Embed Size (px)
Citation preview

THE JOURNAL OF BKXOGICAL CHEMWIYW 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 17. Issue of June 15, pp. 9952-9959,199O Printed in U.S.A.
ATPlO, a Yeast Nuclear Gene Required for the Assembly of the Mitochondrial F1-F. Complex*
(Received for publication, December 26, 1989)
Sharon H. Ackerman and Alexander Tzagoloff From the Department of Biological Sciences, Columbia University, New York, New York 10027
A yeast nuclear gene (ATPlO) is reported whose product is essential for the assembly of a functional mitochondrial ATPase complex. Mutations in ATPIO induce a loss of rutamycin sensitivity in the mitochon- drial ATPase but do not affect respiratory enzymes. This phenotype has been correlated with a defect in the F0 sector of the ATPase. The wild type ATPIO gene has been cloned by transformation of an atpl0 mutant with a yeast genomic library. The gene codes for a protein of M, = 30,293. The primary structure of the ATPlO product is not related to any known subunit of the yeast or mammalian mitochondrial ATPase com- plexes.
To further clarify the role of this new protein in the assembly of the ATPase, an antibody was prepared against a hybrid protein expressed from a trpE/ATPlO fusion gene. The antibody recognizes a 30-kDa protein present in wild type mitochondria. The protein is as- sociated with the mitochondrial membrane but does not co-fractionate either with F1 or with the rutamy- tin-sensitive FI-F. complex. These data suggest that the ATPlO product is not a subunit of the ATPase complex but rather is required for the assembly of the F. sector of the complex.
The ATPase or F1-F, complex of bacteria, mitochondria, and chloroplasts is a multisubunit enzyme composed of two functionally and physically coupled parts. The catalytic por- tion, F1, consists of five non-identical subunits designated 01, 0, y, 6, and t (1). A second set of subunits is associated with the membrane sector, Fo, responsible for the proton translo- eating activity of the complex (2, 3). While the composition of F1 is fairly constant across phylogenetic lines that is not true of F,,. Bacterial F. is made up of only three subunits (1, 4). There are at least six different subunits in the Fo of Saccharomyces cerevisiae (5, 6), and as many as 11 subunits have been reported in the bovine Fo (7).
All the structural genes of the Escherichia coli ATPase are clustered in a single operon (1, 4). This is also true of several other bacterial ATPases (8, 9). In the case of the mitochon- drial complex, the same genes are distributed between the nuclear and mitochondrial genomes. The most hydrophobic subunits of yeast F0 are encoded by mitochondrial DNA (lo- 12). All the F, subunits including CY (13) and fi, (14) and some
* This research was supported by Research Grant HL22174 and National Research Service Award GM12435 from the National Insti- tutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(.s) reported in thispaper has been submitted to the GenBank”‘“/EMBL Data Bank with accession number(s) 505463.
of the more hydrophilic subunits of Fo such as subunit 4 (15) and the oligomycin sensitivity-conferring protein (16), are derived from nuclear genes. These constituents are synthe- sized on cytoplasmic ribosomes, usually as precursors with amino-terminal extensions (13, 14, 17, 18), and are trans- ported into mitochondria where they interact with their part- ner subunits to form the oligomeric enzyme. The assembly of the ATPase and of other mitochondrial inner membrane enzymes is still not well understood. Because of the experi- mental difficulty in demonstrating intermediates in the as- sembly pathway, very little can be said about the temporal sequence in which the subunits interact to form the holoen- zyme. While there is little doubt that protein-protein recog- nition must be a dominant factor in determining assembly of the subunits, the process may also depend on accessory pro- teins that guide the interaction of partner subunits.
To learn more about the mechanism of synthesis of the ATPase we have screened a collection of respiratory-deficient pet’ mutants of S. cereuisiae for defects in the mitochondrial ATPase. We reasoned that the synthesis of the active mem- brane complex would depend not only on the expression of the genes coding for the subunits of F1 and Fo, but also on other proteins that might have a function in assembly of the enzyme. Mutations in the latter class of genes would be expected to express either a deficiency or alteration in the catalytic properties of the ATPase.
In the present article we report a group of mutants (com- plementation group G165) impaired in the assembly of the F1-F. complex. The phenotype of G165 mutants suggests that this gene codes for a protein which is essential for the synthe- sis of a functional F0 unit. The gene, henceforth referred to as ATPlO, has been cloned and characterized. The deduced primary structure of the encoded protein is not homologous to any of the known subunits of the F1-F. complex nor does it co-purify with the complex. Taken together our data indi- cate that the ATPlO product is not a subunit of Fo but rather affects its synthesis or assembly.
MATERIALS AND METHODS
Yeast Strains and Growth Media-The strains of S. cereuisiue used in this study are described in Table I. The atpI mutant El03 is one of a large number of pet mutants derived from the respiratory com- petent haploid yeast D273-lOB/Al by mutagenesis with ethyl meth- anesulfonate (21).
The following tnedia were used for growth of yeast: YPD (2% glucose, 2% peptone, 1% yeast extractj; YEPG (3% glycerol, 2% nentone. 1% veast extract): YPGal (2% galactose, 2% peptone, 1% yeast extract):, WO (2% glucose, 0.67% nitrogen base without amino acids (Difco)). Where required media were supplemented with auxo- trophic requirements at 20 rg/ml. Solid media contained 2% agar.
’ The abbreviations used are: pet, nuclear respiratory-deficient mutant of S. cereuisiae; pO, cytoplasmic petite mutant lacking mito- chondrial DNA; kb, kilobase pair(s); SDS, sodium dodecyl sulfate; bp, base pair.
9952
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from
by guest on February 13, 2019
http://ww
w.jbc.org/
Dow
nloaded from
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Assembly of Yeast Mitochondrial F1-Fo
TABLE I Genotypes and sources of S. cerevisiae strains
Strain Genotype
D273-lOB/Al cu,p+,met6 D273-lOB/Alp’ cu,p0,met6 CBllp” a,p’,adel W303-1A ol,p+,ade2-1,hi.s3-11,15,1eu2-3,112,ura3-1,trp1-1 W303-1B a,p’,ade2-1,his3-11,15,1eu2-3,112,ura3-1,~rp1-1 W303VATPlO a,p+,ade2-l,his3-ll,15,leu2-3,ll2,ur~-l,~rpl-l,a~pl~:LEU2 aW303VATPlO cY,p+,ade2-l,his3-ll,15,leu2-3,ll2,ur~-l,~rpl-l,a~pl~:LEU2 El03 cu,p+,met6,atplO-1 B103 a,p+,adel,atplO-1 E103/Ul a,p+,ura&l,atplO-1
(1 R. Rothstein, Department of Human Genetics, Columbia University, New York, NY.
