Embed Size (px)
Citation preview

Leptin Enhances the Calcification of Vascular CellsArtery Wall as a Target of Leptin
Farhad Parhami, Yin Tintut, Alex Ballard, Alan M. Fogelman, Linda L. Demer
Abstract—Leptin, the product of theob gene, regulates food intake, energy expenditure, and other physiological functionsof the peripheral tissues. Leptin receptors have been identified in the hypothalamus and in extrahypothalamic tissues.Increased circulating leptin levels have been correlated with cardiovascular disease, obesity, aging, infection withbacterial lipopolysaccharide, and high-fat diets. All these conditions have also been correlated with increased vascularcalcification, a hallmark of atherosclerotic and age-related vascular disease. In addition, the differentiation of marrowosteoprogenitor cells is regulated by leptin. Thus, we hypothesized that leptin may regulate the calcification of vascularcells. In this report, we tested the effects of leptin on a previously characterized subpopulation of vascular cells thatundergo osteoblastic differentiation and calcification in vitro. When treated with leptin, these calcifying vascular cellshad a significant 5- to 10-fold increase in alkaline phosphatase activity, a marker of osteogenic differentiation ofosteoblastic cells. Prolonged treatment with leptin enhanced the calcification of these cells, further supporting thepro-osteogenic differentiation effects of leptin. Furthermore, the presence of the leptin receptor on calcifying vascularcells was demonstrated using reverse transcriptase polymerase chain reaction, immunocytochemistry, and Western blotanalysis. We also identified the presence of leptin receptor in the mouse artery wall, localized to subpopulations ofmedial and adventitial cells, and the expression of leptin by artery wall cells and atherosclerotic lesions in mice. Takentogether, these results suggest that leptin regulates the osteoblastic differentiation and calcification of vascular cells andthat the artery wall may be an important peripheral tissue target of leptin action.(Circ Res. 2001;88:954-960.)
Key Words: calcification n calcifying vascular cellsn leptin n atherosclerosis
L eptin, the circulating protein product of theob gene, wasrecently shown to be a satiety factor regulating food
intake and energy expenditure.1 Although it is producedprimarily by adipocytes and has a primary effect on thehypothalamus, it is also produced by nonadipocytic cells1 andtargets extrahypothalamic tissues.2,3 It has effects on lipidmetabolism, hematopoiesis, pancreatic cell function, thermo-genesis, and the response to bacterial lipopolysaccharide.4,5
The wide role of leptin is not surprising given the pattern ofexpression of its receptors in a multitude of tissues, includingbut not limited to the brain, lung, kidney, spleen, heart, andliver.2,3
Previously characterized leptin receptors, which are allproducts of alternative splicing of thedb gene transcript,6
have differential patterns of expression in different tissues.2
They share identical extracellular and membrane-spanningdomains, but the intracellular domain varies in length as aresult of the alternative splicing.3 The long form of thereceptor (OB-Rb) seems to be responsible for generating theintracellular signals controlling hypothalamic responses toleptin. However, little is known about its role in regulatingextrahypothalamic signal transduction.
Circulating leptin concentrations vary with fat mass,7 sexhormone levels,8 exposure to bacterial lipopolysaccharide,5
increased dietary fat,9,10 and age.8,11,12Collectively, the widedistribution of leptin receptor expression and the multipleregulatory mechanisms and factors involved in the control ofleptin levels suggest that leptin is involved in other physio-logical or pathological conditions. Sierra-Honigmann et al4
and Bouloumie et al13 showed that leptin has angiogenicactivity in endothelial cells. Silver et al14 recently showed thatleptin regulates the hepatic clearance of high density lipopro-teins in mice, hence possibly affecting lipid-associated dis-eases such as atherosclerosis.
Recently, we and others showed that vascular calcificationis an actively regulated process involving a subpopulation ofartery wall cells, called calcifying vascular cells (CVC), thatundergo osteoblastic differentiation and form hydroxyapatitemineral.15 Both leptin levels and vascular calcification in-crease with age8,16 and with a high-fat diet.9,17 Because leptinalso modulates the differentiation of marrow osteoprogenitorcells,18 we hypothesized that leptin may modulate the differ-entiation of vascular osteoprogenitor cells. The results in thepresent report indicate, for the first time, that leptin is a potent
Original received January 14, 2000; resubmission received November 2, 2000; revised resubmission received March 21, 2001; accepted April 4, 2001.From the Departments of Medicine (F.P., Y.T., A.B., A.M.F., L.L.D.) and Physiology (L.L.D.), University of California, Los Angeles.Correspondence to Farhad Parhami, PhD, Division of Cardiology, UCLA School of Medicine, Center for the Health Sciences, Room 47-123, 10833
Le Conte Avenue, Los Angeles, CA 90095. E-mail [email protected]© 2001 American Heart Association, Inc.
Circulation Researchis available at http://www.circresaha.org
954
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from

inducer of osteoblastic differentiation and calcification ofCVC in vitro and that the leptin receptor is expressed by CVCand by subpopulations of cells in the mouse artery wall. Inaddition, the in vivo expression of leptin in the artery wall andby human aortic endothelial cells and monocyte/macrophagesin vitro are shown. The presence of leptin and its receptor inthe artery wall suggests that physiological or pathologicalfunctions of artery wall cells may be targets of leptin action.
