Embed Size (px)
Citation preview

RESEARCHPAPER
At the limits: habitat suitabilitymodelling of northern 17-yearperiodical cicada extinctions(Hemiptera: Magicicada spp.)John R. Cooley1,2,3, David C. Marshall1, Chris Simon1,
Michael L. Neckermann,3 and Gerry Bunker3
1Department of Ecology and Evolutionary
Biology, University of Connecticut, Storrs, CT,
USA, 2Department of Systems Engineering,
Shizuoka University, Hamamatsu, Japan,3Cicada Research Consulting, Storrs, CT, USA
ABSTRACT
Aim Adult periodical cicadas emerge as temporally isolated, synchronized multi-species communities (‘broods’) that are for the most part geographically contigu-ous and that fit together in jigsaw-puzzle-like fashion. Some year-classes of 17-yearcicadas have become extinct within historical times. We investigate two generalcauses for these extinctions – anthropogenic habitat destruction and post-glacialclimate change.
Location Periodical cicadas are confined to the eastern United States, east of theGreat Plains. We document the locations of known periodical cicada extinctions intwo broods of 17-year cicadas in Connecticut, Rhode Island, and upstate New York,USA.
Methods Using additional distributional records of 17-year cicadas, we develophabitat suitability models for all 17-year periodical cicadas, using data layers thatreflect both ecological and anthropogenic factors.
Results Climatological data layers related specifically to annual mean temperatureand temperature during the warmest months make the greatest contributions toour models, and data layers most specifically related to deforestation and habitatfragmentation tend to make much smaller contributions. Two well-documentedextinct populations of periodical cicadas occurred in locations where these modelspredict relatively low habitat suitability for 17-year cicadas.
Main conclusions Our results and other circumstantial evidence discount theimportance of anthropogenic habitat destruction in explaining these particularextinctions.
KeywordsBrood, cicada, change, eastern USA habitat loss, hypsithermal, metapopulation.
*Corresponding: John Cooley, Department ofEcology and Evolutionary Biology, University ofConnecticut, Storrs, CT 06269-3048, USA.E-mail: [email protected]
INTRODUCTION
There has been increasing interest in ecological studies that use
modelling to predict and understand the environmental corre-
lates of species distributions (reviewed in Guisan & Zimmer-
man, 2000; Guisan & Thuiller, 2005; Araújo & Guisan, 2006;
Araújo & New, 2007; Hirzel & Le Lay, 2008; Elith & Leathwick,
2009; Franklin, 2009). These recent reviews have evaluated
methodology, provided insight into the determinants of species
distributions and stimulated debates related to ecological niche
theory, and the effects of climate change on species distribu-
tions. Our study borrows techniques from species distribution
modelling and uses them to investigate habitat suitability for an
unusual group of species locked together in time and space –
17-year periodical cicadas in the genus Magicicada.
Cicadas are hemipteran insects generally known for their loud
diurnal sounds. Only the seven Magicicada species from eastern
North America are documented to be periodical, with locally
synchronized adult emergences. Magicicada are known for long
life cycles of 13 or 17 years and periodic mass emergences of up
to millions per acre, as well as for their dependence on dense
populations (adult densities greater than c. 1 m–2) to prevent
annihilation by predators (Marlatt, 1923; Beamer, 1931; Dybas
& Davis, 1962; Lloyd & Dybas, 1966; Dybas, 1969; Karban,
bs_bs_banner
Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2013) 22, 410–421
DOI: 10.1111/geb.12002410 © 2012 Blackwell Publishing Ltd http://wileyonlinelibrary.com/journal/geb

1982a, b; Williams & Simon, 1995; Marshall et al., 2011). The
genus Magicicada is a complex of closely related species belong-
ing to three species groups (-decim, -cassini and -decula),
each with at least two members. Currently, seven species are
described, four with a 13-year life cycle, and three with a 17-year
life cycle, such that each species is more closely related to other
members of its own species group than to others with the same
life cycle (Marshall & Cooley, 2000). Multiple allochronic spe-
ciation events, perhaps in response to climate cues, have been
invoked to explain this unusual situation (Alexander & Moore,
1962; Marshall & Cooley, 2000; Cooley et al., 2001).
Among the more remarkable features of periodical cicada
biology are the ‘broods’, which are regional, often parapatric,
mixed-species year-classes of periodical cicada populations that
are bound to a common emergence timing by their shared strat-
egy of predator satiation. Cicadas in one brood are thus tempo-
rally and spatially isolated from those in other broods. Broods
probably give rise to other broods when life-cycle anomalies
(possibly climate-triggered; Marshall et al., 2003, 2011) cause
large numbers of cicadas to emerge off-schedule (Alexander &
Moore, 1962; Lloyd & Dybas, 1966; Lloyd & White, 1976; Simon
& Lloyd, 1982). In theory, broods that do not maintain high
population densities fail to satiate predators and are eliminated.
Brood declines or extinctions have been a long-running topic of
discussion in the literature on periodical cicadas (Pechumen,
1968, 1984; Maier, 1982; Cooley et al., 2004; Gilbert & Klass,
2006), but well-documented examples of extinction are difficult
to identify against a background of shifting techniques for
keeping records and making maps (Maier, 1985; Marshall, 2001).
In this paper, we consider two potential causes of the decline
of some periodical cicada populations: (1) anthropogenic
habitat fragmentation; and (2) late Holocene climate shifts.
These causes are not mutually exclusive, so we examine them by
developing habitat suitability models based on environmental
variables that reflect climatological, biotic and anthropogenic
factors. Our approach is to represent the known distribution of
periodical cicadas as accurately as possible, to identify possible
environmental correlates of range limits and to observe whether
extinct and declining populations fall outside those environ-
mental tolerances.
MATERIALS AND METHODS
Comparison of recent records with historical records suggests
contraction of the ranges of Broods VI, X, XIII and XIV, espe-
cially along their northern edges (by convention, broods are
designated by Roman numerals). This impression may be due
less to actual range contraction than to the misidentification of
cicada species, the observation of off-schedule ‘straggler’ cicadas
or the tendency for county-based maps to overestimate brood
ranges (Maier, 1985; Marshall, 2001). Unfortunately, many older
records and specimens lack specific locality or collection data,
making it difficult to resolve ambiguities. To avoid such confu-
sion, we applied strict criteria for identifying documented cases
of brood decline and concentrated on two well-documented
cases. Brood VII in upstate New York was once found through-
out the Finger Lakes region but has contracted to a small, albeit
dense, population south of Syracuse (Pechumen, 1968, 1984;
Cooley et al., 2004; Gilbert & Klass, 2006). Brood XI, restricted
to Connecticut, became extinct some time after 1954 (Marlatt,
1923; Dow, 1937; Manter, 1937, 1955, 1974).
