Embed Size (px)
Citation preview

ANA BEATRIZ SOTERO SIQUEIRA
Perfil Enzimático de Dermatófitos e Avaliação da Atividade
Antifúngica de Própolis e Lectinas
RECIFE-PE 2008
UFPE

UNIVERSIDADE FEDERAL DE PERNAMBUCO CENTRO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS DOUTORADO EM CIÊNCIAS BIOLÓGICAS
ANA BEATRIZ SOTERO SIQUEIRA
Perfil Enzimático de Dermatófitos e Avaliação da Atividade Antifúngica de Própolis e Lectinas
RECIFE-PE 2008

ANA BEATRIZ SOTERO SIQUEIRA
Perfil Enzimático de Dermatófitos e Avaliação da Atividade Antifúngica de Própolis e Lectinas
Tese apresentada ao Programa de Pós-Graduação
em Ciências Biológicas, nível Doutorado, como
parte dos requisitos para obtenção de grau de
Doutor em Ciências Biológicas, na área de
Microbiologia
Orientadora: Profa. Dra. Ana Lúcia Figueiredo Porto
RECIFE-PE 2008

Siqueira, Ana Beatriz Sotero Perfil enzimático de dermatófitos e avaliação da
atividade antifúngica de própolis e lectinas. / Ana Beatriz Sotero Siqueira. – Recife: A Autora, 2008.
177 fls. .: il.
Tese (Doutorado em Ciências Biológicas) – UFPE. CCB
1. Dermatófitos 2. Atividade antifúngica 3. Própolis
4. Lectinas I.Título
582.28 CDU (2ª. Ed.) UFPE 579.5 CDD (22ª. Ed.) CCB – 2008 –076


AGRADECIMENTOS
À UNIVERSIDADE FEDERAL DE PERNAMBUCO, especialmente ao Curso de Pós-
Graduação em Ciências Biológicas, pela oportunidade oferecida para a realização do Curso de
Doutorado em Ciências Biológicas, área Microbiologia.
À minha eterna Orientadora Profa Dra. ANA LÚCIA FIGUEIREDO PORTO, que
admiro tanto pela dedicação profissional quanto pelo ser humano generoso e sábio que é.
Obrigada pela confiança, pelo estímulo e pelos ensinamentos que sempre me acompanharão.
À minha segunda eterna orientadora Profa Dra. LUSINETE ACIOLE DE QUEIROZ,
pela atenção, confiança, apoio, ensinamentos e antes e durante este trabalho. “Tic, tic , tic, tac...”
Aos professores do Curso de Doutorado em Ciências Biológicas, pelos ensinamentos
prestados durante as disciplinas.
À PhD. RITA DE CÁSSIA CARVALHO MAIA, querida amiga que tanto admiro,
atenciosa, generosa e que sempre se mostrou disposta a colaborar para meu crescimento
profissional.
À Profª. Dra. CRISTINA MARIA DE SOUZA-MOTTA, pela inesquecível e valiosa
contribuição durante o desenvolvimento deste trabalho.
As professoras Dra. REJANE PEREIRA NEVES e Dra. OLIANE MARIA CORREIA
MAGALHÃES, por estarem sempre disponíveis a transmitir ensinamentos, além da amizade e
respeito.
Ao Prof. PhD. ARMANDO MARSDEN, pela contribuição e companheirismo.
À todos os funcionários do Departamento de Micologia e do Laboratório de
Imunopatologia Keizo Asami, desta Universidade, pela ajuda sempre prestada.
Aos amigos do LAViTE/FIOCRUZ: RITA DE CÁSSIA CARVALHO MAIA,
BARTOLOMEU ACIOLE DOS SANTOS, RAFAEL DHALIA, CARLOS EDUARDO
CALZAVARA SILVA e GERUSA BARROS por me receberem de braços abertos dispostos a
ajudar, num momento importante da minha vida.

Ao meu querido amigo BRUNO SEVERO GOMES, por estar sempre ao meu lado,
dividindo comigo todos os momentos durante esses anos. Obrigada, Bruninho, pela ajuda
indispensável, compreensão, amizade sempre presente, por tudo! Esta conquista é nossa.
À minha querida amiga IDALINA CABUIM, pela colaboração, compreensão,
companheirismo e amizade da qual não esquecerei.
Aos amigos GILMARA ARAÚJO e FILIPE DUARTE, pelo incentivo, carinho e
presença constante.
Aos amigos do Laboratório de Imunopatologia Keizo Asami, CAROLINA
ALBUQUERQUE e RÔMULO LINS pela amizade conquistada e agradável convívio durante
esses anos.
Aos amigos do Laboratório de Micologia Médica, pela amizade e ajuda sempre prestada.
À MINHA FAMÍLIA, pelo amor, paciência, compreensão, presença constante,
enfim...por tudo que faz por mim, impossível de esquecer e grande demais para ser resumido em
poucas palavras.

DEDICO
A Deus, pelo dom da vida e por ser a minha
força em todos os momentos.
Aos meus pais Elinaldo Siqueira da Silva e
Ana Lúcia Sotero Siqueira, por serem a essência
da minha formação pessoal, contribuindo
ativamente para a realização deste dia.
A minha irmã Maria Cecilia Sotero
Siqueira, sempre presente com seu apoio, estímulo
e amizade.
A toda minha grande família por ser um
exemplo de FAMÍLIA e porque é a base necessária
da minha vida
A todos os meus amigos, pelo
companheirismo e estímulo dados todos os dias.

É HOJE Composição: Didi e Maestrinho
A minha alegria atravessou o mar
E ancorou na passarela
Fez um desembarque fascinante
No maior show da terra
Será que eu serei o dono dessa festa
Um rei
No meio de uma gente tão modesta
Eu vim descendo a serra
Cheio de euforia para desfilar
O mundo inteiro espera
Hoje é dia do riso chorar
Levei o meu samba pra mãe de santo rezar
Contra o mal olhado eu carrego meu patuá
Eu levei!
Acredito ser o mais valente nessa luta do rochedo como mar
E como ar!
É hoje o dia da alegria
E a tristeza, nem pode pensar em chegar
Diga espelho meu!
Diga espelho meu
Se há na avenida alguém mais feliz que eu
Diga espelho meu
Se há na avenida alguém mais feliz que eu

LISTA DE FIGURAS
Figura Página
1A Coleta de exsudato resinoso verde por abelha Apis
mellifera..................................................................................
19
1B Coleta de exsudato resinoso vermelho por abelha Apis
mellifera...................................................................................
19
2 Representação esquemática de três tipos de lectinas de
plantas: merolectina, hololectina, quimerolectina...................
65
3 Lectinas com atividade antifúngica e especificidade de
ligação aos carboidratos..........................................................
69
4 Lectinas sem atividade antifúngica e especificidade de
ligação aos carboidratos..........................................................
70

LISTA DE TABELAS
Tabela Página
1 Componentes químicos gerais de amostras de
própolis..........................................................................................
46
2 Compostos químicos da própolis verde do Brasil......................... 57
3 Compostos químicos da própolis vermelha do Brasil................... 62

RESUMO
Dermatófitos são fungos filamentosos capazes de infectar pele, pêlos e unhas. São
classificados nos gêneros Microsporum, Trichophyton e Epidermophyton. Com o
objetivo de analisar o perfil enzimático desses fungos e avaliar atividade antifúngica de
própolis brasileiras (verde e vermelha), como também de lectinas de sementes de
leguminosas do Brasil (DViol, DRL, ConBr e LSL), foram utilizadas trinta amostras de
seis espécies de dermatófitos preservadas na Coleção de Culturas Micoteca - University
of Recife Mycolgy (URM), do Departamento de Micologia, da Universidade Federal de
Pernambuco, sendo cinco amostras de cada espécie: Microsporum canis, Microsporum
gypseum, Trichophyton tonsurans, Trichophyton mentagrophytes, Trichophyton rubrum e
Epidermophyton floccosum. Todas as amostras, preservadas ente 2 e 22 anos, foram confirmadas
taxonomicamente. Foi verificado que o período de estocagem não interferiu na capacidade
crescimento e esporulação dos dermatófitos (p-valor < 0.01) e que a estocagem induz ao
pleomorfismo de M. canis, independente do período. Atividade de proteases foi verificada em
57% das amostras. Todas foram capazes de crescer em meio de ágar-queratina e ágar-gelatina,
indicando atividade de queratinases e colagenases, respectivamente. Em nenhuma amostra foi
constatada atividade de fosfolipases mas em 90% foi observada atividade lipásica. O período de
estocagem não interferiu na atividade de proteases, queratinases, colagenases, fospolipases e
lipases. Para verificação da atividade antifúngica foi utilizado o método publicado no documento
M38-A pelo “Clinical and Laboratory Standards Institute”, utilizando como controle o
itraconazol e a terbinafina. A concentração inibitória mínima foi determinada por leitura visual e
por leitor de microplacas. A concentração fungicida mínima foi determinada pela ausência do
crescimento fúngico em meio de cultura Sabouraud líquido. O extrato alcoólico da própolis
verde apresentou atividade fungistática entre 8 e 1024µg/mL e o da própolis vermelha entre 8 e
512µg/mL, variando para cada espécie de dermatófito analisada. O extrato aquoso da própolis
verde e as soluções de lectinas não apresentaram atividade antifúngica. Foi constatado que as
lectinas utilizadas apresentaram discreto efeito estimulador sobre o crescimento dos dermatófitos
em cultivo, e que a lectina DViol também estimulou a esporulação das amostras de E. floccosum.
Dessa forma, foi demonstrado que não houve interferência para expressão enzimática desses
dermatófitos preservados na Coleção de Cultura Micoteca - URM. Foi verificado também que o
potencial antifúngico dos extratos alcoólicos das própolis verde e vermelha para essas espécies,
sugerem aplicações futuras como tratamento alternativo para dermatofitoses.
Palavras chave: dermatófitos, perfil enzimático, própolis verde brasileira, própolis vermelha
brasileira

ABSTRACT
Dermathophytes are a group of filamentous which infect skin, hair and nails. They are
classified in three genera Microsporum, Trichophyton and Epidermophyton. With the
purpose of evaluating the enzymatic profile of dermatophyte samples and the antifungal
activity of brazilian propolis (green and red) and of lectins from legumes seeds of
Brazil (DViol, DRL, ConBr e LSL), were used 30 samples from six different species of
dermatophytes stored for different periods of time at the Mycotheca Cultures Collection
- University of Recife Mycology (URM), from the Federal University of Pernambuco.
They included five samples as following: Microsporum canis, Microsporum gypseum,
Trichophyton tonsurans, Trichophyton mentagrophytes, Trichophyton rubrum and
Epidermophyton floccosum. It was observed that the storage period (2 to 22 years) did not affect
growth ability and sporulation among the dermatophytes (p value < 0.01) and storage in general,
induces M. canis pleomorphism, independently to the period of time. Protease activity was
observed in 57% of the samples. All of them were capable of growing in agar-keratin and agar-
gelatin, indicating keratinase and collagenase activity, respectively. None of the samples showed
phospholipase activity, but 90% of them showed lipase activity. The storage period did not
intervene on protease, keratinase, collagenase, phospholipase, and lipase production. In order to
evaluate the antifungal activity, the method was applied as suggested in the M38-A file, reported
by the “Clinical and Laboratory Standards Institute”, using itraconazole and terbinafine as
controls. The inhibitory concentration was determined by both, visual examination and by a
microplate reader. The minimal fungicide concentration was determined by the lack of fungal
growth in Sabouraud culture medium. The green propolis alcoholic extract showed a fungistatic
activity between 8 and 1024µg/mL, and the red propolis between 8 and 512µg/mL, varying
according to the dermatophyte species analyzed. The green propolis aquous extract and the
lectins solutions did not show any antifungal activity. It was observed that the lectins showed a
growth stimulatory effect on the dermatophytes studied. Moreover, the lectin DViol also
stimulated sporulation in E. floccosum samples. In the present study was demonstrated that there
was no interference to the enzymatic profile of the dermatophytes sample from the Mycotheca
Culture Collection - University of Recife Mycology (URM). It was also observed that
the antifungal potential of the alcoholic extracts of the green and red propolis to those
species, suggest their future application as an alternative treatment to dermatophytosis.
Keywords: dermatophytes, enzymatic profile, green brazilian propolis, red brazilian
propolis

SUMÁRIO
Página
AGRADECIMENTOS
LISTA DE FIGURAS
LISTA DE TABELAS
RESUMO
ABSTRACT
1. INTRODUÇÃO....................................................................................................... 14
2. REVISÃO DA LITERATURA.............................................................................. 21
2.1. Dermatófitos............................................................................................................ 21
2.2. Preservação.............................................................................................................. 24
2.3. Enzimas................................................................................................................... 27
2.4. Drogas antifúngicas e teste de susceptibilidade antifúngica................................ 33
2.5. Própolis.................................................................................................................... 41
2.5.1. Características gerais............................................................................................ 41
2.5.2. Fonte botânica...................................................................................................... 43
2.5.3. Composição química............................................................................................ 44
2.5.4. Atividades biológicas........................................................................................... 49
2.5.5. Extratos alcoólico e aquoso.................................................................................. 52
2.5.6. Própolis verde....................................................................................................... 55
2.5.6.1. Fonte botânica................................................................................................... 55
2.5.6.2. Composição química......................................................................................... 55
2.5.6.3. Atividades biológicas........................................................................................ 58
2.5.6.4. Extratos alcoólico e aquoso............................................................................... 58
2.5.7. Própolis vermelha................................................................................................. 60
2.5.7.1. Fonte botânica................................................................................................... 60
2.5.7.2. Composição química......................................................................................... 61
2.5.7.3. Atividades biológicas........................................................................................ 61
2.6. Lectinas.................................................................................................................... 63
3. OBJETIVOS............................................................................................................. 71
3.1 Objetivo Geral........................................................................................................... 71
3.2 Objetivos Específicos................................................................................................ 71
4. REFERÊNCIAS BIBLIOGRÁFICAS.................................................................. 72
5. ARTIGOS................................................................................................................. 109

5.1. Taxonomic aspects and enzymatic profile of dermatophytes samples stored at the
Mycotheca Cultures Collection – University Of Recife Mycology
(URM)….......................................................................................................
110
5.2. Atividade antifúngica da própolis verde e da vermelha do Brasil em relação a
espécies de dermatófitos..................................................................................................
127
5.3. Susceptibilidade de espécies de Trichophyton a própolis verde e a vermelha do
Brasil................................................................................................................................
143
5.4 Efeito de lectinas de leguminosas do Brasil sobre o crescimento in vitro de
dermatófitos.....................................................................................................................
160
6. CONCLUSÕES.......................................................................................................... 177

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
14
1. INTRODUÇÃO
Os dermatófitos, agentes etiológicos de dermatofitoses, compreendem um grupo de
fungos filamentosos com propriedades queratinofílicas e queratinolíticas, e são
classificados em três gêneros: Microsporum, Trichophyton e Epidermophyton. São
capazes de infectar pele, pêlos e unhas, sendo diferenciados através de aspectos
macroscópicos, microscópicos e fisiológicos (WEITZMAN; SUMMERBELL 1995;
LARONE 1996; LACAZ et al., 2002). Entretanto, o pleomorfismo comum em dermatófitos
dificulta a manifestação desses aspectos tornando a identificação, por vezes, falha
(RIPPON, 1990; ELEWSKI, 1992; FISHER; COOK, 2001; SIDRIM; ROCHA, 2004;
LACAZ et al., 2002). Um dos parâmetros para classificação taxonômica foi proposto por
Emmons (1934), que estabeleceu como critério a morfologia de esporos e estruturas de
ornamentação, definindo dessa forma os três gêneros (WEITZMAN; SUMMERBELL 1995).
A partir do século XIX a Microbiologia destacou a importância de microrganismos nas
áreas da patologia humana e veterinária, assim como na indústria. O reconhecimento da
importância de fungos e bactérias levou ao aprimoramento de diferentes métodos para isolamento
e cultivo. A contínua obtenção de novas culturas e a necessidade da manutenção para fins de
estudos, conduziu à formação de vastas coleções de culturas de microrganismos em diversos
centros de pesquisas científicas e industriais (FIGUEIREDO, 2001).
A preservação de culturas puras é importante para estudos taxonômicos, identificação de
agentes etiológicos (LIMA; BORBA, 2001; GIRÃO et al., 2004; LIMA et al., 2004; BEZERRA et
al., 2006), atividades de ensino, testes de controle de qualidade de produtos e materiais, entre
outros (MELLO et al., 2003; CANHOS...). São importantes a conservação de aspectos
taxonômicos, fisiológicos, genéticos e a estabilidade metabólica de fungos preservados, pois os
mesmos podem ser utilizados como amostras de referência (MELLO et al., 2003; BAKER;
JEFFRIES, 2006; CANHOS...).
Fungos isolados, identificados e preservados de forma adequada, podem ser armazenados
por longos períodos (CANHOS...). Vários métodos de preservação de culturas de fungos têm sido
descritos, sendo que a escolha depende principalmente da disponibilidade da coleção de culturas e
da manutenção de características biológicas (SMITH; ONIONS, 1994; BUENO; GALLARDO,
1998; PANIZZO et al., 2005; NAKASONE et al...).
Os principais métodos empregados para manutenção de culturas de fungos incluem
repiques periódicos com cultivos a temperatura ambiente (SMITH; ONIONS, 1994; APARECIDO
et al., 2001; DESHMUKH, 2003; BORMAN et al., 2006; SOARES JUNIOR et al., 2007;

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
15
NAKASONE et al...) ou em baixas temperaturas (FIGUEIREDO, 2001; NAKASONE et al...), sob
óleo mineral (BORBA; RODRIGUES, 2000; FIGUEIREDO, 2001; LIMA et al., 2004; BORBA et
al., 2005; BEZERRA et al., 2006), em água destilada e esterilizada (método de Castellani, 1967)
(FIGUEIREDO, 2001; BORMAN et al., 2006), em solo (BAKERSPIEGEL, 1953; FERRETI-DE-
LIMA; MORAES-BORBA, 2001; BORMAN et al., 2006), em areia (FENG; HUA, 2005;
BORMAN et al., 2006), liofilização (FIGUEIREDO, 2001; NAKASONE et al...), congelamento
a -20 ou -80ºC (FIGUEIREDO, 2001; GINDRO; PEZET, 2001; BRILHANTE et al., 2004),
sílica-gel (FIGUEIREDO, 2001) e criopreservação em nitrogênio líquido (JONGS; DAVIS, 1979;
RYBNIKAR, 1995; RYAN et al., 2001; BORMAN et al., 2006).
A Micoteca do Departamento de Micologia, do Centro de Ciências Biológicas da
Universidade Federal de Pernambuco (UFPE), registrada no Commonwealth Mycological Institute
sob a sigla URM (University of Recife Mycology), contem atualmente cerca de 8.000 culturas de
fungos filamentosos e leveduras, mantidas em duplicata, identificadas em nível de espécie e
preservadas pelos métodos de óleo mineral, liofilização e de Castellani (MICOTECA URM...).
Para preservação de dermatófitos, os métodos mais utilizados são os de óleo mineral
(SCHONBORN,1989; DESHMUKH, 2003; BRILHANTE et al., 2004) e/ou de Castellani
(DESHMUKH, 2003; BORMAN et.al., 2006; NAKASONE et al...). A indicação para utilização
do óleo mineral se faz para fungos filamentosos ou com dificuldade para esporulação. Além disso,
o método é recomendado para coleções localizadas nos trópicos, devido as intercorrências
causadas por ácaros. Em relação ao método de Castellani, o objetivo é evitar o pleomorfismo.
Ambos mantêm a viabilidade da cultura por períodos prolongados (FIGUEIREDO, 2001).
Além da natural variabilidade fenotípica de dermatófitos (REBELL; TAPLIN, 1979;
LACAZ et al., 2002), as amostras preservadas podem sofrer alterações, sendo que as mais
observadas são diminuição da esporulação e da pigmentação, como também notável produção de
micélio aéreo de textura algodonosa (BRILHANTE et al., 2004; BORMAN et al., 2006).
Esses fungos apresentam características taxonômicas, antigênicas e patógenas similares,
mas com diferenças nutricionais e enzimáticas (LACAZ et al., 2002).
Os dermatófitos produzem várias enzimas que estão significantemente envolvidas na
obtenção de nutrientes para crescimento do fungo, especialmente na fase micelial (WEITZMAN;
SUMMERBELL, 1995; KITAJMA, 2000).
Enzimas como proteases, queratinases, fosfolipases e lipases são importantes na
patogencidade e podem ser secretadas para o meio de cultura durante o crescimento no substrato
(KURNET; KASAFÍREK, 1988; PAPINI; MANCIANTI, 1995; TRABULSI et al., 1999;
ABDEL-RAHMAN, 2000). Por isso, uso de meio de cultura sólido permite detecção da atividade

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
16
enzimática e vem sendo utilizada por vários autores (HANKIN; ANAGNOSTAKIS, 1975;
PRICE; CAWSON, 1977; PRICE et al., 1982; WAWRZKIEWICK et al., 1991; AOKI et al.,
1994; CORDEIRO NETO, 1997; MUHSIN et al., 1997; FRIEDRICH et al.,1999; DOS SANTOS
et al., 2001; LACAZ et al., 2002; CARVALHO, 2003).
Urease foi a primeira enzima isolada e é utilizada para diferenciação entre Trichophyton
mentagrophytes e T. rubrum (LACAZ et al., 2002).
Proteases, principalmente queratinases, são bastante estudadas e estão envolvidas em
infecções por dermatófitos (WEITZMAN; SUMMERBELL, 1995; SIMPANYA; BAXTER, 1996;
MAIA et al., 2001; ABDEL-RAHMAN 2002). Entretanto outros autores não reconhecem a
participação dessas enzimas no processo de infecções fúngicas, pois alguns fungos não produtores
de tais enzimas foram isolados de dermatomicoses, enquanto outros que a produzem, não causam a
doença (ABDEL-GAWAD, 1997; LACAZ et al., 2002; MUHSIN; HADI, 2002).
Queratinases apresentam habilidade para digerir queratina pela quebra das ligações
dissulfeto. Essas enzimas já foram detectadas de várias espécies de dermatófitos que utilizam a
proteína como fonte de carbono e nitrogênio para crescimento (DESHMUKH; AGRAVAL, 1982;
WAWRZKIEWICZ et al., 1991; LOPEZ-MARTINEZ et al., 1994; AUBAID; MUHSIN, 1998;
MIGNON et al., 1998; VIANI et al., 2001; ABDEL-RAHMAN, 2002).
Colagenases são bastante utilizadas no debridamento químico do colágeno, pela
decomposição das fibras dessa proteína com exclusiva capacidade (VICTORIA...). Essas enzimas
já foram detectadas de espécies de dermatófitos pertencentes aos três gêneros (RIPPON;
GARBER, 1969; LOPEZ-MARTINEZ, 1994).
As lipases auxiliam os dermatófitos a superarem a ação de ácidos graxos fungistáticos
presentes na pele (PAPINI; MANCIANTI, 1995; TRABULSI et al., 1999) e as fosfolipases podem
causar danos à membrana da célula hospedeira pela degradação de fosfolipídeos que são
freqüentemente encontrados fazendo parte da membrana celular (LEHNINGER, 1993; HANEL et
al., 1995; IBRAHIM et al., 1995; STRYER, 2001). Essas enzimas também já foram constatadas
em de amostras de dermatófitos (DAS; BANERJEE, 1977; MAIA et al., 2001).
Durante os últimos anos, vem sendo citado o aumento de infecções fúngicas inclusive de
dermatofitoses, devido ao aumento de casos e da sobrevida de pessoas com doenças
imunossupressoras, tais como nos casos de transplante, câncer e HIV/AIDS (TSANG et al., 1996;
PORRO et al., 1997; SQUEO et al., 1998; GARCÍA et al., 2003; FERNÁNDEZ-TORRES et al.,
2002, FERNÁNDEZ-TORRES et al., 2003a; BERGOLD; GEORGIADIS 2004). Nesses casos, a
doença deixa de ser superficial e atinge camadas mais profundas da derme causando lesões
granulomatosas (ESTUDO...).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
17
Geralmente, as micoses superficiais respondem bem aos antifúngicos de aplicação tópica,
entretanto a terapia local pode ser insuficiente em lesões extensas, profundas, em tinea unguium e
tinea capitis (FERNÁNDEZ-TORRES et al., 2001).
Em infecções mais brandas, o tratamento com antifúngico tópico tem como base o uso de
derivados imidazólicos, dos quais, clotrimazol, oxiconazol, miconazol, isoconazol, cetoconazol,
bifonazol, flutrimazol, erbeconazol são comumentes utilizados (FERNÁNDEZ-TORRES et al.,
2003a). Outros antifúngicos de uso tópico como terbinafina, ciclopirox, olamina,
amorolfina, butenafina também são bastante mencionados (FREEDBERG et al., 2003). Na prática
médica-clínica, para o tratamento de infecções crônicas ou severas, terbinafina, itraconazol e
fluconazol são mais utilizadas (FERNÁNDEZ-TORRES, et al., 2001, FERNÁNDEZ-TORRES et
al., 2003a).
Entretanto, essa micose pode ser resistente a terapia e a resposta ao tratamento pode não ser
adequada (BARCHIESI, et al., 2001; FAVRE et al., 2004). Além disso, sabe-se que o tratamento
prolongado pode gerar efeitos tóxicos ao organismo, visto que os antimicóticos também atuam
sobre outras células (GIORDANI et al., 2001), o que torna necessário o desenvolvimento de novas
estratégias terapêuticas (FOSTEL; LARTEY, 2000; TATSUMI et al., 2002; GARCÍA et al., 2003;
FERNÁNDEZ-TORRES et al., 2006).
Atenção especial tem se voltado para derivados naturais, com base no conhecimento da
produção de compostos antifúngicos pela natureza (WOJTASZEK, 1997; GURGEL et al., 2005).
Com o passar do tempo, produtos naturais como própolis estão sendo cada vez mais utilizados na
medicina (ACKERMANN, 1991; PARK et al., 1998a).
Própolis é um material resinoso de coloração e consistência variada, de composição
química complexa e elaborado por abelhas a partir da coleta de exsudatos de diversas partes das
plantas (BURDOCK, 1998; PARK et al., 1998a; BANKOVA et al., 2002; CASTALDO;
CAPASSO, 2002; BANKOVA, 2005a, BANKOVA, 2005b; FERNANDES JUNIOR et al., 2006).
É utilizada pelas abelhas para proteger a colméia de insetos e microrganismos invasores,
pelo emprego de uma fina camada que reveste tanto a entrada como o interior da colméia.
Também é utilizada para selar rachaduras, manter asséptico o ambiente interno da colméia,
preparar locais assépticos para a postura da abelha rainha e embalsamar invasores (BURDOCK,
1998; KUJUMGIEV et al., 1999; BANKOVA et al., 2000; SAWAYA et al., 2002; KOC et al.,
2005; FUNARI; FERRO, 2006).
A própolis verde do Brasil é nativa da região central do país mas pode ser encontrada em
outras regiões (KUMAZAWA et al., 2003). É elaborada por abelhas Apis mellifera a partir da
coleta de exsudatos de Baccharis dracunculifolia, comumente conhecida como alecrim-do-campo

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
18
ou vassourinha (PARK et al., PARK et al., 2002a, PARK et al., 2002c; ALENCAR et al., 2005a;
BANKOVA, 2005a, BANKOVA, 2005b) (Figura 1A).
A própolis vermelha foi recentemente encontrada em colméias localizadas no caule de
arbustos de manguezais como também ao longo da costa dos rios e mar dos estados da Paraíba,
Pernambuco, Alagoas, Sergipe e Bahia (Brasil). Foi observado que as Apis mellifera coletavam o
exsudato vermelho da superfície e dos buracos no tronco de Dalbergia ecastophyllum conhecida
como rabo-de-bugio, assumindo ser essa é a origem botânica da própolis vermelha (TRUSHEVA
et al., 2006; DAUGSCH et al. 2007) (Figura 1B).
Própolis apresenta diversas propriedades biológicas como antifúngica, antimicrobiana,
antiviral, antiinflamatória, antitumoral, cicatrizante, antioxidante, imunoestimulatória,
hepatoprotetora, entre outras (PARK et al., 1998a; PARK et al., 1998b; BANKOVA et al., 2000;
KOO et al., 2000; MARCUCCI et al., 2001; MURAD et al., 2002; KUMAZAWA et al., 2004;
CUSHNIE; LAMB, 2005; BANKOVA, 2005a; BANKOVA, 2005b). Por isso, vem sendo
utilizada pelo homem e a partir da década de 60 tornou-se objeto de intensos estudos
farmacológicos e químicos (BURDOCK, 1998; BANKOVA et al., 2000; CASTALDO;
CAPASSO, 2002; BANKOVA, 2005a; BANKOVA, 2005b).
A ação antifúngica e a antibacteriana estão entre as mais estudadas (MARCUCCI, 1995;
BURDOCK, 1998). A ação antifúngica da própolis já foi verificada em dermatófitos dos gêneros
Microsporum e Trichophyton (DOBROWOLSKI et al., 1991; SOARES; CURY, 2001; KOC et
al., 2005), em leveduras do gênero Candida (KUJUMGIEV et al., 1999; OTA et al., 2001;
SAWAYA et al., 2002; SANTOS et al., 2005), em Paracoccidioides brasiliensis (MURAD et al.,
2002) Cryptococcus neoformans (FERNANDES et al., 2007) e em outros fungos filamentosos
como espécies de Aspergillus, Penicillium e Cladosporium (ZHENG et al., 1996; AFOLAYAN;
MEYER, 1997). A combinação de drogas antifúngicas com própolis aumenta ainda mais a
atividade sobre os fungos (HEGAZI et al., 2000; SOARES; CURY, 2001).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
19
Figura 1A. Coleta de exsudato resinoso verde por abelha Apis mellifera. Fonte:http://www.bioessens.com/images/propolis.jpg Figura 1B. Coleta de exsudato resinoso vermelho por abelha Apis mellifera. Foto gentilmente cedida pela Pharma Néctar®.
A
B

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
20
Além da própolis, lectinas são investigadas pela participação em diversas atividades
biológicas como antiproliferativa (WANG et al., 1995; WONG; NG, 2003), antitumoral (WANG
et al., 1996; ABDULLAEV; MEJIA, 1997; WANG et al., 2000; NGAI; NG, 2004),
imunomodulatória (WANG et al., 1996; SHE et al., 1998; WANG; NG, 2002; RUBINSTEIN et
al., 2004), estimulação de macrófagos, acumulação de leucócitos (RODRIGUEZ et al., 1992),
indução da produção de interferon gama por linfócitos humanos (BARRAL-NETTO et al.,
1992), liberação de histamina (GOMES et al., 1994), efeitos edematogênicos e migração celular
(BENTO et al., 1993), antifúngica (CIOPRAGA et al., 1999; YE et al., 2001; DAMICO et al.,
2003; TRINDADE et al., 2006), antibacteriana (AYOUBA et al., 1994), anti HIV (PEUMANS;
VAN DAME, 1995; OOI et al., 2000; YE et al., 2001; WONG; NG, 2003; BARRIENTOS;
GRONENBORN, 2005; WONG; NG, 2005), antiinseto (PEUMANS; VAN DAME, 1995),
armazenamento e transporte de carboidratos e proteínas, armazenamento de nitrogênio, defesa da
planta (PEUMANS; VAN DAME, 1995; RÜDIGER, 1997; NOMURA et al., 1998), entre outras.
Lectinas são proteínas ou glicoproteínas não pertencentes ao sistema imunológico, que se
ligam especificamente e reversivelmente a mono e oligossacarídeos livres ou na forma de
glicoconjugados (glicoproteína e glicolipídeo). Dessa forma, combinam-se com moléculas e
estruturas biológicas que contem esses açucares sem alterar a estrutura covalente das ligações
glicosídicas nos sítios de ligação (MOREIRA et al., 1996; MOREIRA et al., 1997; CAVADA et
al., 1998, LIS; SHARON 1998; DAMICO et al., 2003; CHEN et al., 2005; CHUMKHUNTHOD
et al., 2006; WONG et al., 2006). Promovem reações celulares como o reconhecimento celular e
comunicação entre as células (DE SOL et al., 2005). Algumas são metaloproteínas, necessitando
de Ca2+, Mn2+ e Mg2+ para completo funcionamento (CAVADA et al., 1998).
São encontradas em ampla variedade em espécies de animais, plantas, fungos, bactérias e
vírus (GRIFFIN, 1993; PEUMANS; VAN DAMME, 1995; WANG et al., 1996; LIS; SHARON,
1998; CAVADA et al., 1998; NGAI; NG, 2004; XIA; NG, 2005; CHUMKHUNTHOD et al.,
2006), entretanto estão presentes em maior quantidade em grãos de leguminosas e gramíneas
(MOREIRA et al., 1996; RAMOS et al., 1996; CAVADA et al., 1998; INA et al., 2005).
Canavalia brasiliensis, Dioclea violacea e D. rostrata são leguminosas de onde são
extraídas as lectinas ConBr, DVioL e DRL, respectivamente (CAVADA et al., 1996; BARBOSA
et al., 2001), as quais têm afinidade por glicose e manose (CAVADA et al.,. 1993; CAVADA et
al., 1996). Essas lectinas já demonstraram indução da produção de histamina em ratos (GOMES et
al., 1994), produção de óxido nítrico (ANDRADE et al., 1999), além do efeito protetor in vivo
contra infecção por Leishmania amazonensis em camundongos BALB/c (BARRAL-NETTO et

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
21
al., 1996), estimulação linfocitária em humanos (BARRAL-NETTO et al., 1992), estimulação da
produção de macrófagos e linfócitos em camundongos C3H/HeJ (RODRIGUEZ et al., 1992).
Lonchocarpus sericeus é uma leguminosa de onde é extraída a lectina LSL (NAPIMONGA
et al., 2007) com afinidade por N-acetilglicosamina, a qual já demonstrou atividade
antiinflamatória em casos de peritonite em ratos, notável diminuição de bactérias que
colonizavam essa região peritoneal (ALENCAR et al., 2005c) e inibição da migração de
neutrófilos em processos inflamatórios (NAPIMONGA et al., 2007).
Lectinas com propriedades antifúngicas já foram isoladas de sementes de leguminosa (YE
et al., 2001) de frutas (FREIRE et al., 2002; DAMICO et al., 2003; TRINDADE et al., 2006), de
fungos (VIARD et al., 1993; CHUMKHUNTHOD et al., 2006), de urtiga (BROCKAERT et al.,
1989), de batata (GOZIA et al., 1993), de germe de trigo (CIOPRAGA et al., 1999) e de erva
chinesa (YAN et al., 2005).
Entretanto, algumas lectinas de leguminosas não apresentam essa atividade (YE; NG,
2001; NG et al., 2002; WONG; NG, 2003; WONG et al., 2005; WONG et al., 2006) e outras
apresentaram efeito ativador da cinética do crescimento do fungo em cultivo (VIARD et al., 1993;
PÉREZ-SANTIAGO et al., 2000; HERNÁNDEZ et al., 2003).
2. REVISÃO DA LITERATURA
2.1. DERMATÓFITOS
Dermatófitos são fungos que tem afinidade pela queratina da pele, pêlo e unha. Apresentam
ampla distribuição geográfica mas algumas espécies podem prevalecer em determinadas regiões, o
que está associado a variáveis como a adaptação do fungo ao ecossistema, migrações e/ou viagens,
condições sócio-econômicas e o grau de higiene das pessoas (SOYINKA, 1978; AZULAY, 1989;
AL-FOUZAN et al., 1993; OMIDYNIA et al., 1996; ASTE et al., 1997; ALI-SHTAYEH et al.,
1998; SIDRIM; ROCHA, 2004).
Esses fungos são agentes etiológicos de dermatofitoses que são comuns e endêmicas na
América Latina (WANK et al., 1991). A instalação da infecção por dermatófito inicia-se pela
inoculação de artrosporo depositado sobre a pele, favorecido por lesão cutânea ou escoriação pré-
existente (SIDRIM; ROCHA, 2004) e notável habilidade enzimática para degradar a queratina
(ABDEL-RAHMAN, 2000; SIMPANYA, 2000; MACÊDO et al., 2005).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
22
As espécies prevalentes no nordeste do Brasil são Microsporum canis, M. gypseum,
Trichophyton mentagrophytes, T. tonsurans, T. rubrum e Epidermophyton floccosum, com
destaque para M.canis, T. tonsurans e T. rubrum (COSTA et al., 1991; SIDRIM; ROCHA, 2004).
Cerca de 23 a 70% dos casos de dermatofitoses podem ser promovidos por M. canis e T.
tonsurans, juntos (BRILHANTE et al., 2000; FERNANDES et al., 2001; ABDEL-RAHMAN,
2002; DIAS et al., 2003; BARROS, 2004).
A espécie T. rubrum é antropofílica e de distribuição mundial. É responsável por 70% dos
casos de dermatofitoses no mundo (APODACA; MCKERROW, 1990; SAMDANI et al., 1995;
COSTA et al., 2002; ESQUENAZI et al., 2004; SANTOS; HAMDAN, 2005) e por 50% no
nordeste brasileiro (BRILHANTE et al., 2000; DOS SANTOS et al., 2001; PONTES et al., 2002;
BARROS, 2004). É citada como agente etiológico predominante de dermatofitoses de pé,
tornozelo, regiões plantar e palmar no Reino Unido, como também de onicomicose nos Estados
Unidos, Europa (SAMDANI et al., 1995; VENKATESAN et al., 2007) e Brasil (LOPES et al.,
1994; AQUINO et al., 2007).
De acordo com Weitzman e Summerbell (1995) e Lacaz et al. (2002) a História da
Micologia Médica teve início em meados do século XIX a partir de colaborações de três
pesquisadores europeus: Robert Remak, Johann Schönlein e David Gruby. As primeiras
observações de estruturas microscópicas, foram de lesões de favo (tinea capitis), feitas por Robert
Remak em 1835, o qual não reconheceu as estruturas fúngicas e creditou esse reconhecimento a
Schönlein, que descreveu a natureza micótica em 1839. Posteriormente, Remak estabeleceu essa
micose como infecciosa, a partir do cultivo do agente etiológico em fatias de maçã, denominando
de Achorion schoenleinii. Valiosos achados sobre dermatomicologia também foram escritos por
David Gruby entre 1841 e 1844, através de comunicados enviados a French Academy of Science e
publicações nesse mesmo período. Independente dos achados de Remak e Schönlein, Gruby
descreveu o agente etiológico do favo, o parasitismo ectotrix (ao redor do pêlo), os aspectos
clínicos da lesão e estabeleceu a natureza contagiosa dessa doença. Em 1843 denominou o agente
etiológico de Microsporum devido ao pequeno tamanho dos esporos ao redor do pêlo e
posteriormente identificou como M. audouinii.
O gênero Trichophyton foi observado pela primeira vez por Gruby em 1844, a partir de
observações de estruturas fúngicas em amostras clínicas de pêlos e escamas epidérmicas do couro
cabeludo, entretanto esse gênero não foi nomeado. No ano seguinte, Malmsten consultando as
informações de Gruby para publicar sobre aspectos clínicos de lesão de couro cabeludo, observou
que as estruturas fúngicas por ele observadas, eram semelhantes às observadas por Gruby.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
23
Malmsten denominou o agente etiológico de Trichophyton pela afinidade desse fungo por pêlos. A
espécie observada foi T. tonsurans (GEORG, 1956; WEITZMAN; SUMMERBELL, 1995).
O nome Epidermidophyton foi proposto por Langeron em 1879, para designar agente
etiológico de micose de pele. Em 1907, Raymond Sabouraud propôs o nome Epidermophyton pela
constatação de estruturas fúngicas em escamas epidérmicas (McGINNS et al., 1981). Esse fungo é
antropofílico e de um modo geral pode ser agente etiológico de tinea curis, tinea pedis e tinea
corporis (LACAZ et al., 2002; AQUINO et al., 2007).
Em 1910, Sabouraud publicou uma obra clássica, “Les Teignes”, propondo classificação
para dermatófitos a partir de achados microscópicos de parasitismo no pêlo e em escamas
epidérmicas, aspectos clínicos da lesão, métodos de cultivo e observações macroscópicas. Dessa
forma, os dermatótifos foram classificados em quatro gêneros: Achorion, Microsporum,
Trichophyton e Epidermophyton. A base do meio para cultivo (dextrose e ágar) desenvolvido na
época, é utilizada até hoje, embora a composição tenha sido modificada e em homenagem, é
denominado de ágar Sabouraud (WEITZMAN; SUMMERBELL, 1995; LACAZ et al., 2002).
Emmons (1934) propôs uma nova classificação para os dermatófitos, modificando a de
Sabouraud. Estabeleceu como critério a morfologia de esporos e estruturas de ornamentação, no
qual extinguiu o gênero Achorion, proposto por Sabouraud, inserindo-o no gênero Trichophyton.
Essa classificação é atualmente mantida e composta por três gêneros: Microsporum, Trichophyton
e Epidermophyton (LACAZ et al., 2002).
As principais características que distinguem os gêneros Microsporum, Trichophyton e
Epidermophyton são: presença de macroaleurias fusiformes; presença abundante de microaleurias
e presença de macroaleurias em forma de raquete, respectivamente (REBELL; TAPLIN, 1979;
LARONE, 1996; LACAZ et al., 1998; LACAZ et al., 2002).
Cole e Samson (1979) demonstraram que a ontogenia holotálica de esporor de
Microsporum e Trichophyton é a mesma. De Hoog et al. (1998) estudando a taxonomia de fungos
de interesse médico e a filogenia molecular, verificaram que os gêneros Microsporum e
Trichophyton são parafiléticos e o Epidermophyton é polifilético.
Apesar de ser um grupo extenso com características taxonômicas, antigênicas e patógenas
similares, apresentam diferenças nutricionais e enzimáticas (LACAZ et al., 2002). O pleomorfismo
comum em dermatófitos pode dificultar a manifestação de características taxonômicas, refletindo
em dificuldade para identificação (RIPPON, 1990; ELEWSKI, 1992; FISHER; COOK, 2001;
LACAZ et al., 2002; SIDRIM; ROCHA, 2004). Essa alteração genética pode acontecer quando há
repiques sucessivos do fungo para meio artificial ou quando é submetido à condição de

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
24
preservação, na qual há uma diminuição do metabolismo (KRAMER; MIX, 1957; DESHMUKH,
2003).
2.2. PRESERVAÇÃO
O contínuo isolamento de fungos e a necessidade da manutenção para estudos de
taxonomia, patologia ou industriais, conduzem à formação de vastas coleções de culturas de
fungos preservados de tal forma que permita principalmente a conservação de características
taxonômicas, genéticas e atividades biológicas (FIGUEIREDO, 2001; GIRÃO et al., 2004;
DIOGO et al., 2005).
Coleções de culturas de fungos são centros de excelência que atuam como unidades de
conservação e exploração principalmente de recursos taxonômicos, genéticos e metabólicos desses
organismos, cujo principal objetivo é adquirir fungos relevantes para estudos científicos e de
aplicações, oferecendo serviços fundamentais para a comunidade científica e tecnológica
(CANHOS...).
O principal objetivo da preservação de culturas é manter as mesmas em estado viável, sem
mudanças taxonômicas, fisiológicas e genéticas até que sejam requeridas para pesquisas. Para
garantir a completa viabilidade e estabilidade, são utilizados métodos que têm o princípio de
diminuir o metabolismo celular dos fungos (SMITH; ONIONS, 1994; NEUFELD; SARQUIS,
2003).
Os principais métodos empregados para manutenção de culturas de fungos incluem
repiques periódicos com cultivos a temperatura ambiente (SMITH; ONIONS, 1994; APARECIDO
et al., 2001; DESHMUKH, 2003; BORMAN et al., 2006; SOARES JUNIOR et al., 2007;
NAKASONE et al...) ou em baixas temperaturas (FIGUEIREDO, 2001; NAKASONE et al...), sob
óleo mineral (BORBA; RODRIGUES, 2000; FIGUEIREDO, 2001; LIMA et al., 2004; BORBA et
al., 2005; BEZERRA et al., 2006), em água destilada e esterilizada (método de Castellani, 1967)
(FIGUEIREDO, 2001; BORMAN et al., 2006), em solo (BAKERSPIEGEL, 1953; FERRETI-DE-
LIMA; MORAES-BORBA, 2001; BORMAN et al., 2006), em areia (FENG; HUA, 2005;
BORMAN et al., 2006), liofilização (FIGUEIREDO, 2001; NAKASONE et al...), congelamento a
-20 ou -80ºC (FIGUEIREDO, 2001; GINDRO; PEZET, 2001; BRILHANTE et al., 2004), sílica-
gel (FIGUEIREDO, 2001) e criopreservação em nitrogênio líquido (JONGS; DAVIS, 1979;
RYBNIKAR, 1995; RYAN et al., 2001; BORMAN et al., 2006).
De acordo com Figueiredo (2001) as primeiras coleções mundiais eram mantidas por
repicagens constantes. Além de ser um trabalho exaustivo, problemas como perda de viabilidade,

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
25
alterações fisiológicas de aspecto e pigmentação, decréscimo ou perda da capacidade de
esporulação e da patogenicidade apareciam constantemente. Assim, para preservação por longos
períodos, tornou-se necessário desenvolver outros métodos que mantivessem as culturas viáveis e
menos sujeitais a variações.
A Coleção de Culturas Micoteca - University of Recife Mycology (URM) do
Departamento de Micologia, do Centro de Ciências Biológicas da Universidade Federal de
Pernambuco, foi fundada em 1954 pelo Professor Augusto Chaves Batista, utilizando óleo mineral
como método de preservação (SHERF, 1943). Atualmente, o acervo contém aproximadamente
8.000 culturas de fungos, sendo cerca de 1.300 de leveduras e 6.700 de fungos filamentosos, todas
identificadas em nível de espécies e mantidas em duplicata em cada método de preservação. Todos
os fungos dessa Coleção são mantidos em óleo mineral e por liofilização (RAPER;
ALEXANDER, 1945). O método de Castellani é utilizado apenas para Zygomycetes e para as
espécies patógenas ao homem e vegetais (MICOTECA URM...).
Para preservação de dermatófitos, de modo geral, os métodos mais utilizados são os de óleo
mineral (SCHONBORN, 1989; DESHMUKH, 2003; BRILHANTE et al., 2004) e/ou o de
Castellani (DESHMUKH, 2003; BORMAN et al., 2006; NAKASONE et al...). A indicação para
utilização do óleo mineral se faz para fungos filamentosos ou com dificuldade para esporulação.
Além disso, o método é recomendado para coleções localizadas nos trópicos, devido à
intercorrências causadas por ácaros. O método de Castellani tem o objetivo de evitar o
pleomorfismo. Ambos mantêm a viabilidade da cultura por períodos prolongados (FIGUEIREDO,
2001).
De acordo com Fennel (1960), o método de óleo mineral foi inicialmente utilizado por
Lomiére e Chevrotier para aumentar a longevidade e reduzir a variabilidade de cultura de
Gonococcus, passando depois a ser aplicado também para fungos. A experiência demonstrou que
pelo emprego desse método, as repicagens da maioria das espécies pertencentes à Coelomycetes e
alguns basidiomicetos podem ser realizadas em intervalos de 12 a 24 meses, sem problemas.
Para preservação sob óleo mineral, é necessário que culturas fúngicas jovens desenvolvidas
pelo método clássico em tubos contendo meio de cultura solidificado e inclinado a 30º com a
horizontal, sejam submersas em uma camada de óleo mineral esterilizado. Previne-se desta forma
desidratação do meio de cultura, reduzindo a atividade metabólica e o crescimento, por reduzir a
tensão de oxigênio. A reativação é feita pela remoção de um fragmento da cultura e transferência
para meio líquido ou sólido (SMITH; ONIONS, 1994; FIGUEIREDO, 2001; NAKASONE et
al...).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
26
Foi verificado que culturas submersas sob 1 cm de óleo mineral têm o consumo de
oxigênio reduzido cerca de 10% em poucas horas, e que a redução da atividade metabólica é
evidenciada pelo retardamento do crescimento e da esporulação. Camada de óleo superior a 1 cm
sobre a superfície de crescimento é indesejável, pois diminui o tempo de viabilidade das culturas.
Há evidências de que o óleo mineral em condições de total exaustão do oxigênio torna-se tóxico
para a maioria dos microrganismos (FIGUEIREDO, 2001).
Há relatos da aplicação desse método em centros de preservação de fungos como o New
York Botanical Garden (New York, U.S.A.), American Type Culture Collection (Maryland,
U.S.A.), Comunicable Disease Center (Georgia, U.S.A.), Lilly Research Laboratories (Indiana
U.S.A.), Harvard University (Massach. U.S.A.), Agricultural Research Service (Wash U.S.A.),
Commonwealth Mycological Institute (Kew, Inglaterra) (FIGUEIREDO, 2001).
Schonborn (1989) avaliaram amostras de dermatófitos preservados em óleo mineral e
verificaram viabilidade de 50% das culturas preservadas no período de 2 a 15 anos.
Smith e Onions (1994) citam a boa preservação de espécies de Aspergillus e Penicillium
por 40 anos, sob óleo mineral.
Borba e Rodrigues (2000) constataram viabilidade e perda de capacidade de esporulação
em 20 amostras de Coelomycetes, entre 34 analisadas após 50 anos de preservação sob óleo
mineral. Mas, o grau de variações está mais relacionado a espécie do fungo do que com a
preservação.
Deshmukh (2003) constataram a manutenção de características taxonômicas de
Microsporum e Trichophyton preservados em óleo mineral por sete anos e em água destilada e
esterilizada entre sete a dez anos.
Brilhante et al. (2004) constataram pleomorfismo com desenvolvimento de hifas estéreis
em amostras de M. canis estocadas sob óleo mineral por nove meses.
Borman et al. (2006) constataram que M. canis, T. rubrum e T. tonsurans preservados em
água destilada esterilizada por um período de 2 meses a 21 anos, apresentaram aspectos de
pleomorfismo (micélio algodosono, redução da pigmentação e da esporulação) a partir do quinto
ano de preservação em mais de 20% do grupo analisado, com formação de micélio estéril e
dificuldade de esporulação. Dermatófitos criopreservados por até 2 anos, mantiveram
características taxonômicas típicas (BAKER; JEFFRIES et al., 2006).
Lima et al. (2004) detectaram que amostras preservadas sob óleo mineral de Blastomyces
dermatitidis e Paracoccidioides brasiliensis, apresentaram-se incapazes de completar o
dimorfismo após reativação. Borba et al. (2005) constataram alterações morfológicas em amostras
de P. brasiliensis estocadas sob óleo mineral.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
27
Braz (2007) demonstrou preservação de características morfológicas e capacidade de
produção enzimática de 31 amostras de Acremonium sp. preservadas sob óleo mineral na Coleção
de Culturas da Micoteca-URM, entre 2 e 43 anos.
A variedade dos métodos recomendados e descritos na literatura reflete a falta de um
método que seja abrangente e satisfatório para todos os fungos (FIGUEIREDO, 2001), pois a
estocagem pode promover alterações morfológicas, macroscópicas e/ou de componentes da célula
fúngica (BAKER; JEFFRIES, 2006; BORMAN et al., 2006; SOARES JÚNIOR et al., 2007).
2.3. ENZIMAS
Os dermatófitos produzem uma variedade de enzimas extracelulares como proteases,
queratinases, colagenases e lipases capazes de quebrar moléculas maiores existentes na pele e
anexos em moléculas menores para utilização (SKOREPOVÁ; HAUCK, 1987; PAPINI;
MANCIANTI, 1995; MUHSIN; SALIH, 2000; TAKASUKA, 2000; MAIA et al, 2001; ABDEL-
RAHMAN, 2002). A atividade enzimática pode variar de acordo com a espécie de dermatófito
(MUHSIN; SALIH, 2000) e as diferentes expressões enzimáticas podem estar relacionadas com o
aspecto ecológico (antropofílico, zoofílico e geofílico) uma vez que a adaptação paulatina do solo
para o homem e outros animais, envolveu também adequação nutricional (PHILPOT, 1977;
SIMPANYA; BAXTER, 1996, MUHSIN; SALIH, 2000).
Samdani et al. (1995) demonstraram variações da atividade proteolítica entre T.
mentagrophytes (zoofílico) e T. rubrum (antropofílico) isolados de tinea pedis e tinea unguium,
apresentando maior atividade as amostras de T. mentagrophytes.
Simpanya e Baxter (1996) constataram que amostras de M. canis e M. cookei apresentaram
diferentes expressões enzimáticas de proteases, sendo que as da primeira espécie apresentaram
atividade bem maior do que as da segunda. Para esses autores, essa diferença reflete o contraste
ecológico dessas duas espécies, visto que a primeira é zoofílica e a outra é geofílica. Entretanto,
Bruguera et al. (1997) não verificaram diferenças enzimáticas significativas para proteases, lipases
e fosfolipases entre amostras de M. canis e M. gypseum.
A variação interespecífica de proteases e lipases também foi constatada em amostras de M.
gypseum, T. mentagrophytes e E. floccosum. As duas primeiras espécies que são zoofílicas,
apresentaram elevada atividade de proteases e lipases, enquanto que na última foi observada baixa
atividade de proteases e não houve atividade lipásica (MUHSIN; SALIH 2000).
Amostras da mesma espécie também podem apresentar variações quanto a atividade
enzimática. Essa constatação foi observada entre amostras de T. tonsurans para queratinases,

