Embed Size (px)
Citation preview

/ . Embryol. exp. Morph. Vol. 28, 2, pp. 279-312, 1972 2 7 9
Printed in Great Britain
An investigation of inner cell massand trophoblast tissues following their isolation
from the mouse blastocyst
By R. L. GARDNER1
With an Appendix by M. H. JOHNSON1
From the The Physiological Laboratory, Cambridge
SUMMARY
1. Inner cell mass (ICM) and trophoblast were isolated from 3^-day post-coitum mouseblastocysts by microsurgery.
2. Trophoblastic fragments formed vesicles in culture but did not aggregate with other suchfragments. They proved as effective as intact blastocysts in inducing decidua in recipient uteri,but thereafter failed to proliferate.
3. Isolated ICMs remained as solid balls of cells that readily aggregated in pairs or groupsin culture but failed to induce implantation changes in receptive uteri.
4. Possible explanations for the failure of isolated trophoblast to proliferate after implan-tation are discussed. It is argued that presence of ICM tissue is necessary for trophoblastproliferation, and suggested that the ICM exerts its effect by controlling development of theectoplacental cone.
INTRODUCTION
One of the earliest differences to arise between cells of the mammalian embryois evident at the blastocyst stage with formation of the trophectoderm and innercell mass (ICM). Attention has been devoted to establishing whether thisdifferentiation is determined in the egg prior to division (Dalcq, 1957; Seidel,1960; Mulnard, 1965) or by the relations between naive blastomeres duringcleavage (Mintz, 1965; Tarkowski & Wroblewska, 1967; Graham, 1971). Thequestion of the particular properties that serve to distinguish the two tissues inthe early blastocyst has so far been largely neglected.
Earlier studies involved disaggregation of morulae and blastocysts of both therabbit and mouse, and culture of the cells isolated from them (Cole & Paul, 1965;Cole, Edwards & Paul, 1965). Since no attempt was made to separate ICM andtrophoblast tissue prior to disaggregation, the cells could not be assigned un-equivocally to one or other type.
The present study was thus aimed at separating the blastocyst into its con-stituent tissues, and comparing their properties both in culture and after transferto the uterus. This project promised to provide criteria for distinguishing these
1 Authors' address: Physiological Laboratory, Cambridge, CB2 3EG, U.K.

280 R. L. GARDNER AND M. H. JOHNSON
Table 1. Viability of blastocysts transferred after 8^—11 h at room temperature inphosphate-buffered M199 + 10% serum medium
(Recipients allowed to deliver young.)
Blastocysts
Hours in vitro No.at room transferred No. of
Recipient code temp. bilaterally young born Comments
R4 (24. 10. 69) 8 | 8 6 —R3 (6. 11.69) 9 10 7 1 dead,
uncertainif still-born
R3(16. 10. 69) 11 10 5 —R4(16. 10. 69) 11 12 8 —
A further three recipients did not become pregnant.
tissues in investigations into the mechanism of their differentiation, and toclarify their roles in implantation and interrelations in later development. Themouse was chosen because it is the species used in most studies of early mam-malian development and because its blastocyst is very resilient to microsurgery(Gardner, 1968, 1971, 1972; Lin, 1969). A preliminary report of part of thiswork has been published elsewhere (Gardner, 1971).
Previous accounts of the handling and transfer of so-called 'pure' trophoblastof murine origin have dealt exclusively with the ectoplacental cone of the im-planted egg-cylinder (Grobstein, 1950; Billington, 1965; Simmons & Russell,1966; Clarke, 1969). Despite the enveloping cell layer of the unimplanted blasto-cyst being called trophoblast its relationship with the ectoplacental cone isuncertain. This note of caution is justified by the following results.
MATERIALS AND METHODS
Preparation of donor females and recovery of blastocysts
Adult or 3- to 4-week-old PDE random-bred or Er inbred albino mice weresuperovulated by intraperitoneal injection of pregnant mares' serum gonado-trophin (PMSG - Gestyl, Organon) followed 48 h later by human chorionicgonadotrophin (HCG-Pregnyl, Organon). Five i.u. of each hormone were givento adults and 10 i.u. to the immature female mice. PDE females were pairedwith males of the same or CBA/H-T6 inbred strain, and Er females exclusivelywith CBA/H-715 males after administration of HCG. Females with plugs werekilled and their excised uterine horns flushed for blastocysts either between96-102 or 117-122 h after HCG. Since no consistent differences were notedbetween material obtained at opposite extremes of each time range, blastocysts