Materials and MethodsCell CultureCVC clones were isolated from cultures of bovine smooth musclecells harvested from bovine aortic medial explants and cultured aspreviously described.15 Human aortic endothelial cells were isolatedand cultured as previously described.19 Human peripheral bloodmonocytes were isolated as previously described,20 plated in 30%autologous human serum in Iscove’s modified Dulbecco’s medium(IMDM; Irvine Scientific) supplemented with 100 U/mL penicillin,100 mg/mL streptomycin, 0.25mg/mL Fungizone (amphotericin b),2 mmol/L L-glutamine, and 0.22 U/mL insulin, and cultured for 7days before RNA extraction. Human adipocytes were kindly pro-vided by Dr Daniel De Ugarte (Department of Surgery, University ofCalifornia at Los Angeles). Murine recombinant leptin was obtainedfrom BioMol Research Laboratories and was 95% pure by SDS-PAGE, according to the manufacturer.
Alkaline Phosphatase Activity AssayA cell-associated alkaline phosphatase activity assay was performedas previously described.21
Von Kossa Staining for CalcificationCell monolayers were fixed in 0.1% glutaraldehyde for 15 minutes atroom temperature, followed by staining with silver nitrate, aspreviously described.21
45Ca Incorporation45Ca incorporation into matrix of CVC cultures was performed aspreviously described.21
Reverse Transcriptase Polymerase Chain ReactionTotal RNA from tissue or cells grown in duplicate 60-mm tissueculture dishes was isolated using an RNA isolation kit (Strategene).A total of 3 mg of the total RNA was reverse-transcribed in a 50-mLreaction volume, as described previously.22 Primers were constructedfrom bovine leptin receptor (accession number U62385) and mouseleptin receptor (accession number MMU58861), homologous to thelong form of human leptin receptor OB-Rb, and from mouse leptin(accession number U2421.1). Sequences of the primers, synthesizedby Gibco-BRL were as follows: bovine leptin receptor: sense (1-25)59-GTGCCAGCAACTACAGATGCTCTAC-39, antisense (356-380) 59-AGTTCATCCAGGCCTTCTGAGAACG-39; mouse leptin re-ceptor: sense (2872-2893) 59-TGAGATGGTCCCAGCAGC-TATG-39,antisense (3260-3239) 59-CCAAAAGCTCATC-CAACCCCGA-39;and mouse leptin: sense (7-26) 59-TGGAGACCCCTGTGTCGG-TT-39, antisense (479-502) 59-AGCATTCAGGGCTAACAT-CCAACT-39. Polymerase chain reaction (PCR) was performed aspreviously described,22 except the annealing temperature was 65°C andthe total number of cycles was 45 for bovine leptin receptor, 36 formouse leptin receptor, and 41 for mouse leptin. PCR products (expectedlength of 380 base pairs for bovine leptin receptor, 389 base pairs formouse leptin receptor, and 496 base pairs for mouse leptin) wereelectrophoresed on an agarose gel and visualized using ethidiumbromide stain. All PCR products were sequenced by the University ofCalifornia at Los Angeles (UCLA) sequencing core laboratory, and thesequencing results were electronically compared with GenBank entries
using BLAST NIH software. Bovine leptin receptor PCR productshowed the highest similarity to the submitted entry U62385 (Bos taurusobese receptor gene); the mouse leptin receptor PCR product showedthe highest similarity to the submitted entry AF098792 (Mus musculusleptin receptor long isoform Rb gene); and the mouse leptin PCRproduct showed the highest similarity to the submitted entryNM_008493 (Mus musculus leptin mRNA).
ImmunocytochemistryConfluent CVC monolayers were grown in chamber slides and fixedwith 4% paraformaldehyde and stained with the polyclonal rabbitanti-mouse leptin receptor antibody OBR-E1 (Torrey Pines Biolabs)at a concentration of 4mg/mL. This antibody was generated usingEscherichia coli–expressed recombinant OB receptor, amino acids25 to 208, as an immunogen. It was found to react with bothrecombinant and natural mouse OB receptor (per Torrey PinesBiolabs). As a negative control, nonimmunized rabbit IgG atconcentrations equal to the anti-leptin receptor antibody was used.Biotinylated secondary antibody was used for visualization, andcounterstaining with hematoxylin was performed.
Western Blot AnalysisCVC grown in duplicate 100-mm plates were scraped and centri-fuged, and the cell pellet was resuspended in 13 PBS containingprotease inhibitors (0.5mg/mL leupeptin, 0.7mg/mL pepstatin, and0.2 mmol/L phenylmethylsulfonyl fluoride). A partially purifiedmembrane fraction was prepared by freeze-thawing the cells 3 timesin a dry ice–ethanol bath. On visualization under light microscope,all cells were ruptured. The membrane fraction was then collectedafter centrifugation for 10 minutes at 14 000g, resuspended in lysisbuffer containing protease inhibitors under reducing conditions, andelectrophoresed on 8% Tris-glycine gels (Novex). This was followedby transfer to nitrocellulose membranes, as previously described.22
The blots were probed with 2mg/mL of either the polyclonal rabbitanti-mouse leptin receptor antibody OBR-E1 (Torrey Pines Biolabs)or with nonimmunized rabbit IgG. Detection of immunoreactiveproteins was done by enhanced chemiluminescence (Amersham).
