Embed Size (px)
Citation preview

An MEG Study of Picture Naming
Willem J M Levelt Peter Praamstra and Antje S MeyerMax Planck Institute for Psycholinguistics The Netherlands
Paumlivi Helenius and Riitta SalmelinHelsinki University of Technology
Abstract
n The purpose of this study was to relate a psycholinguisticprocessing model of picture naming to the dynamics of corticalactivation during picture naming The activation was recordedfrom eight Dutch subjects with a whole-head neuromagne-tometer The processing model based on extensive naminglatency studies is a stage model In preparing a picturersquos namethe speaker performs a chain of specic operations They arein this order computing the visual percept activating an ap-propriate lexical concept selecting the target word from themental lexicon phonological encoding phonetic encodingand initiation of articulation The time windows for each ofthese operations are reasonably well known and could berelated to the peak activity of dipole sources in the individual
magnetic response patterns The analyses showed a clear pro-gression over these time windows from early occipital activa-tion via parietal and temporal to frontal activation The majorspecic ndings were that (1) a region in the left posteriortemporal lobe agreeing with the location of Wernickersquos areashowed prominent activation starting about 200 msec afterpicture onset and peaking at about 350 msec (ie within thestage of phonological encoding) and (2) a consistent activationwas found in the right parietal cortex peaking at about 230msec after picture onset thus preceding and partly overlap-ping with the left temporal response An interpretation in termsof the management of visual attention is proposed n
INTRODUCTION
Salmelin Hari Lounasmaa and Sams (1994) reported therst successful magnetoencephalograph (MEG) study ofpicture naming The study demonstrated that picturenaming is a feasible task in MEG studies of languageproduction and also that a distinct dynamics of corticalactivation is obtained that progresses bilaterally from theoccipital visual cortex toward the temporal and frontallobes This dynamics must relate to the time course ofprocesses underlying the naming of a picture rangingfrom the visual analysis of the picture to the motorexecution of the picturersquos name Picture naming is anexcellent choice as a language production task It is anatural task it involves all stages of language productionat least to some extent and importantly it is one of thebest-studied tasks in language production research Pic-ture naming is widely used to probe language functionduring brain surgery as pioneered by Peneld andRoberts (1959) and Ojemann (1983) Preceding resectivesurgery for epilepsy or a cerebral tumor the patientunder local anaesthesia is tested for ldquolanguage sitesrdquo inthe relevant areas of the cortex This is done by stimula-tion mapping (ie electrical stimulation of a particularbrain area while the patient names a picture) If thenaming response is erroneous or fully blocked the area
copy 1998 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 105 pp 553ndash567
is mapped as a language site Although phonemic andsemantic errors do occur during stimulation mappingthe typical result of stimulating a language site is fullomission of the naming response The two major reviewsof this work (Ojemann Ojemann Lettich amp Berger 1989Haglund Berger Shamseldin Lettich amp Ojemann 1994)encompassing a total of 126 epilepsy and glioma pa-tients report an average of 2 to 3 detected language sitesper patient (when 10 to 20 sites were tested) Althoughthese sites were largely concentrated in the superiortemporal gyrus the motor strip and premotor Brocarsquosarea of the (tested) left hemisphere language sites alsoappeared in the medial temporal gyrus and the parietaland prefrontal cortices Patients differed substantially intheir ldquolanguage mapsrdquo No single cortical area was al-ways involved for all patients These ndings make itunlikely that picture naming has a uniform cortical or-ganization across speakers Although this is an importantconclusion by itself one would like to see it furtherqualied The failure to name under stimulation (whichdenes a language site) may after all be due to interfer-ence with any of the processing stages involved in pic-ture naming Failing to recognize the object categorizeit semantically retrieve the target word or its phonologi-cal form from the lexicon or program the wordrsquos articu-latory gesture will block the naming response It is most
unlikely that a single language site performs all of thesefunctions
One would like to know how for individual subjectsthese components of the picture naming process aredistributed over the brain Only then we will begin todiscover what is uniform across speakers and what isvariable The obvious rst hunch is that the input andoutput processes in picture naming (perceiving the ob-ject articulating the word) are much less variable in theircerebral localization than the mediating processes oflexical access and of phonological encoding In particu-lar one should expect to nd occipital lobe activation ina picture naming task as well as activation in the facearea of the sensory-motor cortex But what about themore endogenous responses In a positron emissiontomography (PET) study of object naming and recogni-tion Price Moore Humphreys Frackowiak and Friston(1996) found the following areas involved with objectrecognition ventral and dorsal regions of the left middleoccipital lobe bilateral anterior temporal lobes and theleft cerebellum Martin Wiggs Ungerleider and Haxby(1996) also in a PET study found the left middle occipi-tal lobe only active in the naming of animals but not oftools This study as well as the one by DamasioGrabowski Tranel Hichwa and Damasio (1996) suggeststhat the areas involved in the semantic processing ofpictures are different for different categories of picturesto be named (but the studies disagree on what areas arespecically involved in naming tools) It is one reason forus to publish the pictures used in our experiment (seeFigure 7) Price et al (1996) found that the areas involvedin the naming part of the process were in the leftperisylvian area left anterior insula and right cerebellumDronkers (1996) in a study of 25 stroke patients witharticulatory motor planning decits found the left insulain particular its precentral gyrus damaged in all of thesepatients (and in none of the control patients) RecentlyAbdullaev and Posner (1997) reported a high-density (64electrode) event-related potential (ERP) study of thewell-known (but ill-understood) word generation task Inthis time-course study of semantically generated wordsa left temporo-parietal activation probably involvingWernickersquos area was observed around 650 msec afterpresentation of the stimulus word (in addition to theearly left frontal response that is so typical for the verbgeneration task) These and other ndings provide asmany hypotheses about the cerebral areas involved inthe various operations that mediate between the visualpresentation of an object and the articulation of theobjectrsquos name
There are essentially two ways in which the corticallocalization of component processes in picture namingcan be pursued by means of MEG The rst one is tocarefully map the psychological time course of picturenaming (ie its staging in time) onto the speakerrsquos re-corded magnetic brain activity over time during picture
naming In other words one could try to localize thecortical areas that are active during successive stage ofprocessing The second one is more focused but alsomore risky One can try to affect the duration of a singlestage of processing (for instance phonological wordencoding) by means of an independent experimentalvariable and observe which sensors register a covariatemagnetic response It is more risky because factors af-fecting a single processing stage in picture naming usu-ally have relatively small reaction time effects Bothmethods source analysis and the ldquosingle factorrdquo methodrequire detailed knowledge about the time course ofpicture naming and the processing stages involved inthat task Fortunately the psychology of picture namingis well developed
A Process Analysis of Picture Naming
Any sophisticated use of picture naming in localizationstudies must be based on a processing model of the taskitself Luckily picture naming has been the subject ofmyriad experimental studies (see Glaser 1992 for a re-view) and we can now with some condence distin-guish the stages in the naming of a picture (see Figure 1)outlined in the following subsections
Recognizing the Visual Object
During this stage a visual representation of the objectmdashwhich we will call the perceptmdashis computed from thevisual image The percept is entirely alinguistic but doesinvolve such abstract properties as the objectrsquos typicalsize (an elephant is big and a mouse is small) and color(cf Hart amp Gordon 1992) The object representation alsohas a particular orientation with respect to the perceiverThere is a ldquopoint of viewrdquo (Buumllthoff amp Edelmann 1992)
Accessing a Lexical Concept
To refer to the percept the speaker will activate anappropriate lexical concept Which concept is appropri-ate depends on the speakerrsquos perspective in the commu-nicative situation (Levelt 1996) The same percept canbe conceptualized as a VEHICLE a CAR a MERCEDES etc Butthere tend to be ldquobasic levelrdquo conceptualizations such asCAR APPLE FORK COAT VIOLIN (Rosch Mervis GrayJohnson amp Boyes-Braem 1976) A subject is easily in-duced to operate on this basic level in picture namingLexical concepts can be conceived of as nodes in asemantic network with labeled arcs (Roelofs 1992) Forinstance VIOLIN has an is-a relation to MUSICAL INSTRU-
MENT a has-part relation to STRING etc There is alwayssome activation spreading from the target concept tosemantically related concepts
554 Journal of Cognitive Neuroscience Volume 10 Number 5
Selecting the Lemma
When a lexical concept such as VIOLIN is active activa-tion spreads to the corresponding word in the mentallexicon or more specically to the correspondinglemma This is the wordrsquos syntactic representation in-volving the wordrsquos syntactic category (noun for violinor canoe) its gender if it is a nounmdashin gender-markinglanguages (in German feminine for Geige neuter forKanu) and its subcategorization if it is a verb (transitiveintransitive etc) These are the properties of a word thatare essential for grammatical encoding for example em-bedding it in the appropriate syntactic environment(Geige will need a feminine article become head of a
noun phrase etc) All active lexical concepts spreadactivation to their lemmas in the lexicon but eventuallyonly one lemma will get selected following a simplemathematical rule At any one moment the probability ofselecting the target lemma is the ratio of its activationand the total activation of all lemmas involved Thisldquohazard raterdquo determines the expected selection duration(Roelofs 1992)
Accessing the Morpheme(s) and Generating thePhonological Word
The selected lemma (and only the selected lemma)spreads its activation to its word form representation ormorpheme(s) The morpheme(s) represents the wordrsquosphonological shape in particular its segments (for ltca-noegt these are the phonemes k n u) and itsmetrical structure (for ltcanoegt it is the iambic foot ssrsquo)These two sources of information are independently re-trieved (Levelt 1992a Roelofs amp Meyer 1997) and usedto incrementally create a so-called ldquophonological wordrdquo(w) a fully syllabied representation of the word such as
w
s srsquo
k n u
In the process the phonemic segments are one by one(from left to right) associated with the metrical frame ofthe phonological word successively creating the wordrsquosphonological syllables (k and nu in the example)
Retrieving Syllabic Gestural Scores
As soon as a syllable such as nu has been pro-grammed its gestural score is retrieved from the mentalsyllabary (Levelt amp Wheeldon 1994) The syllabary is therepository of abstract motor representations for the fre-quently used syllables in the language These syllabicscores are retrieved one by one as the phonologicalsyllables become incrementally available Accessing thesyllabary is again due to activation spreading In theexample syllable-initial n activates all syllables with nas onset or offset and all syllables with the vowel u asnucleus The target syllablersquos hazard rate determines theexpected selection duration (Roelofs 1997)
Articulation
The wordrsquos articulation is initiated as soon as all of itssyllabic scores have been retrieved (Levelt amp Wheeldon1994) The articulatory system computes least-effort so-lutions for the execution of the (still rather abstract)gestural scores These solutions provide the motor in-structions for the respiratory the laryngeal and the su-
Figure 1 A stage model of picture naming Preparing a name pro-ceeds through stages of visual processing activating a lexical con-cept word selection phonological encoding phonetic encoding andthe initiation of articulation Self-monitoring refers to phonologicalcodes and overt speech
Levelt et al 555
pralaryngeal systems involved in uent articulation ofspeech (Levelt 1989)
Self-Monitoring
The overt speech is self-perceived which activates thespeakerrsquos speech comprehension system In addition thespeaker can monitor a prearticulatory internal speechcode probably the phonological word representation(Wheeldon amp Levelt 1995) If any communicatively seri-ous error is detected the speaker can halt and make aself-repair (Levelt 1983) This ldquolaterdquo feedback loop isprobably the only one involved in the system in spite ofclaims to the contrary in the connectionist literature(Levelt 1992b MacKay 1992)
A comprehensive review of this stage theory with allthe experimental evidence on which it is based can befound in Levelt Roelofs and Meyer (in press)
The Timing of Stages
Apart from this late self-perceiving feedback picturenaming is completely or almost completely a feedfor-ward process This makes picture naming particularlyattractive in the study of the cortical localization oflanguage-processing components In analyzing functionalimaging data with high temporal resolution such asevoked MEG responses one can assume that each com-ponent is active just once We are of course aware ofdiverging claims in the literature (among them Dell1986 Dell Schwartz Martin Saffran amp Gagnon 1997Dell amp OrsquoSeaghdha 1991 Humphreys Riddock amp Quin-lan 1988 1995 Martin Gagnon Schwartz Dell amp Saffran1996 Peterson amp Savoy 1998) but there is an almostuniversal agreement that picture naming is a staged proc-ess where feedback if any is so local that there will notbe measurable activation-deactivation-reactivation pat-terns for any processing component involved
In the MEG experiments to be reported the averagepicture naming latency (ie from picture onset to theonset of articulation) was 538 msec How is this timedistributed over the above processing stages The experi-mental data in the literature do not allow us to distin-guish between the rst two stages (ie deriving thepercept and accessing the lexical concept) The mostrecent estimate is from Thorpe Fize and Marlot (1996)In an ERP gono-go study in which subjects had tocategorize a novel picture as displaying an animal or notthe authors showed that a frontal negativity developedaround 150 msec post-picture onset only on the no-gotrials At that moment apparently the subject recognizedthat the picture contained no animal This picture recog-nition time estimate agrees reasonably well with esti-mates based on the data in Potter (1983) the conceptbeginning to be accessed some 100 msec after pictureonset One should keep in mind though that muchdepends on the quality of the picture The pictures in
our MEG experiment were high-quality line drawings(see Figure 7) Thorpe et al used color photographsprobably also high-quality An additional reason for as-suming that Thorpe et alrsquos estimate holds well for ourmaterials is the following Their average animatenessdecision time was 445 msec One of our pretests (seeldquoMethodsrdquo section) was an objectnon-object decision onthe experimental picture Average object decision timewas 479 msec Most of our pictures had earlier beenused in the Jescheniak and Levelt (1994) experimentsOne of these used a picture recognition task The subjectsaw a word on the screen (for instance dog) then sawthe picture and had to press a ldquoyesrdquo button if the twoagreed a task very close to Thorpe et alrsquos The averageldquoyesrdquo response took 439 msec Because dog (etc) andThorpersquos et alrsquos ldquoanimaterdquo are lexical concepts it is likelythat the 150-msec estimate includes accessing the lexicalconcept (ie it probably covers the rst two stages ofthe naming process)
Turning now to lexical access proper a rst estimateof the duration of lemma selection in naming tasks ispresented in Levelt et al (1991) 115 msec That numberis derived from data obtained in a picture naminglexicaldecision experiment The primary task in the experimentwas picture naming But on one-third of the trials thesubject heard an auditory probe a word or a nonwordIt could be presented at one of three moments afterpicture onset but before the initiation of naming (ie atthree different stimulus onset asynchronies or SOAs) Onthese trials the subject had to perform a secondary taskbefore naming the picture pushing a ldquoyesrdquo or ldquonordquo buttondepending on whether the auditory probe was a wordor not On all relevant items the probe was a word andit could be semantically or phonologically related to thetarget picture or be unrelated to it (for instance if thepicture was one of a sheep the probe word could begoat sheet or house) At some SOAs the lexical decisionlatency to a semantically related probe (goat in theexample) was slower than to an unrelated probe (suchas house in the example) The obvious interpretation forthat nding is that the picture naming had advanced sofar at that SOA that the target lemma was active andcould begin interfering with the probe lemma The com-putational model that provided an optimal t for theselexical decision data yielded the mentioned 115-msecestimate for lemma access
This estimate is in good agreement with the oneprovided by the computational model of lemma selec-tion in Roelofs (1992) Lemma access was the very targetof that paper Roelofs used a pictureword interferencetask in his experiments Subjects named pictures andnaming latency was the dependent measure in theseexperiments At some moment during the task (ie atdifferent SOAs) a visual distracter word was ashed inthe picture and the subject was instructed to ignoreit The distracter could be semantically related to thetarget picture name or unrelated to it (just as in the
556 Journal of Cognitive Neuroscience Volume 10 Number 5
above experiment) Typically naming latencies wereslower when the distracter was semantically related(such as goat when a sheep was displayed) than whenit was an unrelated distracter (such as house) at leastwhen picture and distracter presentation were closelysynchronized The computational model based on thesedata sets gives lemma selection durations in the rangeof 100 to 150 msec This nally corresponds wellwith what Potter (personal communication) derivesfrom the data in Potter and Faulconer (1975) an estimateof 150 msec
Duration estimates for the next stage of lexical encod-ing word form access and phonological encoding canbe derived from the following three sources The rstone is Roelofsrsquos (1997) WEAVER model of phonologicalencoding It is based on myriad pictureword interfer-ence experiments in which auditory phonological dis-tracters were presented during picture naming Based onthese data Roelofs gives an estimate of 265 msec for theinterval between lemma selection and accessing thesyllable score of a monosyllabic word Because this in-volves selecting the syllable node (although not ldquounpack-ingrdquo) it the time for attaching phonemic segments to themetrical frame should be less An estimate for the lattercomes from a second study by Wheeldon and Levelt(1994) In their experiments the subject was providedwith a target phoneme for instance an l or an r The(Dutch) subject would then hear an English word (forexample hitchhiker) and decided whether the Dutchtranslation of that word (lifter in the example case)contained the target phoneme The Dutch word was notovertly pronounced The dependent measure was thepush button (yes) latency For bisyllabic items such aslifter the average difference in phoneme monitoringspeed between the word initial and the word nal pho-neme (l and r for this example) turned out to beabout 125 msec This can be taken as an estimate of theduration to (internally) encode the whole wordphonologically Finally van Turennout Hagoort andBrown (1997) in an ERP study not unlike Thorpe et alrsquoswhere word-initial and word-nal phonemes providedgono-go cues for a semantic picture categorizationcould estimate a duration of about 120 msec forphonological encoding