Inner cell mass and trophoblast tissues 281
Fig. 1. Schematic side view of hanging drop in manipulation chamber to showarrangement of blastocyst and fragment of razor blade for sectioning blastocyst toobtain pure trophoblast. c.s. = coverslip, i.c.m. - inner cell mass, r.b. = fragmentof razor blade, s.p. = suction pipette.
recovered during the former interval (and tissue derived from them) will becalled 3^-day and those recovered during the latter interval will be called 4^-dayblastocysts or fragments.
Conditions for storage, manipulation and culture of blastocysts and fragments
The special conditions imposed by microsurgery required modified methodsfor keeping blastocysts in vitro. The medium could not be maintained in equi-librium with an atmosphere of 5 % CO2 in air. Also, attempts to perform manipu-lations at 37 °C were unsuccessful because of the great increase in adherence ofcells and debris to the instruments at higher temperature. Manipulations werethus carried out at room temperature in medium M 199 plus 10 % inactivatedfoetal calf serum (Microbiological Associates Inc., or Flow Laboratories,Scotland), which contained 100 i.u./ml of sodium benzyl penicillin and wasbuffered at pH 7-0 with 5-8 % (v/v) Sorensen's phosphate buffer (M/15). TO
avoid unnecessary osmotic shocks or changes in pH this medium was usedthroughout. Under these conditions intact blastocysts remained viable for upto 11 h at room temperature (Table 1).
Microsurgery
A Leitz micromanipulator was used. Blastocysts were placed in a drop ofmedium hanging from the coverslip of a chamber (Puliv, Leitz) filled with heavyliquid paraffin. Microsurgery was carried out at x 125, x 300 or x 500 usingbright-field or Heine phase-contrast observation in a Leitz Laborlux microscope.
Trophoblast tissue was obtained free from ICM as follows. Expanded 3^-dayblastocysts surrounded by intact zonae were orientated and held by suctionpipette against the underside of the coverslip of the hanging drop (Fig. 1). A pieceof the cutting edge of a safety razor-blade attached to a mounted needle with

282 R. L. GARDNER AND M. H. JOHNSON
KquaulYasiment
Residual trophoblastdiscarded
Maximal trophoblastfiat: me nt
4- - A
Hquatorial inner cellmass fragment
Minimal inner eelmass fragment
Dissected innercell mass
Fig. 2. Scheme of manipulations carried out on the mouse blastocyst to obtaintrophoblast and ICM tissue. D, Trophoblast tissue; • , inner cell mass tissue.(From Gardner, 1971, with permission of F. Vieweg & Sohn.)
Araldite (CIBA) was arranged vertically on one manipulator unit so that itscutting surface was parallel with the coverslip of the chamber (Fig. 1). Byraising the blade slowly the living blastocyst could be severed, parallel to thesurface of the ICM, either equatorially (to yield equatorial trophoblasticfragments, Fig. 2 A) or close to the embryonic pole (to yield maximal tropho-blastic fragments, Fig. 2B). Sectioning blastocysts through the equator presentedno difficulty. The embryo tended to rotate away from the advancing blade whenit was cut towards one pole. This could be prevented by increasing suction or byusing a needle to help immobilize the embryo. Provided the fragment of razor-blade had been cleaned, siliconed and correctly aligned, blastocysts could besevered with only occasional recourse to the needle to clear one or other frag-ments from the blade. The choice of well-expanded blastocysts enabled tropho-blast to be obtained with little risk of inclusion of ICM cells.
Preliminary attempts to isolate ICM tissue from blastocysts by variouschemical and enzymatic treatments were unsuccessful. ICMs were thus obtainedby penetrating immobilized expanded blastocysts from opposite sides with a pairof fine, siliconed glass needles, tearing the trophoblast open and pinning it outas a sheet against the coverslip. The exposed ICM was then gently scraped fromthe trophoblast, but was only used if the sheet of overlying trophoblast leftbehind was intact. It often proved possible to isolate ICMs relatively freefrom cells of the enveloping trophoblast. However, the procedure extensivelydamaged the corresponding trophoblast, so comparison between trophoblastand ICM tissues from the same blastocyst was not possible.