We began our study by identifying emergence records for
Broods VII and XI that are supported by field notes (C.S.,
unpublished), specimens or previous publications with clearly
articulated methodologies and mapping criteria (e.g. Pechu-
men, 1968, 1984). We then mapped the most recent scheduled or
expected emergences of Broods VII (2001) and XI (2005) using
GIS technology. Details of the Brood VII map and the method-
ology underlying it are published elsewhere (Cooley et al.,
2004). We searched for Brood XI during its June 2005 emergence
at 74 unique localities in Connecticut and Rhode Island. These
localities were chosen because they had been recorded as having
emergences of Brood XI, they were in proximity to past reported
locations or they were within the same watersheds as past veri-
fiable records (Marlatt, 1923; Dow, 1937; Manter, 1937, 1955,
1974). We also interviewed James Slater (now deceased) of the
University of Connecticut, who along with J. A. Manter made
collections of Brood XI on 10–12 June 1954, and we interviewed
Tom Moore of the University of Michigan, who attempted to
find Brood XI during its expected 1971 emergence. Based on this
information, the last known sighting of Brood XI was in 1954
along the edge of a dairy pasture near the present address of 75
Cowles Road, in the town of Willington, CT, on the east side
of the Fenton River near both the Mansfield and Ashford
town lines (41.86° N, 72.22° W; see Appendix S1 in Supporting
Information).
We collected locality information with handheld GPS units
(Garmin, various models), using the WGS84 map datum. At
each location, we searched for physical evidence of periodical
cicadas (emergence holes, turrets, exuviae or body parts), and
we also listened for singing cicadas. We listened for minimum
5-min periods on warm, sunny, calm days only, in wooded
habitat on dates and under conditions appropriate for chorus-
ing, using methods similar to those in other studies (e.g. Mar-
shall et al., 1996; Cooley et al., 2004; Cooley et al., 2009; Cooley
et al., 2011; Marshall et al., 2011), accepting any direct evidence
of Magicicada as a positive record. Lack of evidence constituted
a negative record. In addition to our verified records, we solic-
ited records from the general public by distributing posters with
photographs of periodical cicadas and offers of a $100 cash
reward in local stores and post offices; we also placed illustrated
flyers in mailboxes near the last known collecting locality. Based
on these results and our examination of historical records, we
compiled a list of 19 localities within Broods VII and XI where
we could be confident that a population had once existed and
was now extinct (Appendix S1).
We tested possible explanations for the disappearance of
periodical cicadas in these locations by constructing habitat
suitability models (HSMs), also called ecological niche models
(ENMs) or species distribution models (SDMs; Guisan & Thu-
iller, 2005). For our occurrence data, we used a dataset that
combined all 17-year cicada species in all 17-year broods. This
Seventeen-year cicada habitat suitability
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd 411

choice bears some explanation. Although different microhabitat
preferences have been reported for the -decim, -cassini and
-decula species groups (Dybas & Lloyd, 1974) at the c. 1 km2
resolution of our models, all 17-year Magicicada species appear
to have similar general habitat requirements, are treated indis-
criminately by predators and often co-occur (Williams &
Simon, 1995). Furthermore, the species within a life cycle are so
tightly associated that older records often fail to discriminate
among them, and many older records pre-date the last taxo-
nomic revision of the genus (Alexander & Moore, 1962).
Because of these limitations, and because we are interested in
general predictions about where 17-year cicadas can be
expected to live, our analyses group all 17-year species together
and should not be mistaken for models of individual species or
broods. Based on our records, the bounding box for this study
was defined by 49° N, 25° N, -103° W, -67° W, which includes
the 48 contiguous United States east of the approximate line of
20-inch rainfall. No verified historical periodical cicada records
fall outside this area or within 200 km of its edges, although the
edges are near to some unverified (and likely spurious) histori-
cal records.
We used several versions of our dataset to build models. Our
‘full’ 17-year cicada dataset consisted of 11,392 georeferenced,
verified presence records taken during 17-year Magicicada emer-
gences from 1954–2008 and in the Cicada Central database
(http://hydrodictyon.eeb.uconn.edu/projects/cicada/databases/
databases.php). This dataset represents an extensive sampling
effort using methods similar to those described above and
designed to locate the perimeters of various broods. Each pres-
ence record used in this study was obtained or confirmed by
one or more of the authors. The overall range of 17-year cicadas
delineated by our records is similar to the range shown in Mar-
latt’s periodical cicada maps as edited by Simon (Marlatt, 1923;
Simon, 1988). No records of off-cycle or ‘straggler’ cicadas were
included, and all duplicate or geographically coincident records
were removed. The full dataset did include the records of veri-
fied extinct populations. To speed processing, a ‘thinned’ dataset
(Fig. 1) was created by randomly thinning clumped data points
resulting from the use of an automated GPS datalogger in the
most recent field seasons (Cooley et al., 2011). Thinning was
accomplished via construction of a point density contour map
of the ‘full’ dataset; areas that were most clumped on this map
were thinned by a factor of 16. The final thinned dataset con-
sisted of 1737 positive records.
Because some modelling methods make use of both presence
and absence data, we also constructed a ‘known absence’ dataset.
Although the Cicada Central database contains many absence
(‘negative’) records, these records are organized by brood mem-
bership, so they are not necessarily absence records for 17-year
cicadas in general. For instance, many of the absence records for
Brood X (Cooley et al., 2009) fall within the territory of Brood
XIV (Cooley et al., 2011) and vice versa. To select only records
that represent the complete absence of any 17-year Magicicada
brood, we constructed a detailed hull around all positive records
of 17-year cicadas and buffered it to 1 km. We discarded all
absence records falling within the periphery of the 1-km buff-
ered hull, leaving 1388 records in which 17-year Magicicada
were known to be absent. Because these absence records had the
same datalogger biases as the positive records, plus the addi-
tional bias that our absence records were concentrated on areas
in the immediate periphery of our positive records, we thinned
this dataset to 706 observed negative records, using the data
thinning techniques described above. We then constructed a
raster of inferred absences, using a c. 10-km2 grid cell size con-
tained within the bounding box of this study (see above). This
raster was converted to a point shapefile with geocoordinates
placed at the centre of each raster grid. All such points falling
within a 5-km buffered hull of positive data points, or within
Figure 1 Periodical cicada distributionrecords used to create habitat suitabilitymodels. Hatched areas are estimates ofthe placement of Marlatt’s 17-yearMagicicada records (Marlatt, 1923) asrevised by Simon (1988).
J. R. Cooley et al.
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd412

5 km of an observed absence point, were discarded, leaving 536
inferred absence points uniformly distributed in the study
bounding box and located at least 5 km from any verified pres-
ence or absence record. We used this dataset of 1242 ‘absence’
data points for modelling methods that could make use of both
presence and absence observations.