SOUrCeS
Ref. 19 Ref. 19 Ref. 20 -a -0 This study This study This study This study El03 X W303-1A
9953
Preparation of Yeast Mitochondria and Submitochondrial Particles and Purification of Yeast F1-Yeast was grown aerobically in YPGal media at 30 “C to early stationary phase. In most of the experiments mitochondria were prepared by lysis of yeast spheroplasts according to the procedure of Faye et al. (22) except that Zymolyase 20,000 (Miles Corp.) instead of Glusulase was used for the conversion of cells to spheroplasts. The post-mitochondrial supernatant fractions were clarified of membrane components by centrifugation in a Beck- man 50Ti rotor for 30 min at 40,000 rpm. To assay their ATPase activity, the post-mitochondrial supernatants were dialyzed against 20 mM Tris-Cl, pH 7.5, 2 mM ATP, 0.5 mM EDTA at room temper- ature to remove inorganic phosphate. Alternatively, mitochondria were isolated from yeast cells disrupted with glass beads in a Braun homogenizer (23). The temperature at which mitochondria were prepared is specified for each experiment. Phenylmethylsulfonyl flu- oride was added at a final concentration of 10 pg/ml to the crude homogenate to minimize proteolytic degradation.
Submitochondrial particles were prepared by irradiation of a 3-ml suspension of mitochondria (lo-15 mg/ml) with a single 5-s burst with a Braunsonic 1510,400W sonicator. The suspension was centri- fuged for 30 min at 50,000 rpm and the pellet consisting of the submitochondrial particles was washed once in 0.25 M sucrose, 10 mM Tris acetate, pH 7.5, and suspended at 20 mg/ml in the same buffer.
Wild type yeast F1-ATPase with a specific activity of 20 units/mg of protein was purified from commercial baker’s yeast (Red Star) as described previously (24).
Sucrose Gradient Centrifugation Analysis of F1 Proteins-Mito- chondria prepared at room temperature by the Zymolyase procedure were suspended at a protein concentration of 7-15 mg/ml in a buffer solution containing 10 mM Tris-Cl, pH 8, 2 mM ATP, and 1 mM EDTA. The suspensions were sonically irradiated for 30 s and were clarified of membranes by centrifugation at 50,000 rpm in a Beckman 50Ti rotor for 30 min. The supernatant (0.6 ml) was loaded on a 4.4. ml column of a 6-20% linear sucrose gradient containing 10 mM Tris- Cl, pH 8, 2 mM ATP, 1 mM EDTA, and 0.1% Triton X-100. The gradient was centrifuged at room temperature in a Beckman SW65 Ti rotor for 3 h at 65,000 rpm. Ten fractions (0.5 ml) were collected and the position of the ATPase subunits in the gradient was deter- mined immunochemically.
Cloning of the ATPlO Gene-The ATPlO gene was cloned by transformation of E103/Ul (Lu,ura&l,atplO-1) with a genomic library consisting of partial Sau3A fragments (5-15 kb) of yeast nuclear DNA ligated to the BamHl site of YEp24 (25). This library was kindly provided by Dr. Marian Carlson, Department of Human Genetics, Columbia University. Approximately 3 x 10’ cells were transformed with 5 pg of DNA by the procedure of Beggs (26). Transformants complemented for both the uracil auxotrophy and respiratory deficiency were selected on minimal glycerol medium. One clone was obtained whose transformed phenotype was ascertained to be due to an autonomously replicating plasmid. This plasmid (pG165/ Tl) was used to subclone and characterize the ATPIO gene.
Preparation of Antibodies to the ATPlO Gene Product and to F,- A 465-bp fragment of DNA containing a portion of the ATPlO reading frame was ligated in-frame to the amino-terminal half of the E. coli trpE gene in the expression vector pATH20 (27). A transformant of E. coli harboring this construct was used to express the fusion protein as described previously (28). Following cell lysis, the fusion protein was recovered in the insoluble fraction, and further purified by chromatography on Bio-Gel A-O.5 in the presence of 0.1% sodium
dodecyl sulfate (SDS) and 5 mM P-mercaptoethanol. The partially purified protein (l-2 mg) was solubilized in 0.2% SDS, 1 mM B- mercaptoethanol, and injected into rabbits as an emulsion with com- plete Freund’s adjuvant. A booster of 0.5-l mg of protein in incom- plete Freund’s adjuvant was administered after 3 weeks. Serum was collected a week after the second inoculation over a period of 3 weeks. The same protocol was used to raise antibodies against purified yeast F, and the individual o( and p subunits of F, (24). Antiserum against subunit 4 of the ATPase complex was a generous gift from Dr. Jean Velours, Institut de Biochimie Cellulaire et Neurochimie, CNRS, Bordeaux, France.
Miscellaneous Procedures-Standard techniques were used for re- striction endonuclease analysis of DNA, purification and ligation of DNA fragments, transformation of and recovery of plasmid DN.4 from E. coli, and nick translation of DNA (29). Southern hybridiza- tion was done under the conditions of Myers et al. (30). DNA was sequenced by the method of Maxam and Gilbert (31). For Western analysis, proteins were separated on 10, 12, or 15% polyacrylamide gels run in the electrophoretic system of Laemmli (32) except that the separation buffer was adjusted to pH 8 and the composition of the running buffer was 0.05 M Tris, 0.38 M glycine, and 0.1% SDS. Following electrophoretic transfer of the proteins to nitrocellulose, the blot was processed as described by Schmidt et al. (33). ATPase activity was measured by the calorimetric determination of inorganic phosphate released from ATP (34). A unit of activity is defined as the amount of enzyme which hydrolyzes 1 pmol of ATP/min under the conditions specified. Protein was estimated by the method of Lowry et al. (35). The in vivo incorporation of [35S] methionine into the mitochondrial translation products of wild type and atpI mu- tants was kindly performed by J. Mulero and T. Fox at Cornell University.
RESULTS
Phenotype ofatpl0 Mutants-The ATPase mutants (a@10 mutants) reported here were selected from a pet strain collec- tion by screening representatives from 180 different comple- mentation groups for lesions in mitochondrial ATPase. For these biochemical analyses pet mutants were fractionated to yield mitochondrial and post-mitochondrial supernatant frac- tions both of which were assayed for total and rutamycin- sensitive ATPase. These assays revealed that mutants in some dozen complementation groups were defective in mitochon- drial ATPase. The most common phenotype found is de- scribed by mutants exhibiting substantially reduced levels of ATPase activity in the mitochondrial fraction. One comple- mentation group (G165), however, had a somewhat different phenotype. The mitochondrial ATPase activity was about 50% of the wild type level and the enzyme was only partially inhibited by rutamycin. This latter property of the mutant ATPase was of interest because rutamycin is known to block proton translocation by the F. sector and to inhibit ATPase activity only when F1 is functionally coupled to F,. In other respects the mutant mitochondria had normal properties as indicated by the presence of wild type levels of respiratory chain components such as cytochromes a, b, and c and the in
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from
Assembly of Yeast Mitochondrial FI-Fo
uiuo incorporation of radioactive methionine into the full complement of mitochondrial translation products.
The pet complementation group G165 is composed of three independent isolates of which El03 was the most exten- sively analyzed. Additional mutants, W303VATPlO and aW303VATP10, were constructed by in situ disruption of the ATPlO gene as described in a later section. Both El03 and the disrupted strains grow normally on glucose but grow very poorly on non-fermentable carbon sources such as glycerol. The respiratory defect of G165 mutants is complemented by a p” tester lacking mitochondrial DNA indicating that they have recessive mutations in a nuclear gene which we have designated ATPZO. El03 is genetically stable as evidenced by a low frequency of reversion to respiratory competence and accumulation of only IO-15% cytoplasmic petite derivatives upon long term subculturing.