Northern Blot AnalysisA total of 10 mg of total RNA extracted from tissues or cells waselectrophoresed through a 1% agarose gel and transblotted to a nylonmembrane, as previously described.19 The blot was hybridized witha 32P-labeled human leptin probe generated by PCR using the genesequence for human leptin (accession No. U43653). The sequencesof the primers synthesized by Gibco-BRL were as follows: sense(67-87) 59-GAACCCTGTGCGGATTCTTGT-39; antisense (344-367) 59- AGGTTCTCCAGGTCGTTGGATATT 39. A PCR productof 301 base pairs was gel-purified and used for Northern analysis.The PCR product was sequenced by the UCLA sequencing corelaboratory, and the sequencing results were electronically comparedwith GenBank entries using BLAST NIH software. They showed thehighest similarity to submitted entry XM_004625 (Homo sapiensleptin mRNA).
Leptin RIALeptin RIA was performed on conditioned media from culturedhuman aortic endothelial cells and human monocyte/macrophages.Cells were cultured in 60-mm tissue culture plates as describedabove for 6 days. The media was then replaced with 3 ml of IMDMfor monocyte/macrophages and M199 for endothelial cells, bothcontaining 10% FBS. After 48 hours, conditioned media wascollected, centrifuged to remove detached cells, and stored at270°C. The cell layer was dissolved in 0.4 mol/L NaOH, and theprotein concentration was measured using the Bradford protein assay(Bio-Rad). A total of 100mL of conditioned media from each celltype was used in the human leptin RIA assay from Linco Researchaccording to the manufacturer’s instructions. According the manu-facturer’s information, this RIA assay has 100% specificity for
Parhami et al Role of Leptin in Calcification of Vascular Cells 955
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from

human leptin and,0.2% specificity for leptin from other species.The leptin concentrations obtained were normalized to cellularprotein content.
Tissue Preparation and ImmunohistochemistryNormal aortas were obtained from 6-month-old, female, C57BL/6Jmice. Atherosclerotic aortas were obtained from 6-month-old, fe-male, LDL receptor–null mice fed an atherogenic diet for 4 monthsor from apoE-null mice, the F2 progeny of a C57BL/6J X DBA/2Jintercross (generous gift of Dr Thomas A. Drake, Department ofPathology, UCLA), fed a high-fat atherogenic diet for 4 months. Theaortas were surgically removed, embedded in OCT compound(Fisher Scientific), and frozen in liquid nitrogen. Unfixed 8-mmserial sections were incubated in Tris saline solution containing8 mmol/L Tris-base, 40 mmol/L Tris-HCl, 150 mmol/L NaCl, 5%wt/vol nonfat dry milk, and 1% v/v normal goat serum for 2 hours atroom temperature to block nonspecific binding and then stained forleptin receptor using the OBR-E1 polyclonal rabbit anti-mouseantibody (Torrey Pines Biolabs) at 4mg/mL, for von Willebrandfactor using a polyclonal rabbit anti-human antibody (DAKO Corp)at 1mg/mL, or for leptin using an affinity-purified rabbit anti-mousepolyclonal antibody (OB A-20, Santa Cruz Biotechnology) at 1mg/mL. To control for possible nonspecific staining, serial sectionswere stained with leptin antibody neutralized with blocking peptide(Santa Cruz Biotechnology) according to the manufacturer’s instruc-tions. Biotinylated goat anti-rabbit secondary antibody was used todetect the primary antibodies and visualized using the VectastainABC kit (Vector Labs) followed by Peroxidase Chromogen AEC kit(Biomeda Corp), according to the manufacturer’s instructions. Tis-sue sections were counterstained with hematoxylin.
Statistical AnalysisStatistical analyses were performed using ANOVA.
ResultsLeptin Induces Calcification in Cultures of CVCWe previously found that CVC undergo a differentiationprocess similar to that seen in preosteoblasts, includingsequential upregulation of several osteoblast-specific mark-ers. Alkaline phosphatase, an early marker of osteoblasticdifferentiation in CVC and other osteoblastic cells, mediatesthe differentiation process and provides phosphate ions forcalcium phosphate mineral formation.23,24Treatment of CVCwith leptin for 4 days caused a significant, dose-dependent, 5-to 10-fold induction of alkaline phosphatase activity (Figure1A). After 10 days of leptin treatment, calcification in theCVC cultures was also dramatically increased, as shown byvon Kossa staining (Figure 1B) and a45Ca incorporationassay (Figure 1C). Altogether, these results suggest that leptinstrongly induces osteoblastic differentiation and mineraliza-tion of CVC in culture. Leptin did not significantly alterproliferation or osteopontin expression in CVC (data notshown). In addition, the above CVC responses to leptin wereindependent of signal transducer and activator of transcrip-tion 3 (STAT3) activation, because treatment with leptin for15 minutes or 1 hour did not induce phosphorylation ofSTAT3 in CVC, as determined by Western blot analysisusing an antibody to phosphorylated STAT3 (Santa CruzBiotechnology; data not shown).