Our choice of environmental data layers was complicated
by the wealth of data available and the geographic extent of
the study region. We included 19 explicitly climatological data
layers from the WorldClim dataset in our models (Hijmans
et al., 2005) as well as nine additional ecological data layers from
other sources (Appendix S2). These non-climatological layers
reflected broad patterns in forest cover [National Land Cover
database (NCLD) 2001], forest fragment connectivity (NCLD
FrFrg2i1kml_pff), anthropogenic forest fragmentation (NCLD
FrFrg2i1kml_pfa), forest fragmentation (NCLD forfrg1kml),
anthropogenic biomes (NA Anthrome) and human impact
(Human Impact/Last of the Wild). We also included two ecore-
gion datasets that categorize the study region based on land use,
landform, potential natural vegetation, soils and other factors
(Bailey, 1980; Omernik, 1987). All data layers represent current
patterns, not historical conditions or future projections. From
these data layers, we prepared environmental raster datasets in
ArcGIS 9.3, using a c. 1-km2 grid cell size, clipped to the study’s
bounding box to speed processing, reduce file size and reduce
model errors related to an overly large background dataset
(Anderson & Raza, 2010).
A priori choices among the 28 data layers seemed arbitrary,
given the potential interdependence of the datasets and the scar-
city of information about the physiological ecology of periodical
cicadas. For example, since periodical cicadas emerge in the
spring, it might seem tempting to concentrate on variables
related to spring climate; however, it is possible that the cicadas
are most sensitive to summer conditions (when eggs are hatch-
ing) or winter conditions (when the underground nymphs must
avoid freezing). Thus, for our initial models, we used all 28
environmental data layers, and, in effect, we used initial HSMs to
inform variable culling prior to making final models. For our
final models, we selected three high-performing climatic layers
and three high-performing ecological layers. However, these
were not necessarily the six highest-performing layers in our
initial models; rather, our choices were intended to include both
climatological and non-climatological ecological information
and minimize overlap among variables.
We constructed SDMs using several methods, including
random forest, generalized boosting models, classification tree
analysis, multiple adaptive regression splines, generalized addi-
tive models, generalized linear models and artificial neural net-
works, as implemented in the R library BIOMOD (Guisan &
Thuiller, 2005; Moisen et al., 2006; Thuiller et al., 2009). All
methods were optimized according to the receiver operating
characteristic (ROC) criterion, a 50:50 data split for training and
evaluation, and 100 cross-validation runs. Using BIOMOD’s
‘Ensemble.Forecasting’ command, an ensemble forecast of all the
methods, weighted by model score using a default value (1.6),
was made (see Araújo & New, 2007). This ensemble model was
converted into an ArcGIS raster with c. 10-km2 cell size by
kriging.
We also constructed models using the maximum entropy
methods of MaxEnt 3.3.3k (Phillips et al., 2004, 2006; Peterson
et al., 2007; Phillips & Dudík, 2008). MaxEnt models were run
with a default convergence threshold of 10-5 for 1000 replicates,
each time with 25% of the data set chosen randomly without
replacement as training samples, using the ‘Remove Duplicate
Presence Records’ option to reduce the dataset to one observation
per raster cell. For preliminary models, we used the default value
of 10,000 background samples, but for our final model we
increased the value to 50,000. Other MaxEnt options, such as
regularization parameters, were set to default values. The relative
contributions of the different environmental variables were
determined and a jackknife analysis was performed to deter-
mine which variables contained the most unique information.
While the area under the curve (AUC) criterion (Fielding & Bell,
1997) was used to assess model performance, we do not base our
conclusions on comparisons of AUC scores among models.
To judge the support for either anthropogenic or ecological
explanations for periodical cicada extinctions, we considered the
relative importance of various environmental data layers in all of
our models. We also extracted model scores for extant, extinct,
absent and inferred absent locations and determined whether
the extinct populations were located in areas of high or low
predicted suitability.
RESULTS
Searches for Brood VII periodical cicadas are summarized in
Cooley et al. (2004). Despite searching during appropriate times
and conditions, we found no evidence of periodical cicadas
within the reported range of Brood XI (Fig. 2). Although we
were contacted by members of the public who had seen other
insects, and, later in the season, those who had seen cicadas
belonging to the genus Tibicen, no member of the general public
provided any verifiable evidence of periodical cicadas.
All models are well supported (Table 1) and match, with rea-
sonable accuracy, the known range of 17-year cicadas (compare
the hatched area in Fig. 1 with Fig. 3). Although the environ-
mental variables with the greatest predictive power varied
among our analyses, all of the model building methods tended
to place the greatest importance on climatologically oriented
variables related to temperature and precipitation, whereas less
climatologically oriented variables, such as forest fragmentation
and ecoregions, were rarely high scoring. Within BIOMOD, the
method producing the highest scoring model (using the Predic-
tionBestModel) was the random forest (RF) model (Table 1). In
all models, purely climatological variables reflecting tempera-
ture and precipitation made the highest contributions, while the
layers most associated with anthropogenic deforestation (forest
fragmentation classification and land-cover classification) made
only small contributions (Table 1), although the magnitudes of
the variable contributions should be viewed with caution since
the variables are not necessarily independent.
