Embed Size (px)
Citation preview

PETER CRESSWELL ANTIGEN PRESENTATION
Getting peptides into MHC class II moleculesThe long search for the MHC-linked genes known to be required forantigen processing has turned up HLA-DMA and HLA-DMB, genesthat seem to encode a dimer similar to the MHC class II molecule.
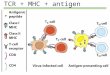
Class II molecules of the major histocompatibilitycomplex (MHC) bind to peptides derived from endo-cytosed proteins and display them on the cell surface,where they are recognized by T lymphocytes that bearthe cell-surface marker CD4. The general principles ofthis phenomenon are reasonably well understood(reviewed in [11). Each MHC class II molecule is aheterodimer of integral transmembrane glycoproteins.The amino-terminal domains of each of the two sub-units, a and 13, combine to form a peptide-bindinggroove, of which the sides are two antiparallel a-helicesand the bottom is made up of a sheet of eight1-strands. Premature occupancy of the class II peptide-binding groove during assembly of the class II moleculewithin the cell is prevented by the early associationof the a and subunits in the endoplasmic reticulumwith a third integral membrane glycoprotein. Thissubunit, the invariant (I) chain, forms a trimer in theendoplasmic reticulum to which three a13 dimers areadded successively.
Complete a13I nonamers are transported from the endo-plasmic reticulum through the Golgi complex and thetrans Golgi network into the endosomal-lysosomalsystem of intracellular vesicles. Entry of the a13Icomplex into the endocytic pathway is mediated by theinvariant chain - specifically by targeting/retentionsignals in its amino-terminal cytoplasmic domain. Theprecise route followed by the at3I complex between thetrans Golgi network and the endosomal system .is amatter of some debate. Electron microscopic evidencefavors the segregation of at3I in the trans Golgi networkaway from the normal constitutive pathway leading tothe plasma membrane, with direct targeting to theendosomal system. Biochemical evidence argues that afraction - perhaps a sizable one - of the total at3Icomplexes traverses the cell surface en route toendosomes.
Within the endosomal-lysosomal system, the invariantchain is proteolytically degraded, releasing a13 dimers.Precisely where this occurs is currently being intensivelyinvestigated. In human B-cell lines transformed with theEpstein-Barr virus, class II molecules accumulate inunusual, membrane-rich acidic vesicles, the 'MHC classII compartments', which contain a number of lysosomalproteins. There is suggestive evidence that these may bethe compartments in which the invariant chain isdegraded, as complexes made up of class II 0a and subunits with partial proteolytic fragments of the invari-ant chain accumulate in relatively dense vesicles.that
also contain lysosomal markers when B-cell lines areincubated with the sulfhydryl-protease inhibitor, leu-peptin [2]. The at3 dimers released following invariantchain degradation are competent to bind peptides, andafter doing so they are transported to the plasma mem-brane. This might involve their transport back to thetrans Golgi network and thence to the cell surface, orthere may be a more direct route. There is, however,much debate at present about the molecular and cellu-lar mechanisms involved in generating the complexes ofclass II molecules and peptides.
A rich history of immunological experiments has shownthat the external foreign antigens that are internalized byantigen-presenting cells constitute the primary source ofthe peptides that - in combination with particular classII molecules - are recognized by CD4-bearing T-helpercells. When the peptides isolated from class II moleculesare analyzed, however, the vast majority are found to bederived from cell-surface glycoproteins or internalizedserum proteins, suggesting that class II-positive antigen-presenting cells continuously sample all of the peptidesgenerated in the endocytic pathway. The introduction offoreign proteins into the pathway results in the genera-tion of novel class II-peptide complexes, which can berecognized by T-helper cells. The peptides that becomeassociated with class II molecules might be generated inthe compartment in which invariant chain degradationoccurs. Alternatively, the peptides could be generatedelsewhere, for example in the lysosomes, and trans-ported to a class II-containing compartment for assemblyinto complexes.