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
28
colagenases e elastases (ABDEL-RAHMAN, 2000), de M. canis para proteases (MAIA et al.,
2001) e de T. rubrum para proteases, queratinases, fosfolipases, lipases e desoxiribonucleases
(DOS SANTOS et al., 2001).
Esse incrível sistema enzimático está direcionado principalmente para a degradação de
queratina, que é proteína estrutural localizada na pele e anexos. Essa escleroproteína contém
cadeias polipeptídicas α queratina (cadeias α) e β queratina (cadeias folhas β) que estão
fortemente unidas por ligações covalentes dissulfeto, mantendo a forma tridimensional da
molécula. Além dessas ligações, interações hidrofóbicas e ligações de hidrogênio estabilizam a
estrutura filamentosa da proteína (COULOMBE; OMARY, 2002). Por essa razão, é extremamente
resistente a degradação por enzimas proteolíticas como tripsina, pepsina e papaína. A alta
resistência a maioria dos microrganismos deve-se, principalmente as ligações dissulfeto, entretanto
os dermatófitos possuem um sistema enzimático capaz de quebrar essas ligações (SOUZA
JUNIOR et al., 1999).
Queratinases pertencem ao grupo das proteases que hidrolisam as ligações peptídicas nas
proteínas (SAID; PIETRO, 2002). Possui alta especificidade por queratina mas também podem
degradar substratos solúveis como caseína, soroalbumina bovina e gelatina, e insolúveis como
elastina, colágeno, lã, cabelo, unha, entre outros (FRIEDRICH; KER, 2003). Tem grande
capacidade para agir em substratos compactos como queratina, quando comparadas com outras
enzimas proteolíticas sendo assim distinguidas de outras proteases (ONIFADE et al., 1998).
Queratinases já foram detectadas de amostras de Microsporum (TAKIUCHI et al., 1984;
MIGNON et al., 1998), de Trichophyton (DESHMUKH; AGRAVAL, 1982; AUBAID; MUHSIN,
1997; ABDEL-RAHMAN, 2000) e de Epidermophyton floccosum (HOPSU-HAVU; TUNNELA,
1977; LOPEZ-MARTINEZ, 1994), os quais utilizam a proteína como fonte de carbono e
nitrogênio para crescimento (WAWRZKIEWICZ et al.,1991; VIANI et al., 2001; MACÊDO et al.,
2005).
O colágeno é uma proteína insolúvel estrutural encontrada na pele, ossos, ligamentos,
cartilagens e outras partes do organismo. Cada molécula de colágeno é formada pelo
entrelaçamento, em tríplice hélice, de três cadeias polipeptídicas chamadas cadeias alfa. É rica em
lisina, prolina, hidroxilisina e hidroxiprolina. Essa estrutura protéica justifica as propriedades
físicas e biológicas do colágeno: rigidez, solidez e estabilidade. Devido a estrutura tridimensional,
é bastante resistente a degradação por enzimas proteolíticas (VARGAS et al., 1997). Existem
cinco tipos de colágeno, sendo que os tipos I e V estão presentes na pele (STRYER, 2001;
CAMPBELL, 2003). A gelatina é uma proteína derivada da hidrólise parcial do colágeno, cujas
ligações moleculares são hidrolisadas, permitindo o seu rearranjo (PEREIRA, et al., 2006).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
29
Colagenases são enzimas proteolíticas capazes de degradar o colágeno, cuja proteína
apresenta alta especificidade por essa enzima. As colagenases de microrganismos possuem
capacidade para hidrolisar o colágeno nativo e desnaturado (gelatina). As proteases, de um modo
geral, não hidrolisam a tripla hélice do colágeno por não degradar essa estrutura (HONDA, 1998;
GOSHEV et al., 2005). Enquanto as colagenases degradam moléculas de colágeno nativo, as
gelatinases degradam principalmente o colágeno desnaturado e o colágeno tipo IV, que está
presente na membrana basal (STRYER, 2001; PEREIRA et al., 2006).
Pesquisas relacionadas com a habilidade de fungos em produzir enzimas extracelulares
como proteases, queratinases e lipases, permitem estabelecer relação com a patogenicidade
(MAGO, 1990).
Vários autores reconhecem o papel importante de proteases sintetisadas por dermatófitos
no processo de infecção do tecido do hospedeiro e desenvolvimento fúngico (MINOCHA et al.,
1972; SKOREPOVÁ; HAUCK, 1987; KURNET; KASAFÍREK, 1988; GRZYWNOWICZ et al.,
1989; SAMDANI et al., 1995; SIMPANYA; BAXTER, 1996; MAIA et al., 2001; ABDEL-
RAHMAN 2002). Além da invasão mecânica promovida pelo micélio no tecido do hospedeiro, a
habilidade que os dermatófitos têm de invadir e subseqüentemente se disseminar no estrato córneo
é governado em parte, por enzimas proteolíticas que contribuem de forma imprescindível
(SAMDANI et al., 1995; ABDEL-RAHMAN, 2000; MAIA et al., 2001).
De acordo Rippon e Garber (1969) a ação de proteases está envolvida na patogenia causada
por dermatófitos, intensificando a reação inflamatória pela indução da quimiotaxia de leucócitos
polimorfonuclear.
Minocha et al. (1972) verificaram a atividade proteolítica de T. rubrum, T. mentagrophytes
e E. floccosum na patogenia de lesões de pele. Para T. rubrum e E. floccosum o eritema intenso
não tem correlação com a atividade proteolítica e quanto mais curta é a duração da lesão, maior é a
atividade das enzimas relacionadas, pois isolados fúngicos de lesões cutâneas extensas,
apresentaram baixa atividade enzimática. Para T. mentagrophytes, quanto maior a atividade
enzimática, maior é a intensidade do eritema e menor é a duração da infecção. Concluíram que
essas enzimas são importantes para a penetração do fungo no tecido queratinizado e que
promovem vasodilatação, melhorando a permeabilidade capilar com intensificação da resposta
inflamatória, mas que a participação na patogenia varia, dependendo da espécie.
Meevootisom e Niederpruem (1979) verificaram que proteases extracelulares e micelial são
produzidas por T. rubrum e devem ter participação na invasão de tecidos do hospedeiro,
contribuindo para hipersensibilidade. Skorepová e Hauck (1987) verificaram que 57,5% de
amostras de T. rubrum isoladas de lesões pustulosas, apresentaram atividade proteolítica.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
30
Alguns fungos de interesse médico produzem proteases que podem atuar como fator de
virulência, devido ao envolvimento com a propagação do fungo no hospedeiro, causando
conseqüente invasão através da degradação de proteínas da pele, anexos ou mucosa (KWON-
CHUNG; BENNETT,1992).
Weitzman e Summerbell (1995) em trabalho de revisão sobre dermatófitos, reafirmaram o
envolvimento de enzimas nas infecções por fungos de interesse médico, em especial por
dermatófitos.
Okafor e Ada (2000) avaliaram a atividade queratinolítica de T. rubrum, T.
mentagrophytes, T. tonsurans, M. audouinii e M. gypseum utilizando cabelo humano como
substrato. Todas cresceram sobre o cabelo, mas apenas M. gypseum e T. mentagrophytes foram
capazes de causar danos estruturais e formar órgãos de perfuração. Macêdo et al. (2005)
constataram a capacidade de perfuração de pêlo in vitro em 94,3% das amostras de E. floccosum
analisadas, sugerindo que essa espécie possa ser agente etiológico de tinea capitis.
Abdel-Rahman (2000) avaliando a expressão de proteases extracelular de T. tonsurans
isolados de tinea capitis, infere que a ação dessas enzimas é importante para a infecção uma vez
que, a presença de estruturas fúngicas, apenas, sobre o tecido queratinizado do hospedeiro, não é
suficiente para desenvolvimento de dermatofitoses.
Atividades de colagenases já foram verificadas em amostras de T. schoenleinii (RIPPON;
GARBER, 1969) e de T. tonsurans (ABDEL-RAHMAN, 2000).
Abdel-Rahman (2002) correlacionou a atividade extracelular de colagenases e elastases de
amostras de T. tonsurans isolados de tinea capitis com a manifestação clínica da doença.
Constatou que quanto maior é a atividade enzimática, mais severa é a infecção e mais curto é o
período da doença. Por outro lado, quanto menor é a atividade dessas enzimas, maior é o período
de infecção e mais branda é a inflamação. É reforçada a participação virulenta e patogênica de
proteases para infecção e disseminação de fungos no estrato córneo. A correlação entre enzimas e
patogenicidade é explicada pela função antigênica enzimática: estimulam a resposta inflamatória e
devido a isso estimulam o organismo reação do hospedeiro infectado.
Macêdo et al. (2005) caracterizaram quanto a fatores de patogenicidade, amostras de E.
floccosum recém isoladas e preservadas na Coleção de Culturas Micoteca - URM e verificaram
que todas apresentavam atividade de proteases pela formação de halo ao redor da colônia.
Viani et al. (2007) avaliaram a atividade de proteases extracelular de M. canis isolados de
gatos com e sem lesão e constataram que a atividade de queratinases e elastases era elevada
(p<0.05) quando comparada aos animais sem lesão.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
31
Mas segundo Kurnet e Kasafírek (1988), é incerta a função precisa de proteases e
queratinases na virulência e patogenia de dermatófitos.
Para Lacaz et al. (2002) a ação dessas enzimas proteolíticas não é responsável por
infecções fúngicas. É citado que alguns fungos não produtores de tais enzimas foram isolados de
dermatomicoses, enquanto outros que a produzem, não causam a doença.
Chaves et al. (2003) trabalhando com espécies de Candida de isolados clínicos de
pacientes com AIDS e com espécie de Candida estocadas na Coleção de Cultura Micoteca - URM,
não detectaram atividade proteásica em meio sólido, em nenhum dos isolados.
Produção de proteases foi verificada em amostras de Chrysosporium tropicum, C.
keratinophilum (ABDEL-GAWAD, 1997) e Curvularia spp. (MUHSIN; SALIH, 2000).
Muhsin e Hadi (2002) constataram produção de queratinases e degradação de pêlos por
62% das amostras de Chrysosporium pannicola. Esses mesmos autores verificaram que amostras
de Aspergillus flavus apresentaram atividade dessa enzima em meio de cultura adicionado de pena
de galinha.
Braz (2007) verificou atividade proteásica em 23 amostras de Acremonium preservadas na
Coleção de Culturas Micoteca - URM e constatou essa atividade na caseína do leite em 10% das
amostras analisadas e na gelatina em 8%.
Atividade de queratinases também já foi constatada em amostras de Penicillium,
Alternaria, Scopulariopsis (RIPPON, 1990; MARCHISIO et al., 1991; KWON-CHUNG;
BENETTT, 1992; AGUT et al., 1995; GUGNANI, 2003) Bacillus e Actinomicetos (ASAHI et al.,
1985; SIESENOP; BOHM, 1995; GRADISAR et al., 2000).
Sabe-se que a pele é a primeira linha de defesa contra infecções por microrganismos
patógenos ao homem (TRUSWELL, 1987; ANDERSON et al., 1988) e que existem ácidos
graxos com ação fungistática que dificultam a infecção por dermatófitos, sendo esse um dos
mecanismos de defesa natural do organismo (NOBRE; VIEGAS, 1972; ZAROR et al., 1987;
MORIELLO; DE BOER, 1991; OGAWA et al., 1998; FERREIRA-NOZAWA, 2003). Tinea
capitis, por exemplo, é reconhecida como uma dermatofitose quase que exclusiva da infância
(LACAZ et al., 2002; HERNÁNDEZ et al., 2004), regredindo espontaneamente na puberdade
devido à produção, sob influência hormonal, de ácidos graxos com propriedade fungistática
(AZULAY, 1989; OGAWA et al., 1998). Além das proteases, as lipases extracelulares também
são citadas pela participação durante a infecção (OGAWA et al., 1998). Acredita-se que as lipases
auxiliam esses fungos a superar a ação dos ácidos graxos fungistático presentes na pele,
colaborando para a virulência nesse grupo de fungos (PAPINI; MANCIANTI, 1995, TRABULSI
et al., 1999).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
32
Nobre e Viegas et al. (1972) determinando atividade lipolítica de amostras de dermatófitos
recém isolados (M. canis, M gypseum, E. floccosum e T. mentagrophytes) e preservados (T.
tonsurans, T. rubrum, T. schoenleinii, T. violaceum e T. megnini), constataram que essa atividade
foi mais evidente em amostras recém isoladas (75 a 100%). Dentre as amostras deste grupo à
atividade foi maior nas de M. canis, E. floccosum e T. mentagrophytes. Todas as amostras
preservadas apresentaram atividade menor quando comparadas com as do outro grupo.
Papini e Mancianti (1995) não constataram atividade lipásica em M. canis, por outro lado,
essa atividade foi verificada nas espécies T. rubrum (DAS; BANERJEE, 1977; DOS SANTOS et
al., 2001), M. gypseum e T. mentagrophytes (MUHSIN; SALIH, 2000).
Costa et al. (1967) foram os primeiros a demonstrarem atividade fosfolipásica utilizando a
lecitina da gema de ovo. Fosfolipases são enzimas que degradam os fosfolipídeos, freqüentemente
encontrados fazendo parte da membrana celular de animais, plantas e microrganismos. Parecem
auxiliar a invasão de células hospedeiras por dermatófitos (DAS; BANERJEE, 1977).
Há relatos sobre a associação entre fosfolipases e fungos patogênicos (HANEL et al.,
1995). A produção dessa enzima por Candida albicans está relacionada com a virulência e
patogenicidade, pois tal enzima pode causar danos à membrana da célula do hospedeiro pela
degradação de fosfolipídeos e lisofosfolipídeos (HANEL et al., 1995; IBRAHIM et al., 1995). A
elevada produção de fosfolipases foi correlacionada com maior capacidade de aderência de C.
albicans, desenvolvimento da doença e elevada taxa de mortalidade em estudos com animais
(MAYSER et al., 1996). Chaves et al. (2003) constataram atividade fosfolipásica nessa espécie,
em 13 amostras preservadas e em quatro recém isoladas de pacientes com AIDS.
Dos Santos et al. (2001) mencionam a falta da atividade de fosfolipases em T. rubrum,
sugerindo a não utilização desse critério como marcador para diferenciação de biotipos. Entretanto
esse parâmetro foi utilizado por Maia et al (2001) para caracterização fenotípica de recém isolados
de M. canis.
Atividade fosfolipásica não foi verificada em amostras de E. floccosum recém isoladas e
preservadas (MACÊDO et al., 2005) como também em amostras preservadas de Acremonium
(BRAZ, 2007).
Hankin e Anagnostakis (1975) detectaram a produção de exoenzimas em meios sólidos e
determinaram as atividades pela hidrólise do substrato, através da formação de halo. A atividade
de proteases é normalmente detectada qualitativamente utilizando como substratos ágar acrescido
de caseína, hemoglobina, gelatina ou outras proteínas. A região de hidrólise, observada em torno
da colônia indica atividade enzimática (AUNSTRUP, 1974).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
33
Muhsin et al. (1997) constataram a atividade lipásica de dermatófitos e leveduras em meio
sólido.
É possível a determinação quantitativa da atividade fosfolipásica em grande número de
amostras de fungos, em meio de cultura sólido contendo lecitina de soja ou gema de ovo como
fonte fosfolipídica (PRICE; CAWSON, 1977).
O uso de meio sólido permite rápida detecção da atividade enzimática (HANKIN;
ANAGNOSTAKIS, 1975), pois durante o crescimento no substrato protéico, os dermatófitos
secretam enzimas para o meio de cultura (KURNET; KASAFIREK, 1988). Essa forma de verificar
a atividade enzimática vem sendo utilizada por outros autores (PRICE; CAWSON, 1977; PRICE
et al., 1982; WAWRZKIEWICK et al., 1991; AOKI et al., 1994; CORDEIRO NETO, 1997;
MUHSIN et al., 1997; FRIEDRICH et al.,1999; DOS SANTOS et al., 2001; LACAZ et al., 2002;
CARVALHO, 2003).
O desenvolvimento de colônias de dermatófitos em meios contendo um único substrato
como fonte nutritiva, demonstra capacidade de sintetizar enzimas para degradar o substrato e
utilizá-lo para o desenvolvimento fúngico (ONIFADE ET AL, 1998; OGAWA ET AL. 1998).
2.4. DROGAS ANTIFÚNGICAS E TESTE DE SUSCEPTIBILIDADE ANTIFÚNGICA
Várias drogas são citadas para o tratamento de dermatofitoses, o qual pode ser de aplicação
tópica e/ou oral (CARAZO et al., 1999; QUINDÓS, 2002; SECCO...). O tratamento tópico é
indicado nos casos de lesões pequenas e localizadas (LLAMBRICH; LECHA, 2002; KARACA;
KOÇ, 2004) cujas drogas mais utilizados pertencem a classe dos derivados imidazólicos
(clotrimazol, oxiconazol, miconazol, isoconazol, cetoconazol, bifonazol, flutrimazol, erbeconazol)
(FERNÁNDEZ-TORRES et al., 2003a). Outros antifúngicos de uso tópico como terbinafina,
ciclopirox, olamina, amorolfina, butenafina também são bastante mencionados (FREEDBERG et
al., 2003).
Entretanto, sabe-se que a escolha da forma ideal depende, dentre outros aspectos, da
localização e extensão da lesão como também da espécie envolvida (FERNANDEZ-TORRES et
al., 2001; FERNANDEZ-TORRES et al., 2002). Às vezes é necessária a administração da droga
por via oral, principalmente em casos de tinea unguium e tinea capitis (FERNANDEZ-TORRES et
al., 2001; PUJOL et al., 2002; FAVRE et al., 2003; FERNANDEZ-TORRES et al., 2003a;
SANTOS et al., 2006). O tratamento por via oral é realizado em casos de lesões extensas ou
disseminadas, utilizando-se principalmente os derivados imidazólicos (fluconazol, itraconazol,
cetoconazol) e alilaminas (terbinafina) (FEEDBERG et al., 2003).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
34
Entre essas, itraconazol e terbinafina são bastante mencionadas principalmente por inibirem
o crescimento do fungo em baixas concentrações (FERNÁNDEZ-TORRES et al., 2001; FAVRE
et al., 2003; BARROS et al., 2006) e apresentarem atividade fungicida (HAZEN, 1998;
FERNÁNDEZ-TORRES et al., 2002; FERNANDEZ-TORRES et al., 2003a; SANTOS;
HAMDAN, 2005). Favre et al. (2003) citam que entre os antifúngicos de uso sistêmico, terbinafina
é bastante eficaz por apresentar atividades fungistática e fungicida em baixas concentrações, como
por exemplo, em 0.06µg/mL e 0.07µg/mL, respectivamente. Fluconazol é citada como a menos
indicada, pois a inibição de 90% do crescimento fúngico é obtida em elevadas concentrações,
como por exemplo, em 64µg/mL para T. mentagrophytes e T. rubrum (SANTOS et al., 2006) e
32µg/mL para outras espécies (FERNÁNDEZ-TORRES et al., 2001; PEREA et al., 2001; FAVRE
et al., 2003; BARROS et al., 2006; SANTOS et al., 2006).
Itraconazol e terbinafina inibem o crescimento fúngico alterando síntese de ergosterol em
nível de membrana celular. Itraconazol é um triazol com três átomos de nitrogênio no anel azólico
(JAIN; SHEGAL, 2001) que inibe a formação de blocos de ergosterol por interferir na demetilação
do lanosterol (DE BACKER et al., 1998). Há inibição da enzima esterol-14-α-desmetilase, que é
um sistema enzimático microssomal dependente do citocromo P450, que prejudica a síntese do
ergosterol na membrana citoplasmática e leva ao acúmulo de 14-α-metilesteróis. Esses
metilesteróis não possuem as mesmas propriedades físicas do ergosterol e levam à formação da
membrana com propriedades alteradas, que não desempenha as funções básicas necessárias ao
desenvolvimento do fungo (JAIN; SHEGAL, 2001; LACAZ et al., 2002; BERGOLD;
GEORGIADIS, 2004). Pode ser fungistático ou fungicida, dependendo da concentração da droga
(OLIVEIRA et al., 2002).
Terbinafina é uma alilamina que inibe a ação da enzima esqualeno epoxidase, impedindo a
formação do ergosterol no início da síntese. Há acúmulo de esqualeno intracelular que é tóxico
para a célula, resultando em rápida atividade fungicida. A elevada concentração de esqualeno
interfere na síntese da estrutura celular do fungo e na função da membrana celular, levando à
ruptura da parede e morte celular (LEYDEN, 1998; DONCKER, 1999; LACAZ et al., 2002;
BERGOLD; GEORGIADIS, 2004).
Devido à variedade de antifúngicos utilizados para tratamento de dermatofitoses, torna-se
evidente a necessidade de um método padronizado para determinação da susceptibilidade
antifúngica, entretanto, ainda não está estabelecido (JESSUP et al., 2000; BARROS et al., 2006;
SANTOS et al., 2006). A utilização de vários métodos pode conferir resultados diferentes
(NORRIS et al., 1999; JESSUP et al., 2000; FERNÁNDEZ-TORRES et al., 2002) e a importância

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
35
da padronização reflete em terapêutica eficaz, a qual é dependente da correta escolha da droga que
apresente atividade em relação ao fungo em questão (JESSUP et al., 2000; BARROS et al., 2006).
Foi proposto em 1998 e aprovado em 2002, pelo “National Committee for Clinical
Laboratory Standards” (NCCLS), atual “Clinical and Laboratory Standards Institute” (CLSI), o
documento M38-A com o protocolo padronizado para avaliação in vitro da atividade antifúngica
de fungos filamentosos que causam infecções invasivas, incluindo espécies de Aspergillus e
Fusarium, Pseudallescheria boydii (Scedosporium apiospermum), Rhizopus arrhizus e Sporothrix
schenckii. O documento inclui a seleção de agentes antifúngicos, preparação de soluções padrão e
diluições de antifúngicos utilizadas para realização, implementação e interpretação dos testes.
Além disso, essa padronização está relacionada com meio de cultura utilizado para crescimento e
estimulação da esporulação do fungo (batata-dextose-ágar), meio de cultura utilizado para teste de
susceptibilidade (RPMI-1640), determinação da concentração inibitória mínima, padronização do
inóculo, período (24 a 50h) e temperatura de incubação (35ºC), leitura e interpretação dos
resultados.
Por causa da falta de padronização para dermatófitos, vários autores citam modificações do
documento M38-A e algumas espécies já foram testadas (FERNÁNDEZ-TORRES et al., 2001;
PEREA et al., 2001; FERNÁNDEZ-TORRES et al., 2002; FERNÁNDEZ-TORRES et al., 2003b;
GHANNOUM et al., 2004; SANTOS; HAMDAN, 2005; BARROS et al., 2006; FERNÁNDEZ-
TORRES et al., 2006).
Algumas modificações referem-se às estruturas fúngicas utilizadas no inóculo (esporos e
fragmentos de hifas), tamanho do inóculo (103 a 104 estruturas fúngicas/mL), período de incubação
(4 a 10 dias), temperatura de incubação (28 a 35ºC), interpretação de resultados (7 a 10 dias),
determinação da concentração inibitória mínima (CIM) que será aquela capaz de inibir de 50 a
100% do crescimento fúngico (NORRIS et al., 1999; JESSUP et al, 2000; FERNÁNDEZ-
TORRES et al., 2001; FERNÁNDEZ-TORRES et al., 2002; FERNÁNDEZ-TORRES et al.,
2003b; GHANNOUM et al., 2004; KARACA; KOÇ, 2004; SANTOS; HAMDAN, 2005;
BARROS et al., 2006; FERNÁNDEZ-TORRES et al., 2006).
Outros métodos têm sido mencionados com algumas alterações, como o proposto pelo
“Antifungal Susceptibility Subcommittee of the European Committee on Antimicrobial
Susceptibility Testing” (AFST-EUCAST, 2002) que é aplicado para leveduras e fungos
filamentosos que não sejam dermatófitos. Algumas modificações sugeridas por BARROS et al.
(2006), incluem crescimento em meio de cultura batata-dextrose-ágar (BDA) suplementado de 2%
de farinha de arroz, adição de 2% de glicose ao RPMI-1640 e temperatura de incubação de 28ºC.
Esses autores compararam o protocolo M38-A (CLSI) com o proposto pelo AFST-EUCAST

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
36
(Discussion Document E.Dis. 7.1 ESCAMID) modificado, em 50 amostras de T. mentagrophytes e
50 de T. rubrum e não observaram diferenças significativas nos resultados.
Pujol et al. (2002) citam procedimento colorimétrico para a avaliação da susceptibilidade
antifúngica de dermatófitos. É utilizado Alamar Blue como indicador que altera a coloração de
azul para vermelho na presença de metabólitos resultantes do crescimento fúngico. Dessa forma,
também é facilitada a leitura da concentração inibitória mínima e podem ser empregadas técnicas
de macro e microdiluição. As drogas antifúngicas são obtidas comercialmente e a diluição em série
é preparada com o indicador que também funciona como diluente. Comparando a concentração
inibitória mínima obtida pelo CLSI e por esse método colorimétrico, foi relatada semelhança de
resultados de 87,7% para itraconazol, 81,6% para anfotericina B, 69,4% para cetoconazol e 67,3%
para fluconazol.
Karaca e Koç (2004) propuseram o método de disco-difusão por ser simples, eficaz e de
baixo custo. A leitura de resultados seguiu a recomendada pelo documento M2-A8 (CLSI, 2003)
para testes de sensibilidades aos antimicrobianos por disco-difusão, através da medição do
diâmetro do halo de inibição do crescimento formado ao redor de cada disco impregnado com a
droga. A comparação em relação ao método proposto pelo CLSI resultou em semelhança e
consistência de resultados. Entretanto, Rex et al. (1993), Hanzen (2000) e Rex et al. (2001) citam
que alguns azólicos não se difundem de forma eficiente no ágar tornando difícil a padronização
desta técnica.
Posteriormente, Fernández-Torres et al. (2006) constataram que pelo método de disco-
difusão em meio composto por RPMI-1640 adicionado de ágar, itraconazol mostrou menor
capacidade de inibição de crescimento para as espécies M. canis, M. gypseum, T. mentagrophytes e
T. rubrum, por apresentar menor halo de inibição.
A concentração inibitória mínima (CIM) é a menor concentração da droga que inibe o
crescimento de microrganismo, mas não indica erradicação. A concentração fungicida mínima
(CFM) é a menor concentração da droga capaz de eliminar o microrganismo. Devido as diferentes
formas de ação, como anteriormente mencionado, terbinafina apresenta CIM e CFM menor do que
itraconazol. Terbinafina é capaz de erradicar o fungo em concentrações baixas podendo apresentar
na mesma concentração atividade fungistática e fungicida (CIM = CFM). Já para itraconazol a
concentração fungicida mínima apresenta-se maior ou igual a concentração inibitória mínima
(CFM≥CIM) (LEYDEN, 1998).
Há controvérsia em relação a determinação da CIM para os azólicos. Alguns autores citam
que deve ser considerada CIM a que inibir pelo menos 50% do crescimento do fungo (PUJOL et
al., 1996; FERNÁNDEZ-TORRES et al., 2001; PEREA et al., 2001; PUJOL et al., 2002;

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
37
KARACA; KOÇ, 2004). Outros citam que tem que ser capaz de inibir pelo menos 80% do
crescimento (HAZEN, 1998; JESSUP et al., 2000; GHANNOUM et al., 2004; SANTOS;
HAMDAN, 2005; BARROS et al., 2006), embora a inibição total seja a mais eficiente (HAZEN,
1998; FERNÁNDEZ-TORRES et al., 2002; FERNÁNDEZ-TORRES et al., 2003a; FAVRE et al.,
2003, GHANNOUM et al., 2004).
De acordo com alguns autores, a concentração inibitória mínima de itraconazol para os
dermatófitos, é a que inibir 80% do crescimento desse grupo de fungos (NORRIS et al., 1999;
JESSUP et al., 2000; JAIN; SHEGAL, 2001; GHANNOUM et al., 2004; SANTOS; HAMDAN,
2005; BARROS et al., 2006; SANTOS et al. 2006). Com base nisso, a CIM é baixa quando
comparada a CFM que é relativamente alta. O coeficiente CIM/CFM apresenta-se alto o que indica
que itraconazol é mais fungistático do que fungicida (HAZEN, 1998; JAIN; SHEGAL, 2001;
GARCÍA-PATOS; APARICIO...; SECCO...).
Ghannoum et al. (2006) promoveram uma avaliação interlaboratorial, seguindo protocolo
M38-A (CLSI, 2002) modificado, para verificar a atividade antifúngica de itraconazol, terbinafina,
ciclopirox, griseofulvina, posaconazol e voriconazol em relação a T. mentagrophytes e T. rubrum.
Constataram que a determinação da concentração que inibe 50% do crescimento foi bastante
variável entre os laboratórios, para a mesma droga e a mesma amostra testada. Já para inibição de
80%, não houve variação significativa.
Santos et al. (2006) utilizaram a inibição de 80% do crescimento com critério de atividade
antifúngica de itraconazol para T. mentagrophytes e T. rubrum, obtendo resultados eficazes.
Vários autores concordam que a concentração inibitória mínima para terbinafina é a que
inibir totalmente o crescimento do fungo (HAZEN, 1998; FERNÁNDEZ-TORRES et al., 2001;
FERNÁNDEZ-TORRES et al., 2002; FERNÁNDEZ-TORRES et al., 2003b; SANTOS;
HAMDAN, 2005; SANTOS et al., 2006; BARROS et al., 2006).
Fernández-Torres et al. (2001) avaliaram a atividade antifúngica de 10 drogas em relação a
508 amostras de dermatófitos, segundo documento M38-A, adotado pelo CLSI (2002). Foram
constatadas concentrações inibitórias mínimas semelhantes entre itraconazol e terbinafina, tais
como 0.22µg/mL e 0.21µg/mL, respectivamente. Todos os antifúngicos mostraram-se eficazes,
com exceção do fluconazol por inibir 80% do crescimento fúngico em elevada concentração.
Ghannoum et al. (2004) verificaram a susceptibilidade antifúngica de M. canis, T.
mentagrophytes, T. tonsurans, T. rubrum e E. floccosum para fluconazol, itraconazol, posaconazol,
voriconazol, terbinafina e griseofulvina. Itraconazol, terbinafina, voriconazol e posaconazol,
destacaram-se por apresentarem concentrações inibitórias mínimas semelhantes e baixas. Foi
verificado que a inibição do crescimento de metade das amostras (CIM50) foi obtida nas

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
38
concentrações de 0.06 e 0.03 µg/mL para itraconazol e terbinafina, respectivamente. A inibição do
crescimento de 90% das amostras (CIM90) foi obtida nas concentrações de 0.125 e 0.03µg/mL,
para as mesmas drogas.
Cetinkaya et al. (2005) avaliaram a atividade antifúngica de itraconazol, terbinafina,
cetoconazol e fluconazol em amostras recém isoladas das mesmas espécies acima citadas. A faixa
de concentração inibitória mínima foi de 0.08 a 0.4µg/mL para itraconazol, 0.04 a 0.2µg/mL para
terbinafina, 0.09 a 1.1µg/mL para cetoconazol e 16.2 a 24µg/mL para fluconazol. Os autores
enfatizam a eficácia antifúngica de itraconazol e terbinafina em concentrações menores quando
comparadas com as outras drogas.
Barros et al. (2006) considerando CIM para itraconazol e terbinafina a que inibir 80% e
100% do crescimento fúngico, respectivamente, constataram elevada atividade inibitória dessas
drogas pela capacidade de inibição de 90% das amostras (CIM90) de Trichophyton sp. nas
concentrações de 0,25 µg/ml e 0,015µg/ml, respectivamente.
A determinação do período de incubação é de extrema importância para determinar CIM e
CFM. Dependendo do período de incubação, certas drogas como a griseofulvina, podem
apresentar ação fungistática ou fungicida. Geralmente, as drogas fungistáticas como itraconazol
estão mais sujeitas as interferências do que as fungicidas como terbinafina, e podem apresentar
ação fungicida devido ao efeito “postantibiotic” (HAZEN 2000), no qual não é observado
crescimento fúngico mesmo após a retirada da droga, devido a diminuição do metabolismo
fúngico (LICATA, et al., 1997). As drogas de ação fungicida, podem apresentar-se fungistáticas
quando o período de incubação for prolongado, entretanto isso é visto como artefato devido a
presença de estruturas fúngicas resistentes (HAZEN 2000).
Dessa forma, o período de incubação para teste de susceptibilidade antifúngica para
dermatófitos, varia de 4 dias (NORRIS et al., 1999; FAVRE et al., 2003; GHANNOUM et al,
2004) a 10 dias (KARACA; KOÇ, 2004; SANTOS; HAMDAN, 2005) sendo o sétimo dia, o
adequado para leitura dos resultados (FERNÁNDEZ-TORRES et al., 2002; FERNÁNDEZ-
TORRES et al., 2003a; KARACA; KOÇ, 2004; SANTOS; HAMDAN, 2005; BARROS et al.,
2006; FERNÁNDEZ-TORRES et al., 2006).
Segundo o CLSI (2002), a preparação do inóculo é importante para eficácia do teste. O
ideal é a utilização de culturas desenvolvidas em meio sólido, visto que a obtenção de esporos é
mais fácil. A utilização de emaranhado de hifas obtido de cultura em meio líquido não é
recomendada porque dificulta o contato e a atuação da droga nas hifas centrais. Dessa forma, pode
haver falsa resistência e capacidade de crescimento do fungo, pois a presença de células restantes e
com metabolismo, ainda que lento, permite o crescimento (HAZEN, 2000).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
39
Um dos aspectos que dificulta a padronização do teste de sensibilidade antifúngica para
dermatófitos, é a pouca esporulação, dependendo da espécie, e a conseqüente dificuldade para a
padronização do inóculo (GHANNOUM et al., 2004). Hazen (2000) cita que para esse grupo de
fungos, as estruturas que compõem o inóculo podem ser fragmentos de hifas, até mesmo porque
essas estruturas são formas infectantes para dermatofitoses e por isso é válido utilizá-las para
avaliação antifúngica.
Perea et al. (2001) constataram susceptibilidade antifúngica de várias espécies de
dermatófitos utilizando como inóculo a mistura de fragmentos de hifas e esporos.
Fernandez-torres et al. (2003b) comparando a susceptibilidade de fragmentos de hifas,
macroaleurias de parede espessa como as de Microsporim canis, M. cookei, M. gypseum, M.
racemosum e de parede delgada como as de Ttrichophyton ajelloi, constataram que a espessura da
parede celular dos esporos interfere na atuação da droga, pois é necessário maior concentração da
droga para inibir o crescimento fúngico.
Para dermatófitos, quando há pouca esporulação, o que predomina é a utilização de esporos
com fragmentos de hifas (NORRIS et al., 1999; JESSUP et al., 2000; FERNÁNDEZ-TORRES et
al., 2001; PETRIKKOU et al., 2001; FERNÁNDEZ-TORRES et al., 2002; FAVRE et al., 2003;
FERNÁNDEZ-TORRES et al., 2003b; KARAKA; KOÇ, 2004; FERNÁNDEZ-TORRES et al.,
2006). Entretanto para as espécies de Trichophyton, que esporulam com mais facilidade, sugere-se
a utilização de esporos (GHANNOUM et al., 2004; SANTOS; HAMDAN 2005; GHANNOUM et
al., 2006; SANTOS et al., 2006).
Além das estruturas, o tamanho do inóculo interfere na determinação da concentração
mínima inibitória (GHANNOUM et al., 2004).
Fernández-Torres et al. (2001) avaliaram a atividade antifúngica de 10 drogas em relação a
508 amostras de dermatófitos, segundo procedimento adotado pelo CLSI (2002), documento M38-
A, com inóculo que variou entre 104 e 106 UFC/ml. Foi constatado que o tamanho do inóculo
exerceu influência para determinação da concentração inibitória mínima, exceto para itraconazol e
terbinafina, que apresentaram CIM semelhantes, 0.22µg/mL e 0.21µg/mL, respectivamente.
Entretanto, Fernández-Torres et al. (2002) constataram diferenças significativas na
determinação da concentração inibitória mínima de itraconazol e terbinafina, em relação à
dermatófitos, utilizando inóculo de 104 UFC/mL, indicando que mesmo no inóculo padronizado, a
resposta às drogas pode ser diferente. As concentrações inibitórias obtidas foram 0.4µg/mL para
itraconazol e 0.04µg/mL para terbinafina. Resultados semelhantes, nas mesmas condições de
inóculo, foram citados por Santos e Hamdan (2005) para itraconazol e terbinafina, sendo as
concentrações de 0.5µg/mL e 0.03µg/mL, respectivamente.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
40
Santos e Hamdan (2005) constataram que diferentes meios de cultura (RPMI 1640,
McVeigh & Morton e ágar Sabouraud) e períodos de incubação (7 e 10dias) também influenciam
na determinação das concentrações inibitórias mínimas e fungicidas, para T. rubrum. A
concentração inibitória mínima obtida em diferentes meios de cultura no mesmo período de
incubação para a mesma amostra de fungo, foi significantemente diferente para azóis e
griseofulvina. Da mesma forma, diferentes períodos de incubação resultaram em concentrações
inibitórias diferentes para cada meio de cultura, quando azóis e griseofulvina são testados, não
havendo alteração para terbinafina. O meio de cultura RPMI 1640 foi o mais adequado, pois os
outros apresentaram discrepância dos resultados.
A variação da temperatura de incubação, entre 28 e 35ºC, parece ser o fator que menos
interfere no teste de sensibilidade antifúngica para dermatófitos (JESSUP et al., 2000;
FERNÁNDEZ-TORRES et al., 2001; FERNÁNDEZ-TORRES et al., 2002; FAVRE et al., 2003;
FERNÁNDEZ-TORRES et al., 2003a; KARACA; KOÇ, 2004; GHANNOUM et al., 2004;
SANTOS; HAMDAN, 2005; BARROS et al., 2006; FERNÁNDEZ-TORRES et al., 2006).
Norris et al. (1999) avaliaram crescimento a 30ºC e 35ºC, de T. rubrum, T. mentagrophytes
e T. tonsurans em placas de microdiluição utilizadas para avaliação de susceptibilidade antifúngica
e verificaram que a temperatura não influenciou no crescimento desses fungos.
Fernández-Torres et al. (2002) não observaram interferência significativa no teste de
susceptibilidade in vitro para dermatófitos realizado nas temperaturas de 28ºC e 37ºC. Por isso
sugeriram que a realização a 28ºC é satisfatória. De forma semelhante, Santos e Hamdan (2005)
citaram que a temperatura de incubação (28º e 35ºC) das placas de microdiluição não interferiu nos
resultados.
Devido ao aumento de casos de dermatofitoses, principalmente, em pessoas
imunocomprometidas (TSANG et al., 1996; PORRO et al., 1997; WALSH; GROLL, 1999;
GIORDANI et al., 2001; FERNÁNDEZ-TORRES et al., 2002; BERGOLD; GEORGIADIS, 2004;
VENKATESAN et al 2007), torna-se evidente a necessidade da padrozinação do método de
susceptibilidade antifúngica para esses fungos, pois a avaliação in vitro de drogas antifúngicas está
relacionada com a contribuição para efetiva terapia principalmente em pacientes com
dermatofitoses crônicas (CETINKAYA et al., 2005).
As dermatofitoses podem ser resistentes à terapia e a resposta ao tratamento pode não ser
adequada (BARCHIESI et al., 2001, FAVRE et al., 2004). A absorção das drogas de uso tópico
através da pele é insignificante e por causa disso é improvável o aparecimento de efeitos tóxicos
ao organismo. Mas quando há rompimento dessa barreira cutânea como em queimaduras e lesões
cutâneas extensas com exposição da derme, poderá haver irritações no local da aplicação

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
41
(FREEDBERG et al., 2003). Entretanto, sabe-se que o tratamento prolongado com uso de drogas
sistêmicas pode gerar efeitos tóxicos como náuseas, vômitos, anemia hemolítica, sonolência,
erupções cutâneas, efeitos antiandrogênicos como diminuição da libido, ginecomastia e
hepatotoxicidade, esta última ocorre entre 1-5% dos casos com raros casos fatais (GIORDANI et
al., 2001; FREEDBERG et al., 2003). Por isso, torna-se necessário o desenvolvimento de novas
estratégias terapêuticas (FOSTEL; LARTEY, 2000; TATSUMI et al., 2002; GARCÍA et al., 2003;
FERNÁNDEZ-TORRES et al., 2006).
Atenção especial tem se voltado para derivados naturais, com base no conhecimento da
produção de compostos antifúngicos pela natureza (WOJTASZEK, 1997; GURGEL et al., 2005).
Com o passar do tempo, produtos naturais como própolis, estão sendo cada vez mais utilizados na
medicina (PARK et al., 1998a).
2.5. PRÓPOLIS 2.5.1. Características Gerais
Própolis é um material resinoso, de consistência viscosa a dura e composição química
complexa, elaborado por abelhas a partir da coleta de exsudatos de diversas partes das plantas
como brotos, botões florais, folhas novas e casca (BURDOCK, 1998; PARK et al., 2000;
BANKOVA et al., 2000; CASTALDO; CAPASSO 2002; PARK et al., 2004; BANKOVA, 2005a;
FERNANDES JÚNIOR et al., 2006). A transformação desse material coletado pelas abelhas em
própolis, ocorre na colméia após mastigação e adição de enzimas salivares que são produzidas
durante a coleta da matéria-prima, como também pela adição de cera (PARK et al.,1997;
BURDOCK, 1998; PINTO et al., 2001; CASTALDO; CAPASSO, 2002; PIETTA et al., 2002;
FUNARI; FERRO, 2006; HEGAZI...).
As abelhas da espécie Apis mellifera, responsáveis pela elaboração da própolis, foram
introduzidas no Brasil em 1893 pelo padre português Antônio Carneiro. Em 1956 houve
introdução de abelhas africanas para melhoria da produção de mel, entretanto um escape acidental
de abelhas rainha promoveu africanização das abelhas, devido à reprodução entre abelhas
européias e africanas. Por causa disso, hoje em dia, as abelhas presentes no Brasil são
reconhecidas como abelhas africanizadas (PEREIRA et al., 2002).
A coloração da própolis depende da procedência, podendo variar entre marrom claro e
escuro, marrom avermelhado, esverdeado, amarelado e avermelhado (BURDOCK, 1998;
SALATINO et al., 2005; DAUGSCH et al., 2007). Possui odor característico de resina aromática

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
42
que pode variar de uma amostra para outra. O ponto de fusão varia entre 60 e 70 ºC podendo
atingir em alguns casos, até 100 ºC (MARCUCCI, 1995). É uma substância rígida, mas quebradiça
quando fria e que se torna dúctil e maleável quando aquecida (BURDOCK, 1998; SALATINO et
al., 2005).
A própolis é utilizada pelas abelhas para proteger a colméia de insetos e microrganismos
invasores, pelo emprego de uma fina camada que reveste tanto a entrada como o interior da
colméia. Também é utilizada para selar rachaduras, manter asséptico o ambiente interno da
colméia, preparar locais assépticos para a postura da abelha rainha e embalsamar invasores
(BURDOCK, 1998; KUJUMGIEV et al., 1999; BANKOVA et al., 2000; SAWAYA et al., 2002;
KOC et al., 2005; SALATINO et al., 2005; FUNARI; FERRO, 2006). O nome deriva do grego,
sendo que pro significa “a favor, em defesa de” e polis “cidade” (MARCUCCI, 1995;
BURDOCK, 1998; SALATINO et al., 2005).
Antes de Cristo, a própolis já era bastante utilizada no Egito para mumificação e também
pelos sacerdotes dotados de conhecimentos médicos e químicos. Praticamente todas as civilizações
antigas conheceram e utilizaram os produtos das abelhas como valiosos recursos na medicina
(PEREIRA et al., 2002). A história da medicina das civilizações Chinesa, Tibetana, Egípcia e
Greco-Romana contém registros com centenas de receitas para curar ou prevenir enfermidades
cujos ingredientes principais são mel, própolis e às vezes as abelhas (PARK et al., 2002b; PARK
et al...). Persas, romanos, gregos e incas também utilizavam para o tratamento de infecções, reduzir
inchaços, aliviar cólica e como cicatrizante. Na II Guerra Mundial, a própolis foi bastante utilizada
em hospitais como antimicrobiano e para alívio de queimaduras (HEGAZI...). Na Europa foi
utilizada como antibacteriano, antieczematoso, antiacne, no tratamento de abscessos gengivais
hemorrágicos, candidíase bucal e halitose. Também se fez presente nos receituários chineses como
medicamento ativo contra moléstias coronarianas e hipertensão (NOTHENBERG, 1997). A partir
da década de 60 tornou-se objeto de intensos estudos farmacológicos e bioquímicos na Europa,
Bulgária, Czechoslovakia e Polônia (BANKOVA, 2005a; SALATINO et al., 2005), sendo que em
1969 o extrato etanólico da própolis passou a ser industrializado farmaceuticamente (HEGAZI...).
No Japão, o uso da própolis foi impulsionado em 1985 após a realização do XXX Congresso
Internacional de Apicultura, sendo hoje, o principal comprador da própolis brasileira, pois
consome 90% da própolis exportada pelo Brasil, devido principalmente as propriedades biológicas
(AGUIAR et al., 2003; SALATINO et al., 2005; MARCUCCI et al., 2007; NASCIMENTO et al.,
2007).
O primeiro “paper review” foi escrito por Ghisalberti em 1979, indicando que era tempo de
colher mais informações sobre a própolis do que recomendá-la na prática médica, uma vez que a

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
43
composição química era muito desconhecida. Vinte anos após, aumentou a quantidade de
informações sobre a composição química e propriedades biológicas, entretanto a aplicação não
obteve tanto avanço (BANKOVA et al., 2000). Uma das causas é a capacidade de panacéia (várias
atividades biológicas), que atrapalha a aceitação e aplicação, uma vez que pode gerar desconfiança
devido à atribuição de várias atividades biológicas simultâneas (PEREIRA, 2002).
Atualmente existem diversos produtos contendo própolis que são comercializados pelas
industrias farmacêuticas e alimentícias em todo mundo, principalmente no Japão. São apresentados
sob várias formas, tais como balas, chocolates, doces, xampus, cremes para pele, soluções anti-
sépticas, pastas de dente entre outras (PARK et al., 1998a).
2.5.2 Fonte Botânica
Em regiões de clima temperado como Europa, América do Norte, oeste da Ásia e norte da
África, as fontes de própolis são poucas, sendo a principal, o botão do álamo Populus sp.
(MARKHAM et al., 1996; WOLLENWEBER; BUCHMANN, 1997; PARK et al., 2000). As
árvores do gênero Populus podem ser encontradas ao longo dos rios ou em regiões pantanosas.
São dotadas de raízes longas que podem causar fraturas na terra pela busca de água e por causa
disso não devem ser plantadas perto de residências ou de sistemas de canalização.
Em regiões de clima tropical o álamo não é nativo, mas existe uma grande diversidade
vegetal para retirada da resina, o que dificulta a correlação com a composição química da própolis.
Entretanto, o melhor indicador da origem botânica própolis é a análise da sua composição química
comparada com a provável fonte vegetal (PARK et al., 2002a). Por apresentar diversas
propriedades biológicas, existe grande interesse da determinação da composição química da
própolis da América do Sul (AGUIAR et al., 2003).
No Brasil, existem diversas espécies vegetais para a retirada de resina. No entanto, poucas
foram identificadas até agora, sendo o alecrim, assa-peixe, aroeira e eucalipto alguns exemplos de
onde as abelhas buscam a matéria-prima para a produção da própolis (PARK et al., 2000). A
principal fonte botânica da própolis verde brasileira é a resina extraída de Baccharis
dracunculifolia (PARK et al., 2002a; KUMAZWA et al., 2003; ALENCAR et al., 2005a),
comumente conhecida como alecrim-do-campo ou vassourinha. É nativa da região central do
Brasil mas pode ser encontrada em qualquer outra região do país (KUMAZWA et al., 2003).
Baccharis dracunculifolia além de ser amplamente encontrada no sul e sudeste do Brasil,
também é encontrada no Uruguai, Argentina, Paraguai e Bolívia, entretanto não predomina
(SALATINO et al., 2005). Ao oeste de Uberlândia (Minas Gerais, Brasil), foram encontradas

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
44
Brachiarea decumbens e Eucalyptus urophilla como fontes botânicas da própolis dessa região
(NASCIMENTO; BENZZAN, 2001).
A principal fonte botânica da própolis vermelha do Brasil é Dalbergia ecastophyllum,
conhecido como rabo-de-bugio, localizados nos estados da região nordeste do país, tais como
Paraíba, Pernambuco, Alagoas, Sergipe e Bahia (DAUGSCH et al., 2007).
Park et al. (2002a) classificaram própolis brasileira a partir de características físico-
químicas e identificaram fontes botânicas da própolis de cinco estados do sul do Brasil, um do
sudeste e seis do nordeste. Esses autores concluíram que própolis dessas regiões tem como fonte
botânica a resina extraída do álamo, de Baccharis dracunculifolia e de Hyptis divaricata,
respectivamente, com aspectos físicos e colorações diferentes.
Marcucci et al. (2007) verificaram que quando a fonte vegetal predominante da região vai
terminando, começam a aparecer novas fontes de resinas o que justifica a presença do álamo na
região sul do Brasil e o surgimento de própolis com composição química proveniente de várias
fontes botânicas.
A fonte botânica da própolis de Cuba é a resina floral de Clusia rosea (CUESTA-RUBIO
et al., 2002). De forma semelhante, a própolis da Venezuela apresenta como fonte botânica
exsudatos resinosos de botões florais de Clusia minor e C. major (TOMÁS-BARBERÁN et al.,
1993). Essas própolis apresentam como principais componentes benzofenonas poliisoprenilados,
oriundos das resinas florais, tornando essa própolis diferente das do Brasil e Europa (TOMÁS-
BARBERÁN et al., 1993; CUESTA-RUBIO et al., 2002).
2.5.3. Composição Química
A composição química varia quali e quantitativamente dependendo do tipo de flora que
esteja próxima a colméia, sendo, portanto, um reflexo da fonte botânica (BURDOCK, 1998;
PARK et al., 2002a; BANKOVA, 2005b; NASCIMENTO et al., 2007). A variabilidade genética
das abelhas (KOO; PARK, 1997), fatores associados à época do ano em que a própolis é produzida
(PARK et al., 1997), técnicas de extração e metodologias utilizadas podem influenciar na
composição (PARK et al., 1998a; KUMAZAWA et al., 2004; BANKOVA et al., 2005a; SOUSA
et al., 2007). Inclusive, numa mesma localidade, a composição pode variar sazonalmente
(SFORCIN et al., 2000).
Em regiões de clima temperado, é observada menor variação de composição química.
Nesse caso, as abelhas coletam a própolis apenas no verão (cerca de quatro meses) e por isso as
variações sazonais na composição da própolis são insignificantes. No Brasil, entretanto, a coleta de