Seventeen-year cicada habitat suitability
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd 413

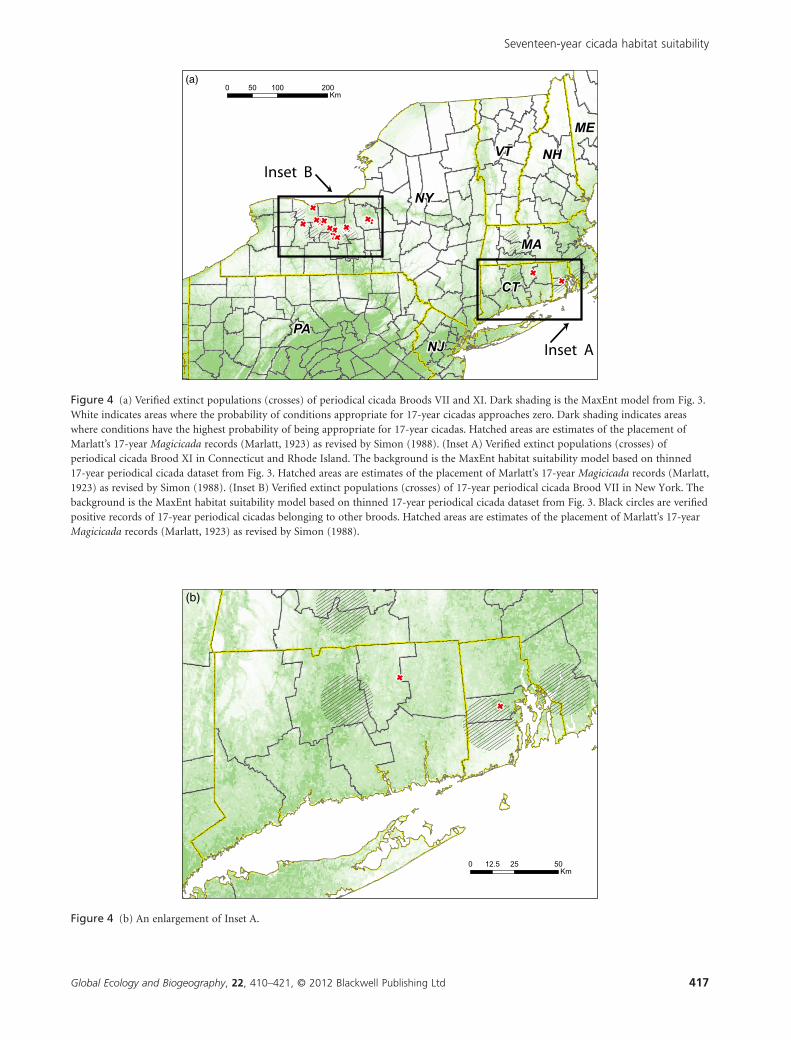
Extinct populations of Broods VII and XI fall in or near areas
where our models predict lower suitability for 17-year cicadas
(Fig. 4). A plot of model values for extant and extinct popula-
tions shows that extinct populations of Broods VII and XI are
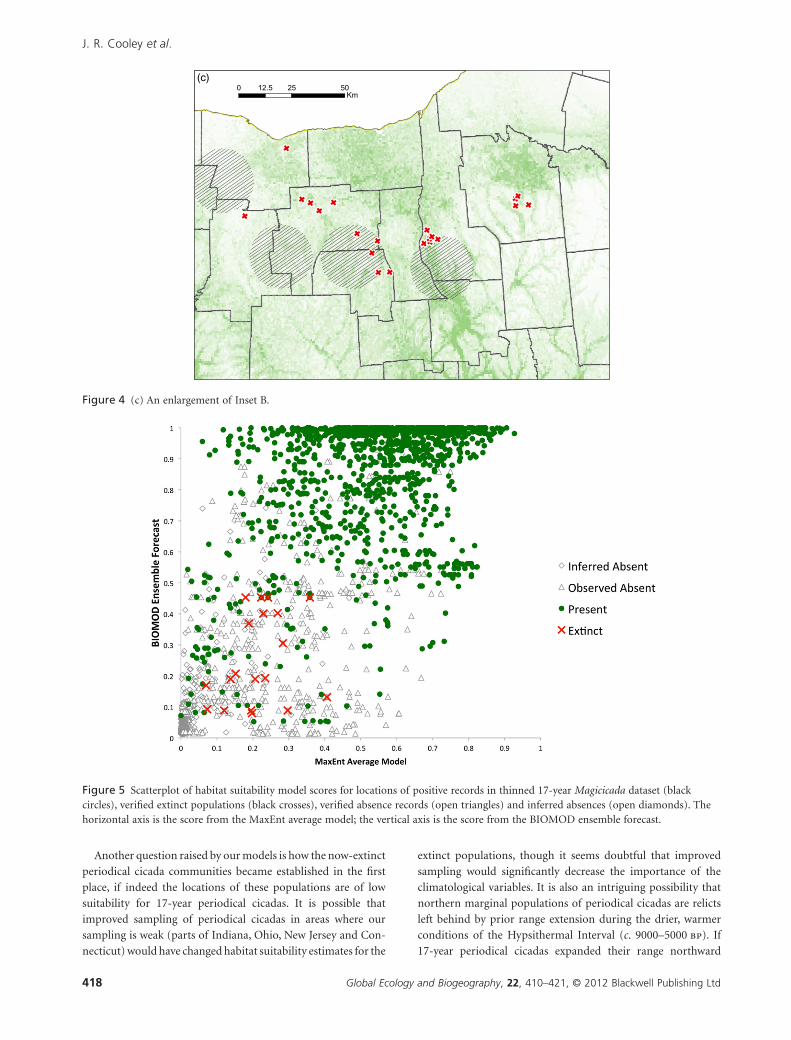
low-value outliers in all models (Fig. 5). These differences are
statistically significant; when model scores for extinct and extant
populations are extracted from the thinned-dataset model,
scores for extinct populations are statistically lower for the
MaxEnt model (Mann–Whitney–Wilcoxon test; W = 2405.5, P <0.001) and the summary statistic for the BIOMOD ensemble
models (Mann–Whitney–Wilcoxon test; W = 801, P < 0.001).
Thus, extinct populations of Broods VII and XI fall into areas
where our models predict a lower probability of conditions
appropriate for 17-year Magicicada.
DISCUSSION
In this paper we investigate factors that affect habitat suitability
for 17-year cicadas in the genus Magicicada. It would be inac-
curate for us to use the term ‘species distribution modelling’ to
describe our efforts, because of the quirky biology of periodical
cicadas: Whereas the 17-year cicadas are a group of closely
related and largely co-distributed species that in some respects
function as a single ecological unit, they do not form a mono-
phyletic clade. Even so, these species are locked together in space
and time, and it is the manner in which their shared ecology
binds them together and limits their distribution that we wished
to capture in our models. While the accuracy of HSMs tends to
be greatest for taxa with small geographic ranges, limited eco-
logical tolerance (Hernandez et al., 2006) and data sets of inter-
mediate prevalence (McPherson et al., 2004), we generated
HSMs that accurately represent the distribution of 17-year peri-
odical cicadas, even though their range is large and ecologically
diverse.
Two aspects of our models are relevant to understanding
extinct populations and broods. First, documented extinct
populations of Broods VII and XI were found in areas where our
models predict a comparatively low probability of conditions
appropriate for 17-year cicadas. Second, climatological variables
(e.g. temperature and precipitation) seem to have the greatest
importance in our models, while non-climatological variables
(e.g. land cover and fragmentation) tend to make smaller model
contributions. Taken at face value, these results suggest that
climatological explanations for the increased susceptibility of
periodical cicadas to extinction along the northern edge of their
distribution are more plausible than non-climatological expla-
nations like ‘ecoregions’, anthropogenic fragmentation, forest
connectivity and land-cover classification (see Appendix S2).
The low contribution of the forest fragmentation data layers
to our models was unexpected. To the extent that fragmentation
involves the reduction of existing forests into smaller and
smaller patches, we expect the probability of local extinction to
increase with increasing fragmentation, because periodical
cicadas depend on high community densities of at least tens of
thousands per hectare that decrease per capita predation risk
(Beamer, 1931; Dybas, 1969; Karban, 1982a; Williams & Simon,
1995; Marshall et al., 2011). In addition, forest patchiness has
been found to affect the spatial distributions of periodical
cicadas (Rodenhouse et al., 1997), and Magicicada fecundity
may be lowered in sparse populations (Karban, 1982b). Thus,
scarcity tends to magnify conditions that further depress popu-
lation size leading to demographic time lags that make it diffi-
cult for declining populations to recover. It is possible that the
measures of fragmentation used in our study fail to capture the
importance of fragmentation to periodical cicadas, either
because the resolution of the data layers is too coarse or because
current levels of fragmentation are irrelevant; instead, informa-
tion about fragmentation that occurred in the past might be
Figure 2 Periodical cicada Brood XI2005 search area in Connecticut andRhode Island in 2005. Cross symbolsdenote the absence of periodical cicadas;no positive records of Brood XI werefound in 2005. Black circles are verifiedpositive records of 17-year periodicalcicadas belonging to other broods.Hatched areas are estimates of theplacement of Marlatt’s Brood XI records(Marlatt, 1923) as revised by Simon(1988).