Inner cell mass and trophoblast tissues 283
Culture of trophoblastic fragments, isolated 1CMs and intact blastocysts
Trophoblastic fragments and intact blastocysts were cultured either in hangingdrops in the oil chambers used for microsurgery, or in microdrops on siliconedcoverslips placed in culture dishes (Falcon Plastic, 60 x 15 mm) filled with heavyliquid paraffin. Isolated ICMs were generally very small and delicate and weretherefore left in the manipulation chambers for culture. Fusion was encouragedby pairing fragments with a blunt glass needle, and with the warm stage ofthe microscope set at 37 °C. This allowed firm contact to be made betweenfragments before they were moved to the incubator.
Cell counts on blastocysts and isolated ICMs
The material was prepared according to the air-drying method of Tarkowski(1966).
Transfer of tissue and blastocysts to the uteri of recipient mice
The 3|-day blastocysts, trophoblastic fragments or isolated ICMs weretransferred to the uteri of mice on the third day of pseudopregnancy, except inone case where a group of trophoblastic fragments were transferred to a recipienton the fourth day. The recipients were albino PDE or Er mice mated withvasectomized males. All ICMs and some trophoblastic fragments were trans-ferred unilaterally, and intact blastocysts introduced into the contralateral hornsserved as a control for uterine reactivity. Uterine transmigration of embryos isa possible complication in such experiments (Runner, 1951; McLaren &Michie, 1954). However, it was not found among 36 foetuses examined followingtransfer of blastocysts of pigmented and albino genotypes to opposite hornsof 6 recipients, nor among 63 implants developed from blastocysts transferredunilaterally to a further 18 recipients (unpublished observations).
Induction of deciduomata
Intraluminal injection of arachis oil between 15.00 and 16.00 h on the fourthday of pseudopregnancy was employed for this purpose (Finn, 1965).
Examination of the transferred tissue
Recipient females that received trophoblast tissue were examined between3 and 8 days after transfer. Horns carrying decidual swellings were fixed inSousa or Bouin's fixative. The dehydrated tissue was cleared in supercedrol(G. T. Gurr), embedded in paraffin wax and serially sectioned at 6-10 jam. Thesections were stained with haemalum and eosin and mounted in DPX (G. T.Gurr).
Females receiving ICM tissue were given i.v. pontamine sky blue 6 BX (G. T.19 EMB 28

