Embed Size (px)
Citation preview

APPLIED ISSUES
Flow-related patterns in abundance and compositionof the fish fauna of a degraded Australian lowland river
PAUL HUMPHRIES* , 1 , PAUL BROWN †, JOHN DOUGLAS †, ANDREW PICKWORTH†, 2 ,
RUSSEL STRONGMAN †, KYLIE HALL † AND LUCIANO SERAFINI*
*Department of Biological Sciences, Cooperative Research Centre for Freshwater Ecology, c/- Murray-Darling Freshwater Research
Centre, Monash University, Albury, NSW, Australia†Department of Primary Industries, Primary Industries Research Victoria, Alexandra, Vic., Australia
SUMMARY
1. A 7-year study was conducted in three hydrologically distinct sections within the highly
regulated, lowland Campaspe River to investigate the influence of hydrology on temporal
and spatial patterns in fish composition, abundance and recruitment. One section had
6 months, one section 2 months and one section no months of increased flow due to
storage releases. The fish fauna of the less regulated, nearby Broken River served as a
reference to which that of the Campaspe River was compared for the last 3 years of the
study to allow insight into the relative effects of hydrology, barriers to movement and
other environmental characteristics. The study included one high-flow year, a moderate-
flow year and five low-flow years.
2. A total of 16 fish species – 10 native and six alien – were caught in the Campaspe River,
although of the native species, only three are considered to have self-sustaining
populations. The remaining species are either itinerants or a result of stocking. Alien
species comprised approximately 64% of the total biomass of all fish caught.
3. Overall composition of the fish fauna did not differ significantly by year, but did by
section of river. Species richness and the abundance of most of the dominant species also
differed significantly by river section, but there was little inter-annual variation in the
abundance of any species, except for European perch and for common carp; the latter
showing an increase in abundance following a high-flow event during the spring of 2000 as
a result of recruitment.
4. Overall faunal composition was not influenced by hydrology. However, multiple
regression indicated that species richness, abundance of the dominant species and
abundance of young-of-year (YOY) of golden perch, European perch and common carp all
were influenced significantly by hydrological variables. The nature of the relationships
was dependent on river section and hydrological season (‘winter’ or ‘spring/summer’). Of
note was the result that the total abundance of fish and that of YOY common carp were
significantly positively related to the number of spells above the threshold for movement
upstream through the lower two weirs in the Campaspe River. Only one significant
Correspondence: Paul Humphries, School of Environmental Sciences, Institute for Land Water and Society, Charles Sturt University,
PO Box 789, Albury, NSW 2640, Australia.
E-mail: [email protected] address: School of Environmental Sciences, Institute for Land Water and Society, Charles Sturt University, PO Box 789,
Albury, NSW 2640, Australia.2Present address: Department of Sustainability and Environment, Arthur Rylah Institute for Environmental Research, PO Box 137,
Heidelberg, Vic. 3084, Australia.
Freshwater Biology (2008) 53, 789–813 doi:10.1111/j.1365-2427.2007.01904.x
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd 789

relationship between hydrological and fish-related variables was found for the upper river
section, whereas seven and five were found for the lower and middle sections respectively.
5. Comparisons with fish collected in the Broken River over 3 years suggest that the fauna
of the Broken River is in a more natural state than that of the Campaspe River. Since the
two rivers do not differ substantially in water quality, and since both contain significant
weirs, which act as barriers to movement of fish, flow regulation is most likely to be the
major reason for the poor state of the fauna in the Campaspe River.
Keywords: alien, Campaspe and Broken Rivers, flow regulation, hydrology, Murray-Darling Basin,recruitment
Introduction
Flow is the overriding force in rivers. Flow defines
river geomorphology and sediment transport, deter-
mines the type, amount and accessibility of habitat for
riverine animals and plants, it drives food webs
through transportation of carbon and it has a major
influence on animal and plant behaviour and life
histories (see Vannote et al., 1980; Frissell et al., 1986;
Junk, Bayley & Sparks, 1989; Calow & Petts, 1992;
Thorp & Delong, 1994; Matthews, 1998; Fausch et al.,
2002). The flow regime of a river can be described by
five major components: magnitude, duration, timing,
frequency and rate of change. These components
operate at a number of temporal and spatial scales,
and riverine biota have evolved in their context, and
must either take advantage of the predictable aspects
of the components, such as annual, widespread
flooding of forests in tropical South America, or deal
as best they can with the less predictable aspects, e.g.
summer spates or supra-seasonal droughts (Poff et al.,
1997; Lake, 2003; Richter et al., 2003). High flows, and
especially spates, seem to play a major role in
influencing the variability in fish assemblages, at least
in the short term (Oberdorff, Hugueny & Vigneron,
2001; Schiemer et al., 2001; Koel & Sparks, 2002; Li &
Gelwick, 2005).
River regulation is widespread throughout the
world and it typically alters one or more of the
components of a flow regime, depending on whether
the regulation exists for irrigation, hydroelectricity
generation, river transport or other purposes (Ward &
Stanford, 1979; Petts, 1984; Calow & Petts, 1992).
Riverine fishes are some of the more conspicuous
victims of river regulation, with flow alteration and
the structures which effect it, being implicated in
disrupting life histories, degrading habitat, impeding
movement, affecting food webs and so on (see Ward
& Stanford, 1979; Petts, 1984; Welcomme, 1985;
Walker, 1986, 1992; Matthews, 1998). The result of
this is, as a general rule, to alter the composition of
fish assemblages; with intolerant species declining in
abundance or becoming locally extinct and opportu-
nistic native and alien species often coming to dom-
inate (e.g. Gehrke et al., 1995; Aarts, Van den Brink &
Nienhuis, 2004; Lamouroux & Cattaneo, 2006). Barri-
ers to movement exacerbate the effects of flow
alteration by preventing movement into and out of
affected areas and preventing recolonization (Mat-
thews, 1998; Lucas & Baras, 2001).
Whilst many studies exist which highlight the likely
effects of river regulation on fish (see above reviews),
most have been of relatively short duration, which
limits their ability to draw meaningful conclusions.
Moreover, few are able to make definite links between
hydrology and characteristics of fish assemblages and
populations, such as composition, abundance/bio-
mass, recruitment and movement, because appropriate
spatial and/or interannual comparisons are unable to
be made (see Cattaneo et al., 2002; Daufresne et al., 2003
for notable exceptions). For example, although the
study by Gehrke et al. (1995) suggests that the more
altered the flow of a river is, the fewer natives species
and the greater the dominance by alien species, the
results are based on two sampling occasions (despite a
good geographic spread) and so questions remain
about temporal variation. It is only by conducting
multi-year studies, under a range of hydrological
conditions, such as floods, droughts and ‘average’
flows, that we are likely to be able to establish how
hydrology influences fish faunal characteristics (Lloyd
et al., 2003; Lytle & Poff, 2003). By so doing, this will
provide the type of knowledge needed for effective
management of regulated rivers.
790 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

Australian lowland rivers and their fish faunas, like
many throughout the world, are in a parlous state
(Murray-Darling Basin Commission, 2004). The highly
regulated Campaspe River, in northern Victoria, is
one example of a severely degraded river system,
with a fish fauna dominated by alien species and few
native species having self-sustaining populations
(Humphries & Lake, 2000; Humphries, Serafini &
King, 2002). The Campaspe River provides an oppor-
tunity to study the effects of flow alteration on a fish
fauna because, below the major regulating structure
(Lake Eppalock) there are three river sections, each
with its own distinct hydrology, all within about 175
river km from its junction with the Murray River at
Echuca. Lake Eppalock stores winter and early spring
runoff, releasing it at a later date, primarily for
irrigation purposes. Unless a major rainfall event
occurs, the ‘upper’ section of the Campaspe River
typically experiences highly regulated, enhanced
flows for 6 months over later spring-early autumn,
whereas the ‘middle’ section only receives enhanced
flow for about 2 months during autumn and the
‘lower’ section receives only minor ‘maintenance’
flows for most of the year.
We present here the results of a 7-year study of the
Campaspe River below Lake Eppalock, in which we
compare the fish faunas in the three hydrologically-
distinct river sections. We were thus able to investi-
gate the role of river section (with contrasting
hydrologies) and year (the period of the study
included one high-flow, one moderate-flow and five
low-flow years), focusing on components of the fauna
which we considered would give us insight into the
role of hydrology in structuring fish assemblages and
populations (species composition, abundance/bio-
mass and recruitment) and correlated these with
hydrological variables. To strengthen our study fur-
ther, we included, in the final 3 years of the study, a
comparison of the fish fauna of the Campaspe River
with that of the much less regulated nearby Broken
River. This has allowed us to speculate on how much
of an effect barriers to movement, since both rivers
contained significant weirs, and other environmental
variables had on characteristics of the fish faunas from
each river.
The current paper thus aims to (i) determine how
the composition, species richness, abundance and
biomass of the fish fauna of the Campaspe River
varied among the three hydrologically distinct sec-
tions of river and among the 7 years of the study, with
the prediction that the greater the degree of flow
alteration, the fewer species and the greater the
dominance of alien species; (ii) relate characteristics
of the fish fauna (specifically, composition, species
richness, abundance of dominant species and juvenile
recruitment) to hydrological variables (high, low,
average and threshold flows for fish passage up-
stream of weirs) of the Campaspe River among the
three river sections and for two seasons (‘winter’ and
‘spring/summer’), with the prediction that the fish-
related variables will respond more to high and
threshold flows because of greater ability for fish to
move between river sections during these times; and
(iii) compare the composition, species richness, abun-
dance and biomass of fish in the Campaspe River with
that of the less regulated and nearby Broken River,
with the prediction that there will be more native
species, and that they will comprise a larger propor-
tion of the fish fauna, in the Broken than in the
Campaspe River. We emphasize spatial and temporal
differences and also speculate on the proximate and
ultimate roles of flow in shaping the Campaspe River
fish fauna.
Methods
Study area and hydrology
The Campaspe River is located in the southern
Murray-Darling Basin and is a tributary of the Murray
River (Fig. 1). Since the early 1960s, c. 50% of the
mean annual discharge of about 2 · 108 m3 has been
stored in Lake Eppalock for later release downstream
(Fig. 2a). The river downstream of Lake Eppalock can
be divided into three sections, based on hydrology
and the presence of weirs: the ‘upper’ section from
Lake Eppalock to Campaspe Weir (c. 9 m high); the
‘middle’ section from Campaspe Weir to Campaspe
Siphon (c. 2 m high) and the ‘lower’ section from
Campaspe Siphon to the confluence with the Murray
River (Fig. 1). There is also a small, c. 1 m high,
gauging weir in the Campaspe River, near Echuca.
The entire river downstream of Lake Eppalock expe-
riences reduced duration of high winter flows due to
the capture of water (Fig. 2a). In addition, the upper
and middle sections of the river experience approx-
imately 6 and 2 months, respectively, of enhanced
summer flows because of irrigation releases. Most of
Regulated river fish fauna 791
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

this irrigation flow is extracted from the river at the
Campaspe Siphon and so the lower section has only
marginally enhanced flows at this time. Prior to our
study, Lake Eppalock filled and spilled approximately
every 2 years during which time high flows would
pass down the entire length of the river (Fig. 2b). This
only occurred once, early in the present study. Before
the construction of Lake Eppalock and the weirs (pre-
regulation), the Campaspe River would have ceased
to flow during the dry summer period in most years.
This no longer occurs.
Long (up to 1 km), deep (up to 8 m) and still or very
slow flowing natural pools are common in the upper
section of the Campaspe River and are separated by
relatively shallow (0.3–1.5 m), still to moderate veloc-
ity (0–0.5 m s)1) run habitats. The middle and lower
sections of the river have no large natural pools and
consist mostly of run habitat, characterized by alter-
nating shallow (<1 m) and deep (1–3 m) regions. The
river immediately downstream of Lake Eppalock is
typically 30–40 m wide, with high banks and few
anastomosing channels or oxbow lakes. Further
downstream, the river becomes less constrained
(40–50 m wide mostly), in places it has multiple
channels and is more likely to move out on to the
floodplain. The dominant instream structure is coarse
woody debris (or ‘snags’) mainly from riparian river
redgums, Eucalyptus camaldulensis (Dehnhardt) and
stands of aquatic macrophytes, mostly common reed,
Phragmites australis (Cav). Records indicate that the
Campaspe River once supported approximately 20
species, however, recent evidence suggests that the
fish fauna is now highly degraded, and dominated by
the alien species common carp, Cyprinus carpio L, and
European perch, Perca fluviatilis L (Humphries &
Lake, 2000; Humphries et al., 2002).
Between 1993 and 2002, approximately 182 000
golden perch juveniles (0.8–1.6 g) were released into
the Campaspe River below Lake Eppalock: 20 000–
45 000 were stocked each year except in 1996 and
1997. Official records indicate that stocking of Murray
cod juveniles in the Campaspe River below Lake
Eppalock only took place in 2001 (5000 upper, 10 000
lower section), 2002 (19 000 juveniles and 25 1 year
olds, middle section) and 2003 (10 000 upper and
middle sections).
The Broken River is a tributary of the Goulburn
River, which itself is a tributary of the Murray River
(Fig. 1). It was chosen as a reference river with which
to compare the fish fauna of the Campaspe River,
because it is less regulated than the Campaspe River,
with a smaller fraction of its mean annual discharge of
2 · 108 m3 diverted for offstream use and responds
more naturally to rainfall events in the catchment than
does the Campaspe River (Smith & Humphries, 1997).
The impoundments, Lakes Nillahcootie (built 1967)
and Mokoan (built 1971), are the primary regulating
bodies in the Broken River. Two weirs (Casey and
Fig. 1 Map of Campaspe and Broken Rivers, showing major
storages and delineating upper, middle and lower sections of
each river included in study.
0
5
10
15
0
5
10
15
Jan Mar May Jul Sep NovMonth
Dis
char
ge (
m3 s
–1)
Pre-dam
Post-dam
(a)
(b)
Fig. 2 Median mean monthly discharges for the (a) upper
section and (b) lower section of the Campaspe River prior to the
construction of Lake Eppalock (pre-dam, 1885–1908) and after
the construction of Lake Eppalock (post-dam, 1977–94).
792 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

