Embed Size (px)
Citation preview

Neuroscience Letters, 6 (1977) 143--149 © ~vier /North-HoHand Scientific Publishers Ltd.
AUDrl~RY-VISUAL INTERACTION IN CAT ORBrFAL.LNSULAR CORTEX
143
JAMES H, FALLON* and LOUIS A. BENEVENTO**, ***
Department of Neurosciences, University of California, San Diego Lc Jolla, Calif. 92093 (~.8.A.)
(Received July l l t h , 1977) (Accepted August 3rd, 1977)
SUMMARY
Extra- and intracellular recordings were made in the orbital and insular cortices of ~he cat. Auditory and visual stimuli were presented alone, simultane. ously and w~.~th the onsets of the stimuli paired at various conditioning-test intervals (At's) up to 1,000 msec. The largest class of interactions (25%) that occurred resulted in an inhibition of the cells' response. Both active inhibition (IPSPs) and presymaptic blockade (suppression without IPSPs at the recorded cell) were common responses. It was also found that the post-synaptic poten- rials induced by one stimulus could enhance or negate the post-synaptic potentials induced by the other stimulus. These simple interactions, which could be inferred by the summation of the cells' response to either stimulus, were ccntmsted wtih numerous complex responses at a wide range of A t's. It is concluded that most of the cells in the orbital and insular cortices of the cat are multimodal, with the interactions of visual and auditory stimuli occurring along multiple pathways with different latencies and functional synapses,
Single units in the orbital-insular cortex of the cat and lateral orbital cortex of the rhesus monkey respond to visual and auditory stimuli [1,5,6,7,12]. The distributio~ of unit responses in the orbital-insular cortex of the cat is dominated by units with bimodal responses (49%) while some of the units responded only to visual stimuli (12%)or auditory stimuli (16%). The present study was undertaken to describe the response characteristics of single units in the orbital-insular cortex of the cat which are bimodal, with special attention to the interaction of visual and auditory stimuli.
Thirteen cats were prepared for surgery and extra- and ~ntracellular physio-
* Supported by University of nlinois Assistantship and USPHS Grant NS-05187 ** Supported by NSF Grants BNS 75-7349 and BNS 75-17890 *** P.O. Box 6998, University of Illinois Medical Center, Chicago, Illinois 60680 U.S.A.

| ITq~J
Fig. 1. Lateral view of left cerebral hemisphere of cat. Black dots in upper drawing illustrate electrode penetrations in orbital (0) and insular (i) cortex. Lower drawing from maps of cortical areas described by Woolsey [ 17 ].
logical recordings were made as previously described [2,6,7,12]. The stimuli were bursts of clicks delivered through matched earphones and bright flashes of white light projected through an oscillating grid pattern of 100% contrast for 5 sec. The stimuli were present alone, simultaneously and with the onsets of the stimuli paired at different conditioning-test inte~als or "At 's" The A t'S varied from At = 1,000 msec (onset of visual stimulus leading onset of auditory stimulus by 1,000 msec) to At = 1,000 msec (onset of visual stimulus trailing onset o:~ auditory :~timulus by 1,000 msec). Intermediate values of At were also used, e.g., At's - 500, 200, 100, 50, 20, 0 , - -20 , - -50, - -100, - -200, --500 msec.
The recordirLg sites were vertified histologically. The recording sites were found to be in ~he auditory cortic~d areas AIII (orbital cortex) and AIV (insular cortex) as illustrated in Fig. 1.
