kis dózis MDMA is maradandó, hosszan fenálló
idegrendszeri károsodásokat okozhat az arra érzékenyekben. Különösen fontos
tehát küzdeni az ecstasy széleskörű fogyasztása ellen és törekedni arra, hogy a
káros hatásairól rendelkezésünkre álló ismeretek eljussanak azok körébe is, akik a
drogfogyasztás által veszélyeztetettek.
89

6 KÖVETKEZTETÉSEK
Az MDMA motoros aktivitás és alvás-ébrenléti hatásaira, valamint
neurotoxicitásának farmakológiai következményeire irányuló állatkísérletes
vizsgálataim új eredményei az alábbiakban foglalhatók össze:
• Az MDMA a kezelés napján lényegesen hosszabb ideig gátolta a REM-alvást a
patkányokban, mint a NREM-1-et és a NREM-2-t.
• Az akut hatásokat 10-12 órával a kezelés után ellentétes hatások követték az
MDMA-val kezelt állatokon.
• A kezelés utáni 3. és 5. napon a paraméterek cirkadián ritmusának zavarát
figyeltük meg az MDMA-val kezelt állatcsoporton.
• A 14. napra a cirkadián ritmus nagyjából normalizálódik, de a kontrollokéhoz
képest nagyobb ingadozások láthatók mind 14., mind a 28. napon az
időtartamok értékeiben.
• Az MDMA növelte a REM fázisban töltött idő hosszát az alvás első óráiban 3
héttel a kezelés után.
• Az MDMA akut hatásai rövidebb ideig tartottak a droggal előkezelt állatokban,
mint a nem előkezeltekben.
• A citalopram rövidebb ideig gátolta a REM-alvást és nem fokozta a passzív
ébrenlétet az MDMA-val előkezelt állatokban.
Az új eredményekből levonható következtetések:
• Megállapítható, hogy egyetlen dózis MDMA hosszútávon befolyásolta - az
azt lassan metabolizáló - DA patkányok motoros aktivitását és
alvásparamétereit.
• Egyetlen dózis MDMA tartós, az újabb MDMA kezelés és a citalopram
akut hatásainak csökkenésével kimutatható változásokat eredményezett a
DA patkányok szerotonerg rendszerének működésében.
• Mindezek felhívják a figyelmet a humán ecstasy-fogyasztás potenciális
veszélyeire.
90

7 ÖSSZEFOGLALÁS
A rekreációs drogként ismert ecstasy (3,4-metilén-dioxi-metamfetamin,
MDMA) népszerűsége ma is töretlenül növekszik a fiatalok körében az egész világon.
Az MDMA akut neurokémiai és tartós, szerotonerg neuronokat károsító hatásait jól
ismerjük, azonban nincsenek kellőképpen felderítve rövid- és hosszútávú alvás-ébrenléti
hatásai. Nem rendelkezünk információval arról sem, hogy miben különböznek az
MDMA-val előkezelt állatokon az újabb MDMA dózis és a szelektíven szerotonin-
visszavételt gátló citalopram akut alvás-ébrenléti hatásai.
Kísérleteinkkel célunk az volt, hogy megállapítsuk, milyen változásokat okoz
egyetlen dózis MDMA a szerotonerg rendszer működésében. Mindezt az MDMA-t
örökletesen lassabban metabolizáló, ezért klinikai szövődményeire különösen érzékeny
emberpopuláció állatmodelljén, a Dark Agouti patkányokon vizsgáltuk.
Tanulmányoztuk az MDMA hatásait a motoros aktivitásra és az alvás-ébrenléti ciklusra
a kezelést követő 4 héten keresztül. Megvizsgáltuk az újabb MDMA dózis akut motoros
aktivitás- és alvás-ébrenléti hatásait 3 héttel korábban előkezelt patkányokon. Emellett
összehasonlítottuk a citalopram akut alváshatásait MDMA-val előkezelt és nem
előkezelt állatokon. Párhuzamosan kezelt csoportokban elemeztük a szerotonin
transzporterek (3H)-paroxetin-kötését a motoros agyterületeken és az occipitális
cortexben, hogy tanulmányozzuk az axonkárosodás mértékét 3 héttel az MDMA kezelés
után.
Az MDMA akutan fokozta a motoros aktivitást és az ébrenlétet, s hosszabb ideig
gátolta a REM-alvást (paradox alvást), mint a lassú hullámú alvást. 10-12 óra múlva
ellentétes, szubakut hatások (csökkent motoros aktivitás, hosszabb alvásidők) követték
az akut hatásokat. A kezelést követő 3. napon a motoros aktivitás csökkent, főként a
sötét (aktív) fázis ideje alatt, míg az 5. napon az ébrenlét időtartamának növekedését és
a REM-alvás idejének csökkenését figyeltük meg. 14 és 28 nappal az MDMA kezelés
után is a motoros aktivitás és az alvásparaméterek cirkadián ritmusának zavarát
tapasztaltuk. 3 héttel a kezelés után a REM-alvás időtartamának növekedése látható az
alvás első óráiban. A szerotonin transzporterek (3H)-paroxetin-kötésének csökkenése az
occipitális cortexben az axonkárosodást bizonyítja, ugyanakkor a motoros
agyterületeken nincs jelentős változás. Az előkezelt állatokon az MDMA motoros
91

aktivitás- és alvás-ébrenléti hatásai rövidebb ideig tartottak. A citalopram a passzív
ébrenlétet nem fokozta az előkezelt állatokon és a REM-alvást gátló hatása kisebb
mértékű volt, mint a nem előkezelteken.
Vizsgálataink bizonyítják, hogy egyetlen dózis MDMA is tartós változásokat
okozhat a motoros aktivitás, a cirkadián ritmus és az alvás-ébrenlét szabályozásában,
ami felhívja a figyelmet az ecstasyfogyasztás potenciális veszélyeire.
92

8 SUMMARY
The recreational drug ecstasy (3,4-methylenedioxymethamphetamine, MDMA)
has become increasingly used by young people all over the world. Despite the well
documented acute neurochemical and long-term serotonergic neurotoxical actions of
MDMA, short- and long-term effects on sleep-wake cycle have not been extensively
explored. No information is available on the acute sleep effects of a second MDMA
dose on MDMA-pretreated animals and the effect of the selective serotonin reuptake
inhibitor citalopram on sleep patterns has not been explored under these conditions
either.
Our aim was to assess possible functional changes in the 5-HT system following
the administration of a single dose of MDMA. In our experiments we used Dark Agouti
rats which provide a model for the genetically-defined, poor metabolizer human sub-
population in which clinical complications of MDMA may be more likely to occur. We
examined the effects of MDMA on motor activity and sleep-wake cycle up to 4 weeks
after treatment in drug-naïve rats and studied the acute effects on rats exposed to the
drug 3 weeks earlier. Additionally we compared the acute effects of citalopram on sleep
patterns in drug-naïve rats and in rats previously exposed to MDMA. Furthermore, in
parallel groups of animals, we quantified axonal damage by measuring (3H)-paroxetine
binding in motor areas and the occipital cortex 3 weeks after MDMA treatment.
In drug-naïve rats, MDMA initially increased motor activity and the awake period
and caused longer inhibition of REM (rapid eye movement) sleep than that of slow
wave sleep. 10-12 hours later adverse subacute effects such as decrease in motor
activity and increase in sleep followed the acute effects. Three days after MDMA
treatment motor activity was reduced mainly during dark (active) phase. Furthermore,
MDMA increased wakefulness and reduced REM-sleep 5 days after treatment.
Circadian patterns of motor activity and sleep/vigilance parameters were also disturbed
14 and 28 days after treatment. An increase in the length of REM was observed early in
the sleep cycle 3 weeks after MDMA treatment. Significant reductions in
[3H]paroxetine binding in the occipital cortex of MDMA-treated animals provided
evidence for axonal damage, although no significant change was observed in motor
areas. In rats exposed to MDMA 3 weeks earlier, acute effects induced by MDMA on
93

motor activity and vigilance had a shorter duration. Both groups of animals exhibited a
decrease in REM following citalopram treatment, although the effects seen in rats
previously exposed to MDMA were attenuated compared to drug-naïve animals.
Futhermore citalopram did not increase passive wakefulness in rats exposed to MDMA
3 weeks earlier.
In conclusion, our findings provide evidence that even a single dose of MDMA
can result in long-term changes in the regulation of circadian rhythms, motor activity
and sleep generation, highlighting the potential dangers of human ecstasy-use.
94