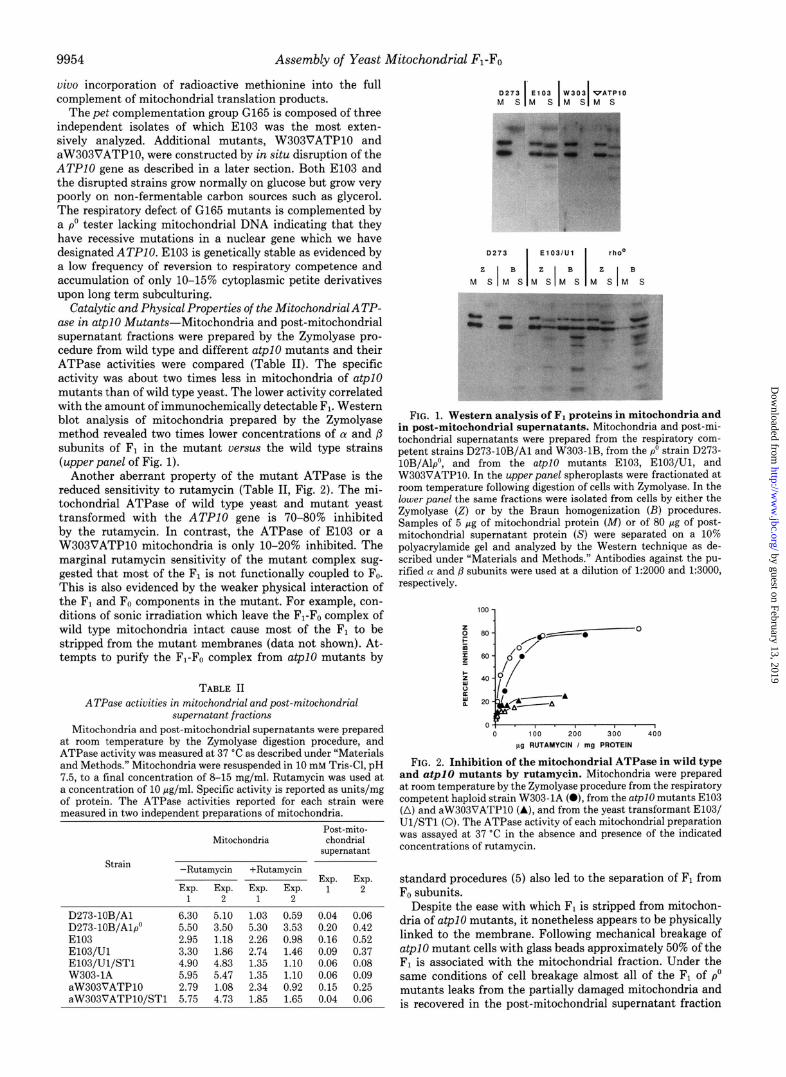
Catalytic and Physical Properties of the MitochondrialATP- ase in atpl0 Mutants-Mitochondria and post-mitochondrial supernatant fractions were prepared by the Zymolyase pro- cedure from wild type and different atpI mutants and their ATPase activities were compared (Table II). The specific activity was about two times less in mitochondria of atpI mutants than of wild type yeast. The lower activity correlated with the amount of immunochemically detectable Fi. Western blot analysis of mitochondria prepared by the Zymolyase method revealed two times lower concentrations of a and /3 subunits of F, in the mutant uersus the wild type strains (upper panel of Fig. 1).
Another aberrant property of the mutant ATPase is the reduced sensitivity to rutamycin (Table II, Fig. 2). The mi- tochondrial ATPase of wild type yeast and mutant yeast transformed with the ATPlO gene is 70-80% inhibited by the rutamycin. In contrast, the ATPase of El03 or a W303VATPlO mitochondria is only lo-20% inhibited. The marginal rutamycin sensitivity of the mutant complex sug- gested that most of the F, is not functionally coupled to Fo. This is also evidenced by the weaker physical interaction of the Fi and F. components in the mutant. For example, con- ditions of sonic irradiation which leave the Fi-F. complex of wild type mitochondria intact cause most of the F, to be stripped from the mutant membranes (data not shown). At- tempts to purify the Fi-F. complex from atpI mutants by
TABLE II
AZ’Pase activities in mitochondrial and post-mitochondrial supernatant fractions
Mitochondria and post-mitochondrial supernatants were prepared at room temperature by the Zymolyase digestion procedure, and ATPase activity was measured at 37 “C as described under “Materials and Methods.” Mitochondria were resuspended in 10 mM Tris-Cl, pH 7.5, to a final concentration of 8-15 mg/ml. Rutamycin was used at a concentration of 10 pg/ml. Specific activity is reported as units/mg of protein. The ATPase activities reported for each strain were measured in two independent preparations of mitochondria.
Post-mito- Mitochondria chondrial
supernatant Strain
-Rutamycin +Rutamycin Exp. Exp.
Exp. Exp. Exp. E~P. 1 2 1 2 1 2
D273-lOB/Al 6.30 5.10 1.03 0.59 0.04 0.06 D273-lOB/Al# 5.50 3.50 5.30 3.53 0.20 0.42 El03 2.95 1.18 2.26 0.98 0.16 0.52 E103/Ul 3.30 1.86 2.74 1.46 0.09 0.37 E103/Ul/STl 4.90 4.83 1.35 1.10 0.06 0.08 W303-1A 5.95 5.47 1.35 1.10 0.06 0.09 aW303VATPlO 2.79 1.08 2.34 0.92 0.15 0.25 aW303VATPlO/STl 5.75 4.73 1.85 1.65 0.04 0.06
FIG. 1. Western analysis of F1 proteins in mitochondria and in post-mitochondrial supernatants. Mitochondria and post-mi- tochondrial supernatants were prepared from the respiratory com- petent strains D273-lOB/Al and W303-lB, from the p0 strain D273- 10B/Alp”, and from the atpI0 mutants E103, E103/Ul, and W303VATPlO. In the upper panel spheroplasts were fractionated at room temperature following digestion of cells with Zymolyase. In the lower panel the same fractions were isolated from cells by either the Zymolyase (2) or by the Braun homogenization (E) procedures. Samples of 5 pg of mitochondrial protein (M) or of 80 pg of post- mitochondrial supernatant protein (S) were separated on a 10% polyacrylamide gel and analyzed by the Western technique as de- scribed under “Materials and Methods.” Antibodies against the pu- rified (Y and p subunits were used at a dilution of 1:2000 and 1:3000, respectively.
100 1 p 80 0
m /a-===-
z 60 0. /O/
2 40
z ;
L 0 I / I
20 0 /s----A A-A
OT I . I I 0 100 200 300 400
pg RUTAMVCIN I mg PROTEIN
FIG. 2. Inhibition of the mitochondrial ATPase in wild type and a@10 mutants by rutamycin. Mitochondria were prepared at room temperature by the Zymolyase procedure from the respiratory competent haploid strain W303-lA (O), from the atpI mutants El03 (A) and aW303VATPlO (A), and from the yeast transformant E103/ Ul/STl (0). The ATPase activity of each mitochondrial preparation was assayed at 37 “C in the absence and presence of the indicated concentrations of rutamycin.
standard procedures (5) also led to the separation of F1 from F0 subunits.