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
45
própolis ocorre durante todo o ano, existindo deste modo, uma variação sazonal na sua
composição (BANKOVA et al., 2000).
Amostras de própolis das regiões sudeste (São Paulo, Minas Gerais e Rio de Janeiro) e sul
(Paraná, Santa Catarina e Rio Grande do Sul) do Brasil apresentam diferentes composições
químicas. Foi verificado que as amostras da região sul contem flavonóides em maior concentração
do que as da outra região, já que o clima se assemelha ao de zonas temperadas da Europa (PARK
et al., 2002a; MARCUCCI et al., 2007).
PARK et al. (1998a) demonstraram que própolis de várias regiões do Brasil (Minas Gerais,
São Paulo, Goiás, Mato Grosso do Sul, Paraná e Rio Grande do Sul) apresentam presença variada
de flavonóides como quercetina, canferol, apigenina, isorhamnetina, pinocembrina, sakuranetina,
isosakuranetina, crisina, acacetina, galangina e canferide e, em maiores concentrações,
sakuranetina e acacetina.
Os primeiros componentes identificados na própolis foram a vanilina, ácido ciâmico e
álcool cinâmico (DIETRICH; KUSTENMACHER, 1911) seguidos de flavonóides e terpenos
(PROPAVK, 1975). Já foram identificadas mais de 300 substâncias e algumas delas estão citadas
na Tabela 1.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
46
COMPOSTOS REFERÊNCIAS BIBLIOGRÁFICAS
Flavonóides
crisina, tectocrisina, canferol, canferide,
pinocembrina, pinobanskina, quercetina,
galangina, acacetina, apigenina, galangina,
quercetina, tectocrisina, isosakuretina,
pinostrobina, isoramnetina, sakuranetina
Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Boudourova-Krasteva et al 1997;
Martos et al 1997; Banskota et al. 1998; Bankova et al.,
2000, Fontana et al., 2000; Castro, 2001; Marcucci et al.,
2001; Park et al., 2002a; Pereira et al., 2002; Funari e
Ferro, 2006.
Ácidos fenólicos
ácido caféico, ácido ferúlico, ácido
cinâmico, artepelin C, ácido cumárico,
derivados prenilados do ácido p-cumárico
dupranina e derivados
Walker e Crane, 1987; Greenaway et al., 1991; Aga et
al., 1994; Marcucci, 1995; Tatefuji et al., 1996;
Boudourova-Krasteva et al., 1997; Matsuno et al., 1997;
Banskota et al., 1998; Tazawa et al., 1998; Bankova et
al., 2000; Fontana et al., 2000; Castro 2001; Marcucci et
al., 2001; Park et al., 2002a; Pereira et al., 2002; Funari
e Ferro, 2006; Sousa et al., 2007.
Aldeídos aromáticos
vanilina, isovanilina, gálico, salicílico,
hidroxicinâmico, isoferúlico, benzaldeído,
aldeído capróico, p-hidroxibenzaldeído
Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Markham et al., 1996; Christov et al.,
1998; Tazawa et al., 1998; Bankova et al., 2000;
Fontana et al., 2000; Pereira et al., 2002.
Álcoois
Triterpenos, fenetílico, prenílico,
isobutenol, benzílico
Christov et al., 1998; Marcucci et al., 1998; Bankova et
al., 2000.
Lignanas;
sesamina, ascantina, sesartenina,
dihidrobenzofuran
Valcic et al., 1998; Bankova et al., 1998; Walker e
Crane, 1987; Greenaway et al., 1991; Marcucci, 1995;
Bankova et al., 1996; Banskota et al., 1998; Christov et
al., 1999; Fontana et al., 2000; Bankova et al., 2000;
Pereira et al., 2002.
Aminoácidos
alanina, arginina, asparagina, ácido
aspártico, cistina, cisteína, glicina,
histidina, triptófano, isoleucina, serina,
lisina, metionina, ornitina, fenilalanina
Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Fontana et al., 2000; Bankova et al.,
2000; Pereira et al., 2002.
continua na página seguinte
Tabela 1. Componentes químicos gerais de amostras de própolis.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
47
Ácidos graxos
araquídico, behênico, cerótico, lignocérico,
palmítico, oleico, láurico, mirístico,
Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Fontana et al., 2000; Bankova et al.,
2000; Pereira et al., 2002.
Terpenos
ácido diterpenos, farnesol, geraniol,
cimeno, limoneno, estireno, naftaleno, β-
bisabolol, derivados de clerodane,
derivados do labdane, β- amirin,
sesquiterpenóides, ledol, spatulenol
Matsuno 1995; Walker e Crane, 1987; Greenaway et
al., 1991; Marcucci, 1995; Bankova et al. 1996;
Matsuno et al. 1997; Banskota et al. 1998; Bankova et
al., 2000; Fontana et al., 2000; Bankova et al., 2000;
Pereira et al., 2002.
Acúcares
ribofuranose, frutose, glucitol, glicose,
talose, sacarose, xilitol, xilose, galactose,
manose, ácido galacturônico, lactose,
maltose, melibiose, eritritol, inositol
Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Bankova et al. 1998; Bankova et al.,
2000; Fontana et al., 2000; Pereira et al., 2002.
Compostos voláteis
Sesquiterpenos, monoterpenos Bankova et al. 1995; Borcic et al. 1996; Bankova et al
1998; Bankova et al., 2000.
Cetonas
Acetofenonas e derivados Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Banskota et al. 1998; Bankova et al.,
2000; Fontana et al., 2000; Bankova et al., 2000; Pereira
et al., 2002.
Vitaminas
A, B1, B2, B6, C e E Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Fontana et al., 2000; Bankova et al.,
2000; Pereira et al., 2002.
Minerais
alumínio, vanádio, ferro, cálcio, silício,
manganês, estrôncio, sódio, potássio,
magnésio, bário, estrôncio, cádmio,
chumbo, cobre, manganês, ferro, cálcio,
vanádio, silício, alumínio, níquel, zinco
Walker e Crane, 1987; Greenaway et al., 1991;
Marcucci, 1995; Fontana et al., 2000; Bankova et al.,
2000; Pereira et al., 2002.
Continuação da Tabela 1.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
48
Entretanto, flavonóides e ácidos fenólicos são os compostos mais importantes, pois são os
maiores responsáveis por várias atividades biológicas, estando dessa forma, interligados
(MARCUCCI et al., 2001; HAVSTEEN, 2002; FERNANDES JÚNIOR et al., 2006; PACKER;
LUZ, 2007).
Flavonóides são pigmentos de plantas responsáveis pela coloração de flores e frutos, sendo
bastante utilizados para classificação taxonômica. Controlam o crescimento da planta e
representam uma barreira química de defesa contra bactérias, fungos, vírus, insetos e animais
herbívoros, principalmente por inibição enzimática que pode ser por competição ou alostérica.
Entretanto, atuam também de forma harmônica com plantas e insetos, atraindo-os e orientando-os
até o néctar, contribuindo enormemente para a polinização. A toxicidade às células humanas é
muito baixa, por isso são componentes presentes em preparações de uso medicinal, como por
exemplo na própolis (HAVSTEEN, 2002). Já foram atribuídas aos flavonóides propriedades
antiinflamatória, antimicrobiana (HAVSTEEN, 1983) antialérgica, antioxidante e antitumoral
(HARBORNE; WILLIAMS, 2000).
Os ácidos fenólicos derivam do ácido benzóico e são responsáveis por diversas atividades
da própolis, principalmente antitumoral. Ácido caféico e ácido gálico são tipos de ácidos fenólicos
que inibem seletivamente o vírus HIV, pois se ligam irreversivelmente a proteína gp120 presente
no vírus, proteína esta essencial para a capacidade de infecção pelo mesmo (VLIETINCK et
al.,1998).
Devido a biodiversidade da flora no Brasil, diferentes compostos químicos podem estar
relacionados as mesmas propriedades biológicas. A diminuição em alguns componentes
biologicamente ativos como os fenólicos são acompanhados pelo aumento de outros, por exemplo,
ácidos diterpênicos. Como a própolis da região sul do Brasil é rica em flavonóides e a da região
sudeste é rica em ácidos fenólicos (SALATINO et al., 2005), pode-se esperar que algumas
atividades biológicas relacionadas a estes compostos, sejam similares em diferentes estações do
ano (BANKOVA et al., 1998). Isto foi comprovado por Sforcin et al. (2000) que demonstraram
não existir diferença estatisticamente significante na atividade antimicrobiana da própolis coletada
de São Paulo durante as quatro estações do ano, em relação a Staphylococcus aureus e Escherichia
coli. A subespécie da abelha também tem influência na composição quantitativa da própolis,
porém o critério de escolha da fonte botânica pelas abelhas permanece incerto (BANKOVA et al.,
1998).
Sabe-se que a própolis européia é constituída principalmente por substâncias resinosas
provenientes de Populus nigra (BANKOVA et al., 2000) por isso, contem fenólicos típicos:

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
49
flavonóides agliconas, flavonas e flavononas, ácidos fenólicos e seus ésteres ácidos aromáticos e
seus ésteres (BANKOVA et al., 2000; BANKOVA et al., 2002).
A própolis do Brasil, em geral, é composta por 50% de resina e bálsamo vegetal, 30% de
cera, 10% de óleos essenciais e aromáticos, 5% de pólen e 5% de outras substâncias variadas,
incluindo restos orgânicos, sendo considerada uma das misturas mais heterogêneas (BURDOCK,
1998; PARK et al., 2002a; PIETTA et al., 2002). Os constituintes principais são os compostos
fenólicos representados pelos flavonóides, ácidos fenólicos e seus ésteres, que são amplamente
distribuídas no reino vegetal, sendo encontradas em diversas partes da planta como em frutas,
sementes, hastes e flores (CUSHNIE; LAMB, 2005). Flavonóides agliconas (canferol, quercetina,
ramnazina) já foram constatados, embora a origem vegetal seja diferente da Europa (MARCUCCI
et al., 2007).
2.5.4. Atividades Biológicas
Além da aplicação industrial, a própolis destaca-se pelas propriedades biológicas como
atividade antimicrobiana (BIANCHINI; BEDENDO, 1998; BANKOVA et al., 2000; SFORCIN et
al., 2000;; MARCUCCI et al 2001; BANKOVA, 2005a, FERNANDES JÚNIOR et al., 2006;
BARROS et al., 2007) antifúngica (BANKOVA et al., 2000; MURAD et al., 2002, CUSHNIE;
LAMB, 2005) antiviral, como para os vírus influenza e os da herpes (BURDOCK, 1998;
BANKOVA et al., 2000; BARROS et al., 2007), anti-HIV (PARK et al., 2000) antiinflamatória
(BANKOVA et al., 2000; REIS et al., 2000; BARROS et al., 2007), antitripranossomal
(PRYTZYK et al., 2003), antioxidante (BANKOVA et al., 2000; KUMAZAWA et al., 2004;
SIMÕES et al., 2004), imunomodulatória (BANKOVA et al., 2000; CHEN et al., 2004; SFORCIN
et al., 2005; BARROS et al., 2007), antitumoral (PARK et al., 1998a, 1998b; KOO et al., 1999;
BANKOVA et al., 2000; KOO et al., 2000; BAZO et al., 2002; AKAO et al., 2003; CHEN et al.,
2004; ISHIKAWA et al., 2004), antiulcerativa (REIS et al., 2000; ALENCAR et al., 2005a;
BARROS et al., 2007; SOUSA et al., 2007), como coadjuvante no tratamento de periodontite
crônica (AMARAL et al., 2006), sequestrante de radicais livres (CHEN et al., 2004),
hepatoprotetora (BANKOVA et al., 2000; BANKOVA, 2005a; BARROS et al., 2007), cicatrizante
e anestésica (PARK et al., 1998a; PARK et al., 2000; PARK et al., 2002b), entre outras.
Os flavonóides, juntamente com ácidos fenólicos e seus ésteres, aldeídos fenólicos e
cetonas são considerados os mais importantes compostos antimicrobianos da própolis
(FERNANDES JÚNIOR et al., 2006). A atividade antibiótica dos flavonóides é decorrente,

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
50
principalmente, da inibição de atividade de proteases (HAVSTEEN 2002) e fosfolipases
extracelulares (D´AURIA et al., 2003).
A atividade antimicrobiana da própolis européia é similar à própolis brasileira (POPOVA
et al., 2004), entretanto, a ação sobre o microrganismo alvo pode variar (MENEZES, 2005).
Mesmo com variação na concentração de flavonóides, própolis de várias regiões do Brasil
(Minas Gerais, São Paulo, Goiás, Mato Grosso do Sul, Paraná e Rio Grande do Sul) demonstraram
atividade antimicrobiana para Streptococcus mutans, a qual foi verificada pela presença de halo
inibitório (0,5 a 1,5mm) ao redor do disco impregnado com extrato alcoólico dessa própolis
(PARK et al., 1998a).
Park et al. (2001) verificaram atividade antimicrobiana semelhante para amostras de
própolis de regiões subtropicais do Brasil (Santa Cruz), do Uruguai e da Argentina (Buenos Aires
e Tucumán), em relação a Staphylococcus aureus. Todas as própolis apresentaram atividades
inibitórias semelhantes (halo de 2mm), exceto a própolis do Uruguai, que inibiu de forma
moderada (halo de 1mm). Além disso, constataram que a resina do álamo Populus sp. presente
nessas regiões, inibiu fortemente (halo de 4mm) o crescimento dessa bactéria.
Popova et al. (2005) verificaram que a atividade antimicrobiana da própolis da Turquia,
com elevada concentração de ácidos fenólicos e flavonóides, é maior do que para própolis com
baixa concentração dessas substâncias. Apesar das diferentes composições químicas, a atividade
antimicrobiana sempre está presente, independente da origem geográfica, da fonte botânica e do
clima da região (BANKOVA, 2005b), uma vez que as abelhas utilizam a própolis como anti-
séptico (MENEZES, 2005).
A maioria dos flavonóides com atividade de proteção a planta contra ataques de
microrganismos são isoflavonóides, flavonas e flavononas (HARBORNE; WILLIAMS 2000). O
efeito antifúngico de sakuranetina (ATKINSON; BLAKEMAN, 1982) e isoflavonóides de
leguminosas (ervilhas, feijão, soja, grão-de-bico) foi verificado pela inibição da germinação de
conídios de Botrytis cinerea na superfície das folhas (HARBONE; WILLIAMS 2000).
Em aplicações agrícolas, foi avaliado o uso da própolis como agente de controle da
germinação de esporos do fungo da ferrugem do cafeeiro (Hemileia vastatrix). A partir da
concentração de 2mL de própolis por litro de água, o controle de germinação de esporos
apresentou-se superior a 99%. Essa dosagem de própolis mostra a viabilidade em uso agrícola
como fungicida natural para fins de cafeicultura e também agricultura orgânica (PEREIRA et al.,
2001).
Devido a habilidade em inibir a germinação de esporos de fungos fitopatógenos, tem se
proposto o uso de flavonóides como agente antifúngico para fungos patógenos ao homem

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
51
(HARBORNE; WILLIAMS, 2000; CUSHNIE; LAMB, 2005). Já foi verificada a atividade
antifúngica de flavonóides em relação a Candida albicans (VALSARAJ et al., 1997; WACHTER
et al., 1999) e Aspergillus flavus (ZHENG et al., 1996).
De um modo geral, as ações antibacteriana e antifúngica da própolis estão entre as mais
avaliadas (MARCUCCI, 1995; BURDOCK et al., 1998), sendo a antimicrobiana mais
popularmente conhecida e a mais comprovada cientificamente (MENEZES, 2005). Marcucci et al.
(2001) constataram atividades antibacteriana (Pseudomonas aeruginosa, Staphylococcus aureus,
Streptococcus faecalis, Escherichia coli) e antitripanossomal da própolis do Paraná.
A ação antifúngica (fungistática e fungicida) é atribuída aos ácidos fenólicos (ácido
cinâmico, ácido felúrico e ácido caféico), terpenos e a flavonóides como crisina, ermanina,
galangina, canferol, pinobanskina e pinocembrina, sendo a atividade fungicida atribuída
principalmente a esse último (PARK et al...). Essas substâncias são encontradas nas própolis verde
(BANKOVA et al., 2000; PARK et al., 2002a; SALATINO et al., 2005; MARCUCCI et al., 2007)
e na vermelha, ambas do Brasil (DAUGSCH et al., 2007; SILVA et al., 2007).
A ação anifúngica da própolis já foi verificada em relação a dermatófitos como M. canis,
M.gypseum (DOBROWOLSKI et al., 1991), Trichophyton mentagrophytes e T.rubrum
(DOBROWOLSKI et al., 1991; SOARES; CURY, 2001; KOC et al., 2005); a leveduras como
Candida albicans (KUJUMGIEV et al., 1999; SAWAYA et al., 2002; SANTOS et al., 2005), C.
tropicalis, C. krusei, C. guilliermondii (OTA et al., 2001; SAWAYA et al., 2002) C. lusitaneae, C.
stellatoidea e C. parapsilosis (SAWAYA et al., 2002), Paracoccidioides brasiliensis (MURAD et
al., 2002) e Cryptococcus neoformans (FERNANDES et al., 2007), como também em relação a
outros fungos filamentosos como Aspergillus flavus (ZHENG et al., 1996); A. tamarii, A. flavus,
Cladosporium sphaerospermum, Penicillium digitatum e P. italicum (AFOLAYAN; MEYER,
1997) A combinação de drogas antimicóticas com a própolis aumenta ainda mais a atividade sobre
os fungos (HEGAZI et al., 2000).
A atividade antifúngica da própolis em relação a dermatófitos e espécies de Candida tem
sido atribuída, em parte, aos flavonóides (CAFARCHIA et al., 1999).
Flavonóides prenilados, flavonas e flavononas apresentaram atividade antifúngica para C.
albicans (VALSARAJ et al., 1997; WACHTER et al., 1999) e A. flavus (CUSHNIE; LAMB,
2005). Galangina, um flavonóide comum presente na própolis, tem apresentado atividade
antifúngica para A. tamarii, A. flavus, Cladosporium sphaerospermum, Penicillium digitatum and
P. italicum (CAFARCHIA et al., 1999).
Kujumgiev et al. (1999) observaram que amostras de própolis de diferentes regiões
geográficas (Brasil, Bulgária, Egito e Mongólia) apresentaram atividade antibiótica para Candida

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
52
albicans, Staphylococus aureus e Escherichia coli, como também antiviral para vírus da influenza
aviária. Para amostras de própolis de região temperada, os flavonóides e ésteres de ácidos
fenólicos foram os responsáveis por essas atividades. Própolis de região tropical apresentaram
traços dessas substâncias mas apresentaram atividades semelhantes, indicando que a atividade
biológica é resultante de uma mistura de componentes e não de um em particular
Da mesma forma, própolis do Brasil e da Bulgária estimularam a atividade fungicida de
macrófagos peritoneais em camundongos, em relação a Paracoccidioides brasiliensis
demonstrando melhor resposta no mecanismo de resistência a esse fungo (MURAD et al., 2002).
Silici et al. (2005) verificaram atividade antifúngica da própolis de apiários da Turquia,
coletadas por abelhas Apis mellifera caucasica em relação a C. albicans, C. glabrata,
Trichosporon spp. e Rhodotorula sp., nas concentrações de 0.03 a 3.75µg/mL.
Oliveira et al. (2006) constataram o efeito fungistático da própolis da região de Maringá na
concentração de 5x10-2 mg/ml sobre espécies de Candida e de Trichosporon assim como, sobre
Geotrichum candidum isodalas de onicomicoses, de acordo com o protocolo de referência M27-A,
segundo CLSI (2002). O efeito fungicida foi obtido na concentração de 20x10-2mg/ml, embora a
susceptibilidade das leveduras tenha sido diferente entre as espécies. A predominância de
flavonóides determinou a propriedade antifúngica e o pH=5,6 do extrato, revelou a proximidade
com o pH da pele indicando a possibilidade tópica de utilização, pois quanto mais distante é esse
valor, maior o grau de agressão à pele. A partir desses dados, foi sugerido o uso da própolis como
uma opção para o tratamento de onicomicoses, pois além de ser um produto natural, é de baixo
custo e não é tóxico.
2.5.5. Extratos Alcoólico e Aquoso
Várias pesquisas têm demonstrado as atividades biológicas da própolis, entretanto a
maioria delas está relacionada ao extrato alcoólico (MARKHAM et al., 1996; MORENO et al.,
1999; KOO et al., 2000; MARCUCCI et al., 2001; SILICI et al., 2005; AMARAL et al., 2006;
FERNANDES JÚNIOR et al., 2006). Foi demonstrado que mais compostos ativos são extraídos da
própolis nativa se o extrato for preparado em álcool etílico em concentrações acima de 50% e por
isso o efeito antibiótico é maior. Quando a água é utilizada como solvente, foi verificado que as
atividades antifúngicas e antibacterianas são mais fracas comparando com extrato etanólico.
(SAWAYA et al., 2002).
O efeito bactericida do extrato etanólico da própolis é promovido pela presença de
ingredientes ativos, principalmente de flavonóides, que em combinação com antimicrobianos

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
53
podem permitir a redução da dose clínica de determinados antibacterianos. Isso reflete na
diminuição da incidência de efeitos colaterais e potencializa a antibioticoterapia no tratamento de
infecções quando a resistência bacteriana torna-se fator determinante (MIRZOEVA et al., 1997).
O extrato alcoólico da própolis de diferentes localidades da Argentina apresentou atividade
antibacteriana na concentração inibitória mínima de 7.8µg/mL para Streptococcus pyogenes e
outras bactérias resistentes a antibióticos (MORENO et al., 1999).
Park et al. (2000) verificaram atividade antimicrobiana de extratos alcoólicos de amostras
de própolis de São Paulo, Paraná, Bahia e Pernambuco em relação as espécies Staphylococcus
aureus e Streptococcus mutans, pela formação de halo transparente ao redor da colônia indicando
inibição de crescimento.
Prytzyk et al. (2003) constataram atividade antifúngica dos extratos etanólicos de diferentes
amostras de própolis de duas regiões da Bulgária (sudeste e oeste) na concentração de 0.5mg/mL,
em relação a C. albicans. O diâmetro do halo de inibição do crescimento desse fungo foi igual a 13
e a 13,7 mm para as própolis das regiões sudeste e oeste, respectivamente. Esses autores
consideraram que a susceptibilidade antifúngica é negativa quando o diâmetro do halo é menor do
que 10mm.
Trichoderma viride, Aspergillus niger e Penicillium chrysogenum não apresentaram
susceptibilidade ao extrato aquoso da própolis da Alemanha, utilizada em concentrações de até
10% m/v. Por outro lado, os dois fungos filamentosos foram sensíveis aos extratos etanólicos
provenientes de outras localidades (Colômbia, Etiópia, Alemanha, Itália, Polônia, Rússia e África
do Sul) em concentrações de 0.5 a 2.5% m/v. Entre os 16 extratos alcoólicos avaliados, T. viride só
não apresentou sensibilidade em cinco (um da Etiópia, um da Colômbia, três da África do Sul).
Entre seis espécies de bactérias avaliadas (Bacillus subtilis, B. megaterium, B. brevis,
Pseudomonas syringae e Escherichia coli), a atividade antibacteriana do extrato aquoso dessa
própolis foi obtida na máxima concentração utilizada (10% m/v) para quatro primeiras espécies e
não foi obtida na última (GAREDEW et al., 2004).
Koc et al. (2005) constataram a atividade antifúngica do extrato etanólico da própolis da
Turquia (“poplar type”) em relação a Trichophyton rubrum e T. mentagrophytes segundo
protocolo M38-A (CLSI, 2002). Foi obtida inibição de 50% e 90% das amostras analisadas nas
concentrações de 0.1µg/mL e 0.2 µg/mL, respectivamente. Além disso, o uso da própolis como
anti-séptico para dermatófitos representa excelente alternativa no caso de tratamento de tinea pedis
(SOARES; CURY, 2001).
De acordo com o documento M27-A (CLSI, 2002) foi atribuída atividade antifúngica ao
extrato etanólico da própolis da Turquia para C. albicans, C. glabrata, Trichosporon spp.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
54
Rhodotorula sp. cuja CIM ≤0.06µg/mL foi a mesma para todas as amostras. A concentração
inibitória mínima para itraconazol foi ≤0.35µg/mL, para todas as amostras, indicando que a
própolis foi tão efetiva quanto o itraconazol (SILICI; KOÇ, 2006).
Quiroga et al. (2006) verificaram a susceptibilidade de espécies de Aspergillus, Fusarium,
Saccharomyces e Rhodotorula mantidas nas coleções de culturas do Instituto de Estudios
Vegetales Dr. A.R. Sampietro e da Universidad Nacional de Tucumán, na Argentina, em relação a
amostras de própolis da região noroeste da Argentina. Os resultados mostraram que esses fungos
foram tão sensíveis a própolis quanto ao cetoconazol e clotrimazol, principalmente pela presença
de pinocembrina e galangina presente no extrato alcoólico. Esses mesmos autores verificaram a
capacidade citotóxica das amostras de própolis nas plantas e constataram a baixa citotoxicidade,
sugerindo a partir desses dados o uso desse produto natural para controle de pragas de plantas.
Alguns trabalhos têm mencionado ausência ou pouca toxicidade da própolis (BURDOCK,
1998; MORENO et al., 2005) inclusive da própolis do Brasil (REIS et al., 2000; RIBEIRO et al.,
2004). Estes resultados já eram esperados, visto que os principais constituintes da própolis
possuem toxicidade relativamente baixa (ADELMANN, 2005).
Entretanto há relatos de dermatite de contato em apicultores espanhóis (CALLEJO et al.,
2001), em pessoas na Rússia e em outras regiões do mundo (MARCUCCI, 1995; BURDOCK,
1998).
Ribeiro et al. (2004) não constataram efeitos tóxicos da própolis sobre o peso corporal e
sobre os metabolismos protéico, lipídico, mineral e de carboidratos de coelhos.
Reis et al. (2000) observaram atividade antiinflamatória do extrato etanólico de amostras de
própolis de regiões de São Paulo, com atividade semelhante a de medicamentos sintéticos
relacionados e a ausência de efeito tóxico.
Experimentos com ratos demonstraram que uma alta dosagem de própolis (700 mg/Kg) não
foi capaz de causar efeitos indesejáveis nesses animais (DOBROWOLSKI et al., 1991). O mesmo
resultado foi observado por pesquisadores russos, os quais demonstraram que a dosagem de 350
mg/Kg administrada em camundongos, não foi capaz de causar efeitos tóxicos. A dose letal
(DL50) para camundongos só foi observada com a ingestão de uma quantidade superior a 7.340
mg/Kg. No entanto, os efeitos indesejáveis podem ser evitados com utilização de dosagens
controladas (RIBEIRO et al., 2004).
Dessa forma, pesquisas científicas vêm corroborando para acentuar o potencial terapêutico
da própolis (MENEZES, 2005).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
55
2.5.6. Própolis Verde
2.5.6.1. Fonte Botânica
Baccharis dracunculifolia é a principal fonte botânica da própolis verde produzida em
Minas Gerais e São Paulo por abelhas Apis mellifera (PARK et al., 2000; PARK et al., 2002a).
Alencar et al. (2005a) constataram 96% de similaridade entre as própolis dessas regiões quanto à
fonte botânica. Entretanto a composição química pode variar mesmo quando se têm abelhas da
mesma espécie e o mesmo vegetal como principal fonte botânica. Além de fatores climáticos já
mencionados, qualquer interferência agrícola acentuada na região, pode interferir.
Sousa et al. (2007) verificaram que a produção da própolis pode ocorrer em qualquer lugar
desde que seja oferecida matéria-prima, mas a produção da própolis verde ocorre apenas em
regiões onde a mata nativa característica de cerrado é predominante. Caso contrário, a própolis não
pode ser classificada como própolis verde, pois além de Baccharis dracunculifolia podem existir
outras fontes que influenciem na composição química.
Isso foi observado em cinco regiões de São Paulo e uma de Minas Gerais, as quais tinham
Apis mellifera e Baccharis dracunculifolia como produtor e fonte botânica, respectivamente. A
própolis verde só foi produzida em duas regiões (Capetinga-MG e no distrito de Chave da
Taquara-SP), devido a presença predominante da respectiva fonte botânica. Nas outras regiões,
havia além do alecrim, culturas de cana-de-açúcar, café e eucalipto, que modificaram a
composição química da própolis verde (SOUSA et al., 2007).
2.5.6.2. Composição Química
A própolis verde brasileira apresenta alta concentração de compostos fenólicos com
predomínio de ácidos fenólicos (NASCIMENTO; BEZZAN, 2001; PINTO et al., 2001; PARK et
al., 2002a; BANKOVA, 2005a; SOUSA et al., 2007), sendo o ácido cinâmico o mais abundante,
principalmente os que contem o grupo prenil, como por exemplo, o artepelin C (PARK et al.,
2002a; SALATINO et al., 2005; SOUSA et al., 2007) (Tabela 2). Além desses, também são
citados terpenos (ácidos diterpênicos) e ácidos aromáticos (ALENCAR et al., 2005a).
Flavonóides diferentes da própolis européia já foram constatados na própolis verde do
Brasil, tais como canferide (BOUDOROVA-KRAVSTEVA et al., 1997; PARK et al., 2002a;

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
56
MARCUCCI et al., 2007), pinobanskina (PARK et al., 2002a; MARCUCCI et al., 2007) crisina,
galangina (MARCUCCI et al., 2007) canferol e isosakuranetina (PARK et al., 2002a).
Mesmo não sendo abundante como na própolis européia, os flavonóides junto com os
ácidos fenólicos, compreendem as classes mais importantes (PINTO et al., 2001; SALATINO et
al., 2005; NASCIMENTO et al., 2007) pois a elas são atribuídas grande parte da bioatividade
contra vários microrganismos patógenos e insetos invasores da colméia (BANSKOTA et al., 1998;
BURDOCK, 1998; HARBORNE; WILLIAMS, 2000) como também as atividades antimicrobiana,
antioxidante, antiviral, antitumoral, antiinflamatória (PINTO et al., 2001; FUNARI; FERRO,
2006).
Além dessas substâncias, também já foram encontrados terpenos (ácidos sesqui, di e
triterpênicos) (SALATINO et al., 2005), acetofenonas (KUMAZAWA et al., 2003; PARK et al.,
2004), lignanas, álcool triterpênico e hidrocarbonos (BANKOVA et al., 2000).
Própolis das regiões de Minas Gerais e São Paulo são ricas em ácidos, entre eles: ácido
caféico, ácido cafeoilquínico, ácido p-cumárico, artepelin C e derivados prenilados (MARCUCCI
et al., 2007).
Dependendo do tipo de solvente empregado na extração ou mesmo das condições
utilizadas no processo, haverá maior ou menor eficiência de extração das substâncias fenólicas
(PINTO et al., 2001). Entretanto, não há dúvidas que em outros ecossistemas, a própolis apresente
diferente composição química, a depender da fonte botânica (BANKOVA et al., 2000;
BANKOVA, 2005b).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
57
COMPOSTO REFERÊNCIA BIBLIOGRÁFICA
Flavonóides
canferol, canferide, pinobanskina,
pinocembrina, acacetina, crisina, galangina,
isossacuranetina, sacuretina
Park et al., 1998a; Park et al., 2002; Funari e Ferro,
2006; Marcucci et al., 2007.
Ácidos fenólicos
ácido cafeico, ácido cafeoilquínico, derivados
prenilados do ácido p-cumárico, ácido
cinâmico, artepelin C, ácido felúrico
Bankova et al., 2000; Marcucci et al., 2001; Pinto et
al., 2001; Alencar et al., 2005a; Funari e Ferro,
2006; Sousa et al., 2007.
Cetonas
Acetofenonas Bankova et al., 2000; Pinto et al., 2001;
Kumazwa et al., 2003; Park et al., 2004.
Aldeídos aromáticos (vanilina e isovanilina)
e lignanas
Bankova et al., 2000.
Álcoois
Triterpenos Bankova et al., 2000; Pinto et al., 2001.
Terpenos
ácidos sesqui e diterpênicos
Bankova et al., 2000; Pinto et al., 2001; Salatino et
al., 2005.
Compostos voláteis (sesquiterpenos) ácidos
graxos, aminoácidos
Bankova et al., 2000.
Oligoelementos
alumíno, vanádio, ferro, cálcio, silício,
manganês, estrôncio,
Bankova et al., 2000; Pinto et al., 2001.
Vitamina B1, B2, B6, E, C e Hidrocarbonos Bankova et al., 2000.
Tabela 2. Compostos químicos da própolis verde do Brasil.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
58
2.5.6.3. Atividades Biológicas
Já foram constatadas várias atividades biológicas relacionadas a própolis verde brasileira,
como antibacteriana (PARK et al., 1998a; BANKOVA et al., 2000, KOO et al., 2000; MATSUI,
et.al., 2004; AMARAL et al., 2006), antiulcerativa (BARROS, 2007), antifúngica (OTA et al.,
2001; SAWAYA et al., 2002; SANTOS et al., 2005), anti-HIV (PARK et al., 2000); antioxidante
(PARK et al., 1998a; REIS et al., 2000; MATSUI et al., 2004) antiinflamatória (KOO et al., 2000;
REIS et al., 2000), anestésica (KOO et al., 2000), hepatoprotetora (MISHIMA et al., 2005;
BANKOVA et al., 2000) citotóxica e antitumoral (BANKOVA et al., 2000; KUMAZWA et al.,
2003; KOO et al., 2000; MISHIMA et al., 2005; PHARMANECTAR...). As atividades
antibacteriana e citotóxica são atribuídas principalmente aos prenilados derivados do ácido p-
cumárico, ao ácido diterpênico e as lignanas, e a antitumoal ao artepelin C e flavonóides
(BANKOVA et al., 2000; KUMAZAWA et al., 2003). Além disso, a atividade biológica dos
flavonóides também está relacionada à capacidade de proteção celular contra radicais livres
(RUSSO et al., 2002). Atividades imunomodulatória e antihepatotóxica são atribuídas ao ácido
cafeiolquínico (BANKOVA et al., 2000).
Há comprovações da eficácia antifúngica da própolis verde de Minas Gerais por inibição de
candidíase oral (SANTOS et al., 2005) e antibacteriana por regressão de 95% de leões de
gengivites (AMARAL et al., 2006), como também da própolis de São Paulo (in vitro e in vivo) em
relação a Candida albicans, C. tropicalis, C. krusei e C. guilliermondi, C. glabrata, C.
parapsilosis e C. lusitaneae (OTA et al., 2001; SAWAYA et al., 2002).
2.5.6.4. Extratos Alcoólico e Aquoso
O extrato etanólico da própolis verde brasileira é rico em ácidos fenólicos, principalmente
o artepelin C e contém maior concentração de flavonóides quando comparando com o extrato
aquoso. O primeiro composto proporciona o sinergismo das ações que incluem antitumoral,
citotóxica, antibacteriana, antifúngica e antiinflamatória. A atividade antibacteriana, nesse caso,
está relacionada a bactérias gram positivas como Staphylococcus aureus e Streptococcus sp. e
gram negativas, como Escherichia coli e a antifúngica, para Candida sp. Já ao artepelin C é
atribuída a atividade antitumoral, citotóxica e antibacteriana, neste último caso, em relação a
Helicobacter pylori, principal agente causador da gastrite (PHARMANECTAR...).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
59
O extrato alcoólico da própolis verde do Brasil contém derivados prenilados do ácido p-
cumárico, ácido cinâmico, artepelin C e ácido felúrico como os principais ácidos fenólicos e,
pinobanskina, pinocembrina, canferol, isosacuretina, quercetina, galangina e canferide como os
principais flavonóides. Entre esses, chamam a atenção à elevada concentração de derivados
prenilados do ácido p-cumárico e de canferide (PARK et al., 2002a).
O extrato aquoso da própolis verde brasileira contém como principais constituintes o ácido
cafeiolquínico e derivados (ácido 3,4-di-cafeiolquínico e ácido 3,5-di-O-cafeiolquínico) como
também o ácido cinâmico e derivados (ácido p-cumárico, artepelin C, drupanina e bacarina)
(NAKAJIMA et al., 2007).
Pinto et al. (2001) avaliaram o efeito do extrato alcoólico da própolis verde de Minas
Gerais sobre bactérias patogênicas isoladas do leite de vacas com mastite, da raça Holandesa, de
uma propriedade da zona da mata de Minas Gerais. A própolis utilizada era proveniente de uma
propriedade rural localizada no Município de Itapecerica (MG), de cor verde e odor balsâmico,
cedida pelo proprietário. Os resultados in vitro demonstraram que o extrato alcoólico inibiu o
crescimento de bactérias gram positivas (Staphylococcus aureus, Staphylococcus sp. coagulase
negativo e Streptococcus agalactiae) na concentração de 100mg/mL pela técnica do antibiograma
em discos de papel de filtro. O extrato aquoso não inibiu nenhuma amostra bacteriana, assim como
o veículo alcoólico puro utilizado como controle. De forma semelhante, Park e Ikegaki (1998c) já
haviam constatado que o extrato aquoso da própolis de São Paulo não exibiu atividade
antibacteriana sobre bactérias gram-positivas e que quando o solvente utilizado possuía uma
porcentagem de água superior a 50%, menor era a atividade antibacteriana do respectivo extrato.
Os autores concluíram que os componentes ativos da própolis responsáveis pela antibacteriana na
própolis não são solubilizados pela água por ser solvente de alta polaridade.
Mello et al. (2006) constataram, segundo procedimento do CLSI, que o extrato etanólico da
própolis verde brasileira industrializada pela Pharma Néctar® inibiu a formação do tubo de
germinação de uma amostra de Candida albicans (ATCC 18804). A ausência do tubo de
germinação, observado por microscopia de luz, foi constatado na concentração de 0,33µg/mL. Por
microscopia eletrônica, foi possível observar hiperplasia e alterações de superfície celular como
destacamento da parece celular na concentração de 0.43µg/mL, indicando que essa própolis atua
em nível de parede celular promovendo aumento da permeabilidade celular e conseqüente ruptura
da membrana com perda de material celular.
Extratos etanólicos da própolis verde brasileira de Minas Gerais demonstraram atividade
antimicrobiana em relação a Staphylococcus aureus, mas extrato aquoso não apresentou essa

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
60
atividade (PARK et al., 1998a). Além disso, também foi observada atividade antioxidante para o
extrato alcoólico (MATSUI et al., 2004).
O ácido cafeoilquínico e seus derivados possuem forte ação antioxidante, sequestrante de
radicais livres, inibição do crescimento de células cancerígenas na leucemia mielóide, indução
daapoptose dessas células e favorecimento da diferenciação celular de granulócitos (BANKOVA
et al., 2000; MISHIMA et al., 2005), ação protetora hepática e das células pancreáticas e
citotóxica, acelerando a redução dos tumores. Foi verificado aumento da eficácia da radioterapia
em cobaias após pré-tratamento com própolis, além de ser observado menor ocorrência de efeitos
adversos em conjunto à estimulação das células sanguíneas (PHARMANECTAR...).
Em geral, a maioria dos compostos da própolis nativa é lipofílica e os mesmos são mais
facilmente extraídos por etanol, por isso, o extrato alcoólico é alvo de várias pesquisas. Isso não
ocorre com o extrato aquoso, embora seja mais facilmente absorvido do que o extrato alcoólico e
apresente componentes importantes como ácido cafeoilquínico. Como a maioria dos compostos da
própolis verde brasileira é hidrofílica, a extração aquosa é mais eficiente nessa própolis do que nas
originárias de outras regiões (MATSUI et al., 2004). Mas, sabe-se que o extrato etanólico
apresenta de um modo geral, maior rendimento de compostos fenólicos e promove mais atividades
biológicas como antimicrobiana, antioxidante (ADELMANN, 2005), antiinflamatória (KOO et al.,
2000; REIS et al., 2000), anestésica e citotóxica (KOO et al., 2000).
2.5.7. Própolis Vermelha
2.5.7.1. Fonte Botânica
Recentemente foi encontrada no Brasil a própolis vermelha em colméias localizadas no
caule de arbustos de manguezais, como também ao longo da costa dos rios e mar dos estados da
Paraíba, Pernambuco, Alagoas, Sergipe e Bahia (Brasil). Foi observado que as abelhas coletavam
o exsudato vermelho da superfície dos buracos feitos por insetos no tronco de Dalbergia
ecastophyllum, conhecida como rabo-de-bugio, sendo essa a origem botânica dessa própolis
(DAUGSCH et al., 2007; SILVA et al., 2007).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
61
2.5.7.2. Composição Química
Silva et al. (2007) relataram que a composição química da própolis vermelha do Brasil
também é variável dependendo da biodiversidade da fonte botânica, mas é notável a
predominância de flavonóides. Trusheva et al. (2006) constataram a presença de elemicina,
eugenol, álcool triterpênico (α-amyrin, β-amyrin, cicloartenol, lupeol) e isoflavonóides
(isosativan, medicarpin) na própolis vermelha de Cuba.
Para a própolis vermelha da região nordeste do Brasil, há relatos da composição química
do extrato alcoólico, com predomínio de flavonóides, entre os quais podem ser citados:
liquiritigenina, daidzeína, dalbergina, isoliquiritigenina, formononetina e biochanina A (DAUGSH
et al., 2007). Foram encontrados outros flavonóides presentes na fonte botânica e na própolis, tais
como rutina, pinobanskina, quercetina, luteolina, pinocembrina, entre outros (KANAZAWA et al.,
2003) (Tabela 3).
Silva et al. (2007) encontraram ácido felúrico, quercetina, crisina e medicarpin em amostras
de própolis vermelha de Alagoas.
2.5.7.3. Atividades Biológicas
Em Cuba, atividades biológicas da própolis vermelha são relatadas há mais de 17 anos,
entre as quais podem ser mencionadas: sequestrante de radicais livres e hepatoprotetora
(GONZALEZ et al., 1994; PASCUAL et al., 1994; GONZALEZ et al., 1995).
Aos flavonóides presentes na própolis vermelha são atribuídas atividades terapêuticas
como antitumoral, hepatoprotetora e antimicrobiana. Foi descoberto que isoliquritigenina inibe o
crescimento de câncer prostático (KANAZAWA et al., 2003) e que liquiritigenina e
isoliquirigenina inibem a xantina oxidase. A inibição da xantina oxidase foi sugerida para uso no
tratamento de hepatites e tumores cerebrais, porque aumenta o nível de xantina oxidase sérica
(KONG et al., 2000).
As amostras de própolis vermelha da região nordeste do Brasil apresentaram atividade
antimicrobiana para Staphylococcus aureus (ATCC 25923), em concentrações próximas a
2.5µg/mL, que variaram a depender da prevalência de D. ecastophyllum nas regiões (DAUGSCH,
et al., 2007).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
62
COMPOSTO REFERÊNCIA BIBLIOGRÁFICA
Flavonóides
pinocembrina, pinobanskina Daugsch et al., 2007.
isovatin, medicarpin
(isofalvonoides)
Silva et al., 2007.
rutina, luteolina,
formononetina
Dugsch et al., 2007.
Quercetina Daugsch et al., 2007; Silva et al., 2007.
Crisina Silva et al., 2007.
Liquititigenina Donnelly et al., 1973; Kong et al., 2000; Daugsch et al., 2007.
daidzeína, fomononetina,
diocanina A (isoflavonoides)
Donnelly et al., 1973; Daugsch et al., 2007.
Dalbergina Donnelly et al., 1973; Daugsch et al., 2007.
Isoloquitigenina Kong et al., 2000; Kanazawa et al., 2003; Daugsch et al., 2007.
Ácido fenólico
ácido felúrico Silva et al., 2007.
Tabela 3. Compostos químicos da própolis vermelha do Brasil.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
63
2.6. LECTINAS
Lectinas são proteínas ou glicoproteínas que se ligam especificamente e reversivelmente a
mono e oligossacarídeos livres ou na forma de glicoconjugados (glicoproteína e glicolipídeo).
Dessa forma, combinam-se com moléculas e estruturas biológicas que contem esses açucares sem
alterar a estrutura covalente das ligações glicosídicas nos sítios de ligação (MOREIRA et al., 1996;
MOREIRA et al., 1997; CAVADA et al., 1998, LIS; SHARON, 1998; DAMICO et al., 2003;
CHEN et al., 2005; CHUMKHUNTHOD et al., 2006; WONG et al., 2006). Algumas são
metaloproteínas, necessitando de Ca2+, Mn2+ e Mg2+ para completo funcionamento (CAVADA et
al., 1998).
A palavra tem origem do latim legere (selecionar) e leva em consideração a capacidade
hemaglutinante seletiva das lectinas para um tipo particular de grupo sangüíneo humano.
Entretanto, desde que o termo deixou de ser restrito a isso, o significado original foi esquecido.
Para substituir, usa-se o termo aglutinina que é usado como sinônimo de lectina e mais adequado,
pois aglutina eritrócitos e outras células (PEUMANS; VAN DAMME, 1995).
São encontradas amplamente em animais, plantas, fungos, bactérias e vírus (GRIFFIN,
1993; PEUMANS; VAN DAMME, 1995; WANG et al., 1996; CAVADA et al., 1998; LIS;
SHARON, 1998; NGAI; NG 2004; XIA; NG, 2005; CHUMKHUNTHOD et al., 2006) entretanto
essas substâncias estão presentes em maior quantidade em grãos de leguminosas e gramíneas
(BARONDES, 1981; GRIFFIN, 1993; MOREIRA et al., 1996; RAMOS et al., 1996; CAVADA et
al., 1998; CAVADA et al., 2000; YE et al., 2001; YE; NG, 2001; LORIS, 2002; NG et al., 2002;
SHARON; LIS, 2004; ALENCAR et al., 2005b). As sementes de leguminosas como as de feijão e
ervilha são fontes ricas de lectinas as quais tem sido amplamente caracterizada em relação a
propriedades química, fisicoquímica, estrutural e biológica (GOZIA et al., 1993; MOREIRA et al.,
1993; CAVADA et al., 1996; MOREIRA et al., 1996; GRANGEIRO et al., 1997; NAPIMOGA et
al., 2007). As partes das plantas mais susceptíveis ao ataque de predadores, como por exemplo as
sementes, são as que contem lectinas em maior concentração. Com exceção de quitinases,
glucanases e glicosidades, as lectinas são as únicas proteínas capazes de reconhecer e ligar-se a
glicoconjugados presentes na superfície de bactérias e fungos ou expostos no trato intestinal de
insetos e mamíferos. Além disso, sabe-se da capacidade de estabilidade da lectina em condições
desfavoráveis, a qual se mantém viável em ampla faixa de pH, elevada temperatura e na presença
de proteases de animais e insetos. Isso leva a crer na função de defesa dessa classe de proteína
(PEUMANS; VAN DAMME, 1995, YE; NG, 2001).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
64
As lectinas de plantas podem sem agrupadas de acordo com a especificidade de ligação
com o açúcar e com as características estruturais. De acordo com a especificidade por
carboidratos, podem ligar-se a D-manose/D-glicose, D-manose, N-acetilglicosamina, N-
acetilgalactosamina, galactose e ácido siálico (PEUMANS; VAN DAMME, 1995; MOREIRA et
al., 1996; NGAI; NG, 2004; WONG; NG, 2005; WONG et al., 2006). De acordo com as
características estruturais, os três maiores grupos são merolectinas, hololectinas e quimerolectinas.
Merolectinas são proteínas que consistem exclusivamente de único domínio de ligação a
carboidrato. Devido ao caráter monovalente, as merolectinas são incapazes de precipitar
glicoconjugados ou aglutinar células (PEUMANS; VAN DAMME, 1995). Exemplo desse grupo
são as haveínas, proteína do látex de seringueira (Hevea brasiliensis), que se liga à quitina. (VAN
PARIJS et al., 1992). Hololectinas são compostas exclusivamente de domínios de carboidratos
ligantes, mas com dois ou mais sítios de ligação, podendo ser idênticos ou parecidos. Por serem di
ou multivalente, podem aglutinar células e/ou precipitar glicoconjugados, comportando-se como
aglutininas e compreende a maioria das lectinas de plantas, pois se comportam como
hemaglutininas. Quimerolectinas são compostas de um ou mais domínios de carboidratos ligantes
e um domínio não relacionado com uma atividade catalítica bem definida e que age independente
dos domínios de carboidratos ligantes. Dependendo do número de sítios de ligação a carboidratos,
as quimerolectinas comportam-se como merolectinas ou hololectinas. Representantes deste grupo
são as proteínas inativadoras de ribossomos (RIP tipo 2), como por exemplo a ricina (toxina da
mamona), que possui dois domínios de ligação para carboidratos comportando-se como
hololectina e um domínio para a inativação do ribossomo (PEUMANS; VAN DAMME, 1995)
(Figura 2). Essa classificação foi ampliada com o conceito de superlectinas, as quais possuem dois
ou mais domínios ligantes a carboidratos, domínios esses com especificidade para açúcares
diferentes (não-relacionados). Posteriormente foi elaborado o termo multilectinas, que são aquelas
que possuem dois ou mais domínios ligantes a carboidratos, idênticos, mas que podem se ligar a
açúcares diferentes (MONTEIRO-MOREIRA, 2002).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
65
Durante muito tempo, as lectinas são analisadas pela participação em diversas atividades
biológicas como antiproliferativa (WANG et al., 1995; WONG; NG, 2003), antitumoral (WANG
et al., 1996; ABDULLAEV; MEJIA, 1997; WANG et al., 2000; NGAI; NG, 2004),
imunomodulatória (WANG et al., 1996; SHE et al., 1998; WANG et al., 2002; RUBINSTEIN et
al., 2004), estimulação de macrófagos, acumulação de leucócitos (RODRIGUEZ et al., 1992),
indução da produção de interferon δ por linfócitos humanos (BARRAL-NETTO et al., 1992),
liberação de histamina (GOMES et al., 1994), efeitos edematogênicos e migração celular (BENTO
et al., 1993), antifúngica (CIOPRAGA et al., 1999; YE et al., 2001; DAMICO et al., 2003;
TRINDADE et al., 2006), antibacteriana (SEQUEIRA; GRAHAM, 1977; AYOUBA et al., 1994),
anti-HIV (PEUMANS; VAN DAMME, 1995; OOI et al., 2000; WANG; NG, 2001; YE et al.,
2001; WONG; NG, 2003; BARRIENTOS; GRONENBORN, 2005; WONG; NG, 2005),
antiinseto (PEUMANS; VAN DAMME, 1995), armazenamento e transporte de carboidratos e
proteínas, armazenamento de nitrogênio, defesa da planta (PEUMANS; VAN DAMME, 1995;
RÜDIGER, 1997; NOMURA et al., 1998), entre outras.
Lectinas com propriedades antifúngicas já foram isoladas de sementes de leguminosa como
feijão Phaseolus vulgaris (YE et al., 2001); de frutas como pitomba Talisia esculenta (FREIRE et
al., 2002), jaca Artocarpus integrifolia, fruta-pão A. incisa (TRINDADE et al 2006) e graviola
Annona muricata (DAMICO et al., 2003); de fungos como Kluyveromyces bulgaricus (VIARD et
Figura 2. Representação esquemática de três tipos de lectinas de plantas: merolectina, hololectina, quimerolectina (Peumans e van Dame, 1995).
MEROLECTINA HOLOLECTINA QUIMEROLECTINA
quitinase – classe I
RIP – tipo II
domínio de ligação ao carboidrato
domínio catalítico
domínio de inativação ribossômica