Gowangardie Weirs, both c. 3 m high) downstream of
the major impoundments, whilst creating pools
upstream and inhibiting fish movement, have a
relatively small effect on the hydrology of the river.
The Broken River was similarly divided into three
sections: Benalla Weir to Casey Weir, the ‘upper’
section; Casey Weir to Gowangardie Weir, the
‘middle’ section and Gowangardie Weir to the
confluence with the Goulburn River at Shepparton,
the ‘lower’ section (Fig. 1). The Broken River channel
width is marginally smaller than the Campaspe
(typically 25–35 m) and has no large natural pools
comparable to those in the upper Campaspe River,
however, the run habitat is similar to the Campaspe,
comprising alternating shallow (<1 m) and deep
(1–3 m) areas with still to moderate velocities
(0–0.5 m s)1). Instream structure similarly comprises
snags, derived from riparian river redgums, and
aquatic macrophytes, dominated by common reed.
Historically, the Campaspe and Broken Rivers
would have had a similar suite of fish species
(Humphries & Lake, 2000). The native fish fauna of
the Broken River has no doubt also suffered as a result
of a variety of anthropogenic disturbances, however,
nowadays there are still approximately 12 native
species extant in the Broken River, with several large
and small native species present in significant
numbers (Humphries & Lake, 2000). Stocking of
golden perch (0.8–1.5 g) also occurred in this river
during the current study at the rate of, typically,
20 000 juveniles annually (Victorian Department of
Primary Industries, unpubl. data). In 1993 there were
no records of golden perch being stocked, and in 2002,
9000 juveniles and 25 1-year-old fish were stocked.
Collection and processing of fish
Fish were collected from two reaches, randomly
chosen from a range of possible reaches, on each
occasion, from within each section of the Campaspe
River bimonthly between October 1995 and June 2000.
Because of changes in funding, between August 2000
and February 2003 the sampling frequency decreased
to four times per year: February, June, October and
December. During the latter period, sampling also
included two randomly chosen reaches within each
section of the Broken River. The Campaspe and
Broken Rivers were sampled within the same week
on each occasion and following identical protocols.
Thus our sampling consisted of a total of eight
replicates per section per year, including four from
each reach. This frequency of sampling is twice and
sometimes four times that of most comparable studies
(e.g. Gehrke et al., 1995) and it encompasses the warm
and cool, wet and dry and non-breeding and breeding
times of the year.
Sampling over the first year of the study using large
and small fyke nets, bait traps, seines, dip nets and
back-pack and boat electrofishers indicated that a
combination of large and small fyke nets was able to
be used in a consistent manner, and was able to be
deployed in all river conditions and captured juve-
niles and adults of the large-bodied species and most
of the smaller-bodied species in similar proportions to
a combination of all the other methods trialled. Fyke
nets were not efficient in catching Australian smelt or
gambusia, which were better collected using light
traps and/or other active methods (see Humphries
et al., 2002 and P. Humphries & A. Richardson,
unpubl. data). Therefore, fish were collected using a
combination of seven large fyke nets (35 mm mesh,
0.5 m diameter mouth, 3 m long, 5 m central wing)
and three small fyke nets [4 mm mesh, 0.5 m mouth
(with 30 mm mesh at front to prevent large fauna
from entering), 1.3 m long, 3 m central wing]. Nets
were set facing downstream at an angle to the bank,
randomly along a 100–350 m reach, approximately
3 h before dusk and retrieved within approximately
4 h after dawn the next day.
Opportunistic electrofishing sampling by backpack
(Smith-Root model 15-C POW and model 12-A;
Smith-Root, Vancouver, WA, U.S.A.) and boat
(Smith-Root model 5.0 GPP; Smith-Root, Vancouver,
WA, U.S.A.) was undertaken between October 1995
and June 2000. However, because this method could
not be carried out consistently due to frequent
difficulties gaining access to the river, the results
from this sampling are only included in a description
of the presence of species in the river as a whole, over
time (Fig. 4) and were not included in any other
analyses.
Fish were anaesthetized, identified to species,
weighed to 1 g (if fish were less than 1 g, they were
pooled and the total weight of that species for a net was
determined) and measured to 1 mm. All fish, except
common carp (which, by law must be destroyed), were
then allowed to recover in clean water, before being
returned to the river. It is recognized that some small
Regulated river fish fauna 793
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

species will not be sampled as efficiently by fyke nets as
other gear (see below in Discussion), but the consis-
tency of sampling for all reaches and sampling occa-
sions using fyke nets allows for meaningful
comparisons throughout the entire study.
Environmental and hydrological data
Duplicate measurements of temperature, dissolved
oxygen, turbidity, conductivity and pH were taken at
each reach on each sampling occasion using a Horiba�
Water Checker U-10 (Horiba, Kyoto, Japan).
Hydrological data were obtained from Goulburn-
Murray Water through gauging stations operated by
Thiess Environmental Services. Gauging stations are
located at Lake Eppalock, Campaspe Weir and
Campaspe Siphon on the Campaspe River (thus
providing flow records for each of the three sections
of the river) and at Benalla and Casey Weir on the
Broken River (only providing flow records for the
upper section and the combined middle and lower
sections) (Fig. 1).
Analysis of fish data
The occurrence of species of fish in the Campaspe
River over time was described by the presence or
absence of species derived from samples collected
using backpack and boat electrofishing and large and
small fyke netting. All other analyses were carried out
on fyke net data only (seven large and three small
fyke nets per reach), since these were employed
consistently throughout the entire study.
Relative contribution of species to the fish fauna
was used as an overall descriptor of the fauna and
was derived from fish abundance and biomass data
for each section of the Campaspe River, calculated
from the summed values of fish collected in two
reaches within that section (from 14 large and six
small fyke nets per section per sampling trip) for the
entire study period (October 1995–February 2003).
Relative contribution of species to the Broken River
fish fauna, between August 2000 and February 2003,
were calculated in the same way and were compared
with data from the Campaspe River obtained during
the same period.
An overall description of the fish fauna of the
Campaspe River over time and space was performed
using non-metric multidimensional scaling (NMDS)
analysis on fish abundance data collected using fyke
nets. Rare species were excluded from the NMDS; rare
species were those that occurred in samples only once.
Data were log10(x + 1) transformed and a Bray-Curtis
dissimilarity matrix calculated. Fifty random starts
were used. To determine whether fish assemblages
differed significantly with ‘Year’ and ‘Section’ two-way
analysis of similarity (ANOSIMANOSIM) was carried out. A
‘year’ was defined as being a spawning year: beginning
in August of 1 year and ending in June of the following.
NMDS and ANOSIMANOSIM were carried out using the
PRIMER V5 package (Clark & Warwick, 2001).
To determine if species richness and abundance
differed significantly over the 7-year study period and
among river sections in the Campaspe River, four of
the bimonthly sampling occasions per year were used
as replicates (June, October, December and February),
since these were sampled for the entire study period.
Two-way ANOVAANOVAs were carried out examining the
effects of ‘Year’ and ‘Section’ on species richness,
mean abundance of total fish, common carp, golden
perch, European perch and flathead gudgeon. These
four species were the only species collected in suffi-
cient abundance to allow such a comparison. Richness
data met the assumptions of ANOVAANOVA and so raw
values were used, whereas abundance data were
log10(x + 1) transformed to satisfy the assumptions of
ANOVAANOVA.
Recruitment
Recruitment was investigated by determining the
number of young-of-year (YOY) fish collected in any
1 year. A ‘recruitment season’ was assumed to begin
approximately 2 months after the commencement of
spawning (although, in the case of golden perch, this
was not possible, because no larvae were ever
collected), when fish would have become juveniles
and it is assumed that they would have already
suffered the bulk of mortality, and to finish before
spawning commenced in the following season. Thus,
the ‘recruitment season’ for common carp and Euro-
pean perch was designated from October to June and
for golden perch, from December to June. YOY carp
were designated as fish up to 125 mm (Brumley, 1996;
Villizi & Walker, 1999), YOY European perch as fish
up to 100 mm (Shafi & Maitland, 1971) and YOY
golden perch as fish up to 200 mm (Anderson,
Morison & Ray, 1992).
794 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