284 R. L. GARDNER AND M. H. JOHNSON
Gurr) approximately 48 h after transfer of the tissue to determine whether im-plantation changes had been initiated (Psychoyos, 1961; Finn & McLaren, 1967).
Deciduomata and implants derived from intact blastocysts were processedhistologically as above.
Assessment of implantation rates by counting decidual swellings-complicatingfactors
More than one implant may occur within a single decidual swelling (author'sunpublished observations). This phenomenon would not be detected in caseswhere derivatives of the ICM are poorly represented or totally lacking (as innearly all implants obtained following transfer of trophoblast). Counts ofdecidual swellings would thus tend to underestimate the rate of implantation.
Decidualization can also occur from the inevitable trauma accompanying thetransfer operation, and inclusion of these swellings would lead to an overestimateof the implantation rate. However, in two cases where such deciduomata wereidentified with certainty, i.e. in horns where all transferred blastocysts had givenrise to decidual swellings containing embryos or embryonic tissues, there was noquestion of their being confused with normal decidua. They were found at theoviducal end of their respective horns (i.e. at a point corresponding to thetransfer site) and were less than half the normal diameter. One was sectioned, andwas not only completely without embryonic tissues but very poorly orientatedin relation to the mesometrial-antimesometrial axis of the uterus. During thisand a succeeding study (in preparation), further examples of probable traumaticdeciduomata were encountered that showed all the above features, particularlythe small size and poor orientation. Only once did a probable deciduomaapproach the size of an adjacent decidual swelling. Yet even here the orientationwas atypical, being exactly perpendicular to the mesometrial-antimesometrialaxis. So, although these possible complications must be borne in mind theireffect on the results will be slight.
RESULTS
1. Observations on trophoblastic fragments in culture
The following remarks apply to both maximal and equatorial fragments(Fig. 2 A, B) unless qualified, and are based on study of 139 maximal and 34equatorial fragments.
Initially, the fragments were solid balls of cells with no visible cavity (Fig. 3 A).When placed in culture the majority sealed and began to cavitate within 30 min,becoming obviously vesicular within l^h (Fig. 3B). In a typical series 85%(56/66) cavitated. The remaining 10 may have been damaged during cutting orhandling. Although the vesiculated fragments could cavitate a second time iftorn open, they did not swell to the same size as intact blastocysts.
Twenty-four maximal fragments were placed in contact in 12 pairs and the
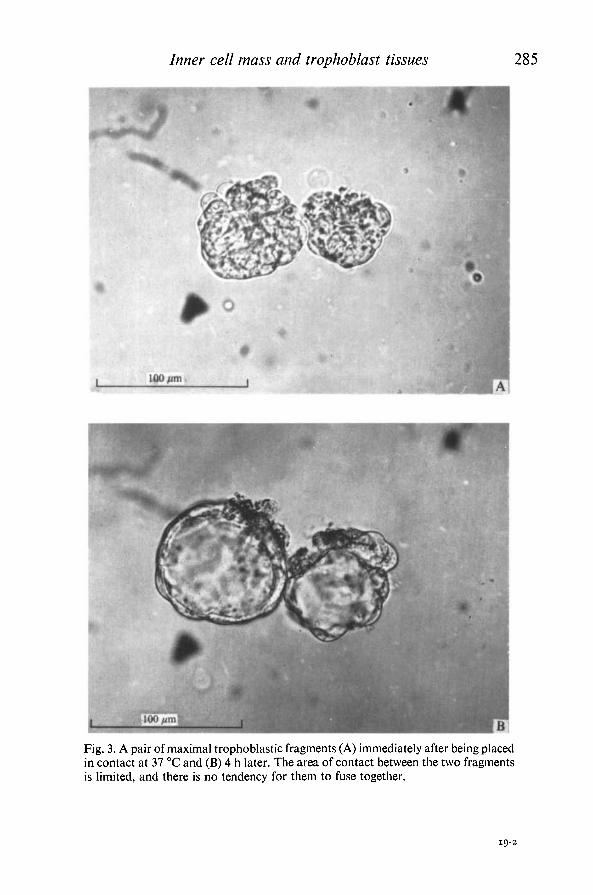
Inner cell mass and trophoblast tissues 285
Fig. 3. A pair of maximal trophoblastic fragments (A) immediately after being placedin contact at 37 °C and (B) 4 h later. The area of contact between the two fragmentsis limited, and there is no tendency for them to fuse together.
19-2

oo ON
Tab
le 2
. Su
mm
ary
of r
esul
ts o
fcul
turi
ng
vari
ous
cate
gori
es o
f fra
gmen
ts
of m
ouse
bla
stoc
ysts
in
cont
act
at 3
7 °C
Exp
eri-
men
tal
seri
es
Bla
stoc
yst
frag
men
ts p
lace
d in
con
tact
Cat
egor
y an
d/?.
c. a
geNo
. an
d ge
noty
pe
No.
ofpa
irs
orgr
oups
in s
erie
s
Post
-cul
tura
lm
orph
olog
y
No.
fuse
dNo
.no
t fu
sed
App
rox.
tim
efo
r su
cces
sful
fusi
on (
h)C
omm
ents
o o d S o a O
II III
IV V VI
VII
3^-d
ay I
CM
s of
sam
ege
noty
pe -
pai
rs
3i-d
ay I
CM
s of
diff
eren
tge
noty
pe -
pai
rs3^
-day
IC
Ms
of s
ame
geno
type
- g
roup
s
2 P
DE
2C
BA
HT
6x
PD
E
1 P
DE
+ 1
CB
AH
T 6
x P
DE
3 P
DE
2 fu
sed
PD
E +
2 f
used
PD
E*
3 C
BA
HT
6xP
DE
3 fu
sed
CB
AH
T6
x P
DE
f(a
bove
) + 2
fus
edC
BA
HT
6x P
DE
3^-d
ay I
CM
s of
diff
eren
tge
noty
pe -
gro
ups
4^-d
ay I
CM
s fr
om
T.T.
blas
tocy
sts
of s
ame
geno
type
- p
airs
4^-d
ay I
CM
s fr
om n
on-
T.T.
bla
stoc
ysts
of
sam
ege
noty
pe -
pai
rs4^
-day
IC
Ms
from
non
-T.
T. b
last
ocys
ts o
f sa
me
geno
type
- g
roup
s
2P
DE
+2
CB
AH
T
2E
r2
CB
AH
T6x
Er
2C
BA
HT
6x
PD
E2
PD
E
3E
r4
PD
E
17 1815 14 1
—
5 1 1 7 1 1
4 1 1 7 1 1
2 +
2 failures =
T.C
.1 failure = T
.C.
3 failures = ?
1 fa
ilure
=
T.C.
2/3
ICM
s =
T.
C.U
nder
10i
4 Und
er 8
1
failu
re =
?
Und
er 6
Und
er 7
6