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
66
al., 1993) e Schizophyllum commune (CHUMKHUNTHOD et al., 2006); de urtiga Urtica dioica
(BROCKAERT et al., 1989), de batata (GOZIA et al., 1993), de germe de trigo (CIOPRAGA et
al., 1999) e de erva chinesa como Astragalus mongholicus (YAN et al., 2005). Essas lectinas são
denominadas, respectivamente: lectinas de Phaseolus vulgaris, TEL, “jackin”, “frutackin”, AML,
Kb-CWL I, SCL, UDA, lectina de batata, WGA, AMML.
A atividade antifúngica de lectinas já foi constatada em concentrações que variaram entre
100µg/mL a 10.58mg/mL, para diferentes espécies de fungos. Há relatos da atividade antifúngica
de lectinas em relação a leveduras como Sacchromyces cerevisiae (VIARD et al., 1993; FREIRE
et al., 2002; TRINDADE et al., 2006), Candida albicans, C. glabrata, Kluyveromyces bulgaricus,
K. lactis, Pichia pastoris, Rhodotorula mucilaginosa, Sacchromyces bayanus, S. uvarurn,
Schizosacchamyces bulgaricus, Schizosacchamyces octosporus (VIARD et al., 1993),
Trichoderma viridae, T. hamatum (BROCKAERT et al., 1989) e em relação a fungos filamentosos
como Aspergillus niger (CHUMKHUNTHOD et al., 2006), Fusarium oxysporum (CIOPRAGA et
al., 1999; YE et al., 2001; FREIRE et al., 2002; DAMICO et al., 2003; YAN et al., 2005), F.
moniliforme (TRINDADE et al., 2006), F. graminearum (CIOPRAGA et al., 1999), F. solani
(DAMICO et al., 2003), Colletorichum sp. (YAN et al., 2005), C. lindemuthianum (BROCKAERT
et al., 1989; FREIRE et al., 2002), C. musae (DAMICO et al., 2003), Phoma betae, Phycomyces
blakesleeanus, Phytophthora erythroseptica, Septoria nodorum (BROCKAERT et al., 1989),
Botrytis cinerea (BROCKAERT et al., 1989; YAN et al., 2005), Coprinus comatus, Rhizoctonia
solani (YE et al., 2001), Drechslera turcia (YAN et al., 2005).
A atividade antifúngica de lectinas pode variar, dependendo da espécie analisada. Yan et al.
(2005) não constataram atividade antifúngica da lectina AMML, com afinidade de ligação para
galactose e derivados, em relação a Rhizoctonia solani e Mycospharella arachidicola na
concentração de até 200µg/mL. Chumkhunthod et al. (2006) também não observaram essa
atividade para lectina SCL com afinidade de ligação para N-acetilgalactosamina, em relação a S.
cerevisiae, Candida albicans, Penicillium diditatum na concentração de até 1mg/ml. Entretanto
esses autores constataram atividade antifúngica dessas mesmas lectinas para outras espécies de
fungos, como mencionado anteriormente. Os exemplos de lectinas, a respectiva especificidade por
carboidratos e a atuação sobre algumas espécies de fungos estão sumarizados na Figura 3.
A inibição do crescimento de fungos pela ação de lectinas, parece ser resultante da inibição
da germinação de esporos como também do crescimento do micélio (GRIFFIN, 1993). O exato
mecanismo de atuação ainda não foi elucidado, mas parece haver alteração na síntese da parede
celular fúngica devido a modificação na síntese de quitina, por deficiência da deposição na parede
celular. Lectinas que se ligam à quitina demonstram importante efeito antifúngico, entretanto, a

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
67
ocorrência ou não dessa alteração é dependente da combinação do fungo com a lectina (GRIFFIN,
1993; PEUMANS; VAN DAMME, 1995).
A parede celular dos fungos é altamente versátil, sendo continuamente expandida durante o
crescimento e extensivamente remodelada durante o desenvolvimento. Quimicamente, a parede
contém entre 80 e 90% de polissacarídeos, sendo o restante constituído de proteínas e lipídeos. Em
algumas espécies são encontrados quantidades apreciáveis de pigmentos (melanina), polifosfatos e
íons inorgânicos. Fisicamente, a parede celular é formada por microfibrilas entrelaçadas, embebida
numa matriz amorfa. Quitina e celulose são os principais componentes microfibrilantes da maioria
dos fungos filamentosos, enquanto que nas leveduras a parede contém principalmente glucanas
não celulósicas. Proteínas e vários polissacarídeos (glucanas, mananas, galactanas e
heteropolissacarídeos) são as principais substâncias cimentantes. Os monossacarídeos mais
freqüentemente encontrados na parede celular dos fungos são: D-glicose, N-acetil-D-glicosamina,
D-manose, D-galactose, D-galactosamina, L-fucose, D-glicosamina, xilose e ácido D-glicurônico.
Desses, glicose, N-acetil-D-glicosamina e manose são encontrados na maioria dos fungos.
Ocasionalmente ramnose, ribose e arabinose podem ser observadas (KITAJMA, 2000).
Entretanto, algumas lectinas de leguminosas não apresentam essa atividade para Fusarium
oxysporum, Botrytis cinerea, Mycosphaerella arachidicola e Coprinus comatus (YE; NG, 2001;
NG et al., 2002; WONG; NG, 2003, 2005; WONG et al., 2006) (Figura 4). Outras, podem
apresentar discreto efeito ativador da cinética do crescimento do fungo em cultivo (VIARD et al.,
1993; PÉREZ-SANTIAGO et al., 2000; HERNÁNDEZ et al., 2003).
Viard et al. (1993) observaram efeito estimulatório da lectina Kb-CWL I com
especificidade de ligação a galactose, sobre Candida albicans e C. tropicalis nas concentrações
entre 0.04 e 0.08mg/mL, havendo aceleração para formação do tubo germinativo como também
indução da formação de micélio.
Pérez-Santiago et al. (2000) verificaram a capacidade de inibição e estimulação do
crescimento de Ustilago maydis (basidiomyceto) por lectinas com especificidade para ligação a N-
acetilgalactosamina, N-acetilglicosamina e galactose, na concentração de 100µg/mL. A única
lectina que inibiu a germinação de esporos e o desenvolvimento do fungo foi a CCL, extraída de
Zea mays (milho), com especificidade de ligação por galactose. As lectinas DBA e PLA extraídas
de Dolichos biflorus e Phaseolus lunatus (feijão lima, comum no México e Argentina)
respectivamente, com especificidades por N-acetilgalactosamina, e a lectina WGA com
especificidade pode N-acetilglicosamina, obtida de Triticum vulgaris (trigo), inibiram a
germinação desse fungo mas não houve inibição do desenvolvimento do micélio nem alterações

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
68
morfológicas. A lectina ConA, obtida de Canavalia ensiformis (feijão-de-porco) e capaz de ligar-
se a glicose e manose, estimulou a germinação dos esporos e o crescimento fúngico.
Hernández et al. (2003) constataram que a ConA tem efeito ativador sobre basidiósporos de
Ustilago maydis após nove horas de incubação, apresentando acentuada capacidade de agregação,
ramificações múltiplas e germinação. Além disso, foram observadas várias alterações
morfológicas como, por exemplo, espessamento da parede celular e deformações no basidiósporo.
Does et al. (1999) avaliaram a atividade antifúngica de UDA e demonstraram que a
inibição do crescimento Botrytis cinerea, Trichoderma viride e Colletotrichum lindemuthianum é
temporária, detectável após seis horas de incubação com a lectina 67µg/mL. Após esse período,
não houve inibição do crescimento fúngico, mesmo na máxima concentração de lectina
(300µg/mL). Esses autores sugerem que o fungo desenvolve algum mecanismo de adaptação.
Canavalia brasiliensis, Dioclea violacea e D. rostrata são leguminosas das quais são
extraídas as lectinas ConBr, DVioL e DRL, respectivamente (CAVADA et al., 1996; BARBOSA
et al., 2001), as quais têm afinidade por glicose e manose (CAVADA et al. 1993, CAVADA et al.,
1996). Essas lectinas já demonstraram indução da produção de histamina em ratos (GOMES et al.,
1994), produção de óxido nítrico (ANDRADE et al., 1999), além do efeito protetor in vivo contra
infecção por Leishmania amazonensis em camundongos BALB/c (BARRAL-NETTO et al.,
1996), estimulação linfocitária em humanos (BARRAL-NETTO et al., 1992), estimulação da
produção de macrófagos e linfócitos em camundongos C3H/HeJ (RODRIGUEZ et al., 1992).
Lonchocarpus sericeus é uma leguminosa da qual é extraída a lectina LSL (NAPIMOGA et
al., 2007) com afinidade por N-acetilglicosamina. Já demonstrou atividade antiinflamatória em
casos de peritonite em ratos, notável diminuição de bactérias que colonizavam essa região
peritoneal (ALENCAR et al., 2005b) e inibição da migração de neutrófilos em processos
inflamatórios (NAPIMOGA et al., 2007).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
69
Origem da
lectina Nome da lectina
Carboidrato de ligação Fungo susceptível Referência bibliográfica
Phaseolus vulgaris (feijão vermelho)
lectina de Phaseolus vulgaris
glicoproteínas: lactoferrina, ovoalbumina e tiroglobulina
Fusarium oxysporum Coprinus comatus Rhizoctonia solani
Ye et al., 2001
semente de Talisia esculenta (pitomba)
TEL glicose, manose, N-acetilglicosamina
Sacchromyces cerevisiae Fusarium oxysporum Colletotrichum lindemuthianum
Freire et al., 2002
Artocarpus integrifolia (jaca) A. incisa (fruta-pão)
jackin
frutackin
N- acetilglicosamina S. cerevisiae Fusarium moniliforme
Trindade et al., 2006
Semente de Annona muricata (graviola)
AML glicose, manose Fusarium oxysporum Fusarium solani Colletotrichum musae
Damico et al., 2003
Kluyveromyces bulgaricus (levedura)
Kb-CWL I Galactose Sacchromyces cerevisiae S. bayanus, S. uvarurn Candida albicans, Candida glabrata Rhodotorula mucilaginosa Pichia pastoris Kluyveromyces lactis K.bulgaricus Schizosacchamyces bulgaricus Sch. Octosporus
Viard et al., 1993
Schizophyllum commune (basidiomiceto)
SCL N-acetilgalactosamina Aspergillus niger Chumkhunthod et al., 2006
Urtica dioica (urtiga)
UDA N- acetilglicosamina Trichoderma viride T. hamatum Colletorichum lindemuthianum Botrytis cincerea Phoma betae Phycomyces blakesleeanus Septoria nodorum Phytophytora erythroseptica
Brockaert et al., 1989
Germe de trigo WGA
N- acetilglicosamina Fusarium oxysporum F. graminearum
Ciopraga et al., 1999
Astragalus mongholicus (erva chinesa)
AMML Galactose Colletorichum sp. Drechslera turcia F.oxysporum, Botrytis cincerea
Yan et al., 2005
Figura 3. Lectinas com atividade antifúngica e especificidade de ligação aos carboidratos.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
70
Origem da lectina Carboidrato de ligação Fungo analisado Referência bibliográfica
Pisum sativum var. macrocarpon (ervilha)
glicose, manose Fusarium oxysporum Rhizoctonia solani Botrytis cinerea Mycosphaerella arachidicola
Ye; Ng, 2001
Castanea mollisima (castanha chinesa)
glicose, manose Coprinus comatus, Mycosphaerella arachidicola Botrytis cinerea
Ng et al., 2002
Vigna sesquipedalis (feijao da terra)
ácido poligalacturônico Fusarium oxysporum Rhizoctonia solan Coprinus comatus
Wong; Ng, 2003
Canavalia gladiata
glicose, manose, ramnose Botrytis cinerea Mycosphaerella arachidicol Fusarium oxysporum
Wong; Ng, 2005
Feijão malhado Galactose Botrytis cinerea Mycosphaerella arachidicola Fusarium oxysporum
Wong et al., 2006
Figura 4. Lectinas sem atividade antifúngica e especificidade de ligação aos carboidratos.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
71
3. OBJETIVOS
3.1 OBJETIVO GERAL
Caracterizar amostras Microsporum canis, Microsporum gypseum, Trichophyton
tonsurans, Trichophyton mentagrophytes, Trichophyton rubrum e Epidermophyton floccosum,
preservadas na Coleção de Culturas Micoteca – University of Recife Mycology (URM) quanto ao
perfil enzimático e avaliar a atividade antifúngica própolis e lectinas.
3.2 OBJETIVOS ESPECÍFICOS
Confirmar as características clássicas de amostras de dermatófitos através de aspectos
macroscópicos, microscópicos e fisiológicos;
Detectar a atividade de proteases, queratinases, colagenases, fosfolipases e lipases;
Correlacionar aspectos taxonômicos e perfil enzimático de amostras dermatófitos preservadas;
Avaliar in vitro a atividade antifúngica do extrato alcoólico e aquoso da própolis verde e do
extrato alcoólico da própolis vermelha em relação a espécies de dermatófitos;
Avaliar in vitro a atividade antifúngica de lectinas de leguminosas em relação a espécies de
dermatófitos

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
72
4. REFERÊNCIAS BIBLIOGRÁFICAS ABDEL-GAWAD KM. Mycological and some physiological studies of keratinophilic and other moulds associated with sheep wool. Microbiological Research. v.152, n.2, p.181-188, jul. 1997. ABDEL-RAHMAN, S.M. Polymorphic exocellular protease expression in clinical isolates of Trichophyton tonsurans. Mycopathologia, v. 150, n. 3, p. 117–120, jun. 2000. ABDEL-RAHMAN, S.M. Trichophyton tonsurans exocellular protease expression: correlation with clinical presentation in tinea capitis. Clinical and Experimental Dermatology, v. 27, n. 4, p. 268-271, jun. 2002. ABDULLAEV, F.I.; MEJIA, E.G. Antitumor effect of plant lectins. Natural Toxins, v. 5, n. 4, p. 157-163. 1997. ACKERMANN, T. Fast Chromatographic study of propolis crudes. Food Chemistry, v. 42, n. 2, p. 135-138. 1991. ADELMANN, J. Própolis: variabilidade composicional, correlação com a flora e bioatividade antimicrobiana / antioxidante. 2005. 186f: il. color, tb. Dissertação (Mestrado em Ciências Farmacêuticas) – Universidade Federal do Paraná, Curitiba, 2005. AFOLAYAN, A.J.; MEYER, J. J. The antimicrobial activity of 3,5,7- trihydroxyflavone isolated from the shoots of Helichrysum aureonitens. Journal of Ethnopharmacology, v. 57, n. 3, p. 177–181, ago. 1997. AFST-EUCAST: Reference Method for Determination of Minimal Inhibitory Concentration (MIC) by Broth Dilution of Fermentative Yeasts. Discussion document E. Dis. 7.1 ESCAMID. Subcommittee of Antifungal Susceptibility Testing of the European Committee on Antibiotic Susceptibility Testing of the European Society of Clinical Microbiology and Infectious Diseases. Taufkirchen. 2002. AGA, H.; SHIBUYA, T.; SUGIMOTO, T.; KURIMOTO, M.; NAKAJIMA, S.H. Isolation and identification of antimicrobial compounds in Brazilian propolis. Bioscience, biotechnology, and Biochemistry, v. 58, n.5, p. 945-946. 1994 AGUIAR, C.L.; ALENCAR, S.M.; PAREDES-GUZMÁN, J.F.; KOO, M.H.; PARK, Y.K. Caracterização físico-química das própolis originárias da região de Mata Atlântica do Estado de Alagoas. Mensagem Doce, n.72, jul. 2003.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
73
AGUT, M.; BAYÓ, M.; LARRONDO. J.; CALVO, M. Keratinophilic fungi from soil of Brittany, France. Mycopathologia, v. 129, n. 2, p. 81-82, jun. 1995. AKAO,Y.; MARUYAMA, H.; MATSUMOTO, K.; OHGUCHI, K.; NISHIZAWA, K.; SAKAMOTO, T.; ARAKI, Y.; MISHIMA, S.; NOZAWA, Y. Cell growth inhibitory effect of cinnamic acid derivatives from propolis on human tumor cell lines. Biological & Pharmaceutical Bulletin, v. 26, n. 7, p. 1057-1059. 2003. AL – FOUZAN, A. S.; NANDA, A.; KUBEC, K. Dermatophytosis of children in Kwait: a prospective survey. International Journal of Dermatology, v. 32, n. 11, p. 798-801, nov. 1993. ALENCAR, N.M.; CAVALCANTE, C.F.; VASCONCELOS, M.P.; LEITE, K.B.; ARAGÃO, K.S.; ASSREUY, A.M.; NOGUEIRA, N.A.; CAVADA, B.S.; VALE, M.R. Anti-inflammatory and antimicrobial effect of lectin from Lonchocarpus sericeus seeds in an experimental rat model of infectious peritonitis. Journal of Pharmacy and Pharmacology, v. 57, n. 7, p. 919-922, jul. 2005b. ALENCAR, S.M.; AGUIAR, C.L.; PAREDES-GUZMÁN, J.; PARK, Y.K. Composição química de Baccharis dracunculifolia, fonte botânica das própolis dos estados de São Paulo e Minas Gerais. Ciência Rural, v. 35, n. 4, p. 909-915, jul./ago. 2005a. ALENCAR, V.B.M., ALENCAR, N.M.N., ASSREUY, A.M.S., MOTA, L.M., BRITO,G.A.C., ARAGÃO, K.S., BITTENCOURT, F.S., PINTO, V.P.T., DEBRAY, M., RIBEIRO, R.A., CAVADA, B.S. Pro-inflammatory effect of Arum maculatum lectin and role of resident cells. The International Journal of Biochemistry & Cell Biology, v. 37, n. 9, p. 1805-1814, set. 2005b. ALI – SHTAYEH, M. S.; ARDA, H. M.; ABU – GHDEIB, S. I. Epidemiological study of tinea capitis in schoolchildren in the Anabus area (West Bank). Mycoses, v. 41, p. 243-248, 1998. AMARAL, R.C.; GOMES, R.T.; ROCHA, W.M.S.; ABREU, S.L.R; SANTOS, V.R. Periodontitis treatment with brazilian green propolis gel. Pharmacology online, v. 3, p. 336-341, 2006. ANDERSON, L.; DIBBLE, M. V.; TURKKI, P. R. Nutrição, 17. ed. Rio de Janeiro: Guanabara, 1988. 737 p. ANDRADE, J.L.; ARRUDA, S.; BARBOSA, T.; PAIM, L.; RAMOS, M.V.; CAVADA, B.S.; BARRAL-NETTO, M. Lectin-induced nitric oxide production. Cellular Immunology, v. 194, n. 1, p. 98-102, maio. 1999.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
74
AOKI, S.; ITO-KUWA, S.; NAKAMURA, K.; NINOMIYA, K.; VIDOTT, V. Extracellular proteolytic activity of Cryptococcus neoformans. Mycophatologia, v. 128, n. 3, p. 143-150, dez. 1994. APARECIDO, C. C.; EGYDIO, A. P. M.; FIGUEIREDO, M. B. Avaliação de três métodos para preservação de fungos fitopatogênicos. Summa Phytopathologica, v.27, n.4, p.421-424, 2001. APODACA, G.; McKERROW, J.H. Expression of proteolytic activity by cultures of Trichophyton rubrum. Journal of Medical and Veterinary Mycology, v. 28, n. 2, p. 159-171. 1990. AQUINO, V.R.; CONSTANTE, C.C.; BAKOS, L. Freqüência das dermatofitoses em exames micológicos em Hospital Geral de Porto Alegre, Brasil. Anais Brasileiros de Dermatologia, v. 82, n. 3, p. 239-244. 2007. ASTE, N.; PAU, M.; BIGGIO, P. Tinea capitis in children in the district of Caligari, Italy. Mycoses, v. 40, p. 231-233, 1997. ASAHI, M.; LINDQUIST, R.; FUKUYAMA, K.; APODACA, G.; EPSTEIN, W.L.; MCKERROW, J.H. Purification and characterization of major extracellular proteinases from Trichophyton rubrum. The Biochemical Journal, v. 232, n. 1, p. 139-144, nov. 1985. ATKINSON, P.; BLAKEMAN, J.P. Seasonal occurrence of an antimicrobial flavanone, sakuranetin, associated with glands on leaves of Ribes nigrum. New Phytologist, v. 92, n.1, p. 63-74, set. 1982. AUNSTRUP, K. Industrial production of proteolytic enzymes. In: SPENCER, B. Industrial Aspects of Biochemistry. Federation of European Biochemical Societies, p. 23-46. 1974. AUBAID, A.H.; MUHSIN, T.M. Partial purification and kinetic studies of exocellular proteinase from Trichophyton mentagrophytes var. erinacei. Mycoses, v. 41, n. 3-4, p. 163-168, mar./apr. 1998. AYOUBA, A.; CAUSSE, H.; VAN DAMME, E.J.M.; PEUMANS, W.J.; CAMBILLAU, C.; ROUGÉ, P. Interactions of plant lectins with the components of the bacterial cell wall peptidoglycan. Biochemical systematics and ecology, v. 22, n. 2, p. 153-159. 1994. AZULAY, D. R. Dermatofitoses. Anais Brasileiros de Dermatologia, v. 64, suppl. 1, p. 93-96, 1989.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
75
BAKER, M.; JEFFRIES, P. Use of Commercially Available Cryogenic Vials for Long-Term Preservation of Dermatophyte Fungi. Journal of Clinical Microbiology, v. 44, n. 2, p. 617–618, fev. 2006. BAKERSPIEGEL, A. Soil as a storage medium for fungi. Mycologia, v. 45, p. 596-604. 1953. BANKOVA, S. Recent trends and important developments in propolis research. Evidence-Based Complementary and Alternative Medicine, v. 2, n. 1, p. 29-32, mar. 2005a BANKOVA, V. Chemical diversity of propolis and the problem of standardization. Journal of Ethnopharmacology, v. 100, n. 1/2, p. 114–117, ago. 2005b. BANKOVA, V., BOUDOUROVA-KRASTEVA, G., POPOV, S., SFORCIN, J. M. E FUNARI, S. R. C. Seasonal variations of the chemical composition of Brazilian propolis. Apidologie, v. 29, p. 361-367. 1998. BANKOVA, V.; CHRISTOV, R.; DELGADO TEJERA, A. Lignans and other constituents of propolis from the Canary Islands. Phytochemistry, v. 49, n. 5, p. 1411-1415. 1998. BANKOVA, V.; CHRISTOV, R.; KUJUMGIEV, A.; MARCUCCI M, C.; POPOV, S. Chemical composition and antibacterial activity of Brazilian propolis. Zeitschrift für Naturforschung, v. 50C, p. 167-172. 1995. BANKOVA, V.; POPOVA, M.; BOGDANOV, S.; SABATINI, A.G. Chemical composition of European propolis: expected and unexpected results. Zeitschrift fur Naturforschung, v. 57c, n. 5/6, p. 530-533, maio/jun. 2002. BANKOVA, V.S.; CASTRO, S.L.; MARCUCCI, M.C. Propolis: recent advances in chemistry and plant. Apidologie, v. 31, n. 1, p. 3-15, jan./fev. 2000 BANSKOTA, A.H.; TEZUKA, Y.; PRASAIN, J.K.; MATSUSHIGE, K.; SAIKI, I. KADOTA, S. Chemical constituents of Brazilian propolis and their cytotoxic activities. Journal of Natural Products, v. 61, n. 7, p. 896-900, jun. 1998. BARBOSA, T.; ARRUDA, S.; CAVADA, B.; GRANGEIRO, T. B.; FREITAS, L. A. R.; BARRAL-NETTO, M. In vivo lymphocyte activation and apoptosis by lectins of Diocleinae subtribe. Memórias do Instituto Oswaldo Cruz, v. 96, n. 5, p. 673-678, jul. 2001.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
76
BARCHIESI, F.; ARZENI, D.; CAMILETTI, V.; SIMONETTI, O.; CELLINI, A.; OFFIDANI, AM.; SCALISE, G. In vitro activity of posaconazole against clinical isolates of dermatophytes. Journal of Clinical Microbiology, v. 39, n. 11, p. 4208-4209, nov. 2001. BARONDES, S.H. Lectins: their multiple endogenous cellular functions. Annual Review of Biochemistry, v. 50, p. 207-231. 1981. BARRAL-NETTO, M.; SANTOS, S.B.; MOREIRA, R.A.; OLIVEIRA, J.T.A.; BARRAL, A.; MOREIRA, L.I.M.; SANTOS, C.F.; CAVADA, B.S. Human lymphocyte stimulation by lectins from Brazilian leguminous seeds. Immunological Investigations, v.21, n.4, p. 297-303. 1992. BARRAL-NETTO, M.; VON SOHSTEN, R. L.; TEIXEIRA, M.; SANTOS, W. L.; POMPEU, M. L.; MOREIRA, R. A.; OLIVEIRA, J. T.; CAVADA, B. S.; FALCOFF, E.; BARRAL, A. In vivo protective effect of the lectin from Canavalia brasiliensis on Balb/ mice infected by Leishmania amazonensis. Acta Tropica, v. 60, n. 4, p. 237-250, fev. 1996. BARRIENTOS, L.G.; GRONENBORN, A.M. The highly specific carbohydrate- binding protein cyanovirin-N: structure, anti-HIV/Ebola activity and possibilities for therapy. Mini Reviews in Medicinal Chemistry, v. 5, n. 1, p. 21-31, jan. 2005. BARROS, E.A. Dermatófitos isolados de amostras clínicas de pacientes atendidos no Laboratório de Micologia Médica da Universidade Federal de Pernambuco. 2004. 50f: il color., tb. Trabalho de conclusão de curso (Especialização em Micologia) – Universidade Federal de Pernambuco, Recife, 2004. BARROS, M.E.S.; SANTOS, D.A.; HAMDAN, J.S. In vitro methods for antifungal susceptibility testing of Trichophyton spp. Mycological Research, v. 110, n. 11, p. 1355-1360, nov. 2006. BARROS, M.P.; SOUSA, J.P.B.; BASTOS, J.K.; ANDRADE, S.F. Effect of Brazilian green propolis on experimental gastric ulcers in rats. Journal of Ethnopharmacology, v. 110, n. 3, p. 567-571, abr. 2007. BAZO, A.P.; RODRIGUES, M.A.M.; SFORCIN, J.M.; VIANA DE CAMARGO, J.L.; RIBEIRO, L.R.; SALVADORI, D.M.F. Protective action of propolis on the rat colon carcinogenesis. Teratogenesis, Carcinogenesis, and Mutagenesis, v. 22, n. 3, p. 183-194, abr. 2002. BENTO, C.A.M.; CAVADA, B.S.; OLIVEIRA, J.T.A.; MOREIRA, R.A.; BARJA-FIDALGO, C. Rat paw edema and leucocyte immigration induced by plant lectins. Agents and Action, v. 38, n. 1/2, p. 48-54, jan. 1993.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
77
BERGOLD, A. M.; GEORGIADIS, S. Novidades em fármacos antifúngicos: uma revisão Visão Acadêmica, v. 5, n. 2, p. 159 -172, jul./dez. 2004 BEZERRA, C.C.F.; LIMA, R.F.; LAZERA, M.S.; WANKE, B.; BORBA, C.M. Viability and molecular authentication of Coccidioides immitis strains from culture collection of the Instituto Oswaldo Cruz, Rio de Janeiro, Brazil Revista da Sociedade Brasileira de Medicina Tropical, v. 39, n. 3, p. 241-244, maio/jun. 2006. BIANCHINI, L.; BEDENDO, I.P. Efeito antibiótico da própolis sobre bactérias fitopatogênicas. Scientia Agricola, v. 55, n. 1, jan./abr. 1998. BORBA, C.M.; RODRIGUES, K.F. Viability and sporulating capability of Coelomycetes preserved under a range of different storage regimes. Revista Iberoamericana de Micología, v. 17, n. 4, p. 142-145, dez. 2000. BORBA C. M.; VINHAS E. A.; LOPES-BEZERRA L. M.; LUCENA-SILVA N. Morphological, biochemical and molecular approaches for comparing typical and atypical Paracoccidioides brasiliensis strains. Antonie Van Leeuwenhoek, v. 88, n. 3/4, p.257-66, out./nov. 2005. BORCIC, I.; RADONIC, A.; GRZUNOV, K. Comparisom of the volatile constituents of propolis gathered in different regions of Croatia. Flavour and Fragrance Journal, v. 11, n. 5, p. 311-313, set. 1996. BORMAN, A.M.; SZEKELY, A.; CAMPBELL, C.K.; JOHNSON, E.M. Evaluation of the viability of pathogenic filamentous fungi after prolonged storage in sterile water and review of recent published studies on storage methods. Mycopathologia, v. 161, n. 6, p. 361-368, jun. 2006. BOUDOROVA-KRAVSTEVA, G.; BANKOVA, V.; SFORCIN, J..M.; NIKOLOVA, N.; POPOV, S. Phenolics from Brazilian propolis. Zeitschrift fur Naturforschung, v. 52C, p. 676-679. 1997. BRAZ, S.C.M. Viabilidade, confirmação taxonômica e caracterização enzimática de espécies de Acremonium (Link, 1809) estocados na Coleção de Culturas Micoteca-URM. 2007. Monografia (Graduação em Bacharelado em Biomedicina) - Universidade Federal de Pernambuco, Recife, 2007. BRILHANTE, R.S.; PAIXAO, G.C.; SALVINO, L.K.; DIOGENES, M.J.; BANDEIRA, S.P.; ROCHA, M.F.; DOS SANTOS, J.B.; SIDRIM, J.J. Epidemiology and ecology of dermatophytoses in the City of Fortaleza: Trichophyton tonsurans as important emerging pathogen of tinea capitis. Revista da Sociedade Brasileira de Medicina Tropical, v. 33, n. 5, p. 417-125, sep./oct. 2000.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
78
BRILHANTE, R.S.N.; CAVALCANTE, C.S.P.; SOARES-JÚNIOR, F.A.; MONTEIRO, A.J.; BRITO, E.H.S.; CORDEIRO, R.A.; SIDRIM, J.J.C.; ROCHA, M.F.G. Evaluation of Microsporum canis in different methodos of storage. Medical Mycology, v. 42, n. 6, p. 499-504. dez. 2004. BROCKAERT, W.F.; VAN PARIJS, J.; LEYNS, F.; JOOS, H.; PEUMANS, W.J. A chitin-binding lectin from stinging nettle rhizomes with antifungal properties. Science, v. 245, n. 4922, p. 1100-1103, set. 1989. BRUGUERA, M.T.; ABARCA, M.L.; BRAGULAT, M.R.; CABAÑES, F.J. Determinación de actividades enzimaticas en cepas de Microsporum canis y Microsporum gypseum. Revista Iberoamericana de Micología, v. 14, p. 119-124. 1997. BUENO, L.; GALLARDO, T. Preservación de hongos filamentosos en agua destilada esterilizada. Revista Iberoamericana Medica, v. 15, p. 166-168. 1998. BURDOCK, G.A. Review of the biological properties and toxicity of bee propolis. Food and Chemical Toxicology, v. 36, n. 4, p. 347- 363. 1998. CAFARCHIA, C.; DE LAURENTIS, N.; MILILLO, M.A.; LOSACCO, V.; PUCCINI, V. Antifungal activity of Apulia region propolis. Parassitologia, v. 41, n. 4, p. 587-590, dez. 1999. CALLEJO, A.; ARMENTIA, A.; LOMBARDERO, M.; MARTÍNEZ, C.; REBOLLO, S.; SEDANO, E.; DE LA FUENTE, R.; FERNÁNDEZ, A. Hypersensitivity to propolis. Alergologia e Inmunologia Clinica, v. 16, p. 113-117. 2001. CAMPBELL, M. K. Bioquímica. 3. ed. São Paulo: Artmed, 2003. p. 133. CANHOS, V.P.; UMINO, C.Y.; MANFIO, G.P. Coleções de microrganismos: coleções de cultura de microrganismos. p. 82-101. Disponível em: <http://www.biota.org.br/pdf/v72cap03. pdf> Acesso em: 23 mar. 2007. CARAZO, J.L.S.; LOSADA, L.O.; SANJUAN, Y.V.P. Tratamiento actual de las micosis superficiales. Revista Iberoamericana de Micología, v. 16, p. S26-S30. 1999. CARVALHO, M.B. Fatores de patogenicidade de Trichophyton rubrum (Castellani) Sabouraud isoldados de culturas de pacientes atendidos no Laboratório de Micologia Médica da Universidade Federal de Pernambuco. 2003. 50f: il. color. Monografia (Graduação em Ciências Biológicas) - Universidade Federal de Pernambuco, Recife, 2003.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
79
CASTALDO, S.; CAPASSO, F. Propolis, an old remedy used in modern medicine. Fitoterapia, v. 73, suppl. 1, p. S1–S6, 2002. CASTELLANI A. Maintenance and cultivation of common pathogenic fungi in man in sterile distilled water. Further researches. American Journal of Tropical Medicine and Hygiene, v. 70, p. 181-184. 1967. CASTRO, S.L. Propolis: biological and pharmacological activities. Therapeutic uses of this bee-product. Annual Review of Biomedical Science, v. 3, p. 49–83. 2001. CAVADA, B.S.; GRANGEIRO, T.B.; RAMOS, M.V.; CORDEIRO, E.F.; OLIVEIRA, J.T.A.; MOREIRA, R.A. Isolation and partial characterization of a lectin from Dioclea rostrata Benth seeds. Revista Brasileira de Fisiologia Vegetal, v. 8, n. 1, p. 31-36. 1996. CAVADA, B.S.; MADEIRA, S.V.F.; CALVETE, J.J.; SOUZA, L.A.; BOMFIM, L.R.; DANTAS, A.R.; LOPES, M.C.; GRANGEIRO, T.B.; FREITAS, B.T.; PINTO, V.P.; LEITE, K.B.; RAMOS, M.V. Purification, chemical, and immunochemical properties of a new lectin from Mimosoideae (Parkia discolor). Preparative Biochemistry & Biotechnology, v. 30, n. 4, p. 271-280, 2000. CAVADA, B. S.; MOREIRA, R. A.; OLIVEIRA, J. T. A.; GRANGEIRO, T. B. Primary strutures and functions of plant lectins. Revista Brasileira de Fisiologia Vegetal, v. 5, p. 193-201. 1993. CAVADA, B.S.; SANTOS, C.F.; GRANGEIRO, T.B.; NUNES, E.P.; SALES, P.V.P.; RAMOS, R.L.; SOUSA, F.A.M.; CRISOSTOMO, C.V.; CALVETE, J.J. Purification and characterization of a lectin from seeds of Vatairea macrocarpa duke. Phytochemistry, v. 49, n. 3, p. 675-680, out. 1998. CETINKAYA, Z.; KIRAZ, N.; KARACA, S.; KULAC, M.; CIFTCI, I.H.; AKTEPE, O.C.; ALTINDIS, M.; KIYILDI, N.; PIYADE, M. Antifungal susceptibilities of dermatophytic agents isolated from clinical specimens. European Journal of Dermatology, v. 15, n. 4, p. 258-261, jul./ago. 2005. CHAVES, G.M.; CAVALCANTI, M.A.Q.; PORTO, A.L.F. Pathogenicity characteristics of stocked and fresh yeast strains. Brazilian Journal of Microbiology, v. 34, n. 3, p. 197-202, jul./set. 2003. CHEN, C.H.; WENG, M.; WU, C.H.; L.IN,l J. Comparison of radical scavenging activity, cytotoxic effects and apoptosis induction in human melanoma cells by Taiwanese propolis from different sources. Evidence-Based Complementary and Alternative Medicine, v. 1, n. 2, p. 175–85, ago. 2004.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
80
CHEN, Z.; SUN, X.; TANG, K. Cloning and expression of a novel cDNA encoding a mannose-binding lectin from Dendrobium officinale. Toxicon, v.45, n. 1, p. 535-540, dez. 2005. CHRISTOV, R.; BANKOVA, V.; HEGAZI, A.; ABD EL HADY, F.; POPOV, S. Chemical composition of Egyptian propolis. Zeitschrift für Naturforschung, v.53C, p. 197-200. 1998. CHRISTOV, R.; BANKOVA, V.; TSVETKOVA, I.; KUJUMGIEV, A.; DELGADO TEJERA A. Antibacterial furofuran lignans from Canary Island propolis. Fitoterapia, v.70, n. 1, p. 89-92, fev. 1999. CHUMKHUNTHOD, P.; RODTONG, S.; LAMBERT, S.J.; FORDHAM-SKELTON, A.P.; RIZKALLAH, P.J.; WILKINSON, M.C.; REYNOLDS, C.D. Purification and characterization of an N-acetyl-D-galactosamine-specific lectin from the edible mushroom Schizophyllum commune. Biochimica et Biophysica Acta (General Subjects), v. 1760, n. 3, p. 326–332, mar. 2006 CIOPRAGA, J., GOZIA, O.; TUDOR,R.; BREZUICA, L.; DOYLE, R.J. Fusarium sp. growth inhibition by wheat germ agglutinin. Biochimica et Biophysica Acta (General Subjects), v. 1428, n. 2/3, p. 424-432, ago. 1999. CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI): Método de referência para testes de diluição em caldo para a determinação da sensibilidade a terapia antifúngica dos fungos filamentosos. Norma aprovada, M38-A, v. 22, n. 16. 2002. CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI): Método de referência para testes de diluição em caldo para determinação da sensibilidade de leveduras à terapia antifúngica. Norma aprovada, M27-A, 2. ed., v. 22, n. 15. 2002. COLE, G.T.; SAMSON, R. A. Patterns of development in conidial fungi. London: Pittman, 1979. 190 p. CORDEIRO NETO, F.; PERSSONI, R.A.B.; FIGUEIREDO-RIBEIRO, R.C.L. Fungos produtores de inulinases isolados da rizosfera de Asteraceae herbácea do cerrado. Revista Brasileira de Ciência do Solo, v. 21, p. 149-153. 1997. COSTA, E. F.; WANKE, B.; MARTINS, E. C. S. Micoses Superficiais e Cutânea.Estudo comparativo entre duas populações: Rio de Janeiro(RJ) e Aracaju (SE). Anais Brasileiros de Dermatologia, v. 66, n. 3, p. 119-122, 1991. COSTA, M.; MISEFARI, A.; AMATO, A. Enzymatic activity of Candida albicans in egg yolk containing media. Atti XIV Congresso Nazionale di Microbiologia (Messina Taormina). 1967.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
81
COSTA, M.; PASSOS, X.S.; , SOUZA, L.K.H.; MIRANDA, A.T.B.; LEMOS, J.A.; OLIVEIRA JÚNIOR, J.G.; RODRIGUES SILVA, M.R.R. Epidemiology and etiology of dermatophytosis in Goiânia, GO, Brazil Revista da Sociedade Brasileira de Medicina Tropical, v. 35, n. 1, p. 19-22, jan./fev. 2002. COULOMBE, P.A.; OMARY, M.B. Hard and soft principles definig structure, function an regulation of keratin intermediate filaments. Current Opinion in Cell Biology, v. 14, n. 1, p. 110-122, fev. 2002. CUESTA-RUBIO, O.; FRONTANA-URIBA, B.A.; RAMIREZ-APAN, T.; CARDENAS, J. Polyisoprenylated benzophenones in Cuban propolis; biological activity of nemorosone. Zeitschrift fur Naturforschung, v. 57c, n. 3/4, p. 372-380, mar./abr. 2002. CUSHNIE, T.P.T.; LAMB, A.J. Antimicrobial activity of flavonoids. International Journal of Antimicrobial Agents, v. 26, n. 5, p. 343–356, nov. 2005. DAMICO, D.C.S.; FREIRE, M.G.M.; GOMES, V.M.; TOYAMA, M.H.; MARANGONI, S.; NOVELLO, J.C.; MACEDO, M.L.R . Isolation and characterization of a lectin from Annona muricata seeds. Journal of Protein Chemistry, v. 22, n. 7/8, p. 655-661, nov. 2003. D’AURIA, F.D.; TECCA, M.; SCAZZOCCHIO, F.; RENZINI, V.; STRIPPOLI, V. 2003. Effect of propolis on virulence factors of Candida albicans. Journal of Chemotherapy, v. 15, n. 5, p. 454-460, out. 2003. DAS, K.S.; BANERJEE, A.B. Lipolytic enzymes of Trichophyton rubrum. Sabouraudia, v. 15, p. 313-323. 1977. DAUGSCH, A.; MORAES, C.S.; FORT, P.; PARK, Y.K. Brazilian Red Propolis - Chemical Composition and Botanical Origin. Evidence-Based Complementary and Alternative Medicine, p.1-7, jul. 2007. DE BACKER, M.; VROEY, M.C.; LESAFFRE, E.; SCHEYS, I.; KEYSER, P. Twelve weeks of continuous oral therapy for toenail onychomycosis caused by dermatophytes: a double-blind comparative trial of terbinafine 250mg/day versus itraconazole 200mg/day. Journal of the American Academy of Dermatology, v. 38, n. 5, suppl. 2, p. S57-S63, maio. 1998 DE HOOG, G.S.; BOUWMAN, B.; GRASER, Y.; HAASE, G.; EL FARI, M.; GERRITS VAN DEN ENDE, H.G.; MELZER-KRICK, B.; UNTEREINER, W.A. Molecular phylogeny and taxonomy of the medically important fungi. Medical Mycology. v. 36, suppl. 1, p. 52-56. 1998.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
82
DE SOL, F.G.; NAGANO, C.; CAVADA, B.S.; CALVETE, J.J. The first crystal structure of a mimosoideae lectin reveals a novel quaternary arrangement of a widespread domain. Journal of Molecular Biology, v. 353, n. 3, p. 574-583, out. 2005. DESHMUKH, S.K. The maintenance and preservation of keratinophilic fungi and related dermatophytes. Mycoses. v. 46, p. 203-207. 2003. DESHMUKH, S.; AGRAVAL, S.C. Degradation of human hair by some dermatophytes and keratinophilic fungi. Mykosen. v. 28, n. 9, p. 463-466, set. 1982. DIETRICH; KUSTENMACHER (1911) apud CIZMÁRIK, J.; MATEL, I. Examination of the chemical composition of propolis I. Isolation and identification of the 3,4-dihydroxycinnamic acid (caffeic acid) from propolis. Cellular and Molecular Life Sciences , v. 26, p. 7, p. 713. 1970. DIOGO, H.C.; SARPIERI, A.; PIRES, M.C. Preservação de fungos em água destilada. Anais Brasileiros de Dermatologia, v. 80, n. 6, p. 591-594, nov./dez. 2005. DIAS, T.; FERNANDES, O.F.; SOARES, A.J.; PASSOS, X.S.; COSTA, M.; HASIMOTO, S.L.K.; SILVA, M.D.O.R. Tinha do couro cabeludo em crianças de Goiânia, Brasil. Revista da Sociedade Brasileira de Medicina Tropical. v. 36, n. 6, p. 653-655, nov./dez. 2003. DOBROWOLSKI, J. W., VOHORA, S. B., SHARMA, K., SHAH, S. A., NAQVI, S. A. H.; DANDIYA, P. C. Antibacterial, antifungal, antiamoebic, antiinflamatory and antipyretic studies on propolis bee products. Journal of Ethnopharmacology, v. 35, n. 1, p. 77-82, out. 1991. DOES, M. P.; HOUTERMAN, P. M.; DEKKER, H. L.; CORNELISSEN, B. J. Processing, targeting and antifungal activity of stinging nettle. Plant Physiology, v.120, n. 2, p. 421-432, jun. 1999. DONCKER, P. Itraconazole and terbinafine in perspective: from petri dish to patient. Journal of the European Academy of Dermatology and Venereology, v. 12, suppl. 1, p. S10-S16, set. 1999. DONNELLY, D. M. X.; KEENAN, P. J.; PRENDERGAST, J. P. 1973. Isoflavonoids of Dalbergia ecastophyllum. Phytochemistry, v. 12, n. 5, p. 1157-1161, maio. 1973 DOS SANTOS, J.I.; VICENTE, E.J.; PAULA, C.R.; GAMBALE, W. Phenotypic Characterization of Trichophyton rubrum isolates from two geographic locations in Brazil. European Journal of Epidemiology, v. 17, n. 8, p. 729-735. 2001.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
83
ELEWSKI, B. E. Topics in Clinical Dermatology - Cutaneous Fungi Infection. New York: Igaku - Shoin, 1992. 255 p. EMMONS, C.W. Dermatophytes natural grouping based on the form of the spore and accessory. Archives of Dermatology and Syphylology, v. 30, p. 337-362. 1934. ESTUDO da biologia celular de fungos patogênicos e da relação parasita – hospedeiro. Disponível em: <http://www.sigma foco.scire.coppe. ufrj.br/UFRJ/SIGMA/projetos/consulta/ relatorio.stm?app=PROJETOS&código=11395&buscas_cruzadas=ON>. Acesso em: 06 jun. 2007. ESQUENAZI, D.; ALVIANO, C.S.; DE SOUZA, W.; ROZENTAL, S. The influence of surface carbohydrates during in vitro infection of mammalian cells by the dermatophyte Trichophyton rubrum. Research in Microbiology, v. 155, n. 3, p. 144–153, abr. 2004. FAVRE, B.; GHANNOUM, M.A.; RYDER, N.S. Biochemical characterization of terbinafine-resistant Trichophyton rubrum isolates. Medical Mycology, v. 42, n. 6, p. 525-529, dez. 2004. FAVRE, B.; HOFBAUER, B.; HILDERING, K.A.; RYDER, N.S. Comparison of In Vitro Activities of 17 Antifungal Drugs against a Panel of 20 Dermatophytes by Using a Microdilution Assay. Journal of Clinical Microbiology, v. 41, n. 10, p. 4817-4819, out. 2003. FENNEL, D. Conservation of fungus cultures. Botanical Review. v. 26, p. 79-141, 1960. FENG, M.G.; HUA, L. Factors a.ecting the sporulation capacity during long-term storage of the aphid-pathogenic fungus Pandora neoaphidis grown on broomcorn millet. FEMS Microbiology Letters, v. 245, n. 2, p. 205–211, abr. 2005. FERNANDES, F.F.; DIAS, A.L.T.; RAMOS, C.L.; IKEGAKI, M.; SIQUEIRA, A.M.; FRANCO, M.C. The in vitro antifungal activity evaluation of propolis G12 ethanol extract on Cryptococcus neoformans. Revista do Instituto de Medicina Tropical de São Paulo, v.49, n. 2, p. 93-95, mar./abr. 2007. FERNANDES, J.; LEOMIL, R.L.; FERNANDES, A.A.H.; SFORCIN, J.M. The antibacterial activity of propolis produced by Apis mellifera L. and brazilian stingless bees. Journal of Venomous Animals and Toxins, v. 7, n. 2, p. 173-182, dec. 2001 FERNANDES JÚNIOR; A.; LOPES, M.M.R.; COLOMBARI, V.; MONTEIRO, A.C.M.; VIEIRA, E.P. Atividade antimicrobiana de própolis de Apis mellifera obtidas em três regiões do Brasil. Ciência Rural, v. 36, n. 1, p. 294-297, jan./fev. 2006.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
84
FERNÁNDEZ-TORRES, B.; CABAÑES, F.J.; CARRILLO-MUÑOZ, A.J.; ESTEBAN, A.; INZA, I.; ABARCA, L.; GUARRO, J. Collaborative evaluation of optimal antifungal susceptibility testing conditions for dermatophytes. Journal of Clinical Microbiology, v. 40, n. 11, p. 3999-4003, nov. 2002. FERNÁNDEZ-TORRES, B.; CARRILLO, A.J.; MARTÍN, E.; DEL PALACIO, A.; MOORE, M.K.; VALVERDE, A.; SERRANO, M.; GUARRO, J. In Vitro Activities of 10 Antifungal Drugs against 508 Dermatophyte Strains. Antimicrobial Agents and Chemotherapy, v. 45, n. 9, p. 2524-2528, set. 2001. FERNÁNDEZ-TORRES, B.; CARRILLO-MUÑOZ, A.; INZA, I.; GUARRO, J. Effect of Culture Medium on the Disk Diffusion Method for Determining Antifungal Susceptibilities of Dermatophytes. Antimicrobial Agents and Chemotherapy, v. 50, n. 6, p. 2222–2224, jun. 2006. FERNÁNDEZ-TORRES, B.; INZA, I.; GUARRO, J. In vitro activities of the new antifungal drug eberconazole and three other topical agents against 200 strains of dermatophytes. Journal of Clinical Microbiology, v. 41, n. 11, p. 5209-5211, nov. 2003a. FERNÁNDEZ-TORRES, B.; INZA, I.; GUARRO, J. Comparasion of in vitro antifungal susceptibilities of conidia and hyphae of dermatophytes with thick-wall macroconidia. Antimicobial Agents and Chemotherapy, v. 47, n. 10, p. 3371-3372, out. 2003b. FERREIRA-NOZAWA, M.S.; NOZAWA1, S.R.; MARTINEZ-ROSSI1, N.M.; ROSSI, A. The dermatophyte Trichophyton rubrum secretes an EDTA-sensitive alkaline phosphatase on high-phosphate medium. Brazilian Journal of Microbiology, v. 34, n. 2, p. 161-164, jun. 2003. FERRETI-DE-LIMA, R.; DE-MORAES-BORBA, C. Viability, morphological characteristics and dimorphic ability of fungi preserved by di.erent methods. Revista Iberoamericana de Micolgía, v. 18, p. 191–196. 2001. FIGUEIREDO, M. B. Métodos de preservação de fungos patogênicos. Biológico, v. 63, n. 1/2, p. 73-82, jan./dez. 2001. FISHER, F.; COOK, N.B. Micologia: Fundamentos e Diagnóstico. Rio de Janeiro: Revinter, 2001, p. 116-147, 137-141, 321-322, 337p. FONTANA, J. D., PASSOS, M., SANTOS, M. H. R. D., FONTANA, C. K., OLIVEIRA, B. H., SCHAUSE, L., PONTAROLO, R., BARBIRATO, M. A., RUGGIERO, M. A. E LANÇAS, F. M. Profiling propolis flavonoids by means of micellar electrokinetic capillary chromatography, capillary gas chromatography and bactericidal action. Chromatographia, v. 52, n. 3/4, p. 147-151; 2000.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
85
FOSTEL, J.; LARTEY, P. Emerging novel antifungal agents. Drug Discovery Today, v. 5, n. 1, p. 25-32, jan. 2000. FREEDBERG, I.M.; EISEN, A.G.; WOLLF, K.; AUSTEN, K.F.; GOLDSMITH, C.A.; KATZ, F.I. Fungal disease with cutaneous involvement. In: ______. Fitgpatrick’s: dermatology in general medicine. 6. ed. New York: Mc Grawhill, 2003. cap. 29, v. 2, p. 1989-2018. FREIRE, M.G.M.; GOMES, V.M.; CORSINI, R.E.; MACHADO, O.L.T.; DE SIMONE, S.G.; NOVELLO, J.C.; MARANGONI, S.; MACEDO, M.L.R. Isolation and partial characterization of a novel lectin from Talisia esculenta seeds that interferes with fungal growth. Plant Physiology and Biochemistry, v. 40, n. 1, p. 61-68, jan. 2002. FRIEDRICH, J.; GRADISAR, H.; MANDIN, D.; CHAUMONT, J.P. Screening fungi for synthesis of keratinolytic enzymes. Letters in Applied Microbiology, v. 28, p. 127-130. 1999. FRIEDCH, J.; KERN, S. Hydrolisis of native proteins by keratinolytic proteases of Doratomyces microsporus. Journal of Molecular Catalysis B: Enzymatic, v. 21, n. 1-2, p. 35-37, jan. 2003. FUNARI, C.S.; FERRO, V.O. Análise de Própolis. Ciência e Tecnologia de Alimentos, v. 26, n. 1, p. 171-178, jan./mar. 2006. GARCÍA, V.M.N.; GONZALEZ, A.; FUENTES, M.; AVILES, M.; RIOS, M.Y.; ZEPEDA, G.; ROJAS, M.G. Antifungal activities of nine traditional Mexican medicinal plants. Journal of Ethnopharmacology, v. 87, n. 1, p. 85–88, jul. 2003. GARCÍA-PATOS, V.; G. APARICIO. Terapéutica sistémica en dermatología pediátrica. Disponível em: <http://www.aeped.es/protocolos/dermatologica/cuatro/terapiasistemica.pdf>. Acesso em: 20 jul. 2007. GAREDEW, A.; SCHMOLZ, E.; LAMPRECHT, I. Microbiological and calorimetric investigations on the antimicrobial actions of different propolis extracts: an in vitro approach. Thermochimica Acta, v. 422, n. 1-2, p. 115–124, nov. 2004. GEORG, L.K. Studies on Trichophyton tonsurans. I. The taxonomy of T. tonsurans. Mycologia, v.48, n. 1, p.65-82, jan./fev. 1956. Disponível em: <http://links.jstor.org/sici?sici=00275514(1956 01%2F02)48%3A1%3C65%3ASOTTIT%3E2.0.CO%3B2-S>. Acesso em 28.jul. 2008. GHANNOUM, M.A.; CHATURVEDI, V.; ESPINEL-INGROFF, A.; PFALLER, M.A.; RINALDI, M.G.; LEE-YANG, W.; WARNOCK, D.W. Intra- and interlaboratory study of a method for testing the antifungal susceptibilities of dermatophytes. Journal of Clinical Microbiology, v. 42, n. 7, p. 2977–2979, jul. 2004.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
86
GHANNOUM, M.A.; ARTHINGTON-SKAGGS, B.; CHATURVEDI, V.; ESPINEL-INGROFF, A.; PFALLER, M.A.; RENNIE, R.; RINALDI, M.G.; WALSH, T.J. Interlaboratory study of quality control isolates for a broth microdilution method (modified CLSI M38-A) for testing susceptibilities of dermatophytes to antifungals. Journal of Clinical Microbiology, v. 44, n. 12, p. 4353-4356, dez. 2006. GIORDANI, R., TREBAUX, J., MASI, M., REGLI, P. Enhanced antifungal activity of ketoconazole by Euphorbia characias latex against Candida albicans. Journal of Ethnopharmacology, v. 78, n. 1, p. 1-5, nov. 2001. GINDRO, K.; PEZET, R. Effects of long-term storage at different temperatures on conidia of Botrytis cinerea Pers.:Fr. FEMS Microbiology Letters, v. 204, n. 1, p. 101-104, out. 2001. GIRÃO, M.D.; PRADO, M.R.; BRILHANTE, R.S.N.; CORDEIRO, R.A.; MONTEIRO, A.J.; SIDRIM, J.J.C; ROCHA, M.F.G. Viabilidade de cepas de Malassezia pachydermatis mantidas em diferentes métodos de conservação. Revista da Sociedade Brasileira de Medicina Tropical, v. 37, n. 3, p. 229-233, mai./jun. 2004. GOMES, J.C.; CAVADA, B.S.; MOREIRA, R.A.; OLIVEIRA, J.T.A. Histamine release induced by glucose (mannose) specific lectins isolated from Brazilian plants. Comparison with concanavalin A. Inflammation Research, v. 41, n. 3-4, p. 132-135, maio. 1994. GONZALEZ, R.; REMIREZ, D.; RODRIGUEZ, S.; GONZALEZ, A.; ANCHETA, O.; MERINO, N.; PASCUAL, C. Hepatoprotective effects of propolis extract on paracetamol-induced liver damage in mice. Phytotherapy Research, v. 8, n. 4, p. 229–232. 1994. GONZALEZ, R.; CORCHO, I.; REMIREZ, D.; RODRIGUEZ, S.; ANCHETA, O.; MERINO, N.; GONZALEZ, A.; PASCUAL, C. Hepatoprotective effects of propolis extract on carbon tetraratchloride-induced liver injury in rats. Phytotherapy Research, v. 9, p. 114–17. 1995. GOSHEV, I.; GOUSTEROVA, A.; VASILEVA-TONKOVA, E.; NEDKOV, P. Characterization of the enzyme complexes produced by two newly isolated thermophylic actinomycete strains during growth on collagen-rich materials. Process Biochemistry, v. 40, n. 5, p. 1627-1631, abr. 2005. GOZIA, O.; CIOPRAGA, J.; BENTIA, T.; LUNGU, M.; ZAMFIRESCU, I.; TUDOR, R.; ROSEANU, A.; NITU, F. Antifungal properties of lectin and new chitinases from potato tubers. Comptes Rendus de l’Académie de Ssiences, Serie III, Sciences de la Vie, v. 316, n. 8, p. 788-92, aug. 1993. GRADISAR, H.; KERN, S.; FRIEDRICH, J. Keratinase of Doratomyces microsporus. Applied Microbiology and Biotechnology, v. 53, n. 2, p. 196-200, fev. 2000.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
87
GRANGEIRO, T.B.; SCHRIEFER, A.S.; CALVETE, J.J.; RAIDA, M.; URBANKE, C.; BARRAL-NETTO, M.; CAVADA, B.S. Molecular cloning and characterization of ConBr, the lectin of Canavalia brasiliensis seeds. European Journal of Biochemistry, v. 248, n. 1, p. 43-48, ago. 1997. GREENAWAY, W.; MAY, J.; SCAYSBROOK, T; WHATLEY, F. R. Identification by Gas- Chromatography Mass-Spectrometry of 150 compounds in propolis. Zeitschrift Fur Naturforschung C, v. 46, p. 111-121. 1991. GRIFFIN, D.H. Fungal Physiology. 2.ed. Wiley-Liss: New York, 1993. 458p. GRZYWNOWICZ, G.; LOBARZEWSKI, J.; WAWRZKIEWICZ, K.; WOLSKI, T. Comparative characterization of proteolytic enzymes from Trichophyton gallinae and Trichophyton verrucosum. Journal of Medical and Veterinary Mycology, v. 27, n. 5, p. 319-328. 1989. GUGNANI, H.C. Nondermatophytic filamentous keratinophilic and their role in the human infection. Polish Journal of Environmental Studies, v. 12, n. 4, p. 461-466. 2003. GURGEL, L.A.; SIDRIM, J.J.C.; MARTINS, D.T.; CECHINEL FILHO, V.; RAO, V.S. In vitro antifungal activity of dragon’s blood from Croton urucurana against dermatophytes. Journal of Ethnopharmacology, v. 97, n. 2, p. 409-412, fev. 2005. HANKIN, L.; ANAGNOSTAKIS, S.L. The use of solid media for detection of enzymes production by fungi. Mycologia, v. 67, n. 3, p.597-607, maio/jun. 1975. HANEL., H.; KIRS, R.; SCHIMIDTS. H. New systematically active antimycotics from the betablocker category. Mycoses, v. 38, n. 7-8, p. 251 – 264, 1995. HAZEN, K.C. Fungicidal versus fungistatic activity of terbinafine and itraconazole: an in vitro comparison. Journal of the American Academy of Dermatology, v. 38, n. 5, p. 37-41, maio. 1998. HAZEN, K.C. Evaluation of in vitro susceptibility of dermatophytes to oral antifungal agents. Journal of the American Academy of Dermatology, v. 43, suppl. 5, p. S125-S129, nov. 2000. HARBORNE, J.B.; WILLIAMS, C.A. Advances in flavonoid research since 1992. Phytochemistry, v. 55, n. 6, p. 481-504, nov. 2000. HAVSTEEN, B.H. The biochemistry and medical significance of the flavonoids. Pharmacology and Therapeutics, v. 96, n. 2, p. 67-202. 2002.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
88
HAVSTEEN, B. Flavonoids, a class of natural products of high pharmacological potency. Biochemical Pharmacolgy, v. 32, n. 9, p. 1141-1148, abr. 1983 HEGAZI, A.G., HADY, F.K.A.E.; ALLAH, F.A.A. Chemical composition and antimicrobial activity of European propolis. Zeitschrift fur Naturforschung, v. 55c, n. 1-2, p. 70-75, jan./fev. 2000. HEGAZI, A.G. Propolis an overview. Disponível em: <http://www.propoleo.cl/cientificospropolis/dr_Ahmed_Hegazi.pdf.> Acesso em: 22 abr. 2007. HERNÁNDEZ, T.; MACHADO, S.; CARVALHO, S.; SELORES, M. Tinhas do couro cabeludo na idade pediátrica. Nascer e Crescer. v. 12, n. 1. 2004. HERNÁNDEZ, E.S.; SANTIAGO, A.P.; CAMPOS, E.P.; ALVA, F.C. Efecto de la concanavalina a en el cultivo del fitopatógeno Ustilago maydis. Interciencia, v. 28, n. 5, p. 276-280, maio. 2003. HONDA, S. Dietary use of collagen and collagen peptides for cosmetics. Food Style, v. 21, p. 54-60. 1998. HOPSU-HAVU, V.K.; TUNNELA, E. Production of elastase, urease and sulphatase by Epidermophyton floccosum (Harz) Langeron et Milochevitch (1930). Mycosen, v. 20, n. 3, p. 91-96, mar. 1977. IBRAHIM, A.S.; MIRBORDE, F.; FILLER, S.G.; BANO, Y.; COLE, G.T.; KITAJIMA, Y.; EDWARDS, J.E.; NOZAWA, Y.; CHAMMOUM, M.A. Evidence implicating phospolipase as a virulence factor of Candida albicans. Infection and Immunity, v. 63, n. 5, p. 1993 - 1998, maio. 1995. INA, C.; SANO, K.; YAMAMOTO-TAKAHASHI, M.; MATSUSHITA-OIKAWA, H.; TAKEKAWA, H.; TAKEHARA, Y.; UEDA, H.; OGAWA, H. Screening for and purification of novel self-aggregatable lectins reveal a new functional lectin group in the bark of leguminous trees. Biochimica et Biophysica Acta (BBA) - General Subjects, v. 1726, n. 1, p. 21-24, out. 2005 ISHIKAWA, M.; KANNO, S.; ASOU, K.; OGINO, M.; TADANO, T.; SATOU, S. Inhibition of growth and induction of apoptosis in human cancer cell lines by Propolis. Journal of Pharmacological Sciences, v. 94, suppl. 1, p. 129-129, 2004. JAIN, S.; SHEGAL, V.N. Itraconazole: an effective oral antifungal for onychomycosis. International Journal of Dermatology, v. 40, n. 1, p. 1-5, jan. 2001.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
89
JESSUP, C.J.; WARNER, J.; ISHAM, N.; HASAN, I.; GHANNOUM, M.A. Antifungal Susceptibility Testing of Dermatophytes: Establishing a Medium for Inducing Conidial Growth and Evaluation of Susceptibility of Clinical Isolates. Journal of Clinical Microbiology, v. 38, n. 1, p. 341–344, jan. 2000. JONGS, S.G.; DAVIS, E.E. Conservation of reference strains of Fusarium in pure culture. Mycopathologia, v. 66, n. 3, p. 153-159, jan. 1979. KANAZAWA, M.; SATOMI, Y.; MIZUTANI, Y.; UKIMURA, O; KAWAUCHI, A.; SAKAI, T.; BABA, M.; OKUYAMA, T., 2003. Isoliquiritigenin inhibits the growth of prostate cancer. European Urology, v. 43, n. 5, p. 580-586, maio. 2003. KARACA, N.; KOÇ, A.N. In vitro susceptibility testing of dermatophytes: comparison of disk diffusion and reference broth dilution methods. Diagnostic Microbiology and Infectious Disease, v. 48, n. 4, p. 259-264, abr. 2004. KOC, A.N.; SILICI, S.; AYANGIL, D.; FERAHBAS, A.; ÇANKAYA, S. Comparison of in vitro activities of antifungal drugs and ethanolic extract of propolis against Trichophyton rubrum and T. mentagrophytes by using a microdilution assay. Mycoses, v. 48, n. 3, p. 205-210, maio. 2005. KONG, L. D.; ZHANG, Y.; PAN, X.; TAN, R. X.; CHENG, C. H. K., 2000. Inhibition of xanthine oxidase by liquiritigenin and isoliquiritigenin isolated from Sinofranchetia chinensis. Cellular and Molecular Life Sciences, v. 57, n. 3, p. 500-505, mar. 2000. KOO, H.; GOMES, B.P.F.A.; ROSALEN, P.L.; AMBROSANO, G.M.B.; PARK, Y.K.; CURY,J.A. In vitro antimicrobial activity of propolis and Arnica montana against oral pathogens. Archives of Oral Biology, v. 45, n. 2, p. 141-148, fev. 2000. KOO, M. H.; PARK, Y. K. Investigation of flavonoid aglycones in propolis collected by two different varieties of bees in the same region. Bioscience, Biotechnology and Biochemistry, v. 61, n. 2, p. 367-369, 1997. KOO, H; PARK, Y.K.; IKEGAKI, M.; CURY, J.A.; ROSALEN, P.L. Effect of Apis mellifera propolis from two Brazilian regions on caries development in desalivated rats. Caries Research, v. 33, n. 5, p. 393-400, 1999. KRAMER, C.L.; MIX, A.J. Deep Freeze Storage of Fungus Cultures. Transactions of the Kansas Academy of Science, v. 60, n. 1, p. 58-64. 1957.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
90
KITAJMA, Y. Structural and biochemical characteristics of pathogenic fungus: cell walls, lipids and dimorphism, and action modes of antifungal agents. Nippon Ishinkin Gakkai Zasshi, v.41, n. 4, p. 211-217, 2000. KUJUMGIEV, A.; TSVETKOVA, I.; SERKEDJIEVA, Y.; BANKOVA, V.; CHRISTOV, R.; POPOV, S. Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. Journal of Ethnopharmacology, v. 64, n. 3, p. 235-240, mar. 1999.
KUMAZAWA, S.; YONEDA, M.; SHIBATA, I.; KANAEDA, J.; HAMASAKA, T.; NAKAYAMA, T. Direct evidence for the plant origin of brazilian propolis by the observation of honeybee behavior and phytochemical analysis. Chemical and Pharmaceutical Bulletin, v. 51, n. 6, p. 740-742., jun. 2003.
KUMAZAWA, S.; HAMASAKA, T.; NAKAYAMA, T. Antioxidant activity of propolis of various geographic origins. Food Chemistry, v. 84, n. 3, p. 329-339, fev. 2004. KURNET, J; KASAFÍREK, E. Preliminary characterization of extracellular proteolytic enzymes of dermatophytes by chromogenic substrates. Journal of Medical and Veterinary Mycology, v. 26, n. 3, p. 187-194. 1988. KWON - CHUNG, K.J.; BENETT, J.E. Medical Mycology. London: Lea & Febiger, 1992. 833 p. LACAZ, C.S.; PORTO, E.; HEINS-VACCARI, M.E.; MELO, N.T. Guia para identificação: fungos, actinomicetos e algas de interesse médico. São Paulo: Sarvier, 1998. 445p. LACAZ, C.S.; PORTO, E.; MARTINS, J.E.C.; HEINS-VACCARI, E.M.; MELO, N.T. Tratado de Micologia Médica. 9. ed. Rio de Janeiro: Sarvier, 2002. 1104p. LARONE, D.H Culture and Identification of Dermatophytes. Clinical Microbiology Newsletter, v. 18, n. 5, p. 33-40, mar. 1996. LLAMBRICH, A.; LECHA, Y.M. Tratamiento actual de las onicomicosis. Revista Iberoamericana de Micología, v. 19, p. 127-129. 2002. LICATA, L.; SMITH, C.E.; GOLDSCHMIDT, R.; BARRETT, J.F.; FROSCO, M. Comparison of the postantibiotic and postantibiotic sub-MIC effects of levofloxacin and ciprofloxacin on Staphylococcus aureus and Streptococcus pneumoniae. Antimicrobial Agents and Chemotherapy, v. 41, n. 5, p. 950-955, maio. 1997. LIMA, R.F.; BORBA, C.M. Viability and morphological characteristics and dimorphic ability by different methods. Revista Iberoamericana de Micología, v. 18, n. 4, p. 191-196, dez. 2001.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
91
LIMA, R.F.; SANTOS, B.M.; SCHFFER, G.G.; LIMA, O.C.; BORBA, C.M. Evaluation of the in vitro and in vivo dimorphism of Sporothrix schenckii, Blastomyces dermatitidis and Paracoccidioides brasiliensis isolates after preservation in mineral oil. Canadian Journal of Microbiology, v. 50, n. 6, p. 445-449, jun. 2004.
LIS, H.; SHARON, N. Lectins: Carbohydrate-specific proteins that mediate cellular recognition. Chemical Reviews, v. 98, n. 2, p. 637-674, abr. 1998. LEHNINGER, A.L. Princípios de Bioquímica. Sarvier: São Paulo, 1993. 725p. LEYDEN, J. Pharmacokinetics and pharmacology of terbinafine and itraconazole. Journal of the American Academy of Dermatology, v. 38, 5 Pt. 3, p. S42-47. 1998. LOPES. J.O.; ALVES, S.H.; BENEVENGA, J.P. Dermatofitoses humanas no interior do Rio Grande do Sul no período de 1988-1992. Revista do Instituto de Medicina Tropical de São Paulo, v. 36, n. 2, p. 115-119, mar./abr. 1994. LOPEZ-MARTINEZ, R.; MANZANO-GAYOSSO, P.; MIER, T.; MENDEZ-TOVAR, L.J.; HERNANDEZ-HERNANDEZ, F. Exoenzymes of dermatophytes isolated from acute and chronic tinea Revista Latinoamericana de Microbiologia, v. 36, n. 1, p. 17-20, jan./mar. 1994. LORIS, R. Principles of structures of animal and plant lectins. Biochimica et Biophysica Acta (General Subjects), v. 1572, n. 2, p. 198-208, set. 2002. MACÊDO, D.P.C.; NEVES, R.P.; MAGALHÃES, O.M.C. SOUZA-MOTTA, C.M.; QUEIROZ, L.A. Pathogenic aspects of Epidermophyton floccosum Langeron et Milochevitch as a possible aethiological agent of tinea capitis. Brazilian Journal of Microbiology, v. 36, n. 1, p. 6-37, jan./mar. 2005. MAGO, K. Subcellular localization of enzymes of phospholipase metabolism in Candida albicans. Journal of Medical and Veterinary Mycology, v. 28, n. 5, p. 355-362, 1990. MAIA, M.L.S.; DOS SANTOS, J.I.; VIANI, F.C.; LARSSON, C.E.; PAULA, C.R.; GAMBALE, W. Phenotypic characterization of Microsporum canis isolates from cats and dogs. Mycoses, v. 44, n. 11/12, p. 480-486, dez. 2001. MARCUCCI, M.C. Propolis: chemical composition, biological properties and therapeutic activity. Apidologie, v. 26, n. 2, p. 83–99. 1995. MARCUCCI, M.C.; FERRERES, F.; GARCÝ´A-VIGUERA, C.; BANKOVA, V.S.; DE CASTRO, S.L.; DANTAS, A.P.; VALENTE, P.H.M.; PAULINO, N. Phenolic compounds from