A number of years ago, Pious and co-workers [3] gener-ated several mutant human Epstein-Barr virus-trans-formed B-cell lines that are not able to generatecomplexes of class II a[3 dimers with antigenic peptidesderived from internalized foreign proteins. This defectcannot be ascribed to a failure of expression of the classII molecules themselves, nor to a defect in their assem-bly with peptides: when the mutant cell lines are incu-bated with free antigenic peptides - rather than withthe proteins from which the peptides are derived -they can stimulate T-cells specific for the classII-peptide complex perfectly well. Genetic data suggestthat the genes involved in this defect map to the MHC,closely linked to the class II-encoding genes HLA-DR,HLA-DQ and HLA-DP. The genes defective in these celllines have now been identified [4,5] and are called HLA-DMA and HLA-DMB [6]; homologous genes also exist inthe mouse. The two genes seem to encode the subunits
© Current Biology 1994, Vol 4 No 6 541
542 Current Biology 1994, Vol 4 No 6
compartments, but with their morphology dramaticallyaltered as a result of the absence of DMA and DMB.Other genetic abnormalities in the T2.DR3 cell line areunlikely to be involved, because both normal morphol-ogy and the epitope recognized by the 16.23 antibody,which is absent from T2.DR3 cells, can be restored bythe introduction into the cells of expression vectorsencoding functional DMA and DMB proteins (L. Denzin,N. Robbins and P.C., unpublished observations). Inaddition, the proteolysis of invariant chain and sub-sequent expression of class II dimers on the cell surfaceis much slower in mutant than in wild-type cells; in thelatter, the process is essentially complete within fourhours [7]. Finally, virtually all of the HLA-DR3 moleculesexpressed on the mutant cells bear the same peptide,which is derived from residues 81-104 of the invariantchain and has been called CLIP, the class II-associatedinvariant chain peptide [8].
of a heterodimer with remarkable structural similarity toclass II molecules themselves. Mutant cell lines defectivein either DMA or DMB expression have been identified,and somatic cell hybrids between the two mutant typesare functionally normal [5]. This genetic complementa-tion is consistent with the suggestion that DMA andDMB proteins normally function as a heterodimer.
How might the DMA and DMB gene products functionin antigen processing? To begin to answer this question,it is important to understand more precisely the pheno-type of the antigen-processing mutant cell lines used toisolate the genes (Table 1). Class II molecules areexpressed on the surfaces of the mutant cells in normalamounts. But these class II molecules are conformation-ally abnormal, in that certain monoclonal antibodiesthat bind normal class II molecules fail to bind to them.One such antibody, called 16.23, which recognizes theproduct of the HLA-DR3 MHC class II allele in wild-typecells, was used in the isolation of the original mutantcell lines, in combination with complement, to selec-tively kill normal cells and leave behind the mutants [3].In addition, the a and ,B subunits of class II moleculesin the mutant cells dissociate in the detergent sodiumdodecyl sulfate (SDS), whereas in wild-type cells a largefraction are stable in SDS, probably because they have anormal complement of associated peptides.
In the mutant cell line T2.DR3, which contains a largehomozygous deletion encompassing both the DMA andDMB genes, class II molecules accumulate in largevacuoles prior to their expression on the cell surface [7].In addition to class II molecules and invariant chain,these vacuoles contain the same range of lysosomalproteins as were identified in the MHC class II compart-ments characteristic of normal Epstein-Barr virus-trans-formed cells. The vacuoles are likely to be MHC class II
On the basis of the phenotype of the mutant cell lines,the DMA and DMB gene products could function in anumber of ways. They might be critical for a late stagein class II transport to the cell surface. This wouldexplain the accumulation of HLA-DR3 molecules inswollen MHC class II compartments in T2.DR3 mutantcells. Invariant chain proteolysis might normally becompleted in a compartment later in the pathway,which is inaccessible to class II molecules in theabsence of DMA/DMB, explaining the reduced rate ofinvariant chain degradation in the mutant cells. Recentevidence suggests that complexes of oL3 class II dimerswith CLIP represent an intermediate in the processing ofclass II molecules ([7]; R. Awa and P.C., unpublishedobservations), and the final stages of processing - CLIPremoval and peptide loading - might occur in thishypothetical late compartment.
Alternatively, DMA and/or DMB might play a directrole in invariant chain dissociation, particularly inCLIP removal. Acidification of purified HLA-DR3-CLIPcomplexes in the detergent N-octyl glucoside permitsCLIP removal and antigenic peptide binding [8]. Thismight be catalyzed in vivo by DMA/DMB, an idea thatis superficially attractive, because DMA/DMB dimerscould form a peptide-binding groove akin to that ofconventional class II molecules [6]. The observationthat DMA and DMB lack the extreme polymorphismcharacteristic of classical class II molecules might beconsistent with the DMA/DMB heterodimer havingaffinity for only a single peptide, namely CLIP, ratherthan the wide range of peptides bound by classicalclass II molecules. It is not, however, obvious why theabsence of DMA/DMB heterodimers with such a func-tion should result in the slower invariant chain degrada-tion and the accumulation of class II molecules inintracellular vesicles seen in the mutant cells. Thedefined signal for endosomal retention and targeting ofclass II molecules resides in the cytoplasmic domain ofthe invariant chain, not in the CLIP region, althoughthere is evidence from murine class II mutants thatadditional endosomal targeting signals may be presentin class II molecules themselves [9].