Relationships between hydrological and fish variables
The Echuca gauging weir is only about 1 m in height,
but its ‘drowning-out’ is a function of discharge of
both the Campaspe River and that of the Murray
River, since the waterlevel backs up the Campaspe,
when the Murray River runs high. Analysis of
hydrological data, consultation with Goulburn-
Murray Water hydrologists and personal observations
suggest that a discharge of approximately
23.15 m3 s)1 or above in the Campaspe River would
make upstream fish passage possible. The Campaspe
Siphon is constructed of a single, concrete apron,
rounded in section and set between relatively steep
banks. It is estimated that this structure, despite being
higher than the Echuca weir, is similarly ‘drowned-
out’ at approximately 23.15 m3 s)1, and personal
observations suggest that the discharge at which
upstream fish passage is possible would be at this
level or marginally lower. Thus, 23.15 m3 s)1 is taken
as the threshold level above which upstream fish
passage from the lower section to the middle section
of the Campaspe Rivers is possible. It is estimated
from field observations and confirmed by hydrolo-
gists from Goulburn-Murray Water, that upstream
fish passage through the 9 m Campaspe Weir would
be possible at around 185.18 m3 s)1, which is close to
minor flood level (c. 243.06 m3 s)1; B. Viney, pers.
comm.). It is at this discharge that there is sufficient
flow over and around the weir to provide a range of
velocities likely to allow fish passage. Thus,
185.18 m3 s)1 is taken as the threshold level above
which upstream fish passage from the middle section
to the upper section of the Campaspe Rivers is
possible. It is presumed, for the purposes of this
study, that downstream passage over all weirs is
possible at all times.
To relate the hydrology of the Campaspe River with
characteristics of the fish fauna (composition, species
richness, abundance and recruitment), a number of
hydrological variables were calculated (Table 1).
Analysis was conducted using the River Analysis
Package 2.0.1 (Marsh, Stewardson & Kennard, 2003).
Two periods of the year were chosen for separate
analysis, representing the ‘winter’ or non-growing
season (April–July) and the ‘spring/summer’ or
growing season (August–March), in a similar manner
to Cattaneo et al. (2002). The hydrology of each river
section – upper, middle and lower – was treated
separately. Hydrological variables were chosen to
take into account high (maximum flow, number and
duration of 10th percentile flows), low (minimum
flow, number and duration of 90th percentile flows),
median and variability of flows, as well as number
and duration of spells above threshold levels of
23.15 m3 s)1 (which is estimated as the flow above
which upstream fish passage is possible over the 1 m
Echuca gauging weir and the 2 m Campaspe Siphon,
see below) and 185.18 m3 s)1 (which is estimated as
the flow above which upstream fish passage is
possible over the 9 m Campaspe Weir).
Aspects of the fish fauna of the Campaspe River
were related to hydrological variables in two ways.
Firstly, a BIOENV analysis was performed on the
NMDS carried out earlier; BIOENV compares envi-
ronmental data, in this case the hydrological variables
described above, with the dissimilarity matrix derived
from the fish data by maximising a rank correlation
between their respective (dis)similarity matrices (PRI-
MER V5 package, Clark & Warwick, 2001). This was
done for the ‘winter’ and ‘spring/summer’ periods
separately. Secondly, a series of stepwise multiple
regressions was carried out to determine the effects of
the hydrological variables (as described above,
Table 1) on species richness, log10(x + 1) transformed
total abundance of all fish and the total abundance of
the dominant species (common carp, European perch,
golden perch, flathead gudgeon) and log10(x + 1)
transformed total abundance of recruits, i.e. YOY
common carp, European perch and golden perch. The
7 years were used as replicates. Each river section and
hydrological season (winter and spring/summer) was
analysed separately.
All analyses, except NMDS, ANOSIMANOSIM and BIOENV
were performed using Excel 2002 (Microsoft Inc.,
Sydney, NSW, Australia) and SPSS Version 14 (SPSS
Inc., Chicago, IL, U.S.A.).
Results
Environmental and hydrological variables
For virtually the entire seven and a half-year period of
the study, the Campaspe River experienced the failure
of normal winter rains (Fig. 3a). Only at the com-
mencement of the study, in the winter of 1996, and to
a lesser degree in the winter of 2000, was there
substantial runoff into the river, causing significant
Regulated river fish fauna 795
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813
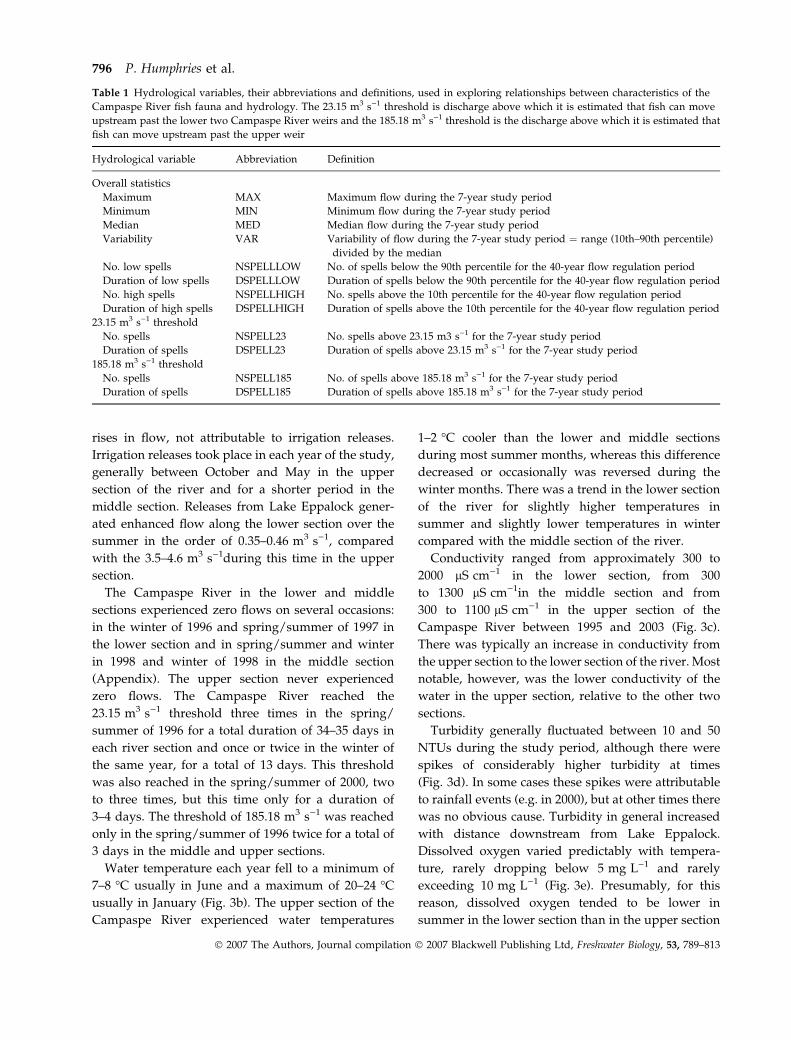
rises in flow, not attributable to irrigation releases.
Irrigation releases took place in each year of the study,
generally between October and May in the upper
section of the river and for a shorter period in the
middle section. Releases from Lake Eppalock gener-
ated enhanced flow along the lower section over the
summer in the order of 0.35–0.46 m3 s)1, compared
with the 3.5–4.6 m3 s)1during this time in the upper
section.
The Campaspe River in the lower and middle
sections experienced zero flows on several occasions:
in the winter of 1996 and spring/summer of 1997 in
the lower section and in spring/summer and winter
in 1998 and winter of 1998 in the middle section
(Appendix). The upper section never experienced
zero flows. The Campaspe River reached the
23.15 m3 s)1 threshold three times in the spring/
summer of 1996 for a total duration of 34–35 days in
each river section and once or twice in the winter of
the same year, for a total of 13 days. This threshold
was also reached in the spring/summer of 2000, two
to three times, but this time only for a duration of
3–4 days. The threshold of 185.18 m3 s)1 was reached
only in the spring/summer of 1996 twice for a total of
3 days in the middle and upper sections.
Water temperature each year fell to a minimum of
7–8 �C usually in June and a maximum of 20–24 �C
usually in January (Fig. 3b). The upper section of the
Campaspe River experienced water temperatures
1–2 �C cooler than the lower and middle sections
during most summer months, whereas this difference
decreased or occasionally was reversed during the
winter months. There was a trend in the lower section
of the river for slightly higher temperatures in
summer and slightly lower temperatures in winter
compared with the middle section of the river.
Conductivity ranged from approximately 300 to
2000 lS cm)1 in the lower section, from 300
to 1300 lS cm)1in the middle section and from
300 to 1100 lS cm)1 in the upper section of the
Campaspe River between 1995 and 2003 (Fig. 3c).
There was typically an increase in conductivity from
the upper section to the lower section of the river. Most
notable, however, was the lower conductivity of the
water in the upper section, relative to the other two
sections.
Turbidity generally fluctuated between 10 and 50
NTUs during the study period, although there were
spikes of considerably higher turbidity at times
(Fig. 3d). In some cases these spikes were attributable
to rainfall events (e.g. in 2000), but at other times there
was no obvious cause. Turbidity in general increased
with distance downstream from Lake Eppalock.
Dissolved oxygen varied predictably with tempera-
ture, rarely dropping below 5 mg L)1 and rarely
exceeding 10 mg L)1 (Fig. 3e). Presumably, for this
reason, dissolved oxygen tended to be lower in
summer in the lower section than in the upper section
Table 1 Hydrological variables, their abbreviations and definitions, used in exploring relationships between characteristics of the
Campaspe River fish fauna and hydrology. The 23.15 m3 s)1 threshold is discharge above which it is estimated that fish can move
upstream past the lower two Campaspe River weirs and the 185.18 m3 s)1 threshold is the discharge above which it is estimated that
fish can move upstream past the upper weir
Hydrological variable Abbreviation Definition
Overall statistics
Maximum MAX Maximum flow during the 7-year study period
Minimum MIN Minimum flow during the 7-year study period
Median MED Median flow during the 7-year study period
Variability VAR Variability of flow during the 7-year study period ¼ range (10th–90th percentile)
divided by the median
No. low spells NSPELLLOW No. of spells below the 90th percentile for the 40-year flow regulation period
Duration of low spells DSPELLLOW Duration of spells below the 90th percentile for the 40-year flow regulation period
No. high spells NSPELLHIGH No. spells above the 10th percentile for the 40-year flow regulation period
Duration of high spells DSPELLHIGH Duration of spells above the 10th percentile for the 40-year flow regulation period
23.15 m3 s)1 threshold
No. spells NSPELL23 No. spells above 23.15 m3 s)1 for the 7-year study period
Duration of spells DSPELL23 Duration of spells above 23.15 m3 s)1 for the 7-year study period
185.18 m3 s)1 threshold
No. spells NSPELL185 No. of spells above 185.18 m3 s)1 for the 7-year study period
Duration of spells DSPELL185 Duration of spells above 185.18 m3 s)1 for the 7-year study period
796 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813
and vice versa in winter. pH was generally between 7
and 8 in all sections of the Campaspe River during the
study and there were no consistent differences among
river sections (Fig. 3f).
Composition of the Campaspe River fish fauna
A total of 16 species, 10 native and six alien, were
recorded from the Campaspe River between October
5
6
7
8
9
O J A S D M A N F J O J A S D M A N F J O J A D
0
(f)
5
10
15 (e)
0
50
100
150 (d)
0500
1000150020002500
05
1015202530
(c)
(b)
1995 1996 1997 1998 1999 2000 2001 2002
(a)
pHD
isso
lved
oxy
gen
(mg
L–1)
Tur
bidi
ty (
NT
U)
Con
duct
ivity
(µS
cm
–1)
Tem
pera
ture
(°C
)D
isch
arge
(m
3 s–1
)
Date
0
10
20
30
40
50
60
Fig. 3 Discharge, temperature, conductivity, turbidity, dissolved oxygen and pH measurements taken for the upper (dotted line),
middle (dashed) and lower (solid) sections of the Campaspe River between October 1995 and February 2003. Vertical dashed line
represents when bimonthly sampling changed to sampling in October, December, February and June.
Regulated river fish fauna 797
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813
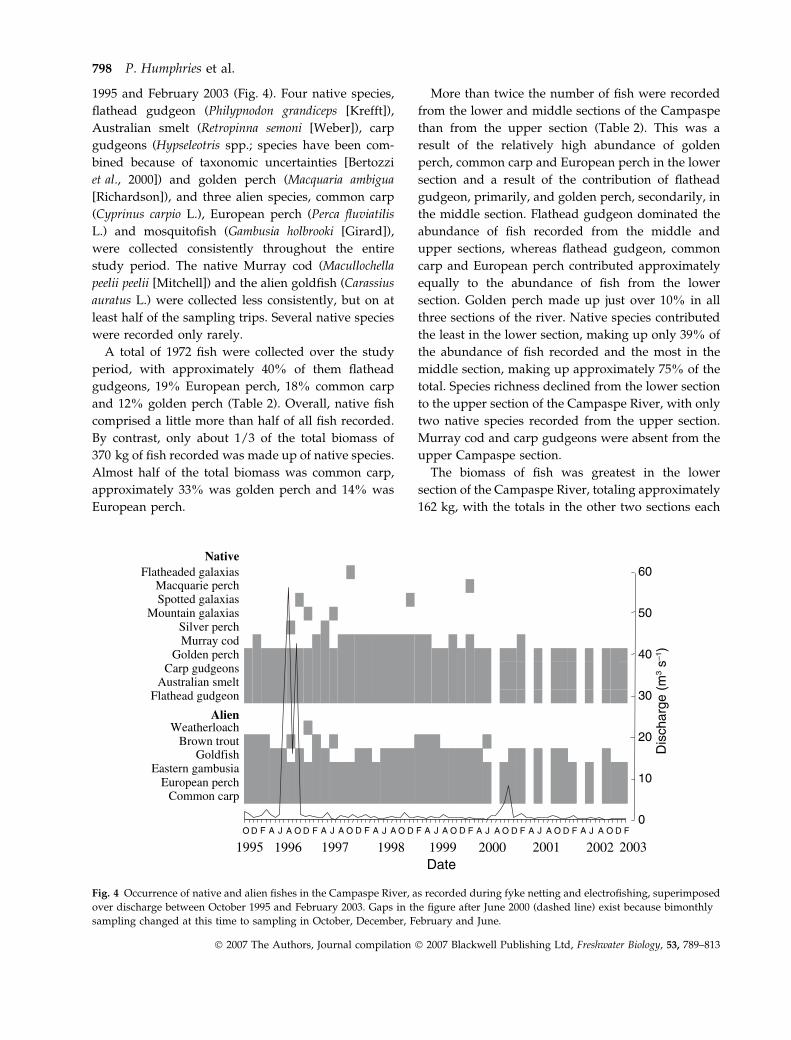
1995 and February 2003 (Fig. 4). Four native species,
flathead gudgeon (Philypnodon grandiceps [Krefft]),
Australian smelt (Retropinna semoni [Weber]), carp
gudgeons (Hypseleotris spp.; species have been com-
bined because of taxonomic uncertainties [Bertozzi
et al., 2000]) and golden perch (Macquaria ambigua
[Richardson]), and three alien species, common carp
(Cyprinus carpio L.), European perch (Perca fluviatilis
L.) and mosquitofish (Gambusia holbrooki [Girard]),
were collected consistently throughout the entire
study period. The native Murray cod (Macullochella
peelii peelii [Mitchell]) and the alien goldfish (Carassius
auratus L.) were collected less consistently, but on at
least half of the sampling trips. Several native species
were recorded only rarely.
A total of 1972 fish were collected over the study
period, with approximately 40% of them flathead
gudgeons, 19% European perch, 18% common carp
and 12% golden perch (Table 2). Overall, native fish
comprised a little more than half of all fish recorded.
By contrast, only about 1/3 of the total biomass of
370 kg of fish recorded was made up of native species.
Almost half of the total biomass was common carp,
approximately 33% was golden perch and 14% was
European perch.
More than twice the number of fish were recorded
from the lower and middle sections of the Campaspe
than from the upper section (Table 2). This was a
result of the relatively high abundance of golden
perch, common carp and European perch in the lower
section and a result of the contribution of flathead
gudgeon, primarily, and golden perch, secondarily, in
the middle section. Flathead gudgeon dominated the
abundance of fish recorded from the middle and
upper sections, whereas flathead gudgeon, common
carp and European perch contributed approximately
equally to the abundance of fish from the lower
section. Golden perch made up just over 10% in all
three sections of the river. Native species contributed
the least in the lower section, making up only 39% of
the abundance of fish recorded and the most in the
middle section, making up approximately 75% of the
total. Species richness declined from the lower section
to the upper section of the Campaspe River, with only
two native species recorded from the upper section.
Murray cod and carp gudgeons were absent from the
upper Campaspe section.
The biomass of fish was greatest in the lower
section of the Campaspe River, totaling approximately
162 kg, with the totals in the other two sections each
Native
Flathead gudgeonAustralian smelt
Carp gudgeonsGolden perch
Murray codSilver perch
Mountain galaxiasSpotted galaxiasMacquarie perch
Flatheaded galaxias
Alien
Common carpEuropean perch
Eastern gambusiaGoldfish
Brown troutWeatherloach
O D F A J A O D F A J A O D F A J A O D F A J A O D F A J A O D F A J A O D F A J A O D F
1995 1996 1997 1998 1999 2000 2001 2002 2003Date
Dis
char
ge (
m3
s–1)
0
10
20
30
40
50
60
Fig. 4 Occurrence of native and alien fishes in the Campaspe River, as recorded during fyke netting and electrofishing, superimposed
over discharge between October 1995 and February 2003. Gaps in the figure after June 2000 (dashed line) exist because bimonthly
sampling changed at this time to sampling in October, December, February and June.
798 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