Left over then are phonetic encoding and the initia-tion of articulation If the above estimates are approxi-mately correct subtraction can provide an estimate forthese nal stages For an average naming latency of 538msec the distribution over the stages would roughly beas follows visual processing plus accessing the lexicalconcept 150 msec lemma selection 125 msec phono-logical encoding 125 msec and phonetic encoding andinitiation of articulation 138 msec This latter numberagrees well with a nal estimate that we have Leveltet al (1991) provide a model estimate for the durationof phonological plus phonetic encoding based on theirpicture naminglexical decision expriments it is 270
msec If we subtract 125 msec for phonological encod-ing we are left with 145 msec for the duration ofphonetic encoding
In the present study we will group activation dataaccording to the following time windows post-pictureonset
0ndash150 msec Visual processing and accessing the lexical concept
150ndash275 msec Lemma selection275ndash400 msec Phonological encoding400ndash600 msec Phonetic and articulatory processing
The last window extends somewhat beyond the averagenaming response to include most of the factual namingresponses (but of course for each response only pre-onset activation data were analyzed) Although thesewindows capture all of the data in the above meta-analy-sis these data do not allow us to estimate windowvariances However our selection of experimental mate-rials (see ldquoMethodsrdquo) aimed at minimizing variances inobject recognition and in total naming latency
Singling Out Phonological Encoding
As mentioned previously one can in principle study thecerebral localization of a single processing stage by in-troducing an independent variable that specically af-fects the duration of that stage Any dependent effect inthe MEG trace should be due to the correspondingprocessing component In the present study we decidedto try this out on the stage of phonological encodingJescheniak and Levelt (1994) discovered that the well-known word frequency effect in picture naming(Oldeld amp Wingeld 1965) is due to accessing thewordrsquos form Pictures whose names are highly frequent(HF) in the speakerrsquos language (such as boat) are namedfaster (by some 30 to 50 msec) than pictures whosenames have low frequency (LF) (such as broom) Theauthors could show that this is not due to forming thepercept or to retrieving the lemma or the wordrsquos sylla-bles The effect arises in accessing the wordrsquos form infor-mation
This fact can now in principle be used to localize theprocess of word form access in the brain if one hasaccess to a temporal high-resolution method such asMEG The idea is this (see Figure 2) When the inde-pendent variable in the picture naming task is targetword frequency (ie HF versus LF picture names) weshould expect that the initial processing stages up to thestage of form access are the same for HF and LF picturesBut from morpheme access on the time course for HFpictures will divert from the time course for LF picturesMore precisely (see Figure 2) the critical stage will berelatively short for the HF pictures and relatively long forthe LF pictures (the time difference dt will be about 30to 50 msec) All subsequent stages will not differ induration but will be desynchronized by dt One can now
Levelt et al 557
trace the activation pattern over the MEG sensors anddetermine whether such desynchronization arises and inparticular where it begins Inversely one can response-lock the activation patterns and determine where desyn-chronization begins running backward in time In theideal case one can ldquosqueeze inrdquo the critical process intime and place But as already mentioned 30 to 50 msecis not much and is easily drowned in experimental noise
Whatever it is worth this single factors or stage focus-ing method (or SF-method) differs essentially from thesubtraction method which is now commonly used inpositron emission tomography (PET) and functionalmagnetic resonance imaging (fMRI) studies There onetries to eliminate a stage completely here all stages arealways present There is reason to have serious doubtsabout the possibility of eliminating a stage entirely (Fris-ton et al 1996) In his classical paper Sternberg (1969)argued for a different approach the additive factorsmethod which leaves all stages intact The SF-method isa derivative of Sternbergrsquos method (see also Dehaene1996)
In the present study eight Dutch subjects repeatedlynamed 24 carefully preselected pictures 12 with LFnames and 12 with HF names while their magnetic brainactivity was recorded in the Neuromag-122 whole-headmagnetometer This was done in two identical and im-mediately adjacent sessions of six randomly dispersedrepetitions of all 24 pictures In a prerun of exactly thesame experiment but without MEG recording and ondifferent subjects a 37-msec word frequency effect hadbeen obtained (see ldquoResultsrdquo)
RESULTS
In the following we will rst in summary fashion de-scribe the unsuccessful single-factors analysis and thenturn in detail to the more revealing sourcetime analysis
Single Factors Analysis
Reaction Times
As mentioned we obtained a solid 37-msec word fre-quency effect in the pre-experiment Figure 3 shows thereaction time curves for high- and low-frequency wordresponses The word frequency effect was stable over
the 12 repetitions (see ldquoMethodsrdquo) This substantial stableword frequency effect was not replicated in the mainexperiment (run on different subjects) Across all sub-jects mean reaction times for high- and low-frequencyitems differed by only 3 msec (535 versus 538 msec)The effects per subject ranged from 20 msec favoringthe low-frequency items to 15 msec favoring the high-frequency items In analyses of variance within subjectsnone of these effects reached statistical signicanceSeparate analyses were carried out for each subject andeach of the two test sessions (ie before versus after theshort break) A signicant (p lt 005) frequency effectfavoring high-frequency items was only obtained for onesubject in one test session
Should this null effect be adduced to the new set ofsubjects we tested or to the experimental situation inthe MEG lab To nd out we retested the same subjects6 months after the main experiment on the same mate-rials but in the Max Planck Institute (ie without MEGrecording) The instructions stressed that it was impor-tant to name the targets correctlymdashspeed was of secon-dary importance Now a signicant word frequencyeffect of 33 msec was obtained for the 24 relevant test
Figure 2 The single-factorsmethod Affecting the durationof a single stage does not af-fect the cortical dynamics be-fore that stage but shifts allactivity following that stageby a constant dt
Figure 3 The word frequency effect in the naming latency for theultimate 12 HF and 12 LF stimuli as measured in the preexperimentover 12 repetitions of the test pictures
558 Journal of Cognitive Neuroscience Volume 10 Number 5
items (F1(1 7) = 1376 MSE = 3771 F2(1 22) = 968MSE = 7965 both p lt 001) Six subjects showed fre-quency effects favoring the high-frequency items in bothhalves of the experiment and two subjects showed themonly in one-half of it In other words during the retestthe subjects of the main experiments behaved verymuch like the subjects of the pretest Hence the nulleffect in the main experiment was not due to the set ofsubjects Was it due to the testing situation The mostnoticeable difference was that naming latencies werevery short in the MEG experiment on average 538 msecThese latencies were manually determined from thevisually displayed acoustic wave form In the preexperi-ment (different subjects) average naming latency hadbeen 592 msec measured by voice key Because thevoice key is triggered at some acoustic threshold voicekey latencies are always slower than latencies measuredon the wave form display this amounts to about 20 msecon average Even with this correction the latencies inthe MEG experiment were faster by 42 msec Is it thecase that our young Dutch subjects own into Helsinkifor a weekend had been so highly motivated during theMEG session that their naming speeds reached ceilinglevels obliterating any difference between HF and LFwords This is unlikely First in their post-test their aver-age voice key latency was 575 msec Implementing the20-msec correction this differs by no more than 17 msecfrom the latencies in the MEG experiment Second wetested a new set of 10 subjects who did the experimenttwice (in two sessions 1 week apart) once under strictspeed conditions and once under the instruction toavoid errors and not to worry about time Although thevoice key averages differed substantially between thesetwo conditions (542 msec versus 618 msec) the fre-quency effect was exactly the same 27 msec Notice thatthe speeded condition was faster by 16 msec than in theMEG experiment (correcting for the 20 msec) So aword frequency effect can be obtained even with veryfast naming responses In short we have no explanationfor the tragic disappearance of the effect in the MEGexperiment It did however undermine our single-factors analysis
Evoked Magnetic Fields
Although the null effect for word frequency eliminatedthe single-factors analysis as displayed in Figure 2 theremight still be a word frequency effect in the evokedmagnetic data For all subjects we visually checked theHF and LF responses recorded by the 122 sensors Thespatio-temporal patterns of evoked magnetic elds dif-fered considerably between subjects except for the rela-tively uniform exogenous visual evoked elds recordedat occipital sensors But none of the subjects showed anysystematic difference between HF and LF activation pat-terns Nevertheless we performed analyses of varianceover HF versus LF curves for all sensors and all subjects
by partitioning all curves in 50-msec windows rangingfrom 0 to 600 msec post-onset Although we found moresignicant word frequency effects than could be ex-pected by chance they were not ubiquitous and showedno interpretable patterns within or over subjects Thiswas further conrmed in the dipole source analysis (seeFigure 4)
Dipole Source Analysis
The main aim of our dipole source analysis was to relatepeak activity of distinct cortical areas to the time win-dows of the stage model of picture naming Thereforedipole source analysis was performed separately for eachsubjectrsquos data using the signals from subjectsrsquo rst test-ing session and averaged over the two frequency condi-tions (see ldquoMethodsrdquo section) Figure 4 shows theoriginal whole-head MEG responses for HF and LF wordsin one subject The data were modeled by eight equiva-lent current dipoles (ECD) identied at distinct latenciesand shown on the surface rendition of the subjectrsquos MRIThe source strengths as a function of time are plottedon the right
The source localization results over all subjects aredisplayed in Figure 5 The four time windows discussedabove correspond to four colors in the diagrams Let usgo over these windows in temporal order
Visual Processing and Accessing the Lexical Concept0 to 150 msec
The visual evoked magnetic elds were adequately mod-eled by about two to three sources in each subjectThese sources were located in the occipital cortex (yel-low sources) with a preponderance of sources in theright hemisphere Note that the model sphere for thesource analyses was tted in such a way that centralparietal and temporal areas were covered optimallyTherefore localization errors are likely to be largest inthe frontal and occipital areas Because the modeling ofvisual-perceptual activity was not the goal of this studyno further attempt was made to determine the locationof these sources more precisely The ndings do agreehowever with the occipital lobe activation adduced toobject recognition in Price et alrsquos (1996) PET study ofpicture naming
Lemma Selection 150 to 275 msec
In this time window magnetic eld patterns were morevariable The sources identied in this time frame (threeto four per subjectmdashblue dots in the second panel) werelargely found in the occipital area (50) and in theparietal and occasionally temporal areas However therewas a remarkable clustering of sources (seven of eightsubjects) in the right parietal cortex along the posteriorend of the superior temporal sulcus
Levelt et al 559
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

unlikely that a single language site performs all of thesefunctions
One would like to know how for individual subjectsthese components of the picture naming process aredistributed over the brain Only then we will begin todiscover what is uniform across speakers and what isvariable The obvious rst hunch is that the input andoutput processes in picture naming (perceiving the ob-ject articulating the word) are much less variable in theircerebral localization than the mediating processes oflexical access and of phonological encoding In particu-lar one should expect to nd occipital lobe activation ina picture naming task as well as activation in the facearea of the sensory-motor cortex But what about themore endogenous responses In a positron emissiontomography (PET) study of object naming and recogni-tion Price Moore Humphreys Frackowiak and Friston(1996) found the following areas involved with objectrecognition ventral and dorsal regions of the left middleoccipital lobe bilateral anterior temporal lobes and theleft cerebellum Martin Wiggs Ungerleider and Haxby(1996) also in a PET study found the left middle occipi-tal lobe only active in the naming of animals but not oftools This study as well as the one by DamasioGrabowski Tranel Hichwa and Damasio (1996) suggeststhat the areas involved in the semantic processing ofpictures are different for different categories of picturesto be named (but the studies disagree on what areas arespecically involved in naming tools) It is one reason forus to publish the pictures used in our experiment (seeFigure 7) Price et al (1996) found that the areas involvedin the naming part of the process were in the leftperisylvian area left anterior insula and right cerebellumDronkers (1996) in a study of 25 stroke patients witharticulatory motor planning decits found the left insulain particular its precentral gyrus damaged in all of thesepatients (and in none of the control patients) RecentlyAbdullaev and Posner (1997) reported a high-density (64electrode) event-related potential (ERP) study of thewell-known (but ill-understood) word generation task Inthis time-course study of semantically generated wordsa left temporo-parietal activation probably involvingWernickersquos area was observed around 650 msec afterpresentation of the stimulus word (in addition to theearly left frontal response that is so typical for the verbgeneration task) These and other ndings provide asmany hypotheses about the cerebral areas involved inthe various operations that mediate between the visualpresentation of an object and the articulation of theobjectrsquos name
There are essentially two ways in which the corticallocalization of component processes in picture namingcan be pursued by means of MEG The rst one is tocarefully map the psychological time course of picturenaming (ie its staging in time) onto the speakerrsquos re-corded magnetic brain activity over time during picture
naming In other words one could try to localize thecortical areas that are active during successive stage ofprocessing The second one is more focused but alsomore risky One can try to affect the duration of a singlestage of processing (for instance phonological wordencoding) by means of an independent experimentalvariable and observe which sensors register a covariatemagnetic response It is more risky because factors af-fecting a single processing stage in picture naming usu-ally have relatively small reaction time effects Bothmethods source analysis and the ldquosingle factorrdquo methodrequire detailed knowledge about the time course ofpicture naming and the processing stages involved inthat task Fortunately the psychology of picture namingis well developed
A Process Analysis of Picture Naming
Any sophisticated use of picture naming in localizationstudies must be based on a processing model of the taskitself Luckily picture naming has been the subject ofmyriad experimental studies (see Glaser 1992 for a re-view) and we can now with some condence distin-guish the stages in the naming of a picture (see Figure 1)outlined in the following subsections
Recognizing the Visual Object
During this stage a visual representation of the objectmdashwhich we will call the perceptmdashis computed from thevisual image The percept is entirely alinguistic but doesinvolve such abstract properties as the objectrsquos typicalsize (an elephant is big and a mouse is small) and color(cf Hart amp Gordon 1992) The object representation alsohas a particular orientation with respect to the perceiverThere is a ldquopoint of viewrdquo (Buumllthoff amp Edelmann 1992)
Accessing a Lexical Concept
To refer to the percept the speaker will activate anappropriate lexical concept Which concept is appropri-ate depends on the speakerrsquos perspective in the commu-nicative situation (Levelt 1996) The same percept canbe conceptualized as a VEHICLE a CAR a MERCEDES etc Butthere tend to be ldquobasic levelrdquo conceptualizations such asCAR APPLE FORK COAT VIOLIN (Rosch Mervis GrayJohnson amp Boyes-Braem 1976) A subject is easily in-duced to operate on this basic level in picture namingLexical concepts can be conceived of as nodes in asemantic network with labeled arcs (Roelofs 1992) Forinstance VIOLIN has an is-a relation to MUSICAL INSTRU-
MENT a has-part relation to STRING etc There is alwayssome activation spreading from the target concept tosemantically related concepts
554 Journal of Cognitive Neuroscience Volume 10 Number 5
Selecting the Lemma
When a lexical concept such as VIOLIN is active activa-tion spreads to the corresponding word in the mentallexicon or more specically to the correspondinglemma This is the wordrsquos syntactic representation in-volving the wordrsquos syntactic category (noun for violinor canoe) its gender if it is a nounmdashin gender-markinglanguages (in German feminine for Geige neuter forKanu) and its subcategorization if it is a verb (transitiveintransitive etc) These are the properties of a word thatare essential for grammatical encoding for example em-bedding it in the appropriate syntactic environment(Geige will need a feminine article become head of a
noun phrase etc) All active lexical concepts spreadactivation to their lemmas in the lexicon but eventuallyonly one lemma will get selected following a simplemathematical rule At any one moment the probability ofselecting the target lemma is the ratio of its activationand the total activation of all lemmas involved Thisldquohazard raterdquo determines the expected selection duration(Roelofs 1992)
Accessing the Morpheme(s) and Generating thePhonological Word
The selected lemma (and only the selected lemma)spreads its activation to its word form representation ormorpheme(s) The morpheme(s) represents the wordrsquosphonological shape in particular its segments (for ltca-noegt these are the phonemes k n u) and itsmetrical structure (for ltcanoegt it is the iambic foot ssrsquo)These two sources of information are independently re-trieved (Levelt 1992a Roelofs amp Meyer 1997) and usedto incrementally create a so-called ldquophonological wordrdquo(w) a fully syllabied representation of the word such as
w
s srsquo
k n u
In the process the phonemic segments are one by one(from left to right) associated with the metrical frame ofthe phonological word successively creating the wordrsquosphonological syllables (k and nu in the example)
Retrieving Syllabic Gestural Scores
As soon as a syllable such as nu has been pro-grammed its gestural score is retrieved from the mentalsyllabary (Levelt amp Wheeldon 1994) The syllabary is therepository of abstract motor representations for the fre-quently used syllables in the language These syllabicscores are retrieved one by one as the phonologicalsyllables become incrementally available Accessing thesyllabary is again due to activation spreading In theexample syllable-initial n activates all syllables with nas onset or offset and all syllables with the vowel u asnucleus The target syllablersquos hazard rate determines theexpected selection duration (Roelofs 1997)
Articulation
The wordrsquos articulation is initiated as soon as all of itssyllabic scores have been retrieved (Levelt amp Wheeldon1994) The articulatory system computes least-effort so-lutions for the execution of the (still rather abstract)gestural scores These solutions provide the motor in-structions for the respiratory the laryngeal and the su-
Figure 1 A stage model of picture naming Preparing a name pro-ceeds through stages of visual processing activating a lexical con-cept word selection phonological encoding phonetic encoding andthe initiation of articulation Self-monitoring refers to phonologicalcodes and overt speech
