Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, July 1992, p. 2188-2195 Vol. 58, No. 70099-2240/92/072188-08$02.00/0Copyright © 1992, American Society for Microbiology
Assessment of Genetic Diversity and Population Structure ofXanthomonas oryzae pv. Oryzae with a Repetitive
DNA ElementJ. E. LEACH,1* M. L. RHOADS,' C. M. VERA CRUZ,"12 F. F. WHITE,' T. W. MEW,2
AND H. LEUNG2'3Department of Plant Pathology, Throckmorton Hall, Kansas State University, Manhattan,Kansas 66506-55021; Department of Plant Pathology, The Intemnational Rice Research
Institute, Manila, Philippines2; and Department of Plant Pathology, WashingtonState University, Pullman, Washington 99164-64303
Received 9 January 1992/Accepted 4 May 1992
A repetitive DNA element cloned from Xanthomonas oryzae pv. oryzae was used to assess the populationstructure and genetic diversity of 98 strains ofX. oryzae pv. oryzae collected between 1972 and 1988 from thePhilippine Islands. Genomic DNA from X. oryzae pv. oryzae was digested with EcoRI and analyzed forrestriction fragment length polymorphisms (RFLPs) with repetitive DNA element as a probe. Twenty-sevenRFLP types were identified; there was no overlap ofRFLP types among the six races from the Philippines. Mostvariability (20 RFLP types) was found in strains of races 1, 2, and 3, which were isolated from tropical lowlandareas. Four RFLP types (all race 5) were found among strains isolated from cultivars grown in the temperatehighlands. The genetic diversity of the total population ofX. oryzae pv. oryzae was 0.93, ofwhich 42% was dueto genetic differentiation between races. The genetic diversities of strains collected in 1972 to 1976, 1977 to 1981,and 1982 to 1986, were 0.89, 0.90, and 0.92, respectively, suggesting a consistently high level of variability inthe pathogen population over the past 15 years. Cluster analysis based on RFLP banding patterns showed fivegroupings at 85% similarity. The majority of strains from a given race were contained within one cluster,except for race 3 strains, which were distributed in three of the five clusters.
The host population in a plant pathogen system influencesthe genetic diversity and population structure of pathogens(5, 13, 15). In rice, single-gene resistance has been theprimary means of control for bacterial blight, caused byXanthomonas oryzae pv. oryzae (ex Ishiyama 1922) (27).There is concern that the widespread use of a few resistancegenes might accelerate the selection of new pathogenic racesand result in destabilization of crop production (4, 21).However, little is known regarding the genetic diversity andpopulation dynamics of X. oryzae pv. oryzae.
In previous studies, X. oryzae pv. oryzae populations inthe Philippines have been characterized on the basis ofpathogenicity patterns on five indica varieties of rice (16).Six races of X. oryzae pv. oryzae are presently recognized.While useful, race groupings provide little insight into thegenetic structure of the bacterial population. Physiologicaltraits provide little additional value in the assessment of X.oryzae pv. oryzae diversity since most strains are similarwith regard to many traits (28). Recently, restriction frag-ment length polymorphism (RFLP) analysis, which exploitsthe abundant variation in the DNA sequence, has been usedto generate a large number of markers for the measurementof genetic diversity in populations (9, 20). We previouslydescribed the existence of a repetitive sequence in the X.oryzae pv. oryzae genome that is in sufficient copy numberand distribution to provide a useful probe to detect restric-tion polymorphisms in the X. oryzae pv. oryzae genome (12).In order to better understand the population structure of X.oryzae pv. oryzae, we have used the repetitive element tomeasure the genetic diversity of the X. otyzae pv. oryzae
* Corresponding author.
populations and relate the diversity to previous character-izations of the pathogen population.
MATERIALS AND METHODS
Bacterial strains and culture conditions. The origin, date ofcollection, source variety, and race grouping for the X.oryzae pv. oryzae strains used in this study are shown inTable 1. All strains were isolated from naturally infectedleaves collected between 1972 and 1988 from different re-gions in the Philippines. Varietal names of the host plantswere obtained whenever possible. The race of each strainwas determined with a set of bacterial blight differentialvarieties (see below). Strains collected before 1982 weremaintained on Wakimoto's medium [0.5 g of Ca(NO3)2.4H20, 0.82 g of Na2HPO4, 5 g of peptone, 20 g of sucrose,300 g of potato, 15 g of agar per liter of water (24)] at -20°Cand revived yearly by transferring to fresh slants. Since1983, all stock cultures and newly collected strains were kepteither in 5% skim milk at -20°C or as lyophilized cultures.The strains were revived in fresh slants of modified Wakim-oto's medium (Wakimoto's medium without potato andsupplemented with 0.05 g of ferrous sulfate per liter) forsubsequent pathogenicity tests and DNA isolation.