Despite the ease with which F1 is stripped from mitochon- dria of atpI mutants, it nonetheless appears to be physically linked to the membrane. Following mechanical breakage of atpI mutant cells with glass beads approximately 50% of the F, is associated with the mitochondrial fraction. Under the same conditions of cell breakage almost all of the Fi of p” mutants leaks from the partially damaged mitochondria and is recovered in the post-mitochondrial supernatant fraction
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Assembly of Yeast Mitochondrial FI-F. 9955
(lowerpanel of Fig. 1). Because p” mutants lack mitochondrial DNA they do not synthesize F. and F1 exists in a soluble and rutamycin-insensitive form in the matrix compartment (36). The mitochondrial localization of Fi in p” mutants is evident when mitochondria are prepared by the less damaging Zym- olyase procedure (Table II, Fig. 1). The fact that the content of Fi in mitochondria of a@lO mutants is about the same independent of their method of preparation indicates that the enzyme is part of a larger membrane complex.
It is interesting to note that a substantial fraction of F1 in both p” and at@0 mutants is recovered in the post-mitochon- drial supernatant fraction even when mitochondria are pre- pared by the Zymolyase method (Table II, Fig. 1). The most reasonable interpretation of these observations is that some F, leakage from mitochondria occurs in both types of mutants during the fractionation procedure. Western blot analyses of the extra-mitochondrial Fi reveal the LY subunit to be slightly smaller in size than the normal mature subunit. The disparity in the size of the 01 subunit is probably caused by proteolytic cleavage of the protein by a cytoplasmic protease. Since the ATPase activity assayed in the post-mitochondrial superna- tant correlates with the amount of immunologically measur- able F1, the proteolytic modification of the 01 subunit must not have any appreciable effect on the hydrolytic activity of the enzyme.
Characterization of F, in atpl0 Mutants-The absence of functionally coupled ATPase in atpI mutants could be a consequence of either defective F1 or Fo. The following evi- dence excludes F, from being responsible for the altered properties of the ATPase. Catalytically active F, extracted by sonic irradiation of El03 mitochondria has the same sedimen- tation properties as the wild type enzyme (Fig. 3) indicating that the atpl0 mutation does not affect assembly of the Fi subunits into a functional oligomeric enzyme. Nor does the mutation appear to affect the total ATPase activity. The ATPase units measured in the mitochondrial and post-mito- chondrial supernatant fractions were 20 and 3, respectively, for the wild type strain D273-10B and 12 and 9, respectively,
UP 12345678910
FIG. 3. Sedimentation properties of F, from wild type and an atpl0 mutant. Mitochondria were prepared at room temperature by the Zymolyase procedure from the atpI0 mutant El03 and from the parental wild type strain D273-lOB/Al. F, was extracted by sonic irradiation of mitochondrial membranes and centrifuged through a 6-20% linear sucrose gradient as described under “Materials and Methods.” The gradients were collected in 10 equal fractions and analvzed for the distribution of the N and fl subunits of F, as described in the legend to Fig. 1. Fraction 1 corresponds to the most dense portion (bottom) of the gradient. Panel A, gradient fractions of the F, extracted from wild type membranes. The two left most lanes were loaded with 0.5 pg of purified (Y and (I subunit as standards. Panel B, gradient fractions of the F, extracted from the mutant membranes. To the leftmost lane was applied 2.5 pg of wild type mitochondria (0273 MITO).
for the mutant E103. The sum of these vaiues, 23 for the wild type strain and 21 for the mutant, are almost identical sug- gesting that there is no appreciable decrease in enzymatically active F, in the mutant.
A third line of evidence indicating that the F, component is not affected by mutations in ATPlO was obtained from reconstitution experiments. The reconstituted ATPase was equally sensitive to rutamycin (70-80% inhibition) when par- tially purified F1 of wild type or El03 was added to wild type membranes depleted of their F, by extraction with NaBr (37).
Reconstitution of Rutamycin-sensitive ATPase in Wild Type and Mutant Membranes-The presence in El03 of F, with physical and functional properties indistinguishable from the wild type enzyme suggested that the ATPase defect of G165 mutants is due to an altered F. component. This was con- firmed by the results of the reconstitutions summarized in Table III. In these experiments wild type F1 was used to reconstitute rutamycin-sensitive ATPase in wild type and mutant membranes depleted of F,. Western analysis of the NaBr-extracted membranes prepared from both strains indi- cated that less than 10% of F1 protein remained bound to the membrane following treatment with NaBr (data not shown). This is in accord with the observed near complete absence of ATPase activity in NaBr-extracted membranes. In two inde- pendent experiments the addition of purified F1 to NaBr- extracted membranes restored 30-40% of the starting ATPase activity of the particles (Table III). The ATPase reconstituted in the wild type mitochondrial membranes displayed the same rutamycin sensitivity as the original particles while the ATP- ase reconstituted in the mutant membranes was only partially sensitive to the antibiotic (Table III). All manipulations sub- sequent to the incubation of F, with the membranes were done at 4 “C. Since soluble F1 is inactivated at 4 “C (38) these experiments, which measure only the activity of membrane- bound enzyme, unambiguously show that the defect in atpl0 mutants affects specifically the F. component of the ATPase.
Cloning and Sequence Determination of ATPlO-To learn
TABLE III
Reconstitution of ATPose activity to FL-depleted membranes Mitochondria were prepared at 4 “C by Braun homogenization as
described under “Materials and Methods” and suspended in 0.25 M sucrose, 10 mM Tris acetate, pH 7.5. F1 was extracted from the wild type (D273-lOB/Al) and the mutant (E103) mitochondria with 3 M NaBr. The extracted membranes were mixed with the indicated amounts of purified wild type F, as previously described (37). The reconstituted membranes were isolated by centrifugation, suspended in sucrose Tris, and assayed for ATPase activity as described in the leaend to Table II. The suecitic activities of the D273-lOB/Al and th‘k El03 mitochondria measured prior to NaBr extraction were 5.40 and 0.77 units/mg, respectively. The results of two separate experi- ments are shown.
Specific activities Reconstitution
Reconstitution -Rutamycin +Rutamycin mixture Exp. Exp.
Exp. Exp. Exp. EXP. 1 2 1 2 1 2
Wild type NaBr membrane
+O.l mg/ml F, +0.3 mg/ml F, +0.6 mg/ml F,
Mutant NaBr mem- brane
+O.l mg/ml F, +0.3 mg/ml F, +0.6 m&/ml F,
% 0.05 0.04 0.03 0.06 0.0 0.0
1.30 1.31 0.34 0.16 24 24 2.10 1.70 0.53 0.20 39 31
1.67 0.22 31
0.02 0.06 0.02 0.05 0.0 0.0
0.20 0.19 0.13 0.08 26 25 0.29 0.29 0.20 0.20 38 38
0.29 0.23 38
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

9956 Assembly of Yeast Mitochondrial Fl-F.
more about the nature and function of the ATPlO product, the gene was cloned by complementation of the a@10 mutant E103/Ul. Transformation of this mutant with a yeast ge- nomic library yielded several respiratory competent clones which were also complemented for the uracil auxotrophy. The respiratory competence and uracil independence were ascer- tained to be a function of the presence of an autonomously replicating plasmid as evidenced by the co-segregation of the Gly’ and Ura+ phenotypes among vegetative progeny of the transformant ElOS/Ul/Tl grown under non-selective condi- tions on rich glucose medium. In addition, plasmid DNA (pG165/Tl) recovered from this clone was able to rescue the respiratory defect of E103/Ul.