Levelt et al 555
pralaryngeal systems involved in uent articulation ofspeech (Levelt 1989)
Self-Monitoring
The overt speech is self-perceived which activates thespeakerrsquos speech comprehension system In addition thespeaker can monitor a prearticulatory internal speechcode probably the phonological word representation(Wheeldon amp Levelt 1995) If any communicatively seri-ous error is detected the speaker can halt and make aself-repair (Levelt 1983) This ldquolaterdquo feedback loop isprobably the only one involved in the system in spite ofclaims to the contrary in the connectionist literature(Levelt 1992b MacKay 1992)
A comprehensive review of this stage theory with allthe experimental evidence on which it is based can befound in Levelt Roelofs and Meyer (in press)
The Timing of Stages
Apart from this late self-perceiving feedback picturenaming is completely or almost completely a feedfor-ward process This makes picture naming particularlyattractive in the study of the cortical localization oflanguage-processing components In analyzing functionalimaging data with high temporal resolution such asevoked MEG responses one can assume that each com-ponent is active just once We are of course aware ofdiverging claims in the literature (among them Dell1986 Dell Schwartz Martin Saffran amp Gagnon 1997Dell amp OrsquoSeaghdha 1991 Humphreys Riddock amp Quin-lan 1988 1995 Martin Gagnon Schwartz Dell amp Saffran1996 Peterson amp Savoy 1998) but there is an almostuniversal agreement that picture naming is a staged proc-ess where feedback if any is so local that there will notbe measurable activation-deactivation-reactivation pat-terns for any processing component involved
In the MEG experiments to be reported the averagepicture naming latency (ie from picture onset to theonset of articulation) was 538 msec How is this timedistributed over the above processing stages The experi-mental data in the literature do not allow us to distin-guish between the rst two stages (ie deriving thepercept and accessing the lexical concept) The mostrecent estimate is from Thorpe Fize and Marlot (1996)In an ERP gono-go study in which subjects had tocategorize a novel picture as displaying an animal or notthe authors showed that a frontal negativity developedaround 150 msec post-picture onset only on the no-gotrials At that moment apparently the subject recognizedthat the picture contained no animal This picture recog-nition time estimate agrees reasonably well with esti-mates based on the data in Potter (1983) the conceptbeginning to be accessed some 100 msec after pictureonset One should keep in mind though that muchdepends on the quality of the picture The pictures in
our MEG experiment were high-quality line drawings(see Figure 7) Thorpe et al used color photographsprobably also high-quality An additional reason for as-suming that Thorpe et alrsquos estimate holds well for ourmaterials is the following Their average animatenessdecision time was 445 msec One of our pretests (seeldquoMethodsrdquo section) was an objectnon-object decision onthe experimental picture Average object decision timewas 479 msec Most of our pictures had earlier beenused in the Jescheniak and Levelt (1994) experimentsOne of these used a picture recognition task The subjectsaw a word on the screen (for instance dog) then sawthe picture and had to press a ldquoyesrdquo button if the twoagreed a task very close to Thorpe et alrsquos The averageldquoyesrdquo response took 439 msec Because dog (etc) andThorpersquos et alrsquos ldquoanimaterdquo are lexical concepts it is likelythat the 150-msec estimate includes accessing the lexicalconcept (ie it probably covers the rst two stages ofthe naming process)
Turning now to lexical access proper a rst estimateof the duration of lemma selection in naming tasks ispresented in Levelt et al (1991) 115 msec That numberis derived from data obtained in a picture naminglexicaldecision experiment The primary task in the experimentwas picture naming But on one-third of the trials thesubject heard an auditory probe a word or a nonwordIt could be presented at one of three moments afterpicture onset but before the initiation of naming (ie atthree different stimulus onset asynchronies or SOAs) Onthese trials the subject had to perform a secondary taskbefore naming the picture pushing a ldquoyesrdquo or ldquonordquo buttondepending on whether the auditory probe was a wordor not On all relevant items the probe was a word andit could be semantically or phonologically related to thetarget picture or be unrelated to it (for instance if thepicture was one of a sheep the probe word could begoat sheet or house) At some SOAs the lexical decisionlatency to a semantically related probe (goat in theexample) was slower than to an unrelated probe (suchas house in the example) The obvious interpretation forthat nding is that the picture naming had advanced sofar at that SOA that the target lemma was active andcould begin interfering with the probe lemma The com-putational model that provided an optimal t for theselexical decision data yielded the mentioned 115-msecestimate for lemma access
This estimate is in good agreement with the oneprovided by the computational model of lemma selec-tion in Roelofs (1992) Lemma access was the very targetof that paper Roelofs used a pictureword interferencetask in his experiments Subjects named pictures andnaming latency was the dependent measure in theseexperiments At some moment during the task (ie atdifferent SOAs) a visual distracter word was ashed inthe picture and the subject was instructed to ignoreit The distracter could be semantically related to thetarget picture name or unrelated to it (just as in the
556 Journal of Cognitive Neuroscience Volume 10 Number 5
above experiment) Typically naming latencies wereslower when the distracter was semantically related(such as goat when a sheep was displayed) than whenit was an unrelated distracter (such as house) at leastwhen picture and distracter presentation were closelysynchronized The computational model based on thesedata sets gives lemma selection durations in the rangeof 100 to 150 msec This nally corresponds wellwith what Potter (personal communication) derivesfrom the data in Potter and Faulconer (1975) an estimateof 150 msec
Duration estimates for the next stage of lexical encod-ing word form access and phonological encoding canbe derived from the following three sources The rstone is Roelofsrsquos (1997) WEAVER model of phonologicalencoding It is based on myriad pictureword interfer-ence experiments in which auditory phonological dis-tracters were presented during picture naming Based onthese data Roelofs gives an estimate of 265 msec for theinterval between lemma selection and accessing thesyllable score of a monosyllabic word Because this in-volves selecting the syllable node (although not ldquounpack-ingrdquo) it the time for attaching phonemic segments to themetrical frame should be less An estimate for the lattercomes from a second study by Wheeldon and Levelt(1994) In their experiments the subject was providedwith a target phoneme for instance an l or an r The(Dutch) subject would then hear an English word (forexample hitchhiker) and decided whether the Dutchtranslation of that word (lifter in the example case)contained the target phoneme The Dutch word was notovertly pronounced The dependent measure was thepush button (yes) latency For bisyllabic items such aslifter the average difference in phoneme monitoringspeed between the word initial and the word nal pho-neme (l and r for this example) turned out to beabout 125 msec This can be taken as an estimate of theduration to (internally) encode the whole wordphonologically Finally van Turennout Hagoort andBrown (1997) in an ERP study not unlike Thorpe et alrsquoswhere word-initial and word-nal phonemes providedgono-go cues for a semantic picture categorizationcould estimate a duration of about 120 msec forphonological encoding
Left over then are phonetic encoding and the initia-tion of articulation If the above estimates are approxi-mately correct subtraction can provide an estimate forthese nal stages For an average naming latency of 538msec the distribution over the stages would roughly beas follows visual processing plus accessing the lexicalconcept 150 msec lemma selection 125 msec phono-logical encoding 125 msec and phonetic encoding andinitiation of articulation 138 msec This latter numberagrees well with a nal estimate that we have Leveltet al (1991) provide a model estimate for the durationof phonological plus phonetic encoding based on theirpicture naminglexical decision expriments it is 270
msec If we subtract 125 msec for phonological encod-ing we are left with 145 msec for the duration ofphonetic encoding
In the present study we will group activation dataaccording to the following time windows post-pictureonset
0ndash150 msec Visual processing and accessing the lexical concept
150ndash275 msec Lemma selection275ndash400 msec Phonological encoding400ndash600 msec Phonetic and articulatory processing
The last window extends somewhat beyond the averagenaming response to include most of the factual namingresponses (but of course for each response only pre-onset activation data were analyzed) Although thesewindows capture all of the data in the above meta-analy-sis these data do not allow us to estimate windowvariances However our selection of experimental mate-rials (see ldquoMethodsrdquo) aimed at minimizing variances inobject recognition and in total naming latency
Singling Out Phonological Encoding
As mentioned previously one can in principle study thecerebral localization of a single processing stage by in-troducing an independent variable that specically af-fects the duration of that stage Any dependent effect inthe MEG trace should be due to the correspondingprocessing component In the present study we decidedto try this out on the stage of phonological encodingJescheniak and Levelt (1994) discovered that the well-known word frequency effect in picture naming(Oldeld amp Wingeld 1965) is due to accessing thewordrsquos form Pictures whose names are highly frequent(HF) in the speakerrsquos language (such as boat) are namedfaster (by some 30 to 50 msec) than pictures whosenames have low frequency (LF) (such as broom) Theauthors could show that this is not due to forming thepercept or to retrieving the lemma or the wordrsquos sylla-bles The effect arises in accessing the wordrsquos form infor-mation
This fact can now in principle be used to localize theprocess of word form access in the brain if one hasaccess to a temporal high-resolution method such asMEG The idea is this (see Figure 2) When the inde-pendent variable in the picture naming task is targetword frequency (ie HF versus LF picture names) weshould expect that the initial processing stages up to thestage of form access are the same for HF and LF picturesBut from morpheme access on the time course for HFpictures will divert from the time course for LF picturesMore precisely (see Figure 2) the critical stage will berelatively short for the HF pictures and relatively long forthe LF pictures (the time difference dt will be about 30to 50 msec) All subsequent stages will not differ induration but will be desynchronized by dt One can now
Levelt et al 557
trace the activation pattern over the MEG sensors anddetermine whether such desynchronization arises and inparticular where it begins Inversely one can response-lock the activation patterns and determine where desyn-chronization begins running backward in time In theideal case one can ldquosqueeze inrdquo the critical process intime and place But as already mentioned 30 to 50 msecis not much and is easily drowned in experimental noise
Whatever it is worth this single factors or stage focus-ing method (or SF-method) differs essentially from thesubtraction method which is now commonly used inpositron emission tomography (PET) and functionalmagnetic resonance imaging (fMRI) studies There onetries to eliminate a stage completely here all stages arealways present There is reason to have serious doubtsabout the possibility of eliminating a stage entirely (Fris-ton et al 1996) In his classical paper Sternberg (1969)argued for a different approach the additive factorsmethod which leaves all stages intact The SF-method isa derivative of Sternbergrsquos method (see also Dehaene1996)
In the present study eight Dutch subjects repeatedlynamed 24 carefully preselected pictures 12 with LFnames and 12 with HF names while their magnetic brainactivity was recorded in the Neuromag-122 whole-headmagnetometer This was done in two identical and im-mediately adjacent sessions of six randomly dispersedrepetitions of all 24 pictures In a prerun of exactly thesame experiment but without MEG recording and ondifferent subjects a 37-msec word frequency effect hadbeen obtained (see ldquoResultsrdquo)
RESULTS
In the following we will rst in summary fashion de-scribe the unsuccessful single-factors analysis and thenturn in detail to the more revealing sourcetime analysis
Single Factors Analysis
Reaction Times
As mentioned we obtained a solid 37-msec word fre-quency effect in the pre-experiment Figure 3 shows thereaction time curves for high- and low-frequency wordresponses The word frequency effect was stable over
the 12 repetitions (see ldquoMethodsrdquo) This substantial stableword frequency effect was not replicated in the mainexperiment (run on different subjects) Across all sub-jects mean reaction times for high- and low-frequencyitems differed by only 3 msec (535 versus 538 msec)The effects per subject ranged from 20 msec favoringthe low-frequency items to 15 msec favoring the high-frequency items In analyses of variance within subjectsnone of these effects reached statistical signicanceSeparate analyses were carried out for each subject andeach of the two test sessions (ie before versus after theshort break) A signicant (p lt 005) frequency effectfavoring high-frequency items was only obtained for onesubject in one test session
Should this null effect be adduced to the new set ofsubjects we tested or to the experimental situation inthe MEG lab To nd out we retested the same subjects6 months after the main experiment on the same mate-rials but in the Max Planck Institute (ie without MEGrecording) The instructions stressed that it was impor-tant to name the targets correctlymdashspeed was of secon-dary importance Now a signicant word frequencyeffect of 33 msec was obtained for the 24 relevant test
Figure 2 The single-factorsmethod Affecting the durationof a single stage does not af-fect the cortical dynamics be-fore that stage but shifts allactivity following that stageby a constant dt
Figure 3 The word frequency effect in the naming latency for theultimate 12 HF and 12 LF stimuli as measured in the preexperimentover 12 repetitions of the test pictures
558 Journal of Cognitive Neuroscience Volume 10 Number 5
items (F1(1 7) = 1376 MSE = 3771 F2(1 22) = 968MSE = 7965 both p lt 001) Six subjects showed fre-quency effects favoring the high-frequency items in bothhalves of the experiment and two subjects showed themonly in one-half of it In other words during the retestthe subjects of the main experiments behaved verymuch like the subjects of the pretest Hence the nulleffect in the main experiment was not due to the set ofsubjects Was it due to the testing situation The mostnoticeable difference was that naming latencies werevery short in the MEG experiment on average 538 msecThese latencies were manually determined from thevisually displayed acoustic wave form In the preexperi-ment (different subjects) average naming latency hadbeen 592 msec measured by voice key Because thevoice key is triggered at some acoustic threshold voicekey latencies are always slower than latencies measuredon the wave form display this amounts to about 20 msecon average Even with this correction the latencies inthe MEG experiment were faster by 42 msec Is it thecase that our young Dutch subjects own into Helsinkifor a weekend had been so highly motivated during theMEG session that their naming speeds reached ceilinglevels obliterating any difference between HF and LFwords This is unlikely First in their post-test their aver-age voice key latency was 575 msec Implementing the20-msec correction this differs by no more than 17 msecfrom the latencies in the MEG experiment Second wetested a new set of 10 subjects who did the experimenttwice (in two sessions 1 week apart) once under strictspeed conditions and once under the instruction toavoid errors and not to worry about time Although thevoice key averages differed substantially between thesetwo conditions (542 msec versus 618 msec) the fre-quency effect was exactly the same 27 msec Notice thatthe speeded condition was faster by 16 msec than in theMEG experiment (correcting for the 20 msec) So aword frequency effect can be obtained even with veryfast naming responses In short we have no explanationfor the tragic disappearance of the effect in the MEGexperiment It did however undermine our single-factors analysis
Evoked Magnetic Fields
Although the null effect for word frequency eliminatedthe single-factors analysis as displayed in Figure 2 theremight still be a word frequency effect in the evokedmagnetic data For all subjects we visually checked theHF and LF responses recorded by the 122 sensors Thespatio-temporal patterns of evoked magnetic elds dif-fered considerably between subjects except for the rela-tively uniform exogenous visual evoked elds recordedat occipital sensors But none of the subjects showed anysystematic difference between HF and LF activation pat-terns Nevertheless we performed analyses of varianceover HF versus LF curves for all sensors and all subjects
by partitioning all curves in 50-msec windows rangingfrom 0 to 600 msec post-onset Although we found moresignicant word frequency effects than could be ex-pected by chance they were not ubiquitous and showedno interpretable patterns within or over subjects Thiswas further conrmed in the dipole source analysis (seeFigure 4)
Dipole Source Analysis
The main aim of our dipole source analysis was to relatepeak activity of distinct cortical areas to the time win-dows of the stage model of picture naming Thereforedipole source analysis was performed separately for eachsubjectrsquos data using the signals from subjectsrsquo rst test-ing session and averaged over the two frequency condi-tions (see ldquoMethodsrdquo section) Figure 4 shows theoriginal whole-head MEG responses for HF and LF wordsin one subject The data were modeled by eight equiva-lent current dipoles (ECD) identied at distinct latenciesand shown on the surface rendition of the subjectrsquos MRIThe source strengths as a function of time are plottedon the right
The source localization results over all subjects aredisplayed in Figure 5 The four time windows discussedabove correspond to four colors in the diagrams Let usgo over these windows in temporal order
Visual Processing and Accessing the Lexical Concept0 to 150 msec
The visual evoked magnetic elds were adequately mod-eled by about two to three sources in each subjectThese sources were located in the occipital cortex (yel-low sources) with a preponderance of sources in theright hemisphere Note that the model sphere for thesource analyses was tted in such a way that centralparietal and temporal areas were covered optimallyTherefore localization errors are likely to be largest inthe frontal and occipital areas Because the modeling ofvisual-perceptual activity was not the goal of this studyno further attempt was made to determine the locationof these sources more precisely The ndings do agreehowever with the occipital lobe activation adduced toobject recognition in Price et alrsquos (1996) PET study ofpicture naming
Lemma Selection 150 to 275 msec
In this time window magnetic eld patterns were morevariable The sources identied in this time frame (threeto four per subjectmdashblue dots in the second panel) werelargely found in the occipital area (50) and in theparietal and occasionally temporal areas However therewas a remarkable clustering of sources (seven of eightsubjects) in the right parietal cortex along the posteriorend of the superior temporal sulcus
Levelt et al 559
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

Selecting the Lemma
When a lexical concept such as VIOLIN is active activa-tion spreads to the corresponding word in the mentallexicon or more specically to the correspondinglemma This is the wordrsquos syntactic representation in-volving the wordrsquos syntactic category (noun for violinor canoe) its gender if it is a nounmdashin gender-markinglanguages (in German feminine for Geige neuter forKanu) and its subcategorization if it is a verb (transitiveintransitive etc) These are the properties of a word thatare essential for grammatical encoding for example em-bedding it in the appropriate syntactic environment(Geige will need a feminine article become head of a