9 IRODALOMJEGYZÉK Aberg, M., D. Wade, E. Wall and S. Izenwasser (2007). "Effect of MDMA (ecstasy)
on activity and cocaine conditioned place preference in adult and adolescent rats." Neurotoxicol Teratol 29(1): 37-46.
Acquas, E., P. Marrocu, A. Pisanu, C. Cadoni, G. Zernig, A. Saria and G. Di Chiara (2001). "Intravenous administration of ecstasy (3,4-methylendioxymethamphetamine) enhances cortical and striatal acetylcholine release in vivo." Eur J Pharmacol 418(3): 207-11.
Aghajanian, G. K., Sanders-Bush, E. (2002). Serotonin. Chapter 2. Neuropsychopharmacology: The Fifth Generation of Progress. K. L. Davis, Charney, D., Coyle, J. T., Nemeroff C., American College of Neuropsychopharmacology.
Alcaro, A., R. Huber and J. Panksepp (2007). "Behavioral functions of the mesolimbic dopaminergic system: an affective neuroethological perspective." Brain Res Rev 56(2): 283-321.
Anderson, G. M., 3rd, G. Braun, U. Braun, D. E. Nichols and A. T. Shulgin (1978). "Absolute configuration and psychotomimetic activity." NIDA Res Monogr(22): 8-15.
Ando, R. D., A. Benko, L. Ferrington, E. Kirilly, P. A. Kelly and G. Bagdy (2006). "Partial lesion of the serotonergic system by a single dose of MDMA results in behavioural disinhibition and enhances acute MDMA-induced social behaviour on the social interaction test." Neuropharmacology 50(7): 884-96.
Arnold, H. M., J. Fadel, M. Sarter and J. P. Bruno (2001). "Amphetamine-stimulated cortical acetylcholine release: role of the basal forebrain." Brain Res 894(1): 74-87.
Azmitia, E. C., Whitaker-Azmitia, P.M. (2000). Anatomy, cell biology, and maturation of the serotonergic system: neurotrophic implications for the actions of psychotropic drugs. In: S.J. Watson(ed.): Psychopharmacology: The Fourth Generation of Progress, Part I., Chapter 42. New York, Lippincott Williams & Wilkins, Online Edition.
Baghdoyan, H. A. and R. Lydic (1999). "M2 muscarinic receptor subtype in the feline medial pontine reticular formation modulates the amount of rapid eye movement sleep." Sleep 22(7): 835-47.
Ball, K. T., D. Budreau and G. V. Rebec (2003). "Acute effects of 3,4-methylenedioxymethamphetamine on striatal single-unit activity and behavior in freely moving rats: differential involvement of dopamine D(1) and D(2) receptors." Brain Res 994(2): 203-15.
Ball, K. T. and G. V. Rebec (2005). "Role of 5-HT2A and 5-HT2C/B receptors in the acute effects of 3,4-methylenedioxymethamphetamine (MDMA) on striatal single-unit activity and locomotion in freely moving rats." Psychopharmacology (Berl) 181(4): 676-87.
Balogh, B., E. Molnar, R. Jakus, L. Quate, H. J. Olverman, P. A. Kelly, S. Kantor and G. Bagdy (2004). "Effects of a single dose of 3,4-methylenedioxymethamphetamine on circadian patterns, motor activity
95

and sleep in drug-naive rats and rats previously exposed to MDMA." Psychopharmacology (Berl) 173(3-4): 296-309.
Bankson, M. G. and K. A. Cunningham (2002). "Pharmacological studies of the acute effects of (+)-3,4-methylenedioxymethamphetamine on locomotor activity: role of 5-HT(1B/1D) and 5-HT(2) receptors." Neuropsychopharmacology 26(1): 40-52.
Bankson, M. G. and B. K. Yamamoto (2004). "Serotonin-GABA interactions modulate MDMA-induced mesolimbic dopamine release." J Neurochem 91(4): 852-9.
Battaglia, G., S. Y. Yeh, E. O'Hearn, M. E. Molliver, M. J. Kuhar and E. B. De Souza (1987). "3,4-Methylenedioxymethamphetamine and 3,4-methylenedioxyamphetamine destroy serotonin terminals in rat brain: quantification of neurodegeneration by measurement of [3H]paroxetine-labeled serotonin uptake sites." J Pharmacol Exp Ther 242(3): 911-6.
Baumann, M. H., X. Wang and R. B. Rothman (2007). "3,4-Methylenedioxymethamphetamine (MDMA) neurotoxicity in rats: a reappraisal of past and present findings." Psychopharmacology (Berl) 189(4): 407-24.
Bengel, D., D. L. Murphy, A. M. Andrews, C. H. Wichems, D. Feltner, A. Heils, R. Mossner, H. Westphal and K. P. Lesch (1998). "Altered brain serotonin homeostasis and locomotor insensitivity to 3, 4-methylenedioxymethamphetamine ("Ecstasy") in serotonin transporter-deficient mice." Mol Pharmacol 53(4): 649-55.
Benzenhofer, U. and T. Passie (2006). "[The early history of "Ecstasy"]." Nervenarzt 77(1): 95-6, 98-9.
Biello, S. M. and R. I. Dafters (2001). "MDMA and fenfluramine alter the response of the circadian clock to a serotonin agonist in vitro." Brain Res 920(1-2): 202-9.
Bódizs, R. (2000). Alvás és álmosság. Alvás, álom, bioritmusok.Budapest, Medicina Könyvkiadó: 21-74.
Bolla, K. I., U. D. McCann and G. A. Ricaurte (1998). "Memory impairment in abstinent MDMA ("Ecstasy") users." Neurology 51(6): 1532-7.
Bonaventure, P., L. Kelly, L. Aluisio, J. Shelton, B. Lord, R. Galici, K. Miller, T. W. Lovenberg, J. Atack and C. Dugovic (2007). "Selective blockade of 5-HT7 receptors enhances 5-HT transmission, antidepressant-like behavior and REM sleep suppression induced by citalopram in rodents." J Pharmacol Exp Ther.
Brown, G. L., F. K. Goodwin, J. C. Ballenger, P. F. Goyer and L. F. Major (1979). "Aggression in humans correlates with cerebrospinal fluid amine metabolites." Psychiatry Res 1(2): 131-9.
Bubar, M. J., K. M. Pack, P. S. Frankel and K. A. Cunningham (2004). "Effects of dopamine D1- or D2-like receptor antagonists on the hypermotive and discriminative stimulus effects of (+)-MDMA." Psychopharmacology (Berl) 173(3-4): 326-36.
Buchert, R., R. Thomasius, B. Nebeling, K. Petersen, J. Obrocki, L. Jenicke, F. Wilke, L. Wartberg, P. Zapletalova and M. Clausen (2003). "Long-term effects of "ecstasy" use on serotonin transporters of the brain investigated by PET." J Nucl Med 44(3): 375-84.
96

Buchert, R., R. Thomasius, F. Wilke, K. Petersen, B. Nebeling, J. Obrocki, O. Schulze, U. Schmidt and M. Clausen (2004). "A voxel-based PET investigation of the long-term effects of "Ecstasy" consumption on brain serotonin transporters." Am J Psychiatry 161(7): 1181-9.
Burgess, C., A. O'Donohoe and M. Gill (2000). "Agony and ecstasy: a review of MDMA effects and toxicity." Eur Psychiatry 15(5): 287-94.
Burns, N., H. J. Olverman, P. A. Kelly and B. C. Williams (1996). "Effects of ecstasy on aldosterone secretion in the rat in vivo and in vitro." Endocr Res 22(4): 601-6.
Byrne, T., L. E. Baker and A. Poling (2000). "MDMA and learning: effects of acute and neurotoxic exposure in the rat." Pharmacol Biochem Behav 66(3): 501-8.
Cadet, J. L. and C. Brannock (1998). "Free radicals and the pathobiology of brain dopamine systems." Neurochem Int 32(2): 117-31.
Cambras, T., J. Vilaplana, A. Campuzano, M. M. Canal-Corretger, M. Carulla and A. Diez-Noguera (2000). "Entrainment of the rat motor activity rhythm: effects of the light-dark cycle and physical exercise." Physiol Behav 70(3-4): 227-32.
Capece, M. L., S. M. Efange and R. Lydic (1997). "Vesicular acetylcholine transport inhibitor suppresses REM sleep." Neuroreport 8(2): 481-4.
Capela, J. P., A. Meisel, A. R. Abreu, P. S. Branco, L. M. Ferreira, A. M. Lobo, F. Remiao, M. L. Bastos and F. Carvalho (2006). "Neurotoxicity of Ecstasy metabolites in rat cortical neurons, and influence of hyperthermia." J Pharmacol Exp Ther 316(1): 53-61.
Chance, M. (1947). "Factors influencing the toxicity of sympathomimetic amines to solitary mice." J Pharmacol Exp Ther 89: 289-296.
Chang, J. Y. and C. Owman (1987). "Involvement of specific receptors and calcium mechanisms in serotonergic contractile response of isolated cerebral and peripheral arteries from rats." J Pharmacol Exp Ther 242(2): 629-36.
Coccaro, E. F. (1989). "Central serotonin and impulsive aggression." Br J Psychiatry Suppl(8): 52-62.
Colado, M. I., T. K. Murray and A. R. Green (1993). "5-HT loss in rat brain following 3,4-methylenedioxymethamphetamine (MDMA), p-chloroamphetamine and fenfluramine administration and effects of chlormethiazole and dizocilpine." Br J Pharmacol 108(3): 583-9.
Colado, M. I., E. O'Shea and A. R. Green (2004). "Acute and long-term effects of MDMA on cerebral dopamine biochemistry and function." Psychopharmacology (Berl) 173(3-4): 249-63.
Colado, M. I., J. L. Williams and A. R. Green (1995). "The hyperthermic and neurotoxic effects of 'Ecstasy' (MDMA) and 3,4 methylenedioxyamphetamine (MDA) in the Dark Agouti (DA) rat, a model of the CYP2D6 poor metabolizer phenotype." Br J Pharmacol 115(7): 1281-9.
Colbron, S., M. Jones and S. M. Biello (2002). "MDMA alters the response of the circadian clock to a photic and non-photic stimulus." Brain Res 956(1): 45-52.
97