Pathogenicity tests and race grouping. The pathogenicity ofeach culture of X. oryzae pv. oryzae was determined byinoculation of susceptible rice varieties IR8 or IR24. Racegrouping was based on virulence tests with the differentialvarieties IR8, IR20, Cas 209, IR1545-339, and DV85, whichcontain the bacterial blight resistance genes Xa-11, Xa-4,Xa-10, xa-5, and both xa-5 and Xa-7, respectively (16). Totest virulence, three plants per variety were grown in thegreenhouse, and the two youngest fully expanded leaves (35
2188
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

X. ORYZAE pv. ORYZAE POPULATION DIVERSITY, STRUCTURE 2189
TABLE 1. X oryzae pv. oryzae strains from the Philippines used in RFLP analysis
Strain Race groupa RFLP typeb Yr of collection Geographic origin (grouping)c Variety (grouping)d
Los Baflos, Laguna (1)Bay, Laguna (1)Los Bafios, Laguna (1)Los Bafios, Laguna (1)Casigan, Narra, Palawan (1)Los Banios, Laguna (1)Malicboy, Pagbilao, Quezon (1)Salvaci6n, Rosales, Pangasinan (1)Los Banios, Laguna (1)Barrio 11, Paoay, Ilocos Sur (1)Buguey, Cagayan (1)Bantay, Ilocos Sur (1)Atimonan, Quezon (1)Tapao, Sinait, Ilocos Sur (1)Culape, Lucban, Quezon (1)Tilib, Lucban, Quezon (1)Palawan (1)Casigan, Narra, Palawan (1)Malinao, Narra, Palawan (1)Isabela (1)Los Bafios, Laguna (1)Los Banios, Laguna (1)Mabitac, Laguna (1)Pototan, Iloilo (3)Los Bafios, Laguna (1)Talavera, Nueva Ecija (1)Ajuy, Iloilo (3)Camansihay, Palo, Leyte (3)Abuyog, Leyte (3)Iraan, Aborlan, Palawan (1)Santa Ignacia, Tarlac (1)Munioz, Nueva Ecija (1)Baliwag, Bulacan (1)Claveria, Misamis Oriental (4)Aborlan (site 3), Palawan (1)Midsayap, North Cotabato (4)Nueva Ecija (1)San Enrique, Negros Occidental (3)Davao (4)Panabo, Davao (4)Kagibikan, Pototan, Iloilo (3)Gines, Zarraga, Iloilo (3)Tramo, Nato Sangay, Camarines Sur (1)Goa, Camarines Sur (1)Moriones, Ocampo, Camarines Sur (1)BRCES,e Pili, Camarines Sur (1)Lopez, Bohol (3)Isabela (1)Isabela (1)Polangui, Albay (1)Bascaran, Daraga, Albay (1)Sorsogon, Sorsogon (1)Pawa, Tabaco, Albay (1)Palawan (1)Palawan (1)Los Bahios, Laguna (1)Los Banios, Laguna (1)Los Bafhos, Laguna (1)Palawan (1)Banaue, Ifugao (2)Banaue, Ifugao (2)Banaue, Ifugao (2)Banaue, Ifugao (2)Banaue, Ifugao (2)Banaue, Ifugao (2)Banaue, Ifugao (2)Banaue, Ifugao (2)Banaue, Ifugao (2)
IR841 (M)IR20(?) dwarf (M)Ketan Lemar (T)Dwarf (M)C22 (M)
Malagkit (T)C4-63G (M)
(T)IR5 (M)IR8 (M)
Wagwag (T)IR20 (M)IR27301-62-2 (M)IR14875-98-5 (M)IR48 (M)IR36 (M)IR480-5-9-3/IR20 (M)
IR30 (M)IR46 (M)
IR60 (M)
IR36 (M)Burik (T)
IR54 (M)IR26 (M)B3 (T)IR30 (M)Malagkit (T)
IR36 (M)IR50 (M)
IR42 (M)Mestiza (T)
IR46 (M)
BPI76 (T)Benzar (T)Balaratawee (T)IR8 (M)IR17525-150-2-2-1-2-3 (M)C4-63G (M)(M)(T)
F2 materials (M)JP5 (T)
IR3155 (M)Kaosiong 68 (M)
Continued on following page
PXO14PXO20PX084PXO1PXO151PXO61PXO165PX039PX0133PX0187PX0132PX0188PX0184PX0192PX035PX036PX068PX0157PXO170PX063PX0126PXO103IRN793PX0134PX086PX0172PX078PX0169PX0175PXO190PX0185PX0186PX0168PX0173PX0189PX0137PX083PXO171PX079PX0142PXO191PX0148PX0166PX0146PX0179PXO141PX0143PX087PX088PX0164PX0178PX0176PX0177PX069PXO70PX0129PXO113PXO125PXO71PX0198PX0197PX0182PXO80PXO107PXO112PXO105PxO110PXO180
111111111111111111122222222222222222223333333333333334444445555S55SS
1111111112222234S678888889999999999101112131415151515151516171718181920212121212121222222222222222222
19721972197619721985197319821972198219861979198619861986197219721974198519841973198119801985198119771985197519841985198619861986198419851986198219761984197519811986198519841984198619801982197419741979198619861986197419741982197819811974198819881986197519751978197519761986
VOL. 58, 1992
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 1-Continued
Strain Race groupa RFLP type' Yr of collection Geographic origin (grouping)' Variety (grouping)d
PX0195 5 22 1988 Banaue, Ifugao (2) (T)PX0194 5 22 1988 Banaue, Ifugao (2) (T)PX0196 5 22 1988 Banaue, Ifugao (2) (T)PX0181 5 22 1986 Banaue, Ifugao (2)PX0163 5 22 1985 Poblacion, Banaue, Ifugao (2) Tinawen (T)PXO111 5 22 1978 Banaue, Ifugao (2)PX0161 5 22 1985 Banaue Development Center, Banaue, Tinawen (T)
Ifugao (2)PX0162 5 22 1985 Bacoor, Banaue, Ifugao (2) Tinawen (T)IRN394 5 23 1981 IRCTNf Banaue, Ifugao (2) Tinawen (T)IRN753 5 23 1985 IRCTN, Banaue, Ifugao (2) Dakpa (T)PXO154 5 23 1981 IRCTN, Banaue, Ifugao (2) C16486 (T)IRN392 5 23 1981 IRCTN, Banaue, Ifugao (2) Gaebyeo (M)PX0145 5 24 1982 Bontoc, Mt. Province (2) (T)PX0144 5 24 1982 Bontoc, Mt. Province (2) (T)PXO130 5 24 1981 Banaue, Ifugao (2) IR line (IRCTN#9745) (M)PX0193 5 25 1988 Banaue, Ifugao (2) (T)PX0183 5 25 1986 Banaue, Ifugao (2)PXO116 6 26 1979 Los Bafios, Laguna (1) CR-157-392-284 (M)PXO117 6 26 1979 Los Bafios, Laguna (1) B-2277C-MR-99-2 (M)PXO114 6 26 1979 Los Banios, Laguna (1)PXO118 6 26 1979 Los Bafios, Laguna (1) B2540b-Pn-20-2 (M)PXO119 6 26 1979 Los Bafios, Laguna (1) 3782-79 (M)PXO115 6 27 1979 Los Bafios, Laguna (1)PX0123 6 27 1980 Los Bafios, Laguna (1) CR-157-392-4 (M)PX0127 6 27 1982 Los Bafios, Laguna (1) IR8 (M)PX0128 6 27 1982 Los Bafios, Laguna (1) IR442 (M)PX099 6 27 1980 Los Banios, Laguna (1) Rexoro (T)PX0124 6 27 1980 Los Bafios, Laguna (1) IR20878-R (M)PX0122 6 27 1980 Los Bafios, Laguna (1) Dourado Precose (T)PXO121 6 27 1979 Los Bafios, Laguna (1)
a Race was determined by inoculation to the rice differential cultivars IR8 (Xa-11), IR20 (Xa-4), IR1545-339 (xa-S), DV85 (xa-5Xa-7), and Cas 209 (Xa-10), asdescribed in Materials and Methods.