approximately 100 kg (Table 2). Common carp con-
tributed the most biomass to the fauna in each river
section (44–48%), with golden perch second
(29–37%). The only other species to contribute more
than 10% was European perch. Native species com-
prised 33–39% of the total biomass of fish from each
reach. Common carp and European perch together
comprised approximately 60% in each reach.
Non-metric multidimensional scaling analysis and
ANOSIMANOSIM of the fish abundance data for the 7 years
and three river sections indicated that there were
different assemblages among sections (global
R ¼ 0.231, P < 0.01), but not among years (global
R ¼ )0.185, P > 0.05) (Fig. 5). Post hoc comparisons
showed that there was no difference in the assem-
blages between the lower and middle sections (R ¼0.086, P > 0.05), but there were differences between
the lower and upper sections (R ¼ 0.285, P < 0.05)
and the middle and upper sections (R ¼ 0.357,
P < 0.01).
1996
1997
1998
1999
2000
2001
2002
1996
1997
1998
1999
2000
2001
2002
1996
1997
1998
19992000
20012002
Stress: 0.2
Lower
Middle
Upper
Fig. 5 Non-metric multidimensional
scaling analysis plot of fish assemblage
data for the three sections and 7 years
from the Campaspe River.
Table 2 Percentage abundance and biomass, total abundance and biomass,% natives, species richness total and% species richness of
natives of fish collected in the Campaspe River between October 1995 and June 2003
Species
Lower Middle Upper Total
No. Biomass (kg) No. Biomass (kg) No. Biomass (kg) No. Biomass (kg)
Native
Flathead gudgeon 25.1 0.3 56.9 0.9 52.8 0.4 43.0 0.5
Golden perch 12.7 37.1 11.6 29.1 12.3 32.6 12.2 33.6
Carp gudgeons 0.7 <0.1 1.7 <0.1 1.0 <0.01
Murray cod 0.4 1.3 2.3 5.8 1.1 2.2
Australian smelt 0.1 <0.1 2.2 <0.1 0.9 <0.01
Silver perch 0.1 <0.1 0.1 0.1 <0.01
Flatheaded galaxias 0.1 <0.1 0.1 <0.01
Alien
Common carp 30.6 46.7 8.1 43.8 10.2 47.9 17.8 46.2
European perch 25.6 11.2 12.3 19.1 18.4 11.9 19.0 13.6
Goldfish 3.3 3.3 1.3 1.2 4.5 2.8 2.7 2.6
Eastern gambusia 1.1 <0.1 3.3 <0.1 1.8 <0.01
Brown trout 1.8 4.5 0.4 1.2
Carp/goldfish hybrid 0.1 <0.1 0.1 <0.01
Total 812 161951 778 104206 381 103068 1972 369225
Natives (%) 39.2 38.8 74.8 35.8 65.1 33 58.3 36.3
Species richness total 12 11 6 13
Species richness natives (%) 58.3 54.5 33.3 53.8
Regulated river fish fauna 799
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

Species richness and abundance of dominant species
Species richness and the abundance of total fish and
flathead gudgeons varied significantly with section of
river, whereas abundance of common carp and
European perch varied with section and year (Table 3,
Fig. 6). In the case of species richness and all species,
the mean square was greatest for section. Interaction
terms for all variables were non-significant and
abundance of golden perch did not vary significantly
with either factor. There were more species in the
lower and middle sections than in the upper section
(Tukey’s test, P < 0.001 and P < 0.01, respectively),
but there were similar numbers of species collected in
the lower and middle sections. For total fish, common
carp and European perch, Tukey’s test indicated that
there were significantly more fish in the lower section
than in the upper section of the Campaspe River
(P < 0.01). There were also more total fish and
flathead gudgeons in the middle section than in the
upper section of the river (P < 0.05 and P < 0.01,
respectively) and more common carp and European
perch in the lower section than in the middle section
(P < 0.05 and P < 0.01 respectively). There was evi-
dence for a gradual, but consistent decline in abun-
dance of European perch from 1996 until 2002,
whereas the abundance of common carp declined
from 1996 to 1999, increased in 2000 and subsequently
declined again to February 2003 (Fig. 6).
Recruitment
Common carp, European perch and golden perch
were the only species which were caught consistently
and in sufficient numbers to adequately examine
patterns in population structure (0+ versus ‡1+) and
recruitment. There were almost no YOY carp collected
in the upper section of the Campaspe River through-
out the entire study, only a small number in the
middle section and highly variable, but sometimes
large numbers in the lower section (Fig. 7a). YOY carp
were highly abundant and dominated the carp pop-
ulation in the 2000/01 season and made up approx-
imately 50% of the fish collected in the 1996/97
season.
In a similar pattern to carp, there were virtually no
YOY European perch in the upper section and very
few in the middle section of the Campaspe River
throughout the study period (Fig. 7b). Only in the first
year of the study were YOY collected in the middle
section, and these made up approximately 1/3 of all
fish collected in that season. By contrast, YOY Euro-
pean perch were collected in most years in the lower
section of the river. Furthermore, apart from the
2000/01 and 2001/02 seasons, the abundance of YOY
European perch in the lower section was relatively
consistent among years.
There were small numbers of YOY golden perch
collected in most years in each of the three sections of
the Campaspe River during the study period (Fig. 7c).
Only in 1996/97 in the lower section were YOY
golden perch reasonably abundant and made up a
substantial proportion of all fish collected.
Relationships between hydrological and fish variables
The BIOENV analysis relating fish assemblage com-
position with hydrological variables, using all 7 years’
data, but for each section separately, indicated that
there were no significant relationships either for the
‘winter’ or the ‘spring/summer’ seasons (Spearman’s
r ¼ 0.127 and r ¼ 0.108 respectively). However, when
multiple regressions were carried out relating species
richness, the abundance of individual species and the
abundance of YOY common carp, European perch
and golden perch with the hydrological variables,
several significant relationships were found (Table 4).
The fish variables that were positively related to high
flow variables were golden perch, European perch
(lower section, winter), and common carp and species
Table 3 Mean squares and significance
levels for two-way A N O V AA N O V As of species
richness and abundance of total fish,
flathead gudgeon, golden perch, common
carp and European perch by ‘Section’ and
‘Year’ for the Campaspe River between
October 1995 and February 2003
Source of
variation d.f.
Species
richness Total
Flathead
gudgeon
Golden
perch
Common
carp
European
perch
Section 2 14.250*** 0.841*** 1.178** 0.207 0.950** 0.845***
Year 6 2.067 0.038 0.407 0.103 0.426** 0.272**
Section · Year 12 1.264 0.103 0.141 0.131 0.145 0.147
Error 63 1.31 0.111 0.206 0.082 0.135 0.087
**P < 0.01, ***P < 0.001.
800 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

richness (middle, spring/summer). The fish variables
that were positively associated with low flow vari-
ables were total fish and YOY common carp (lower
section, in winter), YOY golden perch (upper, winter)
and European perch (middle, spring/summer). Total
fish (lower, winter) and flathead gudgeon (middle,
winter) were significantly associated with variability
in flows, the former positively, and the latter nega-
tively. YOY golden perch (lower, winter) and total fish
(lower, spring/summer) were positively related to the
threshold flow for the lower two weirs (23.15 m3 s)1),
whereas for YOY common carp, the relationship was
less clear; abundance of this group was positively
related to the number of spells over the 23.15 m3 s)1
and negatively related to the duration of these spells
and maximum flow in the lower section in spring/
summer.
Comparison of the Campaspe and Broken River
environments: 2000–2003
The Campaspe and Broken Rivers experienced
drought conditions for most of the period between
October 2000 and February 2003 (Figs 3a & 8a). The
only time when significant rainfall and runoff
occurred was during the spring of 2000, when mean
monthly discharge in the Broken River peaked at
approximately 28.94 m3 s)1 and in the Campaspe
River at approximately 11.57 m3 s)1.
Mean monthly water temperatures were generally
similar for the Campaspe and Broken Rivers, although
the latter reached marginally higher levels in mid-
summer and mid-winter than the former (Figs 3b &
8b). Conductivity was always lower in the Broken
River than the Campaspe River (Figs 3c & 8c). There
was a gradual rise in conductivity over the 3-year
study period in the Broken River, however, levels
never reached 400 lS cm)1, whereas they were rarely
below 400 lS cm)1 in the Campaspe River, even in the
upper section. Turbidity, on the other hand, was often
higher in the Broken River than the Campaspe River
(Figs 3d & 8d): the 3-year mean turbidity in the lower
and upper Broken River was 51.0 ± 5.1 and 34.3 ± 1.8
NTU, respectively, whereas in the lower and upper
Campaspe River it was 39.1 ± 4.0 NTU and 23.1 ± 4.6
NTU respectively. Turbidity levels, like conductivity,
showed an overall rise during the study period.
Dissolved oxygen concentrations were comparable
in the Broken and Campaspe Rivers during the 3-year
comparison, dropping to approximately 5 mg L)1 in
mid-summer and reaching approximately 10 mg L)1
in mid-winter (Figs 3e & 8e). As with pH levels in the
Campaspe River, those in the Broken River were
never lower than 7 and rarely greater than 8 (Figs 3f &
8f).
Comparison of the Campaspe and Broken River fish
faunas: 2000–2003
A total of three native and four alien species were
recorded from the Campaspe River and a total of five
native and four alien species were recorded from the
Broken River between August 2000 and February 2003
(b) Total fish
0
0.5
1.0
1.5
2.0
(e) Common carp
0
0.5
1.0
1.5
2.0
(d) Flathead gudgeon
0
0.5
1.0
1.5
2.0
(c) Golden perch
0
0.5
1.0
1.5
2.0
(f) European perch
0
0.5
1.0
1.5
2.0
1996 1997 1998 1999 2000 2001 2002
Lowersection
Middlesection Upper
section
Year
Abu
ndan
ce o
f fis
h
0123456
Fre
quen
cy(a) Species richness
Fig. 6 Mean + 1SE of (a) species richness and log10(x + 1)
abundance of (b) total fish, (c) golden perch, (d) flathead
gudgeon (e) common carp and (f) European perch for the lower
(black columns), middle (hatched) and upper (white) sections of
the Campaspe River between 1996 and 2002.
Regulated river fish fauna 801
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