J. R. Cooley et al.
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd414

Tab
le1
Hab
itat
suit
abili
tym
odel
sfo
rth
eth
inn
edda
tase
tof
17-y
ear
peri
odic
alci
cada
reco
rds
usi
ng
six
envi
ron
men
talv
aria
bles
.For
Max
En
tm
odel
s,va
riab
leco
ntr
ibu
tion
sar
eex
pres
sed
aspe
rcen
tage
con
trib
uti
onto
the
mod
el(a
nd
thu
sal
lsu
mto
1),a
nd
mod
els
are
give
nan
area
un
der
the
curv
e(A
UC
)sc
ore.
Ast
eris
ks(*
)in
dica
teva
riab
lew
ith
the
hig
hes
tga
inw
hen
use
din
isol
atio
n(t
hat
is,t
he
vari
able
con
tain
ing
the
mos
tu
sefu
lin
form
atio
nby
itse
lf).
For
met
hod
sru
nin
BIO
MO
D,v
aria
ble
con
trib
uti
ons
(usi
ng
the
Var
Impo
rtan
cefu
nct
ion
)ar
e1
min
us
the
corr
elat
ion
betw
een
the
orig
inal
pred
icti
onan
dth
epr
edic
tion
mad
ew
ith
the
vari
able
perm
ute
d(t
hus
they
don
otsu
mto
1;se
eT
huill
eret
al.,
2009
).M
eth
ods
run
inB
IOM
OD
are
give
na
rece
iver
oper
atin
gch
arac
teri
stic
(RO
C)
scor
e.In
both
case
s,h
igh
erva
lues
indi
cate
grea
ter
con
trib
uti
ons
byth
eva
riab
les.
Env
iron
men
talv
aria
bles
wit
hth
egr
eate
stpr
edic
tive
pow
erp
erm
odel
are
indi
cate
din
bold
.
AU
C/R
OC
Max
En
tM
axE
nt
Art
ifici
al
neu
raln
etw
ork
(AN
N)
Cla
ssifi
cati
on
tree
anal
ysis
(CTA
)
Gen
eral
ized
addi
tive
mod
el
(GA
M)
Gen
eral
ized
boos
tin
gm
odel
(GB
M)
Gen
eral
ized
linea
rm
odel
(GLM
)
Mu
ltip
lead
apti
ve
regr
essi
onsp
lines
(MA
RS)
Ran
dom
fore
st
(RF)
28-v
aria
ble
6-va
riab
le2-
fold
50(c
ross
valid
atio
n)
3-sp
line
Poly
nom
ial
0.84
2�
0.00
40.
924
�0.
004
0.45
30.
899
0.42
20.
739
0.39
50.
620
0.99
9
Tem
pera
ture
Seas
onal
ity
(SD
¥10
0)
Wor
ldC
lim4
0.04
00.
077
0.71
40.
581
0.15
60.
121
0.11
90.
280
0.35
7
Max
.tem
pera
ture
of
war
mes
tm
onth
Wor
ldC
lim5
0.30
40.
453
0.53
80.
296
0.39
60.
192
0.35
40.
263
0.24
0
An
nu
alpr
ecip
itat
ion
Wor
ldC
lim12
0.19
60.
264
0.64
00.
564
0.11
70.
358
0.20
70.
453
0.40
6
Eco
regi
ons
ofU
nit
ed
Stat
es
Bai
ley
(198
0)0.
031*
0.13
1*0.
402
0.61
80.
297
0.46
50.
271
0.63
30.
305
An
thro
poge
nic
frag
men
tati
on
1km
2(N
CLD
,200
1)
Nat
ion
alA
tlas
Seam
less
Serv
er
0.04
00.
028
0.14
00.
021
0.01
60.
000
0.02
00.
000
0.02
8
Lan
dco
ver
clas
sifi
cati
on
(NC
LD,2
001)
Nat
ion
alA
tlas
Seam
less
Serv
er
0.04
00.
048
0.06
30.
021
0.03
30.
000
0.03
50.
000
0.01
0
NC
LD,N
atio
nal
Lan
dC
over
Dat
abas
e;se
eH
omer
etal
.(20
12).
Seventeen-year cicada habitat suitability
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd 415

more informative. Unfragmented habitat that has regenerated
during the past century following agricultural abandonment
(such as habitat found broadly throughout the north-eastern
states today) may contain few Magicicada populations, as a
legacy of widespread extinction that occurred long ago during
extensive deforestation. Indeed, the last known location of
Brood XI and some of the areas formerly inhabited by VII are
currently forested and classified as intact and interconnected,
while an early 20th century photograph of this area, taken when
the brood was still in existence, shows patchy woodlots and
fencerows (Appendix S3; Favretti, 2003).
One issue raised by our study is why extinction has been well
documented only for Broods VII and XI, when portions of
Broods VIII, X, XIII and XIV occupy similar latitudes. In fact,
similar declines have been reported on the basis of county-level
maps in Indiana (Brood X; Kritsky, 1987) in areas where our
models predict lower suitability for 17-year Magicicada. It is
possible that 17-year populations are in retreat along the entire
northern edge of their range, and the lack of documentation
may be simply an artefact of missing or ambiguous historical
records. Indeed, along the northern periphery of the general
periodical cicada distribution, recent emergences of Broods X,
XIII, XIV and several disjunct populations of various broods on
Long Island (Simon & Lloyd, 1982) have fallen short of their
historically reported ranges (J.R.C., D.C.M. and C.S. unpub
lished data). Reporting biases may also explain the lack of
well-documented declines; for example, the complete extinction
of a peripheral brood such as VII or XI may be far more likely to
be noticed than a local extinction near the centre of a large
brood.
Figure 3 (a), (b). Species distributionmodels based on a thinned 17-yearMagicicada dataset and six variables.White indicates areas where probabilityof conditions appropriate for 17-yearcicadas approaches zero. Shadingindicates areas where conditions have thehighest probability of being appropriatefor 17-year cicadas, with darker shadingindicating higher probabilities. (a)BIOMOD ensemble forecast. (b) Averageof 1000 replicate MaxEnt models.