b RFLP type was determined from Southern blot analysis of EcoRI-digested total DNA from strains of X. oryzae pv. oryzae with pJEL101 as a probe.c Geographic origins of strains were grouped according to island clusters as follows: 1, Luzon lowland; 2, Luzon highland; 3, Visayas; and 4, Mindanao.d Where available, the name of the variety is given. Modem varieties (M) include released varieties and crosses or lines made by the International Rice Research
Institute and the University of the Philippines, Los Bafios, Philippines. Modem varieties are shorter (semidwarf), less leafy, more responsive to nitrogen, andmore resistant to lodging than traditional varieties (T). -, variety unknown.
e Bicol Rice and Corn Experiment Station.f International Rice Cold Tolerance Screening Nursery.
to 40 days after sowing) were clip inoculated as describedelsewhere (24). Bacteria, grown for 72 h at 28°C on modifiedWakimoto's medium, were suspended in distilled water,adjusted to 109 CFU/ml, and used as an inoculum. Diseasereactions were scored by both percent lesion area and lesionlength 14 days after inoculation (17).DNA analysis. The DNA probe was pJEL101, a pUC18
plasmid containing a 2.4-kb EcoRI-HindIII fragment derivedfrom race 2 strain PX086 of X. oryzae pv. oryzae (12). Theinsert of pJEL101 contains a high-copy-number repetitiveDNA element present in DNA from over 100 X. oryzae pv.oxyzae strains tested (12). The plasmid was maintained inEscherichia coli TB1 under selection of 50 ,g of carbenicillinper ml, extracted by alkaline lysis, and purified by cesiumchloride centrifugation as described previously (14).Genomic DNA from strains of X. oryzae pv. oryzae was
isolated by a lysozyme-sodium dodecyl sulfate (SDS) lysisprocedure of Owen and Borman (25) modified as describedelsewhere (12). Genomic DNA from X. oryzae pv. oryzaewas digested to completion by EcoRI (2 U of enzyme per ptgof DNA) according to the manufacturer's direction (Be-thesda Research Laboratories, Gaithersburg, Md., orPromega Biotech, Madison, Wis.). DNA was fractionated bygel electrophoresis (horizontal 0.7% agarose gel, 20 by 21.5
cm) in Tris-borate buffer (89 mM Tris-HCI, 89 mM boricacid, and 2 mM Na2EDTA, pH 8.0). A 1-kb ladder (BethesdaResearch Laboratories) was included in gels as a size stan-dard. Southern transfer onto a nylon membrane was doneaccording to instructions of the manufacturer (GeneScreenPlus; Du Pont Co., Wilmington, Del.). The entire VJEL101plasmid was labeled (specific activities between 10 and 108cpm/,ug of DNA) with [32P]dCTP by using a nick translationkit (Bethesda Research Laboratories); vector (pUC18) DNAdoes not hybridize with X. oryzae pv. oryzae genomic DNA(12). Hybridization and washing conditions (12) were highstringency. Hybridization was performed at 65°C in a mix-ture of 0.1% SDS, 50 mM sodium phosphate buffer (pH 7.0),1.0 M NaCl, and 300 ,ug of denatured salmon sperm DNAper ml for 18 h. After hybridization, the blots were washedthree times (20 min each) at 65°C in 2x SSC (1x SSC is 0.15M NaCl plus 0.015 sodium citrate) containing 0.1% SDS and5 mM sodium phosphate buffer (pH 7.0) and three times in0.5 x SSC containing 0.1% SDS and 3 mM sodium phosphatebuffer. Autoradiography was done at -80°C with Cronexfilm (Du Pont) with Cronex Lightning-Plus intensifyingscreens (Du Pont). DNA from strains of one race grouptogether with strains representing each of the six describedraces from the Philippines (16) were analyzed in the same
2190 LEACH ET AL.
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

X ORYZAtE pv. ORYZAE POPULATION DIVERSITY, STRUCTURE 2191
gel. Distinct RFLP types were analyzed in a single blot toconfirm that each RFLP type was unique.
Data analysis. To determine the genetic relationshipsamong RFLP types, the presence or absence of bands at 38different positions was converted into binary data; i.e., thepresence or absence of a band was coded as 1 or 0,respectively. Density of the bands was not considered in theanalysis. Band positions more than 2 mm apart were consid-ered different. DNA from a set of four isolates (PX061,PX086, PX0112, and PX099), EcoRI-HindIII-digestedpJEL101, and 1-kb ladder (Bethesda Research Laboratories)were included in each gel as standards. Each strain wastested on at least two blots done at different times to confirmdifferences. The number of bands scored per strain rangedfrom 17 for PX035 to 9 for PX078. Similarity coefficientswere calculated for all pairwise combinations by using asimple matching coefficient (26). Cluster analysis of thesimilarity coefficients was done by both Ward's method (30)and an unweighted pair group method (26) using a Clustan3.2/PC computer program (32).The haplotype diversity approach of Nei and Tajima (23)
was used to analyze the population structure of X. oryzaepv. oryzae. Haplotypic diversity measures the probability ofobtaining two randomly chosen strains with different RFLPpatterns and therefore has the same biological meaning andstatistical properties as genetic diversity. Hereafter, haplo-typic diversity is referred to as genetic diversity in describingpopulations of X. oryzae pv. oryzae. The genetic diversity(H) for each grouping of strains, i.e., race (HR), time ofcollection (HY), and geographic origin (HG), and the geneticdiversity for the total population of X. oryzae pv. oryzae(HT) were estimated by the following equation (22): H =[n/(n - 1)](1 - YXi7), where Xi is the frequency of the ithRFLP type in any one grouping (for determination of HR.HY, or HG), or the average frequency of the ith RFLP typein the entire population (for determination of HT), and n isthe number of strains examined. The extent of geneticdifferentiation among groups was estimated by the coeffi-cient of genetic differentiation G (7): G = (HT - HX)/HT,where HX is the average genetic diversity of strain groupingsby race (HR), time of collection (HY), or geographic origin(HG).