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
92
Brazilian propolis with pharmacological activities. Journal of Ethnopharmacology, v. 74, n. 2, p. 105–112, fev. 2001. MARCUCCI, M.C.; CUSTÓDIO, A.R.; PEREIRA, R.M.S. Própolis tipificada: um novo caminho para a elaboração de medicamentos de origem natural, contendo este produto apícola. Mensagem Doce, n.90, mar. 2007. MARCUCCI, M.C.; RODRIGUEZ, J.; FERRERES, F.; BANKOVA, V.; GROTO, R.; POPOV, S. Chemical composition of Brazilian propolis from São Paulo state. Zeitschrift für Naturforschung, v. 53C, p. 117-119. 1998. MARCHISIO, V.F.; CURETTI, D.; CASSINELLI, C.; BORDESE, C. Keratinolytic and keratinophilic fungi in the soils Papua New Guinea. Mycopathologia, v. 115, n. 2, p. 113-119, ago. 1991. MARKHAM, K.R.; MITCHELL, K.A.; WILKINS, A.L.; DALDY, J.A.; LU, Y. HPLC and GC-MS identification of the major organic constituents in new Zealand propolis. Phytochemistry, v. 42, n. 1, p. 205-211, maio. 1996.
MARTOS, I.; COSSENTINI, M.; FERRERES, F.; TOMAS- BARBERAN, F.A. Flavonoid Composition of Tunisian honey and propolis. Journal of Agricultural and Food Chemistry, v. 54, n.8, p. 2824-2829. 1997 MATSUI, T.; EBUCHI, S.; FUJISE, T.; ABESUNDARA, K..J.; DOI, S.; YAMADA, H.; MATSUMOTO, K. Strong antihyperglycemic effects of water-soluble fraction of Brazilian propolis and its bioactive constituent, 3, 4, 5-tri-ocaffeoylquinic acid. Biological & Pharmaceutical Bulletin, v. 27, n. 11, p. 1797–1803. 2004. MATSUNO, T. A new clerodane diterpenoid isolated from propolis. Zeitschrift für Naturforschung, v. 50C, p. 93-97. 1995. MATSUNO, T.; JUNG, S.K.; MATSUMOTO, Y.; SAITO, M.; MORILAWA, J. Preferential cytotoxicity to tumor cells of 3,5-diprenyl-4-hydroxycinnamic acid artepillin C isolated from propolis. Anticancer Research, v. 17, n 5A, p. 3565-3568, set./out. 1997. MAYSER, P.; LAABS, S.; HEUER, K.; GRÜNDER, K. Detection os extracellular phospholipase activity in Candida albicans and Rhodotorula rubra. Mycopathologia, v. 135, n.3, p. 149-155, jan. 1996. McGINNIS, M.R.; AJELLO, L.; ROGERS, D.P. Proposal to Conserve Epidermophyton Sabouraud, 1907 (Moniliaceae). Taxon, v. 30, n. 1, p. 351-353, fev. 1981.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
93
MEEVOOTISOM, V.; NIEDERPRUM, D.J. Control of exocellular proteases in dermatophytes and especially Trichophyton rubrum. Sabouraudia, v. 17, p. 91 – 106, 1979. MELLO, A.M.; GOMES, R.T.; LARA, R.S.; SILVA, L.G.; ALVES, J.B.; CORTÉS, M.E.; ABREU, S.L.; SANTOS, V.R. The effect of brazilian propolis on the germ tube formation and cell wall of Candida albicans. Pharmacologyonline, v. 3, p. 352-358. 2006. MELLO, S. C. M.; SILVA, J. B. T.; WETZEL, M. M. V. S. Coleções de culturas microbianas sistema nacional de pesquisa agropecuária. Embrapa Recursos genéticos e Biotecnologia, ISSN: 0102-0110, n. 102. Brasília, julho, 2003. MENEZES, H. Própolis: uma revisão dos recentes estudos de suas propriedades farmacológicas. Arquivos do Instituto Biológico, v. 72, n. 3, p. 405-411, jul./set. 2005. MICOTECA URM. Histórico. Disponível em: <http://www.ufpe.br/micoteca/historico.html>. Acesso em: 08 04. 2007. MIGNON, B.; SWINNEN, M.; BOUCHARA, J.P.; HOFINGER, M.; NIKKELS, A.; PIERARD, G.; GERDA,Y. C.; LOSSON, B. Purification and characterization of a 315 kDa keratinolytic subtilisin-like serine protease from Microsporum canis and evidence of its secretion in naturally infected cats. Medical Mycology, v. 36, n. 6, p.395-404, dez. 1998. MINOCHA, Y.; PASRICHA, J.S.; MOHAPATRA, L.N.; KANDHARI. K.C. Proteolytic activity of dermatophytes and its role in the pathogenesis of skin lesions. Sabouraudia, v. 10, p. 79-85. 1972 MIRZOEVA, O. K.; GRISHANIN, R. N.; CALDER, P. C. Antimicrobial action of propolis and some of its components: the effects on growth, membrane potencial and motility of bacteria. Microbiological Research, v. 152, n. 3, p. 239-246, 1997. MISHIMA, S.; INOH, Y.; NARITA, Y.; OHTA, S.; SAKAMOTO, T.; ARAKI, Y.; SUZUKI, K.; AKAO, Y.; NOZAWA, Y. Identification of caffeoylquinic acid derivatives from Brazilian propolis as constituents involved in induction of granulocytic differentiation of HL-60 cells. Bioorganic & Medicinal Chemistry, v. 13, n. 20, p. 5814–5818, out. 2005. MONTEIRO-MOREIRA, A.C.O. Caracterização estrutural de três lectinas apresentando especificidades distintas presentes em sementes de fruta-pão (Artocarpus incisa L.) Tese de Doutorado em Bioquímica Vegetal. Universidade Federal do Ceará. Disponível em: <http://servicos.capes.gov.br/capesdw/Pesquisa.do?autor=MONTEIROMOREIRA&tipoPesqAutor=T&assunto=lectinas&tipoPesqAssunto=T&ies=&tipoPesqIes=T&nivel=&anoBase>. Acesso em 12/01/2008

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
94
MORENO, M.I.N.; ISLÃ, M.I.; CUDMANI, N.G.; VATTUONE, M.A.; SAMPIETRO, A.R. Screening of antibacterial activity of Amaicha del Valle (Tucumán, Argentina) propolis. Journal of Ethnopharmacology, v. 68, n. 1/3, p. 97–102, dez. 1999. MORENO, M.I.N.; ZAMPINI, I.C.; ORDÓÑEZ, R.M.; JAIME, G.S.; VATTUONE, M.A.; ISLA, M.I. Evaluation of the cytotoxicity, genotoxicity, mutagenicity, and antimutagenicity of propolis from Tucuman, Argentina. Journal of Agricultutal and Food Chemistry, v. 53, p. 8957-8962. 2005. MORIELLO, K.A.; DE BOER, D.J. Fungal flora of the hair coat cats with and without dermatophytosis. Journal of Medical and Veterinary Mycology, v. 29, n. 5, p. 285-292, 1991. MOREIRA, R.A.; CORDEIRO, E.F.; RAMOS, M.V.; GRANGEIRO, T.B.; MARTINS, J.L.; OLIVEIRA, J.T.A.; CAVADA, B.S. Isolation and partial characterization of a lectin from seeds of Dioclea violacea. Revista Brasileira de Fisiologia Vegetal, v. 8, n. 1, p. 23-29. 1996 MOREIRA, R.A.; MONTEIRO, A.C.O.; HORTA, A.C.G.; OLIVEIRA, J.T.A.; CAVADA, B.S. Isolation and characterization of Dioclea altissima var. megacarpa seed lectin. Phytochemistry, v. 46, n. 1, p. 139-144, set. 1997. MOREIRA, R.A.; SILVA, L.M.A.; HORTA, A.C.G.; OLIVEIRA, J.T.A.; CAVADA, B.S. Lectin from Canavalia brasiliensis Mart. Behavior during maturation and detection of a lectin precursor. Revista Brasileira de Fisiologia Vegetal, v. 5, n. 2, p. 133-138, 1993. MUHSIN, T.; HADI, R.B. Degradation of keratin substrates by fungi isolated from sewage sludge. Mycopathologia, v. 154, n. 4, p.185-189, ago. 2002. MUHSIN, T.M.; AUBAID, A.H.; AL-DUBOON, A.A. Extracellular enzyme activities of dermatphytes and yeast isolates on solid media. Mycoses, v. 40, n. 11-12, p. 465-469. 1997. MUHSIN, T.M.; SALIH, T.H. Exocellular enzyme activity of dermatophytes and other fungi isolated from ruminants in Southern Iraq. Mycopathologia, v. 150, n. 2, p. 49-52, fev. 2000. MURAD, M.; CALVI, S.A.; SOARES, A.M.V.C.; BANKOVA, V.; SFORCIN, J.M. Effects of propolis from Brazil and Bulgaria on fungicidal activity of macrophages against Paracoccidioides brasiliensis. Journal of Ethnopharmacology, v. 79, n. 3, p. 331-334, mar. 2002. NAPIMOGA, M.H.; CAVADA, B.S.; ALENCAR, N.M.N.; MOTA, M.L.; BITTENCOURT, F.S.; ALVES-FILHO, J.C.; GRESPAN, R.; GONÇALVES, R.B.; CLEMENTE-NAPIMOGA, J.T.; FREITAS, A.; PARADA, C.A.; FERREIRA, S.H.; CUNHA, F.Q. Lonchocarpus sericeus lectin decreases leukocyte migration and mechanical hypernociception by inhibiting cytokine and chemokines production. International Immunopharmacology, v. 7, n. 6, p. 824–835, jun. 2007.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
95
NASCIMENTO, E.A.; BEZZAN, L.C.F. Estudo da própolis do triângulo mineiro. Parte 1: identificação dos constituintes solúveis em metanol/acetato de etila por CG-EM. Mensagem Doce, n. 63, set. 2001. NASCIMENTO, E.A.; MORAIS, S.A.L.; PILÓ-VELOSO, D.; CHANG, R.; REIS, D.C. Um marcador químico de fácil detecção para a própolis de alecrim-do-campo (Baccharis dracunculifolia). Mensagem Doce, n. 91, maio. 2007. NAKAJIMA, Y.; SHIMAZAWA, M.; MISHIMA, S.; HARA, H. Water extract of propolis and its main constituents, caffeoylquinic acid derivatives, exert neuroprotective effects via antioxidant actions. Life Sciences, v. 80, n. 4, p. 370-377, jan. 2007. NAKASONE, K.K.; PETERSON, S.W.; JONG, S.C. Preservation and distribution of fungal cultures. Disponível em: <http://www.fpl.fs.fed.us/documnts/pdf2004/fpl_2004_nakasone001. pdf>. Acesso em: 23 jul. 2007. NEUFELD, P. N.; SARQUIS, M. I. M. Preservação em laboratório de fungos filamentosos pelo método do óleo mineral. Revista Brasileira de Análises Clínicas, v. 35, n. 3, p. 147-150, 2003. NG, T.B.; YU, Y.L.; CHU, K.T. Isolation of a novel legumin-like lectin with potent hemagglutinating activity from seeds of the Chinese chestnut Castanea mollisima. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, v. 133, n. 3, p. 453-460, nov. 2002. NGAI, P.H.K.; NG, T.B. A mushroom (Ganoderma capense) lectin with spectacular thermostability, potent mitogenic activity on splenocytes, and antiproliferative activity toward tumor cells. Biochemical and Biophysical Research Communications, v. 314, n. 4, p. 988-993, fev. 2004. NOBRE, G.; VIEGAS, M.P. Lipolytic activity of dermatophytes. Mycopathologia, v. 46, n. 4, p. 319-323. 1972. NOMURA, K.; ASHIDA, H.; UEMURA, N.; KUSHIBE, S.; OZAKI, T.; YOSHIDA, M. Purification and characterization of a mannose/glucose-specific lectin from Castanea crenata. Phytochemistry, v. 49, n. 3, p. 667-673, out. 1998. NORRIS, H.A.; ELEWSKI, B.E.; GHANNOUM, M.A. Optimal growth conditions for the determination of the antifungal susceptibility of three species of dermatophytes with the use of a microdilution method. Journal of the American Academy of Dermatology, v. 40, n. 6, part. 2, p. Sp-S13, jun. 1999

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
96
NOTHENBERG, M. Própolis enfrenta bem o desafio das pesquisas. Química e Derivados. v. 348, p. 24-28, 1997. OGAWA, H.; SUMMERBELL, R.C.; CLEMONS, K.V.; KOGAS, T.; RAN, Y.P.; RASHID, A.; SOHNLE, P.G.; STEVENSM D.A.; TSUBOI, R. Dermatophytes and host defence in cutaneous mycoses. Medical Mycology, v. 36, suppl. I, p. 166-173, 1998. OKAFOR, J.I.; ADA, N. Keratinolytic activity of five human isolates of the dermatophytes. Journal of Communicable Diseases, v. 32, n. 4, p. 300-305, dez. 2000. OLIVEIRA, A.C.P.; SHINOBU, C.S.; ONGHINI, R.L.; FRANCO, S.L.; SVIDZINSKI, T.I.E. Antifungal activity of propolis extract against yeasts isolated from onychomycosis lesions. Memórias do Instituto Oswaldo Cruz, v. 101, n. 5, p. 493-497, ago. 2006. OLIVEIRA, M.N.; FERREIRO, P.N.S.; MEIRELES, M.C.; FERREIRO, L. Drogas antifúngicas para pequenos e grandes animais. Ciência Rural, v. 32, n. 1, p. 175-184. 2002. OMIDYNIA, E.; FARSHCHIAN, M.; SADJJADI, M.; ZAMANIAN, A.; RASHIDPOURAEI, R. A study of dermatophytoses in Hamadan, the governmentship of West Iran. Mycopathologia, v. 133, n. 1, p. 9-13, jan. 1996. ONIFADE, A.A.; AL-SANE, N.A.; AL-MUSALLAM, A.A.; AL-ZARBAN, S. A review: potentials for biotechnological applications of keratin-degrading microorganisms and their enzymes for nutritional improvement of feathers and others keratins as livestock feed resource. Bioresource Technology, v. 66, n. 1, p.1-11, out. 1998. OOI, L.S.; NG, T.B.; GENG, Y.; OOI, V.E. Lectins from bulbs of the Chinese daffodil Narcissus tazetta (family Amaryllidaceae). Biochemistry and Cell Biology, v. 78, n. 4, p. 463-468. 2000. pegar trab OTA, C.; UNTERKIRCHER, C.; FANTINATO, V.; SHIMIZU, M.T. Antifungal activity of propolis on different species of Candida. Mycoses, v. 44, n. 9/10, p. 375-378, nov. 2001. PACKER, J.F.; LUZ, M.M.S. Método para avaliação e pesquisa da atividade antimicrobiana de produtos de origem natural. Revista Brasileira de Farmacognosia, v. 17, n. 1, p. 102-107, jan./mar. 2007. PANIZZO, M.M.; REVIÁKINA, V.; WILLIAMA, M.;. GONZÁLEZ, G. Mantenimiento y preservación de hongos en agua destilada e aceite mineral. Revista de la Sociedad Venezolana de Microbiología, v. 25, n. 1, p. 37-48, 2005.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
97
PAPINI, R.; MANCIANTI, F. Extracellular enzymatic activity of Microsporum canis isolates. Mycopathologia. v.132, n. 3, p. 129 – 132, out. 1995. PARK, Y.K.; ALENCAR, S.M.; AGUIAR, C.L. Botanical Origin and Chemical Composition of Brazilian Propolis. Journal of Agricultural and Food Chemistry, v. 50, p. 2502-2506. 2002a. PARK, Y.K.; ALENCAR, S.M.; AGUIAR, C.L. Estudo da composição fenólica de méis e própolis oriundos de mesma colméia. Mensagem Doce, n. 67, jul. 2002b. PARK, Y.K.; ALENCAR, S.M.; AGUIAR, C.L.; SCAMPARINI, A.R.P.; GONZÁLEZ, M.; MOLINA, M.A.A. Comparação das características físico-químicas das própolis produzidas na região sub-tropical da América do Sul: evidência fitoquímica de sua origem botânica. Mensagem Doce, n. 61, maio. 2001. PARK, Y.K.; ALENCAR, S.M.; MOURA, F.F.; IKEGAKI, M. Atividades Biológicas da Própolis. Disponível em: <http://www.apisbrasil.com.br/propriedades.html>. Acesso em: 26 jun 2007. PARK, Y.K.; ALENCAR, S.M.; SCAMPARINI, A.R.P.; AGUIAR, C.L. Própolis produzida no sul do Brasil, Argentina e Uruguai: evidências fitoquímicas de sua origem vegetal. Ciência Rural, v. 32, n. 6, p. 997-1003, nov./dez. 2002c. PARK, Y. K.; IKEGAKI, M. Preparation of water and ethanolic extracts of propolis and evaluation of the preparations. Bioscience Biotechnology and Biochemistry, v. 62, n. 11, p. 2230-2232, 1998c. PARK, Y.K.; IKEGAKI, M.; ALENCAR, S.M. Classificação das própolis brasileiras a partir de suas características físico-químicas e propriedades biológicas. Mensagem Doce, n. 58, p. 2-7, set. 2000. PARK, Y.K.; KOO, M.H.; ABREU, J.A.S.; IKEGAKI, M.; CURY, J.A.; ROSALEN, P.L. Antimicrobial activity of propolis on oral microorganisms. Current Microbiology, v. 36, n. 1, p. 24-28, jan. 1998a. PARK, Y.K.; KOO, H; IKEGAKI, M.; CURY, J.A.; ROSALEN, P.L. Effect of propolis on Streptococcus mutans, Actinomyces naeslundii and Staphylococcus aureus. Revista de Microbiologia, v. 29, p. 143-148. 1998b. PARK, Y. K., KOO, M. H., IKEGAKI, M. E CONTADO, J. L. Comparison of the flavonoid aglycone contents of Apis mellifera propolis from various regions of Brazil. Arquivos de Biologia e Tecnologia, v. 40, n. 1, p. 97-106, mar. 1997.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
98
PARK, Y.K.; PAREDES-GUZMAN, J.F.; AGUIAR, C.L.; ALENCAR, S.M.; FUJIWARA, F.Y. Chemical constituents in Baccharis dracunculifolia as the main botanical origin of Southeastern Brazilian propolis. Journal of Agriculture and Food Chemistry, v. 52, n. 5, p. 1100-1103, fev. 2004.
PASCUAL,C.; GONZALEZ, R.; TORRICELLA, R. Scavenging action of propolis extract against oxygen radicals. Journal of Ethnopharmacology. v. 41, n. 1-2, p. 9-13, jan. 1994. PEREA, S.; FOTHERGILL, A.W.; SUTTON, D.A.; RINALDI, M.G. Comparasion of in vitro activies of variconazole and five established antifungal agents against different species of dermatophytes using a broth macrodiluition method. Journal of Clinical Microbiology, v. 39, n. 1, p. 385-388, jan. 2001. PEREIRA, A.C.; CARMO, E.D.; SILVEIRA, V.A.S.; AMADEI, S.U.; ROSA, L.E.B. O papel das MMP-2 e -9 no desenvolvimento do carcinoma Epidermóide. Revista Brasileira de Cancerologia, v. 52, n. 3, p. 257-262, jul./ago./set. 2006. PEREIRA, A. D. S., SEIXAS, F. R. M. S. E NETO, F. R. D. A. Própolis: 100 anos de pesquisa e suas perspectivas futuras. Química Nova, v. 25, n. 2, p. 321-326, mar./abr. 2002. PEREIRA, C.S.; ARAUJO, A.G.; GUIMARÃES, R,J.; PAIVA, L.C. Uso da própolis como inibidor da germinação de esporos de Hemileia vastrix. Mensagem Doce, n. 64, nov. 2001. PÉREZ-SANTIAGO, A.; SAAVEDRA, E.; PÉREZ CAMPOSA, E.; CÓRDOBA, F. Effect of plant lectins on Ustilago maydis in vitro. Cellular and Molecular Life Sciences, v. 57, n. 13/14, p. 1986-1989, dez. 2000. PETRIKKOU, E.; RODRÍGUEZ-TUDELA, J.L.; CUENCA-ESTRELLA, M.; GÓMEZ, A.; MOLLEJA, A.; MELLADO, E. Inoculum Standardization for Antifungal Susceptibility Testing of Filamentous Fungi Pathogenic for Humans. Journal of Clinical Microbiology, v. 39, n. 4, p. 1345-1347, abr. 2001. PEUMANS, W.J.; VAN DAMME, E.J.M. Lectins as plant defense proteins. Plant Physiology, v. 109, n. 2, p. 347-352, out. 1995. PHARMANECTAR: Citroprópolis - própolis de Baccharis dracunculifolia. Disponível em: <http://www.pharmanectar.com.br/cytopropolis.html>. Acesso em 15 mar. 2007. PHILPOT, C.M. Some aspects of the epidemiology of tinea. Mycopathologia, v. 62, n. 1, p. 3-13, jan. 1977. PIETTA, P.G.; GARDANA, C.; PIETTA, A.M. Analytical methods for quality control of propolis. Fitoterapia, v. 73, suppl. 1, p. S7-S20, nov. 2002.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
99
PINTO, M.S.; FARIA, J..; MESSAGE, D; CASSINI, S.T.A; PEREIRA, C.S.; GIOSO, M.M. Effect of green propolis extracts on pathogenic bacteria isolated from milk of cows with mastitis. Brazilian Journal of Veterinary Research and Animal Science, v. 38, n. 6, p. 278-283. 2001. PONTES, Z.; LIMA, E.D.E.O.; OLIVEIRA, N.M.; DOS SANTOS, J.P.; RAMOS, A.L.; CARVALHO, M.F. Onychomycosis in Joao Pessoa City, Brazil. Revista Argentina de Microbiología, v.34, n.2, p.95-99, apr./jun. 2002. POPOVA, M.; BANKOVA, V.; NAYDENSKY, CH.; TSVETKOVA, I.; KUJUMGIEV, A. Comparative study of the biological activity of propolis from different geographic origem: a statistical approach. Macedonian Pharmaceutical Bulletin, v. 50, p. 9-14, 2004. POPOVA, M.; SILICI, S.; KAFTANOGLU, O.; BANKOVA, V. Antibacterial activity of Turkish propolis and its qualitative and quantitative chemical composition. Phytomedicine, v 12, n. 3, p. 221-228, mar. 2005. PORRO, A. M., M. C. N. YOSIOKA, S. K. KAMINSKI, M. DE A. PALMEIRA, O. FISCHMAN, AND M. ALCHORNE. Disseminated dermatophytosis caused by Microsporum gypseum in two patients with the acquired immunodeficiency syndrome. Mycopathologia, v. 137, n. 1, p. 9-12, abr. 1997. PRICE, M.F.; CAWSON, R.A. Phospholipase activity in Candida albicans. Saboraudia, v.15, p. 79-185. 1977. PRICE, M.F.; WILKINSON, I.D.; GENTRY, L.O. Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia, v. 20, p. 7-14. 1982. PROPAVK, S. (1975) apud CIZMÁRIK, J.; MATEL, I. Examination of the chemical composition of propolis I. Isolation and identification of the 3,4-dihydroxycinnamic acid (caffeic acid) from propolis. Cellular and Molecular Life Sciences , v. 26, p. 7, p. 713. 1970 PRYTZYK, E.; DANTAS, A.P.; SALOMÃO, K.; PEREIRA, A.S.; BANKOVA, V.S.; DE CASTRO, S.L.; NETO, F.R.A. Flavonoids and trypanocidal activity of Bulgarian propolis. Journal of Ethnopharmacology, v. 88, n. 2/3, p. 189-193, out. 2003. PUJOL, I.; GUARRO, J.; LLOP, C.; SOLER, L.; FERNÁNDEZ-BALLART, J. Comparison Study of Broth Macrodilution and Microdilution Antifungal Susceptibility Tests for the Filamentous Fungi. Antimicrobial Agents and Chemotherapy, v. 40, n. 9, p. 2106-2110, set. 1996. PUJOL, I.; CAPILLA, J.; FERNÁNDEZ-TORRES, B.; ORTONEDA, M.; GUARRO, J. Use of the sensititre colorimetric microdilution panel for antifungal susceptibility testing of dermatophytes. Journal of Clinical Microbiology, v. 40, n. 7, p. 2618-2621, jul. 2002.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
100
QUINDÓS, G. Las micosis en el amanecer del Siglo XXI. Revista Iberoamericana de Micología, v. 19, p. 1-4. 2002. QUIROGA, E.N.; SAMPIETRO, D.A.; SOBERO, J.R.; SGARIGLIA, M.A.; VATTUONE, M.A. Propolis from the northwest of Argentina as a source of antifungal principles. Journal of Applied Microbiology, v. 101, n. 101, p. 103-110, jul. 2006. RAMOS, M.V.; MOREIRA, R.A.; CAVADA, B.S.; OLIVEIRA, J.T.A.; ROUGE, P. Interaction of lectins from the sub-tribe Diocleinae with specific ligands. Revista Brasileira de Fisiologia Vegetal, v. 8, n. 3, p. 193-199. 1996. RAPER, K. B.; ALEXANDER, D. F. Preservation of molds by the lyophilize process. Mycologia, v. 37, p. 499. 1945. REBELL, G.; TAPLIN, D. Dermatophytes: their recognition and identification. Florida: University of Miami Press, 1979. 124 p. REIS, C.F.M.; CARVALHO, J.C.T.; CAPUTO, L.R.G.; PATRÍCIO, K.C.M.; BARBOSA, M.V.J.; CHIEFF, A.L.; BASTOS, J.K. Atividade antiinflamatória, antiúlcera gástrica e toxicidade subcrônica do extrato etanólico de própolis. Revista Brasileira de Farmacognosia, v. 9/10, p. 43-52. 2000. REX, J. H.; PFALLER, M.A.; RINALDI, M.G.; POLAK, A.; GALGIANI, J.N. Antifungal susceptibility testing. Clinical Microbiology Reviews, v. 6, p. 367-381. 1993. REX, J.H.; PFALLER, M.A.; WALSH, T.J.; CHATURVEDI, V.; ESPINEL-INGROFF, A.; GHANNOUM, M.A.; GOSEY, L.L.; ODDS, F.C.; RINALDI, M.C.; SHEEHAN, D.J.; WARNOCK, D.W. Antifungal Susceptibility Testing: Practical Aspects and Current Challenges. Clinical Microbiology Reviews, v. 14, n. 4, p. 643-658, oct. 2001. RIBEIRO, J.N.; OLIVEIRA, T.T.; NAGEM, T.J.; FLORES, A.V. Ausência de Toxicidade da Própolis. Mensagem Doce, n.77, jul. 2004. RIPPON, J. W. Tratado de Micologia Médica: hongos e actinomicetos patógenos. 3 ed. México: Interamericana, 1990. 855 p.
RIPPON, J.W.; GARBER, D. Dermatophyte infection as a function of mating type and as associates enzymes. Journal of Investigative Dermatology, v. 53, p. 445-448. 1969.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
101
RODRIGUEZ, D.; CAVADA, B. S.; OLIVEITA, J. T. A.; MOREIRA, R. A.; RUSSO, M. Differences in macrophage stimulation and leukocyte accumulation in response to intraperitoneal administration of glucose/mannose-binding lectins. Brazilian Journal of Medical and Biological Research, v. 25, n. 8, p. 823-826. 1992. RUBINSTEIN, N.; ILARREGUI, J.M.; TOSCANO, M.A.; RABINOVICH, G.A. The role of galectins in the initiation, amplication and resolution of the inflammatory response. Tissue Antigens, v. 64, n. 1, p. 1-12, jul. 2004. RÜDIGER, H. Structure and function of plant lectins in Glycosciences. Status and perspectives. Chapman & Hall, Weinheim. p. 415-438. 1997. In Grangeiro et al., 1997 RUSSO, A.; LONGO, R.; VANELLA, A. Antioxidant activity of propolis: role of caffeic acid phenethyl ester and galangin. Fitoterapia, v. 73, suppl.. 1, p. S21-S29, nov. 2002. RYAN, M.J.; JEFFRIES, P.; BRIDGE, P.D.; SMITH, D. Developing cryopreservation protocols to secure fungal gene function. Cryoletters, v. 22, n. 2, p. 115-124, mar./abr. 2001. RYBNIKAR, A. Long-term maintenance of lyophilized fungal cultures of the genera Epidermophyton, Microsporum, Paecilomyces and Trichophyton. Mycoses, v. 38, n. 3-4, p. 145-147, mar./abr. 1995 SAID, S.; PIETRO, R. C. L. R. Enzimas de Interesse Industrial e Biotecnológico. Teresópolis: Livraria e Editora Eventos, 2002. v. 1. 120 p. SALATINO, A.; TEIXEIRA, E.W.; NEGRI, G.; MESSAGE, D. Origin and chemical variation of Brazilian propolis. Evidence-Based Complementary and Alternative Medicine, v 2, n. 1, p. 33-38. 2005. SAMDANI, A.J.; DYKES, P.J.; MARKS, R. The proteolytic activity of strains of T. mentagrophytes and T. rubrum isolated from tinea pedis e tinea unguium infections. Journal of Medical and Veterinary Mycology, v. 33, n. 3, p. 167-170, maio/jun. 1995. SANTOS, D.A.; HAMDAN, J.S. Evaluation of broth microdilution antifungal susceptibility testing conditions for Trichophyton rubrum. Journal of Clinical Microbiology, v. 43, n. 4, p. 1917-1920, abr. 2005. SANTOS, D.A.; BARROS, M.E.S.; HAMDAN, J.S. Establishing a Method of Inoculum Preparation for Susceptibility Testing of Trichophyton rubrum and Trichophyton mentagrophytes. Journal of Clinical Microbiology, v. 44, n. 1, p. 98–101, jan. 2006.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
102
SANTOS, V.R.; PIMENTA, F. J. G. S.; AGUIAR, M.C.F.; DO CARMO, M.A.V.; NAVES, M.D.; MESQUITA, R.A. Oral candidiasis treatment with brazilian ethanol propolis extract. Phytotherapy Research, v. 19, n. 7, p. 652–654, jul. 2005. SAWAYA, A.C.H.F.; PALMA, A.M.; CAETANO, F.M.; MARCUCCI, M.C.; DA SILVA CUNHA, I.B.; ARAÚJO, C.E.P.; SHIMIZU, M.T. Comparative study of in vitro methods used to analyse the activity of propolis extracts with different compositions against species of Candida. Letters in Applied Microbiology, v. 35, n. 3, p. 203–207, set. 2002. SCHONBORN, C. The long time survival of dermatophytes and moulds under paraffin oil. Mycoses, v. 39, n. 2, p. 349-353, jul. 1989. SECCO, L.C. Abordagem introdutória às micoses superficiais e aspectos relacionados. Disponível em: <http://www.racine.com.br/download.asp?idarquivobanco=2848>. Acesso em: 27 jul. 2007. SEQUEIRA, L.; GRAHAM, T.L. Agglutination of avirulent strains of Pseudomonus solanacearum by potato lectin. Physiological Plant Pathol, v. 11, p. 43-54. 1977. SFORCIN, J.M.; FERNANDES JUNIOR., A.; LOPES, C.A.; BANKOVA, V.; FUNARI, S.R. Seasonal effect on Brazilian propolis antibacterial activity. Journal of Ethnopharmacology, v. 73, n. 1-2, p. 243-249, nov. 2000. SFORCIN, J.M.; ORSI, R.O.; BANKOVA, V. 2005. Effect of propolis, some isolated compounds and its source plant on antibody production. Journal of Ethnopharmacology, v. 98, n. 3, p. 301-305, abr. 2005. SHARON, N.; LIS, H. History of lectins: from hemagglutinins to biological recognition molecules. Glicobiology, v. 14, n. 11, p. 53R-64R, 2004. SHE, Q.B.; NG, T.B.; LIU, W.K. Isolation of cicadin, a novel and antifungal peptide from dried juvenile cicadas, Biochemical and Biophysical Research Communications, v. 247, n. 1, p. 106-111, jun. 1998. SHERF, A.F. A method for maintaining Phytomonas sepedomica for long periods without transfer. Phytopathology, v. 31, p. 30-32, 1943. SIDRIM, J. J.C.; ROCHA, M. F. G. Micologia Médica à luz de autores contemporâneos. Rio de Janeiro: Guanabara Koogan, 2004. 388p. SIESENOP, U.; BOHM, K.H. Comparative studies on keratinases production of Trichophyton mentagrophytes strains of animal origin. Mycoses, v. 38, n. 5/6, p. 205-209, maio/jun. 1995.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
103
SILICI, S.; KOÇ, N.A. Comparative study of in vitro methods to analyse the antifungal activity of propolis against yeasts isolated from patients with superficial mycoses. Letters in Applied Microbiology, v. 43, n. 3, p. 318-324, set. 2006. SILICI, S.; KOÇ, N.A.; AYANGIL, D.; ÇANKAYA, S. Antifungal activities of propolis collected by different races of honeybees against yeasts isolated from patients with superficial mycoses. Journal of Pharmacological Sciences, v. 99, n. 1, p. 39-44. 2005. SILVA, B.B.; ROSALEN, P.L.; CURY, J.A.; GAKI, M.; SOUZA, V.C.; ESTEVES, A./ ALENCAR, S.M. Chemical composition and botanical origin of red propolis, a new type of Brazilian propolis. Evidence-Based Complementary and Alternative Medicine, v. 4, p. 1/4, jul. 2007. SIMÕES, L.M.C.; GREGÓRIO, L.E.; DA SILVA FILHO, A.A.; DE SOUZA, M.L.; AZZOLINI, A.E.C.S.; BASTOS, J.K.; LUCISANO-VALIM, Y.M. Effect of Brazilian green propolis on the production of reactive oxygen species by stimulated neutrophils. Journal of Ethnopharmacology, v. 94, n. 1, p. 59-65, set. 2004. SIMPANYA, M.F. Dermatophytes: Their taxonomy, ecology and pathogenicity. Revista Iberoamericana de Micologia, 2000. Disponível em: <http://www.dermatophytes.reviberoammi col.com/p001012.pdf>. Acesso em 12 jan. 2005. SIMPANYA, M.F.; BAXTER, M. Multiple proteinases from two Microsporum species. Journal of Medical and Veterinary Mycology, v. 34, n. 1, p. 31-36, jan. 1996. SKOREPOVÁ, M.; HAUCK, H. Extracellular proteinases of Trichophyton rubrum and the clinical picture of tinea. Mycosen, v.30, n. 1, p.25-27. 1998. SMITH, D.; ONIONS, A.H.S. The preservation and maintenance of living fungi. IMI. Technical Hadbooks. 2. ed. n. 2. Egham: CAB International, 1994, 102 p. SOARES, M.M.S.R.; CURY, A.E. In vitro activity of antifungal and antiseptic agents against dermatophyte isolates from patients with tinea pedis. Brazilian Journal of Microbiology, v. 32, n. 2, p. 130-134, jun. 2001. SOARES JÚNIOR, F.A.; BRILHANTE, R.S.N.; CORDEIRO, R.A.; BRITO, E.H.S. SIDRIM, J.J.C.; ROCHA, M.F.G. Glucose improves the in vitro viability of Microsporum canis and Trichophyton mentagrophytes var. mentagrophytes. Journal of Microbiological Methods, v. 69, n. 1, p. 218-221, abr. 2007.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
104
SOUSA, J.P.B.; FURTADO, N.A.J.C.; JORGE, R.; SOARES, A.E.E.; J.K. BASTOS. Perfis físico-químico e cromatográfico de amostras de própolis produzidas nas microrregiões de Franca (SP) e Passos (MG), Brasil. Revista Brasileira de Farmacognosia, v. 17, n. 1, p. 85-93, jan./mar. 2007 SOUZA JUNIOR, P.; COLARES, R.A.; CALIXTO, R.S. Dermatofitoses, 1999. Disponível em: <http://www.geocities.com/CollegePark/Classroom/6137/dematofi.html>. Acesso em: 28 jul. 2007. SOYINKA, F. Epidemiologic study of dermatophyte infections in Nigeria (clinical survey and laboratory investigations). Mycopathologia, v. 63, n. 2, p. 99-103, 1978. SQUEO, R. F.; BEER, R.; SILVERS, D.; WEITZMAN, I.; GROSSMAN, M. 1998. Invasive Trichophyton rubrum resembling blastomycosis infection in the immunocompromised host. Journal of the American Academy of Dermatology, v. 39, 2 Pt. 2, p. 379-380, ago. 1998. STRYER, L. Bioquímica. 4. ed. Reverté: Barcelona. 2001. 458p. TAKASUKA, T. Amino acid- or protein-dependent growth of Trichophyton mentagrophytes and Trichophyton rubrum. FEMS Immunology and Medical Microbiology, v. 29, n. 4, p. 241-245. 2000. TAKIUCHI, I.; HIGUCHI, D.; SEI, Y.; KOGA, M. Isolation of an extracellular proteinase (keratinase) from Microsporum canis. Sabouraudia, v.20, n.4, p.281-288. 1982. TATEFUJI,T.; IZUMI, N.; OHTA, T., ARAI, SH., IKEDA, M., KURIMOTO, M. Isolation and identification of compounds from Brazilian propolis which enhance macrophage spreading and mobility. Biol. Pharm. Bull, v. 19, n.7, p. 966-970. 1996. TATSUMI, Y.; YOKOO, M.; SENDA, H.; KAKEHI, K. 2002. Therapeutic efficacy of topically applied KP-103 against experimental tinea unguium in guinea pigs in comparison with amorolfine and terbinafine. Antimicrobial Agents and Chemotherapy, v. 46, n. 12, p. 3797-3801, dez. 2002. TAZAWA,S.H.; WARASHINA, T.; NORO, T.; MIYASE, T. Studies on the constituents of Brazilian propolis. Chem. Pharm. Bull, v. 46, p. 1477-1479. 1998. TOMÁS-BARBERÁN, F.A.; GARCÍA-VIGUERA, C.; VIT-OLIVIER, P.; FERRERES, F.; TOMÁS-LORENTE, F. Phytochemical evidence for the botanical origin of tropical propolis from Venezuela . Phytochemistry, v. 34, n. 1, p. 191-196, ago. 1993.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
105
TRABULSI, L.R.; ALTERTHUM, F.; GOMPERTZ, O.F.; CANDEIAS, J.A.N. Microbiologia. São Paulo: Atheneu, 1999. 586 p. TRINDADE, M.B.; LOPES, J.L.S.; SOARES-COSTA, A.; MONTEIRO-MOREIRA, A.C.; MOREIRA, R.A.; OLIVA, M.L.V.; BELTRAMINI, L.M. Structural characterization of novel chitin-binding lectins from the genus Artocarpus and their antifungal activity. Biochimica et Biophysica Acta (Proteins & Proteomics), v. 1764, n. 1, p. 146-152, jan. 2006. TRUSHEVA, B.;POPOVA, M.; BANKOVA, V.; SIMOVA, S.;MARCUCCI, M.C.; MIORIN, P.L.; PASIN, F.R.; TSVETKOVA, I. Bioactive constituents of Brazilian red propolis. Evidence-Based Complementary and Alternative Medicine, v. 3, n. 2, p. 249-254., maio. 2006. TRUSWELL, S. ABC da Nutrição. São Paulo: Manoele, 1987. 93 p. TSANG, P., T. HOPKINS, AND V. JIMENEZ-LUCHO. Deep dermatophytosis caused by Trichophyton rubrum in a patient with AIDS. Journal of the American Academy of Dermatology, v. 34, n. 6, p. 1090–1091, jun. 1996. VALCIC, S.; MONTENEGRO, G.; TIMMERMANN, B. Lignans from Chilean propolis. Journal of Natural Products, v.61, n. 6, p. 771-775. 1998 VALSARAJ, R.; PUSHPANGADAN, P.; SMITT, U.W.; ADSERSEN, A.; CHRISTENSEN, S.B.; SITTIE, A.; NYMAN, U.; NIELSEN, C.; OLSEN, C.E. New anti-HIV-1, antimalarial, and antifungal compounds from Terminalia bellerica. Journal of natural products, v. 60, n. 7, p. 739-742. 1997. VAN PARIJS, J.; JOOSEN, H.M.; PEUMANS, W.J.; GEUNS, J.M.; VAN LAERE, A.J Effect of the Urtica dioica agglutinin on germination and cell wall formation of Phycomyces blakesleeanus Burgeff. Archives of Microbiology, v. 158, n. 1, p. 19-25, jun. 1992. VARGAS, D.M. ; AUDÍ, L.; CARRASCOSA,A. Peptídeos derivados do colágeno: novos marcadores bioquímicos do metabolismo ósseo. Revista da Associação Médica Brasileira, v. 43, n. 4, p. 367-370, out./dez. 1997. VENKATESAN, G.; SINGH, A. J. A. R.; MURUGESAN, A. G.; JANAKI, C.; SHANKAR, S. G. Trichophyton rubrum – the predominant etiological agent in human dermatophytoses in Chennai, India .African Journal of Microbiology Research, v. 1, n. 1, p. 009-012, maio. 2007. VIANI, F.C.; DOS SANTOS, J.I.; PAULA, C.R.; LARSON, C.E.; GAMBALE, W. Production of extracellular enzymes by Microsporum canis and their role in its virulence. Medical Mycology, v.39, n.5, p. 463-468, out. 2001.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
106
VIANI, F.C.; VIANI, P.R.C.; RIVERA, I.N.G.; DA SILVA, R.G.; PAULA, C.R. GAMBALE, W. Extracellular proteolytic activity and molecular analysis of Microsporum canis strains isolated from symptomatic and asymptomatic cats. Revista Iberoamericana de Micología, v. 24, p. 19-23. 2007. VIARD, B.; AL-MAHMOOD, S.; STREIBLOVA, E.; BONALY, R. Alternate interactions of the o-galactose-specific yeast lectin Kb-CWL I with sensitive yeast strains. FEMS Microbiology Letters, v. 107, n. 1, p. 17-24, fev. 1993. VICTORIA, M.B. Curativos. Disponível em : <http://www.fmt.am.gov.br/manual/ curativos. htm>. Acesso em: 20 mar. 2007 VLIETINCK, A. J., BRUYNE, T. D., APERS, S.; PIETERS, L. A. Plant-derived leading compounds for chemotherapy of human immunodeficiency virus (HIV) infection. Planta Medica, v. 64, n. 2, p. 97-109, mar. 1998. WACHTER, G.A.; HOFFMANN, J.J.; FURBACHER, T.; BLAKE, M.E.; TIMMERMANN, B.N. Antibacterial and antifungal flavanones from Eysenhardtia texana. Phytochemistry, v. 52, n. 8, p. 1469-1471, dez. 1999. WALKER, P.; CRANE, E. Constituents of Propolis. Apidologie, v. 18, p. 327-334. 1987. WALSH, T. J.; GROLL, A.H. Emerging fungal pathogens: evolving challenges to immunocompromised patients for the twenty-first century. Transplant Infectious Disease, v. 1, n. 4, p. 247–261, dez. 1999.
WANG, H.X.; GAO, J.; NG, T.B. A new lectin with highly potent antihepatoma Pleurotus ostreatus. Biochemical and Biophysical Research Communications, v. 275, n. 3, p. 810-816, set. 2000.
WANG, H.X.; LIU, W.K.; NG, T.B.; OOI, V.E.C.; CHANG, S.T. The immunomodulatory and antitumor activities of lectins from the mushroom Tricholoma mongolicum. Immunopharmacology, v. 31, p. 205-211. 1996. WANG, H.X.; NG, T.B. Examination of lectins, polysaccharopeptide, alkaloid, coumarin and trypsin inhibitor for inhibitory activity toward immunodeficiency virus reverse transcriptase and glycohydrolases. Planta Medica, v .67, p. 669-672. 2001. WANG, H.X.; NG, T.B. Isolation of cicadin, a novel and antifungal peptide from dried juvenile cicadas. Peptides, v. 23, n. 1, p. 7-11, jan. 2002. WANG, H.X.; NG, T.B.; LIU, W.K.; OOI, V.E.C.; CHANG, S.T. Isolation and characterization of two distinct lectins with antiproliferative activity from the cultured mycelium of the edible