(Table 5). All together, 657 fish, comprising 56.6%
natives, were recorded from the Campaspe River and
a total of 398 fish, comprising 85.7% natives, were
recorded from the Broken River during this time. The
dominant species by number in the Campaspe River
was the flathead gudgeon, with common carp a close
second. The Broken River fish fauna was dominated
by golden perch, with common carp and carp
gudgeon species ranked approximately equal second.
Flathead gudgeon was never collected from the
Broken River, and Murray cod, river blackfish and
crimson-spotted rainbowfish were never collected
from the Campaspe River.
Almost twice the biomass of fish was collected in
the Broken River than the Campaspe River during the
3 years of study (Table 5). By weight, the Campaspe
River fish fauna comprised 28.5% native species,
whereas the Broken River comprised 49.6% native
species. Carp comprised approximately half of the
biomass, with golden perch ranked second, in each
river. Whilst European perch ranked third in biomass
in the Campaspe, Murray cod ranked third in the
Broken River.
The contribution of native species to the abundance
of all fish was highest in the middle section, interme-
diate in the upper section and lowest in the lower
section of the Campaspe River (Table 5). This pattern
can be explained almost entirely by the relatively
large number of carp collected from the lower section
and the relatively large abundance of golden perch
collected from the upper section of this river. The
contribution of native species to the lower section of
the Broken River was also the lowest of all sections,
but was intermediate in the middle and highest in the
upper section. Native species always comprised more
than 75% of the fish fauna in sections of this river. The
increasing percentage of native species from the lower
to upper section of the Broken River was a result of
(a) Common carp
(b) European perch (c) Golden perch
0
20
40
60
80
100
120
140
160
0
20
40
60
0
20
40
60
≥125 mm
<124 mm
01020304050
01020304050
01020304050
≥101 mm
<100 mm
010203040
010203040
010203040
≥201 mm
<200 mm
Lower
Middle
Upper
1995
/96
1996
/97
1997
/98
1998
/99
1999
/00
2000
/01
2001
/02
2002
/03
1995
/96
1996
/97
1997
/98
1998
/99
1999
/00
2000
/01
2001
/02
2002
/03
1995
/96
1996
/97
1997
/98
1998
/99
1999
/00
2000
/01
2001
/02
2002
/03
Spawning season
Abu
ndan
ce o
f fis
h
Fig. 7 Proportion of young-of-year (white bars) and older (black bars) (a) common carp, (b) European perch and (c) golden perch
collected in the Campaspe River between the 1995/96 and 2002/03 spawning seasons. YOY were defined as carp <124 mm (Brumley,
1996; Villizi & Walker, 1999), European perch <100 mm (Shafi & Maitland, 1971) and golden perch <200 mm (Anderson et al., 1992).
802 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813
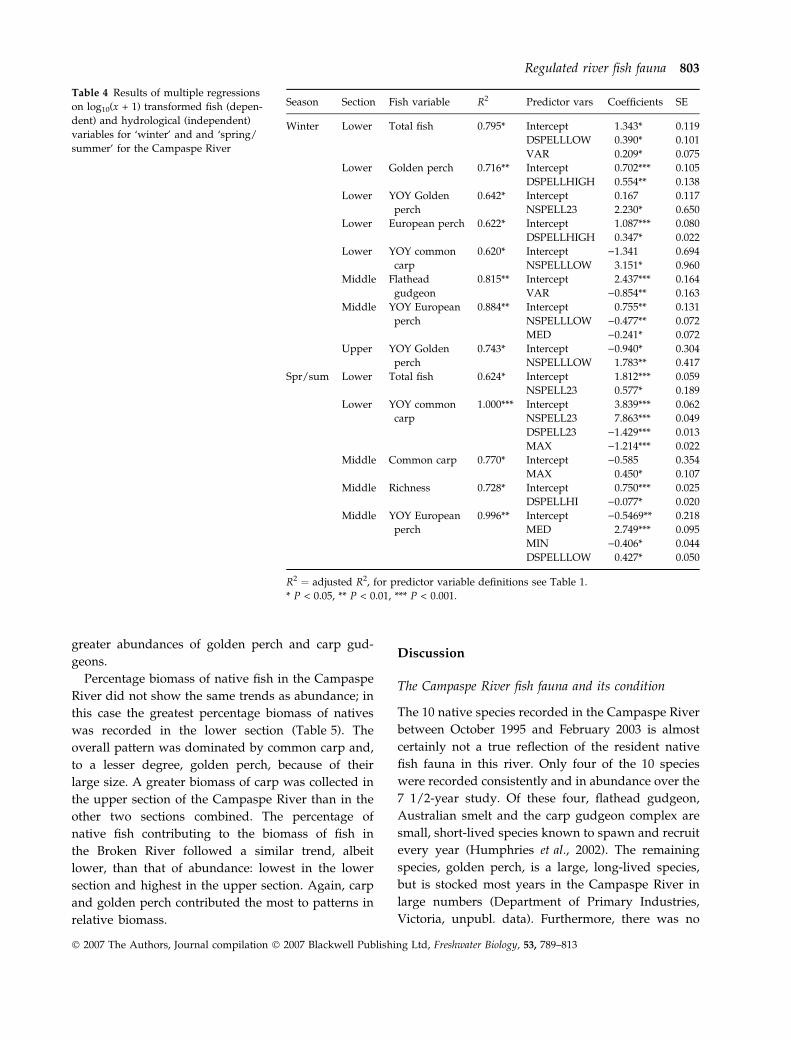
greater abundances of golden perch and carp gud-
geons.
Percentage biomass of native fish in the Campaspe
River did not show the same trends as abundance; in
this case the greatest percentage biomass of natives
was recorded in the lower section (Table 5). The
overall pattern was dominated by common carp and,
to a lesser degree, golden perch, because of their
large size. A greater biomass of carp was collected in
the upper section of the Campaspe River than in the
other two sections combined. The percentage of
native fish contributing to the biomass of fish in
the Broken River followed a similar trend, albeit
lower, than that of abundance: lowest in the lower
section and highest in the upper section. Again, carp
and golden perch contributed the most to patterns in
relative biomass.
Discussion
The Campaspe River fish fauna and its condition
The 10 native species recorded in the Campaspe River
between October 1995 and February 2003 is almost
certainly not a true reflection of the resident native
fish fauna in this river. Only four of the 10 species
were recorded consistently and in abundance over the
7 1/2-year study. Of these four, flathead gudgeon,
Australian smelt and the carp gudgeon complex are
small, short-lived species known to spawn and recruit
every year (Humphries et al., 2002). The remaining
species, golden perch, is a large, long-lived species,
but is stocked most years in the Campaspe River in
large numbers (Department of Primary Industries,
Victoria, unpubl. data). Furthermore, there was no
Table 4 Results of multiple regressions
on log10(x + 1) transformed fish (depen-
dent) and hydrological (independent)
variables for ‘winter’ and and ‘spring/
summer’ for the Campaspe River
Season Section Fish variable R2 Predictor vars Coefficients SE
Winter Lower Total fish 0.795* Intercept 1.343* 0.119
DSPELLLOW 0.390* 0.101
VAR 0.209* 0.075
Lower Golden perch 0.716** Intercept 0.702*** 0.105
DSPELLHIGH 0.554** 0.138
Lower YOY Golden
perch
0.642* Intercept 0.167 0.117
NSPELL23 2.230* 0.650
Lower European perch 0.622* Intercept 1.087*** 0.080
DSPELLHIGH 0.347* 0.022
Lower YOY common
carp
0.620* Intercept )1.341 0.694
NSPELLLOW 3.151* 0.960
Middle Flathead
gudgeon
0.815** Intercept 2.437*** 0.164
VAR )0.854** 0.163
Middle YOY European
perch
0.884** Intercept 0.755** 0.131
NSPELLLOW )0.477** 0.072
MED )0.241* 0.072
Upper YOY Golden
perch
0.743* Intercept )0.940* 0.304
NSPELLLOW 1.783** 0.417
Spr/sum Lower Total fish 0.624* Intercept 1.812*** 0.059
NSPELL23 0.577* 0.189
Lower YOY common
carp
1.000*** Intercept 3.839*** 0.062
NSPELL23 7.863*** 0.049
DSPELL23 )1.429*** 0.013
MAX )1.214*** 0.022
Middle Common carp 0.770* Intercept )0.585 0.354
MAX 0.450* 0.107
Middle Richness 0.728* Intercept 0.750*** 0.025
DSPELLHI )0.077* 0.020
Middle YOY European
perch
0.996** Intercept )0.5469** 0.218
MED 2.749*** 0.095
MIN )0.406* 0.044
DSPELLLOW 0.427* 0.050
R2 ¼ adjusted R2, for predictor variable definitions see Table 1.
* P < 0.05, ** P < 0.01, *** P < 0.001.
Regulated river fish fauna 803
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813
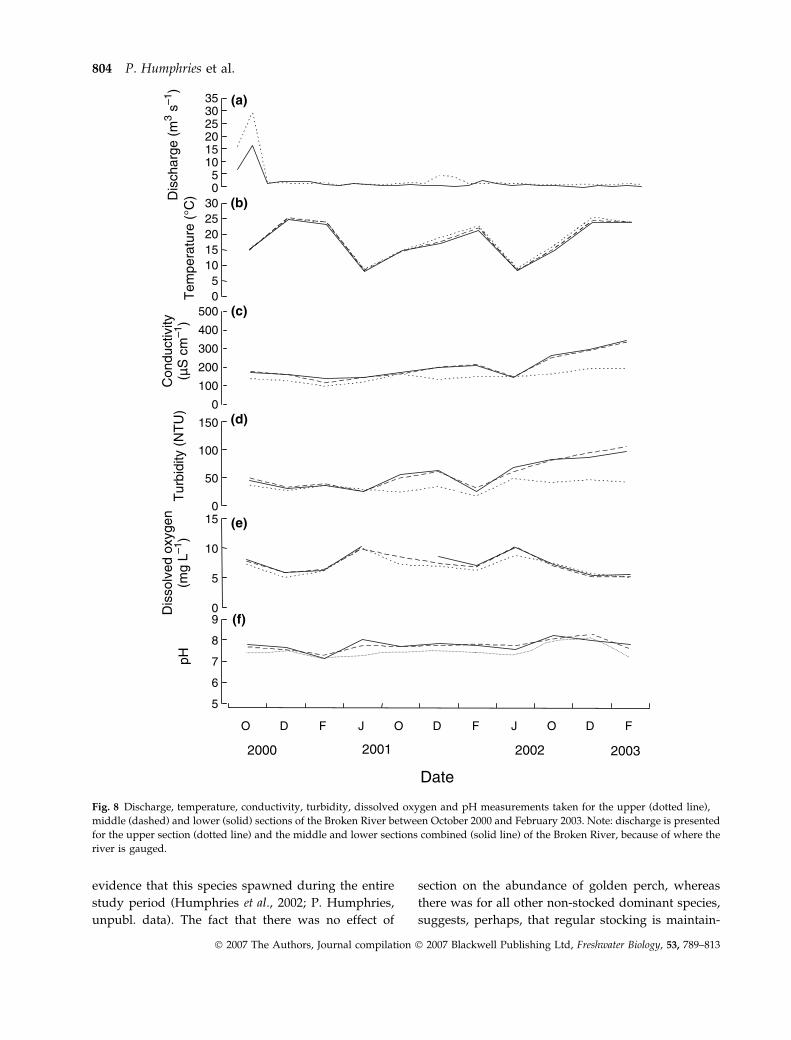
evidence that this species spawned during the entire
study period (Humphries et al., 2002; P. Humphries,
unpubl. data). The fact that there was no effect of
section on the abundance of golden perch, whereas
there was for all other non-stocked dominant species,
suggests, perhaps, that regular stocking is maintain-
5
6
7
8
9
O D F J O D F J O D F
0
50
100
150
0
100
200
300
400
50005
1015202530
2000 2001 2002 2003
(f)
(e)
(d)
(c)
(b)
(a)
pHD
isso
lved
oxy
gen
(mg
L–1)
Tur
bidi
ty (
NT
U)
Con
duct
ivity
(µS
cm
–1)
Tem
pera
ture
(°C
)Dis
char
ge (
m3 s
–1)
0
5
10
15
Date
05
101520253035
Fig. 8 Discharge, temperature, conductivity, turbidity, dissolved oxygen and pH measurements taken for the upper (dotted line),
middle (dashed) and lower (solid) sections of the Broken River between October 2000 and February 2003. Note: discharge is presented
for the upper section (dotted line) and the middle and lower sections combined (solid line) of the Broken River, because of where the
river is gauged.
804 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