RESULTS
RFLP analysis. We examined 98 strains of X. oryzae pv.oryzae for DNA polymorphisms with the repetitive DNAelement in pJEL101. Twenty-seven distinct banding patternswere observed (Fig. 1), and on the basis of comparison withthese patterns, each strain was assigned an RFLP type (1 to27, Table 1). No single RFLP type was present in any twodifferent races (Table 2). Within a race grouping, the RFLPtypes sometimes differed only at one banding position. Forexample, the race 1 strains which were grouped in RFLPtype 1 varied from those in RFLP types 5 and 6 (Fig. lb) atsingle band positions. Other RFLP types within a race weredistinct from one another at several banding positions; forexample, 12 positions vary between RFLP type 1 (race 1)and RFLP type 3 (race 1). In a few cases, patterns betweentwo races were similar; for example, RFLP type 9 (race 2)differed from RFLP type 23 (race 5) at one band position(Fig. lc).On the basis of the occurrence of RFLP types, races 1 and
3 were the most heterogeneous groups, with seven and eightdifferent types in 19 and 15 strains tested, respectively. Race
Rac 1 1 1 1 1 1 1 2 2 2 2 2 3 3 3 3 3 3 3 3 4 5 5 I 5 6 6
6 6
5 j5
1.6
5 1-
a8 -
4
1 5 5 5 5 5 2 B2452w~~~~~~~~~~-ob_
4'* am 4*444.5
-~~~_ _.i_ * --cAw1_ ~
NE
44
3- W- W wM _ _
FIG. 1. Southern blot analysis of total DNA from strains of X.o?yzae pv. oryzae representing the 27 RFLP types. The probe was32P-labeled pJEL101. (A) X. oryzae pv. oryzae strains and RFLPtype are as follows: PXO61, type 1; PX0132, type 2; PX035, type 3;PX036, type 4; PX068, type 5; PX0157, type 6; PXO170, type 7;PX063, type 8; PX078, type 9; PX0137, type 10; PX083, type 11;PXO171, type 12; PX079, type 13; PX0142, type 14; PXO141, type15; PX0143, type 16; PX087, type 17; PX0164, type 18; PX0176,type 19; PX0177, type 20; PX0125, type 21; PXO80, type 22;PX0154, type 23; PX0183, type 25; PXO114, type 26; and PXO115,type 27. Lane J, EcoRI-HindIII-digested pJEL101; lanes k, 1-kbladder. (B) Differences in banding between RFLP types 1 and 6(fragment a) and types 9, 23, and 24 (fragments b and c).
4 was the most homogeneous, with one RFLP type amongthe six strains tested.To determine the genetic relationships among strains of
races, the banding patterns were analyzed by cluster analy-sis. All strains of X. oryzae pv. oryzae were similar to eachother at a level of greater than 70% with the exception of onerace 1 strain, PX035 (similarity level of 63%). At 85%similarity, five clusters of strains were observed in the 27RFLP types (Fig. 2). Races represented in the five clusterswere as follows: cluster I, races 1, 3, and 4; cluster II, races2, 3, and 5; cluster III, race 3; cluster IV, race 6; and clusterV. race 1. Race 3 strains were present in three of the fiveclusters. At this level of similarity (85%), only race 6 wasfound in a unique cluster; at the 88% level of similarity, race4 strains formed a unique cluster. The majority of strainsfrom a given race were contained within one cluster at 85%similarity, i.e., race 1 (18 of 19 strains) in cluster I, race 4 (6
VOL. 58, 1992
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 2. Frequencies of RFLP types ofX oryzae pv. oryzae according to race grouping and the estimated geneticdiversity of each group
Total Frequency of RFLP type in each race grouping"
RFLP typea Type population 1 2 3 4 5 6strainNo. of % of No. of % of No. of % of No. of % of No. of % of No. of % of No. of % ofstrains race strains race strains race strains race strains race strains race strains race
1 PXO61 9 9.2 9 47.32 PX0132 5 5.1 5 26.33 PXO35 1 1.0 1 5.34 PX036 1 1.0 1 5.35 PX068 1 1.0 1 5.36 PXO157 1 1.0 1 5.37 PXO170 1 1.0 1 5.38 PX063 6 6.1 6 31.59 PX078 10 10.2 10 52.610 PX0137 1 1.0 1 5.311 PX083 1 1.0 1 5.312 PXO171 1 1.0 1 5.313 PX079 1 1.0 1 6.714 PX0142 1 1.0 1 6.715 PXO141 6 6.1 6 40.016 PX0143 1 1.0 1 6.617 PX087 2 2.0 2 13.318 PX0164 2 2.0 2 13.319 PX0176 1 1.0 1 6.720 PX0177 1 1.0 1 6.721 PXO125 6 6.1 6 100.022 PXO80 17 17.3 17 65.423 PXO154 4 4.1 4 15.424 PXO145 3 3.1 3 11.525 PX0183 2 2.0 2 7.726 PXO114 5 5.1 5 38.527 PXO115 8 8.1 8 61.5
Total no. of 98 19 19 15 6 26 13strains
Genetic diversity' 0.93 0.73 0.65 0.84 0 0.55 0.51
a RFLP type was determined from Southern blot analysis of EcoRI-digested, total DNA from strains of X. oryzae pv. oryzae by using pJEL101 as a probe.b Race was determined by inoculation to the rice differentials IR8 (Xa-11), IR20 (Xa-4), IR1545-339 (xa-5), DV85 (xa-5Xa-7), and Cas 209 (Xa-10) as described
in Materials and Methods.c Genetic diversity is calculated by the formula [n/(n - 1)1(1 - YXi2), where Xi is the proportion of the ith RFLP type within a group and n is the number of
strains tested in each group.
of 6 strains) in cluster I, race 2 (19 of 19 strains) in cluster II,race 5 (26 of 26 strains) in cluster II, race 3 (8 of 15 strains)in cluster III, and race 6 (13 of 13 strains) in cluster IV.
