Embed Size (px)
Citation preview

Trans. Br. mycol, Soc. 71, (1) 69-74 (1979)
[ 69 ]
Printed in Great Britain
ANALYSIS OF TWO SOUTHERN HEMISPHERE COPROPHILOUSFUNGUS SUCCESSIONS
By L. A. NAGY AND K. M. HARROWER
Botany Department, Australian National University, Canberra ACT 2600, Australia
Studies were made of the fungus succession on kangaroo and rabbit dung collected fromthe same locality. The presence or absence of eight species was assessed over a 74-dayincubation period in moist chambers and the reproductive output of 15 species was assessedover the same incubation period using a ' window trap' procedure.
Although the fungal succession was similar on the two substrates, considerable differenceswere noted in the duration and profuseness of fruiting of species in the two habitats andthe characteristic species pattern which has been drawn up for each substrate is based onreproductive output on that substrate.
The coprophilous fungus succession has been re-latively well-investigated in the northern hemi-sphere (Harper & Webster, 1964; Richardson,1972; Angel & Wicklow, 1974, 1975) but, apartfrom the work of Bell (1975), few investigationshave been carried out in the southern hemisphere.The present study had three main aims: to provideinformation on the coprophilous fungus successionin the southern hemisphere, that would hopefullycomplement northern hemisphere work and pro-vide information concerning the global distributionof coprophilous fungi; to provide information onthe possible substrate preferences of coprophilousfungi by studying the succession on the faeces ofrabbit and kangaroo sharing the same feedingterritory; and to provide information on the copro-philous fungus succession over a relatively longincubation period.
Rabbit faeces were selected in order to provide acomparison with the large body of previous workand kangaroo faeces because the coprophiloussuccession on this substrate has so far been un-described. It was also thought that a comparison ofthe succession on the faeces of an indigenous andan exotic species sharing the same feeding territorymight prove useful.
MATERIALS AND METHODS
Kangaroo and rabbit faeces were collected from theTidbinbilla Nature Reserve (Australian CapitalTerritory) in February, 1977. Every effort wasmade to ensure that the faeces collected were lessthan 24 hold. Using sterilized tweezers, faecalpellets were placed 1 per Petri dish (depth 5 em)on filter paper and kept moist throughout theduration of the experiment with sterile distilledwater. Pellets were incubated in a controlled-
environment chamber using a combined daily lightand temperature regime alternating between 12 hlight at 25° and 12 h darkness at 15° for a periodof 74 days. The passage of light to the pellets wasrestricted by using dishes which were completelyblacked out except for a clear window (2'0 x 3'5 ern)located in the centre of each dish top. Below eachwindow a microscope slide was placed, whichserved as a collection surface for the dischargedreproductive structures of certain coprophilousspecies. At selected times over the incubationperiod, the succession was assessed by recordingthe presence or absence of fungus fruiting bodiesand scoring for the discharged reproductive struc-tures deposited on the microscope slides (the'window-trap' method), Eight species were asses-sed by the presence or absence method and 15species were assessed by the 'window-trap'method.
RESUL TS AND DISCUSSION
The 22 coprophilous fungi identified during thestudy are listed in Table 1. Data obtained byscoring for the presence or absence of fungusfruiting structures on ten kangaroo and ten rabbitpellets over the incubation period are shown inFig. 1. Data obtained for the same ten kangarooand the same ten rabbit pellets using the' win-dow-trap' method are shown in Figs 2 and 3respectively.
The minimum time required to produce maturefruiting bodies, referred to as the minimum fruit-ing time (M,F.T.), for each of the eight speciesassessed by the presence or absence method andfor each of the 15 species assessed by the' window-trap' method are shown in Table 2.
Most of the fungi recorded on rabbit faeces in
0007-1536179/2828-4740 $01.00 © 1979 The British Mycological Society
7° Southern-hemisphere coprophiloussuccessions
Kangaroo dung Rabbit dung
I I I
Pilaira anomala
I5
O ........ur..........r'-....--',.-...,..-+----r..L.
5
o10
10
Saccobolus glaber
Spororm ia intermedia
Delitschia didyma
I IAscobolus sticto ideus
Thelebolus I/ Ol/ US
5
5
OL.--+-..L..1.y.l-..JL,-....ly-----l..----+----r....l.10
. ': [ ,III~ LlllJ--.-....-...---.-....-r-<; 100.
]j 5
=~ 0 ....~........&.r'.....I""r--&.,........~-+-..-'".~ 10
~'-o;.,
~ 0 '-----'t........-'-r''--'T----Ly--.....,.--+-~50 10
'"~
Cop r inus sp.