Tab
le5
Per
cen
tag
eab
un
dan
cean
db
iom
ass,
tota
lab
un
dan
cean
db
iom
ass,%
nat
ives
,sp
ecie
sri
chn
ess
tota
lan
d%
spec
ies
rich
nes
so
fn
ativ
eso
ffi
shco
llec
ted
inth
eC
amp
asp
e
and
Bro
ken
Riv
ers
bet
wee
nJu
ne
2000
and
Feb
ruar
y20
03
Cam
pas
pe
Bro
ken
Lo
wer
Mid
dle
Up
per
To
tal
Lo
wer
Mid
dle
Up
per
To
tal
No
.
Bio
mas
s
(kg
)N
o.
Bio
mas
s
(kg
)
No
.B
iom
ass
(kg
)N
o.
Bio
mas
s
(kg
)
No
.B
iom
ass
(kg
)N
o.
Bio
mas
s
(kg
)N
o.
Bio
mas
s
(kg
)N
o.
Bio
mas
s
(kg
)
Nat
ive
Fla
thea
dg
ud
geo
n27
.40.
874
.31.
758
.90.
448
.60.
8–
––
––
––
–
Go
lden
per
ch3.
633
.56.
718
.712
.627
.96.
827
.850
.521
.756
.544
.945
.150
.049
.537
.8
Car
pg
ud
geo
ns
1.3
<0.
12.
2<
0.1
––
1.2
0.0
12.6
<0.
0122
.2<
0.01
41.0
0.1
9.0
0.1
Mu
rray
cod
––
––
––
–10
.523
.51.
92.
61.
06.
83.
011
.7
Riv
erb
lack
fish
––
––
––
––
––
–1.
00.
10.
5<
0.1
Cri
mso
n-s
po
tted
rain
bo
wfi
sh
––
––
––
–2.
1<
0.01
3.7
<0.
013.
1<
0.01
1.5
<0.
1
Ali
en Co
mm
on
carp
55.8
49.3
8.9
63.7
11.4
61.6
31.2
58.5
24.2
54.8
12.0
52.2
6.7
42.8
12.3
50.3
Eu
rop
ean
per
ch8.
66.
83.
913
.012
.67.
68.
48.
4–
––
–2.
10.
11.
0<
0.1
Go
ldfi
sh3.
09.
61.
72.
84.
62.
53.
04.
6–
–1.
90.
3–
–0.
50.
1
Eas
tern
gam
bu
sia
0.3
<0.
12.
2<
0.1
––
0.8
<0.
1–
–1.
9<
0.01
––
0.5
<0.
1
To
tal
303
3085
317
920
407
175
5642
065
710
7680
9573
529
108
6218
319
561
009
398
1967
21
Nat
ives
(%)
32.3
34.3
83.2
20.4
71.4
28.3
56.6
28.5
75.8
45.2
84.3
47.6
91.3
57.1
85.7
49.6
Sp
ecie
sri
chn
ess
tota
l7
75
75
77
7
Sp
ecie
sri
chn
ess
nat
ives
(%)
42.9
42.9
40.0
42.9
80.0
57.1
71.4
55.6
Regulated river fish fauna 805
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

ing this species throughout the river as a whole.
Considering that approximately 18–20 of the 34
Murray-Darling Basin native fish species probably
once inhabited this river (Humphries & Lake, 2000),
the extant native fish fauna of the Campaspe River is
thus extremely poor. Historically, this river was
known for its abundant Macquarie perch and Murray
cod populations (Anonymous, 1973; Langtry in
Cadwallader, 1977), but it is now virtually unheard-of
for the former, and uncommon for the latter, to be
caught by anglers.
A total of six alien species was recorded during the
study, and those collected consistently are also the
most common alien species encountered in south-
eastern Australia (Cadwallader, 1978; Arthington &
Mitchell, 1986; Koehn, 2002, 2004; Kennard et al.,
2005). Common carp and European perch comprised
approximately 64% of the biomass and 36% of the
abundance of fish collected overall; a figure compa-
rable to studies elsewhere in Australia where these
species occur (Gehrke et al., 1995).
Our results indicated that there were substantial
differences in the fish fauna of each of the hydro-
logically-distinct sections of the Campaspe River and
broadly confirm our predictions related to the degree
of flow alteration and richness and dominance of
native and alien species. Thus, species richness of
natives was greatest in the lower section, lowest in
the upper section and intermediate in the middle
section. These differences were a result of the
presence of carp gudgeons, Murray cod, Australian
smelt, silver perch and flatheaded galaxias in the
lower and middle section, but not in the upper
section. Parallel work on larval fish, using methods
more suitable for capturing Australian smelt, indi-
cated that this pelagic, shoaling species was as
abundant in the upper section of the river as it was
in the other two sections (Humphries et al., 2002). On
the other hand, from the results presented here and
from observations over many years, we believe that
the absence of the other species from samples in the
upper Campaspe is a true reflection of their absence
or rarity in that section (Humphries et al., 2002;
P. Humphries, unpubl. data). Alien species tended to
be more extensive in their spatial distribution in the
Campaspe River than did native species. Common
carp, European perch and goldfish occurred in each
section, albeit in decreasing abundance with increas-
ing distance upstream.
Fish fauna and hydrology
There are few studies in Australia or the rest of the
world where changes to the fish fauna of a river can
be attributed directly to flow alteration (Lloyd et al.,
2003; Lytle & Poff, 2003). The main reason for this is
because of the impracticality of conducting rigorously
designed – controlled and replicated – experiments in
anything but the smallest systems. Nevertheless, we
can gain substantial insight into the influence of flow
by comparisons across broad geographic areas in
streams which differ in their flow regimes and
associated characteristics (Lamouroux & Cattaneo,
2006). Similarly, in our study we have been able to
compare characteristics of the fish fauna of the
Campaspe River among three river sections that
experience contrasting hydrologies. We have also
been able to describe relationships between fish-
related variables and hydrological variables and to
contrast the Campaspe River and the similar, but less
regulated Broken River. An important point to note,
however, is that much of the study reported here
occurred during an extended dry period and that the
patterns observed were without doubt a function of
both natural climatic and anthropogenic influences.
Our analysis of the fish fauna during the 7 1/2 -year
study indicated that the lower section of the Campa-
spe River was consistently the most species rich; again
confirming our prediction that the greater degree of
regulation the fewer native species occurring, since
this section was the least regulated of the three. When
access to the Campaspe River from the Murray River
is possible, this means that this section will potentially
be more accessible than at least the upper section.
These patterns in species richness by section were,
however, not reflected in relationships between
threshold discharge levels for upstream movement
over weirs and species richness in the lower section,
although species richness was positively related to
high flow variables in the middle section of the river.
Thus, our prediction that fish-related variables, such
as richness, would respond to high or threshold flows
was not confirmed. The general lack of significant
relationships between hydrology and richness is not
entirely surprising, given the rarity of the upstream
connection between the Murray River and the lower
Campaspe and between the lower and middle Cam-
paspe (about a month in total in 1996 and for 3–4 days
in 2000) and between the middle and upper sections
806 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

(2–3 days in 1996) and because of the relatively small
pool of species in the river.
Irrespective of the potential for movement between
river sections, greater species richness in the lower
Campaspe River is consistent with the widely held
view of a positive relationship between species rich-
ness and stream order (see Matthews, 1986). How-
ever, what fish are responding to is the subject of
ongoing debate; it may relate to diversity of habitat
(Gorman & Karr, 1978), rates of immigration and
emigration (Power et al., 1988) or drainage basin size
or stream length, width or gradient (Lake, 1982;
Grenouillet, Pont & Herisse, 2004). In contrast to the
Campaspe River, despite the presence of two sub-
stantial weirs, there was no decline in number of
species evident with increasing distance upstream in
the Broken River. Indeed, our prediction that there
would be more native species, and that they would
comprise a larger proportion of the fish fauna in the
Broken than in the Campaspe River, was confirmed. It
is thus unlikely that the decline in species with
distance upstream from the Murray River confluence
can be explained solely by the presence of weirs in the
Campaspe River. We have little doubt, however, that
there will be some species, especially those thought to
move considerable distance as part of their normal life
history (Reynolds, 1983; Koehn, 1997; Koehn & Nicol,
1998; Koehn, Stuart & Crook, 2003), which will not be
in abundance above barriers without regular stocking.
The high degree of regulation in the upper section
of the Campaspe River has been proposed by
Humphries et al. (2002) as having a deleterious effect
on the fish fauna of this system. Differences in
physico-chemical variables between the Campaspe
and Broken Rivers, mainly turbidity and conductivity,
and between the upper and the middle and lower
sections of the Campaspe River, seem unable to
explain the differences observed. In addition, temper-
ature depression, as a result of low-level releases from
Lake Eppalock, is relatively minor and inconsistent,
compared with other systems, like the Goulburn and
Murray Rivers (McMahon & Finlayson, 1995), and is
also unlikely to explain the observed differences. As
circumstantial evidence for the impacts of flow alter-
ation, Humphries et al. (2002) cited the absence of carp
gudgeons, a species considered vulnerable to river
regulation (Gehrke, 1997), in the upper section of the
Campaspe River, the dominance of small, opportu-
nistic, short-lived native species which breed over a
protracted period and the success of generalist alien
species. These authors proposed that the enhanced
flows over the spring/summer period in the upper
section of the Campaspe did not prevent spawning for
most species but reduced the area of slackwater
rearing habitats. Recent experiments in a lowland
river, in which slackwaters were ‘destroyed’, through
diverting current into existing slackwater patches,
showed that simulating the effects of irrigation flows
displaced larvae from slackwaters (Humphries et al.,
2006). River regulation has also been shown generally
to affect recruitment in other rivers systems in the
world (Robinson, Clarkson & Forrest, 1998; Liebig
et al., 1999; Merigoux & Ponton, 1999; Schiemer et al.,
2001).
The Campaspe River’s fish fauna is, however,
depauperate throughout its length downstream of
Lake Eppalock; not just in the upper section, where
most of the irrigation releases occur. It is logical,
especially for highly mobile organisms such as fish,
and for rivers that are longitudinally connected, that
the effects of flow alteration on fish faunas will extend
beyond the immediate area that is heavily regulated.
Some clues to how flow affects this river’s fish fauna
may, perhaps, be gained from sporadic recordings of
species during our study and how they relate to flow
conditions. The only major high flow event which
occurred during the 7 1/2-year study was in the
winter/spring of 1996, although a brief period of
relatively high flow also occurred in late 2000. For
several weeks between August and October of 1996,
Lake Eppalock spilled, the river was very high and
filled billabongs and anabranches and broke its banks
in places, and this was the same time during which
we estimate that threshold levels of discharge were
reached sufficient for fish to move into both the lower
and middle sections of the river and, possibly, for 2–
3 days into the upper from the middle section.
Downstream movement, even from above Lake Ep-
palock, was possible during these high flow events.
During that time, or in the months following the high
flow period, several species were collected that had
not been recorded prior to this, which were mostly not
recorded for the rest of the study and which
responded to high flow events by moving consider-
able distances upstream or downstream. Therefore,
there is qualitative evidence that our prediction that
fish will respond to high or threshold flows through
movement over weirs may have some validity. Under
Regulated river fish fauna 807
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

unregulated conditions, only small rises in flow
would have allowed unimpeded movement through-
out much of the length of the Campaspe River. Since
Lake Eppalock was built, however, movement be-
tween river sections would be limited predominantly
to when the storage filled and spilled; prior to 1996,
approximately every 2 years, but since that time,
never.
Despite the likely effect of high flow events on fish
movement, there was no quantitative relationship
between the overall composition of the Campaspe
River fish fauna and any hydrological variables,
which is contrary to our predictions. This was some-
what unexpected, considering the large inter-annual
differences in discharge during the study. Thus, the
composition was stable over a period, which in-
cluded, as stated above, a high flow year, a moderate
flow year and 5 years when flows were very low.
However, again, there is a relatively small suite of
species which is driving the overall composition, and
these are species – both native and alien – are known
to have broad habitat and water quality tolerances
and life histories which allow them to breed and
recruit under a range of conditions (Humphries et al.,
2002). There has been much effort expended on the
question of how stable fish assemblages are and the
mechanisms behind their variability (see e.g.
Grossman, 1982; Matthews, Cashner & Gelwick,
1988; Schlosser & Ebel, 1989; Grossman, Dowd &
Crawford, 1990; Poff & Allan, 1995; Grossman et al.,
1998). There seems to be general agreement, however,
that flow variability - high flows in general (and short
spates in particular) – affect fish assemblage variabil-
ity (Oberdorff et al., 2001; Schiemer et al., 2001; Koel &
Sparks, 2002; Li & Gelwick, 2005), although this
agreement is not universal (see e.g. Moyle & Vond-
racek, 1985). It appears that the nature of the differ-
ences among the three river sections of the Campaspe
and the nature of the species which remain in this
degraded system outweigh any among-year differ-
ences in their influence on characteristics of the fish
fauna.
When the abundances of individual species and life
stages were examined, however, several were signif-
icantly related to hydrological variables. For example,
the abundance of golden perch in winter in the lower
section was positively related to the duration of 10th
percentile flows and above, suggesting the potential
for movement upstream from the Murray River in
response to high flows. Total fish abundance in the
lower section in winter, on the other hand, was
positively related to the duration of 90th percentile
flows and a measure of variability in flow. The reason
for this is uncertain. Indeed, it is hard to come to
general conclusions regarding fish/hydrology rela-
tionships for the fish fauna, the river and hydrological
seasons. This could be due to the fact that the duration
of the study was still too short for robust investigation
of relationships, the fact that each section of river has
a distinct hydrology and therefore is probably oper-
ating differently and so we would expect relation-
ships between hydrology and fish to be different
among river sections, or that these types of rivers have
high inter-annual variability in flow and antecedent
conditions, perhaps extending for many years, play a
role in the abundances that we see in any one year.
An example, perhaps, of the complexities of fish/
hydrology relationships is of the recruitment of
common carp. Abundance of YOY carp showed two
peaks: a minor one in 1996/97, during a year of high
rainfall, high flows and extended periods of connec-
tion for the river system below Lake Eppalock and a
major one in 2000/01, when rainfall and flows were
much higher than in the last 3 years, but considerably
lower than in 1996. This resulted in the apparently
contradictory positive relationship between YOY carp
abundance and the number of spells of 90th percentile
flows in the lower section of the river. But this is
explainable by the fact that it was the 1996/97 and
2000/01 years which influenced the significance of the
relationship, and that in these two recruitment years,
YOY carp abundance was lowest during the higher of
the two flows and highest during the lower of the two.
Strong carp recruitment was also reported in major
floodplain wetlands of the Murray River in the 2000/
01 breeding season, in response to an environmental
water release (Brown et al., 2005; Crook & Gillanders,
2006). Indeed, this species seems to respond to
inundation of previously dry habitat by spawning in
many locations in the Murray-Darling Basin (King,
Humphries & Lake, 2003; Smith & Walker, 2004;
Driver et al., 2005).
In conclusion, we have found evidence that the
differences that exist in the fish fauna of the three
sections of the Campaspe River are likely to be due to
differences in hydrology and that the fish fauna of
this river as a whole is in a poorer state than the less
regulated Broken River. This adds weight to the call
808 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