Genetic diversity. The genetic diversity (HT) of the totalpopulation of X. oryzae pv. oryzae in the Philippines wasestimated to be 0.93. Genetic diversity calculated for eachrace treated as a subpopulation (HR) was highest for race 3(0.84) and lowest for race 4 (0.0) (Table 2). The averagegenetic diversity (HR) of six race groups was 0.54, and thecoefficient of genetic differentiation (GR) was 0.41.To examine the effect of varietal release over the past 15
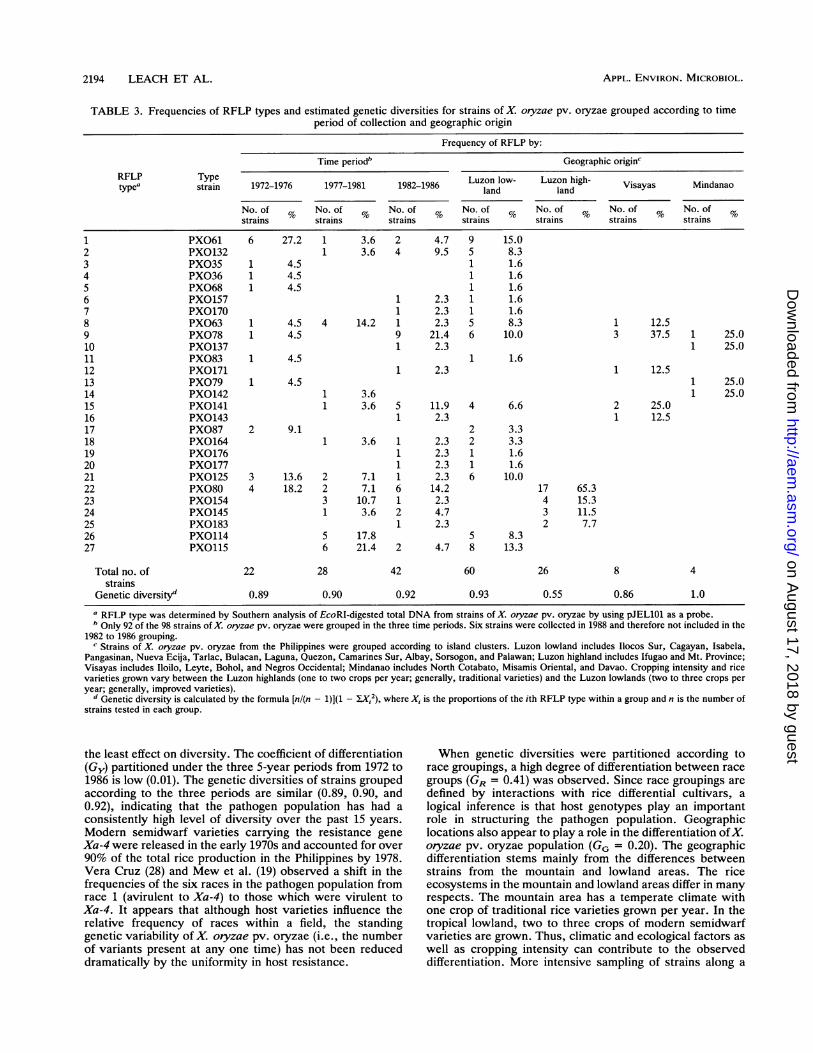
years on the diversity of the population of X. oryzae pv.oryzae, genetic diversities of strains collected in three 5-yearperiods (1972 to 1976, 1977 to 1981, and 1982 to 1986) werecalculated (Table 3). The genetic diversities of strains sam-pled between 1972 and 1976 (25% of the total samples), 1977and 1981 (30% of the total), and 1982 and 1986 (44% of thetotal) were 0.89, 0.90, and 0.92, respectively (Table 3). Thecoefficient of genetic differentiation (Gy) based on the group-ing of time periods was 0.01.On the basis of information collected at the time of
sampling, the varietal origins were classified as either mod-ern (semidwarf) varieties or traditional varieties (Table 1).
Rice breeders consider modern varieties shorter, more re-sponsive to nitrogen, and more resistant to lodging than thetraditional varieties. No difference was observed in thegenetic diversities of strains recovered from traditional(0.90) or modern (0.91) varieties.When strains were grouped according to their geographic
origin in the Philippines, a marked difference in geneticdiversity (HG) was found between strains from the Luzonlowlands (0.93) and the Luzon highlands (0.55) (Table 3).Only four RFLP types were identified among 26 strainsobtained from the highlands, whereas 19 RFLP types werefound among 60 strains tested in the lowlands in LuzonIsland (Table 2). The coefficient of genetic differentiation(GG) based on geographic locations was 0.20.
DISCUSSION
We have used a repetitive DNA element to characterizethe population structure of X. oryzae pv. oryzae in thePhilippines. Two factors aided the analysis. First, the ran-dom genomic distribution of the repetitive element pJEL101(12) provided a single DNA probe to scan for DNA polymor-phisms and saved considerable effort in typing the strains
2192 LEACH ET AL.
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

X. ORYZAE pv. ORYZAE POPULATION DIVERSITY, STRUCTURE 2193
PXO 61 (1)PXO 68 (I)PX0 157()PXOI170()PXO 176 (3)PX0132 (I)PXO 36 (I)PX0142 (3)PXO 164 (3)PXO 87(3)PXO 125 (4)PXO 63 (2)PX0137 (2)PXO171 (2)PX0145 (5)-PXO 83 (2)PX0154 (5)PXO 78(2)PXO 79(3)PXO 80(5)PXO 183 (5)PXO 141 (3)PXO 143 (3)PX0177 (3)PXOI 14(6)PXOI I 5 (6)PXO 35(I)
100 90 80Similarity (N)
FIG. 2. Dendrogram displaying the relationshifRFLP types for strains of X. oryzae pv. oryzae aftanalysis using 32P-labeled pJEL101 as a probe. Thefor each strain is shown in parentheses. Similaritygrouped by using the unweighted pair group metho'means.