I
o 10 20 30 40 50 60 70
5
O'---¥-....L.J+....,-Ly--L,r---+--~
10
5
0L.-..,......L.J+....,-Ly--L,--+---r-1
o 10 20 30 40 50 60 70
Lengt h of incub ation peri od (days)
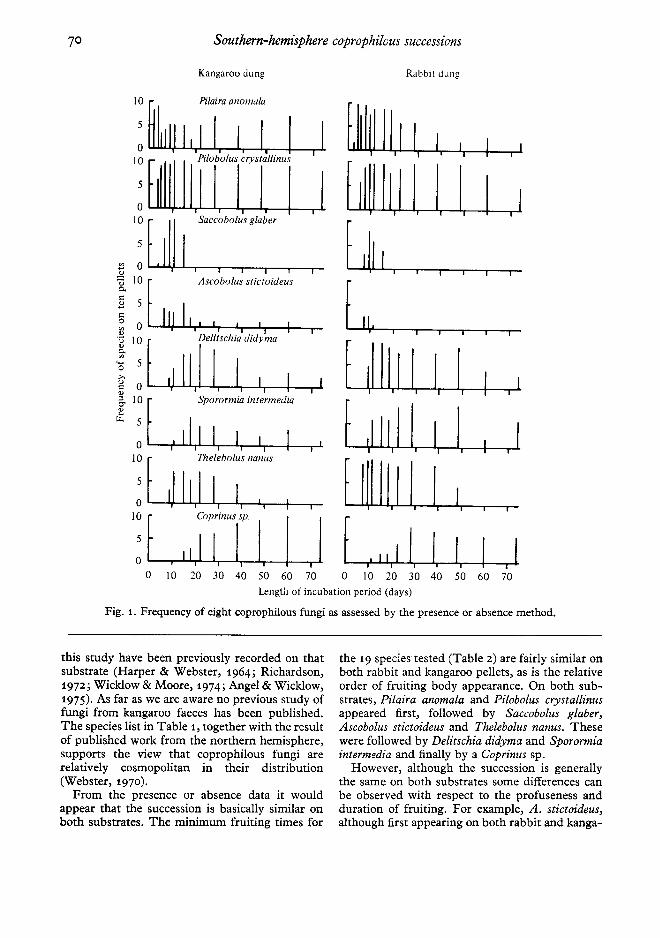
Fig. 1. Frequency of eight coprophilous fungi as assessed by the presenceor absencemethod.
this study have been previously recorded on thatsubstrate (Harper & Webster, 1964; Richardson,1972; Wicklow& Moore, 1974; Angel & Wicklow,1975). As far as we are aware no previous study offungi from kangaroo faeces has been published.The species list in Table 1, together with the resultof published work from the northern hemisphere,supports the view that coprophilous fungi arerelatively cosmopolitan in their distribution(Webster, 1970).
From the presence or absence data it wouldappear that the succession is basically similar onboth substrates. The minimum fruit ing times for
the 19 species tested (Table 2) are fairly similar onboth rabbit and kangaroo pellets, as is the relativeorder of fruiting body appearance. On both sub-strates, Pilaira anomala and Pilobolus crystallinusappeared first, followed by Saccobolus glaber,Ascobolus stictoideus and Th elebolus nanus. Thesewere followed by Delitschia didyma and Sp orormiainterm edia and finally by a Coprinus sp.
However, although the succession is generallythe same on both substrates some differences canbe observed with respect to the profuseness andduration of fruiting. For example, A. stictoideus,although first appearing on both rabbit and kanga-
L. A. Nagy and K. M. Harrower 71
1 ,
Podospora curvula
iAscobolus degluptus
Saccobolus versicolor
Asco bolus albidu s
Pilobolus crys tallinus
ISaccobolus glaber
(
Thelebolus nanus
I I (Sordaria macrospora
200~~~200 ill
"rl=.............,r----r---"T""---r--~--__,-
50U(h----r---..------.----r_r---y--200 iL--.c::;l::::::=:t::::I:::;:=z=:::>...,----.----"T--.,.---~-