for more sensitive management of rivers, especially
for rivers which are heavily regulated and which
have for decades been managed solely as irrigation
conduits. Providing periodic moderate-to-high flows
and threshold flows over barriers or – better still –
allowing fish passage through fishways or removal of
weirs altogether will mean that fish will be able to
move into and out or areas for life history purposes,
for the acquisition of food, to recolonize river sections
and to escape potentially harmful conditions, such as
blackwater events. Despite some progress in under-
standing, our 7 1/2-year study is apparently still of
insufficient duration for proper investigation of fish/
hydrology relationships. It is sobering to realize that
for this to be accomplished, riverine fish studies of
20+ years are probably needed. These are rare in the
world (although see e.g. Daufresne et al., 2003) and
entirely absent in Australia. With the likelihood of
reduced streamflows in south-eastern Australia and
increased temperatures associated with climate
change, it is all the more imperative that long-term
studies are initiated now in order to understand the
link between fish and flow and to make predictions
for the future.
Acknowledgments
We would like to thank the researchers who assisted
in various aspects of the field and laboratory work
associated with this study; especially David Crook,
Alison King, Jane Growns, Simon Lukies, Zeb Tonkin
and Helen Gigney. Rob Cook and Adam Richardson
were a constant source of critical comment and bad
jokes and Nicole McCasker provided helpful com-
ments on the manuscript. Sam Lake, Gerry Quinn,
Terry Hillman, Ben Gawne, Peter E. Davies, Angela
Arthington and John Harris provided support and
guidance, intellectually and administratively. Peter
Cullen and Barry Hart were, amongst others, instru-
mental in helping to get the project going and to keep
it going beyond the normal 3 years. Many landowners
on the Campaspe and Broken Rivers gave up their
valuable time to provide information and advice, as
well as much appreciated hospitality. We would
especially like to thank the Danahers and the Keats
for their consistent congeniality, tolerance and story-
telling. This study was funded jointly by the Cooper-
ative Research Centre for Freshwater Ecology, the
Commonwealth Department of Environment and
Heritage (through Environment Australia and Land
and Water Australia) and Goulburn-Murray Water.
Thiess Environmental Services kindly supplied
hydrological data and the Goulburn-Broken Catch-
ment Management Authority assisted in many ways.
References
Aarts B.G.W., Van den Brink F.W.B. & Nienhuis P.H.
(2004) Habitat loss as the main cause of slow recovery
of fish faunas of regulated large rivers in Europe: the
transversal floodplain gradient. River Research and
Applications, 20, 3–23.
Anderson J.R., Morison A.K. & Ray D.J. (1992) Validation
of the use of thin-sectioned otoliths for determining the
age and growth of golden perch, Macquaria ambigua
(Perciformes: Percichthyidae), in the lower Murray-
Darling Basin, Australia. Australian Journal of Marine
and Freshwater Research, 43, 1103–1128.
Anonymous (1973) Native fish in the Campaspe and
Coliban Rivers, Freshwater Fisheries Newsletter. Fisheries
and Wildlife, Vic., Melbourne, Australia, 5, 18–19.
Arthington A.H. & Mitchell D.S. (1986) In: Aquatic
Invading Species, Ecology of Biological Invasions: An
Australian Perspective (Eds R.H. Groves & J.J. Burdon),
pp. 34–52. Australian Academy of Science, Canberra.
Bertozzi T., Adams M. & Walker K.F. (2000) Species
boundaries in carp gudgeons (Eleotrididae: Hypseleo-
tris) from the River Murray, South Australia: evidence
for multiple species and extensive hybridization.
Marine & Freshwater Research, 51, 805–815.
Brown P., Sivakumaran K.P., Stoessel D. & Giles A.
(2005) Population biology of carp (Cyprinus carpio L.) in
the mid-Murray River and Barmah Forest Wetlands,
Australia. Marine & Freshwater Research, 56, 1151–
1164.
Brumley A.R. (1996) Family Cyprinidae: carps, minnows
etc. In: Freshwater Fishes of South-Eastern Australia (Ed.
R. McDowall), pp. 99–106. Reed, Sydney, NSW.
Cadwallader P.L. (1977) J.O. Langtry’s 1949–50 Murray
River investigations. Fisheries and Wildlife, paper 13,
Fisheries and Wildlife, Melbourne, Vic. 70 pp.
Cadwallader P.L. (1978) Some causes of the decline in the
range and abundance of native fish in the Murray-
Darling river system. Proceedings of the Royal Society of
Victoria, 90, 211–224.
Calow P. & Petts G.E. (1992) The Rivers Handbook. Vol. 1
and 2. Blackwell Scientific Publications, Oxford.
Cattaneo F., Lamouroux N., Breil P. & Capra H. (2002) The
influence of hydrological and biotic processes on brown
trout (Salmo trutta) populations dynamics. Canadian
Journal of Fisheries and Aquatic Sciences, 59, 12–22.
Regulated river fish fauna 809
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

Clark K.R. & Warwick R.M. (2001) PRIMER 5. Plymouth
Marine Laboratory, Plymouth.
Crook D.A. & Gillanders B.M. (2006) Use of otolith
chemical signatures to estimate carp recruitment
sources in the mid-Murray River, Australia. River
Research and Applications, 22, 871–879.
Daufresne M., Roger M.C., Capra H. & Lamouroux N.
(2003) Long-term changes within the invertebrate and
fish communities of the Upper Rhone River : effects of
climatic factors. Global Change Biology, 10, 124–140.
Driver P.D., Harris J.H., Closs G.P. & Koen T.B. (2005)
Effects of flow regulation on carp (Cyprinus carpio L.)
recruitment in the Murray-Darling basin, Australia.
River Research and Applications, 21, 327–335.
Fausch K.D., Torgersen C.E., Baxter C.V. & Li H.W.
(2002) Landscapes to riverscapes: bridging the gap
between research and conservation of stream fishes.
Bioscience, 52, 483–498.
Frissell C.A., Liss W.J., Warren C.E. & Hurley M.D.
(1986) A hierarchical framework for stream habitat
classification. Environmental Management, 10, 199–214.
Gehrke P.C. (1997) Differences in composition and
structure of fish communities associated with flow
regulation in New South Wales rivers. In: Fish and
Rivers in Stress: the NSW Rivers Survey (Eds J.H. Harris
& P.C. Gehrke), pp. 169–200. NSW Fisheries Office of
Conservation and Cooperative Research Centre for
Freshwater Ecology, Canberra.
Gehrke P.C., Brown P., Schiller C.B., Moffatt D.B. &
Bruce A.M. (1995) River regulation and fish commu-
nities in the Murray-Darling River system, Australia.
Regulated Rivers: Research and Application, 11, 363–375.
Gorman O.T. & Karr J.R. (1978) Habitat structure and
stream fish communities. Ecology, 59, 507–515.
Grenouillet G., Pont D. & Herisse C. (2004) Within-basin
fish assemblage structure: the relative influence of
habitat versus stream spatial position on local species
richness. Canadian Journal of Fisheries and Aquatic
Sciences, 61, 93–102.
Grossman G.D. (1982) Dynamics and organisation of a
rocky intertidal fish assemblage: the persistence and
resilience of taxocene structure. American Naturalist,
119, 611–637.
Grossman G.D., Dowd J.F. & Crawford M. (1990)
Assemblage stability in stream fishes: a review. Envi-
ronmental Management, 5, 661–671.
Grossman G.D., Ratajczak R.E., Crawford M. & Freeman
M.C. (1998) Assemblage organisation in stream fishes:
effects of environmental variation and interspecific
interactions. Ecological Monographs, 68, 395–420.
Humphries P. & Lake P.S. (2000) Fish larvae and the
management of regulated rivers. Regulated Rivers:
Research and Management, 16, 421–432.
Humphries P., Serafini L.G. & King A.J. (2002) River
regulation and fish larvae: changes in space and time.
Freshwater Biology, 47, 1307–1330.
Humphries P., Cook R.A., Richardson A.J. & Serafini lG.
(2006) Creating a disturbance: manipulating slackwa-
ters in lowland rivers. River Research and Applications,
22, 525–542.
Junk W.J., Bayley P.B. & Sparks R.E. (1989) The flood
pulse concept in river-floodplain systems. In: Proceed-
ings of the International Large River Symposium (Ed. D.P.
Dodge), Canadian Special Publication Fisheries Aquatic
Sciences, 106, 110–127.
Kennard M.J., Arthington A.H., Pusey B.J. & Harch B.D.
(2005) Are alien fish a reliable indicator of river health?
Freshwater Biology, 50, 174–193.
King A.J., Humphries P. & Lake P.S. (2003) Fish recruit-
ment on flood plains: the roles of patterns of flooding
and life history characteristics. Canadian Journal of
Fisheries and Aquatic Science, 69, 773–786.
Koehn J.D. (1997) Habitats and movements of freshwater
fish in the Murray-Darling Basin. In: Proceedings of the
1995 Riverine Environment Forum (Eds R.J. Banens & R.
Lehane), pp. 27–32. Murray-Darling Basin Commis-
sion, Canberra.
Koehn J.D. (2002) Freshwater invaders in Victoria. Under
Control – Pest Plant and Animal News, 22, 12–14.
Koehn J.D. (2004) Carp (Cyprinus carpio) as a powerful
invader in Australian waterways. Freshwater Biology,
49, 882–894.
Koehn J. & Nicol S. (1998) Habitats and movement
requirements of fish. In: Proceedings of the 1996 Riverine
Environment Forum (eds. R.J. Banens & R. Lehane), pp.
1–6. Murray-Darling Basin Commission, Canberra.
Koehn J.D., Stuart I. & Crook D. (2003) Linking the
ecological importance of downstream fish movements
to management of Murray-Darling Basin fish popu-
lations. In: Downstream Movement of Fish in the
Murray-Darling Basin (Eds M. Lintermans & R. Phil-
lips), pp. 67–79. Murray-Darling Basin Commission,
Canberra.
Koel T.M. & Sparks R.E. (2002) Historical patterns of
river stage and fish communities as criteria for oper-
ations of dams on the Illinois River. River Research and
Applications, 18, 3–19.
Lake P.S. (1982) The relationship between freshwater fish
distribution, stream drainage area and stream length in
some streams of south-east Australia. Bulletin of the
Australian Society of Limnology, 8, 31–37.
Lake P.S. (2003) Ecological effects of perturbation by
drought in flowing waters. Freshwater Biology, 48, 1161–
1172.
Lamouroux N. & Cattaneo F. (2006) Fish assemblages
and stream hydraulics: consistent relations across
810 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