noun phrase etc) All active lexical concepts spreadactivation to their lemmas in the lexicon but eventuallyonly one lemma will get selected following a simplemathematical rule At any one moment the probability ofselecting the target lemma is the ratio of its activationand the total activation of all lemmas involved Thisldquohazard raterdquo determines the expected selection duration(Roelofs 1992)
Accessing the Morpheme(s) and Generating thePhonological Word
The selected lemma (and only the selected lemma)spreads its activation to its word form representation ormorpheme(s) The morpheme(s) represents the wordrsquosphonological shape in particular its segments (for ltca-noegt these are the phonemes k n u) and itsmetrical structure (for ltcanoegt it is the iambic foot ssrsquo)These two sources of information are independently re-trieved (Levelt 1992a Roelofs amp Meyer 1997) and usedto incrementally create a so-called ldquophonological wordrdquo(w) a fully syllabied representation of the word such as
w
s srsquo
k n u
In the process the phonemic segments are one by one(from left to right) associated with the metrical frame ofthe phonological word successively creating the wordrsquosphonological syllables (k and nu in the example)
Retrieving Syllabic Gestural Scores
As soon as a syllable such as nu has been pro-grammed its gestural score is retrieved from the mentalsyllabary (Levelt amp Wheeldon 1994) The syllabary is therepository of abstract motor representations for the fre-quently used syllables in the language These syllabicscores are retrieved one by one as the phonologicalsyllables become incrementally available Accessing thesyllabary is again due to activation spreading In theexample syllable-initial n activates all syllables with nas onset or offset and all syllables with the vowel u asnucleus The target syllablersquos hazard rate determines theexpected selection duration (Roelofs 1997)
Articulation
The wordrsquos articulation is initiated as soon as all of itssyllabic scores have been retrieved (Levelt amp Wheeldon1994) The articulatory system computes least-effort so-lutions for the execution of the (still rather abstract)gestural scores These solutions provide the motor in-structions for the respiratory the laryngeal and the su-
Figure 1 A stage model of picture naming Preparing a name pro-ceeds through stages of visual processing activating a lexical con-cept word selection phonological encoding phonetic encoding andthe initiation of articulation Self-monitoring refers to phonologicalcodes and overt speech
Levelt et al 555
pralaryngeal systems involved in uent articulation ofspeech (Levelt 1989)
Self-Monitoring
The overt speech is self-perceived which activates thespeakerrsquos speech comprehension system In addition thespeaker can monitor a prearticulatory internal speechcode probably the phonological word representation(Wheeldon amp Levelt 1995) If any communicatively seri-ous error is detected the speaker can halt and make aself-repair (Levelt 1983) This ldquolaterdquo feedback loop isprobably the only one involved in the system in spite ofclaims to the contrary in the connectionist literature(Levelt 1992b MacKay 1992)
A comprehensive review of this stage theory with allthe experimental evidence on which it is based can befound in Levelt Roelofs and Meyer (in press)
The Timing of Stages
Apart from this late self-perceiving feedback picturenaming is completely or almost completely a feedfor-ward process This makes picture naming particularlyattractive in the study of the cortical localization oflanguage-processing components In analyzing functionalimaging data with high temporal resolution such asevoked MEG responses one can assume that each com-ponent is active just once We are of course aware ofdiverging claims in the literature (among them Dell1986 Dell Schwartz Martin Saffran amp Gagnon 1997Dell amp OrsquoSeaghdha 1991 Humphreys Riddock amp Quin-lan 1988 1995 Martin Gagnon Schwartz Dell amp Saffran1996 Peterson amp Savoy 1998) but there is an almostuniversal agreement that picture naming is a staged proc-ess where feedback if any is so local that there will notbe measurable activation-deactivation-reactivation pat-terns for any processing component involved
In the MEG experiments to be reported the averagepicture naming latency (ie from picture onset to theonset of articulation) was 538 msec How is this timedistributed over the above processing stages The experi-mental data in the literature do not allow us to distin-guish between the rst two stages (ie deriving thepercept and accessing the lexical concept) The mostrecent estimate is from Thorpe Fize and Marlot (1996)In an ERP gono-go study in which subjects had tocategorize a novel picture as displaying an animal or notthe authors showed that a frontal negativity developedaround 150 msec post-picture onset only on the no-gotrials At that moment apparently the subject recognizedthat the picture contained no animal This picture recog-nition time estimate agrees reasonably well with esti-mates based on the data in Potter (1983) the conceptbeginning to be accessed some 100 msec after pictureonset One should keep in mind though that muchdepends on the quality of the picture The pictures in
our MEG experiment were high-quality line drawings(see Figure 7) Thorpe et al used color photographsprobably also high-quality An additional reason for as-suming that Thorpe et alrsquos estimate holds well for ourmaterials is the following Their average animatenessdecision time was 445 msec One of our pretests (seeldquoMethodsrdquo section) was an objectnon-object decision onthe experimental picture Average object decision timewas 479 msec Most of our pictures had earlier beenused in the Jescheniak and Levelt (1994) experimentsOne of these used a picture recognition task The subjectsaw a word on the screen (for instance dog) then sawthe picture and had to press a ldquoyesrdquo button if the twoagreed a task very close to Thorpe et alrsquos The averageldquoyesrdquo response took 439 msec Because dog (etc) andThorpersquos et alrsquos ldquoanimaterdquo are lexical concepts it is likelythat the 150-msec estimate includes accessing the lexicalconcept (ie it probably covers the rst two stages ofthe naming process)
Turning now to lexical access proper a rst estimateof the duration of lemma selection in naming tasks ispresented in Levelt et al (1991) 115 msec That numberis derived from data obtained in a picture naminglexicaldecision experiment The primary task in the experimentwas picture naming But on one-third of the trials thesubject heard an auditory probe a word or a nonwordIt could be presented at one of three moments afterpicture onset but before the initiation of naming (ie atthree different stimulus onset asynchronies or SOAs) Onthese trials the subject had to perform a secondary taskbefore naming the picture pushing a ldquoyesrdquo or ldquonordquo buttondepending on whether the auditory probe was a wordor not On all relevant items the probe was a word andit could be semantically or phonologically related to thetarget picture or be unrelated to it (for instance if thepicture was one of a sheep the probe word could begoat sheet or house) At some SOAs the lexical decisionlatency to a semantically related probe (goat in theexample) was slower than to an unrelated probe (suchas house in the example) The obvious interpretation forthat nding is that the picture naming had advanced sofar at that SOA that the target lemma was active andcould begin interfering with the probe lemma The com-putational model that provided an optimal t for theselexical decision data yielded the mentioned 115-msecestimate for lemma access
This estimate is in good agreement with the oneprovided by the computational model of lemma selec-tion in Roelofs (1992) Lemma access was the very targetof that paper Roelofs used a pictureword interferencetask in his experiments Subjects named pictures andnaming latency was the dependent measure in theseexperiments At some moment during the task (ie atdifferent SOAs) a visual distracter word was ashed inthe picture and the subject was instructed to ignoreit The distracter could be semantically related to thetarget picture name or unrelated to it (just as in the
556 Journal of Cognitive Neuroscience Volume 10 Number 5
above experiment) Typically naming latencies wereslower when the distracter was semantically related(such as goat when a sheep was displayed) than whenit was an unrelated distracter (such as house) at leastwhen picture and distracter presentation were closelysynchronized The computational model based on thesedata sets gives lemma selection durations in the rangeof 100 to 150 msec This nally corresponds wellwith what Potter (personal communication) derivesfrom the data in Potter and Faulconer (1975) an estimateof 150 msec
Duration estimates for the next stage of lexical encod-ing word form access and phonological encoding canbe derived from the following three sources The rstone is Roelofsrsquos (1997) WEAVER model of phonologicalencoding It is based on myriad pictureword interfer-ence experiments in which auditory phonological dis-tracters were presented during picture naming Based onthese data Roelofs gives an estimate of 265 msec for theinterval between lemma selection and accessing thesyllable score of a monosyllabic word Because this in-volves selecting the syllable node (although not ldquounpack-ingrdquo) it the time for attaching phonemic segments to themetrical frame should be less An estimate for the lattercomes from a second study by Wheeldon and Levelt(1994) In their experiments the subject was providedwith a target phoneme for instance an l or an r The(Dutch) subject would then hear an English word (forexample hitchhiker) and decided whether the Dutchtranslation of that word (lifter in the example case)contained the target phoneme The Dutch word was notovertly pronounced The dependent measure was thepush button (yes) latency For bisyllabic items such aslifter the average difference in phoneme monitoringspeed between the word initial and the word nal pho-neme (l and r for this example) turned out to beabout 125 msec This can be taken as an estimate of theduration to (internally) encode the whole wordphonologically Finally van Turennout Hagoort andBrown (1997) in an ERP study not unlike Thorpe et alrsquoswhere word-initial and word-nal phonemes providedgono-go cues for a semantic picture categorizationcould estimate a duration of about 120 msec forphonological encoding
Left over then are phonetic encoding and the initia-tion of articulation If the above estimates are approxi-mately correct subtraction can provide an estimate forthese nal stages For an average naming latency of 538msec the distribution over the stages would roughly beas follows visual processing plus accessing the lexicalconcept 150 msec lemma selection 125 msec phono-logical encoding 125 msec and phonetic encoding andinitiation of articulation 138 msec This latter numberagrees well with a nal estimate that we have Leveltet al (1991) provide a model estimate for the durationof phonological plus phonetic encoding based on theirpicture naminglexical decision expriments it is 270
msec If we subtract 125 msec for phonological encod-ing we are left with 145 msec for the duration ofphonetic encoding
In the present study we will group activation dataaccording to the following time windows post-pictureonset
0ndash150 msec Visual processing and accessing the lexical concept
150ndash275 msec Lemma selection275ndash400 msec Phonological encoding400ndash600 msec Phonetic and articulatory processing
The last window extends somewhat beyond the averagenaming response to include most of the factual namingresponses (but of course for each response only pre-onset activation data were analyzed) Although thesewindows capture all of the data in the above meta-analy-sis these data do not allow us to estimate windowvariances However our selection of experimental mate-rials (see ldquoMethodsrdquo) aimed at minimizing variances inobject recognition and in total naming latency
Singling Out Phonological Encoding
As mentioned previously one can in principle study thecerebral localization of a single processing stage by in-troducing an independent variable that specically af-fects the duration of that stage Any dependent effect inthe MEG trace should be due to the correspondingprocessing component In the present study we decidedto try this out on the stage of phonological encodingJescheniak and Levelt (1994) discovered that the well-known word frequency effect in picture naming(Oldeld amp Wingeld 1965) is due to accessing thewordrsquos form Pictures whose names are highly frequent(HF) in the speakerrsquos language (such as boat) are namedfaster (by some 30 to 50 msec) than pictures whosenames have low frequency (LF) (such as broom) Theauthors could show that this is not due to forming thepercept or to retrieving the lemma or the wordrsquos sylla-bles The effect arises in accessing the wordrsquos form infor-mation
This fact can now in principle be used to localize theprocess of word form access in the brain if one hasaccess to a temporal high-resolution method such asMEG The idea is this (see Figure 2) When the inde-pendent variable in the picture naming task is targetword frequency (ie HF versus LF picture names) weshould expect that the initial processing stages up to thestage of form access are the same for HF and LF picturesBut from morpheme access on the time course for HFpictures will divert from the time course for LF picturesMore precisely (see Figure 2) the critical stage will berelatively short for the HF pictures and relatively long forthe LF pictures (the time difference dt will be about 30to 50 msec) All subsequent stages will not differ induration but will be desynchronized by dt One can now
Levelt et al 557
trace the activation pattern over the MEG sensors anddetermine whether such desynchronization arises and inparticular where it begins Inversely one can response-lock the activation patterns and determine where desyn-chronization begins running backward in time In theideal case one can ldquosqueeze inrdquo the critical process intime and place But as already mentioned 30 to 50 msecis not much and is easily drowned in experimental noise
Whatever it is worth this single factors or stage focus-ing method (or SF-method) differs essentially from thesubtraction method which is now commonly used inpositron emission tomography (PET) and functionalmagnetic resonance imaging (fMRI) studies There onetries to eliminate a stage completely here all stages arealways present There is reason to have serious doubtsabout the possibility of eliminating a stage entirely (Fris-ton et al 1996) In his classical paper Sternberg (1969)argued for a different approach the additive factorsmethod which leaves all stages intact The SF-method isa derivative of Sternbergrsquos method (see also Dehaene1996)
In the present study eight Dutch subjects repeatedlynamed 24 carefully preselected pictures 12 with LFnames and 12 with HF names while their magnetic brainactivity was recorded in the Neuromag-122 whole-headmagnetometer This was done in two identical and im-mediately adjacent sessions of six randomly dispersedrepetitions of all 24 pictures In a prerun of exactly thesame experiment but without MEG recording and ondifferent subjects a 37-msec word frequency effect hadbeen obtained (see ldquoResultsrdquo)
RESULTS
In the following we will rst in summary fashion de-scribe the unsuccessful single-factors analysis and thenturn in detail to the more revealing sourcetime analysis
Single Factors Analysis
Reaction Times
As mentioned we obtained a solid 37-msec word fre-quency effect in the pre-experiment Figure 3 shows thereaction time curves for high- and low-frequency wordresponses The word frequency effect was stable over
the 12 repetitions (see ldquoMethodsrdquo) This substantial stableword frequency effect was not replicated in the mainexperiment (run on different subjects) Across all sub-jects mean reaction times for high- and low-frequencyitems differed by only 3 msec (535 versus 538 msec)The effects per subject ranged from 20 msec favoringthe low-frequency items to 15 msec favoring the high-frequency items In analyses of variance within subjectsnone of these effects reached statistical signicanceSeparate analyses were carried out for each subject andeach of the two test sessions (ie before versus after theshort break) A signicant (p lt 005) frequency effectfavoring high-frequency items was only obtained for onesubject in one test session
Should this null effect be adduced to the new set ofsubjects we tested or to the experimental situation inthe MEG lab To nd out we retested the same subjects6 months after the main experiment on the same mate-rials but in the Max Planck Institute (ie without MEGrecording) The instructions stressed that it was impor-tant to name the targets correctlymdashspeed was of secon-dary importance Now a signicant word frequencyeffect of 33 msec was obtained for the 24 relevant test
Figure 2 The single-factorsmethod Affecting the durationof a single stage does not af-fect the cortical dynamics be-fore that stage but shifts allactivity following that stageby a constant dt
Figure 3 The word frequency effect in the naming latency for theultimate 12 HF and 12 LF stimuli as measured in the preexperimentover 12 repetitions of the test pictures
558 Journal of Cognitive Neuroscience Volume 10 Number 5
items (F1(1 7) = 1376 MSE = 3771 F2(1 22) = 968MSE = 7965 both p lt 001) Six subjects showed fre-quency effects favoring the high-frequency items in bothhalves of the experiment and two subjects showed themonly in one-half of it In other words during the retestthe subjects of the main experiments behaved verymuch like the subjects of the pretest Hence the nulleffect in the main experiment was not due to the set ofsubjects Was it due to the testing situation The mostnoticeable difference was that naming latencies werevery short in the MEG experiment on average 538 msecThese latencies were manually determined from thevisually displayed acoustic wave form In the preexperi-ment (different subjects) average naming latency hadbeen 592 msec measured by voice key Because thevoice key is triggered at some acoustic threshold voicekey latencies are always slower than latencies measuredon the wave form display this amounts to about 20 msecon average Even with this correction the latencies inthe MEG experiment were faster by 42 msec Is it thecase that our young Dutch subjects own into Helsinkifor a weekend had been so highly motivated during theMEG session that their naming speeds reached ceilinglevels obliterating any difference between HF and LFwords This is unlikely First in their post-test their aver-age voice key latency was 575 msec Implementing the20-msec correction this differs by no more than 17 msecfrom the latencies in the MEG experiment Second wetested a new set of 10 subjects who did the experimenttwice (in two sessions 1 week apart) once under strictspeed conditions and once under the instruction toavoid errors and not to worry about time Although thevoice key averages differed substantially between thesetwo conditions (542 msec versus 618 msec) the fre-quency effect was exactly the same 27 msec Notice thatthe speeded condition was faster by 16 msec than in theMEG experiment (correcting for the 20 msec) So aword frequency effect can be obtained even with veryfast naming responses In short we have no explanationfor the tragic disappearance of the effect in the MEGexperiment It did however undermine our single-factors analysis
Evoked Magnetic Fields
Although the null effect for word frequency eliminatedthe single-factors analysis as displayed in Figure 2 theremight still be a word frequency effect in the evokedmagnetic data For all subjects we visually checked theHF and LF responses recorded by the 122 sensors Thespatio-temporal patterns of evoked magnetic elds dif-fered considerably between subjects except for the rela-tively uniform exogenous visual evoked elds recordedat occipital sensors But none of the subjects showed anysystematic difference between HF and LF activation pat-terns Nevertheless we performed analyses of varianceover HF versus LF curves for all sensors and all subjects
by partitioning all curves in 50-msec windows rangingfrom 0 to 600 msec post-onset Although we found moresignicant word frequency effects than could be ex-pected by chance they were not ubiquitous and showedno interpretable patterns within or over subjects Thiswas further conrmed in the dipole source analysis (seeFigure 4)
Dipole Source Analysis
The main aim of our dipole source analysis was to relatepeak activity of distinct cortical areas to the time win-dows of the stage model of picture naming Thereforedipole source analysis was performed separately for eachsubjectrsquos data using the signals from subjectsrsquo rst test-ing session and averaged over the two frequency condi-tions (see ldquoMethodsrdquo section) Figure 4 shows theoriginal whole-head MEG responses for HF and LF wordsin one subject The data were modeled by eight equiva-lent current dipoles (ECD) identied at distinct latenciesand shown on the surface rendition of the subjectrsquos MRIThe source strengths as a function of time are plottedon the right