A partial restriction map for pG165/Tl indicates this re- combinant plasmid to have a nuclear DNA insert of approx- imately 8 kb (Fig. 4). Different regions of the nuclear DNA insert were transferred to the yeast/E. coli shuttle vector YEp352 (39) and the resultant constructs were tested for their ability to confer respiratory competence to E103/Ul. The results of the subcloning experiments indicated that the small- est region of DNA capable of complementing the atpI0 mu- tation is defined by a 2.2-kb SmaI-BglII fragment spanning the unique SstI site of pG165/ST9.
The DNA insert of pG165/ST9 was sequenced by the method of Maxam and Gilbert (31). All of the restriction sites used for 5’-end labeling were crossed from distal sites and most of the sequence was confirmed from the complementary strands by the strategy shown in Fig. 5. The sequence revealed the presence in the insert of a 792-bp open reading frame capable of coding for a protein with a molecular weight of 30,293. The sequence of this putative gene, which we propose to be ATPIO, starts with an ATG initiation codon at nucleo- tide +l and ends with an opal termination codon at nucleotide +793 of the sequence reported in Fig. 6. Two other reading frames were found in the complementary strand of the pG165/ ST9 insert. Neither of these, however, contained the SstI restriction site found to be essential for complementation of the mutant (Fig. 4). The putative secondary structure of the
ST9 +
ST5 -
1.0 kb -
ST3 -
ST2 -
ST1 +
SE Bsea sp ESp S
FIG. 4. Restriction maps of pG165/Tl and derivative plas- mids. A partial restriction map of the nuclear DNA insert of pG165/ Tl is shown above the circular map of YEp24. The bars in the upper part of the figure denote regions of the insert in pG165/Tl subcloned in the yeast/E. coli shuttle vector YEp352. Complementation of the atpI mutation in E103/Ul is indicated by the plus signs and lack of complementation by the minus signs. The location of the A’I’PlO gene is indicated by the solid bar in the insert of pG165/Tl. The restriction sites for SstI (S), EcoRI (E), EglII (Bg), SphI (Sp), and SmaI (Sm) have been marked.
1.0 kb t i
ATP 10 URF 1 URFZ ( j-------)
-I
FIG. 5. Sequencing strategy. The restriction sites used for 5’- end labeling and the approximate extents of the sequences obtained are indicated by the arrows in the lower part of the figure. The locations of the ATPI gene and of the two open reading frames are indicated by the open and solid bars, respectively. The restriction sites are denoted by the following symbols: Bg111 (+), TuqI (A), Hue111 (A), EcoRI (O), Hind111 (m), XbaI (tl), SstI (0).
ATPlO product as predicted by the procedures of Chou and Fasman (40) and Garnier et al. (41) indicates that the first 24 amino acids form an amphipathic (Y helix. This domain con- tains 33% basic and hydroxylated residues consistent with the notion that this sequence is a mitochondrial import signal (42).
Construction of an atpl0 Deletion Allele-To confirm that the ATPlO gene had been correctly identified, the chromo- somal copy of the gene was disrupted by a one-step gene replacement method (43) in the respiratory competent haploid strains W303-IA and W303-1B. The construction of the deletion allele is shown in the upper part of Fig. 7. A 900-bp Hind111 DNA fragment containing the region coding for the carboxyl-terminal two-thirds of the protein was transferred to the Hind111 site of YEp352H (this plasmid is similar to YEp352 except that the multiple cloning region of the latter plasmid has been replaced with a unique Hind111 site). The resultant plasmid was digested with a combination of SstI and XbaI to remove a 30-bp fragment internal to the ATPlO reading frame. The gapped plasmid was ligated to a 3-kb SstI- XbaI fragment containing the yeast LEU2 gene to yield the disrupted allele, atplO::LEU2.
The above plasmid was used to isolate a linear 3.9-kb Hind111 fragment containing the disrupted gene and 3’-flank- ing sequences. Following transformation of the respiratory competent haploid yeast strains W303-1A and W303-1B with the linear Hind111 fragment, clones were selected on minimal glucose medium supplemented with all the auxotrophic re- quirements of the W303 strains except leucine. Most of the leucine-independent transformants were respiratory defective and were complemented by a p” tester strain but not the original mutant (E103, B103). The results of these crosses confirmed the genetic linkage of the disrupted atplOxLEU2 allele to the c&p10 mutation of E103.
Southern hybridization analysis of the genomic DNA from two Gly-,Leu+ transformants (W303VATPlO and aW303VATPlO) confirmed the successful substitution of the wild type gene by the disrupted allele. In these analyses, wild type and mutant DNAs were digested with EcoRI alone and with a combination of EcoRI and KpnI and hybridized to a nick-translated 465-bp EcoRI-SstI fragment internal to the ATPlO gene. The probe detects a 1.6kb band in the EcoRI- KpnI digest and a 2.2-kb band in the EcoRI digest of the mutant DNA (Fig. 7). The sizes of these fragments are con- sistent with the restriction map of the atplO::LEU2 allele.
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

FIG. 6. Nucleotide sequence of ATPIO. The entire sequence of the insert of pG165/ST9 is shown. The 792-bp reading frame identi- fied as the ATPIO gene starts with the ATG codon at +1 and ends with an opal termination codon at nucleotide +793. The amino acid sequence of the encoded protein is shown above the nucleotide se- quence. The sequences of the putative proteins encoded by the two open reading frames are shown under the nucleotide sequence. Only the sense strands of the coding regions are reported. Both DNA strands are reported for the region between ATPIO and the two open reading frames. The locations of the BglII, S&I, EcoRI, and Hind111 sites have been marked for reference.
The smaller 1.4-kb band detected in the two digests of wild type DNA corresponds to a fragment extending from the EcoRI site inside the ATPlO reading frame to an unmapped EcoRI site downstream of the gene.