J. R. Cooley et al.
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd416
(a)
Figure 4 (a) Verified extinct populations (crosses) of periodical cicada Broods VII and XI. Dark shading is the MaxEnt model from Fig. 3.White indicates areas where the probability of conditions appropriate for 17-year cicadas approaches zero. Dark shading indicates areaswhere conditions have the highest probability of being appropriate for 17-year cicadas. Hatched areas are estimates of the placement ofMarlatt’s 17-year Magicicada records (Marlatt, 1923) as revised by Simon (1988). (Inset A) Verified extinct populations (crosses) ofperiodical cicada Brood XI in Connecticut and Rhode Island. The background is the MaxEnt habitat suitability model based on thinned17-year periodical cicada dataset from Fig. 3. Hatched areas are estimates of the placement of Marlatt’s 17-year Magicicada records (Marlatt,1923) as revised by Simon (1988). (Inset B) Verified extinct populations (crosses) of 17-year periodical cicada Brood VII in New York. Thebackground is the MaxEnt habitat suitability model based on thinned 17-year periodical cicada dataset from Fig. 3. Black circles are verifiedpositive records of 17-year periodical cicadas belonging to other broods. Hatched areas are estimates of the placement of Marlatt’s 17-yearMagicicada records (Marlatt, 1923) as revised by Simon (1988).
(b)
Figure 4 (b) An enlargement of Inset A.
Seventeen-year cicada habitat suitability
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd 417
Another question raised by our models is how the now-extinct
periodical cicada communities became established in the first
place, if indeed the locations of these populations are of low
suitability for 17-year periodical cicadas. It is possible that
improved sampling of periodical cicadas in areas where our
sampling is weak (parts of Indiana, Ohio, New Jersey and Con-
necticut) would have changed habitat suitability estimates for the
extinct populations, though it seems doubtful that improved
sampling would significantly decrease the importance of the
climatological variables. It is also an intriguing possibility that
northern marginal populations of periodical cicadas are relicts
left behind by prior range extension during the drier, warmer
conditions of the Hypsithermal Interval (c. 9000–5000 bp). If
17-year periodical cicadas expanded their range northward
(c)
Figure 4 (c) An enlargement of Inset B.
Figure 5 Scatterplot of habitat suitability model scores for locations of positive records in thinned 17-year Magicicada dataset (blackcircles), verified extinct populations (black crosses), verified absence records (open triangles) and inferred absences (open diamonds). Thehorizontal axis is the score from the MaxEnt average model; the vertical axis is the score from the BIOMOD ensemble forecast.
J. R. Cooley et al.
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd418

during these milder conditions, then, as conditions cooled,
northern limital populations could become vulnerable to extinc-
tion as the range of 17-year cicadas returned to a state of climatic
equilibrium and populations became isolated or dwindled below
levels required for predator satiation (see Araújo & Pearson,
2005). If this hypothesis is correct, then local population extinc-
tions should have occurred more often along the northern
Magicicada range limit than along the southern margin, which is
defined by 13-year cicada populations. Now that climatic condi-
tions are warming again, one might predict an expansion, but
climatic extremes (e.g. early warmth followed by late cold shocks)
associated with recent warming may have more of an effect than
the average warming trend (Parmesean, 2006; Knapp et al.,
2008).
An unusual quality of periodical cicadas is that multiple sym-
patric, synchronized species usually emerge together.
While different Magicicada species have somewhat different
habitat preferences (Dybas & Lloyd, 1962, 1974), predators
appear to treat the different periodical cicada species inter-
changeably. In a mosaic of upland forests (preferred by Magici-
cada species in the -decim complex) and lowland drainages
(preferred by Magicicada species in the -cassini complex), one or
more rare species may avoid predator annihilation due to the
local presence of a dense congener (Lloyd & White, 1983). In this
way, two synchronized species may survive predator pressure
where one alone would not. If so, then northern periodical cicada
communities (e.g. Massachusetts, Connecticut, upstate New
York, Michigan and central Wisconsin) may be especially suscep-
tible to extinction, since they typically contain only a single
species – Magicicada septendecim. Future studies could test this
hypothesis more carefully by specifically comparing extinction
rates observed along the northern edge of the 17-year range,
where only M. septendecim is present, and in the south-western
sector, where only Magicicada cassini is present, to regionally
adjacent locations where populations contain multiple species.
The peculiar biology of periodical cicadas means that migra-
tion among spatially-and temporally separated patches should
also be considered. Although the historical treatment of peri-
odical cicada broods is that they are evolutionary units, it may be
more appropriate to consider them as metapopulations, in
which local population patches are interdependent and bound
to a common emergence schedule by predation pressure. Local
patches of the same brood may exchange migrants, and in those
places where broods are adjacent, temporal migration among
patches may occur when cicadas emerge off-cycle (‘stragglers’).
Such spatial and temporal exchanges may be a key factor in
reducing the probability that predators can find and eliminate
all patches of periodical cicadas in a given region. This may be
particularly important where Broods XIV, X, VI and II overlap
(Lloyd & White, 1976; Simon & Lloyd, 1982; Heliövaara et al.,
1994). However, northern populations of 17-year cicadas appear
to be the ones for which replenishment from neighbouring
populations seems least likely, because northern broods and
populations tend to be isolated from all others.
Why are some northern 17-year periodical cicada populations
going extinct? Although their extensive range and broad toler-
ances would seem to make these insects unlikely candidates for a
successful modelling approach, our models do seem to capture
the essentials of the 17-year cicada distribution. Surprisingly,
these models discount the importance of factors such as present-
day deforestation or forest fragmentation in documented Magici-
cada extinctions.Yet as compelling as they seem, these models are
missing something important: They are entirely lacking the kinds
of ecological interactions, such as predator satiation, known to be
important for these species. An extinction that appears to be
imminent underscores this point. Raccoon Grove Forest Pre-
serve, located 36 miles south of Chicago in Will County, IL, is
found within periodical cicada Brood XIII and in an area where
our models predict suitable conditions for periodical cicadas. In
this location in 1956, Dybas & Davis (1962) measured the highest
Magicicada population density estimate ever recorded for a
natural population, over 1.5 million adults per acre for M. cassini.
Yet during the 2007 emergence of Brood XIII, only a handful of
cicadas were reported from the site (L. Yang, University of Cali-
fornia, Davis, CA, USA, unpublished data). While development
appears to have encroached on the forests surrounding Raccoon
Grove, the preserve boundaries have not changed since 1956, and
the overall forest block remains substantial (around 1 km2,
judging from aerial photographs). Although additional factors
may be involved, including a bout of Dutch elm disease that killed
many trees on the floodplain woods after the 1956 emergence
(White & Lloyd, 1975), the increasing isolation of Raccoon Grove
suggests a role for the kinds of species interactions and metap-
opulation dynamics discussed above. The likely extinction at
Raccoon Grove suggests two cautions about HSMs: first, the
accuracy of such models and the validity of conclusions drawn
from them may vary over the range of the model, especially for
widespread species; second, until we find a way to incorporate
species interactions into them, such models will involve signifi-
cant oversimplifications.