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
107
mushroom Tricholoma mongolicum. International Journal of Peptide Research, v. 46, n. 6, p. 508–513, dez. 1995. WANK, N. C. F.; MONTEIRO, P. C. F.; WANKE, B.; NOGUEIRA, C. M.; PEREZ, M. A. Dermatofitoses no Rio de Janeiro. Estudo de fatores de risco em população adulta. Anais Brasileiros de Dermatologia, v. 66, n. 4, p. 171-174, 1991. WAWRZKIEWICZ, K.; WOLSKI, T.; LOBARZEWSKI, J. Sreening the keratinolytic activity of dermatophytes in vitro. Mycopathologia, v. 114, n. 1, p. 1-8, abr. 1991. WEITZMAN, I.; SUMMERBELL, R.C. The Dermatophytes. Clinical Microbiology Reviews, v. 8, p. 240–259, abr. 1995. WOJTASZEK, P. Oxidative burst: an early plant response to pathogen infection. Biochemistry Journal, v. 322, Pt. 3, p. 681–692, mar. 1997. WOLLENWEBER, E.; BUCHMANN, S.L. Feral honey bees in the Sonoran Desert: propolis sources other than poplars Populus spp. Zeitschrift fur Naturforschung C, v.52, n.7-8, p.530-535, 1997. WONG, J.H.; NG, T.B. Isolation and characterization of a glucose/mannose/rhamnos especific lectin from the knife bean Canavalia gladiata. Archives of Biochemistry and Biophysics, v. 439, n. 1, p. 91-98, jul. 2005. WONG, J.H.; NG, T.B. Purifcation of a trypsin-stable lectin with antiproliferative and HIV-1 reverse transcriptase inhibitory activity. Biochemical and Biophysical Research Communications, v. 301, n. 2, p. 545-550, fev. 2003. WONG, J.H.; WONG, C.C.T.; NG, T.B. Purification and characterization of a galactose-specific lectin with mitogenic activity from pinto beans. Biochimica et Biophysica Acta (General Subjects), v. 1760, n. 5, p. 808–813, maio. 2006. XIA, L.; NG, T.B. An antifungal protein from flageolet beans. Peptides, v.26, p. 2397-2403. 2005. YAN, Q.; JIANG, Z.; YANG, S.; DENG, W.; HAN, L. A novel homodimeric lectin from Astragalus mongholicus with antifungal activity. Archives of Biochemistry and Biophysics, v. 442, n. 1, p. 72-81, out. 2005. YE, X.; NG, T.B. Isolation of lectin and albumin from Pisum sativum var. macrocarpon ser. cv. sugar snap. The International Journal of Biochemistry & Cell Biology, v. 33, n. 1, p. 95-102, jan. 2001.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
108
YE, X.Y.; NG, T.B.; TSANG, P.W.K.; WANG, J. Isolation of a homodimeric lectin with antifungal and antiviral activities from red kidney bean (Phaseolus vulgaris). Journal of Protein Chemistry, v. 20, n. 5, p. 367-375, jul. 2001. ZAROR, I.; OTTH, L. SÁNCHES, J.W. Susceptibilidad de dermatofitos, Candida y otros fungos frente a ácidos grasos y escualeno. Revista Argentina de Micología, v. 10, n. 1, p. 6-9, jan./fev. 1987. ZHENG, W.F.; TAN, R.X.; YANG, L.; LIU, Z.L. Two flavones from Artemisia giraldii and their antimicrobial activity. Planta Medica, v. 62, n. 2, p. 160-162, abr. 1996.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
109
5. ARTIGOS 5.1. Taxonomic aspects and enzymatic profile of dermatophytes samples stored at the Mycotheca
Cultures Collection – University of Recife Mycology (URM).
5.2. Atividade antifúngica da própolis verde e da vermelha do Brasil em relação a espécies de
dermatófitos.
5.3. Susceptibilidade de espécies de Trichophyton a própolis verde e a vermelha do Brasil.
5.4. Efeito de lectinas de leguminosas do Brasil sobre o crescimento in vitro de dermatófitos.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
110
TAXONOMIC ASPECTS AND ENZYMATIC PROFILE OF DERMATOPHYTES SAMPLES
STORED AT THE MYCOTHECA CULTURES COLLECTION – UNIVERSITY OF RECIFE
MYCOLOGY (URM)
Artigo submetido para:
MYCOPATHOLOGIA
Qualis A
Fator de impacto 0,915 (Journal of Citation Report - 2006)

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
111
TAXONOMIC ASPECTS AND ENZYMATIC PROFILE OF DERMATOPHYTES SAMPLES
STORED AT THE MYCOTHECA CULTURES COLLECTION – UNIVERSITY OF RECIFE
MYCOLOGY (URM)
Ana Beatriz Sotero Siqueira*1,2, Idalina Inês F. N. Cambuim1, Rita de Cássia Maia3, Viviana
Giampaoli4, Cristina Souza-Motta1, Lusinete Aciole de Queiroz1, Ana Porto2
1Department of Mycology, Federal University of Pernambuco, Recife-Brazil 2 Keizo Asami Immunopathology Laboratory, Federal University of Pernambuco, Recife-Brazil 3 Aggeu Magalhães Research Center – CPqAM / FIOCRUZ, Recife-Brazil 4 Statistics Department, University of São Paulo, Brazil
*Email for correspondence: [email protected]

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
112
ABSTRACT
Dermatophytes, the etiologic agents of dermatophytosis, are keratinophilic fungi classified in three
genera Microsporum, Trichophyton and Epidermophyton. These fungi are able to produce
a variety of enzymes, which are secreted to the extracellular medium in order to facilitate
substrate degradation. With the purpose of evaluating taxonomic characteristics and the
enzymatic profile of dermatophyte samples stored for different periods of time at the
Mycotheca Culture Collection - University of Recife Mycology (URM), from the Federal
University of Pernambuco, 30 samples from six different species of dermatophytes were
used, being five from each of the following: Microsporum canis, Microsporum gypseum,
Trichophyton tonsurans, Trichophyton mentagrophytes, Trichophyton rubrum and
Epidermophyton floccosum. Every sample, preserved from 2 to 22 years, was taxonomically
confirmed. It was observed that the storage period did not affect growth ability and sporulation
among the dermatophytes (p value < 0.01) and storage in general, induces M. canis pleomorphism,
albeit the period of time. Protease activity was observed in 57% of the samples. All of them were
capable of growing in agar-keratin and agar-gelatin, indicating keratinase and collagenase activity,
respectively. None of the samples showed phospholipase activity, but 90% of them showed lipase
activity. The storage period did not intervene on protease, keratinase, collagenase, phospholipase,
and lipase production. Considering this, the fungal preservation at the Mycotheca Culture
Collection - URM, has been shown efficient since the storage time period did not interfere in
confirming taxonomic data and enzyme expression of the different genera.
Key words: enzymatic profile, stored dermatophytes, taxonomic characteristics

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
113
INTRODUCTION
The dermatophytes are a group of closely related fungal species that have the capacity to
invade keratinized tissue of humans and others animals and produce dermatophytosis. The
organisms belong to three genera including Microsporum, Trichophyton and Epidermophyton. The
identification is based mainly on its morphological and physiological aspects [1, 2], even though
the pleomorphism, common in dermatophytes, difficult the manifestation of these aspects making
the identification, sometimes, flawed. These fungi show similar taxonomic, pathogenic and
antigenic characteristics, but with nutritional and enzymatic differences [3].
These fungi produce a variety of enzymes for the degradation of proteins, including
hydrolytic enzymes like protease, ketarinase, phospholipase and lipase, which are important in the
pathogenicity and can be secreted to the culture medium. Enzymatic detection has been used on
taxonomic studies for species differentiation as well in isolates from the same species allowing the
evidence of variability in epidemiological studies [4, 5, 6, 7].
The preservation of fungi which are pathogenic to man and animals is important for
research and biotechnology, thus, determining the correct preservation method for each fungal
species and periodical monitoring to check their identity has become a requirement [8, 9].
Conservation of morphological, physiological, genetic and metabolic stability is crucial for many
purposes and is vital for isolates used as medical reference strains, for chemotaxonomic studies, or
in the commercial production of biochemicals [10].
Numerous methods for the storage of fungal cultures have been evaluated, and many have
been found to be suitable for high quality storage of at least certain fungal strains. Methods to date
include storage in sterile water, saline, grain or soil at room temperature, storage under mineral or
paraffin oil, freezing at -20 or -80 Cº, freeze-drying, cryopreservation in liquid nitrogen, and
subculturing in sterile distilled water, under mineral oil, drying on silica gel, soil storage or by
freeze-drying [11, 12].
The Mycotheca of the Department of Mycology from the Center for Biological Sciences, of
the Federal University of Pernambuco, registered on the Commonwealth Mycological Institute
under URM (University of Recife Mycology) comprises to date, around 8.000 filamentous and
yeast fungal cultures, kept in duplicate, identified at the species level and preserved by freeze-
drying, mineral oil and distilled sterile water
This work main goal is to evaluate the taxonomic characteristics and enzymatic profile of
the dermatophyte samples stored for different periods of time at the Mycotheca Cultures
Collection – URM, Department of Mycology, Federal University of Pernambuco, Brazil.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
114
MATERIAL E METHODS
Fungal Samples
A total of 30 dermatophytes cultures were tested, including 5 strains of each of the
following species: Microsporum canis, M. gypseum, Trichophyton mentagrophytes, T. tonsurans,
T. rubrum and Epidermophyton floccosum. These fungi were obtained from the culture collection
and were preserved in mineral oil [13]. Most of them were clinical isolates, while some were
isolated from soil. They were subsequently maintained at the Mycotheca Cultures Collection –
URM (Table 1).
Taxonomic Characteristics Evaluation
Methods for evaluation of purification and taxonomic aspects of fungi were used as
described by some authors [3, 14]. The isolated colonies were cultured on Sabouraud Dextrose
Agar medium (SDA) and Dermatophyte Test Medium (DTM) on Petri dishes incubated at 30ºC
for 15 days. The microscopic characteristics were observed in accordance with Ridell technique on
SDA, DTM and Potato-Dextrose-Agar medium (PDA) [3].
In order to differentiate T. mentagrophytes from T. rubrum species, the urease activity test
was employed [3, 15]. Colony fragments from T. mentagrophytes and T. rubrum were plated onto
the urea agar medium by Christensen and observed until the seventh day in order to verify de color
switch from yellow to a fucsia pink color, indicating the presence of the enzyme.
Fungal stock cultures were kept on SDA slants at 4 ºC for the enzymes assays.
Evaluation of the Enzymatic Profile
Methods to detect the enzymatic activity of protease, keratinase, collagenase,
phospholipase and lipase were carried out in Petri dishes. After the medium was autoclaved and
cooled to approximately 50ºC, it was distributed on Petri dishes. Fragments of 5 mm were
removed from young colonies grown on Petri dishes containing SDA and each fragment was
placed on the center of the Petri dish containing the specific solid media. After the inoculation, the
isolates were maintained for 20 days at 30ºC.
Protease assay was carried out as described by Kantarcioglu and Yücel [16]. The substrate
skimmed milk medium was prepared and consisted of two phases: 1st phase - skimmed milk,
10g% in distilled water; 2nd phase - agar 2g% in distilled water. When transparent halo around of
the colony was present, acidified solution of mercury chlorate was added to ascertain whether the
halo occurred by acidic or alkaline metabolic products from milk, or if there was a true proteolysis
indicating that casein was really degraded. To detect the enzymatic activity the activity zone (ZA)
was calculated represented by the colony diameter, divided by the diameter of the colony plus the
diameter of transparent halo. The ZA value represents the enzymatic activity when: ZA is between

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
115
0,9 and 1,0 = very low enzymatic activity (+); between 0,89 and 0,80 = low enzymatic activity (+
+); between 0,79 and 0,70 = high enzymatic activity (+ + +) and < 0,69 = very high enzymatic
activity (+ + + +).
Keratinase and collagenase assays were performed [17], when using keratin and gelatin as
substrates, respectively. Preparation of soluble keratin was obtained by heating chicken feathers in
DMSO (dimethyl sulphoxide) at 100ºC for 2 hours, being precipitated thereafter with acetone.
Soluble keratin at 0,012% was added to the agar medium, containing agar 2% in distilled water.
The test medium to verify the collagenase activity contained gelatin, 1% and agar, 2% in distilled
water. The ability of utilizing keratin or gelatin substrates as the only sources of carbon and
nitrogen, and growth on solid medium containing these substances, indicate keratinase and
collagenase activity, respectively.
To detection of phospholipase activity, two methods were used. For the first method [18],
fragments of cultures were inoculated in the center of Petri dishes containing the phospholipase
medium, which consisted of 0,2% soy lecithin, peptone, 1%, glucose, 2% sodium chloride, 5,73 %,
calcium chloride 0,05% and agar 2%, in distilled water. The second method [19] differs from the
first by using two egg yolks, replacing the “egg yolk” of Difco Laboratories. The formation of an
opaque halo around the colonies indicated phospholipase activity, which was measured. This was
accomplished by calculating the activity zone (Pz), represented by the colony diameter, divided by
the diameter of the colony plus the zone of precipitation. When the Pz was equal to 1.0, the
samples were considered negative and when the Pz was less than 1.0, the phospholipase activity
was considered as positive.
The lipase assay was evaluated as previously described [6]. The lipase medium consisted of
peptone, 1%, sodium chloride, 0,5%, calcium chloride, 0,01% and agar, 2%, in distilled water.
Tween 20 was autoclaved separately and was used as a substrate at a final concentration of 1%.
Formation of a precipitate halo around the colonies indicated lipase production, which was
measured.
Statistical Analysis
In order to evaluate the relationship between dermatophytes samples growth and keratinase
and collagenase production, the Kruskal-Wallis [20] was used.
To analyze the influence of the storage time associated to enzymatic activity, the Gamma
family generalized models were applied [21].

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
116
RESULTS
Taxonomic Characteristics
All dermatophytes samples preserved from 2 to 22 years were analyzed taxonomically,
considering their macroscopic and microscopic characteristics. Regarding texture, the samples
showed typical characteristics to each species, with two M. gypseum (3060 e 3645) samples and all
M. canis samples showing cotton-like colony texture. The two M. gypseum samples had been
preserved for 18 and 12 years respectively, and the M. canis samples had been preserved from 2 to
9 years.
Regarding the surface, 60% of the M. canis (4678, 4225, 4962), M. gypseum (6034, 3645,
4279) and 80% of the T. mentagrophytes (4156, 3239, 2875, 2875) samples were lobated, and had
been stored for a period of 2 to 21 years. The T. tonsurans, T. rubrum and E. floccosum samples
showed discrete surface appearance differences, with the first two been preserved for 2 to 5 years
and the last for 3 to 17 years.
When observing microscopic characteristics, the DTM and PDA media, were considered
the most suitable to induce sporulation. Only one sample of M. canis sporulated in SDA medium
and all of them sporulated on DTM and PDA media. One sample of M. gypseum (3060) was
subjected to microcultivation in SDA and DTM media for over a month and sporulation was not
observed, which was accomplished only in PDA medium. None of the E. floccosum samples
sporulated in SDA medium, but all of them in DTM and PDA media. The sporulation period
varied between 7 and 30 days, depending on the species being cultivated and the culture medium
used, with an average of 14 to 21 days, with one E. flocossum sample sporulating on the 30 day
(Table 2).
Some of the M. canis (4962) and M. gypseum (4964, 3645, 4279) samples were found
piriform and claviform microconidia. In one T. mentagrophytes (2875) sample was observed
predominantly piriform and claviform microconidia. In order to differentiate that species from T.
rubrum, was used the urease test, which turned positive in the fifth day. The other T.
mentagrophytes samples also produced these samples between the third and fifth days, whereas T.
rubrum samples did not produce it until the seventh day.
Considering the sample storage time, it was observed that the fungus growth and
sporulation abilities in culture medium was determined by the species (p value < 0.01) and was not
observed a relationship between the species and storage period (p value < 0.05).
Enzymatic Profile Evaluation
Among 30 dermatophytes samples were observed protease activity in 57% of them. All
samples were capable of growing in a culture medium containing agar-keratin and agar-gelatin.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
117
Phospholipase activity was not observed in any if the samples, while lipase activity was observed
in 90% of the samples.
A high protease activity was observed in three M. canis (4156, 4678, 4158), two M.
gypseum (3060, 4279), two T. mentagrophytes (4156, 4422), one T. tonsurans (4945) and four E.
floccosum (3345, 4151, 4799, 4798) samples. A weak activity was observed in one M. canis
(4962), one M. gypseum (6034), two T. mentagrophytes (3239, 2875) and one T. rubrum (4725)
sample. All other samples did not show any protease activity.
The keratinase production was observed in all samples with the average of the colony
diameter of 6,59; 6,53; 5,29; 4,86; 4,68; 3,60 centimeters for M. gypseum, M canis, T. rubrum, T.
mentagrophytes, T. tonsurans and E. floccosum, respectively (Kruskal-Wallis test 15.7159, p-value
= 0.0016).
Collagenase production was also observed in all samples with the average of the colony
diameter of 7,73; 6,50; 5,63; 5,45; 4,63; 2,89 centimeters for M. canis, M. gypseum, T. rubrum, T.
mentagrophytes, T. tonsurans and E. floccosum, respectively (Kruskal-Wallis test 20.3303, p-value
< 2.2e-16).
The precipitation indicating lipase activity reached diameters between 3,1x 10-3 and 1,55
cm. The samples that showed higher precipitates were two M. canis (4678, 4158), two M. gypseum
(4964, 3645), three T. mentagrophytes (4156, 4422, 2875), two T. tonsurans (4965, 4945), two T.
rubrum (4724, 4728) and three E. floccosum (3345, 4799, 3195) samples. Such activity was not
observed in one M. gypseum (4279), T. mentagrophytes (2785) and T. tonsurans (4968) sample.
The precipitate diameter averages were 5.318, 4.70, 4.60, 3.79, 3.52, 3.142 centimeters observed
in M. canis, T. rubrum, T. mentagrophytes, M. gypseum, T. tonsurans, E. floccosum, respectively
(Kruskal-Wallis test 4.8344, p-value = 0.456). The presence of lipase and protease activity was
observed to be inversely proportional, especially for M. gypseum and T. rubrum species.
When analyzing the enzymatic activity, was observed that the storage period did not
influenced the production of protease (p-value >0.8), keratinase (p-value >0.9), collagenase (p-
value >0.7), phospholipase (p-value >0.7) and lipase (p-value >0.7). Although, it has been shown
that the fungal species influenced the activity of those enzymes (p-value <0.01).
The results obtained in this section can be observed on Table 3.
DISCUSSION
Texture variations and surface aspect observed in the dermatophytes samples analyzed are
typical of pleomorphism and are in agreement with the description others authors [1, 2]. Those
genetic alterations can happen with multiple fungal passages on culture medium or when the

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
118
fungus is subjected to a preservation condition with a metabolism reduction, showing very few to
none pigmentation [11].
The species that varied the most regarding the macroscopic findings were M. canis, M.
gypseum and T. mentagrophytes. It has been shown that some preserved dermatophyte species,
especially M. canis and T. mentagrophytes, has shown taxonomic variations quite unlike the
parental strains [11, 12].
Pleomorphic colonies observed in SDA medium could have bare mycelial areas, which,
eventually, would cover the whole colony and impair sporulation [2, 11]. Although the storage
period has not caused loss of the sporulation ability, it was observed that it became more difficult
to sporulate in SDA medium, which is usually used for maintenance [1, 22, 23]. The PDA medium
is well known and recognized to induce sporulation in dermatophytes [2, 24, 25].
Microconidia were observed in M. canis and M.gypseum samples similarly to the
Trichophyton sp samples what already have been mentioned by other authors [14, 23]. The
ontogeny of the holothallic conidia of Microsporum and Trichophyton is essentially the same [26].
Their only difference is the macroconidial cell-wall thickness and presence of echinulations in
Microsporum species, which are absent in Trichophyton species [1, 14].
According to the results obtained, it was observed that the preservation seems to alter
macroscopic aspects expression typical of each species, but the storage period did not influence
such finding. Moreover, the storage procedure can induce taxonomic changes and/or alterations in
fungal cell components [12].
The M. canis samples preserved less than 10 years and two M. gypseum samples preserved
over 10 years, showed pleomorphic characteristics. Confirming that the storage period did not
influence the maintenance of the macroscopic aspects in this genus, showing different outcomes to
the preservation process. Similar observations have been described by others authors [12, 27].
The Trichophyton samples preserved for up to 5 years conserved the typical macroscopic
characteristics. In T. mentagrophytes, samples preserved for over 10 years, were pleomorphic,
nevertheless, not much can be established about the other species since they had been preserved
for up to 4 years. In this particular case, it was observed that the T. mentagrophytes samples had
been influenced by the storage time. This has been discussed in a similar fashion by other authors
[11, 12].
The urease production by T. rubrum samples and the lack of production by T.
mentagrophytes samples, as described in the present work, allow stating that these characteristics
are specie-related. The ability to hydrolyze urea is considered the biochemical evidence most

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
119
widely used to differentiate T. mentagrophytes from T. rubrum, with the first being able to
hydrolyze urea [1, 3, 14].
The lack of association between species and storage period, demonstrated in the present
work, shows that the time influenced equally all samples and none in a special manner.
The use of solid medium allows the detection of an enzymatic activity since dermatophytes
secrete enzymes into the culture medium during growth what improves the substrate assimilation
[6, 28, 29].
The results obtained in this work show that dermatophytes are able to synthesize different
types of enzymes and that the variability in enzymatic activity was observed among different
species, as well as among samples of the same species [4, 7, 30].
The different enzymatic profile could be associated with the ecologic aspect
(anthropophilic, zoophilic and geophilic) of each species. The gradual adaptation of the soil to the
man and other animals involved nutritional adjustment along with the ability to break and utilize
bigger molecules from the skin and annexes in smaller molecules to digest [5, 30].
The findings obtained in this work, regarding the protease activity, are in agreement with
other authors, who also reported this activity in a majority of M. canis e M. gypseum samples [ 7,
31]. It was observed a variation on protease activity among Tricophyton species and an increased
activity of this enzyme in T. mentagrophytes. Similar results have been reported [4, 6, 30]. The
expression of this enzyme’s activity in a T. tonsurans and a T. rubrum sample could be explained
through the anthropophilic behavior of these specie, since they are well adapted to human
parasitism and therefore, could have lost or attenuated their synthesis capability [32, 33].
The development of colonies from all dermatophytes samples in agar-keratin and agar-
gelatin media, demonstrates the presence of keratinases and collagenases, respectively. The
presence of one substrate as a sole nutrient source, demonstrates de ability of those samples to
synthesize enzymes to degrade the substrate and utilize it to the fungal development [34]. These
findings are in agreement with other authors, who detected keratinase and collagenase activity in
dermatophyte samples [6, 33]. However, Abdel-Rahman [4] did not detect the keratinase activity
in a Trichophyton sp. sample among 12 samples studied. The higher keratinase and collagenase
activity was observed among Microsporum samples, and the ecologic aspects aforementioned also
corroborate this finding.
The lack in the phospholipase detection in dermatophytes is in agreement with Dos Santos
et al. [6] who suggest that this criterion should not be taken into consideration as a biotype
differentiation marker. Nevertheless, this parameter was utilized by Maia et al. [7] to characterize
phenotypically new M. canisi isolates.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
120
The lipase activity was observed in the majority of dermatophyte samples studied and
similar results have been reported [35]. Other authors mentioned the finding of lipase activity in
dermatophytes specimens such as M. gypseum, T. mentagrophytes [30] and T. rubrum [6].
The data reported in this work shows an inverse correlation between proteases and lipases
activity that can be associated with a compensatory balance between the hyper-activation of one
enzyme and lower activity of the other. It has been established that the skin is the first line of
defense against infections caused by pathogenic microorganisms and also that there are fatty
acids with fungal-static action [36, 37] that complicate infections by dermatophytes,
characterizing a natural defense mechanism of the organism. Tinea capitis, as an example, is well
known as a dermatophytosis occurring almost exclusively in infants, spontaneously decreasing in
puberty due to the production, under hormonal control, of fatty acids with fungal-static properties
[34, 37, 38]. Consequently, those fungi that depend on the enzymatic activity to establish the
inflammatory process, especially extracellular protease and lipase [34, 36] are likely to
compensate the higher activity of one enzyme by the lowering of another.
It was observed that the dermatophyte preservation in the Mycotheca Culture Collection -
URM was efficient, since the period of storage did not influenced data collecting to confirm
taxonomic characteristics of the different species, neither to the expression of enzyme activities.
The utilization of these parameters, to evaluate fungal preservation efficiency, have been
mentioned by other authors [12, 27, 39].
Based on the results obtained, it was concluded that different fungi species vary their
responses to one or more parameters, but that a group of characteristics is common to a specific
group.
REFERENCES
1. Weitzman I, Summerbell RC. The Dermatophytes. Clinical Microbiology Reviews 1995; 8 (2):
240–259.
2. Larone DH. Culture and Identification of Dermatophytes. Clinical Microbiology Newsletter.
1996; 18 (5): 33-40.
3. Lacaz CS, Porto E, Martins JEC, Heins-Vaccari EM, Melo NT. Tratado de Micologia
Médica. Rio de Janeiro: Sarvier, 2002; 252-340.
4. Abdel-Rahman SM. Polymorphic exocellular protease expression in clinical isolates of
Trichophyton tonsurans. Mycopathologia 2000; 150 (3): 117-120.
5. Abdel-Rahman SM. Trichophyton tonsurans exocellular protease expression: correlation with
clinical presentation in tinea capitis. Clinical and Experimental Dermatology 2002; 27 (4): 268-
271.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
121
6. Dos Santos JI, Vicente EJ, Paula CR, Gambale W. Phenotypic Characterization of Trichophyton
rubrum isolates from two geographic locations in Brazil. European Journal of Epidemiology 2001;
17 (8): 729-735.
7. Maia MLS, Dos Santos JI, Viani FC, Larsson CE, Paula CR, Gambale W. Phenotypic
characterization of Microsporum canis isolates from cats and dogs. Mycoses 2001; 44 (11-12):
480-486.
8. Lima RF, Borba CM. Viability and morphological characteristics and dimorphic ability by
different methods. Revista Iberoamericana de Micología 2001; 18 (4): 191-196.
9. Bezerra CCF, Lima RF, Lazera MS, Wanke B, Borba C.M. Viability and molecular
authentication of Coccidioides immitis strains from culture collection of the Instituto Oswaldo
Cruz, Rio de Janeiro, Brazil. Revista da Sociedade Brasileira de Medicina Tropical 2006; 39 (3):
241-244.
10. Baker M, Jeffries P. Use of commercially available cryogenic vials for long-term preservation
of dermatophyte fungi. Journal of Clinical Microbiology 2006; 44 (2): 617-618.
11. Deshmukh SK. The maintenance and preservation of keratinophilic fungi and related
dermatophytes. Mycoses 2003; 46: 203-207.
12. Borman AM, Szekely A, Campbell CK, Johnson EM. Evaluation of the viability of pathogenic
filamentous fungi after prolonged storage in sterile water and review of recent published studies on
storage methods. Mycopathologia 2006; 161 (6): 361-368.
13. Sherf AF. A method for maintaining Phytomonas sepedomica for long periods without
transfer. Phytopathology 1943; 31: 30-32.
14. Rebell G, Taplin D. Dermatophytes: their recognition and identification. Florida:
University of Miami Press, 1979; 124.
15. Philpot C. The differentiation of Trichophyton mentagrophytes from T. rubrum by a
simple urease test. Sabouraudia 1967; 5: 189-193.
16. Kantarcioglu, AS, Yücel, A. Phospholipase and protease activities in clinical Candida
isolates with reference to the sources of strains. Mycoses 2001; 45 (14): 160-165.
17. Wawrzkiewicz K, Wolski T, Lobarzewski J. Sreening the keratinolytic activity of
dermatophytes “in vitro”. Mycopathologia 1991; 114 (1): 1-8.
18. Price MF, Cawson RA. Phospholipase activity in Candida albicans. Saboraudia 1977; 15: 79-
185.
19. Price MF, Wilkinson ID, Gentry LO. Plate method for detection of phospholipase activity in
Candida albicans. Sabouraudia 1982; 20: 7-14.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
122
20. Hollander M, Wolfe DA. Nonparametric Statistical Methods. New York: John Wiley & Sons,
1999; 817.
21. Mccullagh P, Nelder JA. Generalized Linear Models. London: Chapman and Hall. 1989; 540.
22. Hainer BL. Dermatophyte Infections. American Family Physician 2003; 67 (1): 101-108.
23. Costa M, Passos XS, Souza LKH, Miranda ATB, Lemos JA, Oliveira Júnior JG, Rodrigues
Silva MRR. Epidemiology and etiology of dermatophytosis in Goiânia, GO, Brazil. Revista da
Sociedade Brasileira de Medicina Tropical 2002; 35(1): 19-22.
24. Fernandez-Torres B, Inza I, Guarro J. In Vitro Activities of the New Antifungal Drug
Eberconazole and Three Other Topical Agents against 200 Strains of Dermatophytes. Journal Of
Clinical Microbiology 2003; 41 (11): 5209-5211.
25. Barros MES, Santos DA, Hamdan JS. “In vitro” methods for antifungal susceptibility testing
of Trichophyton spp. Mycological Research 2006; 110 (11): 1355-1360.
26. Cole GT, Samson R A. Patterns of development in conidial fungi. London: Pittman, 1979;
190.
27. Brilhante, R.S.N.; Cavalcante, C.S.P.; Soares-Júnior, F.A.; Monteiro, A.J.; Brito, E.H.S.;
Cordeiro, R.A.; Sidrim, J.J.C.; Rocha, M.F.G. Evaluation of Microsporum canis in different
methodos of storage. Medical Mycology 2004; 42 (6): 499-504.
28. Hankin L, Anagnostakis SL. The use of solid media for detection of enzym production by
fungi. Mycologia 1975; 67: 597-607.
29. Friedrich J, Gradisar H, Mandin D, Chaumont JP. Screening fungi for synthesis of
keratinolytic enzymes. Letters in Applied Microbiology 1999; 28: 127-130.
30. Muhsin TM, Salih TH. Exocellular enzyme activity of dermatophytes and other fungi isolated
from ruminants in Southern Iraq. Mycopathologia 2000; 150 (2): 49-52.
31. Bruguera MT, Abarca ML, Bragulat MR, Cabañes FJ. Determinación de actividades
enzimaticas en cepas de Microsporum canis y Microsporum gypseum. Revista Iberoamericana de
Micología 1997; 14: 119-124.
32. Skorepová M, Hauck, H. Extracellular proteinases of Trichophyton rubrum and the
clinical picture of tinea. Mycosen 1998; 30 (1): 25-27.
33. Takasuka, T. Amino acid- or protein-dependent growth of Trichophyton mentagrophytes and
Trichophyton rubrum. FEMS Immunology and Medical Microbiology 2000; 29 (4): 241-245.
34. Ogawa H, Summerbell RC, Clemons KV, Kogas T, Ran YP, Rashid A, Sohnle PG, Stevensm
DA, Tsuboi R. Dermatophytes and host defence in cutaneous mycoses. Medical Mycology 1998;
36 (suppl. I): 166-173.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
123
35. Muhsin TM, Aubaid AH, Al-Duboon AA. Extracellular enzyme activities of dermatophytes
and yeast isolates on solid media. Mycoses 1997; 40 (11-12): 465-469.
36. Zaror I, Otth L, Sánches JW. Susceptibilidad de dermatofitos, Candida y otros fungos frente a
ácidos grasos y escualeno. Revista Argentina de Micología 1987; 10 (1): 6-9
37. Hernández T, Machado S, Carvalho S, Selores M. Tinhas do couro cabeludo na idade
pediátrica. Nascer e Crescer 2004; 13 (1): 23-26.
38. Azulay DR. Dermatofitoses. Anais Brasileiros de Dermatologia 1989; 64 (suppl. 1): 93-96.
39. López-Martinez R, Hernández-Hernández F, Bazán-Moraand E, Castañón-Olivares LR.
Comparative study of two culture conservation methods in medical mycology. World Journal of
Microbiology and Biotechnology 1999; 15 (4): 471-474.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
124
Table 1. Dermatophyte species preserved in mineral oil and storaged at the Mycotheca
Cultures Collection - University of Recife Mycology (URM).
Specie Registration Nº Storage year Storage period Substrate (URM) (year)
Microsporum canis M. canis M. canis M. canis M. canis M. gypseum M. gypseum M. gypseum M. gypseum M. gypseum Trichophyton mentagrophytes T. mentagrophytes T. mentagrophytes T. mentagrophytes T. mentagrophytes T. tonsurans T. tonsurans T. tonsurans T. tonsurans T. tonsurans T. rubrum T. rubrum T. rubrum T. rubrum T. rubrum Epidermophyton floccosum E. floccosum E. floccosum E. floccosum E. floccosum
4962 4225 4154 4678 4158 4964 3060 6034 3645 4279 4156 3239 2785 4422 2875 4965 4963 4968 4945 4946 4725 4724 4727 4728 4569 3345 4151 4799 4798 3195
2005 1999 1999 2003 1998 2005 1989 2005 1995 2000 1999 1992 1985 2002 1986 2005 2005 2005 2004 2004 2003 2003 2003 2003 2004 1993 1999 2004 2004 1990
2 8 8 4 9 2
18 2
12 7 8
15 22 5
21 2 2 2 3 3 4 4 4 4 3
14 8 3 3
17
ES ES ES ES ES ES UN ES ES BR BR ES SO ES UN ES ES ES ES ES ES ES ES ES US ES ES ES ES ES
ES: Epidermal scales; UN: Unkown; BR: Beaches of Recife; SO: Soil; US: ungueal scales

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
125
SDA: Sabouraud Dextrose Agar medium
DTM: Dermatophyte Test Medium
PDA: Potato Dextrose Ágar médium
- : spore absence
Table 2. Sporulation period in different culture media from dermatophytes
samples stored in mineral oil at the Mycotheca Cultures Collection -
University of Recife Mycology (URM).
Microsporum canis M. canis M. canis M. canis M. canis M. gypseum M. gypseum M. gypseum M. gypseum M. gypseum Trichophyton mentagrophytes T. mentagrophytes T. mentagrophytes T. mentagrophytes T. mentagrophytes T. tonsurans T. tonsurans T. tonsurans T. tonsurans T. tonsurans T. rubrum T. rubrum T. rubrum T. rubrum T. rubrum Epidermophyton floccosum E. floccosum E. floccosum E. floccosum E. floccosum
4962 4225 4154 4678 4158 4964 3060 6034 3645 4279 4156 3239 2785 4422 2875 4965 4963 4968 4945 4946 4725 4724 4727 4728 4569 3345 4151 4799 4798 3195
Specie Registration Nº Sporulation (days) (URM) SDA DTM PDA
21 - - - -
14 -
14 14 14 14 - -
14 21 14 14 14 14 21 7 7
14 14 7 - - - - -
21 21 21 21 21 14 - 7 7 7
14 21 14 14 21 21 21 21 21 21 14 14 14 14 14 14 21 30 14 14
21 21 21 21 21 14 21 14 14 14 14 21 14 14 21 14 14 14 14 21 14 14 14 14 14 14 21 30 14 14

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
126
Table 3. Enzymatic profile of dermatophytes samples stored in mineral oil at the Micotheca
Cultures Collection - University of Recife Mycology (URM) between 2 and 22 years.
Dermatophyte (URM)
Protease Keratinase Collagenase Phospholipase Lipase (ZA) (φ colony) (φ colony) (φ halo) (φ precipitated)
Enzyme
Microsporum canis 4962 4225 4154 4678 4158 Microsporum gypseum 4964 3060 6034 3645 4279 Trichophyton mentagrophytes 4156 3239 2785 4422 2875 Trichophyton tonsurans 4965 4963 4968 4945 4946 Trichophyton rubrum 4725 4724 4727 4728 4569 Epidermophyton floccosum 3345 4151 4799 4798 3195
+ + -
+ + + + + + + + + + + +
-
+ + + + + +
- + + + +
+ + + + + +
- + + + +
+ + - - -
+ + + + -
+ + - - - -
+ + + + + + + + + + + + + + + +
-
7,35 7,65 6,85 6,65 4,14
6,25 5,15 6,45 7,35 7,75
5,53 4,75 6,20 3,75 4,05
3,85 5,11 3,85 5,95 4,65
4,44 4,40 6,74 5,75 5,15
4,75 4,15 2,75 3,23 3,11
7,75 7,95 7,12 7,25 7,12
7,15 4,65 6,55 7,25 6,90
6,25 4,48 7,95 3,35 5,22
5,94 5,25 3,85 5,45 4,15
5,94 515 4,47 6,35 6,25
2,75 3,40 2,25 3,41 2,65
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
9,3 x 10-3 8 x 10-3
6,8 x 10-3 1,55 1,02
1,45 5,1 x 10-3 7,5 x 10-3
1,09 -
1,40 6,5 x 10-3
- 1,50 1,77
1,30 3,1 x 10-3
- 1,15
3,7 x 10-3
8,3 x 10-3 1,44
6 x 10-3 1,14
6,7 x 10-3
1,30 7,1 x 10-3
1,05 4,7 x 10-3
1,10
ZA: activity zone, φ: diameter in centimeters, - : absence of enzymatic activity, + +: weak proteasic activity, + + + +: very strong proteasic activity

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
127
ATIVIDADE ANTIFÚNGICA DA PRÓPOLIS VERDE E DA VERMELHA DO BRASIL EM
RELAÇÃO A ESPÉCIES DE DERMATÓFITOS
Artigo a ser submetido para:
JOURNAL OF ETHNOPHARMACOLOGY
Qualis A
Fator de impacto 1,625 (Journal of Citation Report - 2006)

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
128
ATIVIDADE ANTIFÚNGICA DA PRÓPOLIS VERDE E DA VERMELHA DO BRASIL EM
RELAÇÃO A ESPÉCIES DE DERMATÓFITOS
Ana Beatriz Sotero Siqueira*a,b, Bruno Severo Gomesa,b, Rita Maiac, Sheila Abreud, Cristina
Souza-Mottaa, Lusinete Aciole de Queiroza, Ana Lucia F. Portob,e.
aDepartamento de Micologia, Universidade Federal de Pernambuco (UFPE), Recife-Brasil bLaboratório de Imunopatologia Keizo Asami (LIKA), UFPE, Recife-Brasil cCentro de Pesquisas Aggeu Magalhães – CPqAM / FIOCRUZ, Recife-Brasil dPharma Nectar®, Belo Horizonte-Brasil eDepartamento de Morfologia e Fisiologia Animal, Universidade Federal Rural de Pernambuco
(UFRPE) / LIKA-UFPE, Recife-Brasil
*Email para correspondência: [email protected]