50i-----r',~ ~odospor~ deciPie~s
liL---r--~~~I i--.--~--,---,-------,------,-
50 l-----r:=-----r--<==:er=====r===a.,----r--....,...-
No. asci 100 iL__'-_-"'T"oI:::===lt:r==t:::;::==:::;===""'f===""'-
No. asci
No. asci
No. asci
No. asci
No. asci
No. asci
No. asci
No . sporangia
Sporormia intermedia
Sporormia ambigua
Coniochaeta hansenii-I
i Ascobolus immersus
5 L---.---.-~d i II i'-----r--~----.----r---r----.
51l-...---'-'---,---,---,-----,-----F====='=
5i'-----r------r---o---r---r----r----r-
No. asci
No. spores
No. spores
No. spores
I ,100 i---..-----r-==c:;~===!:;===1,.-----,I--,--JNo. asci
Coniochaeta discopora
I10o
No. spo res
i I20 30 40 50 60 70Length of incubation period (days)
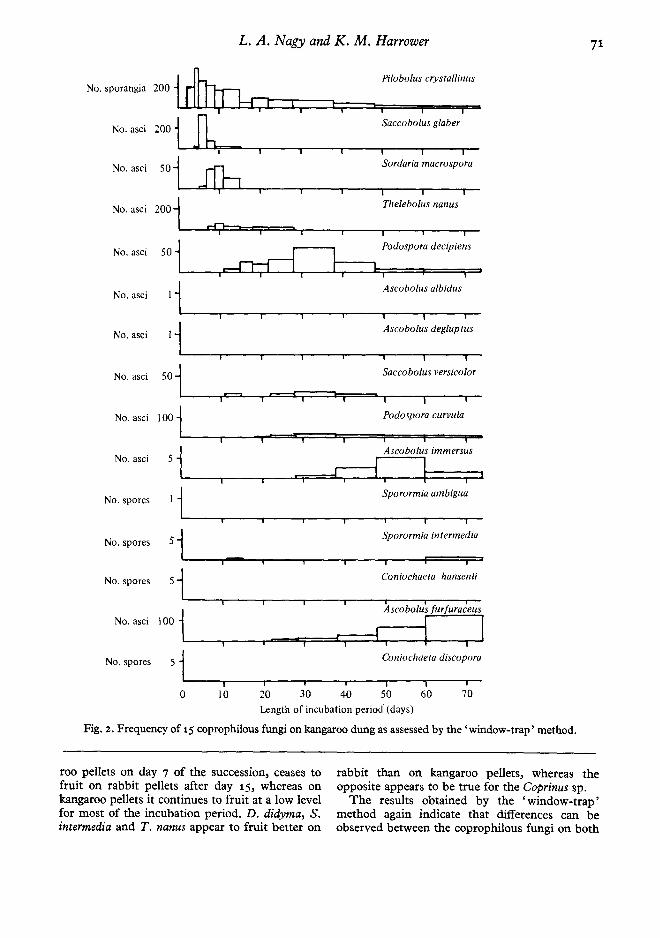
Fig. 2. Frequency of 15 coprophilous fungi on kangaroo dung as assessed by the 'window-trap ' method.
roo pellets on day 7 of the succession, ceases tofruit on rabbit pellets after day 15, whereas onkangaroo pellets it continues to fruit at a low levelfor most of the incubation period. D. didyma, S.intermedia and T. nanus appear to fruit better on
rabbit than on kangaroo pellet s, whereas theoppos ite appears to be true for the Coprinus sp.
The results obtained by the 'window-trap 'method again indicate that differences can beobserved between the coprophilous fungi on both
72 Southern-hemisphere coprophilous successions
,70
I60
I I rSaccobolus versicolor
i
Coniochaeta discospora
r I IAscobolus furfuraceus
r rConoichaeta hansenii
Sporormia intermedia
Sporormia ambigua
iiiAscobolus immersus
I , rPodospora curvula
rAscobolus albidus
Pilobolus crystallinus
IPodospora decipiens
I IAscobolus degluptus
I IThelebolus nanus
ISordaria macrospora
I i ISaccobolus glaber
200 ~t::::::J::;;:::::=::::;:::=::::I::;:=="""'==-
200 il...--'::::L..y::=--r-'----r--~---.---.___-......-No. asci
No. asci 50i
I
No. asci 20oLITilo
No. asci 50i~
No. asci IUr
No. asci1i
0I
No. asci 50i~i I
No. asci IOOl ~No. asci
5i""'f""==' I
No. spores Ii dJr-=,No. spores
51 pIi
No. spores5i D,
No. asci looi
~No. spores
5i 0r I0 10 20 30 40
No. sporangia
50
Length of incubation period (days)
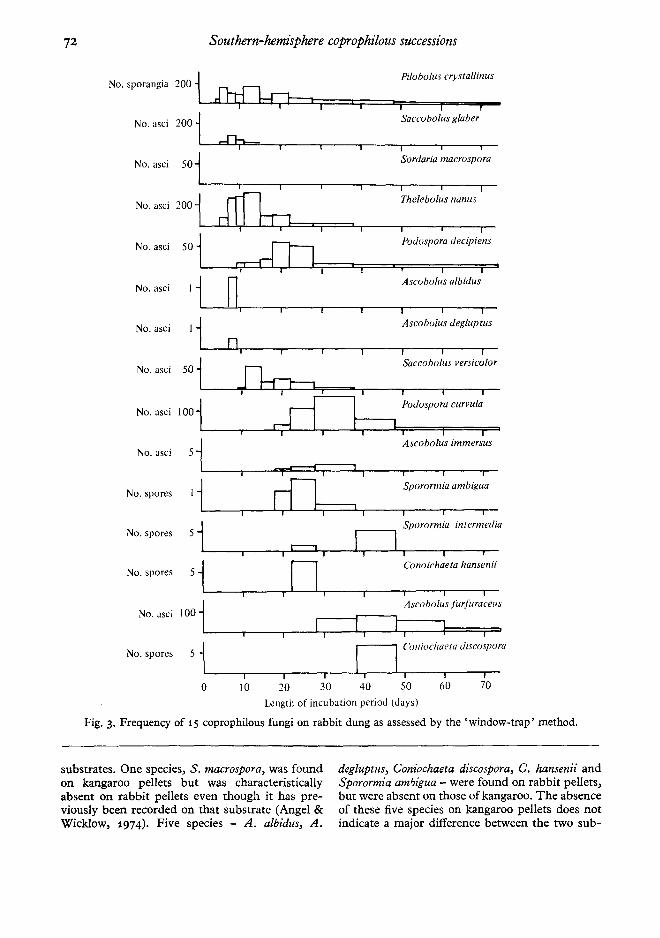
Fig. 3. Frequency of 15 coprophilous fungi on rabbit dung as assessed by the' window-trap' method.