Cole, J. C. and H. R. Sumnall (2003). "The pre-clinical behavioural pharmacology of 3,4-methylenedioxymethamphetamine (MDMA)." Neurosci Biobehav Rev 27(3): 199-217.
Commins, D. L., G. Vosmer, R. M. Virus, W. L. Woolverton, C. R. Schuster and L. S. Seiden (1987). "Biochemical and histological evidence that methylenedioxymethylamphetamine (MDMA) is toxic to neurons in the rat brain." J Pharmacol Exp Ther 241(1): 338-45.
Craig, D. A. and G. R. Martin (1993). "5-HT1B receptors mediate potent contractile responses to 5-HT in rat caudal artery." Br J Pharmacol 109(3): 609-11.
Crespi, D., T. Mennini and M. Gobbi (1997). "Carrier-dependent and Ca(2+)-dependent 5-HT and dopamine release induced by (+)-amphetamine, 3,4-methylendioxymethamphetamine, p-chloroamphetamine and (+)-fenfluramine." Br J Pharmacol 121(8): 1735-43.
Dafters, R. I. (1995). "Hyperthermia following MDMA administration in rats: effects of ambient temperature, water consumption, and chronic dosing." Physiol Behav 58(5): 877-82.
Dafters, R. I. and S. M. Biello (2003). "The effect of 3,4-methylenedioxymethamphetamine ('Ecstasy') on serotonergic regulation of the mammalian circadian clock mechanism in rats: the role of dopamine and hyperthermia." Neurosci Lett 350(2): 117-21.
Dafters, R. I. and E. Lynch (1998). "Persistent loss of thermoregulation in the rat induced by 3,4-methylenedioxymethamphetamine (MDMA or "Ecstasy") but not by fenfluramine." Psychopharmacology (Berl) 138(2): 207-12.
Davis, W. M. and R. F. Borne (1984). "Pharmacologic investigation of compounds related to 3,4-methylenedioxyamphetamine (MDA)." Subst Alcohol Actions Misuse 5(2): 105-10.
Day, J. and H. C. Fibiger (1992). "Dopaminergic regulation of cortical acetylcholine release." Synapse 12(4): 281-6.
de la Torre, R. and M. Farre (2004). "Neurotoxicity of MDMA (ecstasy): the limitations of scaling from animals to humans." Trends Pharmacol Sci 25(10): 505-8.
de la Torre, R., M. Farre, B. O. Mathuna, P. N. Roset, N. Pizarro, M. Segura, M. Torrens, J. Ortuno, M. Pujadas and J. Cami (2005). "MDMA (ecstasy) pharmacokinetics in a CYP2D6 poor metaboliser and in nine CYP2D6 extensive metabolisers." Eur J Clin Pharmacol 61(7): 551-4.
de la Torre, R., M. Farre, J. Ortuno, M. Mas, R. Brenneisen, P. N. Roset, J. Segura and J. Cami (2000). "Non-linear pharmacokinetics of MDMA ('ecstasy') in humans." Br J Clin Pharmacol 49(2): 104-9.
Doyon, S. (2001). "The many faces of ecstasy." Curr Opin Pediatr 13(2): 170-6. Dugovic, C., A. Wauquier, J. E. Leysen, R. Marrannes and P. A. Janssen (1989).
"Functional role of 5-HT2 receptors in the regulation of sleep and wakefulness in the rat." Psychopharmacology (Berl) 97(4): 436-42.
Escobedo, I., E. O'Shea, L. Orio, V. Sanchez, M. Segura, R. de la Torre, M. Farre, A. R. Green and M. I. Colado (2005). "A comparative study on the acute and long-term effects of MDMA and 3,4-dihydroxymethamphetamine (HHMA) on brain monoamine levels after i.p. or striatal administration in mice." Br J Pharmacol 144(2): 231-41.
98

EU, K. K. (2006). A kábítószer-probléma Európában. Éves jelentés. Luxemburg, Kábítószer és Kábítószer-függőség Európai Megfigyelőközpontja: 47-57.
Feige, B., U. Voderholzer, D. Riemann, R. Dittmann, F. Hohagen and M. Berger (2002). "Fluoxetine and sleep EEG: effects of a single dose, subchronic treatment, and discontinuation in healthy subjects." Neuropsychopharmacology 26(2): 246-58.
Ferrington, L., E. Kirilly, D. E. McBean, H. J. Olverman, G. Bagdy and P. A. Kelly (2006). "Persistent cerebrovascular effects of MDMA and acute responses to the drug." Eur J Neurosci 24(2): 509-19.
Fischer, H. S., G. Zernig, D. S. Schatz, C. Humpel and A. Saria (2000). "MDMA ('ecstasy') enhances basal acetylcholine release in brain slices of the rat striatum." Eur J Neurosci 12(4): 1385-90.
Fitzgerald, J. L. and J. J. Reid (1994). "Sympathomimetic actions of methylenedioxymethamphetamine in rat and rabbit isolated cardiovascular tissues." J Pharm Pharmacol 46(10): 826-32.
Fletcher, P. J., J. Sinyard and G. A. Higgins (2006). "The effects of the 5-HT(2C) receptor antagonist SB242084 on locomotor activity induced by selective, or mixed, indirect serotonergic and dopaminergic agonists." Psychopharmacology (Berl) 187(4): 515-25.
Fone, K. C., S. R. Beckett, I. A. Topham, J. Swettenham, M. Ball and L. Maddocks (2002). "Long-term changes in social interaction and reward following repeated MDMA administration to adolescent rats without accompanying serotonergic neurotoxicity." Psychopharmacology (Berl) 159(4): 437-44.
Frazer, A., Hensler, J. G. (1999). Serotonin: Involvement in Physiological Function and Behavior. Basic Neurochemistry, Lippincott Williams and Wilkins. Part Two. Intercellular Signaling.
Frederick, D. L. and M. G. Paule (1997). "Effects of MDMA on complex brain function in laboratory animals." Neurosci Biobehav Rev 21(1): 67-78.
Freudenmann, R. W., F. Oxler and S. Bernschneider-Reif (2006). "The origin of MDMA (ecstasy) revisited: the true story reconstructed from the original documents." Addiction 101(9): 1241-5.
Fürst, Z. (2006). Farmakológia. Budapest, Medicina Könyvkiadó. Gardani, M., R. N. Blance and S. M. Biello (2005). "MDMA alters the response of
the mammalian circadian clock in hamsters: effects on re-entrainment and triazolam-induced phase shifts." Brain Res 1046(1-2): 105-15.
Gartside, S. E., R. McQuade and T. Sharp (1996). "Effects of repeated administration of 3,4-methylenedioxymethamphetamine on 5-hydroxytryptamine neuronal activity and release in the rat brain in vivo." J Pharmacol Exp Ther 279(1): 277-83.
Gerra, G., A. Zaimovic, M. Ferri, U. Zambelli, M. Timpano, E. Neri, G. F. Marzocchi, R. Delsignore and F. Brambilla (2000). "Long-lasting effects of (+/-)3,4-methylenedioxymethamphetamine (ecstasy) on serotonin system function in humans." Biol Psychiatry 47(2): 127-36.
Gerra, G., A. Zaimovic, G. Giucastro, D. Maestri, C. Monica, R. Sartori, R. Caccavari and R. Delsignore (1998). "Serotonergic function after (+/-)3,4-methylene-dioxymethamphetamine ('Ecstasy') in humans." Int Clin Psychopharmacol 13(1): 1-9.
99

Geyer, M. A. (1996). "Serotonergic functions in arousal and motor activity." Behav Brain Res 73(1-2): 31-5.
Gordon, C. J., W. P. Watkinson, J. P. O'Callaghan and D. B. Miller (1991). "Effects of 3,4-methylenedioxymethamphetamine on autonomic thermoregulatory responses of the rat." Pharmacol Biochem Behav 38(2): 339-44.
Green, A. R., A. J. Cross and G. M. Goodwin (1995). "Review of the pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine (MDMA or "Ecstasy")." Psychopharmacology (Berl) 119(3): 247-60.
Green, A. R. and I. S. McGregor (2002). "On the anxiogenic and anxiolytic nature of long-term cerebral 5-HT depletion following MDMA." Psychopharmacology (Berl) 162(4): 448-50.
Green, A. R., A. O. Mechan, J. M. Elliott, E. O'Shea and M. I. Colado (2003). "The pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine (MDMA, "ecstasy")." Pharmacol Rev 55(3): 463-508.
Green, A. R., E. O'Shea and M. I. Colado (2004a). "A review of the mechanisms involved in the acute MDMA (ecstasy)-induced hyperthermic response." Eur J Pharmacol 500(1-3): 3-13.
Green, A. R., V. Sanchez, E. O'Shea, K. S. Saadat, J. M. Elliott and M. I. Colado (2004b). "Effect of ambient temperature and a prior neurotoxic dose of 3,4-methylenedioxymethamphetamine (MDMA) on the hyperthermic response of rats to a single or repeated ('binge' ingestion) low dose of MDMA." Psychopharmacology (Berl) 173(3-4): 264-9.
Griebel, G. (1995). "5-Hydroxytryptamine-interacting drugs in animal models of anxiety disorders: more than 30 years of research." Pharmacol Ther 65(3): 319-95.
Grinspoon, L. and J. B. Bakalar (1986). "Can drugs be used to enhance the psychotherapeutic process?" Am J Psychother 40(3): 393-404.
Gudelsky, G. A. and B. K. Yamamoto (2007). "Actions of 3,4-methylenedioxymethamphetamine (MDMA) on cerebral dopamineric, serotonergic and cholinergic neurons." Pharmacol Biochem Behav.
Gunn, J. A. and M. R. Gurd (1940). "The action of some amines related to adrenaline. Cyclohexylalkylamines." J Physiol 97(4): 453-470.
Henry, J. A., K. J. Jeffreys and S. Dawling (1992). "Toxicity and deaths from 3,4-methylenedioxymethamphetamine ("ecstasy")." Lancet 340(8816): 384-7.
Herin, D. V., S. Liu, T. Ullrich, K. C. Rice and K. A. Cunningham (2005). "Role of the serotonin 5-HT2A receptor in the hyperlocomotive and hyperthermic effects of (+)-3,4-methylenedioxymethamphetamine." Psychopharmacology (Berl) 178(4): 505-13.
Holschneider, D. P. a. S., J.C. (2000). Monoamine Oxidase: Basic and Clinical Perspectives Psychopharmacology: The Fourth Generation of Progress, Part I., Chapter 46 S. J. Watson. New York, Lippincott Williams & Wilkins, Online Edition.
Hornung, J. P. (2003). "The human raphe nuclei and the serotonergic system." J Chem Neuroanat 26(4): 331-43.
Hoyer, D., D. E. Clarke, J. R. Fozard, P. R. Hartig, G. R. Martin, E. J. Mylecharane, P. R. Saxena and P. P. Humphrey (1994). "International
100

Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin)." Pharmacol Rev 46(2): 157-203.
Huxster, J. K., A. Pirona and M. J. Morgan (2006). "The sub-acute effects of recreational ecstasy (MDMA) use: a controlled study in humans." J Psychopharmacol 20(2): 281-90.
Imperato, A., M. C. Obinu and G. L. Gessa (1993). "Effects of cocaine and amphetamine on acetylcholine release in the hippocampus and caudate nucleus." Eur J Pharmacol 238(2-3): 377-81.
Ivarsson, M., L. M. Paterson and P. H. Hutson (2005). "Antidepressants and REM sleep in Wistar-Kyoto and Sprague-Dawley rats." Eur J Pharmacol 522(1-3): 63-71.
Jacobs, B. L. and C. A. Fornal (1997). "Serotonin and motor activity." Curr Opin Neurobiol 7(6): 820-5.
Jacobs, B. L. and C. A. Fornal (1999). "Activity of serotonergic neurons in behaving animals." Neuropsychopharmacology 21(2 Suppl): 9S-15S.
Jacobs, B. L., Fornal, C. A. (2000). Serotonin and Behavior. A General Hypothesis. Psychopharmacology: The Fourth Generation of Progress.
Jones, D. C., S. S. Lau and T. J. Monks (2004). "Thioether metabolites of 3,4-methylenedioxyamphetamine and 3,4-methylenedioxymethamphetamine inhibit human serotonin transporter (hSERT) function and simultaneously stimulate dopamine uptake into hSERT-expressing SK-N-MC cells." J Pharmacol Exp Ther 311(1): 298-306.
Kalant, H. (2001). "The pharmacology and toxicology of "ecstasy" (MDMA) and related drugs." Cmaj 165(7): 917-28.
Kalivas, P. W., P. Duffy and S. R. White (1998). "MDMA elicits behavioral and neurochemical sensitization in rats." Neuropsychopharmacology 18(6): 469-79.
Kantor, S., K. Gerber, P. Halasz and G. Bagdy (2000). "The role of 5-HT2A, 5-HT2C and 5HT3 serotonin receptors in the regulation of sleep." J. Physiol. 526(Suppl): 66P-67P.
Kantor, S., R. Jakus, B. Balogh, A. Benko and G. Bagdy (2004). "Increased wakefulness, motor activity and decreased theta activity after blockade of the 5-HT2B receptor by the subtype-selective antagonist SB-215505." Br J Pharmacol 142(8): 1332-42.
Kantor, S., R. Jakus, R. Bodizs, P. Halasz and G. Bagdy (2002). "Acute and long-term effects of the 5-HT2 receptor antagonist ritanserin on EEG power spectra, motor activity, and sleep: changes at the light-dark phase shift." Brain Res 943(1): 105-11.
Kantor, S., R. Jakus, E. Molnar, N. Gyongyosi, A. Toth, L. Detari and G. Bagdy (2005). "Despite similar anxiolytic potential, the 5-hydroxytryptamine 2C receptor antagonist SB-242084 [6-chloro-5-methyl-1-[2-(2-methylpyrid-3-yloxy)-pyrid-5-yl carbamoyl] indoline] and chlordiazepoxide produced differential effects on electroencephalogram power spectra." J Pharmacol Exp Ther 315(2): 921-30.
Kirilly, E., A. Benko, L. Ferrington, R. D. Ando, P. A. Kelly and G. Bagdy (2006). "Acute and long-term effects of a single dose of MDMA on aggression in Dark Agouti rats." Int J Neuropsychopharmacol 9(1): 63-76.
101

Kirilly, E., Molnar, E., Balogh, B., Kantor, S., Hansson, S. R., Palkovits, M., Bagdy, G. (in press). "Decrease in REM latency and changes in sleep quality parallel serotonergic damage after MDMA: a longitudinal study over 180 days." Int J Neuropsychopharmacol. EPub: 2008 Feb 8;:1-15.
Kish, S. J., Y. Furukawa, L. Ang, S. P. Vorce and K. S. Kalasinsky (2000). "Striatal serotonin is depleted in brain of a human MDMA (Ecstasy) user." Neurology 55(2): 294-6.
Kuschinsky, W. (1991). "Coupling of function, metabolism, and blood flow in the brain." Neurosurg Rev 14(3): 163-8.
Kuypers, K. P., M. Wingen, N. Samyn, N. Limbert and J. G. Ramaekers (2007). "Acute effects of nocturnal doses of MDMA on measures of impulsivity and psychomotor performance throughout the night." Psychopharmacology (Berl) 192(1): 111-9.
Le Bars, D. (1994). Serotonin and pain. Neuronal Serotonin. N. N. Osborne, Hamon, M. New York, Wiley: 171-229.
Leonardi, E. T. and E. C. Azmitia (1994). "MDMA (ecstasy) inhibition of MAO type A and type B: comparisons with fenfluramine and fluoxetine (Prozac)." Neuropsychopharmacology 10(4): 231-8.
Levitt, P., J. E. Pintar and X. O. Breakefield (1982). "Immunocytochemical demonstration of monoamine oxidase B in brain astrocytes and serotonergic neurons." Proc Natl Acad Sci U S A 79(20): 6385-9.
Liechti, M. E. and F. X. Vollenweider (2001). "Which neuroreceptors mediate the subjective effects of MDMA in humans? A summary of mechanistic studies." Hum Psychopharmacol 16(8): 589-598.
Linnoila, M., M. Virkkunen, T. George and D. Higley (1993). "Impulse control disorders." Int Clin Psychopharmacol 8 Suppl 1: 53-6.
Liu, Y. and R. H. Edwards (1997). "The role of vesicular transport proteins in synaptic transmission and neural degeneration." Annu Rev Neurosci 20: 125-56.
Lowry, C. A. (2002). "Functional subsets of serotonergic neurones: implications for control of the hypothalamic-pituitary-adrenal axis." J Neuroendocrinol 14(11): 911-23.
Maldonado, E. and J. F. Navarro (2001). "MDMA ("ecstasy") exhibits an anxiogenic-like activity in social encounters between male mice." Pharmacol Res 44(1): 27-31.
Malpass, A., J. M. White, R. J. Irvine, A. A. Somogyi and F. Bochner (1999). "Acute toxicity of 3,4-methylenedioxymethamphetamine (MDMA) in Sprague-Dawley and Dark Agouti rats." Pharmacol Biochem Behav 64(1): 29-34.
Marston, H. M., M. E. Reid, J. A. Lawrence, H. J. Olverman and S. P. Butcher (1999). "Behavioural analysis of the acute and chronic effects of MDMA treatment in the rat." Psychopharmacology (Berl) 144(1): 67-76.
Martin, J. R., M. Bos, F. Jenck, J. Moreau, V. Mutel, A. J. Sleight, J. Wichmann, J. S. Andrews, H. H. Berendsen, C. L. Broekkamp, G. S. Ruigt, C. Kohler and A. M. Delft (1998). "5-HT2C receptor agonists: pharmacological characteristics and therapeutic potential." J Pharmacol Exp Ther 286(2): 913-24.
102

Mayerhofer, A., K. A. Kovar and W. J. Schmidt (2001). "Changes in serotonin, dopamine and noradrenaline levels in striatum and nucleus accumbens after repeated administration of the abused drug MDMA in rats." Neurosci Lett 308(2): 99-102.
McCann, U. D., V. Eligulashvili, M. Mertl, D. L. Murphy and G. A. Ricaurte (1999). "Altered neuroendocrine and behavioral responses to m-chlorophenylpiperazine in 3,4-methylenedioxymethamphetamine (MDMA) users." Psychopharmacology (Berl) 147(1): 56-65.
McCann, U. D., V. Eligulashvili and G. A. Ricaurte (2000). "(+/-)3,4-Methylenedioxymethamphetamine ('Ecstasy')-induced serotonin neurotoxicity: clinical studies." Neuropsychobiology 42(1): 11-6.
McCann, U. D., M. Mertl, V. Eligulashvili and G. A. Ricaurte (1999). "Cognitive performance in (+/-) 3,4-methylenedioxymethamphetamine (MDMA, "ecstasy") users: a controlled study." Psychopharmacology (Berl) 143(4): 417-25.
McCann, U. D. and G. A. Ricaurte (2007). "Effects of (+/-) 3,4-methylenedioxymethamphetamine (MDMA) on sleep and circadian rhythms." ScientificWorldJournal 7: 231-8.
McCann, U. D., A. Ridenour, Y. Shaham and G. A. Ricaurte (1994). "Serotonin neurotoxicity after (+/-)3,4-methylenedioxymethamphetamine (MDMA; "Ecstasy"): a controlled study in humans." Neuropsychopharmacology 10(2): 129-38.
McCann, U. D., S. O. Slate and G. A. Ricaurte (1996). "Adverse reactions with 3,4-methylenedioxymethamphetamine (MDMA; 'ecstasy')." Drug Saf 15(2): 107-15.
McCann, U. D., Z. Szabo, U. Scheffel, R. F. Dannals and G. A. Ricaurte (1998). "Positron emission tomographic evidence of toxic effect of MDMA ("Ecstasy") on brain serotonin neurons in human beings." Lancet 352(9138): 1433-7.
McCann, U. D., Z. Szabo, E. Seckin, P. Rosenblatt, W. B. Mathews, H. T. Ravert, R. F. Dannals and G. A. Ricaurte (2005). "Quantitative PET studies of the serotonin transporter in MDMA users and controls using [11C]McN5652 and [11C]DASB." Neuropsychopharmacology 30(9): 1741-50.
McEntee, W. J. and T. H. Crook (1991). "Serotonin, memory, and the aging brain." Psychopharmacology (Berl) 103(2): 143-9.
McKenna, D. J. and S. J. Peroutka (1990). "Neurochemistry and neurotoxicity of 3,4-methylenedioxymethamphetamine (MDMA, "ecstasy")." J Neurochem 54(1): 14-22.
McNamara, M. G., J. P. Kelly and B. E. Leonard (1995). "Some behavioural and neurochemical aspects of subacute (+/-)3,4-methylenedioxymethamphetamine administration in rats." Pharmacol Biochem Behav 52(3): 479-84.
Meston, C. M. and B. B. Gorzalka (1992). "Psychoactive drugs and human sexual behavior: the role of serotonergic activity." J Psychoactive Drugs 24(1): 1-40.
Metzger, R. R., G. R. Hanson, J. W. Gibb and A. E. Fleckenstein (1998). "3-4-Methylenedioxymethamphetamine-induced acute changes in dopamine transporter function." Eur J Pharmacol 349(2-3): 205-10.
103