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
129
Resumo
Relevância Etnofarmacológica: Própolis é um material elaborado por abelhas a partir de exsudatos
resinosos de plantas. Algumas amostras de própolis do Brasil já apresentaram atividade
antifúngica.
Objetivo: Verificar a antifúngica da própolis verde e da vermelha do Brasil em relação a amostras
de Microsporum canis, M. gypseum e Epidermophyton floccosum.
Material e Métodos: Foi analisada a atividade antifúngica in vitro do extrato alcoólico e aquoso da
própolis verde e do extrato alcoólico da própolis vermelha em relação a espécies de
demrmatpofitos, sendo utilizado como controle itraconazol e terbinafina.. A concentração
inibitória mínima foi determinada de acordo com o método de adotado pelo “Clinical and
Laboratory Standards Institute”. A concentração fungicida mpinima foi determinada pela ausência
de crescimento em meio de cultura Sabouraud líquido.
Resultados: As espécies pertencentes ao gênero Microsporum foram susceptíveis a atividade
fungistática do extrato alcoólico da própolis verde e da vermelha nas concentrações entre 64-
1024µg/ml e 16-512µg/ml, respectivamente. As amostras de E. floccosum foram susceptíveis a
atividade fungistática desses mesmos extratos entre 8-512µg/ml e 16-256µg/ml, respectivamente.
A atividade fungicida do extrato alcoólico da própolis verde e da vermelha foi constatada entre
256-1024µg/ml e 128-1024µg/ml, respectivamente, para as amostras de dermatófitos utilizadas. O
extrato aquoso da própolis verde não apresentou atividade antifúngica.
Conclusões: Os dados obtidos demonstram que as amostras de M. canis e E. floccosum foram mais
susceptíveis aos extratos alcoólicos das própolis utilizadas do que as de M. gypseum e que o
potencial antifúngico dos extratos alcoólicos das própolis verde e vermelha para essas espécies,
sugerem aplicações futuras como tratamento alternativo para dermatofitoses causadas por essas
espécies.
Palavras chaves: Antifúngico; Dermatófito; Própolis verde; Própolis vermelha

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
130
1. Introdução
Os dermatófitos, agentes etiológicos de dermatofitoses, são fungos com propriedades
queratinofílicas e queratinolíticas, capazes de infectar pele, pêlos e unhas. Microsporum
canis, M. gypseum e Epidermophyton floccosum são exemplos de espécies zoofílica, geofílica e
antropofílica, respectivamente. Esses fungos apresentam distribuição geográfica cosmopolita,
entretanto podem ser observadas variações na prevalência da espécie de região para região, tendo
esse fato importância epidemiológica e terapêutica (Weitzman e Summerbell, 1995; Aquino et al.,
2007).
Os casos de infecções causadas por dermatófitos têm aumentado consideravelmente (Porro
et al., 1997) e medicamentos de uso tópico e oral têm sido empiricamente administrados (Roberts
et al., 1994). Entretanto, alguns fungos são significativamente diferentes a susceptibilidade a essas
drogas (Jessup et al., 2000).
As dermatofitoses podem ser resistentes à terapia e a resposta ao tratamento pode não ser
adequada (Favre et al., 2003), por isso, torna-se necessário o desenvolvimento de novas estratégias
terapêuticas (García et al., 2003).
Atenção especial tem se voltado para derivados naturais, com base no conhecimento da
produção de compostos antifúngicos pela natureza (Gurgel et al., 2005). Com o passar do tempo,
produtos naturais como própolis, estão sendo cada vez mais utilizados na medicina (Park et al.,
1998).
Própolis é um material resinoso elaborado por abelhas a partir da coleta de exsudatos
resinosos de diversas partes das plantas como brotos, botões florais, folhas novas e casca
(Burdock, 1998; Park et al., 2004; Bankova, 2005). Mais de 300 substancias já foram identificadas
em diferentes amostras de própolis, tais como flavonóides, ácidos fenólicos, ácidos aromáticos e
ácidos diterpênicos, que são os principais componentes responsáveis pelas atividades biológicas
das própolis (Bankova e Marcucci, 2000; Park et al., 2004).
Própolis possui propriedades antibacteriana, antifúngica, antiviral, antiinflamatória,
antiulcerativa, antitumoral, antioxidante, anestésica local, hepatoprotetora, imunoestimulatória,
cicatrizante, entre outras (Banskota et al., 1998; Park et al., 1998; Suzuki et al., 2002; Koc et al.,
2005).
Alguns autores têm relatado a atividade antifúngica da própolis da Turquia em relação a
Trichophyton rubrum e T. mentagrophytes (Koc et al., 2005). Própolis do Brasil, provenientes de
São Paulo, Paraná e Ceará (Kujumgiev et al., 1999; Ota et al., 2001; Sawaya et al., 2002; Oliveira
et al., 2006) e da Bulgária, Mongólia e Egito (Kujumgiev et al., 1999) apresentaram atividade
antifúngica em relação a espécies de Candida. A susceptibilidade de Paracoccidioides brasiliensis

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
131
para própolis do Brasil (Botucatu) e da Bulgária (Murad et al., 2002) como também de
Cryptococcus neoformans em relação à própolis verde brasileira (Fernandes et al., 2007), já foi
verificada.
A própolis verde do Brasil é rica em derivados prenilados do ácido cinâmico e é elaborada
por abelhas Apis mellifera a partir da coleta da matéria-prima de Baccharis dracunculifolia
(Bankova e Marcucci, 2000). Já foi verificada atividade in vitro em relação a bactérias gram
positivas e gram negativas e in vivo por redução de 95% de casos de gengivites (Amaral et al.,
2006) e inibição de candidíase oral (Santos et al., 2005).
A própolis vermelha brasileira apresenta elevada concentração de flavonóides e é
encontrada em colméias ao longo do mar e costa dos rios, como também em arbustos de
manguezais da região Nordeste do Brasil. O material coletado pelas abelhas Apis mellifera é
proveniente de exsudato vermelho da superfície de Dalbergia ecastophyllum, sendo essa a fonte
botânica dessa própolis. Já foi demonstrada atividade antimicrobiana in vitro da própolis vermelha,
em relação a Staphylococcus aureus (Daugsch et al., 2007).
Este trabalho teve como objetivo avaliar in vitro as atividades fungistática e fungicida do
extrato aquoso da própolis verde e dos extratos alcoólicos da própolis verde e da vermelha, ambas
do Brasil, em relação a amostras de Microsporum canis, M. gypseum e Epidermophyton
floccosum.
2. Material e Métodos
2.1. Amostras fúngicas
Quinze amostras de dermatófitos foram analisadas, sendo cinco de Microsporum canis,
cinco de M. gypseum e cinco de Epidermophyton floccosum. Esses fungos foram obtidos da
Coleção de Culturas Micoteca-University of Recife Mycology (URM) e estavam preservados sob
óleo mineral (Sherf, 1943). A maioria das amostras foi isolada de escamas epidérmicas (Tabela
1). A amostra Trichophyton mentagrophytes (INCQS 40004, ATCC 9533) foi utilizada como
controle.
2.2. Procedimentos para avaliação da atividade antifúngica
O procedimento adotado foi realizado de acordo com a padronização publicada no
documento M38-A (2002) pelo “Clinical and Laboratory Standards Institute” (CLSI, 2002) e
adequado para dermatófitos segundo Fernández-Torres et al. (2002) e Barros et al. (2006).
2.2.1. Meio de cultura
O meio de cultura utilizado para o procedimento foi o RPMI 1640 (Sigma-Aldrich, USA)
com L-glutamina e sem bicarbonato de sódio, tamponado a pH 7.0 com ácido

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
132
morfolinepropanesulfônico (MOPS) a 0.165 M (Sigma-Aldrich, USA). O meio foi esterilizado por
filtração em membrana com porosidade de 0,22µm (Millipore).
2.2.2. Antifúngicos
As drogas utilizadas para efeito comparativo foram itraconazol (Jansen-Cilag, São Paulo,
Brasil) e terbinafina (Novartis, São Paulo, Brasil) as quais foram dissolvidas em dimetilsulfóxido
(DMSO) (Gibco, Minas Gerais, Brasil). Foram utilizadas 10 concentrações entre 0.03 e 16µg/ml.
2.2.3. Própolis
Foram utilizados extratos alcoólico e aquoso da própolis verde e extrato alcoólico da
própolis vermelha, fornecidos secos pela Pharma Néctar®, Belo Horizonte, Brasil. A própolis
verde foi obtida de Minas Gerais e a própolis vermelha da Paraíba, estados do Brasi. Após a
solubilização dos extratos nos referidos solventes, água e álcool etílico a 70% (Sawaya et al.,
2002), foram preparadas concentrações de 2 a 1024µg/ml e de 4 a 2048µg/ml, respectivamente
para os extratos alcoólico e aquoso.
2.2.4. Preparação do inóculo
As amostras de Microsporum e E. floccosum foram semeadas em meio de Batata-Dextrose-
Ágar contido em tubos de ensaio os quais foram mantidos a temperatura ambiente (28+ 2ºC) até a
esporulação de cada amostra. Posterirmente foram acrescentados 5ml de água destilada
esterilizada nos tubos de ensaio e com auxílio de pipeta Pasteur foi promovido desprendimento de
esporos e fragmentos de hifas. Após 20 minutos em repouso, o sobrenadante foi aspirado para
outro tubo de ensaio esterilizado e agitado em vortex por 15 segundos. A densidade das
suspensões foi ajustada em espectrofotômetro a 530nm, para obter entre 65-70% de transmitância.
Posteriormente, as suspensões foram diluídas (1:50) em RPMI 1640 para obter a concentração de 2
vezes maior do que a necessária.
2.2.5. Procedimento do teste
Em placas esterilizadas de microtitulação com tampa (TPP, Suíça), contendo 96 poços com
formato de U (fundo chato) foram depositados 100µl das drogas nas fileiras de 1 a 10, sendo que
cada concentração foi depositada em uma fileira. Nos poços das fileiras 11 e 12 foram depositados
100µl do meio RPMI 1640, os quais foram os controles de crescimento e o de esterilização do
meio, respectivamente. Essas placas foram armazenadas a -20ºC até o momento do uso.
No momento adequado, foi depositado em cada poço 100µl do inóculo padronizado como
citado anteriormente (item 2.2.4) e as placas de microtitulação foram incubadas a temperatura
ambiente (28+ 2ºC) por sete dias para interpretação dos resultados.
A determinação da concentração inibitória mínima dos extratos das própolis e dos
antifúngicos foi realizada por observação visual de cada poço que apresentava redução de

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
133
crescimento fúngico. Levando em consideração o crescimento total (100%) no poço controle, o
percentual de redução de crescimento foi atribuído aos demais poços. A inibição do crescimento
também foi verificada por leitura das placas de microtitulação pelo programa Microplate Manager,
modelo BenchMark Plus (Bio-Rad Laboratories Inc.) a 530nm.
Para itraconazol e terbinafina a concentração inibitória mínima (CIM) foi a que promoveu
inibição de 80% e 100% do crescimento fúngico, respectivamente (Santos e Hamdan, 2005). Para
as própolis a CIM considerada foi a que inibiu 100% do crescimento fúngico.
Para determinação da concentração fungicida mínima, cada conteúdo dos poços que
apresentava redução de 80% e 100% do crescimento fúngico, foi transferido da placa de
microtitulação para tubos de ensaio contendo 5ml de Sabouraud líquido. Os tubos foram mantidos
a temperatura ambiente (28 + 2ºC) por sete dias para verificação de desenvolvimento fúngico. A
concentração fungicida mínima correspondeu a menor concentração fungistática que não permitiu
o crescimento do fungo (Favre et al., 2003).
2. 3. Análise de dados
Todo experimento foi realizado em duplicata. Foram determinadas as concentrações
inibitória e fungicida mínima (CIM e CFM) de cada antifúngico e dos extratos das própolis, as
médias geométricas das concentrações inibitória e fungicida mínima (CIMMG e CFMMG) em
relação a espécie, a determinação da concentração capaz de inibir metade e todas as amostras
analisadas (CIM50 e CIM100, respectivamente).
3. Resultados
Os resultados referentes a atividade antifúngica estão nas Tabelas 2 e 3. A média dos
resultados referentes a inibição do crescimento das amostras de dermatófitos estão nos Gráficos 1
e 2. As amostras de dermatófitos analisadas não foram susceptíveis ao extrato aquoso da própolis
verde.
O extrato alcoólico da própolis verde apresentou atividade fungistática entre 64-1024µg/ml
para M. canis, 256-1024 µg/ml para M. gypseum e 8-512 µg/ml para E. floccosum. A atividade
fungicida do extrato alcoólico da própolis verde foi constatada em três amostras de M. canis
(URM 4225, URM 4154 e URM 4678), cuja CFM=256µg/ml para a primeira e de 1024µg/ml para
as demais. Entre as amostras de M. gypseum, duas (URM 4279 e URM 3645) foram susceptíveis a
atividade fungicida do extrato alcoólico da própolis verde, cuja CFM=1024µg/ml. Todas as
amostras de E. floccosum foram susceptíveis a atividade fungicida do extrato alcoólico da própolis
verde na maior concentração analisada (1024µg/ml).
A atividade fungistática do extrato alcoólico da própolis vermelha foi verificada nas
concentrações de 16-128µg/ml para M. canis, 64-512µg/ml para M. gypseum e 16-256µg/ml para

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
134
E. floccosum. A atividade fungicida foi verificada entre 128-256µg/ml, 128-1024 e 128-512µg/ml,
respectivamente para as mesmas espécies acima citadas.
4. Discussão
A atividade antifúngica de itraconazol e terbinafina verificada neste trabalho tem sido
mencionada de forma similar por outros autores (Favre et al., 2003; Barros et al., 2006).
As concentrações inibitórias que não foram determinadas, como citados na Tabela 2,
demonstram sensibilidade da maioria das amostras analisadas de M. canis, M. gypseum e E.
floccosum para uma mesma concentração de própolis.
A susceptibilidade das espécies de dermatófitos aos extratos alcoólicos das própolis
utilizadas, pode ser explicada pela composição química das própolis. Já foi demonstrado que mais
compostos ativos são extraídos da própolis nativa se o extrato for preparado com álcool etílico e
em concentrações acima de 50%. Quando a água é utilizada como solvente, é citado que a
atividade antifúngica e a antibacteriana, por exemplo, são mais fracas quando comparada com o
extrato alcoólico (Sawaya et al., 2002).
O extrato alcoólico da própolis verde brasileira é rico em ácidos fenólicos e flavonóides,
havendo predomínio de ácidos fenólicos dos quais o ácido cinâmico é o mais abundante,
principalmente os que contem o grupo prenil, como por exemplo, o artepelin C (Park et al., 2002;
Sousa et al., 2007). Além desses, também são citados terpenos, ácidos aromáticos e flavonóides
tais como canferide, pinobanskina, pinocembrina, crisina, galangina, canferol e isosakuranetina,
entre outros (Park et al., 2002; Marcucci, 2006).
A maior susceptibilidade ao extrato alcoólico da própolis vermelha, verificada neste
trabalho, também está relacionada com a composição química dessa própolis, pois há predomínio
de flavonóides em vez de ácidos fenólicos. Entre os flavonóides, podem ser citados:
liquiritigenina, daidzeína, dalbergina, isoliquiritigenina, formononetina e biochanina A, rutina,
pinobanskina, quercetina, luteolina, pinocembrina, entre outros (Trusheva et al., 2007).
De um modo geral, a atividade antifúngica da própolis é atribuída ao ácido cinâmico, ácido
felúrico, ácido caféico, terpenos e principalmente aos flavonóides, tais como crisina, ermanina,
galangina, canferol, pinobanskina e pinocembrina, sendo a atividade fungicida atribuída
principalmente a esse último (Burdok et al., 1998).
A atividade antifúngica da própolis verde constatada neste trabalho já havia sido verificada
em relação a Candida albicans (Santos et al., 2005) e a Cryptoccoccus neoformans (Fernandes et
al., 2007). Para o extrato alcoólico da própolis vermelha já foi verificada atividade antibacteriana,
em relação a Staphylococcus aureus (Daugsch et al. 2007).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
135
As elevadas concentrações inibitórias mínimas obtidas neste trabalho e a não obtenção da
média geométrica da concentração fungicida do extrato alcoólico da própolis verde para M. canis e
M. gypseum, refletem a composição química da própolis do Brasil, que é diferente da própolis
européia, onde predomina flavonóides (Bankova et al., 2000). Koc et al. (2005) verificaram
susceptibilidade de dermatófitos em relação à própolis da Turquia em baixas concentrações
(0.025-0.8µg/ml). Por outro lado, amostras de Candida sp. avaliadas por outros autores foram
susceptíveis ao extrato alcoólico da própolis verde de São Paulo nas concentrações entre 3-
12mg/ml (Ota et al., 2001) e 6-12mg/ml (Sawaya et al., 2002). Entretanto, é citado que quanto
maior é a concentração de flavonóides no extrato de própolis, maior é a atividade antifúngica (Koc
et al., 2005).
A menor sensibilidade de M. gypseum aos antifúngicos e ao extrato alcoólico das própolis
utilizadas, pode está associada com a capacidade de esporulação e a quantidade de esporos
produzidos. De acordo com Fernández-Torres et al. (2003) a parede celular do esporo do
dermatófito é mais espessa do que da hifa, havendo necessidade de maior concentração do
antifúngico para inibição do crescimento. Devido a facilidade da produção de esporos por
amostras dessa espécie, esse fato pode ter ocorrido.
Soares e Cury (2001) sugerem a associação de antifúngicos com própolis como uma
excelente alternativa para o tratamento de dermatofitoses causadas por T. rubrum, T.
mentagrophytes e E. floccosum. A susceptibilidade das amostras de Microsporum e E. floccosum,
utilizadas neste trabalho, a atividade antifúngica do extrato alcoólico da própolis verde e da
vermelha, pode reforçar essa possibilidade.
5. Conclusões
Os dados obtidos demonstram que as amostras de M. canis e E. floccosum foram mais
sensíveis a atividade antifúngica do extrato alcoólico das própolis verde e da vermelha do que as
de M. gypseum. O potencial antifúngico da própolis vermelha para essas espécies sugerem
aplicações futuras como tratamento alternativo para dermatofitoses causadas por M. canis, M.
gypseum ou E. floccosum.
Agradecimentos
Agradecemos a concessão de bolsa pela Coordenação de Aperfeiçoamento de Pessoal de
Nível Superior (CAPES) como também pelo suporte financeiro e infra-estrutura da Universidade
Federal de Pernambuco (UFPE), Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq) e Rede de Coleções de Culturas de Microrganismos do Norte e Nordeste do Brasil
(RENNEBRA).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
136
Referências
Amaral, R.C., Gomes, R.T., Rocha, W.M.S., Abreu, S.L.R., Santos, V.R., 2006. Periodontitis
treatment with brazilian green propolis gel. Pharmacology online 3: 336-341.
Aquino, V.R., Constante, C.C., Bakos, L., 2007. Frequency of dermatophytosis in mycological
examinations at a general hospital in Porto Alegre, Brazil. Anais Brasileiros de Dermatologia.
82,239-244.
Bankova, V., Marcucci, M.C., 2000. Standardization of propolis: present status and perspectives.
Bee World 81, 182–8.
Bankova, V.S., Castro, S.L., Marcucci, M.C., 2000. Propolis: recent advances in chemistry and
plant. Apidologie 31, 3-15.
Banskota, A.H., Tezuka, Y., Prasain, J.K., Matsushige, K., Saiki, I., Kadota, S., 1998. Chemical
constituents of Brazilian propolis and their cytotoxic activities. Journal of Natural Products 61,
896–900.
Barros, M.E.S., Santos, D.A., Hamdan, J.S., 2006. “In vitro” methods for antifungal susceptibility
testing of Trichophyton spp. Mycological Research 110, 1355-1360.
Burdock, G.A., 1998. Review of the biological properties and toxicity of bee propolis. Food and
Chemical Toxicology 36, 347- 363.
Clinical and Laboratory Standards Institute - CLSI (2002) Reference Method for Broth Dilution
Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard document M38-A.
National Committee for Clinical Laboratory Standards, Wayne, PA, USA
Daugsch, A., Moraes, C.S., Fort, P. and Park, Y.K., 2007. Brazilian Red Propolis - Chemical
Composition and Botanical Origin. Evidence-based Complementary and Alternative Medicine,
doi:10.1093/ecam/nem057.
Favre, B., Hofbauer, B., Hildering, K.A., Ryder, N.S., 2003. Comparison of in vitro activities of
17 antifungal drugs against a panel of 20 dermatophytes by using a microdilution assay. Journal of
Clinical Microbiology 41, 4817- 4819.
Fernandes, F.F., Dias, A.L.T., Ramos, C.L., Ikegaki, M., Siqueira, A,M, Franco, M.C., 2007. The
“in vitro” antifungal activity evaluation of propolis G12 ethanol extract on Cryptococcus
neoformans. Revista do Instituto de Medicina Tropical de São Paulo 49: 93-95.
Fernández-Torres, B., Cabanes, F.J., Carrillo-Muñoz, A.J., Esteban, A., Inza, I., Abarca, L.,
Guarro, J., 2002. Collaborative evaluation of optimal antifungal susceptibility testing conditions
for dermatophytes. Journal of Clinical Microbiology 40, 3999-4003.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
137
Fernández-Torres, B., Inza, I., Guarro, J., 2003. “In vitro” activities of the new antifungal drug
eberconazole and three other topical agents against 200 strains of dermatophytes. Journal of
Clinical Microbiology 41, 5209-5211.
García, V.M.N., Gonzalez, A., Fuentes, M., Aviles, M., Rios, M.Y., Zepada, G. and Rojas M.G.,
(2003). Antifungal activities of nine traditional Mexican medicinal plants. Journal of
Ethnopharmacology 87, 85–88.
Gurgel, L.A., Sidrim, J.J.C., Martins, D.T., Cechinel Filho, V., Rao, V.S., 2005. In vitro antifungal
activity of dragon’s blood from Croton urucurana against dermatophytes. Journal of
Ethnopharmacology 97, 409-412.
Jessup, C.J., Warner, J., Isham, N., Hasan, I., Ghannoum, M.A., 2000. Antifungal susceptibility
testing of dermatophytes: establishing a medium for inducing conidial growth and evaluation of
susceptibility of clinical isolates. Journal of Clinical Microbiology 38, 341– 4.
Koc, A.N., Silici, S., Ayangil, D., Ferahbas, A., Çankaya, S., 2005. Comparison of in vitro
activities of antifungal drugs and ethanolic extract of propolis against Trichophyton rubrum and T.
mentagrophytes by using a microdilution assay. Mycoses 48, 205-210.
Kujumgiev, A., Tsvetkova, I., Serkedjieva, Y., Bankova, V., Christov and R. Popov, S., 1999.
Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. Journal of
Ethnopharmacology 64, 235-240.
Marcucci, M.C. 2006. Própolis tipificada: um novo caminho para a elaboração de medicamentos
de origem natural, contendo este produto apícola. Revista Fitos 1, 36-46.
Murad, M., Calvi, S.A., Soares, A.M.V.C., Bankova, V., Sforcin, J.M., 2002. Effects of propolis
from Brazil and Bulgaria on fungicidal activity of macrophages against Paracoccidioides
brasiliensis. Journal of Ethnopharmacology 79, 331-334.
Oliveira, A.C.P., Shinobu, C.S., Onghini, R.L., Franco, S.L., Svidzinski, T.I.E., 2006. Antifungal
activity of propolis extract against yeasts isolated from onychomycosis lesions. Memórias do
Instituto Oswaldo Cruz 101, 493-497.
Ota, C., Unterkircher, C., Fantinato, V., Shimizu, M.T., 2001. Antifungal activity of propolis on
different species of Candida. Mycoses 44, 375-378
Park, Y.K., Alencar, S.M., Aguiar, C.L., 2002. Botanical Origin and Chemical Composition of
Brazilian Propolis. Journal of Agricultural and Food Chemistry 50, 2502-2506.
Park, Y.K., Koo, M.H., Abreu, J.A.S., Ikegaki, M., Cury, J.A., Rosalen, P.L., 1998. Antimicrobial
activity of propolis on oral microorganisms. Current Microbiology 36, 24-28.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
138
Park, Y.K., Paredes-Guzman, J.F., Aguiar, C.L., Alencar, S.M., Fujiwara, F.Y., 2004. Chemical
constituents in Baccharis dracunculifolia as the main botanical origin of Southeastern Brazilian
propolis. Journal of Agriculture and Food Chemistry 52, 1100-1103.
Popova, M., Silici, S., Kaftnoglu, O., Bankova, V., 2005. Antibacterial activity of Turkish propolis
and its qualitative and quantitative chemical composition. Phytomedicine 12: 221- 228.
Porro, A.M., Yoshioka, M.C.N., Kaminski, S.K., Palmeira, M.C.A, Fischman, O., Alchorne,
M.M.A., 1997. Disseminated dermatophytosis caused by Microsporum gypseum in two patients
with the acquired immunodeficiency syndrome. Mycopathologia 137, 9–12.
Santos, D.A., Hamdan, J.S., 2005. Evaluation of broth microdilution antifungal susceptibility
testing conditions for Trichophyton rubrum. Journal of Clinical Microbiology 43, 1917-1920.
Santos, V.R., Pimenta, F. J. G. S., Aguiar, M.C.F., Do Carmo, M.A.V., Naves, M.D. and
Mesquita, R.A. 2005. Oral candidiasis treatment with brazilian ethanol propolis extract.
Phytotherapy Research 19, 652–654.
Sawaya, A.C.H.F., Palma, A.M., Caetano, F.M., Marcucci, M.C., Da Silva Cunha, I.B., Araújo,
C.E.P., Shimizu, M.T., 2002. Comparative study of in vitro methods used to analyse the activity of
propolis extracts with different compositions against species of Candida. Letters in Applied
Microbiology 35, 203–207.
Sherf, A.F., 1943. A method for maintaining Phytomonas sepedomica for long periods without
transfer. Phytopathology 3, 30-32.
Soares, M.M.R.S., Cury, A.E., 2001. “In vitro” activity of antifungal and antiseptic agents against
dermatophyte isolates from patients with tinea pedis. Brazilian Journal of Microbiology 32, 130-
134.
Sousa, J.P.B., Furtado, N.A.J.C., Jorge, R., Soares, A.E.E., Bastos, J.K., 2007. Perfis físico-
químico e cromatográfico de amostras de própolis produzidas nas microrregiões de Franca (SP) e
Passos (MG), Brasil. Revista Brasileira de Farmacognosia 17, 85-93.
Suzuki, I., Hayashi, I., Takaki, T., Groveman, D.S., Fujimiya, Y., 2002. Antitumor and
anticytopenic effects of aqueous extracts of propolis in combination with chemotherapeutics
agents. Cancer Biotherapy and Radiopharmaceuticals 17, 553-562
Trusheva, B., Popova, M., Bankova, V., Simova, S., Marcucci, M.C., Miorin, P.L., Pasin, F.R.,
Tsvetkova, I., 2006. Bioactive Constituents of Brazilian Red Propolis. Evidence-Based
Complementary and Alternative Medicine 3, 249-254.
Weitzman I, Summerbell RC., 1995. The Dermatophytes. Clinical Microbiology Reviews 8, 240–
259.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
139
Espécie Nº registro Ano de estoque Período de estoque Substrato
(URM) (ano)
Microsporum canis
M. canis
M. canis
M. canis
M. canis
M. gypseum
M. gypseum
M. gypseum
M. gypseum
M. gypseum
Epidermophyton floccosum
E. floccosum
E. floccosum
E. floccosum
E. floccosum
4962
4225
4154
4678
4158
4964
3060
6034
3645
4279
3345
4151
4799
4798
3195
2005
1999
1999
2003
1998
2005
1989
2005
1995
2000
1993
1999
2004
2004
1990
2
8
8
4
9
2
18
2
12
7
14
8
3
3
17
EE
EE
EE
EE
EE
EE
DE
EE
EE
PR
EE
EE
EE
EE
EE
EE: escamas epidérmicas; DE: desconhecido; PR: praias de Recife
Tabela 1. Amostras de Microsporum canis, M. gypseum e Epidermophyton floccosum
preservadas sob óleo mineral e estocados entre 2 e 18 anos, na Coleção de Culturas
Micoteca - University of Recife Mycology (URM).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
140
Tabela 2. Determinação da concentração mínima inibitória de itraconazol, terbinafina e extratos
alcoólicos das própolis verde e vermelha do Brasil, em relação a amostras de Microsporum canis, M.
gypseum e Epidermophyton floccosum preservadas na Coleção de Culturas Micoteca - University of
Recife Mycology (URM).
* Média dos resultados
CIMs: concentrações inibitórias mínimas
CIM50 e CIM100: concentração mínima capaz de inibir 50% e 100% das amostras de Microsporum, canis, M.
gypseum e Epidermophyton floccosum, respectivamente
CIMMG: média geométrica da concentração inibitória mínima de todas as amostras de cada espécie
ND : não determinado
Espécie de dermatófito
(número de amostras testadas)
Microsporum canis (5)
M. gypseum (5)
Epidermophyton floccosum (5)
Antifúngico
(inibição do crescimento)
Itraconazol (80%)
Terbinafina (100%)
Própolis verde (100%)
Própolis vermelha (100%)
Itraconazol (80%)
Terbinafina (100%)
Própolis verde (100%)
Própolis vermelha (100%)
Itraconazol (80%)
Terbinafina (100%)
Própolis verde (100%)
Própolis vermelha (100%)
*CIMs
(µg/mL)
0.125 – 1
0.06 – 0.125
256 – 1024
64 – 128
0.25 – 16
0.25 – 16
1024
128 – 512
0.03
0.03 – 0.125
256 – 512
32 – 256
*CIM50
(µg/mL)
0.125
0.06
256
ND
0.5
0.25
ND
128
ND
0.03
ND
128
*CIM100
(µg/mL)
1
0.125
1024
128
16
16
1024
512
ND
0.06
ND
ND
*CIMMG
(µg/mL)
0.21
0.03
588.13
111.43
1.74
0.57
1024
222.86
0.03
0.04
445.72
147.03

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
141
* Média dos resultados
CFMs: concentrações fungicidas mínima
CFM50 e CFM100: concentração mínima capaz de inibir 50% e 100% das amostras de Microsporum canis, M.
gypseum e E. floccosum, respectivamente
CFMMG: média geométrica da concentração fungicida mínima de todas as amostras de cada espécie
ND : não determinado
Espécie de dermatófito
(número de amostras testadas)
Microsporum canis (5)
M. gypseum (5)
Epidermophyton floccosum (5)
Antifúngico
(inibição de crescimento)
Itraconazol
Terbinafina
Própolis verde
Própolis vermelha
Itraconazol
Terbinafina
Própolis verde
Própolis vermelha
Itraconazol
Terbinafina
Própolis verde
Própolis vermelha
*CFMs
(µg/mL)
0.25 – 2
0.125 – 0.5
256 – 1024
128 – 256
0.5 – 16
0.03 – 16
1024
128 - 1024
0.03 – 0.25
0.03 – 0.25
1024
128 – 512
*CFM50
(µg/mL)
0.25
0.125
256
ND
1
0.25
1024
256
0.06
0.125
ND
256
*CFM100
(µg/mL)
2
0.5
ND
256
16
16
ND
1024
0.125
ND
ND
ND
*CFMMG
(µg/mL)
0.66
0.19
ND
147.03
2
0.43
ND
337.79
0.07
0.16
1024
294.06
Tabela 3. Determinação da concentração mínima fungicida de itraconazol, terbinafina e extratos
alcoólicos das própolis verde e vermelha do Brasil, em relação a amostras de Microsporum canis e
M. gypseum e Epidermophyton floccosum preservadas na Coleção de Culturas Micoteca - University
of Recife Mycology (URM).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
142
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0 2 4 8 16 32 64 128 256 512 1024
Microsporum canisM. gypseumEpidermophyton floccosum
Concentração (µg/mL) do extrato alcoólico da própolis verde
Gráfico 1. Média da inibição do crescimento de amostras de Microsporum canis, M. gypseum e
Epidermophyton floccosum preservadas na Coleção de Culturas Micoteca - University of Recife
Mycology (URM), em diferentes concentrações do extrato alcoólico da própolis verde.
D.O
. 530
nm
Concentração (µg/mL) do extrato alcoólico da própolis vermelha
Gráfico 2. Média da inibição do crescimento de amostras de Microsporum canis, M. gypseum e
Epidermophyton floccosum preservadas na Coleção de Culturas Micoteca - University of Recife
Mycology (URM), em diferentes concentrações do extrato alcoólico da própolis vermelha.
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0 2 4 8 16 32 64 128 256 512 1024
Microsporum canisM. gypseumEpidermophyton floccosumD
.O. 5
30nm

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
143
SUSCEPTIBILIDADE DE ESPÉCIES DE Trichophyton A PRÓPOLIS VERDE E A
VERMELHA DO BRASIL
Artigo a ser submetido para:
LETTERS IN APPLIED MICROBIOLOGY
Qualis A
Fator de impacto 1,593 (Journal of Citation Report - 2006)

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
144
SUSCEPTIBILIDADE DE ESPÉCIES DE Trichophyton A PRÓPOLIS VERDE E A
VERMELHA DO BRASIL
Ana Beatriz Sotero Siqueira*1,2, Bruno Severo Gomes1,2, Gilmara Araújo1, Sheila Abreu3, Rita
Maia4, Cristina Souza-Motta1, Lusinete Aciole de Queiroz1, Ana Lucia F. Porto2,5
1Departmento de Micologia, Universidade Federal de Pernambuco (UFPE), Recife-Brasil 2 Laboratório de Imunopatologia Keizo Asami (LIKA), UFPE, Recife-Brasil 3 Pharma Néctar®, Belo Horizonte-Brasil 4 Centro de Pesquisa Aggeu Magalhães – CPqAM / FIOCRUZ, Recife-Brazil 5 Departamento de Morfologia e Fisiologia Animal, Universidade Federal Rural de Pernambuco
(UFRPE), Recife – PE, Brasil
*Email para correspondência: [email protected]

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
145
RESUMO
Objetivos: A atividade antifúngica de extratos da própolis verde e da vermelha do Brasil foi
constatada in vitro em relação a diferentes espécies de Trichophyton.
Material e Resultados: Foi analisada atividade antifúngica in vitro dos extratos aquoso e alcoólico
da própolis verde e do extrato alcoólico da própolis vermelha do Brasil, em relação a amostras de
Trichophyton rubrum, T. tonsurans e T. mentagrohytes, sendo utilizado como controle itraconazol
e terbinafina. A concentração inibitória mínima foi determinada de acordo com o método de
microdiluição adotado pelo “Clinical and Laboratory Standards Institute”. A concentração
fungicida mínima foi determinada pela ausência de crescimento em meio de cultura Sabouraud
líquido. Os dados obtidos demonstraram que a atividade antifúngica do extrato alcoólico da
própolis verde foi verificada nas concentração de 64 a 1024µg ml-1, enquanto que para o extrato
alcoólico da própolis vermelha foi de 8 a 1024µg ml-1.
Conclusão: A atividade antifúngica do extrato alcoólico da própolis vermelha foi mais eficiente do
que o da verde em todas as amostras de Trichophyton analisadas.
Significância e Impacto do estudo: O potencial antifúngico dos extratos alcoólicos das própolis
verde e da vermelha para essas espécies, sugere aplicações futuras como tratamento alternativo
para dermatofitoses causadas por T. rubrum, T. tonsurans e T. mentagrophytes.
Palavras chaves: antifúngico, dermatófito, própolis verde brasileira, própolis vermelha brasileira

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
146
INTRODUÇÃO
Os dermatófitos são fungos filamentosos pertencentes aos gêneros Trichophyton,
Microsporum e Epidermophyton capazes de infectar pele, pêlos e unhas. Os casos de infecções
causadas por esses fungos têm aumentado consideravelmente sendo que os agentes etiológicos
mais comuns desse tipo de micose superficial são as espécies pertencentes ao gênero Trichophyton
(Weitzman e Summerbell 1995; Lacaz et al., 2002).
Além dos fungos apresentarem diferentes susceptibilidades às drogas, a resposta ao
tratamento pode não ser eficaz devido a resistência ao medicamento (Favre et al. 2004).
No desenvolvimento de novas estratégias terapêuticas para tratamento de micoses, aquelas
direcionadas para os derivados naturais vêm se destacando principalmente pela capacidade de
produção de compostos antifúngicos pela natureza (Gurgel et al. 2005; Fernandes Júnior et al.
2006) e entre essas, a própolis está sendo cada vez mais utilizada na medicina (Kujumgiev et al.
1999; Santos et al. 2005).
Própolis é um material resinoso elaborado por abelhas a partir da coleta de exsudato
resinoso de diversas partes das plantas como brotos, botões florais, folhas novas e casca (Castaldo
e Capasso 2002; Park, et al. 2004). Já foram identificadas mais de 300 substancias em diferentes
amostras de própolis, tais como flavonóides, ácidos fenólicos, ácidos aromáticos e ácidos
diterpênicos que são os principais componentes responsáveis pelas atividades biológicas da
própolis (Bankova 2005).
A própolis possui propriedades antibacteriana, antifúngica, antiviral, antiinflamatória,
antiulcerativa, antitumoral, antioxidante, anestésica local, hepatoprotetora, imunoestimulatória,
cicatrizante, entre outras (Park et al. 1998; Castaldo e Capasso 2002; Suzuki et al. 2002; Fernandes
Júnior et al. 2006).
A atividade antifúngica da própolis de várias regiões do mundo tem sido recentemente
mencionada. Já foi constatada a atividade antifúngica da própolis da Turquia em relação aos
dermatófitos Trichophyton rubrum e T. mentagrophytes (Koc et al. 2005). A própolis do Brasil,
como as provenientes de São Paulo, Paraná e Ceará, e de outras regiões do mundo como as da
Bulgária, Mongólia e Egito apresentaram atividade antifúngica em relação a espécies de Candida
(Kujumgiev et al. 1999; Ota et al. 2001; Sawaya et al. 2002; Oliveira et al. 2006). Já foi constatada
a susceptibilidade de Paracoccidioides brasiliensis para própolis de Botucatu (Brasil) e da
Bulgária (Murad et al. 2002) como também de Cryptococcus neoformans em relação a própolis
verde brasileira (Fernandes et al. 2007).
A própolis verde do Brasil é nativa da região central do país, é elaborada por abelhas Apis
mellifera a partir da coleta de exsudatos de Baccharis dracunculifolia e é rica em derivados

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
147
prenilados do ácido cinâmico (Park et al. 2004). Dessa própolis já foi verificada atividade in vitro
em relação a bactérias gram positivas e gram negativas e in vivo por redução de 95% de casos de
gengivites por bactérias (Amaral et al. 2006) além de inibição de candidíase oral (Santos et al.
2005).
A própolis vermelha brasileira é encontrada em colméias ao longo do mar e da costa dos
rios, como também em arbustos de manguezais da região Nordeste do Brasil e contém maior
concentração de flavonóides do que a própolis verde. O material coletado por Apis mellifera é
proveniente de exsudato vermelho da superfície de Dalbergia ecastophyllum, sendo essa a fonte
botânica dessa própolis (Trusheva et al. 2006; Daugsch et al. 2007). Já foi demonstrada atividade
antimicrobiana in vitro dessa própolis em relação a Staphylococcus aureus (Daugsch et al. 2007).
Este trabalho teve como objetivo avaliar in vitro a atividade antifúngica do extrato aquoso
da própolis verde e dos extratos alcoólicos das própolis verde e vermelha do Brasil, em relação a
amostras de Trichophyton mentagrophytes, T. tonsurans e T. rubrum.
MATERIAL E MÉTODOS
Amostras fúngicas
Quinze amostras de dermatófitos foram analisadas, sendo cinco de Trichophyton rubrum,
cinco de T. tonsurans e cinco de T. mentagrophytes. Esses fungos foram obtidos da Coleção de
Culturas Micoteca-University of Recife Mycology (URM) e estavam preservados sob óleo mineral
(Sherf 1943). A maioria das amostras foi isolada de escamas epidérmicas (Tabela 1). A amostra
Trichophyton mentagrophytes (INCQS 40004, ATCC 9533) foi utilizada como controle.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
148
Procedimentos para avaliação da atividade antifúngica
Foi utilizado o método descrito no documento M38-A (2002) pelo “Clinical and
Laboratory Standards Institute” (CLSI, 2002) e adequado para dermatófitos segundo Fernández-
Torres et al. (2002) e Barros et al. (2006).
EE: escamas epidérmicas; DE: desconhecido; PR: praias do Recife; SO: solo; EU:
escamas ungueais
Tabela 1. Espécies de dermatófitos preservados sob óleo mineral na Coleção
de Culturas Micoteca - University of Recife Mycology (URM).
Espécie No. registro Ano de Período Substrato (URM) estoque de estoque (ano)
Trichophyton rubrum
T. rubrum
T. rubrum
T. rubrum
T. rubrum
T. tonsurans
T. tonsurans
T. tonsurans
T. tonsurans
T. tonsurans
T. mentagrophytes
T. mentagrophytes
T. mentagrophytes
T. mentagrophytes
T. mentagrophytes
4725
4724
4727
4728
4569
4965
4963
4968
4945
4946
4156
3239
2785
4422
2875
2003
2003
2003
2003
2004
2005
2005
2005
2004
2004
1999
1992
1985
2002
1986
4
4
4
4
3
2
2
2
3
3
8
15
22
5
21
EE
EE
EE
EE
EU
EE
EE
EE
EE
EE
PR
EE
SO
EE
DE

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
149
Meio de cultura
O meio de cultura utilizado para este procedimento foi o RPMI 1640 (Sigma-Aldrich,
USA) com L-glutamina e sem bicarbonato de sódio, tamponado a pH 7.0 com ácido
morfolinepropanesulfônico (MOPS) a 0.165 M (Sigma-Aldrich, USA). O meio foi esterilizado por
filtração em membrana com porosidade de 0,22µm (Millipore).
Antifúngicos
Foram preparadas concentrações entre 0.03 e 16µg ml-1 de itraconazol (Jansen-Cilag,
Brasil) e terbinafina (Novartis, Brasil) após solubilização em dimetilsulfóxido (DMSO) (Gibco,
Minas Gerais, Brasil), as quais foram utilizadas para efeito comparativo.
Própolis
Foram utilizados extratos alcoólico e aquoso da própolis verde e extrato alcoólico da
própolis vermelha, fornecidos liofilizados pela Pharma Néctar®, Belo Horizonte, Brasil. A
própolis verde é obtida de Minas Gerais e a própolis vermelha da Paraíba, estados do Brasil. Após
a solubilização dos extratos nos referidos solventes, água e álcool etílico a 70% (v/v) (Sawaya et
al. 2002), foram preparadas concentrações entre 0.03 a 1024µg ml-1 e 0.03 a 2048µg ml-1,
respectivamente para os extratos alcoólico e aquoso.
Preparação do inóculo
As amostras de Trichophyton foram semeadas em meio Batata-Dextrose-Ágar contido em
tubos de ensaio os quais foram mantidos a temperatura ambiente (28+ 2ºC) até a esporulação de
cada amostra. Posteriormente foram acrescentados 5ml de água destilada esterilizada nos tubos de
ensaio e com auxílio de pipeta Pasteur foi promovido desprendimento de esporos e fragmentos de
hifas. Após 20 minutos em repouso, o sobrenadante foi aspirado para outro tubo de ensaio
esterilizado e agitado em vortex por 15 segundos. A densidade das suspensões foi ajustada em
espectrofotômetro a 530nm, para obter entre 65-70% de transmitância. Posteriormente, as
suspensões foram diluídas (1:50) em RPMI 1640 para obter a concentração de 2 vezes maior do
que a necessária.
Procedimento do teste
Em placas esterilizadas de microtitulação com tampa (TPP, Suíça), contendo 96 poços com
formato de U (fundo chato) foram depositados 100µl das drogas nas fileiras de 1 a 10, sendo que
cada concentração foi depositada em uma fileira. Nos poços das fileiras 11 e 12 foram depositados
100µl do meio RPMI 1640, os quais foram os controles de crescimento e o de esterilização do
meio, respectivamente. Essas placas foram armazenadas a -20ºC até o momento do uso.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
150
No momento adequado, foi depositado em cada poço 100µl do inóculo padronizado como
citado anteriormente (item 2.2.4) e as placas de microtitulação foram incubadas a temperatura
ambiente (28+ 2ºC) por sete dias para interpretação dos resultados.
A determinação da concentração inibitória mínima dos extratos das própolis e dos
antifúngicos foi realizada por observação visual de cada poço que apresentava redução de
crescimento fúngico. Levando em consideração o crescimento total (100%) no poço controle, o
percentual de redução de crescimento foi atribuído aos demais poços. A inibição do crescimento
também foi verificada por leitura das placas de microtitulação pelo programa Microplate Manager,
modelo BenchMark Plus (Bio-Rad Laboratories Inc.) a 530nm.
Para itraconazol e terbinafina a concentração inibitória mínima (CIM) foi a que promoveu
inibição de 80% e 100% do crescimento fúngico, respectivamente (Santos e Hamdan, 2005). Para
as própolis a CIM considerada foi a que inibiu 100% do crescimento fúngico.
Para determinação da concentração fungicida mínima, cada conteúdo dos poços que
apresentava redução de 80% e 100% do crescimento fúngico, foi transferido da placa de
microtitulação para tubos de ensaio contendo 5ml de Sabouraud líquido. Os tubos foram mantidos
a temperatura ambiente (28 + 2ºC) por sete dias para verificação de desenvolvimento fúngico. A
concentração fungicida mínima correspondeu a menor concentração fungistática que não permitiu
o crescimento do fungo (Favre et al., 2003).
Análise de dados
Todo experimento foi realizado em duplicata. Foram determinadas as concentrações
inibitória e fungicida mínima (CIM e CFM) de cada antifúngico e própolis, as médias geométricas
das concentrações inibitória e fungicida mínima (CIMMG e CFMMG) em relação a cada espécie,
a determinação da concentração capaz de inibir metade e todas as amostras analisadas (CIM50 e
CIM100, respectivamente).
RESULTADOS
Estão nas Tabelas 2 e 3 os resultados referentes a atividade antifúngica determinados por
observação visual. Está nos Gráfico 1 e 2 a média dos resultados referentes à inibição do
crescimento das amostras de dermatófitos. As amostras de dermatófitos analisadas não foram
susceptíveis ao extrato aquoso da própolis verde.
O extrato alcoólico da própolis verde apresentou atividade fungistática entre 64-512 µg ml-
1 para T. rubrum e entre 128-1024 µg ml-1 para T. tonsurans e T. mentagrophytes. A atividade
fungicida do extrato alcoólico da própolis verde apresentou CFM=1024 µg ml-1 para T. rubrum e
T. tonsurans e CFM=512 µg ml-1 para T. mentagrophytes.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
151
A atividade fungistática do extrato alcoólico da própolis vermelha foi verificada nas
concentrações de 8-128 µg ml-1 para T. rubrum, 32-128 µg ml-1 para T. ronsurans e 16-128 µg ml-
1 para T. mentagrophytes. A atividade fungicida foi verificada entre 128-256 µg ml-1, 128-1024 e
256-512 µg ml-1, respectivamente para as mesmas espécies acima citadas.
* Média dos resultados
CIMs: concentrações inibitórias mínimas
CIM50 e CIM100: concentração mínima capaz de inibir 50% e 100% das amostras de Trichophyton
rubrum, T. tonsurans e T. mentagrophytes, respectivamente
CIMMG: média geométrica da concentração inibitória mínima de todas as amostras de cada espécie
ND : não determinado
Espécie de dermatófito
(No. de amostras testadas)
Trichophyton rubrum (5)
T. tonsurans (5)
T. mentagrophytes (5)
Antifúngico
(inibição do crescimento)
Itraconazol (80%)
Terbinafina (100%)
Própolis verde (100%)
Própolis vermelha (100%)
Itraconazol (80%)
Terbinafina (100%)
Própolis verde (100%)
Própolis vermelha (100%)
Itraconazol (80%)
Terbinafina (100%)
Própolis verde (100%)
Própolis vermelha (100%)
*CIMs
(µg ml-1)
0.03
0.03
256 – 512
64 – 128
0.03 - 0.125
0.03
512 – 1024
128
0.03 – 0.125
0.06 – 0.125
512 – 1024
128
*CIM50
(µg ml-1)
ND
ND
ND
64
ND
ND
512
ND
0.06
0.125
512
ND
*CIM100
(µg ml-1)
0.03
0.03
512
128
0.125
0.03
1024
128
0.125
1
1024
128
*CIMMG
(µg ml-1)
0.03
0.03
445.72
97
0.03
0.03
776.04
128
0.08
0.16
776.04
128
Tabela 2. Determinação da concentração inibitória mínima de itraconazol, terbinafina e extratos
alcoólicos das própolis verde e vermelha do Brasil, em relação a amostras de Trichophyton rubrum,
T. tonsurans e T. mentagrophytes preservadas na Coleção de Culturas Micoteca - University of
Recife Mycology (URM).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
152
Espécie de dermatófito
(No. de amostras testadas)
Trichophyton rubrum (5)
T. tonsurans (5)
T. mentagrophytes (5)
Antifúngico
(inibição de crescimento)
Itraconazol
Terbinafina
Própolis verde
Própolis vermelha
Itraconazol
Terbinafina
Própolis verde
Própolis vermelha
Itraconazol
Terbinafina
Própolis verde
Própolis vermelha
*CFMs
(µg ml-1)
0.25 – 0.5
0.03 – 0.125
1024
128 – 256
0.25 – 1
0.03 – 0.25
1024
128 - 1024
0.25 – 2
0.06 – 2
512 – 1024
256 – 512
*CFM50
(µg ml-1)
0.25
0.06
ND
128
0.5
0.03
ND
512
0.25
0.125
ND
256
*CFM100
(µg ml-1)
0.5
0.125
1024
256
0.25
0.25
1024
1024
2
0.25
1024
512
*CFMMG
(µg ml-1)
0.38
0.07
1024
194.01
0.66
0.06
1024
512
0.5
0.28
891.44
337.79
* Média dos resultados
CFMs: concentrações fungicidas mínima
CFM50 e CFM100: concentração mínima capaz de inibir 50% e 100% das amostras de Trichophyton
rubrum, T. tonsurans e T. mentagrophytes, respectivamente
CFMMG: média geométrica da concentração fungicida mínima de todas as amostras de cada espécie
ND : não determinado
Tabela 3. Determinação da concentração mínima fungicida de itraconazol, terbinafina e extratos
alcoólicos das própolis verde e vermelha do Brasil, em relação a amostras de Trichophyton rubrum,
T. tonsurans e T. mentagrophytes preservadas na Coleção de Culturas Micoteca - University of
Recife Mycology (URM).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
153
Concentração (µg ml-1) do extrato alcoólico da própolis vermelha
Gráfico 2. Média da inibição do crescimento de amostras de Trichophyton rubrum, T.
tonsurans e T. mentagrophytes preservadas na Coleção de Culturas Micoteca – University
of Recife Mycology (URM), em diferentes concentrações do extrato alcoólico da própolis
vermelha.
D.O
. 530
nm
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0 2 4 8 16 32 64 128 256 512 1024
Trichophyton rubrumT. tonsuransT. mentagrophytes
Concentração (µg ml-1) do extrato alcoólico da própolis verde
Gráfico 1. Média da inibição do crescimento de amostras de Trichophyton rubrum, T.
tonsurans e T. mentagrophytes preservadas na Coleção de Culturas Micoteca – University
of Recife Mycology (URM), em diferentes concentrações do extrato alcoólico da própolis
verde.
D.O
. 530
nm
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0 2 4 8 16 32 64 128 256 512 1024
Trichophyton rubrumT. tonsuransT. mentagrophytes