The source localization results over all subjects aredisplayed in Figure 5 The four time windows discussedabove correspond to four colors in the diagrams Let usgo over these windows in temporal order
Visual Processing and Accessing the Lexical Concept0 to 150 msec
The visual evoked magnetic elds were adequately mod-eled by about two to three sources in each subjectThese sources were located in the occipital cortex (yel-low sources) with a preponderance of sources in theright hemisphere Note that the model sphere for thesource analyses was tted in such a way that centralparietal and temporal areas were covered optimallyTherefore localization errors are likely to be largest inthe frontal and occipital areas Because the modeling ofvisual-perceptual activity was not the goal of this studyno further attempt was made to determine the locationof these sources more precisely The ndings do agreehowever with the occipital lobe activation adduced toobject recognition in Price et alrsquos (1996) PET study ofpicture naming
Lemma Selection 150 to 275 msec
In this time window magnetic eld patterns were morevariable The sources identied in this time frame (threeto four per subjectmdashblue dots in the second panel) werelargely found in the occipital area (50) and in theparietal and occasionally temporal areas However therewas a remarkable clustering of sources (seven of eightsubjects) in the right parietal cortex along the posteriorend of the superior temporal sulcus
Levelt et al 559
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

pralaryngeal systems involved in uent articulation ofspeech (Levelt 1989)
Self-Monitoring
The overt speech is self-perceived which activates thespeakerrsquos speech comprehension system In addition thespeaker can monitor a prearticulatory internal speechcode probably the phonological word representation(Wheeldon amp Levelt 1995) If any communicatively seri-ous error is detected the speaker can halt and make aself-repair (Levelt 1983) This ldquolaterdquo feedback loop isprobably the only one involved in the system in spite ofclaims to the contrary in the connectionist literature(Levelt 1992b MacKay 1992)
A comprehensive review of this stage theory with allthe experimental evidence on which it is based can befound in Levelt Roelofs and Meyer (in press)
The Timing of Stages
Apart from this late self-perceiving feedback picturenaming is completely or almost completely a feedfor-ward process This makes picture naming particularlyattractive in the study of the cortical localization oflanguage-processing components In analyzing functionalimaging data with high temporal resolution such asevoked MEG responses one can assume that each com-ponent is active just once We are of course aware ofdiverging claims in the literature (among them Dell1986 Dell Schwartz Martin Saffran amp Gagnon 1997Dell amp OrsquoSeaghdha 1991 Humphreys Riddock amp Quin-lan 1988 1995 Martin Gagnon Schwartz Dell amp Saffran1996 Peterson amp Savoy 1998) but there is an almostuniversal agreement that picture naming is a staged proc-ess where feedback if any is so local that there will notbe measurable activation-deactivation-reactivation pat-terns for any processing component involved
In the MEG experiments to be reported the averagepicture naming latency (ie from picture onset to theonset of articulation) was 538 msec How is this timedistributed over the above processing stages The experi-mental data in the literature do not allow us to distin-guish between the rst two stages (ie deriving thepercept and accessing the lexical concept) The mostrecent estimate is from Thorpe Fize and Marlot (1996)In an ERP gono-go study in which subjects had tocategorize a novel picture as displaying an animal or notthe authors showed that a frontal negativity developedaround 150 msec post-picture onset only on the no-gotrials At that moment apparently the subject recognizedthat the picture contained no animal This picture recog-nition time estimate agrees reasonably well with esti-mates based on the data in Potter (1983) the conceptbeginning to be accessed some 100 msec after pictureonset One should keep in mind though that muchdepends on the quality of the picture The pictures in
our MEG experiment were high-quality line drawings(see Figure 7) Thorpe et al used color photographsprobably also high-quality An additional reason for as-suming that Thorpe et alrsquos estimate holds well for ourmaterials is the following Their average animatenessdecision time was 445 msec One of our pretests (seeldquoMethodsrdquo section) was an objectnon-object decision onthe experimental picture Average object decision timewas 479 msec Most of our pictures had earlier beenused in the Jescheniak and Levelt (1994) experimentsOne of these used a picture recognition task The subjectsaw a word on the screen (for instance dog) then sawthe picture and had to press a ldquoyesrdquo button if the twoagreed a task very close to Thorpe et alrsquos The averageldquoyesrdquo response took 439 msec Because dog (etc) andThorpersquos et alrsquos ldquoanimaterdquo are lexical concepts it is likelythat the 150-msec estimate includes accessing the lexicalconcept (ie it probably covers the rst two stages ofthe naming process)
Turning now to lexical access proper a rst estimateof the duration of lemma selection in naming tasks ispresented in Levelt et al (1991) 115 msec That numberis derived from data obtained in a picture naminglexicaldecision experiment The primary task in the experimentwas picture naming But on one-third of the trials thesubject heard an auditory probe a word or a nonwordIt could be presented at one of three moments afterpicture onset but before the initiation of naming (ie atthree different stimulus onset asynchronies or SOAs) Onthese trials the subject had to perform a secondary taskbefore naming the picture pushing a ldquoyesrdquo or ldquonordquo buttondepending on whether the auditory probe was a wordor not On all relevant items the probe was a word andit could be semantically or phonologically related to thetarget picture or be unrelated to it (for instance if thepicture was one of a sheep the probe word could begoat sheet or house) At some SOAs the lexical decisionlatency to a semantically related probe (goat in theexample) was slower than to an unrelated probe (suchas house in the example) The obvious interpretation forthat nding is that the picture naming had advanced sofar at that SOA that the target lemma was active andcould begin interfering with the probe lemma The com-putational model that provided an optimal t for theselexical decision data yielded the mentioned 115-msecestimate for lemma access
This estimate is in good agreement with the oneprovided by the computational model of lemma selec-tion in Roelofs (1992) Lemma access was the very targetof that paper Roelofs used a pictureword interferencetask in his experiments Subjects named pictures andnaming latency was the dependent measure in theseexperiments At some moment during the task (ie atdifferent SOAs) a visual distracter word was ashed inthe picture and the subject was instructed to ignoreit The distracter could be semantically related to thetarget picture name or unrelated to it (just as in the
556 Journal of Cognitive Neuroscience Volume 10 Number 5
above experiment) Typically naming latencies wereslower when the distracter was semantically related(such as goat when a sheep was displayed) than whenit was an unrelated distracter (such as house) at leastwhen picture and distracter presentation were closelysynchronized The computational model based on thesedata sets gives lemma selection durations in the rangeof 100 to 150 msec This nally corresponds wellwith what Potter (personal communication) derivesfrom the data in Potter and Faulconer (1975) an estimateof 150 msec
Duration estimates for the next stage of lexical encod-ing word form access and phonological encoding canbe derived from the following three sources The rstone is Roelofsrsquos (1997) WEAVER model of phonologicalencoding It is based on myriad pictureword interfer-ence experiments in which auditory phonological dis-tracters were presented during picture naming Based onthese data Roelofs gives an estimate of 265 msec for theinterval between lemma selection and accessing thesyllable score of a monosyllabic word Because this in-volves selecting the syllable node (although not ldquounpack-ingrdquo) it the time for attaching phonemic segments to themetrical frame should be less An estimate for the lattercomes from a second study by Wheeldon and Levelt(1994) In their experiments the subject was providedwith a target phoneme for instance an l or an r The(Dutch) subject would then hear an English word (forexample hitchhiker) and decided whether the Dutchtranslation of that word (lifter in the example case)contained the target phoneme The Dutch word was notovertly pronounced The dependent measure was thepush button (yes) latency For bisyllabic items such aslifter the average difference in phoneme monitoringspeed between the word initial and the word nal pho-neme (l and r for this example) turned out to beabout 125 msec This can be taken as an estimate of theduration to (internally) encode the whole wordphonologically Finally van Turennout Hagoort andBrown (1997) in an ERP study not unlike Thorpe et alrsquoswhere word-initial and word-nal phonemes providedgono-go cues for a semantic picture categorizationcould estimate a duration of about 120 msec forphonological encoding
Left over then are phonetic encoding and the initia-tion of articulation If the above estimates are approxi-mately correct subtraction can provide an estimate forthese nal stages For an average naming latency of 538msec the distribution over the stages would roughly beas follows visual processing plus accessing the lexicalconcept 150 msec lemma selection 125 msec phono-logical encoding 125 msec and phonetic encoding andinitiation of articulation 138 msec This latter numberagrees well with a nal estimate that we have Leveltet al (1991) provide a model estimate for the durationof phonological plus phonetic encoding based on theirpicture naminglexical decision expriments it is 270
msec If we subtract 125 msec for phonological encod-ing we are left with 145 msec for the duration ofphonetic encoding
In the present study we will group activation dataaccording to the following time windows post-pictureonset
0ndash150 msec Visual processing and accessing the lexical concept
150ndash275 msec Lemma selection275ndash400 msec Phonological encoding400ndash600 msec Phonetic and articulatory processing
The last window extends somewhat beyond the averagenaming response to include most of the factual namingresponses (but of course for each response only pre-onset activation data were analyzed) Although thesewindows capture all of the data in the above meta-analy-sis these data do not allow us to estimate windowvariances However our selection of experimental mate-rials (see ldquoMethodsrdquo) aimed at minimizing variances inobject recognition and in total naming latency
Singling Out Phonological Encoding
As mentioned previously one can in principle study thecerebral localization of a single processing stage by in-troducing an independent variable that specically af-fects the duration of that stage Any dependent effect inthe MEG trace should be due to the correspondingprocessing component In the present study we decidedto try this out on the stage of phonological encodingJescheniak and Levelt (1994) discovered that the well-known word frequency effect in picture naming(Oldeld amp Wingeld 1965) is due to accessing thewordrsquos form Pictures whose names are highly frequent(HF) in the speakerrsquos language (such as boat) are namedfaster (by some 30 to 50 msec) than pictures whosenames have low frequency (LF) (such as broom) Theauthors could show that this is not due to forming thepercept or to retrieving the lemma or the wordrsquos sylla-bles The effect arises in accessing the wordrsquos form infor-mation
This fact can now in principle be used to localize theprocess of word form access in the brain if one hasaccess to a temporal high-resolution method such asMEG The idea is this (see Figure 2) When the inde-pendent variable in the picture naming task is targetword frequency (ie HF versus LF picture names) weshould expect that the initial processing stages up to thestage of form access are the same for HF and LF picturesBut from morpheme access on the time course for HFpictures will divert from the time course for LF picturesMore precisely (see Figure 2) the critical stage will berelatively short for the HF pictures and relatively long forthe LF pictures (the time difference dt will be about 30to 50 msec) All subsequent stages will not differ induration but will be desynchronized by dt One can now
Levelt et al 557
trace the activation pattern over the MEG sensors anddetermine whether such desynchronization arises and inparticular where it begins Inversely one can response-lock the activation patterns and determine where desyn-chronization begins running backward in time In theideal case one can ldquosqueeze inrdquo the critical process intime and place But as already mentioned 30 to 50 msecis not much and is easily drowned in experimental noise
Whatever it is worth this single factors or stage focus-ing method (or SF-method) differs essentially from thesubtraction method which is now commonly used inpositron emission tomography (PET) and functionalmagnetic resonance imaging (fMRI) studies There onetries to eliminate a stage completely here all stages arealways present There is reason to have serious doubtsabout the possibility of eliminating a stage entirely (Fris-ton et al 1996) In his classical paper Sternberg (1969)argued for a different approach the additive factorsmethod which leaves all stages intact The SF-method isa derivative of Sternbergrsquos method (see also Dehaene1996)
In the present study eight Dutch subjects repeatedlynamed 24 carefully preselected pictures 12 with LFnames and 12 with HF names while their magnetic brainactivity was recorded in the Neuromag-122 whole-headmagnetometer This was done in two identical and im-mediately adjacent sessions of six randomly dispersedrepetitions of all 24 pictures In a prerun of exactly thesame experiment but without MEG recording and ondifferent subjects a 37-msec word frequency effect hadbeen obtained (see ldquoResultsrdquo)
RESULTS
In the following we will rst in summary fashion de-scribe the unsuccessful single-factors analysis and thenturn in detail to the more revealing sourcetime analysis
Single Factors Analysis
Reaction Times
As mentioned we obtained a solid 37-msec word fre-quency effect in the pre-experiment Figure 3 shows thereaction time curves for high- and low-frequency wordresponses The word frequency effect was stable over
the 12 repetitions (see ldquoMethodsrdquo) This substantial stableword frequency effect was not replicated in the mainexperiment (run on different subjects) Across all sub-jects mean reaction times for high- and low-frequencyitems differed by only 3 msec (535 versus 538 msec)The effects per subject ranged from 20 msec favoringthe low-frequency items to 15 msec favoring the high-frequency items In analyses of variance within subjectsnone of these effects reached statistical signicanceSeparate analyses were carried out for each subject andeach of the two test sessions (ie before versus after theshort break) A signicant (p lt 005) frequency effectfavoring high-frequency items was only obtained for onesubject in one test session
Should this null effect be adduced to the new set ofsubjects we tested or to the experimental situation inthe MEG lab To nd out we retested the same subjects6 months after the main experiment on the same mate-rials but in the Max Planck Institute (ie without MEGrecording) The instructions stressed that it was impor-tant to name the targets correctlymdashspeed was of secon-dary importance Now a signicant word frequencyeffect of 33 msec was obtained for the 24 relevant test
Figure 2 The single-factorsmethod Affecting the durationof a single stage does not af-fect the cortical dynamics be-fore that stage but shifts allactivity following that stageby a constant dt
Figure 3 The word frequency effect in the naming latency for theultimate 12 HF and 12 LF stimuli as measured in the preexperimentover 12 repetitions of the test pictures
558 Journal of Cognitive Neuroscience Volume 10 Number 5
items (F1(1 7) = 1376 MSE = 3771 F2(1 22) = 968MSE = 7965 both p lt 001) Six subjects showed fre-quency effects favoring the high-frequency items in bothhalves of the experiment and two subjects showed themonly in one-half of it In other words during the retestthe subjects of the main experiments behaved verymuch like the subjects of the pretest Hence the nulleffect in the main experiment was not due to the set ofsubjects Was it due to the testing situation The mostnoticeable difference was that naming latencies werevery short in the MEG experiment on average 538 msecThese latencies were manually determined from thevisually displayed acoustic wave form In the preexperi-ment (different subjects) average naming latency hadbeen 592 msec measured by voice key Because thevoice key is triggered at some acoustic threshold voicekey latencies are always slower than latencies measuredon the wave form display this amounts to about 20 msecon average Even with this correction the latencies inthe MEG experiment were faster by 42 msec Is it thecase that our young Dutch subjects own into Helsinkifor a weekend had been so highly motivated during theMEG session that their naming speeds reached ceilinglevels obliterating any difference between HF and LFwords This is unlikely First in their post-test their aver-age voice key latency was 575 msec Implementing the20-msec correction this differs by no more than 17 msecfrom the latencies in the MEG experiment Second wetested a new set of 10 subjects who did the experimenttwice (in two sessions 1 week apart) once under strictspeed conditions and once under the instruction toavoid errors and not to worry about time Although thevoice key averages differed substantially between thesetwo conditions (542 msec versus 618 msec) the fre-quency effect was exactly the same 27 msec Notice thatthe speeded condition was faster by 16 msec than in theMEG experiment (correcting for the 20 msec) So aword frequency effect can be obtained even with veryfast naming responses In short we have no explanationfor the tragic disappearance of the effect in the MEGexperiment It did however undermine our single-factors analysis
Evoked Magnetic Fields
Although the null effect for word frequency eliminatedthe single-factors analysis as displayed in Figure 2 theremight still be a word frequency effect in the evokedmagnetic data For all subjects we visually checked theHF and LF responses recorded by the 122 sensors Thespatio-temporal patterns of evoked magnetic elds dif-fered considerably between subjects except for the rela-tively uniform exogenous visual evoked elds recordedat occipital sensors But none of the subjects showed anysystematic difference between HF and LF activation pat-terns Nevertheless we performed analyses of varianceover HF versus LF curves for all sensors and all subjects
by partitioning all curves in 50-msec windows rangingfrom 0 to 600 msec post-onset Although we found moresignicant word frequency effects than could be ex-pected by chance they were not ubiquitous and showedno interpretable patterns within or over subjects Thiswas further conrmed in the dipole source analysis (seeFigure 4)
Dipole Source Analysis
The main aim of our dipole source analysis was to relatepeak activity of distinct cortical areas to the time win-dows of the stage model of picture naming Thereforedipole source analysis was performed separately for eachsubjectrsquos data using the signals from subjectsrsquo rst test-ing session and averaged over the two frequency condi-tions (see ldquoMethodsrdquo section) Figure 4 shows theoriginal whole-head MEG responses for HF and LF wordsin one subject The data were modeled by eight equiva-lent current dipoles (ECD) identied at distinct latenciesand shown on the surface rendition of the subjectrsquos MRIThe source strengths as a function of time are plottedon the right
The source localization results over all subjects aredisplayed in Figure 5 The four time windows discussedabove correspond to four colors in the diagrams Let usgo over these windows in temporal order
Visual Processing and Accessing the Lexical Concept0 to 150 msec