substrates. One species, S. macrospora, was foundon kangaroo pellets but was characteristicallyabsent on rabbit pellets even though it has pre-viously been recorded on that substrate (Angel &WickIow, 1974). Five species - A. albidus, A.
degluptus, Coniochaeta discospora, C. hansenii andSporormia ambigua - were found on rabbit pellets,but were absent on those of kangaroo. The absenceof these five species on kangaroo pellets does notindicate a major difference between the two sub-

L. A. Nagy and K. M. Harrower 73
Table 1. Coprophilous fungus species identified during study
Kangaroo Rabbit Previous recordpellets pellets on rabbit pellets*
Pilaira anomala (Ces.) Schroet. + + 1*Pilobolus crystallinus (Wigg.) Tode + + 1Rhizopus nigricans Ehrenberg + .+Mucor mucedo (L.) Brefeld + + 1Ascobolus albidus Crouan + 2*A. degluptus Speg, +A i furfuraceus Pers . ex Fr. + + 2A. immersus Pers . per Pers. + + 2,4*A. stictoideus Speg. + + 2Saccobolus glaber (Pers.) Lamb. + +S. versicolor (Karst.) Karst. + + 2
Thelebolus nanus Heimerl + + 2Coniochaeta discospora (Auersw.) Cain + 2,4C. hansenii (Oud.) Cain + 2
Delitschia didyma Auersw. + +Podospora curuula (DeBary) Niessl + + 2
P. decipiens (Wint.) Niessl + + 2
Sordaria humana (Fckl) Wint. + + 3*S. macrospora Auersw. + 4Sporormia ambigua Niessl + 2S. intermedia Auersw. + + 1,2Coprinus sp. + +
* 1, Harper & Webster (1964) ; 2, Richardson (1972); 3, Wicklow & Moore (1974); 4, Angel &Wicklow (1975).
Table 2 . Minimum fruiting times (day s) of 19 coprophilous fungi
Presence or absence method ' Window-trap ' method,..-------" \
Kangaroo pellets Rabbit pellets Kangaroo pellets Rabbit pellets
Pilaira anomala 2 2Pilobolus cry stallinus 4 5 4 5Ascobolus albidus 9A. degluptu s 9A. furfuraceus 28 38A. immersus 38 22A. stictoideus 7 7Saccobolus glaber 5 7 5 7S. versicolor 15 11
Thelebolus nanus 9 7 9 7Coniochaeta discospora 48C. hansenii 28Delitschia didyma 9 9Podospora curvula 22 22P . decipiens 15 11
Sordaria macrospora 7Sporormia ambigua 22S. intermedin 11 9 60 28Coprinus sp, 15 11
strates, as even on rabbit pellets these five specieswere only recorded at low frequencies. Speciessuch as P. crystallinus, S. glaber, Sordaria m:lcro-spora, A. immersus and Ai furfuraceus fruited better
on kangaroo pellets than on rabbit pellets, whereasthe reverse was found to be true for species likeT. nanus, Saccobolus versicolor, Podospora curvulaand S. ambigua.