Meyer-Bernstein, E. L. and L. P. Morin (1996). "Differential serotonergic innervation of the suprachiasmatic nucleus and the intergeniculate leaflet and its role in circadian rhythm modulation." J Neurosci 16(6): 2097-111.
Milroy, C. M., J. C. Clark and A. R. Forrest (1996). "Pathology of deaths associated with "ecstasy" and "eve" misuse." J Clin Pathol 49(2): 149-53.
Modi, G. M., P. B. Yang, A. C. Swann and N. Dafny (2006). "Chronic exposure to MDMA (Ecstasy) elicits behavioral sensitization in rats but fails to induce cross-sensitization to other psychostimulants." Behav Brain Funct 2: 1.
Monaca, C., B. Boutrel, R. Hen, M. Hamon and J. Adrien (2003). "5-HT 1A/1B receptor-mediated effects of the selective serotonin reuptake inhibitor, citalopram, on sleep: studies in 5-HT 1A and 5-HT 1B knockout mice." Neuropsychopharmacology 28(5): 850-6.
Monti, J. M. and H. Jantos (1992). "Dose-dependent effects of the 5-HT1A receptor agonist 8-OH-DPAT on sleep and wakefulness in the rat." J Sleep Res 1(3): 169-175.
Monti, J. M. and H. Jantos (2006). "Effects of the 5-HT(7) receptor antagonist SB-269970 microinjected into the dorsal raphe nucleus on REM sleep in the rat." Behav Brain Res 167(2): 245-50.
Monti, J. M. and H. Jantos (2006). "Effects of the serotonin 5-HT2A/2C receptor agonist DOI and of the selective 5-HT2A or 5-HT2C receptor antagonists EMD 281014 and SB-243213, respectively, on sleep and waking in the rat." Eur J Pharmacol 553(1-3): 163-70.
Monti, J. M. and D. Monti (2000). "Role of dorsal raphe nucleus serotonin 5-HT1A receptor in the regulation of REM sleep." Life Sci 66(21): 1999-2012.
Monti, J. M., A. Ponzoni, H. Jantos, P. Lagos, R. Silveira and P. Banchero (1999). "Effects of accumbens m-chlorophenylbiguanide microinjections on sleep and waking in intact and 6-hydroxydopamine-treated rats." Eur J Pharmacol 364(2-3): 89-98.
Morgan, M. J. (2000). "Ecstasy (MDMA): a review of its possible persistent psychological effects." Psychopharmacology (Berl) 152(3): 230-48.
Nair, S. G. and G. A. Gudelsky (2005). "3,4-Methylenedioxymethamphetamine (MDMA) enhances the release of acetylcholine by 5-HT4 and D1 receptor mechanisms in the rat prefrontal cortex." Synapse 58(4): 229-35.
Nair, S. G. and G. A. Gudelsky (2006). "3,4-Methylenedioxymethamphetamine enhances the release of acetylcholine in the prefrontal cortex and dorsal hippocampus of the rat." Psychopharmacology (Berl) 184(2): 182-9.
Nash, J. F., Jr., H. Y. Meltzer and G. A. Gudelsky (1988). "Elevation of serum prolactin and corticosterone concentrations in the rat after the administration of 3,4-methylenedioxymethamphetamine." J Pharmacol Exp Ther 245(3): 873-9.
Nash, J. F. and B. K. Yamamoto (1992). "Methamphetamine neurotoxicity and striatal glutamate release: comparison to 3,4-methylenedioxymethamphetamine." Brain Res 581(2): 237-43.
Navarro, J. F. and E. Maldonado (1999). "Behavioral profile of 3,4-methylenedioxy-methamphetamine (MDMA) in agonistic encounters between male mice." Prog Neuropsychopharmacol Biol Psychiatry 23(2): 327-34.
104

Neckelmann, D., B. Bjorvatn, A. A. Bjorkum and R. Ursin (1996). "Citalopram: differential sleep/wake and EEG power spectrum effects after single dose and chronic administration." Behav Brain Res 79(1-2): 183-92.
Nelson, W., Y. L. Tong, J. K. Lee and F. Halberg (1979). "Methods for cosinor-rhythmometry." Chronobiologia 6(4): 305-23.
Nixdorf, W. L., K. B. Burrows, G. A. Gudelsky and B. K. Yamamoto (2001). "Enhancement of 3,4-methylenedioxymethamphetamine neurotoxicity by the energy inhibitor malonate." J Neurochem 77(2): 647-54.
Nofzinger, E. A., C. F. Reynolds, 3rd, M. E. Thase, E. Frank, J. R. Jennings, A. L. Fasiczka, L. R. Sullivan and D. J. Kupfer (1995). "REM sleep enhancement by bupropion in depressed men." Am J Psychiatry 152(2): 274-6.
O'Loinsigh, E. D., G. Boland, J. P. Kelly and K. M. O'Boyle (2001). "Behavioural, hyperthermic and neurotoxic effects of 3,4-methylenedioxymethamphetamine analogues in the Wistar rat." Prog Neuropsychopharmacol Biol Psychiatry 25(3): 621-38.
O'Shea, E., R. Granados, B. Esteban, M. I. Colado and A. R. Green (1998). "The relationship between the degree of neurodegeneration of rat brain 5-HT nerve terminals and the dose and frequency of administration of MDMA ('ecstasy')." Neuropharmacology 37(7): 919-26.
Oliver von Bohlen und Halbach, R. D. (2006). Neurotransmitters. Neurotransmitters and Neuromodulators (Second Edition). R. D. Oliver von Bohlen und Halbach, Wiley-VCH Verlag GmbH & Co. KGaA, Online Edition: 45-142.
Pace-Schott, E. F., Hobson, J. A. (2002). Basic Mechanisms of Sleep: New Evidence on the Neuroanatomy and Neuromodulation of the NREM-REM Cycle. Chapter 128. Neuropsychopharmacology: The Fifth Generation of Progress. K. L. Davis, Charney, D., Coyle, J. T., Nemeroff C., American College of Neuropsychopharmacology.
Parrott, A. C. (2001). "Human psychopharmacology of Ecstasy (MDMA): a review of 15 years of empirical research." Hum Psychopharmacol 16(8): 557-577.
Parrott, A. C. (2002). "Recreational Ecstasy/MDMA, the serotonin syndrome, and serotonergic neurotoxicity." Pharmacol Biochem Behav 71(4): 837-44.
Parrott, A. C. (2004). "Is ecstasy MDMA? A review of the proportion of ecstasy tablets containing MDMA, their dosage levels, and the changing perceptions of purity." Psychopharmacology (Berl) 173(3-4): 234-41.
Parrott, A. C. and J. Lasky (1998). "Ecstasy (MDMA) effects upon mood and cognition: before, during and after a Saturday night dance." Psychopharmacology (Berl) 139(3): 261-8.
Peroutka, S. J. (1987). "Incidence of recreational use of 3,4-methylenedimethoxymethamphetamine (MDMA, "ecstasy") on an undergraduate campus." N Engl J Med 317(24): 1542-3.
Piper, B. J., J. B. Fraiman and J. S. Meyer (2005). "Repeated MDMA ("Ecstasy") exposure in adolescent male rats alters temperature regulation, spontaneous motor activity, attention, and serotonin transporter binding." Dev Psychobiol 47(2): 145-57.
Piper, B. J. and J. S. Meyer (2004). "Memory deficit and reduced anxiety in young adult rats given repeated intermittent MDMA treatment during the periadolescent period." Pharmacol Biochem Behav 79(4): 723-31.
105