Tab
le 2
. (c
ont.)
Exp
eri-
lllC
Ill
dl
seri
es
VII
I
I X X
Bla
stoc
yst
frag
men
ts Ai
Cat
egor
y an
d p.
c. a
ge
4-H
ay a
nd 3
i-da
y IC
Ms
of d
iffer
ent
geno
type
s -
pair
s
3-̂-d
ay m
inim
al I
CM
frag
men
ts o
f sa
me
geno
type
- p
airs
3^-d
ay m
axim
altr
opho
blas
tic f
ragm
ents
of s
ame
geno
type
- p
airs
plac
ed i
n co
ntac
t
No.
and
gen
otyp
e
1, 4
£CB
AH
T6
x PD
E
(T.T
.)+
1, 3̂
-Er
l,4i
PD
E(T
.T.)
+1
,3i
CB
AH
T6x
PD
E2
CB
A x
PD
E2
PD
E
2CB
AH
T6x
PD
E
No.
of
pair
s or
in s
erie
s
1 8 13 4 12
Post
-cul
tura
lm
orph
olog
y
("N
o
fuse
d 1 4 2— —
A
T\T
n.L
NU
.
not
fuse
d— 4 11 4 12
App
rox.
tim
ef7
"\t"
cn
r'(r *^
a ccfi
I 1
fusi
on (
h)
4i 4 or
und
er
? — —
Com
men
ts
3 fa
ilure
s =
T.C
.1
failu
re =
?— —
*W
^^ 1 f v SO a a <~»
Abb
revi
atio
ns:
T.T.
= b
last
ocys
ts i
n w
hich
abe
mbr
yoni
c po
lar
trop
hobl
ast
had
unde
rgon
e gi
ant
cell
tran
sfor
mat
ion
(Dic
kson
,19
63).
T.C.
= c
onta
min
ated
with
trop
hobl
ast
cells
. T
his
was
infe
rred
fro
m t
he fa
ct t
hat
1 or
mor
e pe
riph
eral
cel
ls h
ad v
esic
ulat
ed,
or b
ecau
se a
sm
all
ecce
ntri
c ca
vity
had
dev
elop
ed.
* T
wo
alre
ady
fuse
d pa
irs
of I
CM
s de
rived
fro
m S
erie
s 1.
f
A g
roup
of
thre
e an
d an
alr
eady
fus
ed p
air
plac
ed i
n co
ntac
t.
oo

OO oo
Tab
le 3
. Im
plan
tati
on
of t
roph
obla
st
and
ICM
ti
ssue
of
3\-d
ay
mou
se b
last
ocys
ts i
n th
e ut
eri
of p
seud
opre
gnan
t m
ice
(Onl
y re
cipi
ents
with
one
or
mor
e im
plan
ts i
nclu
ded.
)
T")
nr
Cat
egor
y
A.
Max
imal
tro
pho-
blas
tic f
ragm
ents
Tot
alB
. E
quat
oria
ltr
opho
blas
tic
frag
men
tsT
otal
C.
Fus
ed p
airs
of I
CM
sT
otal
D.
Inta
ct b
last
o-cy
sts,
in
vitr
ofo
r eq
uiva
lent
ti
me
To
tal
A
1S
Su
e Gen
otyp
e
CB
AH
T6
xP
DE
PD
E
PD
EP
DE
CB
AH
T6
xP
DE
PD
EC
BA
HT
6x
PD
E
PD
EC
BA
HT
6x
PD
E
No
.tr
ansf
erre
d
14 26 40 12 5 13 30 9 8 17 32 20 52
Gen
otyp
e
PD
EP
DE
Er
PD
EE
r
PD
EP
DE
PD
EP
DE
Ute
rine
No
. of
fem
ales
2 3 5 1 1 1 3 2 2 4 6 4 10
fost
er-m
othe
rs
"Mr»
r»
f1N
U.
\JL
reci
pien
tho
rns
3 5 8 2 1 2 5 2 2 4 7 4 11
Day
s p.
c.at
tra
nsfe
r
21 21 21 31 21 21 21 21 21
No
ofde
cidu
alsw
ellin
gs
12 14 26 5 1 2 8 0 0 0 25 17 42
d iTl
> >—i
O o 00 o