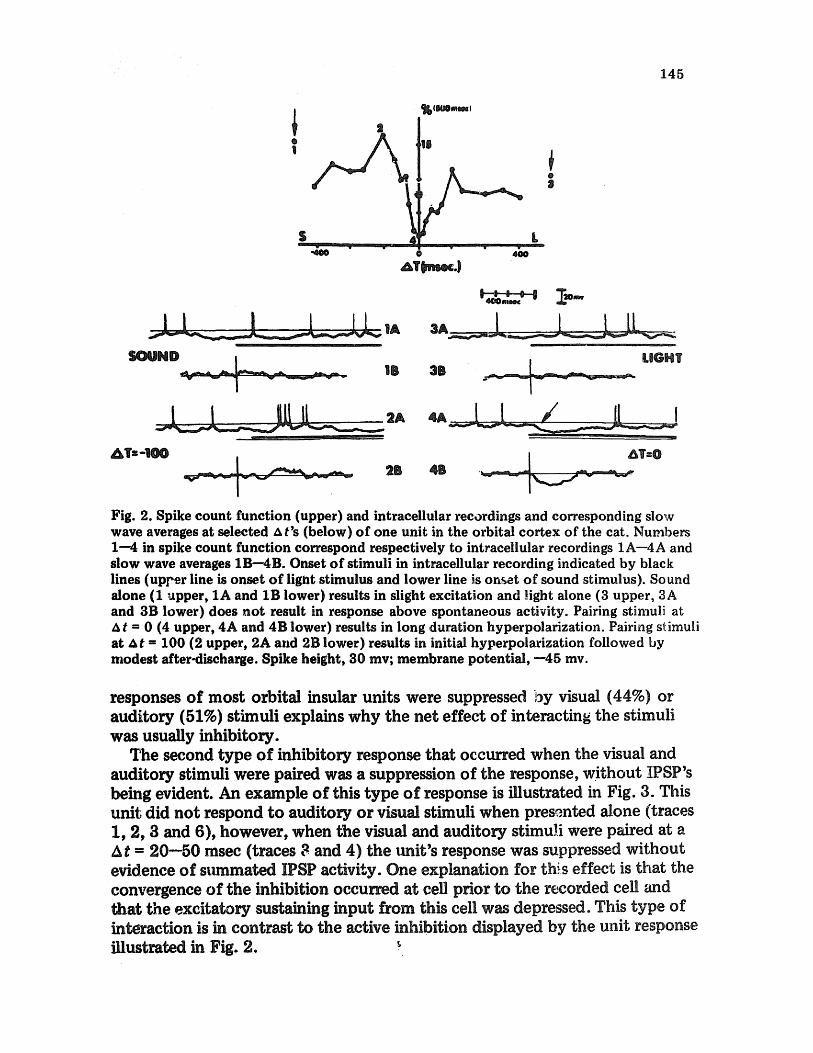
The majorit3' of the interaction~ (approx. 25% of a total of 123 cells) that occurred resuliied in a net inhibition of the cells response at short At's (0--100 msec). The inhibition occurred in two major forms. The first effect was an active inhibitioa at the recorded cell, resulting in summated IPSP (inhibitory post-synaptic potential) activity as illustrated in Fig. 2. This unit demonstrated no overt response to either a visua~ or auditory stimulus when presen*ed alone (auditory; traces 1A and 1B, visual; tra~es 3A and 3B) however, when the visual and auditory stimuli were presented simultaneously at At = 0 (traces 4A and 4B), a net inhibition ensued with striking IPSP activity evident (arrow in trace 4A). These results could ~ p l y that the visual and auditory input coverage on a cell prior to the recorded cell and that the summation of visual and auditory input provided a suprathreshold response on a cell that had an inhibitory input to the celt from which the recording was made. In some units, in which both a visual and auditory stimulus induced IPSP activity in a cell, the interaction of the two stimuli produced more profound IPSP activity. The fact that the
145
|
$ m
-4OO 0 , ~ T ~ . )
i
A1':-100
~(IWOmwe)
L
, ,
~T"O
Fig. 2. Spike count function (upper) and intracellular recordings and corresponding slow wave averages at selected A t's (below) of one unit in the orbital cortex of the cat. Numbers 1--4 in spike count function correspond respectively to intracellular recordings 1A--4A and slow wave averages 1B--4B. Onset of stimuli in intracellular recording indicated by black lines (up/,er line is onset of light stimulus and lower line is onset of sound stimulus). Sound alone (1 upper, 1A and 1B lower) results in slight excitation and light alone (3 upper, 3A and 3B lower) does not result in response above spontaneous activity. Pairing stimuli at A t = 0 (4 upper, 4A and 4B lower) results in long duration hyperpolarization. Pairing stimuli at At ffi 100 (2 upper, 2A and 2B lower) results in initial hyperpolarization followed by modest after-discharge. Spike height, 30 my; membrane potential, --45 my.
responses of most orbital insular units were suppressed by visual (44%) or auditory (51%) stimuli explains why the net effect of interacting the stimuli was usually inhibitory.