Poland, R. E., P. Lutchmansingh, J. T. McCracken, J. P. Zhao, G. L. Brammer, C. S. Grob, K. B. Boone and R. N. Pechnick (1997). "Abnormal ACTH and prolactin responses to fenfluramine in rats exposed to single and multiple doses of MDMA." Psychopharmacology (Berl) 131(4): 411-9.
Popa, D., C. Lena, V. Fabre, C. Prenat, J. Gingrich, P. Escourrou, M. Hamon and J. Adrien (2005). "Contribution of 5-HT2 receptor subtypes to sleep-wakefulness and respiratory control, and functional adaptations in knock-out mice lacking 5-HT2A receptors." J Neurosci 25(49): 11231-8.
Portas, C. M., B. Bjorvatn and R. Ursin (2000). "Serotonin and the sleep/wake cycle: special emphasis on microdialysis studies." Prog Neurobiol 60(1): 13-35.
Price, L. H., G. A. Ricaurte, J. H. Krystal and G. R. Heninger (1989). "Neuroendocrine and mood responses to intravenous L-tryptophan in 3,4-methylenedioxymethamphetamine (MDMA) users. Preliminary observations." Arch Gen Psychiatry 46(1): 20-2.
Quate, L., D. E. McBean, I. M. Ritchie, H. J. Olverman and P. A. Kelly (2004). "Acute methylenedioxymethamphetamine administration: effects on local cerebral blood flow and glucose utilisation in the Dark Agouti rat." Psychopharmacology (Berl) 173(3-4): 287-95.
Reneman, L., J. Booij, K. de Bruin, J. B. Reitsma, F. A. de Wolff, W. B. Gunning, G. J. den Heeten and W. van den Brink (2001). "Effects of dose, sex, and long-term abstention from use on toxic effects of MDMA (ecstasy) on brain serotonin neurons." Lancet 358(9296): 1864-9.
Reneman, L., E. Endert, K. de Bruin, J. Lavalaye, M. G. Feenstra, F. A. de Wolff and J. Booij (2002). "The acute and chronic effects of MDMA ("ecstasy") on cortical 5-HT2A receptors in rat and human brain." Neuropsychopharmacology 26(3): 387-96.
Reneman, L., J. B. Habraken, C. B. Majoie, J. Booij and G. J. den Heeten (2000). "MDMA ("Ecstasy") and its association with cerebrovascular accidents: preliminary findings." AJNR Am J Neuroradiol 21(6): 1001-7.
Ricaurte, G. A., K. T. Finnegan, I. Irwin and J. W. Langston (1990). "Aminergic metabolites in cerebrospinal fluid of humans previously exposed to MDMA: preliminary observations." Ann N Y Acad Sci 600: 699-708; discussion 708-10.
Ricaurte, G. A., A. L. Markowska, G. L. Wenk, G. Hatzidimitriou, J. Wlos and D. S. Olton (1993). "3,4-Methylenedioxymethamphetamine, serotonin and memory." J Pharmacol Exp Ther 266(2): 1097-105.
Ricaurte, G. A. and U. D. McCann (2001). "Experimental studies on 3,4-methylenedioxymethamphetamine (MDA, "ecstasy") and its potential to damage brain serotonin neurons." Neurotox Res 3(1): 85-99.
Ricaurte, G. A., J. Yuan and U. D. McCann (2000). "(+/-)3,4-Methylenedioxymethamphetamine ('Ecstasy')-induced serotonin neurotoxicity: studies in animals." Neuropsychobiology 42(1): 5-10.
Risbrough, V. B., V. L. Masten, S. Caldwell, M. P. Paulus, M. J. Low and M. A. Geyer (2006). "Differential contributions of dopamine D1, D2, and D3 receptors to MDMA-induced effects on locomotor behavior patterns in mice." Neuropsychopharmacology 31(11): 2349-58.
106

Robbins, T. W., Everitt B. J. (2000). Central Norepinephrine Neurons and Behavior. Psychopharmacology: The Fourth Generation of Progress. S. J. Watson. New York, Lippincott & Williams, Online Edition.
Robinson, T. E., E. Castaneda and I. Q. Whishaw (1993). "Effects of cortical serotonin depletion induced by 3,4-methylenedioxymethamphetamine (MDMA) on behavior, before and after additional cholinergic blockade." Neuropsychopharmacology 8(1): 77-85.
Ross, R. J., P. J. Gresch, W. A. Ball, L. D. Sanford and A. R. Morrison (1995). "REM sleep inhibition by desipramine: evidence for an alpha-1 adrenergic mechanism." Brain Res 701(1-2): 129-34.
Rothman, R. B. and M. H. Baumann (2002). "Therapeutic and adverse actions of serotonin transporter substrates." Pharmacol Ther 95(1): 73-88.
Rudnick, G. and J. Clark (1993). "From synapse to vesicle: the reuptake and storage of biogenic amine neurotransmitters." Biochim Biophys Acta 1144(3): 249-63.
Ruggiero, D. A., M. D. Underwood, P. M. Rice, J. J. Mann and V. Arango (1999). "Corticotropic-releasing hormone and serotonin interact in the human brainstem: behavioral implications." Neuroscience 91(4): 1343-54.
Sanders-Bush, E., Canton, H. (1998). Serotonin Receptors: Signal Transduction Pathways. In: S.J. Watson (ed.): Psychopharmacology: The Fourth Generation of Progress. Chapter 41. New York, Lippincott Williams & Wilkins Publishers.
Saper, C. B., T. C. Chou and T. E. Scammell (2001). "The sleep switch: hypothalamic control of sleep and wakefulness." Trends Neurosci 24(12): 726-31.
Sari, Y. (2004). "Serotonin1B receptors: from protein to physiological function and behavior." Neurosci Biobehav Rev 28(6): 565-82.
Scearce-Levie, K., S. S. Viswanathan and R. Hen (1999). "Locomotor response to MDMA is attenuated in knockout mice lacking the 5-HT1B receptor." Psychopharmacology (Berl) 141(2): 154-61.
Schifano, F. (2000). "Potential human neurotoxicity of MDMA ('Ecstasy'): subjective self-reports, evidence from an Italian drug addiction centre and clinical case studies." Neuropsychobiology 42(1): 25-33.
Schifano, F. (2004). "A bitter pill. Overview of ecstasy (MDMA, MDA) related fatalities." Psychopharmacology (Berl) 173(3-4): 242-8.
Schifano, F., L. Di Furia, G. Forza, N. Minicuci and R. Bricolo (1998). "MDMA ('ecstasy') consumption in the context of polydrug abuse: a report on 150 patients." Drug Alcohol Depend 52(1): 85-90.
Schmidt, C. J. (1987). "Neurotoxicity of the psychedelic amphetamine, methylenedioxymethamphetamine." J Pharmacol Exp Ther 240(1): 1-7.
Schmidt, C. J., G. M. Abbate, C. K. Black and V. L. Taylor (1990). "Selective 5-hydroxytryptamine2 receptor antagonists protect against the neurotoxicity of methylenedioxymethamphetamine in rats." J Pharmacol Exp Ther 255(2): 478-83.
Schmidt, C. J., C. K. Black, G. M. Abbate and V. L. Taylor (1990). "Methylenedioxymethamphetamine-induced hyperthermia and neurotoxicity are independently mediated by 5-HT2 receptors." Brain Res 529(1-2): 85-90.
107

Schmidt, C. J., L. Wu and W. Lovenberg (1986). "Methylenedioxymethamphetamine: a potentially neurotoxic amphetamine analogue." Eur J Pharmacol 124(1-2): 175-8.
Schwartz, P. J., T. A. Wehr, N. E. Rosenthal, J. J. Bartko, D. A. Oren, C. Luetke and D. L. Murphy (1995). "Serotonin and thermoregulation. Physiologic and pharmacologic aspects of control revealed by intravenous m-CPP in normal human subjects." Neuropsychopharmacology 13(2): 105-15.
Seiden, L. S., W. L. Woolverton, S. A. Lorens, J. E. Williams, R. L. Corwin, N. Hata and M. Olimski (1993). "Behavioral consequences of partial monoamine depletion in the CNS after methamphetamine-like drugs: the conflict between pharmacology and toxicology." NIDA Res Monogr 136: 34-46; discussion 46-52.
Selken, J. and D. E. Nichols (2007). "Alpha1-adrenergic receptors mediate the locomotor response to systemic administration of (+/-)-3,4-methylenedioxymethamphetamine (MDMA) in rats." Pharmacol Biochem Behav 86(4): 622-30.
Semple, D. M., K. P. Ebmeier, M. F. Glabus, R. E. O'Carroll and E. C. Johnstone (1999). "Reduced in vivo binding to the serotonin transporter in the cerebral cortex of MDMA ('ecstasy') users." Br J Psychiatry 175: 63-9.
Series, H. G., P. J. Cowen and T. Sharp (1994). "p-Chloroamphetamine (PCA), 3,4-methylenedioxy-methamphetamine (MDMA) and d-fenfluramine pretreatment attenuates d-fenfluramine-evoked release of 5-HT in vivo." Psychopharmacology (Berl) 116(4): 508-14.
Shankaran, M. and G. A. Gudelsky (1998). "Effect of 3,4-methylenedioxymethamphetamine (MDMA) on hippocampal dopamine and serotonin." Pharmacol Biochem Behav 61(4): 361-6.
Shankaran, M. and G. A. Gudelsky (1999). "A neurotoxic regimen of MDMA suppresses behavioral, thermal and neurochemical responses to subsequent MDMA administration." Psychopharmacology (Berl) 147(1): 66-72.
Sharpley, A. L., D. J. Williamson, M. E. Attenburrow, G. Pearson, P. Sargent and P. J. Cowen (1996). "The effects of paroxetine and nefazodone on sleep: a placebo controlled trial." Psychopharmacology (Berl) 126(1): 50-4.
Shulgin, A. T. (1986). "The background and chemistry of MDMA." J Psychoactive Drugs 18(4): 291-304.
Slikker, W., Jr., R. R. Holson, S. F. Ali, M. G. Kolta, M. G. Paule, A. C. Scallet, D. E. McMillan, J. R. Bailey, J. S. Hong and F. M. Scalzo (1989). "Behavioral and neurochemical effects of orally administered MDMA in the rodent and nonhuman primate." Neurotoxicology 10(3): 529-42.
Smith, M. I., D. C. Piper, M. S. Duxon and N. Upton (2002). "Effect of SB-243213, a selective 5-HT(2C) receptor antagonist, on the rat sleep profile: a comparison to paroxetine." Pharmacol Biochem Behav 71(4): 599-605.
Smith, S. E., R. O. Pihl, S. N. Young and F. R. Ervin (1987). "A test of possible cognitive and environmental influences on the mood lowering effect of tryptophan depletion in normal males." Psychopharmacology (Berl) 91(4): 451-7.
Spanos, L. J. and B. K. Yamamoto (1989). "Acute and subchronic effects of methylenedioxymethamphetamine [(+/-)MDMA] on locomotion and
108