DISPATCH 543
A third model that might explain the mutant phenotypeis that DMA and DMB could be involved in the genera-tion of the endocytic peptides bound by conventionalclass II molecules. It has always been a reasonableproposition that some chaperone-like molecule mightbind the peptides that are generated in an unfriendlyproteolytic environment, such as the lysosome, anddeliver them to a compartment - perhaps the MHCclass II compartment - containing empty class II mol-ecules. Conditions in the lysosome, such as extremeacidity, could favor the binding of large peptides toDMA/DMB. Conditions in the MHC class II compart-ment, such as a somewhat reduced acidity, might favorthe release of these peptides to class II molecules, per-haps coordinated with CLIP removal by a mechanismdirectly involving DMA/DMB.
Some support for a mechanism involving an essentialdirect interaction of class II molecules with somethingthat promotes CLIP removal and/or peptide loadingcomes from a new mutant cell line [10]. Cells of this lineexpress a structurally altered mutant HLA-DR3 mol-ecule, which has lost the capacity for CLIP dissociationand peptide loading in vivo. The mutated molecule hasan extra Nlinked carbohydrate side chain in the mem-brane-proximal domain of its a chain, at a positionso far removed from the peptide-binding groove thatit would be unlikely to affect peptide binding directly.But the glycan moiety might affect the interaction ofthe DR3 molecule with a molecule essential for promo-ting peptide-binding, which could be the putativeDMA/DMB dimer.
Our current understanding is limited by the lackof reagents for detecting the DMA and DMB proteins.When suitable antibodies become available, questionscan be asked about the assembly, transport andsubcellular localization of DMA and DMB, whether theybind proteins or peptides, and whether they interact
physically with class II molecules, invariant chain orCLIP. Until then, we can only speculate about theprecise role played by DMA and DMB in antigenprocessing.
References1. CRESSWELL P: Assembly, transport and function of MHC class II
molecules. Annu Rev Immunol 1994, 12:259-293.2. MARIC MA, TAYLOR MD, BLUM JS: Endosomal aspartic
proteinases are required for invariant-chain processing.Proc NatlAcad Sct USA 1994, 91:2171-2175.
3. MELLINS E, SMITH L, ARP B, COTNER T, CELIS E, PIOUS D:Defective processing and presentation of exogenous antigensin mutants with normal HLA class II genes. Nature 1990,343:71-74.
4. MORRIS P, SHAMAN J, ATTAYA, M, AMAYA M, GOODMAN S,BERGMAN C, MONACO JJ, MELINS, E: An essential role for HLA-DM in antigen presentation by class 11 major histocompatibilitymolecules. Nature 1994, 368:551-554.
5. FLING SP, ARP B, PIOUS D: HLA-DMA and -DMB genes are bothrequired for MHC class II/peptide complex formation inantigen-presenting cells. Nature 1994, 368:554-558.
6. KELLY AP, MONACO JJ, CHO S, TROWSDALE J: A new human HLAclass II-related locus, DM. Nature 1991, 353:571-576.
7. RIBERDY JM, AWA RR, GEUZE HJ, CRESSWELL P: Transport andintracellular distribution of MHC class H molecules and associ-ated invariant chain in normal and antigen-processing mutantcell lines. J Cell Biol 1994, in press.
8. RIBERDY JM, NEWCOMB JR, SURMAN MJ, BARBOSA JA, CRESSWELL P:HLA-DR molecules from an antigen-processing mutant cell lineare associated with invariant chain peptides. Nature 1992,360:474-477.
9. CHERVONSKY AV, GORDON L, SANT AJ: A segment of the MHCclass H 3 chain plays a critical role in targeting class II mole-cules to the endocytic pathway. Int Immunol 1994, in press.
10. MELLINS E, CAMERON P, AMAYA M, GOODMAN S, PIOUS D, SMITH L,ARP B: A mutant human histocompatibility leukocyte antigenDR molecule associated with invariant chain peptides.JExp Med 1994, 179:541-549.
Peter Cresswell, Section of Immunobiology, HowardHughes Medical Institute, Yale University School ofMedicine, 310 Cedar Street, New Haven, Connecticut06510, USA.