ACKNOWLEDGEMENTS
This paper is dedicated to the memory of the late Charles Rem-
ington, of Yale University, who introduced the first author to
periodical cicadas and their vulnerability to extinction. The
National Geographic CRE-sponsored project ‘Making modern
maps of Magicicada emergences’ provided partial funding for
this project. Kathy Hill assisted in collecting records. Cory Merow
and anonymous referees provided valuable comments on earlier
drafts. The UConn Bioinformatics Facility provided computing
resources for this study. This material is based upon work par-
tially supported by the National Science Foundation under grant
nos NSF DEB 04–22386, DEB 05–29679 to Chris Simon. Any
opinions, findings, and conclusions or recommendations
expressed in this material are those of the authors and do not
necessarily reflect the views of the National Science Foundation.
REFERENCES
Alexander, R.D. & Moore, T.E. (1962) The evolutionary rela-
tionships of 17-year and 13-year cicadas, and three new
Seventeen-year cicada habitat suitability
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd 419

species. (Homoptera: Cicadidae, Magicicada). University of
Michigan Museum of Zoology Miscellaneous Publication, 121,
1–59.
Anderson, R.P. & Raza, A. (2010) The effect of the extent of the
study region on GIS models of species geographic distribu-
tions and estimates of niche evolution: preliminary tests with
montane rodents (genus Nephelomys) in Venezuela. Journal of
Biogeography, 37, 1378–1393.
Araújo, M.B. & Guisan, A. (2006) Five (or so) challenges for
species distribution modelling. Journal of Biogeography, 33,
1677–1688.
Araújo, M.B. & New, M. (2007) Ensemble forecasting of
species distributions. Trends in Ecology and Evolution, 22,
42–47.
Araújo, M.B. & Pearson, R.G. (2005) Equilibrium of species’
distributions with climate. Ecography, 28, 693–695.
Bailey, R.G. (1980) Description of the ecoregions of the United
States. US Department of Agriculture Miscellaneous Publica-
tion 1391. US Forest Service, Washington, DC.
Beamer, R.H. (1931) Notes on the 17-year cicada in Kansas.
Journal of the Kansas Entomological Society, 4, 53–58.
Cooley, J.R., Simon, C., Marshall, D.C., Slon, K. & Ehrhardt, C.
(2001) Allochronic speciation, secondary contact, and
reproductive character displacement in periodical cicadas
(Hemiptera: Magicicada spp.): genetic, morphological,
and behavioural evidence. Molecular Ecology, 10, 661–671.
Cooley, J.R., Marshall, D.C. & Simon, C. (2004) The historical
contraction of periodical cicada Brood VII (Hemiptera: Cica-
didae: Magicicada). Journal of the New York Entomological
Society, 112, 198–204.
Cooley, J.R., Kritsky, G., Zyla, J.D., Edwards, M.J., Simon,
C., Marshall, D.C., Hill, K.B.R. & Krauss, R. (2009) The dis-
tribution of periodical cicada Brood X. The American Ento-
mologist, 55, 106–112.
Cooley, J.R., Kritsky, G., Edwards, M.D., Zyla, J.D., Marshall,
D.C., Hill, K.B.R., Bunker, G.J., Neckermann, M.L. & Simon,
C. (2011) Periodical cicadas (Magicicada spp.): the distribu-
tion of Broods XIV in 2008 and ‘XV’ in 2009. The American
Entomologist, 57, 144–151.
Dow, R. (1937) Brood XI is not extinct – yet. Bulletin of the New
England Museum of Natural History, 84, 3–4.
Dybas, H.S. (1969) The 17-year cicada: a four year mistake?
Bulletin of the Field Museum of Natural History, 40, 10–12.
Dybas, H.S. & Davis, D.D. (1962) A population census of
seventeen-year periodical cicadas (Homoptera: Cicadidae:
Magicicada). Ecology, 43, 432–444.
Dybas, H.S. & Lloyd, M. (1962) Isolation by habitat in two
synchronized species of periodical cicadas (Homoptera:
Cicadidae: Magicicada). Ecology, 43, 444–459.
Dybas, H.S. & Lloyd, M. (1974) The habitats of 17-year periodi-
cal cicadas (Homoptera: Cicadidae: Magicicada spp.). Ecologi-
cal Monographs, 44, 279–324.
Elith, J. & Leathwick, J.R. (2009) Species distribution models:
ecological explanation and prediction across space and
time. Annual Review of Ecology, Evolution and Systematics, 40,
677–697.
Favretti, R.J. (2003) Mansfield four corners: what it used to be and
George Washington didn’t see. Bibliopola Press, Storrs, CT.
Fielding, A.H. & Bell, J.F. (1997) A review of the methods for the
assessment of prediction errors in conservation presence/
absence models. Environmental Conservation, 24, 38–49.
Franklin, J. (2009) Mapping species distributions: spatial inference
and prediction. Cambridge University Press, Cambridge.
Gilbert, C. & Klass, C. (2006) Decrease in geographic range of
the finger lakes brood (Brood VII) of the periodical cicada
(Hemiptera: Cicadidae: Magicicada spp.). Journal of the New
York Entomological Society, 114, 78–85.
Guisan, A. & Thuiller, W. (2005) Predicting species distribution:
offering more than simple habitat models. Ecology Letters, 8,
993–1009.
Guisan, A. & Zimmerman, N.E. (2000) Predictive habitat distri-
bution models in ecology. Ecological Modelling, 135, 147–186.
Heliövaara, K., Väisänen, R. & Simon, C. (1994) Evolutionary
ecology of periodical insects. Trends in Ecology and Evolution,
9, 475–480.
Hernandez, P.A., Graham, C.H., Master, L.L. & Albert, D.L.
(2006) The effect of sample size and species characteristics
on performance of different species distribution modeling
methods. Ecography, 29, 773–785.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G. & Jarvis, A.
(2005) Very high resolution interpolated climate surfaces for
global land areas. International Journal of Climatology, 25,
1965–1978.
Hirzel, A.H. & Le Lay, G. (2008) Habitat suitability modeling
and niche theory. Journal of Applied Ecology, 45, 1372–1381.
Homer, C.H., Fry, J.A. & Barnes, C.A. (2012) The National Land
Cover Database. US Geological Survey Fact Sheet 2012–3020.
Karban, R. (1982a) Population ecology of periodical cicadas. PhD
Thesis, University of Pennsylvania, Philadelphia.
Karban, R. (1982b) Increased reproductive success at high den-
sities and predator satiation for periodical cicadas. Ecology, 63,
321–328.
Knapp, A.K., Beier, C., Briske, D.D., Classen, A.T., Luo, Y., Reich-
stein, M., Smith, M.D., Smith, S.D., Bell, J.E., Fay, P.A., Heisler,
J.L., Leavitt, S.W., Sherry, R., Smith, B. & Weng, E. (2008)
Consequences of more extreme precipitation regimes for ter-
restrial ecosystems. Bioscience, 58, 1–11.
Kritsky, G. (1987) An historical analysis of periodical cicadas in
Indiana (Homoptera: Cicadidae). Proceedings of the Indiana
Academy of Sciences, 97, 295–322.