The visual evoked magnetic elds were adequately mod-eled by about two to three sources in each subjectThese sources were located in the occipital cortex (yel-low sources) with a preponderance of sources in theright hemisphere Note that the model sphere for thesource analyses was tted in such a way that centralparietal and temporal areas were covered optimallyTherefore localization errors are likely to be largest inthe frontal and occipital areas Because the modeling ofvisual-perceptual activity was not the goal of this studyno further attempt was made to determine the locationof these sources more precisely The ndings do agreehowever with the occipital lobe activation adduced toobject recognition in Price et alrsquos (1996) PET study ofpicture naming
Lemma Selection 150 to 275 msec
In this time window magnetic eld patterns were morevariable The sources identied in this time frame (threeto four per subjectmdashblue dots in the second panel) werelargely found in the occipital area (50) and in theparietal and occasionally temporal areas However therewas a remarkable clustering of sources (seven of eightsubjects) in the right parietal cortex along the posteriorend of the superior temporal sulcus
Levelt et al 559
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

above experiment) Typically naming latencies wereslower when the distracter was semantically related(such as goat when a sheep was displayed) than whenit was an unrelated distracter (such as house) at leastwhen picture and distracter presentation were closelysynchronized The computational model based on thesedata sets gives lemma selection durations in the rangeof 100 to 150 msec This nally corresponds wellwith what Potter (personal communication) derivesfrom the data in Potter and Faulconer (1975) an estimateof 150 msec
Duration estimates for the next stage of lexical encod-ing word form access and phonological encoding canbe derived from the following three sources The rstone is Roelofsrsquos (1997) WEAVER model of phonologicalencoding It is based on myriad pictureword interfer-ence experiments in which auditory phonological dis-tracters were presented during picture naming Based onthese data Roelofs gives an estimate of 265 msec for theinterval between lemma selection and accessing thesyllable score of a monosyllabic word Because this in-volves selecting the syllable node (although not ldquounpack-ingrdquo) it the time for attaching phonemic segments to themetrical frame should be less An estimate for the lattercomes from a second study by Wheeldon and Levelt(1994) In their experiments the subject was providedwith a target phoneme for instance an l or an r The(Dutch) subject would then hear an English word (forexample hitchhiker) and decided whether the Dutchtranslation of that word (lifter in the example case)contained the target phoneme The Dutch word was notovertly pronounced The dependent measure was thepush button (yes) latency For bisyllabic items such aslifter the average difference in phoneme monitoringspeed between the word initial and the word nal pho-neme (l and r for this example) turned out to beabout 125 msec This can be taken as an estimate of theduration to (internally) encode the whole wordphonologically Finally van Turennout Hagoort andBrown (1997) in an ERP study not unlike Thorpe et alrsquoswhere word-initial and word-nal phonemes providedgono-go cues for a semantic picture categorizationcould estimate a duration of about 120 msec forphonological encoding
Left over then are phonetic encoding and the initia-tion of articulation If the above estimates are approxi-mately correct subtraction can provide an estimate forthese nal stages For an average naming latency of 538msec the distribution over the stages would roughly beas follows visual processing plus accessing the lexicalconcept 150 msec lemma selection 125 msec phono-logical encoding 125 msec and phonetic encoding andinitiation of articulation 138 msec This latter numberagrees well with a nal estimate that we have Leveltet al (1991) provide a model estimate for the durationof phonological plus phonetic encoding based on theirpicture naminglexical decision expriments it is 270
msec If we subtract 125 msec for phonological encod-ing we are left with 145 msec for the duration ofphonetic encoding
In the present study we will group activation dataaccording to the following time windows post-pictureonset
0ndash150 msec Visual processing and accessing the lexical concept
150ndash275 msec Lemma selection275ndash400 msec Phonological encoding400ndash600 msec Phonetic and articulatory processing
The last window extends somewhat beyond the averagenaming response to include most of the factual namingresponses (but of course for each response only pre-onset activation data were analyzed) Although thesewindows capture all of the data in the above meta-analy-sis these data do not allow us to estimate windowvariances However our selection of experimental mate-rials (see ldquoMethodsrdquo) aimed at minimizing variances inobject recognition and in total naming latency
Singling Out Phonological Encoding
As mentioned previously one can in principle study thecerebral localization of a single processing stage by in-troducing an independent variable that specically af-fects the duration of that stage Any dependent effect inthe MEG trace should be due to the correspondingprocessing component In the present study we decidedto try this out on the stage of phonological encodingJescheniak and Levelt (1994) discovered that the well-known word frequency effect in picture naming(Oldeld amp Wingeld 1965) is due to accessing thewordrsquos form Pictures whose names are highly frequent(HF) in the speakerrsquos language (such as boat) are namedfaster (by some 30 to 50 msec) than pictures whosenames have low frequency (LF) (such as broom) Theauthors could show that this is not due to forming thepercept or to retrieving the lemma or the wordrsquos sylla-bles The effect arises in accessing the wordrsquos form infor-mation
This fact can now in principle be used to localize theprocess of word form access in the brain if one hasaccess to a temporal high-resolution method such asMEG The idea is this (see Figure 2) When the inde-pendent variable in the picture naming task is targetword frequency (ie HF versus LF picture names) weshould expect that the initial processing stages up to thestage of form access are the same for HF and LF picturesBut from morpheme access on the time course for HFpictures will divert from the time course for LF picturesMore precisely (see Figure 2) the critical stage will berelatively short for the HF pictures and relatively long forthe LF pictures (the time difference dt will be about 30to 50 msec) All subsequent stages will not differ induration but will be desynchronized by dt One can now
Levelt et al 557
trace the activation pattern over the MEG sensors anddetermine whether such desynchronization arises and inparticular where it begins Inversely one can response-lock the activation patterns and determine where desyn-chronization begins running backward in time In theideal case one can ldquosqueeze inrdquo the critical process intime and place But as already mentioned 30 to 50 msecis not much and is easily drowned in experimental noise
Whatever it is worth this single factors or stage focus-ing method (or SF-method) differs essentially from thesubtraction method which is now commonly used inpositron emission tomography (PET) and functionalmagnetic resonance imaging (fMRI) studies There onetries to eliminate a stage completely here all stages arealways present There is reason to have serious doubtsabout the possibility of eliminating a stage entirely (Fris-ton et al 1996) In his classical paper Sternberg (1969)argued for a different approach the additive factorsmethod which leaves all stages intact The SF-method isa derivative of Sternbergrsquos method (see also Dehaene1996)
In the present study eight Dutch subjects repeatedlynamed 24 carefully preselected pictures 12 with LFnames and 12 with HF names while their magnetic brainactivity was recorded in the Neuromag-122 whole-headmagnetometer This was done in two identical and im-mediately adjacent sessions of six randomly dispersedrepetitions of all 24 pictures In a prerun of exactly thesame experiment but without MEG recording and ondifferent subjects a 37-msec word frequency effect hadbeen obtained (see ldquoResultsrdquo)
RESULTS
In the following we will rst in summary fashion de-scribe the unsuccessful single-factors analysis and thenturn in detail to the more revealing sourcetime analysis
Single Factors Analysis
Reaction Times
As mentioned we obtained a solid 37-msec word fre-quency effect in the pre-experiment Figure 3 shows thereaction time curves for high- and low-frequency wordresponses The word frequency effect was stable over
the 12 repetitions (see ldquoMethodsrdquo) This substantial stableword frequency effect was not replicated in the mainexperiment (run on different subjects) Across all sub-jects mean reaction times for high- and low-frequencyitems differed by only 3 msec (535 versus 538 msec)The effects per subject ranged from 20 msec favoringthe low-frequency items to 15 msec favoring the high-frequency items In analyses of variance within subjectsnone of these effects reached statistical signicanceSeparate analyses were carried out for each subject andeach of the two test sessions (ie before versus after theshort break) A signicant (p lt 005) frequency effectfavoring high-frequency items was only obtained for onesubject in one test session
Should this null effect be adduced to the new set ofsubjects we tested or to the experimental situation inthe MEG lab To nd out we retested the same subjects6 months after the main experiment on the same mate-rials but in the Max Planck Institute (ie without MEGrecording) The instructions stressed that it was impor-tant to name the targets correctlymdashspeed was of secon-dary importance Now a signicant word frequencyeffect of 33 msec was obtained for the 24 relevant test
Figure 2 The single-factorsmethod Affecting the durationof a single stage does not af-fect the cortical dynamics be-fore that stage but shifts allactivity following that stageby a constant dt
Figure 3 The word frequency effect in the naming latency for theultimate 12 HF and 12 LF stimuli as measured in the preexperimentover 12 repetitions of the test pictures
558 Journal of Cognitive Neuroscience Volume 10 Number 5
items (F1(1 7) = 1376 MSE = 3771 F2(1 22) = 968MSE = 7965 both p lt 001) Six subjects showed fre-quency effects favoring the high-frequency items in bothhalves of the experiment and two subjects showed themonly in one-half of it In other words during the retestthe subjects of the main experiments behaved verymuch like the subjects of the pretest Hence the nulleffect in the main experiment was not due to the set ofsubjects Was it due to the testing situation The mostnoticeable difference was that naming latencies werevery short in the MEG experiment on average 538 msecThese latencies were manually determined from thevisually displayed acoustic wave form In the preexperi-ment (different subjects) average naming latency hadbeen 592 msec measured by voice key Because thevoice key is triggered at some acoustic threshold voicekey latencies are always slower than latencies measuredon the wave form display this amounts to about 20 msecon average Even with this correction the latencies inthe MEG experiment were faster by 42 msec Is it thecase that our young Dutch subjects own into Helsinkifor a weekend had been so highly motivated during theMEG session that their naming speeds reached ceilinglevels obliterating any difference between HF and LFwords This is unlikely First in their post-test their aver-age voice key latency was 575 msec Implementing the20-msec correction this differs by no more than 17 msecfrom the latencies in the MEG experiment Second wetested a new set of 10 subjects who did the experimenttwice (in two sessions 1 week apart) once under strictspeed conditions and once under the instruction toavoid errors and not to worry about time Although thevoice key averages differed substantially between thesetwo conditions (542 msec versus 618 msec) the fre-quency effect was exactly the same 27 msec Notice thatthe speeded condition was faster by 16 msec than in theMEG experiment (correcting for the 20 msec) So aword frequency effect can be obtained even with veryfast naming responses In short we have no explanationfor the tragic disappearance of the effect in the MEGexperiment It did however undermine our single-factors analysis
Evoked Magnetic Fields
Although the null effect for word frequency eliminatedthe single-factors analysis as displayed in Figure 2 theremight still be a word frequency effect in the evokedmagnetic data For all subjects we visually checked theHF and LF responses recorded by the 122 sensors Thespatio-temporal patterns of evoked magnetic elds dif-fered considerably between subjects except for the rela-tively uniform exogenous visual evoked elds recordedat occipital sensors But none of the subjects showed anysystematic difference between HF and LF activation pat-terns Nevertheless we performed analyses of varianceover HF versus LF curves for all sensors and all subjects
by partitioning all curves in 50-msec windows rangingfrom 0 to 600 msec post-onset Although we found moresignicant word frequency effects than could be ex-pected by chance they were not ubiquitous and showedno interpretable patterns within or over subjects Thiswas further conrmed in the dipole source analysis (seeFigure 4)
Dipole Source Analysis
The main aim of our dipole source analysis was to relatepeak activity of distinct cortical areas to the time win-dows of the stage model of picture naming Thereforedipole source analysis was performed separately for eachsubjectrsquos data using the signals from subjectsrsquo rst test-ing session and averaged over the two frequency condi-tions (see ldquoMethodsrdquo section) Figure 4 shows theoriginal whole-head MEG responses for HF and LF wordsin one subject The data were modeled by eight equiva-lent current dipoles (ECD) identied at distinct latenciesand shown on the surface rendition of the subjectrsquos MRIThe source strengths as a function of time are plottedon the right
The source localization results over all subjects aredisplayed in Figure 5 The four time windows discussedabove correspond to four colors in the diagrams Let usgo over these windows in temporal order
Visual Processing and Accessing the Lexical Concept0 to 150 msec
The visual evoked magnetic elds were adequately mod-eled by about two to three sources in each subjectThese sources were located in the occipital cortex (yel-low sources) with a preponderance of sources in theright hemisphere Note that the model sphere for thesource analyses was tted in such a way that centralparietal and temporal areas were covered optimallyTherefore localization errors are likely to be largest inthe frontal and occipital areas Because the modeling ofvisual-perceptual activity was not the goal of this studyno further attempt was made to determine the locationof these sources more precisely The ndings do agreehowever with the occipital lobe activation adduced toobject recognition in Price et alrsquos (1996) PET study ofpicture naming
Lemma Selection 150 to 275 msec
In this time window magnetic eld patterns were morevariable The sources identied in this time frame (threeto four per subjectmdashblue dots in the second panel) werelargely found in the occipital area (50) and in theparietal and occasionally temporal areas However therewas a remarkable clustering of sources (seven of eightsubjects) in the right parietal cortex along the posteriorend of the superior temporal sulcus
Levelt et al 559
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

trace the activation pattern over the MEG sensors anddetermine whether such desynchronization arises and inparticular where it begins Inversely one can response-lock the activation patterns and determine where desyn-chronization begins running backward in time In theideal case one can ldquosqueeze inrdquo the critical process intime and place But as already mentioned 30 to 50 msecis not much and is easily drowned in experimental noise
Whatever it is worth this single factors or stage focus-ing method (or SF-method) differs essentially from thesubtraction method which is now commonly used inpositron emission tomography (PET) and functionalmagnetic resonance imaging (fMRI) studies There onetries to eliminate a stage completely here all stages arealways present There is reason to have serious doubtsabout the possibility of eliminating a stage entirely (Fris-ton et al 1996) In his classical paper Sternberg (1969)argued for a different approach the additive factorsmethod which leaves all stages intact The SF-method isa derivative of Sternbergrsquos method (see also Dehaene1996)
In the present study eight Dutch subjects repeatedlynamed 24 carefully preselected pictures 12 with LFnames and 12 with HF names while their magnetic brainactivity was recorded in the Neuromag-122 whole-headmagnetometer This was done in two identical and im-mediately adjacent sessions of six randomly dispersedrepetitions of all 24 pictures In a prerun of exactly thesame experiment but without MEG recording and ondifferent subjects a 37-msec word frequency effect hadbeen obtained (see ldquoResultsrdquo)
RESULTS
In the following we will rst in summary fashion de-scribe the unsuccessful single-factors analysis and thenturn in detail to the more revealing sourcetime analysis
Single Factors Analysis
Reaction Times
As mentioned we obtained a solid 37-msec word fre-quency effect in the pre-experiment Figure 3 shows thereaction time curves for high- and low-frequency wordresponses The word frequency effect was stable over
the 12 repetitions (see ldquoMethodsrdquo) This substantial stableword frequency effect was not replicated in the mainexperiment (run on different subjects) Across all sub-jects mean reaction times for high- and low-frequencyitems differed by only 3 msec (535 versus 538 msec)The effects per subject ranged from 20 msec favoringthe low-frequency items to 15 msec favoring the high-frequency items In analyses of variance within subjectsnone of these effects reached statistical signicanceSeparate analyses were carried out for each subject andeach of the two test sessions (ie before versus after theshort break) A signicant (p lt 005) frequency effectfavoring high-frequency items was only obtained for onesubject in one test session
Should this null effect be adduced to the new set ofsubjects we tested or to the experimental situation inthe MEG lab To nd out we retested the same subjects6 months after the main experiment on the same mate-rials but in the Max Planck Institute (ie without MEGrecording) The instructions stressed that it was impor-tant to name the targets correctlymdashspeed was of secon-dary importance Now a signicant word frequencyeffect of 33 msec was obtained for the 24 relevant test
Figure 2 The single-factorsmethod Affecting the durationof a single stage does not af-fect the cortical dynamics be-fore that stage but shifts allactivity following that stageby a constant dt
Figure 3 The word frequency effect in the naming latency for theultimate 12 HF and 12 LF stimuli as measured in the preexperimentover 12 repetitions of the test pictures
558 Journal of Cognitive Neuroscience Volume 10 Number 5
items (F1(1 7) = 1376 MSE = 3771 F2(1 22) = 968MSE = 7965 both p lt 001) Six subjects showed fre-quency effects favoring the high-frequency items in bothhalves of the experiment and two subjects showed themonly in one-half of it In other words during the retestthe subjects of the main experiments behaved verymuch like the subjects of the pretest Hence the nulleffect in the main experiment was not due to the set ofsubjects Was it due to the testing situation The mostnoticeable difference was that naming latencies werevery short in the MEG experiment on average 538 msecThese latencies were manually determined from thevisually displayed acoustic wave form In the preexperi-ment (different subjects) average naming latency hadbeen 592 msec measured by voice key Because thevoice key is triggered at some acoustic threshold voicekey latencies are always slower than latencies measuredon the wave form display this amounts to about 20 msecon average Even with this correction the latencies inthe MEG experiment were faster by 42 msec Is it thecase that our young Dutch subjects own into Helsinkifor a weekend had been so highly motivated during theMEG session that their naming speeds reached ceilinglevels obliterating any difference between HF and LFwords This is unlikely First in their post-test their aver-age voice key latency was 575 msec Implementing the20-msec correction this differs by no more than 17 msecfrom the latencies in the MEG experiment Second wetested a new set of 10 subjects who did the experimenttwice (in two sessions 1 week apart) once under strictspeed conditions and once under the instruction toavoid errors and not to worry about time Although thevoice key averages differed substantially between thesetwo conditions (542 msec versus 618 msec) the fre-quency effect was exactly the same 27 msec Notice thatthe speeded condition was faster by 16 msec than in theMEG experiment (correcting for the 20 msec) So aword frequency effect can be obtained even with veryfast naming responses In short we have no explanationfor the tragic disappearance of the effect in the MEGexperiment It did however undermine our single-factors analysis