Localization of the ATPlO Product in Mitochondria-To
Mitochondrial F1-Fo
probe
Bs EHXS H II II ,
- w303
ATPlO -
ATPlO +
ECORl
1 2 3 4
4.3 kb -
9957
1.0 kb I I
K E S I I
.,_______ 2----- :‘: W303VATPlO n-------7
LElJ2 ATPlO -+ -
EcoFil and Kpnl
1 2 3 4
1.3 kb-
FIG. 7. Disruption of the ATPlO gene. The upper part of the figure shows the restriction maps of the wild type ATF’ZO and dis- rupted atpZO::LEU2 allele. The 3-kb fragment of DNA with the LEU2 gene is shown by the dashed lines. The coding regions of ATPlO and LEU2 are depicted by the solid and open bars, respectively, and the direction of transcription by the arrows. The locations of the BglII (Bg), EcoRI (E), Hind111 (H), XbaI (X), and SstI (S) sites are marked on the two maps. The lower part of the figure shows the results of the Southern analysis of wild type and mutant genomic DNA. Nuclear DNA was prepared from the Leu’ transformants W303VATPlO (lane I) and aW303VATPlO (lane 2), and from the two respiratory com- petent strains W303-1A (lane 3) and D273-lOB/Al (lane 4). The four DNAs were digested either with EcoRI alone or with a combination of EcoRI and KpnI. The blot was hybridized to the nick-translated 465-bp EcoRI-SstI fragment (probe) shown in the upper part of the figure. The migration of known size standards is shown in the left hand margin.
determine whether the protein encoded by ATPIO is a mito- chondrial protein, a polyclonal antibody was prepared against a hybrid protein consisting of residues 66-221 of the ATPlO product fused to the amino-terminal 321 residues of compo- nent I of anthranilate synthetase. The construction of the E. coli plasmid with the trpE/ATPlO fusion gene from which the protein was expressed is described under “Materials and Methods.”
The results of the Western analysis shown in Fig. 8 indicate that the ATPlO product is located in mitochondria. The antiserum to the hybrid protein detects a 30-kDa protein in wild type mitochondria and submitochondrial particles. The identity of this protein as the product of ATPlO was confirmed by its absence in aW303VATP10, a strain with the atplO::LEU2 allele, and overexpression of the protein in the transformant ElOS/Ul/STl harboring the ATPlO gene on a multicopy plasmid.
The fact that the ATPlO product is also detected in sub- mitochondrial particles prepared by a procedure which re- moves soluble matrix constituents suggests that it is associ- ated with the membrane. This conclusion was strengthened by the results obtained when mitochondria were extracted with different concentrations of deoxycholate. The ATPlO protein was not extracted with high concentrations (1 M) of
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

9958 Assembly of Yeast Mitochondrial F,-F0
12 3456
67 kDa -
43 kDa - II)
30 kDa - *“*- - M
FIG. 8. Western blot analysis of the ATPIO protein in yeast mitochondria and submitochondrial particles. Mitochondria were prepared at 4 “C by the Zymolyase method from the respiratory competent strains D273-lOB/Al and W303-IA, from the atplo mu- tant aW303VATP10, and from the transformant ElOS/Ul/STl har- boring the ATPIO gene on a multicopy plasmid. Following separation of proteins on a 12% polyacrylamide gel and transfer to nitrocellulose, the Western blot was reacted with a 1:lOO dilution of the antiserum against the trpE/ATPlO product. Lnne 1; 10 pg of D273-lOB/Al submitochondrial particles; lane 2, 20 pg of aW303VATPlO mito- chondria: lane 3. 20 LL~ of W303-IA mitochondria: lane 4: 20 uz of E103/Ul/STl nhtochondria; lane 5, 20 wg of D2’73-lOBjA1 &:to- chondria; lone 6, 1 pg of trpE/ATPIO fusion protein. The migration of known protein standards is indicated in the left hand margin.
salt alone but was extracted with a combination of 1 M salt and 0.5% deoxycholate (data not shown). These conditions are comparable to those needed for the extraction of mito- chondrial inner membrane components such as cytochrome oxidase and ubiquinol-cytochrome c reductase (44). Lower concentrations of deoxycholate (O.l-0.2%) which effectively disrupt mitochondria did not solubilize any appreciable amounts of the protein from the membrane. A Kyte-Doolittle hydrophobicity plot (45) of the ATPlO product is consistent with it being a membrane protein since there are two hydro- phobic domains near the carboxyl terminus which could serve as membrane spanning regions.
The ATPlO Product Is Not a Subunit of F,,-The absence in atpI mutants of functional F. combined with the hydro- phobicity of the ATPlO product suggested it might be a component of the F. sector of the ATPase. Several lines of evidence make this unlikely. First we were unable to detect the protein in preparations of t.he ATPase complex. For these analyses the ATPase was purified by extraction of submito- chondrial particles with Triton X-100 and size fractionation of the extract on a sucrose gradient (5). Even more telling, none of the ATPlO product is extracted from suhmitochon- drial particles with detergent under conditions which solubi- lize appreciable amounts of the rutamycin-sensitive ATPase. In the experiment of Fig. 9 approximately 30% of subunit 4 (an indicator of FJ but no detectable ATPlO product was extracted with 0.25% Triton X-100. In contrast, the ATPIO product is quantitatively extracted from wild type mitochon- dria with 3 M NaBr which leave an intact F. in the membrane (Fig. 9). These conditions are identical to those used to deplete wild type membranes of endogenous F, in the reconstitution experiments reported in Table III. The ATPlO encoded pro- tein, therefore, appears to be required for the synthesis rather than function of Fo.
DISCUSSION
To define the genetic information needed for the expression and assembly of the yeast mitochondrial F,-F. complex, we have screened a collection of pet mutants for lesions in this
FIG. 9. Extraction of the ATP4 and ATPlO products. Mito- chondria (MITO) and submitochondrial particles (SMP) were pre- pared from wild type strain D273-lOB/Al as described under “Ma- terials and Methods.” The submitochondrial particles were extracted with 0.25% Triton X-100 and centrifuged for 20 min at 50,000 rpm in a 50Ti rotor to yield pellet (TRITON MEMB) and supernatant (TRITON SUP) fractions. Mitochondria were extracted with NaBr as described in the legend to Table III. The resultant NaBr-extracted pellet (N&r MEMO) and NaBr extract (NaBr SUP) were dialyzed against 10 mM Tris acetate, pH 7.5, prior to gel electrophoresis. The pellets obtained after extraction with either Triton X-100 or NaBr were resuspended in a volume of 0.25 M sucrose, 10 mM Tris acetate, pH 7.5, corresponding to the starting particles and equal volumes of each pellet and supernatant were loaded on a 15% polyacrylamide gel. The amount of protein loaded per lane was 30 pg of either mitochondria or submitochondrial particles, or of the combined pellet and supernatant fractions. The blot in panel A was probed with a 1:lOO dilution of antibody against the ATPlO product and in panel B with a 1:lOOO dilution of antibody against the ATE’4 product.
inner membrane enzyme. The atpl0 mutants reported here were chosen for study because of the reduced ATPase activity of their mitochondria and the unusually poor inhibition of the enzyme by rutamycin. The 2-fold lower ATPase activity can be explained by the comparable decrease of immunologi- cally detectable F, in the mutant mitochondria. Despite the lower ATPase of mutant mitochondria, the total ATPase units measured in mitochondria and post-mitochondrial su- pernatant fractions is similar in atpI mutants and in wild type yeast.
The marginal inhibition of the ATPase by rutamycin sug- gests that atpl0 mutations affect the coupling of F1 to Fo. This conclusion is supported by other lines of evidence. Con- sistent with a lesion in functional coupling is the more fragile physical association of F, with the membrane sector. Thus conditions of sonic irradiation which fail to detach F1 from wild type mitochondrial membranes achieve substantial re- lease of F, from the mutant ATPase complex. The lability of the ATPase is also evidenced by our inability to detect a complex of F1 and F,, when submitochondrial particles were extracted with Triton and the extract sedimented through a sucrose gradient. The loose physical coupling of F, to F. in the mutant explains the recovery of 40-50% of F, in the post- mitochondrial fraction. Presumably this fraction of F1 repre- sents leakage of either a free or a weakly membrane-bound pool of intramitochondrial F1.