Inner cell mass and trophoblast tissues 289
members of each pair held together at 37 °C until they adhered. These pairs offragments remained discrete despite incubation for up to 14 h, though usuallycavitating and showing a limited area of mutual adhesion (Fig. 3 A, B). In no casewas fusion or aggregation observed, and members of a pair could easily beseparated with a glass needle (Table 2, row X).
2. Fate oftrophoblastic fragments transferred to the uteri of pesudopregnant mice
Sixty-five maximal fragments which had been cultured for 2-5£ h were trans-ferred into seven recipients. Thirty-three equatorial fragments, of which onlyfive had been cultured for 2 h, were transferred into a further four recipients.Implantation was assessed by looking for discrete decidual swellings, and onlyfemales with one or more such swellings have been included in Table 3. A pro-portion of both the maximal and equatorial trophoblastic fragments hadimplanted.
All but one of the 34 implants examined 4-7 days after transfer of trophoblastappeared similar (Table 4A, B). Embryo, amnion, allantois, yolk sac, Reichert'smembrane and ectoplacental tissue were absent. A chamber, correspondingroughly to the position the embryo would occupy in a normal implant, hadalmost invariably formed, apparently by pycnosis of the central decidual cells.Occasionally it had a somewhat spongy or reticular appearance due to thepersistence of a network of elongated cells (Fig. 4). In other decidua the cham-ber was virtually acellular, and filled with an homogeneous ground substancebroken up by a complex fibrin-like network (Fig. 5), or even with maternal bloodcells. A second consistent feature, particularly of the older implants, was a com-pact mass of eosinophilic material of unknown origin and composition whichlay between the mesometrial border of the chamber and the zone rich in maternalsinusoids (Figs. 4, 5). This material sometimes attained large dimensions andsmall dispersed foci could occasionally be seen in younger implants. Poly-mo rphonuclear leucocytes were present in and around it.
The third distinctive feature was the presence in three-quarters of the implantsof cells indistinguishable from the trophoblastic giant cells that surroundnormal conceptuses. Their presence constituted the only difference in cellularcomposition between decidua induced by trophoblast and deciduomata evokedby oil. The number of giant cells was similar between 4 and 7 days after transferand was trivial compared with the number found in normal implants of equiv-alent ages (Table 4A, B; Noyes, 1959; Snell & Stevens, 1966). Some of themwere pycnotic, though viable ones persisted in some of the older implants(Fig. 6).
The single exceptional implant consisted of a small yolk-sac-like structuresurrounded by a thickened Reichert's membrane, and was framed by a networkof healthy giant cells (Fig. 7). This implant was presumably derived from a frag-ment contaminated with ICM tissue.
The transferred trophoblast thus only gave rise to a very few giant cells which