74 Southern-hemisphere coprophilous successions
It appears from the 'window-trap' data thatcertain members of the succession exhibit fluctua-tions in their reproductive output. These fluctua-tions in reproductive output over time can be seenfrom the total 'window-trap' data (Figs. 2, 3) aswell as from individual pellet data not shown here.The phenomenon is further exemplified by theshorter assessment intervals used in the earlierphases of the succession. However, slight fluctua-tions were also observed in the later phases of thesuccession.
The reasons for fluctuations in reproductiveoutput are not clear. It cannot yet be establishedwhether these fluctuations are a result of inter-specific interactions or a species characteristicphenomenon, which may be observed even inaxenic culture. The latter may suggest some degreeof cyclic allocation of energy between vegetativegrowth and reproductive growth. A measure ofreproductive output in axenic cultures using the'window-trap' method would prove interestingand is currently under study.
There is a good correlation between the presenceor absence data and the' window-trap' data, withrespect to the minimum fruiting time, the relativeorder of appearance and the relative intensity offruiting on rabbit versus kangaroo pellets. How-ever, on its own, the presence or absence datacould be interpreted erroneously. Its disadvantageis that it is a qualitative method of assessment andtherefore requires some quantitative method toassist correct interpretation of the succession. Toillustrate the inadequacy of presence or absencedata the case of P . crystallinus could be considered.From presence or absence data it would appearthat, on both rabbit and kangaroo pellets, P.crystallinus starts fruiting within 3-5 days, reachesa maximum within 7-9 days and then remains atapproximately this high level throughout theassessment period.
This interpretation is somewhat inaccurate whenone also takes into consideration the quantitative'window-trap' data. These data show that theinitial maximum in reproductive output of spo-rangia is between 5 and 6 days and that after thismaximum the intensity offruiting of P. crystallinusdrops off considerably during the remainder of theassessment period. The' window-trap' data thushelps to clarify the presence or absence data.
From the' window-trap' data one can suggest agroup of coprophilous fungi that, in terms ofquantity and/or duration of reproductive output,
play a dominant role in the succession. Thesedominant members form a characteristic sequenceon each of the two substrates investigated. Forkangaroo pellets this characteristic sequence is P.crystallinus ~ S, macrospora ~ P. decipiens ~ A.immersus """* A. furfuraceus. The correspondingsequence for rabbit pellets is P. crystallinus """* T.nanus """* P. decipiens >- P. curuula >» A. fur-furaceus.
It appears from Figs 2 and 3 that the copro-philous fungus succession decreases in generalactivity after about 50 days incubation. No addi-tional species were recorded by the 'window-trap'method on either kangaroo or rabbit pellets after50 days incubation. There may be several reasonsfor this. One reason may be the reduction in avail-able nutrients or growth factors. This is question-able, as even after 70-80 days incubation a pellet isstill recognizably a pellet, with at least two-thirdsof its original dry weight still present (Harrower &Nagy, unpubl.) ,
An alternative explanation, is that the observeddecrease in general activity may be producedby the transfer of pellets from the field to thelaboratory.
The authors are indebted to Mrs V. Keraitis forcompetent technical assistance.
REFERENCES
ANGEL, K. & WICKLOW, D. T. (1974), Decompositionof rabbit faeces; an indication of the significance ofthe coprophilous microflora in energy flow schemes.Journal of Ecology 62,429-437.
ANGEL, K. & WICKLOW, D. T. (1975). Relationshipbetween coprophilous fungi and faecal substrates inColorado grassland. My cologia 67, 63-74.
BELL, A. (1975). Fungal succession on dung of thebrush-tailed opossum in New Zealand. New ZealandJournal of Botany 13, 437-462.
HARPER, J. E. & WEBSTER, J. (1964). An experimentalanalysis of the coprophilous fungus succession.Transactions of the British Mycological Society 47,511-530.
RICHARDSON, M. J. (1972). Coprophilous ascomyceteson different dung types. Transactions of the BritishMycological Society 58, 37-48.
WEBSTER, J. (1970). Coprophilous fungi. Transactionsof the British Mycological Society 54, 161-180.
WICKLOW, D. T. & MOORE, V. (1974). Effectof incuba-tion temperature on the coprophilous fungal succes-sion. Transactions of the British Mycological Society62, 411-415.
(A ccepted for publication 31 May 1978)





