serotonin syndrome behavior in the rat." Pharmacol Biochem Behav 32(4): 835-40.
Sprague, J. E., S. L. Everman and D. E. Nichols (1998). "An integrated hypothesis for the serotonergic axonal loss induced by 3,4-methylenedioxymethamphetamine." Neurotoxicology 19(3): 427-41.
Steinbusch, H. W. (1981). "Distribution of serotonin-immunoreactivity in the central nervous system of the rat-cell bodies and terminals." Neuroscience 6(4): 557-618.
Stone, D. M., K. M. Merchant, G. R. Hanson and J. W. Gibb (1987). "Immediate and long-term effects of 3,4-methylenedioxymethamphetamine on serotonin pathways in brain of rat." Neuropharmacology 26(12): 1677-83.
Stone, D. M., D. C. Stahl, G. R. Hanson and J. W. Gibb (1986). "The effects of 3,4-methylenedioxymethamphetamine (MDMA) and 3,4-methylenedioxyamphetamine (MDA) on monoaminergic systems in the rat brain." Eur J Pharmacol 128(1-2): 41-8.
Svensson, K., P. Alfoldi, M. Hajos, G. Rubicsek, A. M. Johansson, A. Carlsson and F. Obal, Jr. (1987). "Dopamine autoreceptor antagonists: effects on sleep-wake activity in the rat." Pharmacol. Biochem. Behav. 26(1): 123-9.
Taguchi, K., J. Atobe, M. Kato, T. Chuma, T. Chikuma, T. Shigenaga and T. Miyatake (1998). "The effect of methamphetamine on the release of acetylcholine in the rat striatum." Eur J Pharmacol 360(2-3): 131-7.
Takeuchi, Y., M. Kojima, T. Matsuura and Y. Sano (1983). "Serotonergic innervation on the motoneurons in the mammalian brainstem. Light and electron microscopic immunohistochemistry." Anat Embryol (Berl) 167(3): 321-33.
Thomas, D. R., S. Melotto, M. Massagrande, A. D. Gribble, P. Jeffrey, A. J. Stevens, N. J. Deeks, P. J. Eddershaw, S. H. Fenwick, G. Riley, T. Stean, C. M. Scott, M. J. Hill, D. N. Middlemiss, J. J. Hagan, G. W. Price and I. T. Forbes (2003). "SB-656104-A, a novel selective 5-HT7 receptor antagonist, modulates REM sleep in rats." Br J Pharmacol 139(4): 705-14.
Timar, J., S. Gyarmati, A. Szabo and S. Furst (2003). "Behavioural changes in rats treated with a neurotoxic dose regimen of dextrorotatory amphetamine derivatives." Behav Pharmacol 14(3): 199-206.
Tohyama, M., Takatsuji, K. (1998). Atlas of Neuroactive Substances and Their Receptors in the Rat. New York, Oxford University Press.
Topp, L., J. Hando, P. Dillon, A. Roche and N. Solowij (1999). "Ecstasy use in Australia: patterns of use and associated harm." Drug Alcohol Depend 55(1-2): 105-15.
Tortella, F. C., E. Echevarria, R. H. Pastel, B. Cox and T. P. Blackburn (1989). "Suppressant effects of selective 5-HT2 antagonists on rapid eye movement sleep in rats." Brain Res 485(2): 294-300.
Tucker, G. T., M. S. Lennard, S. W. Ellis, H. F. Woods, A. K. Cho, L. Y. Lin, A. Hiratsuka, D. A. Schmitz and T. Y. Chu (1994). "The demethylenation of methylenedioxymethamphetamine ("ecstasy") by debrisoquine hydroxylase (CYP2D6)." Biochem Pharmacol 47(7): 1151-6.
Verkes, R. J., H. J. Gijsman, M. S. Pieters, R. C. Schoemaker, S. de Visser, M. Kuijpers, E. J. Pennings, D. de Bruin, G. Van de Wijngaart, J. M. Van
109

Gerven and A. F. Cohen (2001). "Cognitive performance and serotonergic function in users of ecstasy." Psychopharmacology (Berl) 153(2): 196-202.
Wallace, T. L., G. A. Gudelsky and C. V. Vorhees (2001). "Alterations in diurnal and nocturnal locomotor activity in rats treated with a monoamine-depleting regimen of methamphetamine or 3,4-methylenedioxymethamphetamine." Psychopharmacology (Berl) 153(3): 321-6.
Westlund, K. N., R. M. Denney, R. M. Rose and C. W. Abell (1988). "Localization of distinct monoamine oxidase A and monoamine oxidase B cell populations in human brainstem." Neuroscience 25(2): 439-56.
White, C. R. and R. S. Seymour (2005). "Allometric scaling of mammalian metabolism." J Exp Biol 208(Pt 9): 1611-9.
Williams, J. A. and P. B. Reiner (1993). "Noradrenaline hyperpolarizes identified rat mesopontine cholinergic neurons in vitro." J Neurosci 13(9): 3878-83.
Wilson, S. J., J. E. Bailey, A. S. Rich, M. Adrover, J. Potokar and D. J. Nutt (2004). "Using sleep to evaluate comparative serotonergic effects of paroxetine and citalopram." Eur Neuropsychopharmacol 14(5): 367-72.
Young, S. N., S. E. Smith, R. O. Pihl and F. R. Ervin (1985). "Tryptophan depletion causes a rapid lowering of mood in normal males." Psychopharmacology (Berl) 87(2): 173-7.
Yuan, J., B. J. Cord, U. D. McCann, B. T. Callahan and G. A. Ricaurte (2002). "Effect of depleting vesicular and cytoplasmic dopamine on methylenedioxymethamphetamine neurotoxicity." J Neurochem 80(6): 960-9.
Zifa, E. and G. Fillion (1992). "5-Hydroxytryptamine receptors." Pharmacol Rev 44(3): 401-58.
110

10 AZ ÉRTEKEZÉS TÉMÁJÁHOZ KÖZVETLENÜL
KAPCSOLÓDÓ KÖZLEMÉNYEK, KIVONATOK,
ELŐADÁSOK ÉS POSZTEREK
Közlemények
1. Balogh B, Molnár E, Jakus R, Quate L, Olverman HJ, Kelly PA, Kántor S,
Bagdy G (2004). Effects of a single dose of 3,4-methylenedioxymethampheta-
mine on circadian patterns, motor activity and sleep in drug-naive rats and rats
previously exposed to MDMA. Psychopharmacology (Berl) 173: 296-309.
IF= 3,146
2. Eszter Kirilly, Eszter Molnar, Brigitta Balogh, Sandor Kantor, Stefan R.
Hansson, Miklos Palkovits, Gyorgy Bagdy: Decrease in REM latency and
changes in sleep quality parallel serotonergic damage after MDMA: a
longitudinal study over 180 days. Int J Neuropsychopharmacology. EPub: 2008
Feb 8;:1-15. IF= 5,184
Kivonatok
1. B. Balogh, E. Molnar, L. Quate, H. J. Olverman, P. A. T. Kelly, S. Kantor, G.
Bagdy: Acute and long-term consequences of 3,4-methylenedioxy-
methamphetamine administration to rats on motor activity, local cerebral
glucose utilization and [3H]paroxetine binding. FENS Abstracts, 2, A132.2,
2004.
2. B. Balogh, R. Jakus, E. Molnar, S. Kantor, G. Juhasz, G. Bagdy: Sleep and
vigilance effects of 3,4-ethylenedioxymethamphetamine in rats in a 28 days
long, 24-hour continous monitoring study. Fundamental and Clinical
Pharmacology 18, suppl.1. pp. 150, 2004.
111

3. B. Balogh, R. Jakus, S. Kantor, M. Graf, A. Benko, G. Bagdy: Effects of
3,4-Methylenedioxymethamphetamine on motor activity, sleep and
circadian patterns in drug-naive rats and rats previously exposed to
MDMA. Fundamental and Clinical Pharmacology 18, suppl.1. pp. 104, 2004.
4. N. Gyöngyösi, B. Balogh, S. Kántor, G. Bagdy: Effects of the 5-HT1B receptor
agonist CP94253 in rats treated with MDMA (Ecstasy) 6 months earlier.
Clinical Neuroscience 58, 1, 2005. B.88.
Előadások és poszterek
1. Balogh Brigitta, Molnár Eszter, Kántor Sándor, Bagdy György: Az Ecstasy
(MDMA) akut és krónikus hatásai patkányok alvás-ébrenléti ciklusára. IV.
Magyar Viselkedés-élettani konferencia, Budapest, 2003. október 30.
2. Balogh Brigitta, Jakus Rita, Molnár Eszter, Kántor Sándor, Graf Márton, Bagdy
György: Az ecstasy (MDMA) motoros aktivitás- és alvás/ébrenléti hatásainak
tanulmányozása Dark Agouti patkánytörzsön. PhD Napok 2004, Semmelweis
Egyetem, Budapest, 2004. április 8.
3. Bagdy György, Balogh Brigitta, Kántor Sándor: Az MDMA (Ecstasy) akut és
hosszú távú hatásai a napszaki ritmusra és az alvásra. A Magyar Alvástársaság
szimpóziuma, Budapest, 2004. június 18.
4. B. Balogh, E. Molnar, L. Quate, H. J. Olverman, P. A. T. Kelly, S. Kantor, G.
Bagdy: Acute and long-term consequences of 3,4-methylenedioxy-
methamphetamine administration to rats on motor activity, local cerebral
glucose utilization and [3H]paroxetine binding. FENS 2004, Lisszabon, July 10-
14, 2004.
5. B. Balogh, R. Jakus, S. Kantor, M. Graf, A. Benko, G. Bagdy: Effects of
3,4-Methylenedioxymethamphetamine on motor activity, sleep and
112