Leptin Receptor Is Expressed in CVCTo examine the expression of leptin receptor in CVC, weperformed reverse transcriptase PCR (RT-PCR) using RNAisolated from confluent cultures of CVC with primers specific
for OB-Rb. The presence of a PCR product of the expectedsize of 380 base pairs indicated that OB-Rb was expressed inCVC (Figure 2A). Immunocytochemistry performed on non-permeabilized confluent cultures of CVC with an antibody to
Figure 1. Effect of leptin on alkaline phosphatase activity andcalcification in calcifying vascular cell cultures. A, CVC wereincubated for 4 days with control buffer (0) or the indicated con-centrations of leptin in DMEM containing 5% FBS. Alkalinephosphatase activity was measured in total cell homogenates,and the results from a representative of 5 experiments areshown as the mean6SD of quadruplicate determinations.P,0.005 for all leptin vs buffer-treated samples. B, CVC mono-layers were treated as described in A for 10 days. Von Kossasilver nitrate staining shows calcification as a black stain.Results from a representative of 3 experiments are shown(phase-contrast magnification, 340). The number in the bottomcorner of each panel represents the concentration of leptin usedin mg/mL. C, CVC were treated with control buffer or leptin for10 days, as described above, and then a 45Ca incorporationassay was performed. Results from a representative of 3 experi-ments are shown as the mean6SD of 6 determinations. P,0.01for all leptin vs buffer-treated samples.
956 Circulation Research May 11, 2001
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from
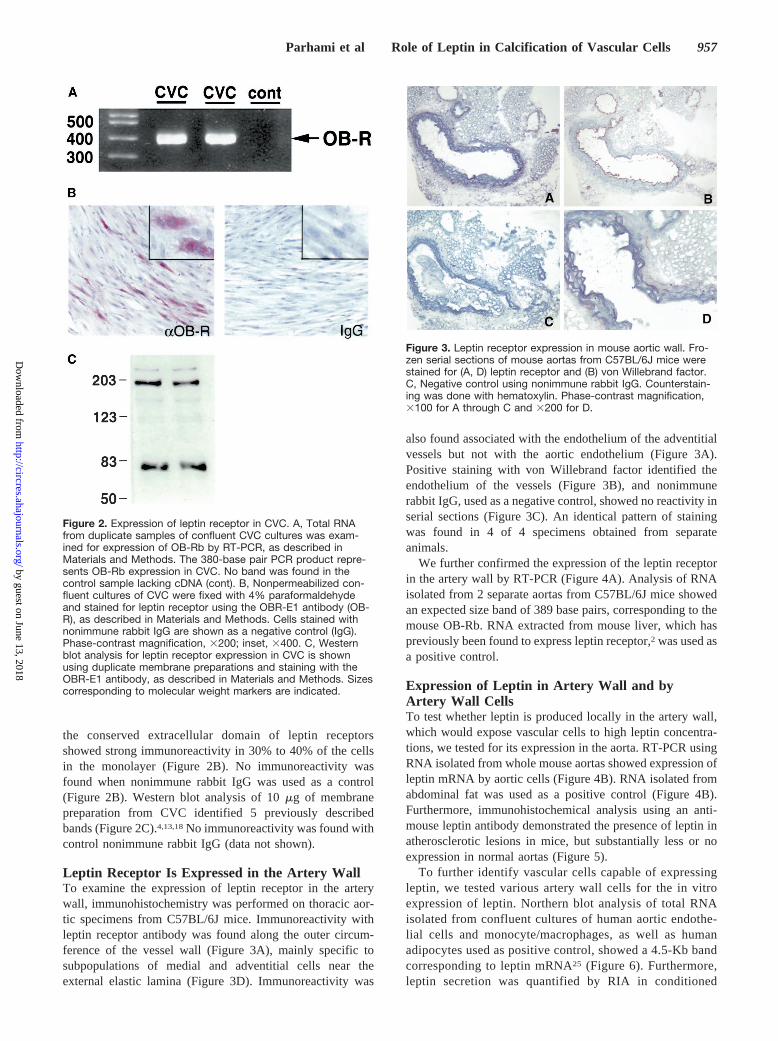
the conserved extracellular domain of leptin receptorsshowed strong immunoreactivity in 30% to 40% of the cellsin the monolayer (Figure 2B). No immunoreactivity wasfound when nonimmune rabbit IgG was used as a control(Figure 2B). Western blot analysis of 10mg of membranepreparation from CVC identified 5 previously describedbands (Figure 2C).4,13,18No immunoreactivity was found withcontrol nonimmune rabbit IgG (data not shown).
Leptin Receptor Is Expressed in the Artery WallTo examine the expression of leptin receptor in the arterywall, immunohistochemistry was performed on thoracic aor-tic specimens from C57BL/6J mice. Immunoreactivity withleptin receptor antibody was found along the outer circum-ference of the vessel wall (Figure 3A), mainly specific tosubpopulations of medial and adventitial cells near theexternal elastic lamina (Figure 3D). Immunoreactivity was
also found associated with the endothelium of the adventitialvessels but not with the aortic endothelium (Figure 3A).Positive staining with von Willebrand factor identified theendothelium of the vessels (Figure 3B), and nonimmunerabbit IgG, used as a negative control, showed no reactivity inserial sections (Figure 3C). An identical pattern of stainingwas found in 4 of 4 specimens obtained from separateanimals.