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
154
DISCUSSÃO
A atividade antifúngica de itraconazol e terbinafina obtida em baixas concentrações,
verificada neste trabalho, tem sido mencionada de forma similar por outros autores (Fernández-
Torres et al. 2001; Favre et al. 2004; Barros et al. 2006).
As concentrações inibitórias que não foram determinadas, como citado na Tabela 2,
demonstram sensibilidade da maioria das amostras analisadas para uma mesma concentração de
própolis.
A atividade antifúngica da própolis verde e da vermelha do Brasil em relação a espécies de
Trichophyton, verificada neste trabalho, já havia sido constatada de forma semelhante com outras
própolis provenientes de São Paulo e do Paraná em relação a Trichophyton mentagrophytes e
T.rubrum (Soares e Cury 2001) e com própolis da Turquia, em relação a essas mesmas espécies
(Koc et al. 2005).
Sabe-se que pinocembrina, crisina (flavonóides) e ácido cinâmico (ácido fenólico) são os
principais responsáveis pela atividade antifúngica da própolis (Burdock 1998; Cushnie e Lamb
2005) e que a extração em álcool etílico promove a obtenção de mais compostos ativos da própolis
nativa. Dessa forma, a atividade fungistática e fungicida do extrato alcoólico da própolis verde e
da vermelha, constatada neste trabalho pode ser explicada pela composição química das própolis.
O extrato alcoólico da própolis verde do Brasil é rico em ácidos fenólicos, principalmente em
artepelin C, e apresenta maior concentração de flavonóides do que o extrato aquoso. Os ácidos
fenólicos e flavonóides do extrato alcoólico da própolis verde promovem, de um modo geral, o
sinergismo das atividades biológicas, incluindo a antifúngica, já o artepelin C em especial, é citado
principalmente pela atividade antitumoral (Banskota et al. 2000; Mello et al. 2006). Nakajima et
al. (2007) identificaram como os principais constituintes do extrato aquoso da própolis verde do
Brasil o ácido cafeiolquínico e derivados, e o ácido cinâmico e derivados (ácido p-cumárico,
artepelin C, bacarina), os quais já tinham sido identificados no extrato alcoólico. Entretanto como
o extrato alcoólico apresenta maior rendimento de compostos fenólicos, as atividades biológicas
são mais evidenciadas. Quando a água é utilizada como solvente, é citado que as atividades
antifúngicas e também as antibacterianas são mais fracas comparando com o extrato alcoólico
(Sawaya et al. 2002).
Outros autores mencionam a atividade antifúngica do extrato alcoólico da própolis verde
do Brasil em relação a espécies de Candida (Ota et al. 2001; Sawaya et al. 2002) e a atividade
antibacteriana do extrato alcoólico da própolis vermelha do Brasil em relação a Staphylococcus
aureus (Daugsch et al. 2007).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
155
As atividades fungistática e fungicida da própolis vermelha obtidas em concentrações
menores do que a própolis verde, verificada neste trabalho, pode está relacionada com a
composição química dessa própolis, pois há predomínio de flavonóides (pinocembrina e
pinobanskina), em vez de ácidos fenólicos (Kanazawa et al. 2003; Trusheva et al. 2006).
Provavelmente a maior concentração de flavonóides no extrato alcoólico da própolis vermelha
utilizada neste trabalho, tenha refletido a maior atividade antifúngica. Outros autores também
correlacionam a composição química da própolis com atividade antimicrobiana (Katircioglu e
Mercan 2006) e antifúngica (Havsteen 2002; Fernandes Júnior et al. 2006; Quiroga et al. 2006).
Os resultados referentes a maior susceptibilidade de T. rubrum a atividade fungistática dos
extratos alcoólicos das própolis utilizadas, estão de acordo com os de Soares e Cury (2001). Esses
autores constataram susceptibilidade de dermatófitos à atividade fungistática do extrato alcoólico
da própolis do Brasil fornecida pela Uniflora Apicultores Associados LTDA® nas concentrações
de 125µg ml-1 para T.rubrum e de 250µg ml-1 para T. mentagrophytes. Koc et al. 2005 também
constataram maior susceptibilidade das amostras de T. rubrum ao extrato alcoólico da própolis da
Turquia, na concentração de 0.025- 0.4µg ml-1, enquanto que para as amostras de T.
mentagrophytes foi de 0.1-0.8µg ml-1. Entretanto, as elevadas concentrações inibitórias mínimas
obtidas neste trabalho, refletem a composição química da própolis do Brasil, que é diferente da
própolis européia, o que é comprovado por outros autores que determinaram susceptibilidade de
amostras de Candida sp. aos extratos alcoólicos de própolis brasileiras provenientes do estado de
São Paulo, nas concentrações entre 3-12mg ml-1 (Ota et al. 2001) e 6-12mg ml-1 (Sawaya et al.
2002).
A semelhante susceptibilidade de T. tonsurans e T. mentagrophytes à atividade fungistática
dos extratos alcoólicos das própolis verde e vermelha utilizadas, demonstra a eficiente atividade
antifúngica pela inibição do crescimento de espécies diferentes de dermatófitos. Resultados
semelhantes foram citados por Koc et al. (2005) que verificaram atividade fungistática do extrato
alcoólico da própolis da Turquia na concentração de 0.076µg ml-1 para T. rubrum e 0.1µg ml-1
para T. mentagrophytes.
A atividade fungistática do extrato alcoólico das própolis verde e da vermelha em relação
às amostras de Trichophyton utilizadas demonstra a possibilidade da utilização dessa própolis em
associação com antifúngicos sintéticos, como alternativa para o tratamento de dermatofitoses por
espécies desse gênero. Esses resultados são corroborados por Soares e Cury (2001) que sugerem a
associação de antifúngicos com própolis como uma excelente alternativa para o tratamento de
dermatofitoses por T. rubrum, T. mentagrophytes e E. floccosum. De forma semelhante Stepanović
et al. (2003) constataram o sinergismo entre o extrato alcoólico da própolis da Sérvia e

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
156
antifúngicos, pela potencialização dessas drogas para inibição do crescimento de amostras de C.
albicans.
CONCLUSÕES
De acordo com os resultados obtidos é possível concluir que o extrato alcoólico da própolis
vermelha apresentou atividade antifúngica mais eficiente do que o da verde. As amostras de T.
rubrum foram mais sensíveis a atividade antifúngica do extrato alcoólico da própolis verde e da
vermelha e as amostras de T. tonsurans e T. mentagrophytes foram susceptíveis ao extrato
alcoólico da própolis verde em concentrações inibitórias mínimas semelhantes. O potencial
antifúngico da própolis verde e da vermelha para essas espécies, sugere aplicações futuras como
tratamento alternativo para dermatofitoses causadas por T. rubrum, T. tonsurans e T.
mentagrophytes
Agradecimentos
Agradecemos a concessão de bolsa pela Coordenação de Aperfeiçoamento de Pessoal de
Nível Superior (CAPES) como também pelo suporte financeiro e infra-estrutura da Universidade
Federal de Pernambuco (UFPE), Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq) e Rede de Coleções de Culturas de Microrganismos do Norte e Nordeste do Brasil
(RENNEBRA).
REFERÊNCIAS
Amaral, R.C., Gomes, R.T., Rocha, W.M.S., Abreu, S.L.R. and Santos, V.R. (2006) Periodontitis
treatment with brazilian green propolis gel. Pharmacology online 3, 336-341.
Bankova S. (2005) Recent trends and important developments in propolis research. Evid. Based
Complement. Altern. Med 2, 29-32.
Banskota, A.H., Tezuka, Y. and Kadota, S. (2000) Recent progress in pharmacological research of
propolis. Phytother Res 15, 561–71.
Barros, M.E.S., Santos D.A. and Hamdan, J.S. (2006) In vitro methods for antifungal
susceptibility testing of Trichophyton spp. Mycol Res 110, 1355-1360.
Burdock, G.A. (1998) Review of the biological properties and toxicity of bee propolis. Food Chem
Toxicol 36, 347- 363.
Castaldo, S. and Capasso, F. (2002) Propolis, an old remedy used in modern medicine. Fitoterapia
73, S1–S6.
Clinical and Laboratory Standards Institute - CLSI (2002) Reference Method for Broth Dilution
Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard document M38-A.
National Committee for Clinical Laboratory Standards, Wayne, PA, USA

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
157
Cushnie, T.P.T. and Lamb, A.J. (2005) Antimicrobial activity of flavonoids. Int J Antimicrob
Agents 26, 343–356.
Daugsch, A., Moraes, C.S., Fort, P. and Park, Y.K. (2007) Brazilian Red Propolis - Chemical
Composition and Botanical Origin. Evid Based Complement Alternat Med,
doi:10.1093/ecam/nem057.
Favre, B., Ghannoum, M.A. and Ryder, N.S. (2004) Biochemical characterization of terbinafine-
resistant Trichophyton rubrum isolates. Med Mycol 42, 525-529.
Fernandes, F.F., Dias, A.L.T., Ramos, C.L., Ikegaki, M., Siqueira, A,M and Franco, M.C. (2007)
The “in vitro” antifungal activity evaluation of propolis G12 ethanol extract on Cryptococcus
neoformans. Rev. Inst. Med. trop. S. Paulo 49: 93-95.
Fernandes Júnior, A., Lopes, M.M.R., Colombari, V., Monteiro, A.C.M. and Vieira, E.P. (2006)
Atividade antimicrobiana de própolis de Apis mellifera obtidas em três regiões do Brasil. Ciência
Rural 36, 294-297.
Fernández-Torres, B., Cabanes, F.J., Carrillo-Muñoz, A.J., Esteban, A., Inza, I., Abarca, L. and
Guarro, J. (2002) Collaborative evaluation of optimal antifungal susceptibility testing conditions
for dermatophytes. J Clin Microbiol 40, 3999-4003.
Gurgel, L.A., Sidrim, J.J.C., Martins, D.T., Cechinel Filho, V. and Rao, V.S. (2005) In vitro
antifungal activity of dragon’s blood from Croton urucurana against dermatophytes. J
Ethnopharmacol 97, 409-412.
Havsteen, B.H. (2002) The biochemistry and medical significance of the flavonoids. Pharmacol
Ther 96, 67-202.
Katircioglu, H. and Mercan, N. (2006) Antimicrobial activity and chemical compositions of
Turkish propolis from different regions. African J Biotechnol 5, 1151-1153.
Koc, A.N., Silici, S., Ayangil, D., Ferahbas, A.and Çankaya, S. (2005) Comparison of in vitro
activities of antifungal drugs and ethanolic extract of propolis against Trichophyton rubrum and T.
mentagrophytes by using a microdilution assay. Mycoses 48, 205-210.
Kujumgiev, A., Tsvetkova, I., Serkedjieva, Y., Bankova, V., Christov and R. Popov, S. (1999)
Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. J
Ethnopharmacol 64, 235-240.
Lacaz, C.S., Porto, E., Martins, J.E.C., Heins-Vaccari, E.M. and MELO, N.T. (2002) Tratado de
Micologia Médica. Rio de Janeiro: Sarvier, pp.252-340.
Mello, A.M., Gomes, R.T., Lara, R.S., Silva, L.G., Alves, J.B., Cortés, M.E., Abreu, S.L. and
Santos, V.R. (2006) The effect of brazilian propolis on the germ tube formation and cell wall of
Candida albicans. Pharmacologyonline 3, 352-358.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
158
Murad, M., Calvi, S.A., Soares, A.M.V.C., Bankova, V. and Sforcin, J.M. (2002) Effects of
propolis from Brazil and Bulgaria on fungicidal activity of macrophages against Paracoccidioides
brasiliensis. J Ethnopharmacol 79, 331-334.
Nakajima, Y., Shimazawa, M., Mishima, S. and Hara, H. (2007) Water extract of propolis and its
main constituents, caffeoylquinic acid derivatives, exert neuroprotective effects via antioxidant
actions. Life Sci 80, 370-377.
Oliveira, A.C.P., Shinobu, C.S., Onghini, R.L., Franco, S.L. and Svidzinski, T.I.E. (2006)
Antifungal activity of propolis extract against yeasts isolated from onychomycosis lesions. Mem
Inst Oswaldo Cruz 101, 493-497.
Ota, C., Unterkircher, C., Fantinato,V. and Shimizu, M.T. (2001) Antifungal activity of propolis
on different species of Candida. Mycoses 44, 375-378.
Park, Y.K., Koo, M.H., Abreu, J.A.S., Ikegaki, M., Cury, J.A. and Rosalen, P.L. (1998)
Antimicrobial activity of propolis on oral microorganisms. Curr Microbiol 36, 24-28.
Park, Y.K., Paredes-Guzman, J.F., Aguiar, C.L., Alencar, S.M. and Fujiwara, F.Y. (2004)
Chemical constituents in Baccharis dracunculifolia as the main botanical origin of Southeastern
Brazilian propolis. J Agric Food Chem 52, 1100-1103.
Porro, A.M., Yoshioka, M.C., Kaminski, S.K., Palmeira, M., Fischman, O. and Alchorne, M.M.
(1997) Disseminated dermatophytosis caused by Microsporum gypseum in two patients with the
acquired immunodeficiency syndrome. Mycopathologia 137, 9-12.
Quiroga, E.N., Sampietro, D.A., Sobero, J.R., Sgariglia, M.A. and Vattuone, M.A. (2006) Propolis
from the northwest of Argentina as a source of antifungal principles. J Appl Microbiol 101, 103-
110.
Santos, V.R., Pimenta, F. J. G. S., Aguiar, M.C.F., Do Carmo, M.A.V., Naves, M.D. and
Mesquita, R.A. (2005) Oral candidiasis treatment with brazilian ethanol propolis extract. Phytother
Res 19, 652–654.
Sawaya, A.C.H.F., Palma, A.M., Caetano, F.M., Marcucci, M.C., Da Silva Cunha, I.B., Araújo,
C.E.P. and Shimizu, M.T. (2002) Comparative study of in vitro methods used to analyse the
activity of propolis extracts with different compositions against species of Candida. Lett Appl
Microbiol 35, 203–207.
Sherf, A.F. (1943) A method for maintaining Phytomonas sepedomica for long periods without
transfer. Phytopathology 31, 30-32.
Soares, M.M.S.R. and Cury, A.E. (2001) In vitro activity of antifungal and antiseptic agents
against dermatophyte isolates from patients with tinea pedis. Braz J Microbiol 32, 130-134.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
159
Suzuki, I., Hayashi, I., Takaki, T., Groveman, D.S. and Fujimiya, Y. (2002) Antitumor and
anticytopenic effects of aqueous extracts of propolis in combination with chemotherapeutics
agents. Cancer Biother Radiopharm 17, 553-562.
Trusheva, B., Popova, M., Bankova, V., Simova, S., Marcucci, M.C., Miorin, P.L., Pasin, F.R. and
Tsvetkova, I. (2006) Bioactive constituents of Brazilian red propolis. Evid Based Complement
Alternat Med 3, 249-254.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
160
EFEITO DE LECTINAS DE LEGUMINOSAS DO BRASIL SOBRE O CRESCIMENTO IN
VITRO DE DERMATÓFITOS
Artigo a ser submetido para:
CELLULAR AND MOLECULAR LIFE SCIENCES
Qualis A
Fator de impacto 4,655 (Journal of Citation Report – 2006)

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
161
EFEITO DE LECTINAS DE LEGUMINOSAS DO BRASIL SOBRE O CRESCIMENTO IN
VITRO DE DERMATÓFITOS
Ana Beatriz Sotero Siqueira*1,2, Bruno Severo Gomes1,2, Rita Maia3, Cristina Souza-Motta1,
Benildo Souza Cavada5, Lusinete Aciole de Queiroz1, Ana Lucia F. Porto2,4
1Departmento de Micologia, Universidade Federal de Pernambuco (UFPE), Recife-Brasil 2 Laboratório de Imunopatologia Keizo Asami (LIKA), UFPE, Recife-Brasil 3 Centro de Pesquisa Aggeu Magalhães – CPqAM / FIOCRUZ, Recife-Brasil 4 Departamento de Morfologia e Fisiologia Animal, Universidade Federal Rural de Pernambuco
(UFRPE), Recife – PE, Brasil 5 Laboratório de Moléculas Biologicamente Ativas, Universidade Federal do Ceará, Fortaleza –
CE, Brasil
*Email para correspondência: [email protected]

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
162
RESUMO
Foi verificado o efeito das lectinas DViol, DRL, ConBr e LSL obtidas de sementes de leguminosas
sobre o crescimento de dermatófitos obtidos da Coleção de Culturas Micoteca – University of
Recife Mycology. O procedimento foi realizado em placas de microtitulação e os resultados
interpretados por leitura visual e pelo leitor de microplacas a 530nm. Foram utilizadas
concentrações de lectinas de 0.5 a 256µg/mL e o inóculo padronizado entre 65-70% de
transmitância. Foi verificado que as lectinas não apresentaram atividade antifúngica mas todas
apresentaram efeito estimulador sobre o crescimento desses fungos. Apenas a lectina DViol
estimulou a produção de esporos de Epidermophyton floccosum.
Palavras chaves: lectinas, leguminosa, Dviol, dermatófitos, Epidermophyton floccosum

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
163
INTRODUÇÃO
Lectinas são proteínas ou glicoproteínas que se ligam especificamente e reversivelmente a
mono e oligossacarídeos livres ou na forma de glicoconjugados (glicoproteína e glicolipídeo).
Dessa forma, combinam-se com moléculas e estruturas biológicas que contem esses açucares sem
alterar a estrutura covalente das ligações glicosídicas nos sítios de ligação [1, 2, 3].
São encontradas amplamente em animais, plantas, fungos, bactérias e vírus [2, 4, 5, 6]
entretanto essas substâncias estão presentes em maior quantidade em grãos de leguminosas e
gramíneas [1, 7, 8, 9]. As sementes de leguminosas como as de feijão e ervilha são fontes ricas de
lectinas as quais tem sido amplamente caracterizada em relação a propriedades química,
fisicoquímica, estrutural e biológica [1, 10, 11, 12]. As partes das plantas mais susceptíveis ao
ataque de predadores, como por exemplo, as sementes, são as que contem lectinas em maior
concentração. Com exceção de quitinases, glucanases e glicosidades, as lectinas são as únicas
proteínas capazes de reconhecer e ligar-se a glicoconjugados presentes na superfície de bactérias e
fungos ou expostos no trato intestinal de insetos e mamíferos [4, 8].
Dioclea violacea, D. rostrata e Canavalia brasiliensis são leguminosas das quais são
extraídas as lectinas DViol, DRL, ConBr respectivamente, as quais têm afinidade por glicose e
manose [11, 14, 15]. Essas lectinas já demonstraram indução da produção de histamina em ratos
[16], efeito protetor in vivo contra infecção por Leishmania amazonensis em camundongos
BALB/c [17], estimulação linfocitária em humanos [18], estimulação da produção de macrófagos
e linfócitos em camundongos C3H/HeJ [19].
Lonchocarpus sericeus é uma leguminosa da qual é extraída a lectina LSL com afinidade
por N-acetilglicosamina [12]. Já demonstrou atividade antiinflamatória em casos de peritonite em
ratos, notável diminuição de bactérias que colonizavam essa região peritoneal [9] e inibição da
migração de neutrófilos em processos inflamatórios [12].
Alguns trabalhos relatam a atividade antifúngica de lectinas de sementes de leguminosas
tais como Phaseolus vulgaris [8]; de frutas como Talisia esculenta [20], Artocarpus integrifolia,
A. incisa [21] e Annona muricata [22]; de fungos como Kluyveromyces bulgaricus [23] e
Schizophyllum commune [24]; de Urtica dioica [25]; de batata [26]; de germe de trigo [27] e de
Astragalus mongholicus [28]. Essas lectinas são denominadas, respectivamente: lectinas de
Phaseolus vulgaris, TEL, “jackin”, “frutackin”, AML, Kb-CWL I, SCL, UDA, lectina de batata,
WGA, AMML.
A susceptibilidade a atividade antifúngica de lectinas já foi constatada em leveduras como
Sacchromyces cerevisiae [20, 21, 23], S. bayanus, S. uvarurn, Candida albicans, Candida
glabrata, Rhodotorula mucilaginosa, Pichia pastoris, Kluyveromyces bulgaricus, K. lactis,

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
164
Schizosacchamyces bulgaricus, Schizosacchamyces octosporus [23] e em fungos filamentosos
como Coprinus comatus, Rhizoctonia solani [8], Fusarium oxysporum [8, 22, 27, 28],
Colletorichum lindemuthianum [20], F. moniliforme [21], Colletorichum musae, F. solani [22],
Aspergillus niger [24], Phoma betae, Phycomyces blakesleeanus, Septoria nodorum, Phytophthora
erythroseptica [25], F. graminearum [27], Colletorichum sp., Drechslera turcia [28], Botrytis
cincerea [25, 28], Trichoderma viride, T. hamatum [25].
Entretanto, outras lectinas não apresentam atividade antifúngica [13, 29, 30, 31, 32] e
algumas demonstram efeito ativador da cinética do crescimento do fungo [23, 33, 34].
Os dermatófitos são fungos filamentosos pertencentes aos gêneros Trichophyton,
Microsporum e Epidermophyton, possuem propriedades queratinofílicas e queratinolíticas e são
capazes de infectar pele, pêlos e unhas [35]. São rotineiramente identificados através de aspectos
macroscópicos, microscópicos e fisiológicos entretanto, o pleomorfismo que é comum em
dermatófitos pode dificultar a manifestação desses aspectos [36, 37].
Este trabalho teve como objetivo verificar o efeito de lectinas de sementes de leguminosas
sobre o crescimento de amostras de dermatófitos.
MATERIAL E METODOS
Fungos
Foram utilizadas 30 de seis espécies de dermatófitos, sendo cinco amostra de cada espécie:
Microsporum canis, Microsporum gypseum, Trichophyton tonsurans, Trichophyton
mentagrophytes, Trichophyton rubrum e Epidermophyton floccosum. Tais fungos obtidos da
Coleção de Culturas da Micoteca University of Recife Mycology - URM, da Universidade Federal
de Pernambuco e estavam preservados sob óleo mineral [38]. A maioria das amostras foi isolada
de dermatofitoses enquanto que as demais foram isoladas do solo e de praias da cidade do Recife
(Tabela 1).
Meio de cultura
O meio de cultura utilizado para a avaliação do efeito das lectinas sobre o crescimento dos
dermatófitos foi o RPMI 1640 (Sigma-Aldrich, USA) com L-glutamina e sem bicarbonato de
sódio, tamponado a pH 7.0 com ácido morfolinepropanesulfônico (MOPS) a 0.165 M (Sigma-
Aldrich, USA). Esse meio de cultura foi esterilizado por filtração em membrana com porosidade
de 0,22µm (Millipore).
Lectinas
Foram utilizadas quatro lectinas extraídas de leguminosas. As lectinas foram DViol, DRL,
ConBr e LSL, extraídas respectivamente de Dioclea violacea, Dioclea rostrata, Conavalia
brasiliensis e Lonchocarpus sericeus. A primeira lectina foi obtida de sementes de leguminosas do

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
165
estado do Rio Grande do Sul (Brasil) e as demais do estado do Ceará (Brasil). Essas lectinas
foram purificadas [1, 11, 39, 40] e gentilmente fornecidas pelo Laboratório de Moléculas
Biologicamente Ativas da Universidade Federal do Ceará. Após solubilização em água destilada e
esterilizada, foram preparadas dez concentrações entre 0.5 e 256µg/mL, seguido de diluições
seriadas para atingir a concentração final de 10 vezes maior do que a concentração utilizada no
procedimento. Posteriormente, foram diluídas (1:5) em RPMI 1640 para obter a concentração de
duas vezes maior do que a necessária.
Preparação do inóculo
As amostras de dermatófitos foram semeadas em meio de Batata-Dextrose-Ágar contido
em tubos de ensaio os quais foram mantidos a temperatura ambiente (28+ 2ºC) até a esporulação
de cada amostra. Posterirmente foram acrescentados 5ml de água destilada esterilizada nos tubos
de ensaio e com auxílio de pipeta Pasteur foi promovido desprendimento de esporos e fragmentos
de hifas. Após 20 minutos em repouso, o sobrenadante foi aspirado para outro tubo de ensaio
esterilizado e agitado em vortex por 15 segundos. A densidade das suspensões foi ajustada em
espectrofotômetro a 530nm, para obter entre 65-70% de transmitância. Posteriormente, as
suspensões foram diluídas (1:50) em RPMI 1640 para obter a concentração de 2 vezes maior do
que a necessária.
Avaliação do efeito de lectinas sobre o crescimento de dermatófitos
O procedimento foi realizado em placas esterilizadas de microtitulação com tampa (TPP,
Suíça), contendo 96 poços com formato de U (fundo chato) nas quais foram depositados 100µL
das lectinas nas fileiras de 1 a 10, sendo que cada concentração foi depositada em uma fileira. Nos
poços das fileiras 11 e 12 foram depositados 100µL do meio RPMI 1640, os quais foram o
controle de crescimento e o de esterilização do meio, respectivamente. Essas placas foram
armazenadas a -20ºC até o momento do uso. No momento adequado, 100µL do inóculo
padronizado foi depositado nos poços das fileiras 1 a 11.
Após inoculação, as placas de microtitulação foram incubadas a temperatura ambiente (28+
2ºC) por sete dias para interpretação dos resultados [41]. A avaliação macroscópica do crescimento
das amostras nos poços das placas de microtitulação foi realizada por observação visual de cada
poço, comparando com o crescimento no poço controle. Além disso, essas placas foram
submetidos ao leitor de microplacas, programa Microplate Manager, modelo BenchMark Plus
(Bio-Rad Laboratories Inc.) a 530nm.
Para avaliação microscópica, foram retirados fragmentos das colônias desenvolvidas nas
placas de microtitulação e corados com azul de Aman. As amostras que apresentaram
características microscópicas típicas da espécie foram semeadas em meio Sabouraud-Dextrose-

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
166
Ágar contido em placas de Petri, para observação de características macroscópicas, as quais foram
identificadas segundo os autores [36, 37].
Análise dos dados
Todo experimento foi realizado em duplicata. De cada concentração, foi determinada a
média do crescimento fúngico.
RESULTADOS
Nenhuma das lectina testadas apresentou atividade antifúngica para as amostras de
dermatófitos analisadas. Entretanto de acordo com os resultados obtidos no sétimo dia de
incubação, tanto pela avaliação visual quanto pelo leitor de microplacas, foi constatado que todas
as amostras de dermatófitos utilizadas apresentaram aumento do crescimento diretamente
proporcional ao aumento da concentração das lectinas utilizadas (Figuras 1A, 1B, 2A e 2B).
Foi realizada observação microscópica de fragmentos das colônias dos dermatófitos
desenvolvidos nas placas de microtitulação e foi observado que apenas as amostras de E.
floccosum no sétimo dia de incubação, produziram em grande quantidade esporos característicos
na presença da lectina DViol (256µg/mL). Foram observadas macroaleurias claviformes, isoladas,
sem ornamentações na parede celular, sem ou com septos (Figura 3A). Não foram observados
clamidósporos nem microaleurias. Não houve produção de esporos de E. floccosum na ausência
dessa lectina nem quando essa espécie foi submetida à incubação em concentrações menores de
DViol (Figura 3B). Para as demais amostras de Microsporum e Trichophyton na presença das
lectinas utilizadas (DViol, DRL, ConBr e LSL) em todas as concentrações testadas, não houve
produção de esporos.
A avaliação macroscópica das culturas de E. floccosum em meio de ágar Sabouraud, após
incubação com a lectina DViol, revelou características da espécie analisada, tais como superfície
de coloração branca (Seguy, 1936 - Pl XLVI 680 albus), textura cotonosa, margem regular,
aspecto plano (URM 4799, URM 4798, URM 4151) ou com sulcos radiais (URM 3195, URM
3345) (Figura 4A). O reverso apresentou-se plano com pigmentação variando entre amarelo
(Seguy, 1936 - PlXV 214 orange), amarelo mostarda (Seguy - PlXV 215 orange) e amarelo-
amarronzado (Seguy - PlXV 215 orange, PlXXIII 337 vert). As colônias com pigmentação
amarela foram URM 3195 e URM 4799, a amarelo-mostarda foi URM 4798 e as amarronzadas
foram URM 4151 e URM 3345 (Figura 4B).
DISCUSSÃO
Lectinas de sementes de leguminosas sem atividade antifúngica e com especificidade de
ligação à glicose e manose também já foram relatadas por outros autores, tais como a lectina de

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
167
ervilha Pisum sativum var. macrocarpon [13], de castanha chinesa Castanea mollisima [29] e de
feijão Canavalia gladiata [32].
Entretanto, há relatos de outras lectinas que podem se ligar à glicose, manose e N-
acetilglicosamina com atividade antifúngica. Entre essas, a lectina TEL já promoveu inibição de
57% do crescimento de Sacchromyces cerevisiae, Fusarium moniliforme e Colletotrichum
lindemuthianum na concentração de 280µg/mL [20] e a lectina AML (glicose/manose) que
promoveu inibição de 41% do crescimento de F. solani, F. oxysporum e Colletotrichum musae na
concentração de 100µg/mL [22]. Trindade et al. [21] verificaram a atividade antifúngica das
lectinas “jackin” e “frutackin” (N-acetilglicosamina) a 2.25mg/mL para S. cerevisiae e Fusarium
moniliforme.
O estímulo do crescimento dos dermatófitos utilizados neste trabalho pelas lectinas DViol,
DRL, ConBr e LSL é corroborado por Viard et al. [23], que também observaram o efeito
estimulador da lectina Kb-CWL I, obtida de Kluveromyces bulgaris e com afinidade de ligação à
galactose, sobre o crescimento in vitro de Candida albicans e C. tropicalis.
De acordo com os resultados obtidos neste trabalho, referente ao período de esporulação
nos meios de SDA, DTM e BDA, foi verificado que E. floccosum é um fungo que esporula
lentamente, mas em contato com a lectina DViol, esse período diminuiu e produção de esporos foi
estimulada. De forma semelhante, outros autores [33, 34] verificaram que a lectina Concanavalina
A (ConA), com a afinidade de ligação por glicose e manose, estimulou a germinação de esporos de
Ustilago maydis. Hernández et al. [34] constataram algumas alterações morfológicas decorrentes
da atuação da ConA sobre basidioporos de Ustilago maydis, tais como espessamento da parede
celular e deformação desses esporos. Entretanto, nenhuma alteração microscópica foi verificada
nas amostras de E. floccosum utilizadas neste trabalho.
A obtenção de grande quantidade de esporos de E. floccosum resultante da estimulação
pela lectina DViol a 250µg/mL, está de acordo com os resultados obtidos por Pérez-Santiago et al.
[33]. Esses autores verificaram que a lectina Con A (glicose/manose) aumentou cerca de 30% a
esporulação de Ustilago maydis a 100µg/mL
A análise da composição química da parece celular dos dermatófitos indica que entre os
carboidratos, há predomínio de glicose, manose, glicomanose e N-acetilglicosamina [43]. Shah e
Knight [44] constataram diferenças significativas na elevada concentração de glicose na parede
celular de E. floccosum. Talvez esse carboidrato tenha favorecido o efeito da lectina DViol
utilizada neste trabalho sobre essa espécie de dermatófito.
De acordo com os resultados obtidos foi concluído que as lectinas DViol, DRL, ConBr e LSL
não apresentaram atividade antifúngica nas concentrações testadas, mas todas estimularam o

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
168
crescimento das amostras de dermatófitos em todas as concentrações utilizadas. Apenas a lectina
DViol estimulou a esporulação das amostras de E. floccosum.
REFERÊNCIAS
1 Moreira, R.A., Cordeiro, E.F., Ramos, M.V., Grangeiro, T.B., Martins, J.L., Oliveira, J.T.A. and
Cavada, B.S. (1996) Isolation and partial characterization of a lectin from seeds of Dioclea
violacea. R. Bras. Fisio. Veg. 8, 23-29.
2 Cavada, B.S., Santos, C.F., Grangeiro, T.B., Nunes, E.P., Sales, P.V.P., Ramos, R.L., Sousa,
F.A.M., Crisostomo, C.V. and Calvete, J.J. (1998) Purification and characterization of a lectin
from seeds of Vatairea macrocarpa duke. Phytochem. 49, 675-680.
3 Wong, J.H., Wong, C.C.T. and Ng, T.B. (2006) Purification and characterization of a galactose-
specific lectin with mitogenic activity from pinto beans. Biochem. Biophysi. Acta. 1760, 808–813,
4 Peumans, W.J. and Van Damme, E.J.M. (1995) Lectins as plant defense proteins. Plant Physiol.
109, 347-352.
5 Ngai, P.H.K. and Ng, T.B. (2004) A mushroom (Ganoderma capense) lectin with spectacular
thermostability, potent mitogenic activity on splenocytes, and antiproliferative activity toward
tumor cells. Biochem. Biophys. Res. Commun. 314, 988-993.
6 Lis, H. and Sharon, N. (2003) Lectins: carbohydrate-specific proteins that mediate cellular
recognition. Chem. Rev. 98, 637–674.
7 Cavada, B.S., Madeira, S.V.F., Calvete, J.J., Souza, L.A., Bomfim, L.R., Dantas, A.R., Lopes,
M.C.; Grangeiro, T.B., Freitas, B.T., Pinto, V.P.; Leite, K.B. and Ramos, M.V. (2000)
Purification, chemical, and immunochemical properties of a new lectin from Mimosoideae (Parkia
discolor). Prep. Biochem. Biotechnol. 30, 271-280.
8 Ye, X.Y., Ng, T.B., Tsang, P.W.K. and Wang, J. (2001) Isolation of a homodimeric lectin with
antifungal and antiviral activities from red kidney bean (Phaseolus vulgaris). J. Protein. Chem. 20,
367-375.
9 Alencar, N.M., Cavalcante, C.F., Vasconcelos, M.P., Leite, K.B., Aragão, K.S., Assreuy, A.M.,
Nogueira, N.A., Cavada, B.S. and Vale, M.R. (2005) Anti-inflammatory and antimicrobial effect
of lectin from Lonchocarpus sericeus seeds in an experimental rat model of infectious peritonitis.
J. Pharm. Pharmacol. 57, 919-922
10 Moreira, R.A., Silva, L.M.A., Horta, A.C.G., Oliveira, J.T.A. and Cavada, B.S. (1993) Lectin
from Canavalia brasiliensis Mart. Behavior during maturation and detection of a lectin precursor.
R. Bras. Fisio. Veg. 5, 133-138.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
169
11 Cavada, B.S., Grangeiro, T.B., Ramos, M.V., Cordeiro, E.F., Oliveira, J.T.A. and Moreira,
R.A. (1996) Isolation and partial characterization of a lectin from Dioclea rostrata Benth seeds. R.
Bras. Fisio. Veg. 8, 31-36.
12 Napimoga, M.H., Cavada, B.S., Alencar, N.M.N., Mota, M.L., Bittencourt, F.S., Alves-Filho,
J.C., Grespan, R., Gonçalves, R.B., Clemente-Napimoga, J.T., Freitas, A., Parada, C.A., Ferreira,
S.H. and Cunha, F.Q. (2007) Lonchocarpus sericeus lectin decreases leukocyte migration and
mechanical hypernociception by inhibiting cytokine and chemokines production. Int.
Immunopharmacol. 7, 824–835.
13 Ye, X. and Ng, T.B. (2001) Isolation of lectin and albumin from Pisum sativum var.
macrocarpon ser. cv. sugar snap. Int. J. Biochem. Cell. Biol. 33, 95-102.
14 Cavada, B. S., Moreira, R. A., Oliveira, J. T. A. and Grangeiro, T. B. (1993) Primary structures
and functions of plant lectins. R. Bras. Fisio. Veg. 5, 193-201.
15 Barbosa, T., Arruda, S., Cavada, B., Grangeiro, T. B., Freitas, L. A. R. and Barral-Netto, M.
(2001) In vivo lymphocyte activation and apoptosis by lectins of Diocleinae subtribe. Mem. Inst.
Oswaldo Cruz 96, 673-678
16 Gomes, J.C., Cavada, B.S., Moreira, R.A. and Oliveira, J.T.A. (1994) Histamine release
induced by glucose (mannose)-specific lectins isolated from Brazilian plants. Comparison with
concanavalin A. Agents Actions 41, 132-135.
17 Barral-Netto, M., Von Sohsten, R. L., Teixeira, M., Santos, W. L., Pompeu, M. L., Moreira, R.
A., Oliveira, J. T., Cavada, B. S., Falcoff, E. and Barral, A. (1996) In vivo protective effect of the
lectin from Canavalia brasiliensis on Balb/ mice infected by Leishmania amazonensis. Acta
Tropica. 60, 237-250.
18 Barral-Netto, M.; Santos, S.B., Moreira, R.A., Oliveira, J.T.A., Barral, A., Moreira, L.I.M.,
Santos, C.F. and Cavada, B.S. (1992) Human lymphocyte stimulation by lectins from Brazilian
leguminous seeds. Immunol. Invest. 21, 297-303
19 Rodriguez, D., Cavada, B. S., Oliveira, J. T. A., Moreira, R. A. and Russo, M. (1992)
Differences in macrophage stimulation and leukocyte accumulation in response to intraperitoneal
administration of glucose/mannose-binding lectins. Braz. J. Med. Biol. Res. 25, 823-826.
20 Freire, M.G.M., Gomes V.M., Corsini, R.E., Machado, O.L.T., De Simone, S.G., Novello,
J.C., Marangoni, S. and Macedo, M.L.R. (2002) Isolation and partial characterization of a novel
lectin from Talisia esculenta seeds that interferes with fungal growth. Plant. Physiol. Biochem. 40,
61-68.
21 Trindade, M.B., Lopes, J.L.S., Soares-Costa, A., Monteiro-Moreira, A.C., Moreira, R.A.,
Oliva, M.L.V. and Beltramini, L.M. (2006) Structural characterization of novel chitin-binding

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
170
lectins from the genus Artocarpus and their antifungal activity. Biochem. Biophysi. Acta. 1764,
146-152.
22 Damico, D.C.S., Freire, M.G.M., Gomes, V.M., Toyama, M.H., Marangoni, S., Novello, J.C.
and Macedo, M.L.R. (2003) Isolation and characterization of a lectin from Annona muricata seeds.
J. Protein. Chem. 22, 655-661.
23 Viard, B., Al-Mahmood, S., Streiblova, E. and Bonaly, R. (1993) Alternate interactions of the
galactose-specific yeast lectin Kb-CWL I with sensitive yeast strains. FEMS Microbiol. Lett. 107,
17-24.
24 Chumkhunthod, P., Rodtong, S.; Lambert, S.J., Fordham-Skelton, A.P., Rizkallah, P.J.,
Wilkinson, M.C. and Reynolds, C.D. (2006) Purification and characterization of an N-acetyl-D-
galactosamine-specific lectin from the edible mushroom Schizophyllum commune. Biochem.
Biophysi. Acta. 1760, 326–332.
25 Brockaert, W.F., Van Parijs, J., Leyns, F., Joos, H. and Peumans, W.J. (1989) A chitin-binding
lectin from stinging nettle rhizomes with antifungal properties. Science. 245, 1100-1103.
26 Gozia, O., Ciopraga, J., Bentia, T., Lungu, M., Zamfirescu, I., Tudor, R., Roseanu, A. and
Nitu, F. (1993) Antifungal properties of lectin and new chitinases from potato tubers. C. R. Acad.
Sci. III. 316, 788-92.
27 Ciopraga, J., Gozia, O., Tudor, R., Brezuica, L. and Doyle, R.J. (1999) Fusarium sp. growth
inhibition by wheat germ agglutinin. Biochem. Biophysi. Acta. 1428, 424-432.
28 Yan, Q., Jiang, Z., Yang, S., Deng, W. and Han, L. (2005) A novel homodimeric lectin from
Astragalus mongholicus with antifungal activity. Arch. Biochem. Biophys. 442, 72-81.
29 Ng, T.B., Yu, Y.L. and Chu, K.T. (2002) Isolation of a novel legumin-like lectin with potent
hemagglutinating activity from seeds of the Chinese chestnut Castanea mollisima. Comp Biochem
Physiol. C. Toxicol. Pharmacol. 133, 453-460.
30 Wong, J.H. and Ng, T.B. (2003) Purification of a trypsin-stable lectin with antiproliferative
and HIV-1 reverse transcriptase inhibitory activity. Biochem. Biophys. Res. Commun. 301, 545-
550.
31 Wong, J.H. and Ng, T.B. (2005) Isolation and characterization of a glucose/mannose/rhamnos
especific lectin from the knife bean Canavalia gladiata. Arch. Biochem. Biophys. 439, 91-98.
32 Wong, J.H., Wong, C.C.T. and Ng, T.B. (2006) Purification and characterization of a
galactose-specific lectin with mitogenic activity from pinto beans. Biochem. Biophysi. Acta 1760,
808–813.
33 Pérez-Santiago, A., Saavedra, E., Pérez Camposa, E. and Córdoba, F. (2000) Effect of plant
lectins on Ustilago maydis “in vitro”. Cell. Mol. Life Sci. 57, 1986-1989.

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
171
34 Hernández, E.S., Santiago, A.P., Campos, E.P. and Alva, F.C. (2003) Efecto de la
concanavalina A en el cultivo del fitopatógeno Ustilago maydis. Interciencia 28, 276-280.
35 Larone, D.H (1996) Culture and Identification of Dermatophytes. Clin. Microbiol. News. 18,
33-40..
36 Otcenasek, M. and Dvorak, J. (1973). Pictorial Dictionary of Medical Mycology.
TheHague: W. Junk B. V., 1973. 229p.
37 Rebell, G. and Taplin, D. (1979) Dermatophytes: their recognition and identification. 124p.
University of Miami Press (ed), Florida.
38 Sherf, A.F. (1943) A method for maintaining Phytomonas sepedomica for long periods without
transfer. Phytopathol. 31, 30-32.
39 Moreira, R. A. and Cavada, B. S. (1984) Lectin from Canavalia brasiliensis (MART).
Isolation, characterization and behavior during germination, Biol. Plant. (Prague) 26, 113- 120.
40 Alencar, N.M., Teixeira, E.H., Assreuy, A.M.S., Cavada, B.S., Flores, C,A, and Ribeiro, R.A.
(1999) Leguminous lectins as tools for studying the role of sugar residues in leukocyte
recruitment. Mediators. Inflamm. 8, 107–13.
41 Fernández-Torres, B., Cabañes, F.J., Carrillo-Muñoz, A.J., Esteban, A., Inza, I., Abarca, L. and
Guarro, J. (2002) Collaborative evaluation of optimal antifungal susceptibility testing conditions
for dermatophytes. J. Clin. Microbiol. 40, 3999-4003.
42 Segyu, E. (1936) Code Universal des Coulours. 68p: il. Paul Lechevalier (ed), Paris.
43 Kitajma, Y. (2000) Structural and biochemical characteristics of pathogenic fungus: cell walls,
lipids and dimorphism, and action modes of antifungal agents. Nippon Ishinkin Gakkai Zasshi. 41,
211-217.
44 Shah, V.K. and Knight, S.G. (1968) Chemical composition of hyphal walls of dermatophytes
Arch. Biochem. Biophys. 127, 229-234

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
172
EE: escamas epidérmicas; DE: desconhecido; PR: praias de Recife; SO: solo; EU: escamas ungueais
Espécie Nº registro Ano de estoque Período de estoque Substrato (URM) (ano)
Microsporum canis M. canis M. canis M. canis M. canis M. gypseum M. gypseum M. gypseum M. gypseum M. gypseum Trichophyton mentagrophytes T. mentagrophytes T. mentagrophytes T. mentagrophytes T. mentagrophytes T. tonsurans T. tonsurans T. tonsurans T. tonsurans T. tonsurans T. rubrum T. rubrum T. rubrum T. rubrum T. rubrum Epidermophyton floccosum E. floccosum E. floccosum E. floccosum E. floccosum
4962 4225 4154 4678 4158 4964 3060 6034 3645 4279 4156 3239 2785 4422 2875 4965 4963 4968 4945 4946 4725 4724 4727 4728 4569 3345 4151 4799 4798 3195
2005 1999 1999 2003 1998 2005 1989 2005 1995 2000 1999 1992 1985 2002 1986 2005 2005 2005 2004 2004 2003 2003 2003 2003 2004 1993 1999 2004 2004 1990
2 8 8 4 9 2
18 2
12 7 8
15 22 5
21 2 2 2 3 3 4 4 4 4 5
14 8 3 3
17
EE EE EE EE EE EE DE EE EE AP AP EE SO EE DE EE EE EE EE EE EE EE EE EE EU EE EE EE EE EE
Tabela 1. Espécies de dermatófitos preservados sob óleo mineral na Coleção de
Cultura Micoteca - University of Recife Mycology (URM).

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
173
Figura 1. Crescimento de amostras de dermatófitos obtidas da Coleção de Culturas Micoteca – University of Recife Mycology em diferentes concentrações da lectina Dviol (A) e DRL (B).
Microsporum canis M. gypseum
Trichophyton rubrum
T. mentagrophytes
T. tonsurans Epidermophyton floccosum
Concentração (µg/mL) da lectina DViol
D.O
530
nm
A
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 0.5 1 2 4 8 16 32 64 128 256
B
D.O
530
nm
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 0.5 1 2 4 8 16 32 64 128 256
Concentração (µg/mL) da lectina DRL
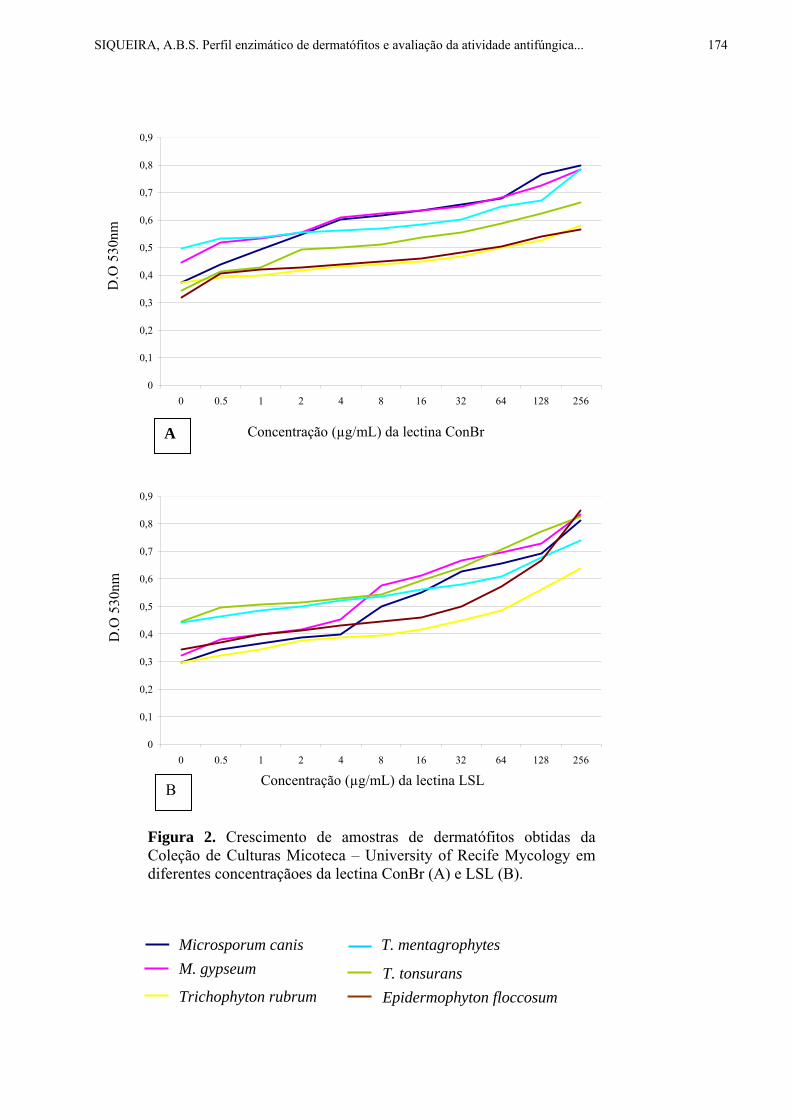
SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
174
Figura 2. Crescimento de amostras de dermatófitos obtidas da Coleção de Culturas Micoteca – University of Recife Mycology em diferentes concentraçãoes da lectina ConBr (A) e LSL (B).
Microsporum canis M. gypseum
Trichophyton rubrum
T. mentagrophytes
T. tonsurans Epidermophyton floccosum
Concentração (µg/mL) da lectina ConBr A
D.O
530
nm
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 0.5 1 2 4 8 16 32 64 128 256
Concentração (µg/mL) da lectina LSL B
D.O
530
nm
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 0.5 1 2 4 8 16 32 64 128 256

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
175
Figura 3. (A) Estruturas microscópicas de Epidermophyton floccosum após incubação por sete dias a temperatura ambiente com a lectina DViol a 256µg/mL: macroaleurias claviformes, isoladas, sem ornamentações na parede celular, com ou sem septos (seta). (B) Filamentos micelianos septados e hialinos de E. floccosum na ausência da lectina DViol
B
A

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
176
Figura 4A. Aspectos macroscópicos do verso da cultura de Epidermophyton floccosum em meio de cultura ágar Sabouraud: coloração branca, superfície plana (A’ – URM 4798) e com sulcos radiais (B’ – URM 3345).
A’ B’
Figura 4B. Aspectos macroscópicos do reveso da cultura de Epidermophyton floccosum em meio de cultura ágar Sabouraud: pigmentação amarela (A’ – URM 3195), amarela-mostarda (B’ – URM 4798) e amarela-amarronzada (C’ – URM 3345).
A’ B’ C’

SIQUEIRA, A.B.S. Perfil enzimático de dermatófitos e avaliação da atividade antifúngica...
177
6. CONCLUSÕES
Os resultados obtidos permitem concluir que:
1. As amostras analisadas de Microsporum canis, M. gypseum e Trichophyton
mentagrophytes apresentaram aspectos macroscópicos de pleomorfismo;
2. As amostras analisadas de T. tonsurans, T. rubrum e Epidermophyton floccosum
apresentaram aspectos taxonômicos característicos das espécies;
3. O período de estocagem não impediu a expressão de aspectos taxonômicos para a
identificação das amostras de dermatófitos utilizadas;
4. O método de preservação utilizado pela Coleção de Culturas Micoteca-URM (UFPE)
mostrou-se eficiente;
5. As amostras de dermatófitos analisadas apresentaram perfil enzimático característico das
espécies avaliadas;
6. Os extratos alcoólicos das própolis verde e vermelha apresentaram atividade antifúngica
em relação aos dermatófitos avaliados;
7. As amostras de M. canis e T. rubrum foram mais susceptíveis a atividade antifúngica do
extrato alcoólico da própolis verde e da vermelha;
8. As amostras de T. tonsurans e T. mentagrophytes foram susceptíveis ao extrato alcoólico
da própolis verde em concentrações inibitórias mínimas semelhantes;
9. O potencial antifúngico do extrato alcoólico da própolis vermelha foi maior do que o da
verde;
10. As lectinas DViol, DRL, ConBr e LSL não apresentaram atividade antifúngica nas
concentrações testadas;
11. Todas as lectinas estimularam o crescimento das amostras de dermatófitos em todas as
concentrações utilizadas;
12. Apenas a lectina DViol estimulou a esporulação das amostras de E. floccosum.





