compared with the use of single-copy or lo,DNA probes. Second, the background inform;the time, location of collection, and host origi:of X. oryzae pv. oryzae from the Philippinesfrom the International Rice Research InpJEL101 as a hybridization probe, we obseassociation between RFLP types and raceRFLP type was shared among the six racesthe criteria used in this study (Table 2); ho'RFLP types were observed within some racsome hybridization patterns were very siistrains of different races. In an asexual haplcnonrandom associations of genes can be du(epistatic selection of the genotypes involveabsence of epistatic selection, a fast-evolvwould be found nested within a more static phoryzae pv. oryzae, we observe much greatemolecular phenotypes than in pathogenic patlence loci and the genomic distribution ofelement are not functionally related, our obs4suggest that the molecular phenotypes, a:pJEL101 hybridization patterns, change at a havirulence loci. However, comparisons betmphenotype and avirulence loci must take int
measurement of diversity at avirulence loci is limited by thenumber of rice differential cultivars available for definingrace. Alternatively, the repetitive element may play a role inrace differentiation. Although we have no evidence to sug-gest that the repetitive element might be functionally asso-ciated with avirulence, sequence analysis of pJEL101showed that it contained structural features common inprokaryotic transposable elements (33). Kearney et al. (10)demonstrated that a transposable element from Xanthomo-nas campestns pv. vesicatoria could insert into and inacti-vate a plasmid-borne avirulence gene, thereby affecting theinteraction of the pathogen with pepper plants containingresistance genes corresponding to the avirulence gene. Workis in progress to determine whether the repetitive elementcontained in pJEL101 could direct or influence the evolutionof races of X. oryzae pv. oryzae.Although the functional relationship between avirulence
and repetitive elements remains unknown, the dendrogramconstructed on the basis of RFLP banding patterns providesnew insights into the evolutionary relationships amongstrains of X. oryzae pv. oryzae (Fig. 2). Race 1 (cluster I)was the predominant race before the widespread use of thebacterial blight resistance gene Xa-4 (18). Races 3 and 4,which are virulent to Xa-4, are also contained in cluster I.PX035 (cluster V) is classified as race 1 on the basis of host
J reactions but is different from other race 1 strains withrespect to RFLP patterns and serological reactions (3). It ispossible that PX035 will be grouped as a different race whentested against new differential hosts. In fact, strains withvery different RFLP patterns proved to be good candidates
....L......... | for a new race group and were useful in screening for new70 60 sources of resistance (29).
Race 3, which includes the most diverse group of strains,)s among the 27 is present in all clusters except IV (race 6 strains only) and Ver Southern blot (single strain PX035). It is possible that race 3 phenotypesrace designation arose in separate, unrelated populations. Alternatively, it iscoefficients were possible that race 3 has existed long enough to accumulate aJ with arithmetic high degree of genetic variation. Nitrosoguanidine mutagen-
esis experiments yielded mutants which changed from race 2to race 3 (1, 11), indicating that the differences between thetwo phenotypes may be controlled by a single avirulencelocus. Additional DNA markers, including cloned avirulence
v-copy-number genes (10a) and other repetitive DNA elements (2, 12), areation regarding being used to further elucidate the phylogenetic relationshipsns of 98 strains between races.were available The genetic diversity (HT = 0.93) estimated for the totalistitute. Using population of X. oryzae pv. oryzae indicates a high level of-rved a strong variability in the pathogen population. The high level ofgroupings. No diversity among X. oryzae pv. oryzae strains has been con-on the basis of firmed with different DNA probes (23a). Such a high degree ofwever, several variability has also been observed for other phytopathogenic.e groups, and bacteria. Denny et al. (8) examined 17 Pseudomonas synngaemilar between pv. tomato and six P. syningae pv. syringae strains by RFLP)id population, analysis and found each strain to have a unique RFLP. Usinge to chance or nine DNA probes, seven of which encode information re-d (31). In the quired for virulence and the hypersensitive response, Cook eting phenotype al. (6) identified 28 RFLP patterns among 62 strains ofienotype. In X. Pseudomonas solanacearum representing three races and.r variations in five biovars. It would be of interest to determine whether the:terns. If aviru- high level of diversity in the genomes of plant pathogenicthe repetitive bacteria is the consequence of host genotype variability.ervation would By partitioning genetic diversities within race groupings,s revealed by time periods, and geographic locations of our limited sampleiigher rate than of X. oryzae pv. oryzae strains, we gain clues to the relativeveen molecular significance of these factors in shaping the population struc-;o account that ture of the pathogen. Of the three factors examined, time had
VOL. 58, 1992
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
TABLE 3. Frequencies of RFLP types and estimated genetic diversities for strains of X. oryzae pv. oryzae grouped according to timeperiod of collection and geographic origin
Frequency of RFLP by:
Time periodb Geographic origin'
RFLP Type Luzon low- Luzon high-typea strain 1972-1976 1977-1981 1982-1986 Land land Visayas Mindanao
No. of % No. of % No. of % No. of % No. of % No. of %strains strains strains strains strains strains strains
1 PXO61 6 27.2 1 3.6 2 4.7 9 15.02 PX0132 1 3.6 4 9.5 5 8.33 PXO35 1 4.5 1 1.64 PX036 1 4.5 1 1.65 PX068 1 4.5 1 1.66 PX0157 1 2.3 1 1.67 PXO170 1 2.3 1 1.68 PX063 1 4.5 4 14.2 1 2.3 5 8.3 1 12.59 PX078 1 4.5 9 21.4 6 10.0 3 37.5 1 25.010 PX0137 1 2.3 1 25.011 PX083 1 4.5 1 1.612 PXO171 1 2.3 1 12.513 PX079 1 4.5 1 25.014 PX0142 1 3.6 1 25.015 PXO141 1 3.6 5 11.9 4 6.6 2 25.016 PX0143 1 2.3 1 12.517 PX087 2 9.1 2 3.318 PX0164 1 3.6 1 2.3 2 3.319 PX0176 1 2.3 1 1.620 PX0177 1 2.3 1 1.621 PXO125 3 13.6 2 7.1 1 2.3 6 10.022 PXO80 4 18.2 2 7.1 6 14.2 17 65.323 PXO154 3 10.7 1 2.3 4 15.324 PXO145 1 3.6 2 4.7 3 11.525 PX0183 1 2.3 2 7.726 PXO114 5 17.8 5 8.327 PXO115 6 21.4 2 4.7 8 13.3
Total no. of 22 28 42 60 26 8 4strains
Genetic diversityd 0.89 0.90 0.92 0.93 0.55 0.86 1.0
a RFLP type was determined by Southern analysis of EcoRI-digested total DNA from strains of X. oryzae pv. oryzae by using pJEL101 as a probe.h Only 92 of the 98 strains of X. oryzae pv. oryzae were grouped in the three time periods. Six strains were collected in 1988 and therefore not included in the
1982 to 1986 grouping.c Strains of X. oryzae pv. oryzae from the Philippines were grouped according to island clusters. Luzon lowland includes Ilocos Sur, Cagayan, Isabela,
Pangasinan, Nueva Ecija, Tarlac, Bulacan, Laguna, Quezon, Camarines Sur, Albay, Sorsogon, and Palawan; Luzon highland includes Ifugao and Mt. Province;Visayas includes Iloilo, Leyte, Bohol, and Negros Occidental; Mindanao includes North Cotabato, Misamis Oriental, and Davao. Cropping intensity and ricevarieties grown vary between the Luzon highlands (one to two crops per year; generally, traditional varieties) and the Luzon lowlands (two to three crops peryear; generally, improved varieties).