The second type of inhibitory response that occurred when the visual and auditory stimuli were paired was a suppression of the response, without IPSP's being evident. An example of this type of response is illustrated in Fig. 3. This unit did not respond to auditory or visual stimuli when presented alone (traces 1, 2, 3 and 6), however, when the visual and auditory stimu!i were paired at e A t - 20--50 msec (traces 3 and 4) the unit's response was suppressed without evidence of summated IPSP activity. One explanation for this effect is that the com, ergence of the inhibition occurred at cell prior to the recorded cell and that the excitatory sustaining input from this cell was depressed. This type of interaction is in contrast to the active inhibition displayed by the unit response illustrated in Fig. 2.

146
TABLE I
CLASSIFICATION OF UNITS ON THE BASIS OF RESPONSIVENESS TO VISUAL AND AUDITORY STIMULI
Responds only to Visual Stimuli 12 (12%) Responds only to Auditory Stimuli 16 (16%)
sponds to Visual and Auditory ~imuli 48 (49%) Unresponsive 22 (23%)
T h e aud i to ry-v i sua l i n t e rac t ions t h a t resu l ted in a n e t i nh ib i t i on o f t h e un i t ' s r e sponse was b y far t he m o s t c o m m q n t y p e seen. O t h e r t y p e s o f i n t e r a c t i o n s were e n c o u n t e r e d ; in fact , t h e r e w e ~ so m a n y ca tegor ies o f i n t e r a c t i o n s t h a t no individual c a t e g o r y o f r e sponse , exc lud ing t h e ca tegor ies m e n t i o n e d above ,
3
%( |000 m~ec)
b
-400 0 400 T Ira,e,=)
• i ~ .... i ~IIIII i~ ii iiiii ~ i ~I~III i i •
~ ~ • i i I II~II Ii ~I~I iii iii i
• i . ~ =2o.,,,, 2oomwc
Fig. 3. Spike count function (above)and intracellular recordings at selected A t's (below) . . . . . . . . . . . . , 50 ;
do not ~t •--50 l t ion ,
147
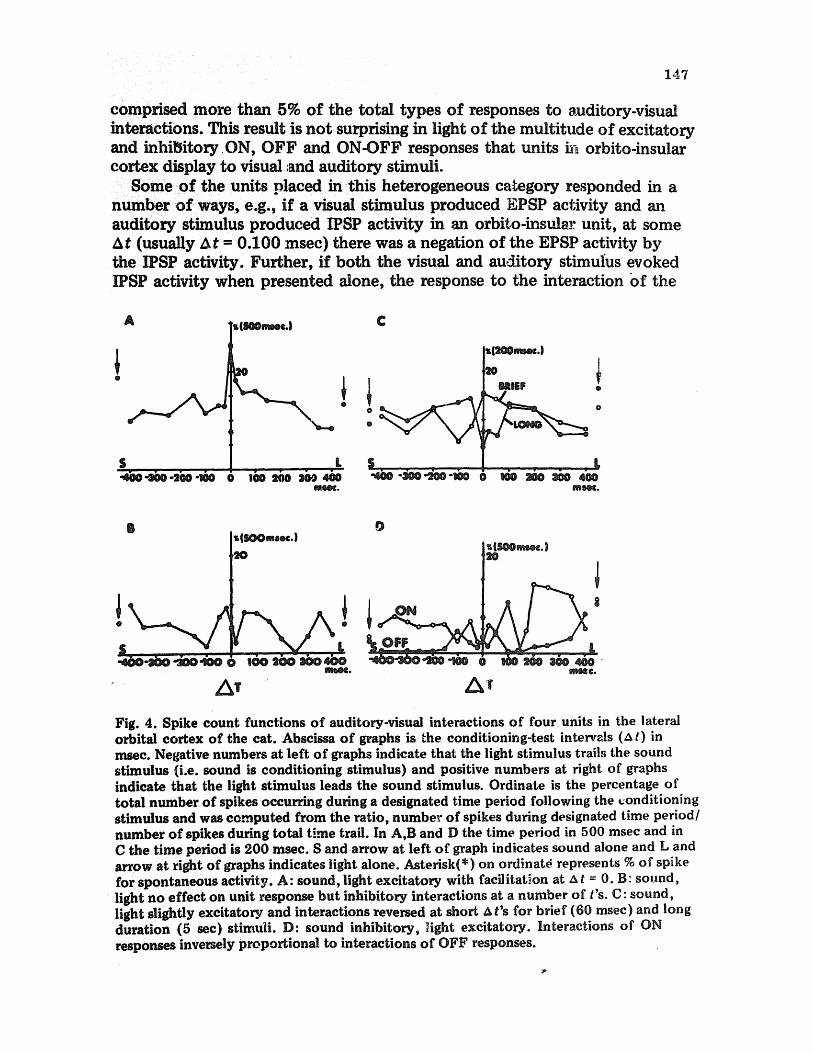
comprised more than 5% of the total types of responses to auditory-visual interactions. This result is not surprising in light of the multitude of excitatory and inhibitory ON, OFF and ON-OFF responses that units in orbito-insular cortex display to visual and auditory stimuli.