Tab
le 4
. Su
mm
ary
of m
ain
feat
ures
of
uter
ine
impl
ants
dev
elop
ed fr
om
trop
hobl
asti
c fr
agm
ents
Exp
erim
enta
l g
rou
p
Age
of
imp
lan
tsin
day
s
Per
iod
in d
ays
betw
een
tran
sfer
and
To
tal
no
.of
No.
w
ith
cent
ral
No.
wit
heo
sino
-ph
ilic
No.
of
impl
ants
acc
ord
ing t
o n
o.
of t
rop
ho
bla
stic
gia
nt c
ells
/im
plan
t
p.c.
au
top
sy
dec
idu
a*
cham
ber
m
ass
1-6
Abu
n-7-
12
13-1
8
dan
tr O > o tn d o O
A.
Eq
uat
ori
al f
ragm
ents
7
i 4
1
—
—
—
—
1
only
tra
nsfe
rred
to
8i
5
7
6
—
15
—
If
—
reci
pien
ts
(sm
all
cham
bers
)
B.
Max
imal
fra
gmen
ts
9i
6
23
23
20
6
15
2
—
—on
ly t
ran
sfer
red
10
i 7
3
3
2
1
1
—
—
1$
to r
ecip
ient
s
C.
Max
imal
tro
ph
ob
last
ic
9i
6
6
6
61
5
—
—fr
agm
ents
to o
ne
ho
rn
10
i 7
6
6
6
2
3of
eac
h o
f a
seri
es o
f H
i 8
12
10
11
8
2
reci
pien
ts r
ecei
ving
inta
ct b
last
ocys
ts i
no
ther
ho
rn
D.
As
C b
ut
whe
re a
ll
Hi
8
3||
3
3
—
3
—
—
—in
tact
bla
stoc
ysts
had
fail
ed t
o i
mpl
ant
* T
his
excl
udes
obv
ious
dec
iduo
mat
a.
t T
hirt
een
out
of
14 g
iant
cel
ls d
egen
erat
ing.
X D
eriv
ativ
es o
f th
e IC
M p
rese
nt (
Fig
. 7)
. § O
ne w
as m
orph
olog
ical
ly n
orm
al f
oetu
s (F
ig.
8 A
, B
).il
The
se t
hre
e im
pla
nts
fou
nd i
n r
ecip
ient
R2 (
14.
5. 7
0) i
n w
hich
int
act
blas
tocy
sts
had
fai
led
to
im
plan
t (s
ee T
abl e
5).
1 —
—
—
—
2§
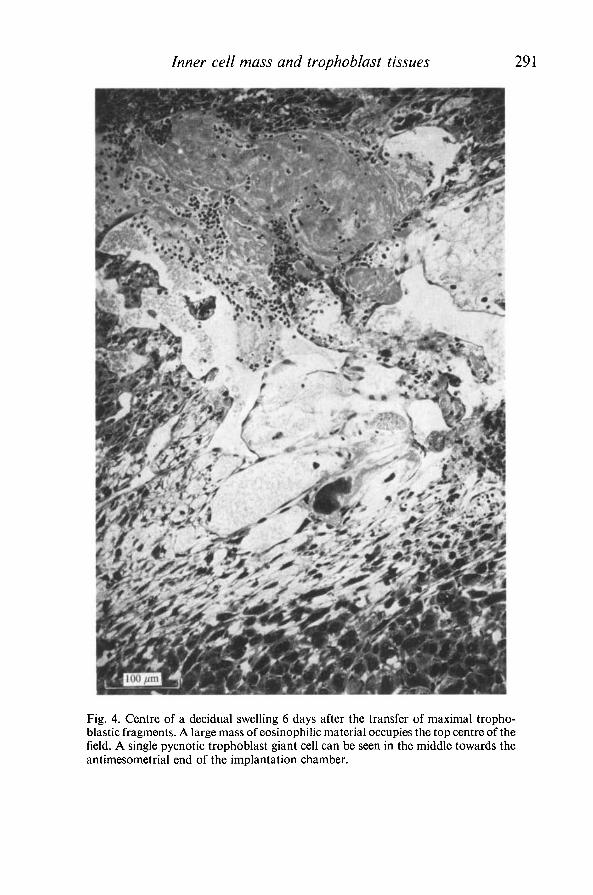
Inner cell mass and trophoblast tissues 291
Fig. 4. Centre of a decidual swelling 6 days after the transfer of maximal tropho-blastic fragments. A large mass of eosinophilic material occupies the top centre of thefield. A single pycnotic trophoblast giant cell can be seen in the middle towards theantimesometrial end of the implantation chamber.

292 R. L. GARDNER AND M. H. JOHNSON
Fig. 5. Another decidual swelling from the same recipient that carried the decidualsite shown in Fig. 4. A large eosinophilic mass lies mesometrially as before. However,the chamber is larger than the previous one and is virtually free of cells.

Inner cell mass and trophoblast tissues 293
Fig. 6. Centre of a decidual swelling 7 days after transfer of maximal trophoblasticfragments. A rather loosely arranged eosinophilic mass occupies most of the centre.The arrow indicates the position of a viable trophoblastic giant cell.

294 R. L. GARDNER AND M. H. JOHNSON
Fig. 7. Exceptional trophoblastic implant 7 days after transfer of maximal tropho-blastic fragments. Note the abundant giant cells, absence of ectoplacental cone, andthe central vesicle consisting of a thick Reichert's membrane lined presumably withcells of the distal endoderm. (Mesometrium towards the top of the figure.)