d Genetic diversity is calculated by the formula [nl(n - 1)1(1 - l;X/), where Xi is the proportions of the ith RFLP type within a group and n is the number ofstrains tested in each group.
the least effect on diversity. The coefficient of differentiation(Gy) partitioned under the three 5-year periods from 1972 to1986 is low (0.01). The genetic diversities of strains groupedaccording to the three periods are similar (0.89, 0.90, and0.92), indicating that the pathogen population has had aconsistently high level of diversity over the past 15 years.Modem semidwarf varieties carrying the resistance geneXa-4 were released in the early 1970s and accounted for over90% of the total rice production in the Philippines by 1978.Vera Cruz (28) and Mew et al. (19) observed a shift in thefrequencies of the six races in the pathogen population fromrace 1 (avirulent to Xa-4) to those which were virulent toXa-4. It appears that although host varieties influence therelative frequency of races within a field, the standinggenetic variability of X. oryzae pv. oryzae (i.e., the numberof variants present at any one time) has not been reduceddramatically by the uniformity in host resistance.
When genetic diversities were partitioned according torace groupings, a high degree of differentiation between racegroups (GR = 0.41) was observed. Since race groupings aredefined by interactions with rice differential cultivars, alogical inference is that host genotypes play an importantrole in structuring the pathogen population. Geographiclocations also appear to play a role in the differentiation ofX.oryzae pv. oryzae population (GG = 0.20). The geographicdifferentiation stems mainly from the differences betweenstrains from the mountain and lowland areas. The riceecosystems in the mountain and lowland areas differ in manyrespects. The mountain area has a temperate climate withone crop of traditional rice varieties grown per year. In thetropical lowland, two to three crops of modern semidwarfvarieties are grown. Thus, climatic and ecological factors aswell as cropping intensity can contribute to the observeddifferentiation. More intensive sampling of strains along a
2194 LEACH ET AL.
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

X. ORYZAE pv. ORYZAE POPULATION DIVERSITY, STRUCTURE 2195
transect across the mountain and lowland areas is beingmade to investigate the fine-scale variation of X. otyzae pv.oryzae under different ecological conditions (23a).The ability to describe pathogen populations over time and
space and to understand the impacts of host genotype,climate, and cropping intensity on pathogen variability canbe of practical use for the deployment of host resistance. Atpresent, the release of new varieties is based primarily onmultilocation yield trials and on the observed level of diseaseand insect resistance under trial conditions. Detailed de-scriptions of race composition, genetic variability, and thegenetic potential to change in the pathogen population havenot been available to research scientists, farmers, or exten-sion workers in planning varietal release. The availability ofsuitable probes such as the repetitive element in pJEL101 tocharacterize different pathogen populations from broad geo-graphic areas will provide useful information for the testingand release of resistant germ plasm among rice-growingcountries.
ACKNOWLEDGMENTS
This research is supported by grants from the Rockefeller Foun-dation (RF86058 #56 to KSU and RF86058 #44 to IRRI), theKansas Agricultural Experiment Station (contribution no. 92-341-J)and Washington State Agricultural Research Center (contributionno. PPNS no. 0116).We thank Rebecca Nelson and Scot Hulbert for their helpful
comments on the manuscript.
REFERENCES1. Ardales, E., N. Chua, J. Leach, and H. Leung. 1988. Isolation of
chemically-induced mutants of Xanthomonas campestns pv.oryzae with changed race-specificity, p. 103. Fifth InternationalCongress on Plant Pathology, Kyoto, Japan. (Abstr.)
2. Baraoidan, M. R., R. Nelson, J. E. Leach, T. W. Mew, and H.Leung. 1990. Use of putative transposable elements as probesfor population studies of the bacterial blight pathogen of rice.Phytopathology 80:982. (Abstr.)
3. Benedict, A. A., A. M. Alvarez, J. Berestecky, W. Imanaka,C. Y. Mizumoto, L. W. Pollard, T. M. Mew, and C. F. Gonzalez.1989. Pathovar-specific monoclonal antibodies for Xanthomo-nas campestns pv. oryzae and for Xanthomonas campestris pv.oryzicola. Phytopathology 79:322-328.
4. Browning, J. A., and K. J. Frey. 1969. Multiline cultivars as ameans of disease control. Annu. Rev. Phytopathol. 7:355-382.