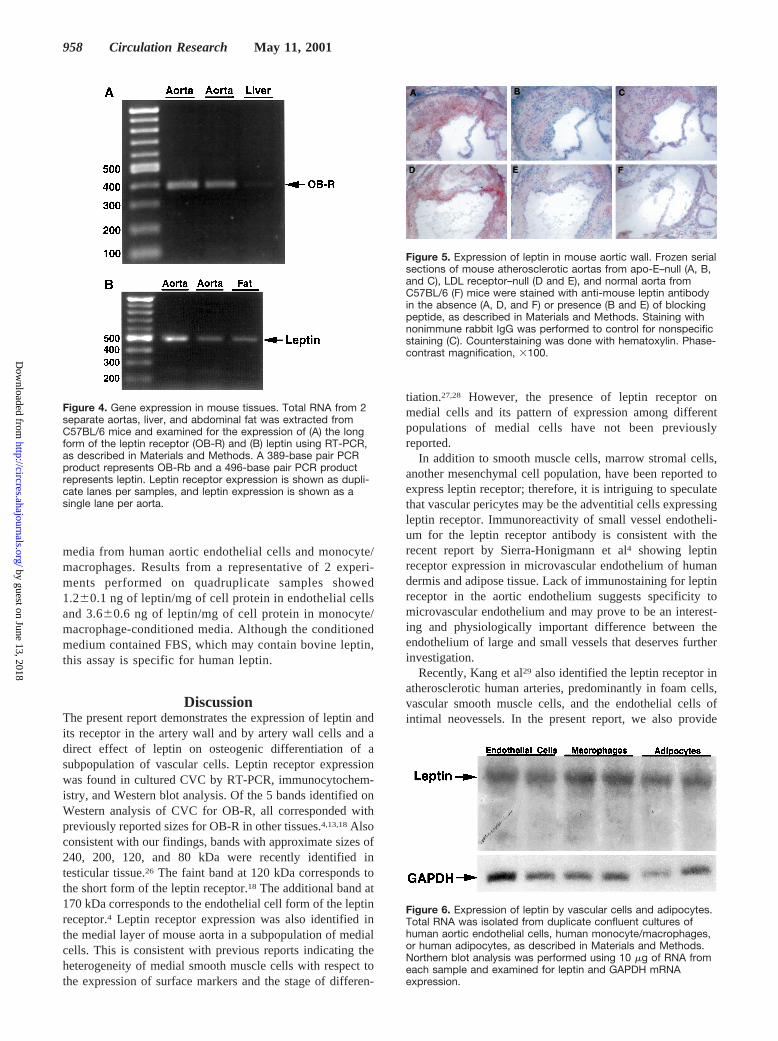
We further confirmed the expression of the leptin receptorin the artery wall by RT-PCR (Figure 4A). Analysis of RNAisolated from 2 separate aortas from C57BL/6J mice showedan expected size band of 389 base pairs, corresponding to themouse OB-Rb. RNA extracted from mouse liver, which haspreviously been found to express leptin receptor,2 was used asa positive control.
Expression of Leptin in Artery Wall and byArtery Wall CellsTo test whether leptin is produced locally in the artery wall,which would expose vascular cells to high leptin concentra-tions, we tested for its expression in the aorta. RT-PCR usingRNA isolated from whole mouse aortas showed expression ofleptin mRNA by aortic cells (Figure 4B). RNA isolated fromabdominal fat was used as a positive control (Figure 4B).Furthermore, immunohistochemical analysis using an anti-mouse leptin antibody demonstrated the presence of leptin inatherosclerotic lesions in mice, but substantially less or noexpression in normal aortas (Figure 5).
To further identify vascular cells capable of expressingleptin, we tested various artery wall cells for the in vitroexpression of leptin. Northern blot analysis of total RNAisolated from confluent cultures of human aortic endothe-lial cells and monocyte/macrophages, as well as humanadipocytes used as positive control, showed a 4.5-Kb bandcorresponding to leptin mRNA25 (Figure 6). Furthermore,leptin secretion was quantified by RIA in conditioned
Figure 2. Expression of leptin receptor in CVC. A, Total RNAfrom duplicate samples of confluent CVC cultures was exam-ined for expression of OB-Rb by RT-PCR, as described inMaterials and Methods. The 380-base pair PCR product repre-sents OB-Rb expression in CVC. No band was found in thecontrol sample lacking cDNA (cont). B, Nonpermeabilized con-fluent cultures of CVC were fixed with 4% paraformaldehydeand stained for leptin receptor using the OBR-E1 antibody (OB-R), as described in Materials and Methods. Cells stained withnonimmune rabbit IgG are shown as a negative control (IgG).Phase-contrast magnification, 3200; inset, 3400. C, Westernblot analysis for leptin receptor expression in CVC is shownusing duplicate membrane preparations and staining with theOBR-E1 antibody, as described in Materials and Methods. Sizescorresponding to molecular weight markers are indicated.