Some of the units placed in this heterogeneous caLegory responded in a number of ways, e.g., if a visual stimulus produced EPSP ac~;ivity and an auditory stimulus produced IPSP activity in an o r b i ~ o - i n s ~ unit, at some At (usually At = 0.100 msee) there was a negation of the EPSP activity by the IPSP activity. Further, if both the visual and auditory stimulus evoked IPSP activity when presented alone, the response to the interaction of the
A ,LIS~,,Mec.)
! o !
lWI~o
C
°
-300-'a)0-100 0 100 200 ~ 400 ~ .
B
t ~:)SO0,,,,®c.)
~ o
OFF
Fig. 4. Spike count functions of auditory-visual interactions of four units in the lateral orbital cortex of the cat. Abscissa of graphs is Lhe conditioning-test intervals (4 t) in msec. Negative numbers at left of graphs indicate that the light stimulus trails the sound stimulus {i.e. sound is conditioning stimulus) and positive numbers at right of graphs indicate that the light stimulus leads the sound stimulus. Ordinate is the percentage of total number of spikes occurring during a designated time period following the ~onditioning stimulus and was computed from the ratio, number of spikes during designated time period/ number of spikes during total time trail. In A,B and D the time period in 500 msec and in C the time period is 200 msec. S and arrow at left of graph indicates sound alone and L and arrow at right of graphs indicates light alone. Asterisk(*) on ordinat~ represents % of spike for spontaneous activity. A: Sound, light excitatory with facilitation at A t = 0. B: sound, light no effect on unit response but inhibitory interactions at a number of t's. C: sound, light slightly excitatory and interactions reversed at short ~ t's for brief (60 msec) and long duration (5 sec) stimuli. D: sound inhibitory, ~ight excitatory. Interactions of ON responses inversely proportional to interactions of OFF responses.

auditow,vis~ infraction could be variable at a number of At's, witha net facilitafi0n 0r suppression of response(Fig' 4B)i The responses could be variable ~d inve~ely related, depending on whether brief (60 msec)or longer
sec) duration st~ were U~ (Fig' 4C), or whether the ON responses and OFF respon~ were comp~ (Fig, 4D),
The finding that audit0ry-visual interactions occur at a wide range of At's and in such a heterogeneous fashion may reflect the difference in arrival of
along multiple afferent pathways having different functional synapses.