circadian patterns in drug-naive rats and rats previously exposed to
MDMA. EPHAR 2004, Porto, July 14-17, 2004.
6. B. Balogh, R. Jakus, E. Molnar, S. Kantor, G. Juhasz, G. Bagdy: Sleep
and vigilance effects of 3,4-Methylenedioxymethamphetamine in rats in
a 28 days long, 24-hour continous monitoring study. Serotonin Club Meeting
2004, Porto, July 18-19, 2004.
7. B. Balogh, E. Molnar, R. Jakus, L. Quate, H. J. Olverman, P. A. T. Kelly, S.
Kantor, G. Bagdy: Acute and long-term consequences of 3,4-
methylenedioxymethamphetamine administration to rats on motor activity,
sleep, local cerebral glucose utilization and [3H]paroxetine binding. IBRO
CEERC Summer School, Sensory and integrative neuroscience: from receptors
to behavior, Moscow, Russia, August 18-31, 2004.
8. Balogh, B., Molnar, E., Quate, L., Olverman, H. J., Kelly, P. A. T., Kantor, S.,
Bagdy, G.: Acute and long-term effects of 3,4-
methylenedioxymethamphetamine on motor activity, local cerebral glucose
utilization and [3H]paroxetine binding in motor areas. 10th International
Symposium on Motor Control, Sofia, Bulgaria, September 25-27, 2004.
9. Gyöngyösi Norbert, Balogh Brigitta, Jakus Rita, Kántor Sándor, Bagdy
György: Az MDMA (Ecstasy) hatásai a vigilanciára hosszú távú követéses
vizsgálatban, Magyar Kísérletes és Klinikai Farmakológiai Társaság VI.
Kongresszusa, Debrecen, 2004. december 9-11.
10. N. Gyöngyösi, B. Balogh, S. Kántor, G. Bagdy: Effects of the 5-HT1B receptor
agonist CP94253 in rats treated with MDMA (Ecstasy) 6 months earlier. 11th of
Congress of the Hungarian Society of Neuroscience, Pécs, January 25-29, 2005.
113

11. N. Gyongyosi, B. Balogh, S. Kantor, G. Bagdy: Long term effects of MDMA on
5-HT1B receptor functions. Spring Symposium of the Hungarian Society for
Experimental and Clinical Pharmacology, Budapest, June 6-7, 2005.
12. T Kitka, N Gyongyosi, B Balogh, S Kantor, G Bagdy. Suppression of REM as a
Measure of Antidepressant Pharmacotherapy: Studies with Citalopram.
Pharmacy: Smart Molecules for Therapy, Semi-centennial conference of
Semmelweis University, Faculty of Pharmacy, October 12-14, 2005.
13. T Kitka, Z Kátai, E Kirilly, B Balogh, T. Renoir, L. Lanfumey, M. Hamon, S.
Kantor, G. Bagdy. Partial serotonergic lesion attenuates REM sleep reduction by
citalopram. Joint Meeting of the Slovak Physiological Society and The
Physiological Society and The Federation of European Physiological Societies,
Bratislava, September 11-14, 2007.
114

11 AZ ÉRTEKEZÉS TÉMÁJÁHOZ KÖZVETLENÜL
NEM KAPCSOLÓDÓ KÖZLEMÉNYEK, KIVONATOK,
ELŐADÁSOK ÉS POSZTEREK
Közlemények
1. Rita Jakus, Marton Graf, Romeo D. Ando, Brigitta Balogh, Istvan Gacsalyi,
Gyorgy Levay, Sandor Kantor, Gyorgy Bagdy: Effect of two non-competitive
AMPA receptor antagonists GYKI 52466 and GYKI 53405 on spike-wave
discharges, vigilance and behavior in a genetic rat model of absence epilepsy.
Brain Research, 1008/2: 236-244, 2004. IF= 2,389
2. Sandor Kantor, Rita Jakus, Brigitta Balogh, Anita Benko, Gyorgy Bagdy:
Increased wakefulness, motor activity and decreased theta activity after selective
blockade of the 5-HT2B receptor by the subtype-selective antagonist SB-215505.
Br J Pharmacol, 142(8):1332-42, 2004. IF= 3,325
Kivonatok
1. R. Jakus, M. Graf, R.D. Ando, B. Balogh, I. Gacsalyi, G. Levay,
S. Kantor and G. Bagdy: Effects of AMPA receptor antagonists GYKI
52466 and GYKI 53405 on spike-wave discharges, vigilance and
behaviour in a genetic rat epilepsy model. Fundamental and Clinical
Pharmacology 18, suppl.1. pp. 48, 2004.
2. R. Jakus, M. Graf, G. Juhasz, B. Balogh, G. Levay, K. Gerber, and G. Bagdy:
Activation of 5-HT2C receptors inhibit, and 5-HT1A and 5-HT7 receptors
activate the generation of spike-wave discharges in a genetic rat epilepsy model.
Fundamental and Clinical Pharmacology 18, suppl.1. pp. 41, 2004.
115

3. S. Kantor, R. Jakus, B. Balogh, A. Benko, G. Bagdy: Increased wakefulness,
motor activity and decreased theta activity after selective blockade of the 5-
HT2B receptor by the subtype-selective antagonist SB-215505. Journal of Sleep
Research 13, suppl. 1., P. 382, 2004.
Előadások és poszterek
1. Kántor Sándor, Balogh Brigitta, Jakus Rita, Bagdy György: A szerotonerg
rendszer szerepe az alvás szabályozásában. A Magyar Alvástársaság
szimpóziuma, Budapest, 2004. június 18.
2. R. Jakus, M. Graf, R.D. Ando, B. Balogh, I. Gacsalyi, G. Levay,
S. Kantor and G. Bagdy: Effects of AMPA receptor antagonists GYKI
52466 and GYKI 53405 on spike-wave discharges, vigilance and
behaviour in a genetic rat epilepsy model. EPHAR 2004, Porto, July 14-17,
2004.
3. R. Jakus, M. Graf, G. Juhasz, B. Balogh, G. Levay, K. Gerber,
and G. Bagdy: Activation of 5-HT2C receptors inhibit, and 5-HT1A and
5-HT7 receptors activate the generation of spike-wave discharges in a
genetic rat epilepsy model. EPHAR 2004, Porto, July 14-17, 2004.
4. S. Kantor, R. Jakus, B. Balogh, A. Benko, G. Bagdy: Increased wakefulness,
motor activity and decreased theta activity after selective blockade of the 5-
HT2B receptor by the subtype-selective antagonist SB-215505. 17th Congress
of the ESRS, Prague, Czech Republic, October 5–9, 2004.
5. Bagdy Gy., Gyöngyösi N., Kitka T., Lazáry J., Balogh B., Benkő A., Andó
R.D., Kántor S. Klinikai hatások előrejelzése a központi idegrendszerre ható
gyógyszerek fejlesztésében: alvás és EEG vizsgálatok rágcsálókban. MBKE
Gyógyszerbiokémiai Szakosztály XXI. Munkaértekezlet, Balatonőszöd, 2006.
május 22-24.
116

6. Bagdy Gy., Gyöngyösi N., Kitka T., Balogh B., Benkő A., Andó R.D., Kántor
S. A klinikai hatások előrejelzése a központi idegrendszerre ható gyógyszerek
kutatásában: rágcsálókon végzett EEG-vizsgálatok. Az MFT II. tavaszi
experimentális szimpóziuma. Pécs, 2006. június 3.
117

12 KÖSZÖNETNYILVÁNÍTÁS
Kutatásaim eredménye ez a doktori disszertáció, amelynek elkészítéséért
köszönettel tartozom mindazoknak, akik segítségükkel, személyes támogatásukkal,
ösztönzésükkel hozzájárultak e munka elkészültéhez. Elsőként szeretném megköszönni
témavezetőmnek, Dr. Bagdy Györgynek, aki szakmai irányításával támogatott, értékes
tanácsaival segítette munkámat. Köszönöm Dr. Tímár Júliának a dolgozatom házi
bírálatát, áldozatkész segítségét, javaslatait, melyekkel rendkívül sokat segített a
dolgozatom jobbá tételében. A kísérleti módszerek elsajátításában nyújtott
nélkülözhetetlen segítségükért és értékes tanácsaikért köszönettel tartozom Dr. Kántor
Sándornak, Dr. Graf Mártonnak, Dr. Jakus Ritának, Dr. Anheuer Egonné Zsuzsának és
Nagy Rezsőné Nórának. Köszönöm segítségét Dr. Módosné Ányok Editnek, aki sok-
sok adminisztrációs és kémiai problémát segített megoldani. Köszönettel tartozom
munkatársaimnak: Molnár Eszternek, Kirilly Eszternek, Kitka Tamásnak, Gyöngyösi
Norbertnek, Dr. Juhász Gabriellának, Andó Rómeó Dénesnek, Benkő Anitának és a
többi munkatársamnak is. Köszönöm odaadó segítségét és tanácsait Dr. Paul A. T.
Kelly-nek, akinek edinburgh-i laboratóriumában 9 hónapot töltöttem, köszönettel
tartozom edinburgh-i munkatársaimnak, Dr. Henry J. Olverman-nak, Dr. Linda
Ferrington-nak és Neil Dawson-nak is segítségükért. Külön köszönet illeti Sidló
Zsuzsannát értekezésemmel kapcsolatos, rendkívül értékes szakmai és stilisztikai
tanácsaiért. Köszönöm férjemnek, Dr. Tamás Péternek kutatómunkám alatt és
értekezésem megírása közben nyújtott hasznos tanácsait, biztatását, támogatását. Végül,
de nem utolsó sorban szeretnék köszönetet mondani szüleimnek, kisfiamnak, férjem
édesanyjának, testvéremnek és családjának kitartó türelmükért és támogatásukért a
kutatómunkám hosszú évei alatt.
118













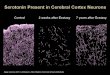














![[MDMA]MDMA Neurochemistry](https://img.dokumen.tips/doc/110x75/577dab601a28ab223f8c57f3/mdmamdma-neurochemistry.jpg)