Figure 3. Leptin receptor expression in mouse aortic wall. Fro-zen serial sections of mouse aortas from C57BL/6J mice werestained for (A, D) leptin receptor and (B) von Willebrand factor.C, Negative control using nonimmune rabbit IgG. Counterstain-ing was done with hematoxylin. Phase-contrast magnification,3100 for A through C and 3200 for D.
Parhami et al Role of Leptin in Calcification of Vascular Cells 957
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from
media from human aortic endothelial cells and monocyte/macrophages. Results from a representative of 2 experi-ments performed on quadruplicate samples showed1.260.1 ng of leptin/mg of cell protein in endothelial cellsand 3.660.6 ng of leptin/mg of cell protein in monocyte/macrophage-conditioned media. Although the conditionedmedium contained FBS, which may contain bovine leptin,this assay is specific for human leptin.
DiscussionThe present report demonstrates the expression of leptin andits receptor in the artery wall and by artery wall cells and adirect effect of leptin on osteogenic differentiation of asubpopulation of vascular cells. Leptin receptor expressionwas found in cultured CVC by RT-PCR, immunocytochem-istry, and Western blot analysis. Of the 5 bands identified onWestern analysis of CVC for OB-R, all corresponded withpreviously reported sizes for OB-R in other tissues.4,13,18Alsoconsistent with our findings, bands with approximate sizes of240, 200, 120, and 80 kDa were recently identified intesticular tissue.26 The faint band at 120 kDa corresponds tothe short form of the leptin receptor.18 The additional band at170 kDa corresponds to the endothelial cell form of the leptinreceptor.4 Leptin receptor expression was also identified inthe medial layer of mouse aorta in a subpopulation of medialcells. This is consistent with previous reports indicating theheterogeneity of medial smooth muscle cells with respect tothe expression of surface markers and the stage of differen-
tiation.27,28 However, the presence of leptin receptor onmedial cells and its pattern of expression among differentpopulations of medial cells have not been previouslyreported.
In addition to smooth muscle cells, marrow stromal cells,another mesenchymal cell population, have been reported toexpress leptin receptor; therefore, it is intriguing to speculatethat vascular pericytes may be the adventitial cells expressingleptin receptor. Immunoreactivity of small vessel endotheli-um for the leptin receptor antibody is consistent with therecent report by Sierra-Honigmann et al4 showing leptinreceptor expression in microvascular endothelium of humandermis and adipose tissue. Lack of immunostaining for leptinreceptor in the aortic endothelium suggests specificity tomicrovascular endothelium and may prove to be an interest-ing and physiologically important difference between theendothelium of large and small vessels that deserves furtherinvestigation.
Recently, Kang et al29 also identified the leptin receptor inatherosclerotic human arteries, predominantly in foam cells,vascular smooth muscle cells, and the endothelial cells ofintimal neovessels. In the present report, we also provide
Figure 5. Expression of leptin in mouse aortic wall. Frozen serialsections of mouse atherosclerotic aortas from apo-E–null (A, B,and C), LDL receptor–null (D and E), and normal aorta fromC57BL/6 (F) mice were stained with anti-mouse leptin antibodyin the absence (A, D, and F) or presence (B and E) of blockingpeptide, as described in Materials and Methods. Staining withnonimmune rabbit IgG was performed to control for nonspecificstaining (C). Counterstaining was done with hematoxylin. Phase-contrast magnification, 3100.
Figure 6. Expression of leptin by vascular cells and adipocytes.Total RNA was isolated from duplicate confluent cultures ofhuman aortic endothelial cells, human monocyte/macrophages,or human adipocytes, as described in Materials and Methods.Northern blot analysis was performed using 10 mg of RNA fromeach sample and examined for leptin and GAPDH mRNAexpression.
Figure 4. Gene expression in mouse tissues. Total RNA from 2separate aortas, liver, and abdominal fat was extracted fromC57BL/6 mice and examined for the expression of (A) the longform of the leptin receptor (OB-R) and (B) leptin using RT-PCR,as described in Materials and Methods. A 389-base pair PCRproduct represents OB-Rb and a 496-base pair PCR productrepresents leptin. Leptin receptor expression is shown as dupli-cate lanes per samples, and leptin expression is shown as asingle lane per aorta.
958 Circulation Research May 11, 2001
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from

evidence that leptin is produced in vitro by cells of the arterywall and in atherosclerotic lesions. Although leptin proteinwas not evident on immunohistochemistry of the normalmouse aorta, it is possible that the endothelial cells secretedthe protein into the circulation. In contrast, in atheroscleroticlesions, leptin protein may be trapped in the abnormal matrix,and it may also be produced by monocytes and macrophageswithin the lesion. Altogether, it is clear that the leptin receptoris present on artery wall cells; therefore, the role of leptin inregulating the function of these cells and the artery walldeserves further elucidation. Interestingly, Nishina et al30
reported that mice lacking leptin (ob/ob) or leptin receptor(db/db) had reduced atherosclerotic lesion areas comparedwith controls, which was partially contributed to by theincreased HDL levels in these mice. However, because leptinmay regulate inflammation and immunity,31 it is intriguing tospeculate that the reduced lesion area reported by Nishina etal30 may by partially due to the absence of leptin and itsproinflammatory effects in the mutant mice.