spatial scales and regions. River Research and Applica-
tions, 22, 727–737.
Li R.Y. & Gelwick F.P. (2005) The relationship of
environmental factors to spatial and temporal varia-
tion of fish assemblages in a flooplain river in Texas,
USA. Ecology of Freshwater Fish, 14, 319–330.
Liebig H., Cereghino R., Lim P., Belaud A. & Lek S.
(1999) Impact of hydropeaking on the abundance of
juvenile brown trout in a Pyrenean stream. Archive fur
Hydrobiologie, 144, 439–454.
Lloyd N., Quinn G., Thoms M., Arthington A., Gawne B.,
Humphries P. & Walker K. (2003) Does Flow Modifica-
tion Cause Geomorphological and Ecological Responses in
rivers? A Literature Review from an Australian Perspective.
Technical report 1/2004, CRC for Freshwater Ecology,
Canberra.
Lucas M.C. & Baras E. (2001) Migrations of Freshwater
Fishes. Blackwell Science, Oxford, 420 pp.
Lytle D.A. & Poff N.L. (2003) Adaptation to natural flow
regimes. Trends in Ecology and Evolution, 19, 94–100.
Marsh N.A., Stewardson M.J. & Kennard M.J. (2003)
River Analysis Package. Cooperative Research Centre for
Catchment Hydrology, Monash University, Mel-
bourne, Vic..
Matthews W.J. (1986) Fish faunal ‘‘breaks’’ and stream
order in the eastern and central United States. Envi-
ronmental Biology of Fishes, 17, 81–92.
Matthews W.J. (1998) Patterns in Freshwater Fish Ecology.
Kluwer, Boston, 784 pp.
Matthews W.J., Cashner R.C. & Gelwick F.P. (1988)
Stability and persistence of fish faunas and assem-
blages in three Midwestern streams. Copeia, 1988, 945–
955.
McMahon T.A. & Finlayson B.L. (1995) Reservoir system
management and environmental flows. Lakes and
Reservoirs: Research and Management, 1, 65–76.
Merigoux S. & Ponton D. (1999) Spatio-temporal distri-
bution of young fish in tributaries of natural and flow-
regulated sections of a neotropical river in French
Guiana. Freshwater Biology, 42, 177–198.
Moyle P.B. & Vondracek B. (1985) Persistence and
structure of the fish assemblage in a small California
stream. Ecology, 66, 1–13.
Murray-Darling Basin Commission (2004) Native Fish
Strategy for the Murray-Darling Basin. MDBC, Canberra,
64 pp.
Oberdorff T., Hugueny B. & Vigneron T. (2001) Is
assemblage variability related to environmental vari-
ability? An answer for riverine fish. Oikos, 93, 419–
428.
Petts G.E. (1984) Impounded Rivers: Perspectives for Ecolog-
ical Management. John Wiley & Sons, Chichester, 326
pp.
Poff N.L. & Allan J.D. (1995) Functional organisation of
stream fish assemblages in relation to hydrological
variability. Ecology, 76, 606–627.
Poff N.L., Allan J.D., Bain M.B., Karr J.R., Prestegaard
K.L., Richter B.D., Sparks R.E. & Stromberg J.C. (1997)
The natural flow regime: a paradigm for river flow
conservation and restoration. Bioscience, 47, 769–784.
Power M.E., Stout R.J., Cushing C.E., Harper P.P., Hauer
F.R., Matthews W.J., Moyle P.B., Statzner B. & Wais de
Badgen I.R. (1988) Biotic and abiotic controls in river
and stream communities. Journal of the North American
Benthological Society, 7, 456–479.
Reynolds L.F. (1983) Migration patterns of five fish
species in the Murray-Darling River System. Austra-
lian Journal of Marine and Freshwater Research, 24, 857–
871.
Richter B.D., Mathews R., Harrison D.L. & Wigington R.
(2003) Ecologically sustainable water management:
managing river flows for ecological integrity. Ecological
Applications, 13, 206–224.
Robinson A.T., Clarkson R.W. & Forrest R.E. (1998)
Dispersal of larval fishes in a regulated river tributary.
Transactions of the American Fisheries Society, 127, 772–
786.
Schiemer F., Keckeis H., Winkler G. & Flore L. (2001)
Large rivers: the relevance of ecotonal structure and
hydrological properties for the fish fauna. Archive fur
Hydrobiologie Supplement, 135, 487–508.
Schlosser I.J. & Ebel K.K. (1989) Effects of flow regime
and cyprinid predation on a headwater stream.
Ecological Monographs, 59, 41–57.
Shafi M. & Maitland P.S. (1971) The age and growth of
perch (Perca fluviatilis L.) in two Scottish lakes. Journal
of Fish Biology, 3, 39–57.
Smith G. & Humphries P. (1997) Environmental flows
research on the Campaspe River in north east Victoria.
In: Proceedings of the Australian National Conference on
Large Dams, pp. 41–46. Australian National Committee
on Large Dams Inc., Tatura, Victoria, Australia.
Smith B.B. & Walker K.F. (2004) Reproduction of com-
mon carp in South Australia, shown by young-of-the-
year samples, gonadosomatic index and the histolog-
ical staging of ovaries. Transactions of the Royal Society
of South Australia, 128, 249–257.
Thorp J.H. & Delong M.D. (1994) The riverine productivity
model: an heuristic view of carbon sources and organic
processing in large-river ecosystems. Oikos, 70, 305–308.
Vannote R.L., Minshall G., Cummins K.W., Sedell J.R. &
Cushing C.E. (1980) The river continuum concept.
Canadian Journal of Fisheries and Aquatic Sciences, 37,
130–137.
Villizi L. & Walker K.F. (1999) Age and growth of the
common carp Cyprinus carpio, in the River Murray,
Regulated river fish fauna 811
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813

Australia: validation, consistency of age interpretation,
and growth models. Environmental Biology of Fishes, 54,
77–106.
Walker K.F. (1986) The Murray-Darling river system. In:
The Ecology of River Systems (Eds B.R. Davies & K.F.
Walker), pp. 631–659. Dr W. Junk, Dordrecht.
Walker K.F. (1992) The River Murray, Australia: a
semiarid lowland river. In: The Rivers Handbook. Vol.
1. (Eds P. Calow & G.E. Petts), pp. 472–492. Blackwell
Scientific Publications, Oxford.
Ward J.V. & Stanford J.A. (1979) Ecological factors
controlling stream zoobenthos and emphasis on ther-
mal modification in regulated streams. In: The Ecology
of Regulated Streams (Eds J.V. Ward & J.A. Stanford),
pp. 33–55. Plenum Press, New York.
Welcomme R.L. (1985) River Fisheries. Food and Agricul-
ture Organization of the United Nations, Rome, FAO
Fisheries Technical Paper No. 262. pp.
(Manuscript accepted 3 October 2007)
812 P. Humphries et al.
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813
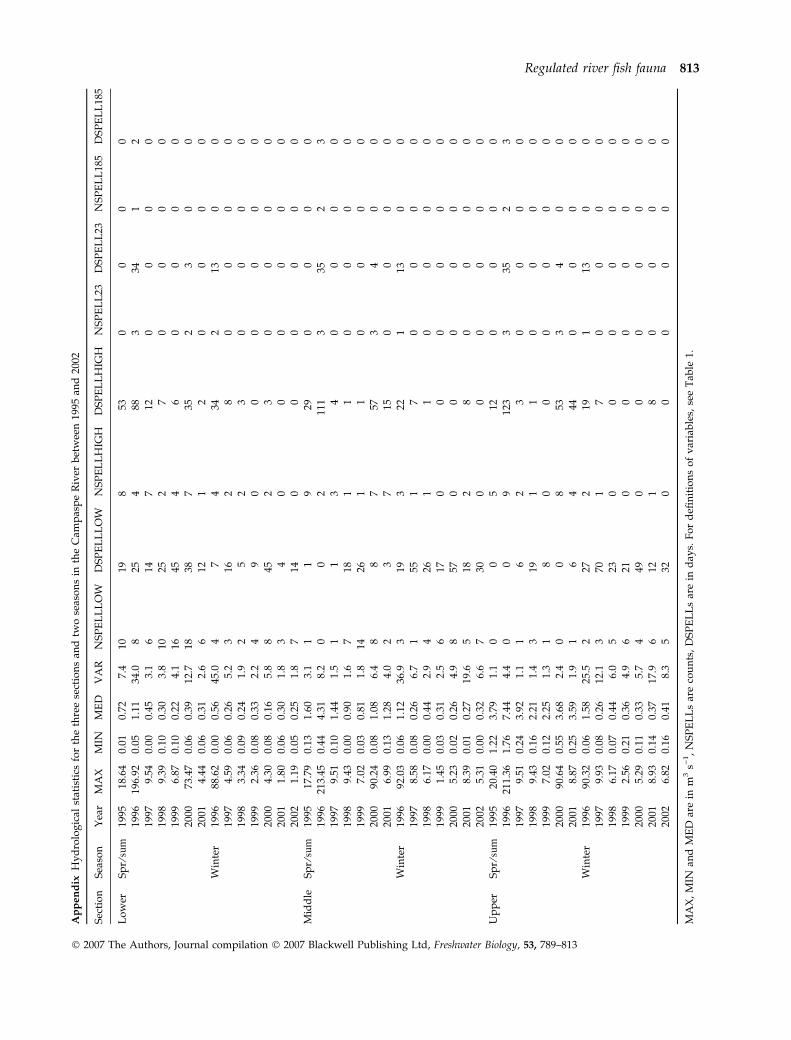
Ap
pe
nd
ixH
yd
rolo
gic
alst
atis
tics
for
the
thre
ese
ctio
ns
and
two
seas
on
sin
the
Cam
pas
pe
Riv
erb
etw
een
1995
and
2002
Sec
tio
nS
easo
nY
ear
MA
XM
INM
ED
VA
RN
SP
EL
LL
OW
DS
PE
LL
LO
WN
SP
EL
LH
IGH
DS
PE
LL
HIG
HN
SP
EL
L23
DS
PE
LL
23N
SP
EL
L18
5D
SP
EL
L18
5
Lo
wer
Sp
r⁄sum
1995
18.6
40.
010.
727.
410
198
530
00
019
9619
6.92
0.05
1.11
34.0
825
488
334
12
1997
9.54
0.00
0.45
3.1
614
712
00
00
1998
9.39
0.10
0.30
3.8
1025
27
00
00
1999
6.87
0.10
0.22
4.1
1645
46
00
00
2000
73.4
70.
060.
3912
.718
387
352
30
020
014.
440.
060.
312.
66
121
20
00
0W
inte
r19
9688
.62
0.00
0.56
45.0
47
434
213
00
1997
4.59
0.06
0.26
5.2
316
28
00
00
1998
3.34
0.09
0.24
1.9
25
23
00
00
1999
2.36
0.08
0.33
2.2
49
00
00
00
2000
4.30
0.08
0.16
5.8
845
23
00
00
2001
1.80
0.06
0.30
1.8
34
00
00
00
2002
1.19
0.05
0.25
1.8
714
00
00
00
Mid
dle
Sp
r⁄sum
1995
17.7
90.
131.
603.
11
19
290
00
019
9621
3.45
0.44
4.31
8.2
00
211
13
352
319
979.
510.
101.
441.
51
13
40
00
019
989.
430.
000.
901.
67
181
10
00
019
997.
020.
030.
811.
814
261
10
00
020
0090
.24
0.08
1.08
6.4
88
757
34
00
2001
6.99
0.13
1.28
4.0
23
715
00
00
Win
ter
1996
92.0
30.
061.
1236
.93
193
221
130
019
978.
580.
080.
266.
71
551
70
00
019
986.
170.
000.
442.
94
261
10
00
019
991.
450.
030.
312.
56
170
00
00
020
005.
230.
020.
264.
98
570
00
00
020
018.
390.
010.
2719
.65
182
80
00
020
025.
310.
000.
326.
67
300
00
00
0U
pp
erS
pr
⁄sum
1995
20.4
01.
223.
791.
10
05
120
00
019
9621
1.36
1.76
7.44
4.4
00
912
33
352
319
979.
510.
243.
921.
11
62
30
00
019
989.
430.
162.
211.
43
191
10
00
019
997.
020.
122.
251.
31
80
00
00
020
0090
.64
0.55
3.68
2.4
00
853
34
00
2001
8.87
0.25
3.59
1.9
16
444
00
00
Win
ter
1996
90.3
20.
061.
5825
.52
272
191
130
019
979.
930.
080.
2612
.13
701
70
00
019
986.
170.
070.
446.
05
230
00
00
019
992.
560.
210.
364.
96
210
00
00
020
005.
290.
110.
335.
74
490
00
00
020
018.
930.
140.
3717
.96
121
80
00
020
026.
820.
160.
418.
35
320
00
00
0
MA
X,
MIN
and
ME
Dar
ein
m3
s)1,
NS
PE
LL
sar
eco
un
ts,
DS
PE
LL
sar
ein
day
s.F
or
defi
nit
ion
so
fv
aria
ble
s,se
eT
able
1.
Regulated river fish fauna 813
� 2007 The Authors, Journal compilation � 2007 Blackwell Publishing Ltd, Freshwater Biology, 53, 789–813





