5. Burdon, J. J., and A. M. Jarosz. 1990. Disease in mixedcultivars, composites, and natural plant populations: someepidemiological and evolutionary consequences, p. 215-228. InA. H. D. Brown, M. T. Clegg, A. L. Kahler, and B. S. Weir(ed.), Plant population genetics, breeding, and genetic re-sources. Sinauer Associates Inc., Sunderland, Mass.
6. Cook, D., E. Barlow, and L. Sequeira. 1989. Genetic diversity ofPseudomonas solanacearum: detection of restriction fragmentlength polymorphisms with DNA probes that specify virulenceand the hypersensitive response. Mol. Plant-Microbe Interact.2:113-121.
7. Crow, J. F. 1986. Basic concepts in population, quantitative andevolutionary genetics. W. H. Freeman, San Francisco.
8. Denny, T. P., M. N. Gilmor, and R. K. Selander. 1988. Geneticdiversity and relationships of two pathovars of Pseudomonassyringae. J. Gen. Microbiol. 134:1949-1960.
9. Gabriel, D. W., J. E. Hunter, M. T. Kingsley, J. W. Miller, andG. R. Lazo. 1988. Clonal population structure of Xanthomonascampestns and genetic diversity among citrus canker strains.Mol. Plant-Microbe Interact. 1:59-65.
10. Kearney, B., P. C. Ronald, D. Dahlbeck, and B. J. Staskawicz.1988. Molecular basis for evasion of plant host defence inbacterial spot disease of pepper. Nature (London) 332:541-543.
10a.Leach, J. E., et al. Unpublished data.
11. Leach, J. E., S. Kelemu, E. Ardales, and H. Leung. 1989.Bacterial blight of rice, p. 199-206. International Rice ResearchInstitute, Manila, Philippines.
12. Leach, J. E., F. W. White, M. L. Rhoads, and H. Leung. 1990.A repetitive DNA sequence differentiates Xanthomonascampestris pv. oryzae from other pathovars of Xanthomonascampestris. Mol. Plant-Microbe Interact. 3:238-246.
13. Leonard, K. J., and R. J. Czochor. 1980. Theory of geneticinteractions among populations of plants and their pathogens.Annu. Rev. Phytopathol. 18:237-258.
14. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
15. Marshall, D. R. 1977. The advantages and hazards of genetichomogeneity. Ann. N.Y. Acad. Sci. 287:1-20.
16. Mew, T. W. 1987. Current status and future prospects ofresearch on bacterial blight of rice. Annu. Rev. Phytopathol.25:359-382.
17. Mew, T. W., R. C. Reyes, and C. M. Vera Cruz. 1989. Screeningfor bacterial blight resistance in rice, p. 338-341. In Z. Klement,K. Rudolph, and D. C. Sands (ed.), Methods in phytobacteri-ology. Akademiae Kiado, Budapest.
18. Mew, T. W., and C. M. Vera Cruz. 1985. Virulence of Xanth-omonas campestris pv. oryzae in the Philippines. Phytopathol-ogy 75:1316. (Abstr.)
19. Mew, T. W., C. M. Vera Cruz, and E. S. Medalla. 1990. Ananatomy of the bacterial blight epidemics in the Philippines.Proc. 3rd Int. Conf. Plant Prot. Tropics, 20-23 March 1990,Malaysia.
20. Michelmore, R. W., and S. Hulbert. 1987. Molecular markers forgenetic analysis of phytopathogenic fungi. Annu. Rev. Phyto-pathol. 25:383-404.
21. National Academy of Sciences. 1972. Genetic vulnerability ofmajor crops. National Academy of Sciences USA, Washington,D.C.
22. Nei, M. 1987. Molecular evolutionary genetics. Columbia Uni-versity Press, New York.
23. Nei, M., and F. Tajima. 1981. DNA polymorphism detectable byrestriction endonucleases. Genetics 97:145-163.
23a.Nelson, R. J. IRRI, Los Bafios, Philippines. Personal commu-nication.
24. Ou, S. H. 1985. Rice diseases. Commonwealth MycologicalInstitute, Kew, Surrey, United Kingdom.
25. Owen, R. J., and P. Borman. 1987. A rapid biochemical methodfor purifying high molecular weight bacterial chromosomalDNA for restriction enzyme analysis. Nucleic Acids Res. 15:3631.
26. Sneath, P. H. A., and R. R. Sokal. 1973. Numerical taxonomy.W. H. Freeman and Co., San Francisco.
27. Swings, J., M. Van den Mooter, L. Vauterin, B. Hoste, M. Gillis,T. W. Mew, and K. Kersters. 1990. Reclassification of the causalagents of bacterial blight (Xanthomonas campestris pv. oryzae)and bacterial leaf streak (Xanthomonas campestris pv. oryzi-cola) of rice as pathovars of Xanthomonas oryzae (ex Ishiyama1922) sp. nov., nom. rev. Int. J. Syst. Bacteriol. 40:309-311.
28. Vera Cruz, C. M. 1984. Bacteriological and pathological varia-tion ofXanthomonas campestris pv. oryzae (Ishiyama) Dye, thepathogen of bacterial blight of rice. M.S. thesis. University ofthe Philippines, Los Banios, Philippines.
29. Vera Cruz, C. M., R. Nelson, H. Leung, J. Leach, and T. W.Mew. 1992. Reaction of rice cultivars from Ifugao Province,Philippines to indigenous strains of the bacterial blight patho-gen. Int. Rice Res. Newsl. 17(2):8-9.
30. Ward, J. H. 1963. Heirarchical grouping to optimize an objec-tive function. J. Am. Stat. Assoc. 58:236-244.
31. Whittam, T. S., H. Ochman, and R. K. Selander. 1983. Geo-graphic components of linkage disequilibrium in natural popu-lations of Eschenichia coli. Mol. Biol. Evol. 1:67-83.
32. Wishart, D. 1987. Clustan user manual, 4th ed. University of St.Andrews, Edinburgh.
33. Yun, C. H. 1991. Molecular characterization of a repetitiveelement of Xanthomonas oryzae pv. oryzae. Ph.D. thesis.Kansas State University, Manhattan.
VOL. 58, 1992
on August 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from





