The present results also indicate that leptin regulates theosteogenic differentiation of a subpopulation of vascularcells. Leptin induced alkaline phosphatase activity and sub-sequent mineralization in CVC. The pro-osteogenic differen-tiation effects of leptin are consistent with findings ofThomas et al,18 who recently reported that leptin induces theosteoblastic differentiation of marrow stromal cells, whichalso express leptin receptors.
We also examined the effect of leptin on 2 major signalingmolecules, STAT3 and cAMP. Leptin induces the phosphor-ylation and activation of STAT3 in several cell types,including endothelial cells and oocytes.4,32 However, leptindid not induce the activation of STAT3 in CVC, as indicatedby a lack of phosphorylation of STAT3 in leptin-treated cells.We previously reported that the induction of cAMP levels inCVC enhances their osteogenic differentiaton.22 TreatingCVC with leptin did not have a significant effect on cAMPlevels (data not shown). These results suggest that thepro-osteogenic effects of leptin on CVC are independent ofSTAT3 or cAMP.
The present report further supports the role of leptin as aregulator of peripheral tissue physiology by providing ana-tomical evidence of leptin receptor expression in the aorticwall and biological activity promoting the calcification ofcells derived from vascular tissue. This is a significant findinggiven the importance of vascular calcification in the diseasesof the artery wall, including atherosclerosis, and the aging-associated impairment of its elastic functions.33,34
AcknowledgmentsThis work was supported in part by NIH grant HL30568 and theLaubisch Fund. F. Parhami is a recipient of a Career DevelopmentAward from the Claude D. Pepper Older American IndependenceCenter at UCLA. The authors would like to thank Dr Thomas A.Drake for insightful suggestions, Vien Le and Tony Mottino forexpert technical assistance, and the UCLA Biomedical TechnologyResearch and Instructional Production Facility for assistance withgraphics.
References1. Tartaglia LA. The leptin receptor.J Biol Chem. 1997;272:6093–6096.2. Fei H, Okano HJ, Li C, Lee GH, Zhao C, Darnell R, Friedman JM.
Anatomic localization of alternatively spliced leptin receptors (Ob-R) inmouse brain and other tissues.Proc Natl Acad Sci U S A. 1997;94:7001–7005.
3. Barr VA, Lane K, Taylor SI. Subcellular localization and internalizationof the four human leptin receptor isoforms.J Biol Chem. 1999;274:21416–21424.
4. Sierra-Honigmann MR, Nath AK, Murakami C, Garcia-Cardena G,Papapetropoulos A, Sessa WC, Madge LA, Schechner JS, Schwabb MB,Polverini PJ, Flores-Riveros JR. Biological action of leptin as anangiogenic factor.Science. 1998;281:1683–1686.
5. Francis J, Mohankumar PS, Mohankumar SMJ, Quadri SK. Systemicadministration of lipopolysaccharide increases plasma leptin levels.Endocrine. 1999;10:291–295.
6. Lee GH, Proenca R, Montez JM, Carroll KM, Darvishzadeh JG, Lee JI,Friedman JM. Abnormal splicing of the leptin receptor in diabetic mice.Nature. 1996;379:632–635.
7. Moller N, O’Brien P, Nair KS. Disruption of the relationship between fatcontent and leptin levels with aging in humans.J Clin Endocrinol Metab.1998;83:931–934.
8. Baumgartner RN, Waters DL, Morley JE, Patrick P, Montoya GD, GarryPJ. Age-related changes in sex hormones affect the sex differences inserum leptin independently of changes in body fat.Metabolism. 1999;48:378–384.
9. Ahren B. Plasma leptin and insulin in C57BL/6J mice on a high-fat diet:relation to subsequent changes in body weight.Acta Physiol Scand.1999;165:233–240.
10. Ahren B, Mansson S, Gingerich RL, Havel PJ. Regulation of plasmaleptin in mice: influence of age, high-fat diet, and fasting.Am J Physiol.1997;273:R113–R120.
11. Li H, Matheny M, Tumer N, Scarpace PJ. Aging and fasting regulation ofleptin and hypothalamic neuropeptide Y gene expression.Am J Physiol.1998;275:E405–E411.
12. Li H, Matheny M, Nicolson M, Tumer N, Scarpace PJ. Leptin geneexpression increases with age independent of increasing adiposity in rats.Diabetes. 1997;46:2035–2039.
13. Bouloumie A, Drexler CAH, Lafontan M, Busse R. Leptin, the product ofOb gene, promotes angiogenesis.Circ Res. 1998;83:1059–1066.
14. Silver DL, Jiang X, Tall A. Increased high density lipoprotein (HDL),defective hepatic catabolism of Apo A-I and ApoA-II, and decreasedApoA-I mRNA in ob/obmice.J Biol Chem. 1999;274:4140–4146.
15. Watson KE, Bostrom K, Ravindranath R, Lam T, Norton B, Demer LL.TGF-b1 and 25-hydroxycholesterol stimulate osteoblast-like vascularcells to calcify.J Clin Invest. 1994;93:2106–2113.