Evoked Magnetic Fields
Although the null effect for word frequency eliminatedthe single-factors analysis as displayed in Figure 2 theremight still be a word frequency effect in the evokedmagnetic data For all subjects we visually checked theHF and LF responses recorded by the 122 sensors Thespatio-temporal patterns of evoked magnetic elds dif-fered considerably between subjects except for the rela-tively uniform exogenous visual evoked elds recordedat occipital sensors But none of the subjects showed anysystematic difference between HF and LF activation pat-terns Nevertheless we performed analyses of varianceover HF versus LF curves for all sensors and all subjects
by partitioning all curves in 50-msec windows rangingfrom 0 to 600 msec post-onset Although we found moresignicant word frequency effects than could be ex-pected by chance they were not ubiquitous and showedno interpretable patterns within or over subjects Thiswas further conrmed in the dipole source analysis (seeFigure 4)
Dipole Source Analysis
The main aim of our dipole source analysis was to relatepeak activity of distinct cortical areas to the time win-dows of the stage model of picture naming Thereforedipole source analysis was performed separately for eachsubjectrsquos data using the signals from subjectsrsquo rst test-ing session and averaged over the two frequency condi-tions (see ldquoMethodsrdquo section) Figure 4 shows theoriginal whole-head MEG responses for HF and LF wordsin one subject The data were modeled by eight equiva-lent current dipoles (ECD) identied at distinct latenciesand shown on the surface rendition of the subjectrsquos MRIThe source strengths as a function of time are plottedon the right
The source localization results over all subjects aredisplayed in Figure 5 The four time windows discussedabove correspond to four colors in the diagrams Let usgo over these windows in temporal order
Visual Processing and Accessing the Lexical Concept0 to 150 msec
The visual evoked magnetic elds were adequately mod-eled by about two to three sources in each subjectThese sources were located in the occipital cortex (yel-low sources) with a preponderance of sources in theright hemisphere Note that the model sphere for thesource analyses was tted in such a way that centralparietal and temporal areas were covered optimallyTherefore localization errors are likely to be largest inthe frontal and occipital areas Because the modeling ofvisual-perceptual activity was not the goal of this studyno further attempt was made to determine the locationof these sources more precisely The ndings do agreehowever with the occipital lobe activation adduced toobject recognition in Price et alrsquos (1996) PET study ofpicture naming
Lemma Selection 150 to 275 msec
In this time window magnetic eld patterns were morevariable The sources identied in this time frame (threeto four per subjectmdashblue dots in the second panel) werelargely found in the occipital area (50) and in theparietal and occasionally temporal areas However therewas a remarkable clustering of sources (seven of eightsubjects) in the right parietal cortex along the posteriorend of the superior temporal sulcus
Levelt et al 559
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

items (F1(1 7) = 1376 MSE = 3771 F2(1 22) = 968MSE = 7965 both p lt 001) Six subjects showed fre-quency effects favoring the high-frequency items in bothhalves of the experiment and two subjects showed themonly in one-half of it In other words during the retestthe subjects of the main experiments behaved verymuch like the subjects of the pretest Hence the nulleffect in the main experiment was not due to the set ofsubjects Was it due to the testing situation The mostnoticeable difference was that naming latencies werevery short in the MEG experiment on average 538 msecThese latencies were manually determined from thevisually displayed acoustic wave form In the preexperi-ment (different subjects) average naming latency hadbeen 592 msec measured by voice key Because thevoice key is triggered at some acoustic threshold voicekey latencies are always slower than latencies measuredon the wave form display this amounts to about 20 msecon average Even with this correction the latencies inthe MEG experiment were faster by 42 msec Is it thecase that our young Dutch subjects own into Helsinkifor a weekend had been so highly motivated during theMEG session that their naming speeds reached ceilinglevels obliterating any difference between HF and LFwords This is unlikely First in their post-test their aver-age voice key latency was 575 msec Implementing the20-msec correction this differs by no more than 17 msecfrom the latencies in the MEG experiment Second wetested a new set of 10 subjects who did the experimenttwice (in two sessions 1 week apart) once under strictspeed conditions and once under the instruction toavoid errors and not to worry about time Although thevoice key averages differed substantially between thesetwo conditions (542 msec versus 618 msec) the fre-quency effect was exactly the same 27 msec Notice thatthe speeded condition was faster by 16 msec than in theMEG experiment (correcting for the 20 msec) So aword frequency effect can be obtained even with veryfast naming responses In short we have no explanationfor the tragic disappearance of the effect in the MEGexperiment It did however undermine our single-factors analysis
Evoked Magnetic Fields
Although the null effect for word frequency eliminatedthe single-factors analysis as displayed in Figure 2 theremight still be a word frequency effect in the evokedmagnetic data For all subjects we visually checked theHF and LF responses recorded by the 122 sensors Thespatio-temporal patterns of evoked magnetic elds dif-fered considerably between subjects except for the rela-tively uniform exogenous visual evoked elds recordedat occipital sensors But none of the subjects showed anysystematic difference between HF and LF activation pat-terns Nevertheless we performed analyses of varianceover HF versus LF curves for all sensors and all subjects
by partitioning all curves in 50-msec windows rangingfrom 0 to 600 msec post-onset Although we found moresignicant word frequency effects than could be ex-pected by chance they were not ubiquitous and showedno interpretable patterns within or over subjects Thiswas further conrmed in the dipole source analysis (seeFigure 4)
Dipole Source Analysis
The main aim of our dipole source analysis was to relatepeak activity of distinct cortical areas to the time win-dows of the stage model of picture naming Thereforedipole source analysis was performed separately for eachsubjectrsquos data using the signals from subjectsrsquo rst test-ing session and averaged over the two frequency condi-tions (see ldquoMethodsrdquo section) Figure 4 shows theoriginal whole-head MEG responses for HF and LF wordsin one subject The data were modeled by eight equiva-lent current dipoles (ECD) identied at distinct latenciesand shown on the surface rendition of the subjectrsquos MRIThe source strengths as a function of time are plottedon the right
The source localization results over all subjects aredisplayed in Figure 5 The four time windows discussedabove correspond to four colors in the diagrams Let usgo over these windows in temporal order
Visual Processing and Accessing the Lexical Concept0 to 150 msec
The visual evoked magnetic elds were adequately mod-eled by about two to three sources in each subjectThese sources were located in the occipital cortex (yel-low sources) with a preponderance of sources in theright hemisphere Note that the model sphere for thesource analyses was tted in such a way that centralparietal and temporal areas were covered optimallyTherefore localization errors are likely to be largest inthe frontal and occipital areas Because the modeling ofvisual-perceptual activity was not the goal of this studyno further attempt was made to determine the locationof these sources more precisely The ndings do agreehowever with the occipital lobe activation adduced toobject recognition in Price et alrsquos (1996) PET study ofpicture naming
Lemma Selection 150 to 275 msec
In this time window magnetic eld patterns were morevariable The sources identied in this time frame (threeto four per subjectmdashblue dots in the second panel) werelargely found in the occipital area (50) and in theparietal and occasionally temporal areas However therewas a remarkable clustering of sources (seven of eightsubjects) in the right parietal cortex along the posteriorend of the superior temporal sulcus
Levelt et al 559
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567
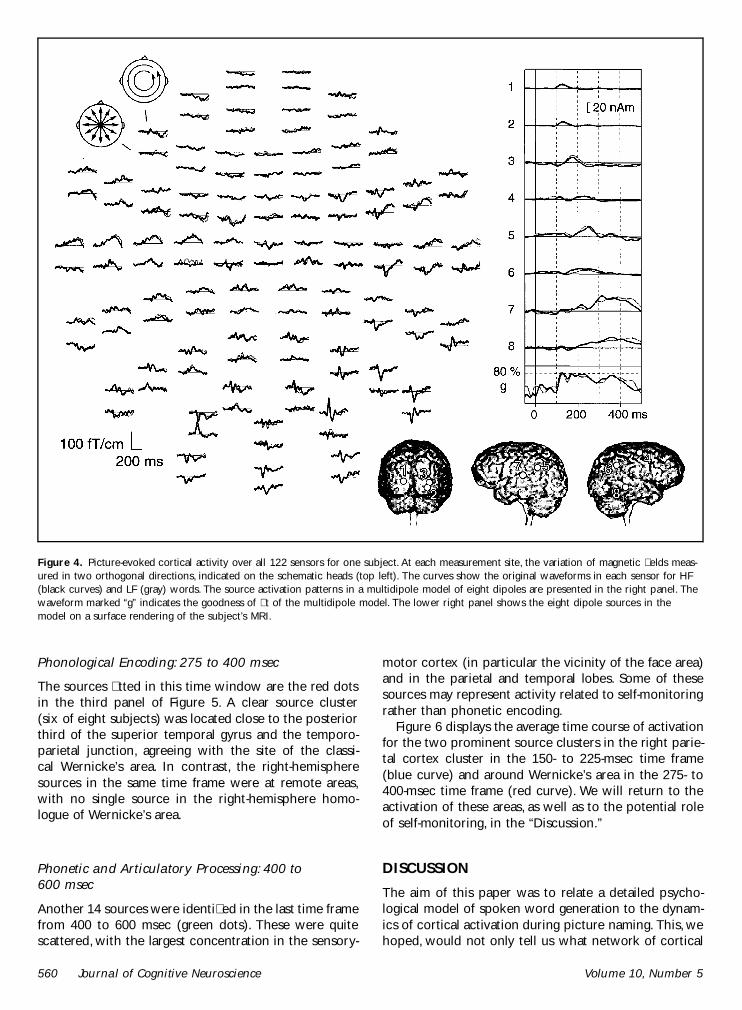
Phonological Encoding 275 to 400 msec
The sources tted in this time window are the red dotsin the third panel of Figure 5 A clear source cluster(six of eight subjects) was located close to the posteriorthird of the superior temporal gyrus and the temporo-parietal junction agreeing with the site of the classi-cal Wernickersquos area In contrast the right-hemispheresources in the same time frame were at remote areaswith no single source in the right-hemisphere homo-logue of Wernickersquos area
Phonetic and Articulatory Processing 400 to600 msec
Another 14 sources were identied in the last time framefrom 400 to 600 msec (green dots) These were quitescattered with the largest concentration in the sensory-
motor cortex (in particular the vicinity of the face area)and in the parietal and temporal lobes Some of thesesources may represent activity related to self-monitoringrather than phonetic encoding
Figure 6 displays the average time course of activationfor the two prominent source clusters in the right parie-tal cortex cluster in the 150- to 225-msec time frame(blue curve) and around Wernickersquos area in the 275- to400-msec time frame (red curve) We will return to theactivation of these areas as well as to the potential roleof self-monitoring in the ldquoDiscussionrdquo
DISCUSSION
The aim of this paper was to relate a detailed psycho-logical model of spoken word generation to the dynam-ics of cortical activation during picture naming This wehoped would not only tell us what network of cortical
Figure 4 Picture-evoked cortical activity over all 122 sensors for one subject At each measurement site the variation of magnetic elds meas-ured in two orthogonal directions indicated on the schematic heads (top left) The curves show the original waveforms in each sensor for HF(black curves) and LF (gray) words The source activation patterns in a multidipole model of eight dipoles are presented in the right panel Thewaveform marked ldquogrdquo indicates the goodness of t of the multidipole model The lower right panel shows the eight dipole sources in themodel on a surface rendering of the subjectrsquos MRI
560 Journal of Cognitive Neuroscience Volume 10 Number 5
areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

areas is involved in this production task but also andmore interestingly which loci are involved in successivefunctional stages in word generation During each ofthese stages a different highly specic computation isperformed involving different knowledge sources (vis-ual conceptual lexical phonological phonetic artic-ulatory) Each of these successive computations isindispensable if any of them fails naming will deterio-rate or block altogether But because the operations areso wildly different it is a reasonable working hypothesisthat they all involve different dedicated neural popula-tions
The two methods we applied to localize these dedi-cated regions were not equally successful The rst onethe single-factors method failed to work because of anunhappy complication in the experiment The inde-pendent variable word frequency was not effective forreasons that evade us in spite of careful pre- and post-tests Still we believe the logic of the method portrayedin Figure 2 to be correct To relate any stage-type proc-essing model to high temporal-resolution imaging datathe single-factors method is to be preferred over theusual subtraction methodology Notice that generally itis not required to have reliable estimates of the precise
Figure 5 Dipole sources ofevoked responses collectedover eight subjects Yellowdots Visual processing and ac-cessing the lexical concept 0ndash150 msec Blue dots Lemmaselection 150ndash275 msec Reddots Phonological encoding275ndash400 msec Green dotsPhonetic and articulatory proc-essing 400ndash600 msec Thegray dots represent othersources (ie those that donot have their peak activationin the relevant time frame)
Figure 6 The average timecourse of activation for thecluster of right parietalsources in the second timewindow (blue curve) and forthe cluster of Wernicke area di-poles in the third time win-dow (red curve) Plots wereobtained by reading the laten-cies at which the activationswere 0 25 50 75 and 100of the maximum in the dipolewave forms of the individualsubjects and then averagingthe latencies over those sub-jects who showed activity inthe relevant area
Levelt et al 561
time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

time course (over stages) of the process To apply theSF-method one only needs some independent experi-mental variable that can sufciently affect the durationof a single stage That can always be pretested in areaction time experiment
The second approach was to relate a dipole sourceanalysis to the precise temporal staging of the processas measured in reaction time experiments This type ofanalysis is more demanding of the psychological modelIt requires rather solid estimates of the durations of thesuccessive stages This is not normally given for psycho-logical stage models But for the task at hand picturenaming so much is available in terms of temporal mea-surements that reasonable estimates could in fact bemade of the time windows for successive stages Thatallowed us to detect stage-bound dipole sources Theresults were encouraging As a general pattern ourndings conform to Salmelin et alrsquos (1994)mdasha steadyprogression of activation from the occipital to the parie-tal-temporal and frontal areas of the brain But therewere substantial differences in detail First Salmelinet alrsquos data showed stronger involvement of lateralmotorpremotor areas in both hemispheres than thepresent data Second only the present data show con-centrated right-parietal activity (second time frame) Andthird only the present data show a denite clustering ofdipoles in the left posterior sylvian region (third timeframe) Still both experiments involved rather straight-forward picture naming tasks What could have causedthese and other differences in the patterns of evokedbrain activation The present experiment was not meantas a replication of the original study it served its ownpurpose In particular the single-factors word frequencyanalysis put specic requirements on the task and thestimulus materials It was for instance specic to thepresent experiment that (1) all picture names weremonomorphemic (2) all picture names were monosyl-labic (3) pretests had selected for pictures with smallvariance in recognition times (4) pretests had removedall pictures that showed naming uncertainty (5) pictureswere repeatedly presented during a session (six times)(6) a limited number of different pictures was used (42test plus ller stimuli as opposed to 200 pictures in theoriginal study) and (7) speed instructions were given tothe subjects It is hard to adduce differences in resultsspecically to any one of these procedural differencesbetween the experiments they have probably beenworking in concert The lesson is rather that even subtledifferences in tasks and experimental materials maycause substantial variation in evoked brain activationpatterns Here we analyze the present results in theirown right
Let us rst return to the right parietal lobe clusteringof dipoles which are active during the second timeperiod (150 to 275 msec) We had no a priori reason toexpect this area to be involved in lemma selection butit still may be in a task where lemma selection is based
on the recognition of visual objects There are two setsof relevant ndings in the literature The rst concernsthe role of the right parietal cortex in working memorytasks In a PET study Klingberg Kawashima and Roland(1996) found evidence for right inferior parietal activa-tion (on the border of angular and supramarginal gyri)involved in auditory visual and tactile short-term mem-ory tasks with detection tasks as controls Salmon et al(1996) also in a PET study found activation in the rightinferior parietal region during a verbal memory task inwhich subjects apparently used a visual imagery strategy(see Ungerleider 1995 for a review of cortical mecha-nisms for memory) In our experiment the subject sawthe same picture repeatedly in fact six times during thesession on which our dipole analyses are based Al-though on average 41 other pictures intervened be-tween two presentations of the same picture somesubjects may have tried to retrieve the response theyproduced earlier for the same now recognized pictureThe problem though with such a strategy is that it willincrease rather than decrease naming latency (cf Leveltamp Wheeldon 1994) If such an involvement of short-termvisual-verbal retention would nevertheless account forright parietal activation in our task it should entirelydisappear in a task where pictures are presented justonce A more likely interpretation of the right parietalcluster in our data derives from a second set of studieson the management of visual attention Faillenot ToniDecety Greacutegoire and Jeannerod (1997) also in a PETstudy observed activation in the right hemisphere at theborder of Brodmannrsquos areas (BA) 19 and 7 during a visualshape-matching task They attributed this activation tothe shifting of attention to the location of an expectedobject part or property This is in good agreement witha PET study by Fink et al (1996) in which the rightparietal region (in particular BA 19 and 39) is involvedin visually attending to global features of a stimulusHusain Shapiro Martin and Kennard (1997) showed thatdamage to this area affects the patientrsquos ability to disen-gage attention from one visual stimulus to the nextLemma access in our task may gain by active manage-ment of visual attention Global features of the pic-ture will often be informative of its semantic categoryattending to them may speed up the zooming in on theappropriate lemma In addition a strategy of rapidlydisengaging visual attention from the picture just namedin order to be ready for the next one may have contrib-uted to the short naming latencies that we observed inthe present experiment
The sources showing the remarkable clustering in theleft temporal cortex were active during the stage ofphonological encoding It strongly suggests Wernickersquosarea to be a favorite site for phonological encoding inword production This nding certainly agrees with theOjemann et al (1989) and Haglund et al (1994) statisticswhich show a high concentration of language sites injust this area Our results can now interpret these par-
562 Journal of Cognitive Neuroscience Volume 10 Number 5
ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

ticular sites as ones probably dedicated to phonologicalencoding From other recent work we know thatphonological encoding not lexical selection is the trou-blemaker in both the tip-of-the-tongue phenomenon(Vigliocco Antonioni amp Garrett 1997) and in at least onetype of anomia (Badecker Miozzo amp Zanuttini 1995)Our results point to Wernickersquos area as the cortical siteof these kinds of trouble These results also complementexisting evidence for the involvement of the left supe-rior temporal gyrus in auditory phonological processing(Boatman Lesser amp Gordon 1995 Deacutemonet et al 1992)It should be noticed though that in their PET study ofpicture and color naming Price et al (1996) did notobserve any activation in Wernickersquos area for phonologi-cal retrieval But that negative result may have been dueto the subtraction used In their experiment the controltask for picture or color naming was just viewing thepicture and saying ldquoyesrdquo Preparing the word ldquoyesrdquo needsphonological encoding just as well as preparing a pic-ture or color name Finally the present data agree withthe already mentioned ERP results of Abdullaev andPosner (1997) a Wernicke area activation in the verbgeneration task We can now interpret that nding as dueto phonological encoding of the response word