Lloyd, M. & Dybas, H.S. (1966) The periodical cicada problem.
II. Evolution. Evolution, 20, 466–505.
Lloyd, M. & White, J.A. (1976) Sympatry of periodical cicada
broods and the hypothetical four-year acceleration. Evolution,
30, 786–801.
Lloyd, M. & White, J.A. (1983) Why is one of the periodical
cicadas (Magicicada septendecula) a comparatively rare
species? Ecological Entomology, 8, 293–303.
McPherson, J., Jetz, W. & Rogers, D.J. (2004) The effects of
species’ range sizes on the accuracy of distribution models:
ecological phenomenon or statistical artefact? Journal of
Applied Ecology, 41, 811–823.
J. R. Cooley et al.
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd420

Maier, C. (1982) Abundance and distribution of the seventeen-
year periodical cicada, Magicicada septendecim (Linnaeus)
(Hemiptera: Cicadidae- Brood II), in Connecticut. Proceed-
ings of the Entomological Society of Washington, 84, 430–439.
Maier, C. (1985) Brood VI of 17-year periodical cicadas, Magici-
cada spp. (Hemiptera: Homoptera: Cicadidae): new evidence
from Connecticut (USA), the hypothetical 4-year decelera-
tion, and the status of the brood. Journal of the New York
Entomological Society, 93, 1019–1026.
Manter, J.A. (1937) The Periodical Cicada: Occurrence of Brood
XI in Connecticut in 1937. In: 14th Annual Conference of
Connecticut Entomologists, p. 153. Connecticut State Ento-
mologist’s Report, Bulletin 408, Connecticut Agricultural
Experiment Station, New Haven, CT.
Manter, J.A. (1955) Magicicada septendecim Linn., Brood XI in
Connecticut (Hemiptera: Cicadidae). Psyche, 62, 157–158.
Manter, J.A. (1974) Brood XI of the periodical cicada seems
doomed. 25th Anniversary Memoirs of the Connecticut Ento-
mological Society (ed. by R.L. Beard), pp. 99–100. Connecticut
Entomological Society, New Haven.
Marlatt, C. (1923) The periodical cicada. US Department of
Agriculture, Bureau of Entomology Bulletin 71.
Marshall, D.C. (2001) Periodical cicada (Homoptera: Cicadidae)
life-cycle variations, the historical emergence record, and the
geographic stability of brood distributions. Annals of the Ento-
mological Society of America, 94, 386–399.
Marshall, D.C. & Cooley, J.R. (2000) Reproductive character
displacement and speciation in periodical cicadas, with
description of a new species, 13-year Magicicada neotredecim.
Evolution, 54, 1313–1325.
Marshall, D.C., Cooley, J.R., Alexander, R.D. & Moore, T.E.
(1996) New records of Michigan Cicadidae (Homoptera),
with notes on the use of songs to monitor range changes.
Great Lakes Entomologist, 29, 165–169.
Marshall, D.C., Cooley, J.R. & Simon, C. (2003) Holocene
climate shifts, life-cycle plasticity, and speciation in periodical
cicadas: a reply to Cox and Carlton. Evolution, 57, 433–437.
Marshall, D.C., Cooley, J.R. & Hill, K.B.R. (2011) Developmen-
tal plasticity of life-cycle length in thirteen-year periodical
cicadas (Hemiptera: Cicadidae). Annals of the Entomological
Society of America, 104, 443–450.
Moisen, G.G., Edwards, T.C.J. & Osborne, P.E. (2006) Further
advances in predicting species distributions. Ecological Mod-
elling, 199, 129–131.
Omernik, J.M. (1987) Ecoregions of the coterminous United
States. Annals of the Association of American Geographers, 77,
118–125.
Parmesean, C. (2006) Ecological and evolutionary responses to
recent climate change. Annual Review of Ecology and System-
atics, 37, 637–639.
Pechumen, L.L. (1968) The periodical cicada, Brood VII
(Homoptera: Cicadidae: Magicicada). Transactions of the
American Entomological Society, 94, 137–153.
Pechumen, L.L. (1984) The periodical cicada: Brood VII revis-
ited (Homoptera: Cicadidae). Entomological News, 96, 59.
Peterson, A.T., Papes, M. & Eaton, E. (2007) Transferrability and
model evaluation in ecological niche modeling: a comparison
of GARP and Maxent. Ecography, 30, 550–560.
Phillips, S.J. & Dudík, M. (2008) Modeling of species distribu-
tions with Maxent: new extensions and a comprehensive
evaluation. Ecography, 31, 161–175.
Phillips, S.J., Dudík, M. & Schapire, R.E. (2004) A maximum
entropy approach to species distribution modeling. Proceed-
ings of the 21st International Conference on Machine Learning
(ed. by R. Greiner and D. Schuurmans), pp. 655–662. ACM
Press, New York, NY.
Phillips, S.J., Anderson, R.P. & Schapire, R.E. (2006) Maximum
entropy modeling of species geographic distributions. Ecologi-
cal Modelling, 190, 231–259.
Rodenhouse, N.L., Bohlen, P.J. & Barrett, G.W. (1997) Effects of
woodland shape on the spatial distribution and density of
17-year periodical cicadas (Homoptera: Cicadidae). American
Midland Naturalist, 137, 124–135.
Simon, C. (1988) Evolution of 13- and 17-year periodical
cicadas. Bulletin of the Entomological Society of America, 34,
163–176.
Simon, C. & Lloyd, M. (1982) Disjunct synchronic population
of 17-year periodical cicadas: relicts or evidence of poly-
phyly? Journal of the New York Entomological Society, 90, 275–
301.
Thuiller, W., Lafourcade, B., Engler, R. & Araújo, M.B. (2009)
BIOMOD – a platform for ensemble forecasting of species
distributions. Ecography, 32, 369–373.
White, J.A. & Lloyd, M. (1975) Growth rates of 17- and 13-year
periodical cicadas. American Midland Naturalist, 94, 127–143.
Williams, K.S. & Simon, C. (1995) The ecology, behavior, and
evolution of periodical cicadas. Annual Review of Entomology,
40, 269–295.
SUPPORTING INFORMATION
Additional supporting information may be found in the online
version of this article at the publisher’s web-site.
Appendix S1. Verified, extinct populations of periodical cicada
Broods VII and XI.
Appendix S2. Environmental layers used to construct habitat
suitability models for 17-year periodical cicadas.
Appendix S3. Fenton River Valley, looking east towards Willing-
ton, CT along the Boston Turnpike.
BIOSKETCH
John R. Cooley, researcher at the University of
Connecticut, studies the behaviour and evolution of
cicadas. Current information about the cicada mapping
project may be found at http://www.magicicada.org.
Editor: Antoine Guisan
Seventeen-year cicada habitat suitability
Global Ecology and Biogeography, 22, 410–421, © 2012 Blackwell Publishing Ltd 421





