16. Agatston AS, Janowitz WR, Hildner FJ, Zusmer NR, Viamonte M,Detrano R. Quantification of coronary artery calcium using ultrafastcomputed tomography.J Am Coll Cardiol. 1990;15:827–832.
17. Towler DA, Bidder M, Latifi T, Coleman T, Semenkovich CF. Diet-induced diabetes activates an osteogenic gene regulatory program in theaortas of low density lipoprotein receptor-deficient mice.J Biol Chem.1998;273:30427–30434.
18. Thomas T, Gori F, Khosla S, Jensen MD, Burguera B, Riggs BL. Leptinacts on human marrow stromal cells to enhance differentiation to osteo-blasts and to inhibit differentiation to adipocytes.Endocrinology. 1999;140:1630–1638.
19. Parhami F, Fang ZT, Fogelman AM, Andalibi A, Territo MC, BerlinerJA. Minimally modified low density lipoprotein-induced inflammatoryresponses in endothelial cells are mediated by cyclic adenosine mono-phosphate.J Clin Invest. 1993;92:471–478.
20. Colotta F, Peri G, Villa A, Mantovani A. A rapid killing of actinomycinD-treated tumor cells by human mononuclear cells.J Immunol. 1984;132:936–944.
21. Parhami F, Morrow AD, Balucan J, Leitinger N, Watson AD, Tintut Y,Berliner JA, Demer LL. Lipid oxidation products have opposite effects oncalcifying vascular cell and bone cell differentiation.Arterioscler ThrombVasc Biol. 1997;17:680–687.
22. Tintut Y, Parhami F, Bostrom K, Jackson SM, Demer LL. cAMP stim-ulates osteoblast-like differentiation of calcifying vascular cells.J BiolChem. 1998;273:7547–7553.
23. Anderson HC. Molecular biology of matrix vesicles.Clin Orthop. 1995;314:266–280.
Parhami et al Role of Leptin in Calcification of Vascular Cells 959
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from

24. Narisawa S, Hofmann MC, Ziomek CA, Millan JL. Embryonic alkalinephosphatase is expressed at M-phase in the spermatogenic lineage of themouse.Development. 1992;116:159–165.
25. Bradley RL, Kokkotou EG, Maratos-Flier E, Cheatham B. Melanin-concentrating hormone regulates leptin synthesis and secretion in ratadipocytes.Diabetes. 2000;49:1073–1077.
26. El-Hefnawy T, Ioffe S, Dym M. Expression of the leptin receptor duringgerm cell development in the mouse testis.Endocrinology. 2000;141:2624–2630.
27. Gittenberger-de Groot AC, DeRuiter MC, Bergwerff M, PoelmannRE. Smooth muscle cell origin and its relation to heterogeneity indevelopment and disease.Arterioscler Thromb Vasc Biol. 1999;19:1589 –1594.
28. Shanahan CM, Weissberg PL. Smooth muscle cell heterogeneity: patternsof gene expression in vascular smooth muscle cells in vitro and in vivo.Arterioscler Thromb Vasc Biol. 1998;18:333–338.
29. Kang SM, Kwon HM, Hong BK, Kim D, Kim IJ, Choi EY, Jang Y, KimHS, Kim MS, Kwon HC. Expression of leptin receptor (Ob-R) in human
atherosclerotic lesions: potential role in intimal neovascularization.Yonsei Med J. 2000;41:68–75.
30. Nishina PM, Naggert JK, Verstuyft J, Paigen B. Atherosclerosis in genet-ically obese mice: the mutation obese, diabetes, fat, tubby, and lethalyellow. Metabolism. 1994;43:554–558.
31. Fantuzzi G, Faggioni R. Leptin in the regulation of immunity, inflam-mation, and hematopoiesis.J Leukocyte Biol. 2000;68:437–446.
32. Matsuoka T, Tahara M, Yokoi T, Masumoto N, Takeda T, Yamaguchi M,Tasaka K, Kurachi H, Murata Y. Tyrosine phosphorylation of STAT3 byleptin through leptin receptor in mouse metaphase 2 stage oocyte.Biochem Biophys Res Commun. 1999;256:480–484.
33. Stanford W, Thompson BH, Weiss RM. Coronary artery calcification:clinical significance and current methods of detection.Am J Radiol.1993;161:1139–1146.
34. Parhami F, Demer LL. New concepts in regulation of vascular calcifi-cation. In: Fuster V, ed.The Vulnerable Plaque: Understanding, Identi-fication, and Modification.Armonk NY: Futura Publishing Co; 1999:383–391.
960 Circulation Research May 11, 2001
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from

Farhad Parhami, Yin Tintut, Alex Ballard, Alan M. Fogelman and Linda L. DemerLeptin Enhances the Calcification of Vascular Cells: Artery Wall as a Target of Leptin
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2001 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/hh0901.0909752001;88:954-960; originally published online April 27, 2001;Circ Res.
http://circres.ahajournals.org/content/88/9/954World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 13, 2018http://circres.ahajournals.org/
Dow
nloaded from





