Still one should be careful not to draw the reverseconclusion (ie to argue from our data that the solefunction of Wernickersquos area in word production is tophonologically encode the word) The red curve in Fig-ure 6 shows that on average dipoles in the criticalcluster show an activation pattern that transcends thetime window for phonological encoding which couldsignal the involvement of Wernickersquos area in temporallyadjacent processes To be precise the rise and fall to 25levels of activation in the cluster encompass a time spanof 250 msec quite a bit more than the estimated 125-msec duration of phonological encoding Although thismay be due to normal variability in the data over trialsand subjects or to some temporal overlap of phonologi-cal encoding in Wernickersquos area with pre- and post-phonological encoding in nearby cortical areas oneshouldnrsquot exclude the possibility that Wernickersquos area isalso involved with other aspects of the production proc-ess A particularly relevant aspect is self-monitoring (seeFigure 1) which is indeed temporally contiguous withphonological encoding It is known that self-monitoringtakes self-produced overt speech as input as well asldquointernal speechrdquo more precisely the phonological code(Wheeldon amp Levelt 1995) Self-monitoring supposedlyinvolves the speakerrsquos normal speech comprehensionsystem (Levelt 1989) Recent PET studies by McGuireSilbersweig and Frith (1996) have indeed shown thatself- and other-monitoring of overt speech involve thesame temporal lobe areas Monitoring of self-producedovert speech cannot be discerned in the data analyzedhere because we didnrsquot include the stage of overt articu-lation But internal speech monitoring can begin duringthe phase of phonological encoding (ie as early as 275
msec after picture onset) This may involve Wernickersquosarea which is known to be essential to speech compre-hension (Deacutemonet et al 1992) In addition McGuireet al found right temporal lobe activation in self-monitoring That may also have been the case for oursubjects involving some or all of the red- and green-marked right temporal lobe sources in Figure 5
More generally these considerations show that activesources cannot be univocally assigned to one compo-nent process or another when these processes overlapin time However using as paradigms well-dened taskssuch as picture naming for which explicit extensivelytested psycholinguistic processing theories are availableit becomes possible to suggest correlations betweendistinct stages of cognitive processing and cortical dy-namics
METHODS
Subjects
The subjects were eight healthy right-handed students(four male four female) between 21 and 30 years of age(mean 24) All subjects had participated in earlier psy-cholinguistic experiments using reaction time methodsand EEG recordings They were all native speakers ofDutch and had normal or corrected-to-normal eyesight
Materials
The experimental materials included 4 practice and 42experimental pictures (see Figure 7) Half the experi-mental pictures had high-frequency and the others low-frequency names (mean word form frequenciesaccording to CELEX 9996 versus 408 per million) Allpicture names were monosyllabic The items constituteda selection from the set used in Jescheniak and Leveltrsquos(1994) study of word frequency effects The pictureswere presented as white line-drawings on a black back-ground All pictures were drawn to t into a quadraticframe with sides corresponding to a visual angle of 6degon the subjectrsquos screen
With these materials two pretests were carried outThe goal of the rst pretest was to establish that thefrequency effect was stable across a fairly large numberof item repetitions Ten subjects named each of theexperimental pictures 12 times At the beginning of theexperiment the subjects studied a booklet that includedthe drawings and the names they should use to referto them For each subject a different test list of 12blocks was created In each block all experimental pic-tures occurred once The order of the pictures within ablock was random and different for each block andsubject The rst test block was preceded by presenta-tion of the practice items There was a short break afterthe sixth block The seventh block was again precededby the four practice items On each trial a xation cross
Levelt et al 563
was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

was presented for 200 msec After a blank interval of 600msec a drawing was presented until a voice key wastriggered by the onset of the subjectrsquos naming responseMaximum picture presentation time was 1300 msec Thenext trial began 1300 msec after speech onset or after
2600 msec if no response was made within the responseinterval
A highly signicant frequency effect of 31 msec wasobtained (means 607 versus 576 msec F1(1 9) = 4084MSE = 1384 F2(1 40) = 1239 MSE = 9697 both p lt
Figure 7 The 24 experimen-tal pictures with their high- orlow-frequency Dutch namesthe 18 llers and the 4 prac-tice items that were used inthe main experiment and therst pretest
564 Journal of Cognitive Neuroscience Volume 10 Number 5
001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

001) The interaction of blocks and frequency was notsignicant Thus the frequency effect was stable across12 picture repetitions (see Figure 3)
The goal of the second pretest was to rule out thatthe frequency effect was due to differences in visualandor conceptual processing of the pictures Ten sub-jects performed an object decision task on the experi-mental pictures and a set of 42 drawings of nonobjectssimilar to those used in Kroll and Potterrsquos (1984) studyof object naming On each trial the subject indicated bypressing one of two response buttons whether or notthe drawing represented an existing object
The experiment included 12 test blocks in each ofwhich all objects and nonobjects appeared once Deci-sion times were faster by 7 msec for high-frequency thanfor low-frequency pictures This difference was signi-cant by subjects but not by items (means 475 versus482 msec F1(1 9) = 713 MSE = 429 p lt 005 F2(140) = 101 MSE = 6903) The interaction of frequencyand blocks was not signicant Thus the pictures withhigh-frequency names were slightly easier to recognizethan those with low-frequency names But it is also clearthat the 31-msec word frequency effect obtained in thepretest was largely independent of visualconceptualprocessing
The main experiment included the 42 experimentalitems of the pretest However just to be sure not toinvolve any frequency-dependent visualconceptualpreparation we identied a subset of 12 high- and 12low-frequency items that were matched for decisiontimes in the second pretest and all analyses wereconned to these pictures (see Figure 7) (To identify thematched subset pairs of high- and low-frequency pic-tures were formed that did not differ by more than 5msec in average decision time Those pictures for whichno decision-time matched partner could be found wereexcluded) The mean word form frequencies for thehigh- and low-frequency items in the subset were 11554and 412 per million Reanalyses of the results of the rstpretest showed that the frequency effect for these itemswas 37 msec (means 573 versus 610 msec F1(1 9) =3403 MSE = 2380 F2(1 22) = 1318 MSE = 7306 bothp lt 001) The interaction of frequency and blocks wasnot signicant (see Figure 7) In analyses per subjectseven out of ten subjects showed a signicant (p lt 005)frequency effect
Task Procedure
Before the start of the measurements subjects studiedthe pictures and their names from a sheet of paper asthe subjects in the rst pretest had done They wereinstructed to name the pictures as fast as possible Sub-sequently they were brought into the shielded MEGroom and seated before a white screen (distance 80 cm)on which the stimuli were projected by a Sony VPL-350QM LCD data projector The trial structure was exactly
the same as in the rst pretest Each subject receivedone training block to get acquainted with the procedureand to ensure a clear pronunciation without too muchmovement of jaw and mouth The experimental sessionincluded 12 blocks and was divided in two sessions of6 blocks each separated by a short break of about 10min Each block had a duration of about 2 min and 20sec
Recording Procedure
The subjects were tested individually in a magneticallyshielded room Magnetic signals were recorded with a122-channel whole-head magnetometer (Neuromag-122Ahonen et al 1993) using a bandpass of 003 to 90 HzThe signals were recorded continuously digitized at arate of 297 Hz and stored on magneto-optical disks foroff-line analyses Along with the magnetic signals theelectrooculogram (EOG) was recorded using bipolarhorizontal and vertical EOG derivations
For the alignment of MEG and magnetic resonanceimage (MRI) data three coils were attached to the sub-jectrsquos head Coil positions relative to nasion andpreauricular points were measured with a 3-D digitizer(Isotrak 3S1002 Polhemus Navigation Sciences Col-chester VT) The coil positions with respect to the mag-netometer were determined from the magnetic signalsproduced by the coils MRIs were obtained with a 15 TSiemens Magnetom
MEG Data Analysis
Evoked magnetic elds following picture presentationand preceding the naming response were averagedstimulus-locked relative to picture onset Trials with EOGactivity exceeding 150 mV were excluded from the aver-age (~10) The averaged epochs encompassed -100 to900 msec The data consisted of averages over 110 to144 trials yielding waveforms with good signal-to-noiseratio (which were also highly reproducible between thetwo measurement sessions) The procedure involved theidentication of the dipolar eld patterns by scanningthrough the data (ie the magnetic eld patterns esti-mated from the measured values at each sensor cfHaumlmaumllaumlinen 1995) Stable eld patterns were modeledat the time instant at which they were clearest andshowed least interference with other source areas bytting an equivalent current dipole (ECD) to the localeld pattern measured by a subset of sensors The 3-Dlocation of an ECD represents the center of gravity ofthe active area and its amplitude and orientation esti-mate the magnitude and average direction of the postsy-naptic currents within the area The ECDs thus obtainedwere integrated in a multidipole source model derivedby tting the dipoles to the entire spatiotemporal eldpattern keeping the locations and orientations of thesources xed (Haumlmaumllaumlinen Hari Ilmoniemi Knuutila amp
Levelt et al 565
Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

Lounasmaa 1993) The source analyses employed aspherical head model of homogeneous conductivity Theresulting source models typically explained 80 to 90 ofthe variance in the data with the best t usually in thetime range between 200 and 500 msec Occasionallytting the dipoles to the entire eld pattern resulted inmore than a single peak activation for a dipole We tookthis seriously because there is no a priori theoreticalreason why a particular cortical area would not be reac-tivated at a later stage in the process the serial modelonly excludes reactivation through feedback We usedthe following criterion for multiple activation of a dipoleA second moment of activation is only reported if therelevant peak in the waveform exceeds 75 of the rstmaximum and the two peaks are separated by at least100 msec and the activity returns to baseline level dur-ing this interval Using this criterion four source wave-forms with two distinct maxima were identied in ourdata
Acknowledgments
This study was supported by the Academy of Finland and bythe EUrsquos Human Capital and Mobility program through theLarge-Scale Facility BIRCH at the Low Temperature LaboratoryHelsinki University of Technology We are grateful to the lateEddy Roskam for advice on the analyses of variance to PeterWittenburg for preparing the data for the analyses of varianceto Ger Desserjer and Godelieve Hofstee for assistance in pre-paring the stimulus materials for running the pre- and post-experiments and for assistance in the data analysis to KimmoUutela for preparing the stimulus sequence in Helsinki to RiittaHari for comments on the manuscript and to Chris Frith forcomments on the right parietal lobe cluster
Reprint requests should be sent to Willem J M Levelt MaxPlanck Institute for Psycholinguistics PO Box 310 6500 AHNijmegen The Netherlands or via e-mail pimmpinl
REFERENCES
Abdullaev Y G amp Posner M I (1997) Time course of activat-ing brain areas in generating verbal associations Psycho-logical Science 8 56ndash59
Ahonen A L Haumlmaumllaumlinen M S Kajola M J KnuutilaJ E T Laine P P Lounasmaa O V Parkkonen L T Si-mola J T amp Tesche C D (1993) 122-Channel SQUID in-strument for investigating the magnetic signals from thehuman brain Physica Scripta T49 198ndash205
Badecker W Miozzo M amp Zanuttini R (1995) The two-stage model of lexical retrieval Evidence from a case ofanomia with selective preservation of grammatical genderCognition 57 193ndash216
Boatman D Lesser R P amp Gordon B (1995) Auditoryspeech processing in the left temporal lobe An electricalinterference study Brain amp Language 51 269ndash290
Buumllthoff H H amp Edelmann S (1992) Psychophysical sup-port for a two-dimensional view interpolation theory ofobject recognition Proceedings of the National Academyof Sciences of the United States of America 89 60ndash64
Damasio H Grabowski T J Tranel D Hichwa R D ampDamasio A R (1996) A neural basis for lexical retrievalNature 380 499ndash505
Dehaene S (1996) The organization of brain activations innumber comparison Event-related potentials and the addi-tive-factors method Journal of Cognitive Neuroscience 847ndash68
Dell G S (1986) A spreading activation theory of retrievalin sentence production Psychological Review 93 283ndash321
Dell G S amp OrsquoSeaghdha P G (1991) Mediated and conver-gent lexical priming in language production A commenton Levelt et al Psychological Review 98 604ndash614
Dell G S Schwartz M F Martin N Saffran E M amp Gag-non D A (1997) Lexical access in normal and aphasicspeech Psychological Review 104 801ndash838
Deacutemonet J-F Chollet F Ramsay S Cardebat D NespoulosJ Lamp Wise R (1992) The anatomy of phonological and se-mantic processing in normal subjects Brain 115 1753ndash1768
Dronkers N F (1996) A new brain region for coordinatingspeech articulation Nature 384159ndash161
Faillenot I Toni I Decety J Greacutegoire M-C amp JeannerodM (1997) Visual pathways for object-oriented action andobject recognition Functional anatomy with PET CerebralCortex 7 77ndash85
Fink G R Halligan P W Marshall J C Frith C D Frack-owiak R S J amp Dolan R J (1996) Where in the braindoes visual attention select the forest and the trees Na-ture 382 626ndash628
Friston K J Price C J Fletcher P Moore C FrackowiakR S J amp Dolan R J (1996) The trouble with cognitivesubtraction Neuroimage 4 97ndash104
Glaser W R (1992) Picture naming Cognition 42 61ndash105Haglund M M Berger M S Shamseldin M Lettich E amp
Ojemann G A (1994) Cortical localization of temporallobe language sites in patients with gliomas Neurosur-gery 34 567ndash576
Haumlmaumllaumlinen M (1995) Functional localization based on meas-urements with a whole-head magnetometer system BrainTopography 7 283ndash289
Haumlmaumllaumlinen M Hari R Ilmoniemi R J Knuutila J ampLounasmaa O V (1993) MagnetoencephalographymdashTheory instrumentation and applications to noninvasivestudies of the working human brain Reviews of ModernPhysics 65 413ndash497
Hart J amp Gordon B (1992) Neural subsystems for objectknowledge Nature 359 60ndash64
Humphreys G W Lam C amp Lloyd-Jones T J (1995) An in-teractive activation approach to object processing Effectsof structural similarity name frequency and task in normal-ity and pathology Memory 3 535ndash586
Humphreys G W Riddock M J amp Quinlan P T (1988) Cas-cade processes in picture identication Cognitive Neuro-psychology 5 67ndash103
Husain M Shapiro K Martin J amp Kennard C (1997) Ab-normal temporal dynamics of visual attention in spatial ne-glect patients Nature 385 154ndash156
Jescheniak J D amp Levelt W J M (1994) Word frequency ef-fects in speech production Retrieval of syntactic informa-tion and of phonological form Journal of ExperimentalPsychology Learning Memory and Cognition 20 824ndash843
Klingberg T Kawashima R amp Roland P E (1996) Activa-tion of multi-modal cortical areas underlies short-termmemory European Journal of Neuroscience 8 1965ndash1971
Kroll J F amp Potter M C (1984) Recognizing words pic-tures and concepts A comparison of lexical object and re-ality decisions Journal of Verbal Learning and VerbalBehavior 23 39ndash66
566 Journal of Cognitive Neuroscience Volume 10 Number 5
Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567

Levelt W J M (1983) Monitoring and self-repair in speechCognition 14 41ndash104
Levelt W J M (1989) Speaking From intention to articula-tion Cambridge MA MIT Press
Levelt W J M (1992a) Accessing words in speech produc-tion Stages processes and representations Cognition 421ndash22
Levelt W J M (1992b) The perceptual loop theory not dis-conrmed A reply to MacKay Consciousness amp Cogni-tion 1 226ndash230
Levelt W J M (1996) Perspective taking and ellipsis in spa-tial descriptions In P Bloom M A Peterson L Nadel ampM F Garrett (Eds) Language and space (pp 77ndash107)Cambridge MA MIT Press
Levelt W J M Roelofs A amp Meyer A S (in press) A theoryof lexical access in speech production Behavioral andBrain Sciences
Levelt W J M Schriefers H Vorberg D Meyer A S Pech-mann T amp Havinga J (1991) The time course of lexicalaccess in speech production A study of picture namingPsychological Review 98 122ndash142
Levelt W J M amp Wheeldon L (1994) Do speakers have ac-cess to a mental syllabary Cognition 50 239ndash269
MacKay D A (1992) Awareness and error detection Newtheories and research paradigms Consciousness amp Cogni-tion 1 199ndash225
Martin A Wiggs C L Ungerleider L G amp Haxby J V(1996) Neural correlates of category-specic knowledgeNature 649ndash652
Martin N Gagnon D A Schwartz M F Dell G S amp Saf-fran E M (1996) Phonological facilitation of semantic er-rors in normal and aphasic speakers Language andCognitive Processes 11 257ndash282
McGuire P K Silbersweig D A amp Frith C D (1996) Func-tional neuroanatomy of verbal self-monitoring Brain 119907ndash917
Ojemann G A (1983) Brain organization for language fromthe perspective of electrical stimulation mapping Behav-ioral and Brain Sciences 6 189ndash206
Ojemann G A Ojemann J Lettich E amp Berger M (1989)Cortical language localization in left dominant hemi-sphere Journal of Neurosurgery 71 316ndash326
Oldeld R C amp Wingeld A (1965) Response latencies innaming objects Quarterly Journal of Experimental Psy-chology17 273ndash281
Peneld W amp Roberts L (1959) Speech and brain mecha-nisms Princeton NJ Princeton University Press
Peterson R R amp Savoy P (1998) Lexical selection andphonological encoding during language production Evi-dence for cascaded processing Journal of ExperimentalPsychology Learning Memory and Cognition 24 539ndash557
Pollen D A amp Trachtenberg M C (1972) Some problems ofoccipital alpha block in man Brain Research 41 303ndash314
Potter M C (1983) Representational buffers The see-mindhypothesis in picture perception reading and visualsearch In K Rayner (Ed) Eye movements in reading Per-ceptual and language processes (pp 423ndash437) New YorkAcademic Press
Potter M C amp Faulconer B A (1975) Time to understandpictures and words Understanding without words Na-ture 253 437ndash438
Price C J Moore C J Humphreys G W FrackowiakR S J amp Friston K J (1996) The neural regions sustain-ing object recognition and naming Proceedings of theRoyal Society London B 263 1501ndash1507
Roelofs A (1992) A spreading-activation theory of lemma re-trieval in speaking Cognition 42 107ndash142
Roelofs A (1997) The WEAVER model of word-form encod-ing in speech production Cognition 64 249ndash284
Roelofs A amp Meyer A S (1998) Metrical structure in plan-ning the production of spoken words Journal of Experi-mental Psychology Learning Memory and Cognition24 1ndash18
Rosch E Mervis C B Gray W Johnson D amp Boyes-BraemP (1976) Basic objects in natural categories Cognitive Psy-chology 8 382ndash439
Salmelin R Hari R Lounasmaa O V amp Sams M (1994) Dy-namics of brain activation during picture naming Nature368 463ndash465
Salmon E Van der Linden M Collette F Delore GMaquet P Degueldre C Luxen A amp Franck G (1996) Re-gional brain activity during working memory tasks Brain119 1617ndash1625
Sternberg S (1969) The discovery of processing stages Ex-tensions of Dondersrsquo method In W G Koster (Ed) Atten-tion and performance Vol 2 (pp 276ndash315) AmsterdamNorth-Holland
Thorpe S Fize D amp Marlot C (1996) Speed of processingin the human visual system Nature 381 520ndash522
Ungerleider L G (1995) Functional brain imaging studies ofcortical mechanisms for memory Science 270 769ndash775
van Turennout M Hagoort P amp Brown C M (1997) Elec-trophysiological evidence on the time course of semanticand phonological processes in speech production Jour-nal of Experimental Psychology Learning Memory andCognition 23 787ndash806
Vigliocco G Antonioni T amp Garrett M F (1997) Grammati-cal gender is on the tip of Italian tongues PsychologicalScience 8 314ndash317
Wheeldon L amp Levelt W J M (1995) Monitoring the timecourse of phonological encoding Journal of Memoryand Language 34 311ndash334
Levelt et al 567





