A number of th~o-cort i ca l and cortico~ortical pathways to the orbito- ins~ cortex have been previously described. Evidence was presented that auditory-v~ual convergence may occur prio~r to the cell from which the record- ings were made (Fig. 3) or on the cell in orbital-insular cortex itself (Fig. 2). One possible route whereby multimodal information reaches orbital-insular cortex is via the projection from the magnocellular medial geniculate nucleus [2,8,9,14] which is known to have cells with multimodal properties [16]. A convergence of visual and auditory input could also be relayed to the orbital- insular cortex via cortico~ortical projections [3,4,10,11,13,15].
REFERENCES
1 Benevento, L.A., Fallon, J.H. Davis, B. and Rezak, M., Auditory-visual interaction in single units in the cortex of the superior temporal sulcus and the orbital frontal cortex of the macaque monkey, (submitted for publication).
2 Benevento; L.A. and ~ 0 , P.R., An intracellular study of th~amocortical synapses in the orbito-insular cortex, Exp. Neurol., 46 (19"/5) 634--643.
3 Bignall, K.E., Effects of subcortical ablations on polysensory cortical responses and interactions in the cat, Exp. Neurol., i8 (1967) 56--6"/.
4 Bignall, K.E., Singer, P. and Herman, C., Interaction of cortical and peripheral inputs to polysensory areas of the cat neocortex, Exp. Neurol., 18 (1967) 194--209.
5 Fallon, J.H. and Benevento, L.A., Physiological and anatomical correlates of visual and auditory modalities in the inferior convexity of orbital cortex in rhzsus monkey, Soc. Neuroscience Abs., (1974) p. 193.
6 Fal~on, J,H. and Benevento, L.A.,: Extra~- and intracellular recordings in single units of the lateral olbital and adjacent cortices in the cat and rhesusmonkey, Soe, Neuroscience Abs, (19"/5) . . . .
7 Fallon, J.H. and Benevento, L.A.~0 Frequ~mcy'dependent inhibition to tones in the cat insular.cortex (AIV), (submitted for publication).

149
8 Graybiel, A.M., The thalamo-cortical projection of the so-called posterior nuclear group A study with anterograde degeneration methods in the cat, Brain Res., 49 (1973) 229-- 244.
9 Heath, C.J. and Jones, E.G., An experimental study of ascending connections from the posterior group of thalamic nuclei in the cat, J. comp. Neurol., 141 (1971) 397--426.
10 Imbert, M.~ Bignall, K.E. and Buser, P., Neocortical inte.rconnections in the cat, J. Neurophysiol., 29 (196S) 382--395.
11 Kawamura, K., Corticocortical fiber connections of the cat cerebrum. I. The temporal region, Brain Res., 51 (1973) 1--21.
12 Loe, P,R. and Benevento, L.A., Auditory-visual interaction in single units in the orbito- insular cortex of the cat, Electroenceph. clin. Neurophysiolo, 26 (1969) 395--398.
13 Paula-Barbosa, M.M., Feyo, P.B. and Sousa-Pinto, A., The association connections of the suprasylvian fringe (SF) and other areas of the cat auditory cm~tex, Exp. Brain Res., 23 (1975) 535--554.
14 Raczkowski, P., Diamond, I.T. and Winer, J., Organization of the thalarnocortical auditory system in the cat studied with horseradish peroxidase, Brain Res., 101 (1975) 345--354.
15 Thompson, R.F., Smith, H.E. and Bliss, D., Auditory, somatic sensory and visual response interactions and interrelationships in association and primary cortical fields of the cat, J. Neurophysiol., 26 (1962) 365--378.
16 Wespic, J.G., Multimodal sensory activation of cells in the magnocellular medial geniculate nucleus, Exp. Neurol., 15 (1966) 299--318.
17 Woolsey, C.N., Organization of cortical auditory system: A review and synthesis. In G.L. Rasmussen and W.F. Windle (Eds.), Neural Mechanisms of the Auditory and Vestibular Systems, Thomas, Springfield, Illinois, 1960, pp. 165--180.





