The rutamycin insensitivity and the lability of the complex as evidenced by the relative ease with which F, dissociates from the membrane could reflect defective F, and/or Fo. Since we were unable to detect any significant difference in the ATPase activity, size, and other properties of F1 from wild
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Assembly of Yeast Mitochondrial FI-Fo
type and from an atpl0 mutant it seems reasonably certain that atpI mutations exert their effect on the F. component. This is supported by the inability of sodium bromide-ex- tracted submitochondrial particles from atpl0 mutants to confer any appreciable degree of rutamycin sensitivity on wild type F,.
affecting the assembly of mitochondrial membrane enzymes is not without precedent. Recent evidence suggests that as- sembly of the subunit polypeptides of the yeast mitochondrial
The phenotype of atpl0 mutants is very similar to that reported for mutants lacking subunit 4 of the yeast ATPase complex (46) and is consistent with the notion that the biochemical lesion is confined to the F. component. To assess whether ATPlO codes for a subunit of F, or for a protein which affects the synthesis or assembly of a functional Fo without it necessarily being a constituent of the complex, the gene was cloned and its sequence determined. The primary structure of the protein deduced from the DNA sequence does not match any previously characterized subunit of the yeast or mammalian ATPase complex. This evidence, however, does not exclude the ATPlO product from being part of the enzyme since not all the nuclear-encoded subunits of the yeast mito- chondrial complex have been sequenced.
The localization and possible association of the ATPlO product with the F. sector of the yeast complex was further studied with an antibody prepared against a fusion protein of component I of E. coli anthranilate synthetase and the ATPlO product. The antibody reacts with a mitochondrial protein of 30 kDa, a size commensurate with the molecular weight predicted from the sequence of ATPIO. The fact that this protein is not detected in an atpl0 mutant with a disrupted copy of the gene and is more abundant in a transformant with the ATPlO gene on a multicopy plasmid confirms that the antibody recognizes the correct protein.
Even though the ATPlO protein is associated with the mitochondrial membrane several observations argue against it being a subunit of the F1-Fo complex. First, concentrations of Triton X-100 which effectively solubilize the FL-F0 complex from submitochondrial particles leave all the ATPlO product bound to the membrane. The antibody also fails to react with any protein in purified F1 or the rutamycin-sensitive complex. Conversely, high concentrations of sodium bromide quanti- tatively extract the ATPlO product even though the F. sector of the ATPase co-fractionates with the membrane fraction. In addition to pointing out the differences in the solubility properties of the ATPase complex and of the ATPlO encoded protein, this last observation shows that the ability of F. to bind and to confer rutamycin sensitivity on FL does not depend on the presence of the ATPlO product. The ATPlO product is most likely required during ATPase biosynthesis.
Several functions of the ATPIO product are compatible with the phenotype elicited by mutations in this gene. The protein could be a factor that enhances translation of a mitochondrially encoded subunit of Fo. Such translation fac- tors have been reported for cytochrome b (28,47) and for the mitochodrially derived subunits of cytochrome oxidase (48, 49). The mitochondrial genome of yeast codes for three sub- units of the ATPase (10-12) each of which is needed for the synthesis of a functional F. unit (12, 50, 51). Even though pulse labeling of the atplO mutant El03 in the presence of cycloheximide indicates a normal pattern of labeling of all the mitochondrial translation products, an effect of the mutation on translation of subunit 8 of the ATPase cannot be excluded because of the difficulty in detecting this mitochondrial trans- lation product. Alternatively, the ATPIO product could be important in assembling the subunits of F. by acting as a scaffolding factor or a chaperonin. The existence of proteins
ubiquinol-cytochrome c reductase requires the expression of at least two nuclear gene products (52, 53).
REFERENCES
1. Walker, d. E., Fearnley, I. M., Gay, N. J., Gibson, B. W., Northrop, F. D., Powell, S. J., Runswick, M. J., Saraste, M., and Tybulewicz, V. L. J. (1985) J. Mol. Biol. 184,677-701
2. c-,,,, P r ,,a*,\ d”.,,, Pm, Rin*hom r;n Lxl_,,,t 3.
824
“ I ”UO, Ah “ . ,‘Y”L,rl,rrru. . ,L”. YL”C, IC , * I . “ V I YYL ~I-7
Amzel, M. L., and Pedersen, P. L. (1983) Annu. Rev. Biochem. 52, 801-
eu. 47,285-312 em. 246.7328-7336
Biochem. Biophys. 184,464-475 ri-Sala, F., Friedl, P., and Galante, Y. M. (1983)
4. Futai, M., and Kanazawa, H. (1983) Microbial. RI 5. Tzagoloff, A., and Meagher, P. (1971) J. Biol. Ch. 6. Ryrie, I. J. (1977) Arch. 7. Montecuccq, C., Dabber
Eur. J. Bcochem. 132, 189-194 8. Ohta, S., Yohda, M., Ishizuka, M., Hirata, H., Hamamoto, T., Otawara-
Hamamoto, Y., Matsuda, K., and Kagawa, Y. (1988) Biochim. Biophys. Acta 933, 1 41-155
9. Brusilow, W. ! j. A., Scarpetta, M.iAibF3awthorne, C. A., and Clark, W. P. (1989) J. Biol. Chem. 264. 1528
., Borst, P., and Bos, J. L. (1979) Proc. 10. Hensgens, L. A. M., Grive1h.L. A Natl. Acad. Sci. U. S’. A. 76, 1663-1667
11. Macino, G., and Tzagoloff, A. (1980) Cell 20,507-517 12. Macreadie, I. G., Novitski, C. E., Maxwell, R. J., John, U., Ooi, B.-G.,
McMullen,, G. L., Lukins, H. B., Linnane, A. W., and Nagley, P. (1983) Nucleic Aads Res 11_44?&4451
13. Takeda. M.. Chen. - -, _ _ _ _ - .
W.-J., Salzgaber, J., and Douglas, M. G. (1986) J. Biol. !6-15133 Chmi261,15ii
14. Takeda. M.. Vassar
15.
16. 17.
18.
19. 20.
21.
22. 23. 24. 25.
Faye, G., Kujawa, C., and Fukuhara, H. (1974) J. Mol. Biol. 88, 185-203 Tzagoloff, A. (1971) J. Biol. Chem. 246.3050-3056 Tzagoloff, A. (1978) Methods Enzymol. LV, 351-358 Botstein, D., and Davis, R. W. (1982) in The Molecular Biology of the Yeast
Saccharomyces cereuisiae: Metabolism and Gene Expression (Strathern, J. N., Jones, E. W., and Broach, J. R., eds) pp. 607-636, Cold Spring Harbor, Cold Spring Harbor, NY
26. Beggs, J. D. (1978) Nature 275,104-109 27. Koerner, T. J., Myers, A. M., Hill, J., and Tzagoloff, A. (1990) Methods
Enzymol., in press 28. 29.
Dieckmann, C. L., and Tzagoloff, A. (1985) J. Biol. Chem. 260,1513-1520 Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular, Cloning: A
k&ratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor,
,otti. A.. and Douelas. M. G. (1985) J. Biol. Chem. 260.
us, J., Durrens, P., Aigle, M., and Guerin, B. (1988) Eur. J. Biochem. 170,637-642
Lee, M., Jones, D., and Mueller, D. M. (1988) Nucleic Acids Res. 16, 8181 Maccecchini, M.-L., Rudin, Y., Blobel, G., and Schatz, G. (1979) Proc. N&l.
Acad. Sri 11 S A 76. 343-341 twin, A.
. - -. . , i __ S.. Greeor. I.. Mason. T. L.. Nelson. N.. and Schatz. G. (1980)
Proc. Natl: Acad. .%i.‘U. S. A.‘77. %%Anni . , __-- _““- Tzaeoloff. A.. Akai. A.. and Fourv. F. (1976) FEBS L&t. 65,391-395 ten Berge: A.‘M. A.‘, Zdutewelle, 61, and Needleman, R. B. (1974) Mol. Gen.
Genet. 131, 113-121 Tzagoloff, A., Akai, A., and Needleman, R. B. (1975) J. Bacterial. 122,
826~831
30. Myers, A. M., Pape, L., K., and Tzagoloff, A. (1985) EMBO J. 4,2087-2092 31. M;;;m;,t. M., and Gilbert, W. (1977) Proc. Natl. Acad. Sci. U. S. A. 74,
32. Laemmli, U. K. (1970) Nature 227,680-685 33. Schmidt, R. J., Myers, A. M., Gillham, N. W., and Boynton, J. E. (1984)
Mol. Biol. Euol. 1, 317-334 34. King, E. J. (1932) Biochem. J. 26, 292-297 35. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J.
Biol. Chem. 193,265-275 36. Schatz, G. (1968) J. Biol. Chem. 243,2192-2199 37. Tzagoloff, A. (1970) J. Biol. Chem. 245.1545-1551 38. Penefsk 39. Hill J. I?
H. S., and Warner, R. C. (1965) J. Biol. Chen. 240,4694-4702
163-16; M yers, A. M., Koerner, T. J., and Tzagoloff, A. (1986) Yeast 2,
40. Chou! P. Y., and Fasman, G. D. (1978) Adun. Enzymol. 47,45-148 41. Ga9r;;m;nJ., Osguthorpe, D. J., and Robson, B. (1978) J. Mol. Btol. 120,
42. H&l, F.-U., Pfanner, N., Nicholson, D. W., and Neupert, W. (1989) Biochim. Biophys. Acta 988, l-45
43. Rothstein, R. J. (1983) Methods Enzymol. 101,202-211 44. Tzagoloff, A. (1969) J. Viol. Chem. 244,5020-5026 45. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157, 105-132 46. Paul, M.-F., Velours, J., Arselin de Chateaubodeau, G., Aigle, M., and
Guerin, 9. (1989) Eur. J. Biochem. 186, 163-171 47. Rod& C., and Fox, T. D. (1987) Mol. Gen. &net. 206‘45-50 48. Costanzo. M. C., Seaver, E. C., and Fox, T. D. (1986) EMBO J. 5, 3637-
%A 1 -_.- 49. Strick, C. A., and Fox, T. D. (1987) 1Mol. Cell. Biol. 7, 2728-2734 50. Foury, F., and Tzagoloff, A. (1976) Ew. J. Biochem. 68, 113-119 51. Coruzzi, G., Trembath, M. K., and Tzagoloff, A. (1978) Eur. J. Biochem.
92.279-287 52. Wu, M., and Tzagoloff, A. (1989) J. Biol. Chem. 264, 11122-11130 53. Crivellone, M., Gampel, A., Muroff, I., Wu, M., and Tzagoloff, A. (1987) in
Cytochrome Systems: Molecular Biology and Bioenergetics (Papa, S., C$tmn~or%, and Ernster, L., eds) pp. 67-77, Plenum Publishing Corp.,
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

S H Ackerman and A Tzagoloffcomplex.
ATP10, a yeast nuclear gene required for the assembly of the mitochondrial F1-F0
1990, 265:9952-9959.J. Biol. Chem.
http://www.jbc.org/content/265/17/9952Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/17/9952.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 13, 2019http://w
ww
.jbc.org/D
ownloaded from

Additions and Corrections
Vol. 264 (1989) 16093-16099 Vol. 265 (1990) 9952-9959
Isolation and characterization of a novel large protease accumulated in mammalian cells in the presence of inhibitors.
Akihiko Tsuji and Kotoku Kurachi
Page 16093, right-hand column, line 10: The sentence beginning in the middle of this line should be modified, as follows:
Electron microscopic images of large multicatalytic proteases clearly show either cylinder-shaped complexes made of four rings or discs or hollow, egg-shaped complexes (Kopp, F., Steiner, R., Dahlmann, B., Kuehn, L., and Reinauer, H. (1986) Biochim. Biophys. Acta 872, 253-260; Tanaka, K., Yoshi- mura, T., Ichihara, A., Ikai, A., Nichigai, M., Morimoto, Y., Sato, M., Tanaka, N., Katsube, Y., Kameyama, K., and Tak- agi, T. (198813) J. Mol. Biol. 203, 985-996).
Vol. 265 (1990) 15850-15853
A fraction of CD3c subunits exists as disulfide-linked dimers in both human and murine T lymphocytes.
Yang-Jiu Jin, Shigeo Koyasu, Philippe Moingeon, Ruth Steinbrich, George E. Tarr, and Ellis L. Reinherz
. Page 15852, Table I: This table was incorrectly aligned. A corrected table is shown below:
TABLE I Microsequencing analysis of Jurkat-derived CNBr fragments
from PVDF blots of TCR immunoprecipitates identifies the 20.kDa protein as CD3c
Franment SeclUenCe” Position nab
1 5 1 (M)GGITQTP%SI 6-18 2 CM) E 101-102 3 CM) DV 103-105 4 (M)SVATIVIVDICI 106-118
Unassigned residues (M) - P P - V A G F - L - -
“Cycle numbers are indicated above the sequences. Residues in bold-face type are those that were judged to be significantly above background. Initial signal level was 6-10 pmol.
*Position numbers from the NH2 terminus of mature protein in Ref. 24 were employed.
ATPlO, a yeast nuclear gene required for the assembly of the mitochondrial FI-F. complex.
Sharon H. Ackerman and Alexander Tzagoloff
Page 9953: Table I should indicate that CBllp”, W303-IA, aW303VATP10, and B103 are of the “a” mating type.
Page 9956 (last sentence of the second paragraph): the SmaI-BglII insert of pG165/ST9 is 2.6 kb, not 2.2 kb.
Page 9957: the last 53 nucleotides in Fig. 6 originate from the YEp24 vector.
We suggest that subscribers photocopy these corrections and insert the photocopies at the appropriate places where the article to be corrected originally appeared. Authors are urged to introduce these corrections into any reprints they distribute. Secondary (abstract) services are urged to carry notice of these corrections as prominently as they carried the original abstracts.
20713





