Inner cell mass and trophoblast tissues 295
were never observed in division. One explanation for the failure of the tropho-blast to proliferate might be that the endocrine status of the recipients differs inthese experiments from that of normal pregnancy. Hence the following experi-ments were undertaken to determine whether trophoblastic fragments developedsimilarly in recipients made pregnant by simultaneously transferring intactblastocysts.
3. Transfer of trophoblastic fragments and intact blastocysts to opposite horns ofpseudopregnant mice
Thirty-six maximal trophoblastic fragments were transferred unilaterally tosix females, and 5-6 blastocysts of the same age and genotype were injected intothe contralateral horns. Five recipients had implants at autopsy. It was foundthat trophoblastic fragments devoid of ICM tissue are as effective decidualstimuli as whole blastocysts (Table 5 A).
All implants developed from intact blastocysts contained foetuses. With twoexceptions, the form of the trophoblastic implants was similar to those describedpreviously as regards central chamber, eosinophilic mass and scarcity oftrophoblastic giant cells (Table 4C). The absence of giant cells from most of theoldest implants in this series suggests that death or dispersal of these cells mayoccur 7-8 days after transfer.
The two implants which were not typical of transferred trophoblastic frag-ments were adjacent in one horn of a recipient killed 8 days after transfer. Onecontained a well-developed foetus and the other a mass of giant cells (Fig. 8 A, B).The former might have been due to regulation by a fragment from which allICM cells had not been excluded or to accidental transfer of a blastocyst. It wasprobably responsible for the occurrence of the second exception. Thus the giantcells of the first implant were not enclosed by viable decidual tissue on the sidebordering the second implant, since a wide track of entirely pycnotic decidualtissue extended to the centre of the latter (Fig. 8B). Though no giant cells werenoted actually in the boundary zone they were close enough on both sides tosuggest very strongly that they had invaded the second implant rather than havingoriginated there.
One recipient had implants in the experimental horn only (Table 5) and theseconformed to the typical pattern (Table 4D).
In a further experiment a single recipient (Rx (29. 5. 70), Table 5B) was killedonly 3 days after transfer of blastocysts to one horn and trophoblastic fragmentsto the other. Two implants found in the experimental horn already appearedabnormal (Fig. 9A-C). Each consisted of a small localized net or cord oftrophoblast cells near the antimesometrial end of a uterine crypt. The uterineepithelium was missing immediately to either side of them. Neither possesseda cavity which, since all the control implants did and the entire tract wasfixed as one, is unlikely to have been an artifact of fixation. One implantcontained 25 cells and the second roughly 36 (one or two sections being folded

vo
Tab
le 5
. Im
plan
tati
on
of m
axim
al
trop
hobl
asti
c fr
agm
ents
in
the
ute
ri o
f pse
udop
regn
ant
mic
e th
at r
ecei
ved
inta
ct b
last
ocys
ts
in t
he c
ontr
alat
eral
hor
n
(Onl
y re
cipi
ents
with
one
or
mor
e im
plan
ts h
ave
been
inc
lude
d in
this
tab
le.)
Exp
.G
roup
A B
Cod
e
Rx
(14.
5.
70)
R2
(14.
5.
70)
R3
(14.
5.
70)
Rx
(18.
5.
70)
R2
(18.
5.
70)
Rx
(29.
5.
70)
Rec
ipie
nt
Gen
otyp
e
PD
E
PD
E
PD
E
PD
E
PD
E
Er
* d
= t
raum
atic
dec
iduo
ma.
Ute
rine
hor
n
Rig
htL
eft
Rig
htL
eft
Rig
htL
eft
Rig
htL
eft
Rig
htL
eft
Rig
htL
eft
MT
F =
max
imal
Typ
e
MT
FB
'cys
tM
TF
B'c
yst
B'c
yst
MT
FM
TF
B'c
yst
B'c
yst
MT
F
MT
FB
'cys
t
trop
hobl
asti
c
Tra
nsfe
rred
tis
sue
Gen
otyp
e
CB
AH
T6x
PD
EC
BA
HT
6xP
DE
CB
AH
T6x
PD
EC
BA
HT
6xP
DE
CB
AH
T6x
PD
EC
BA
HT
6xP
DE
CB
AH
T6x
PD
EC
BA
HT
6xP
DE
CB
AH
T6x
PD
EC
BA
HT
6xP
DE
PDE
PDE
peri
od (
h)
n H H 1* H H H H Hfr
agm
ent.
B'c
yst
= b
last
ocys
t.
No.
ferr
ed 5 6 6 6 6 7 6 6 ON ON 6 6
"No
deci
dual
site
s at
auto
psy
5 5 3 0 6 7 6+
ld*
6 6+
ld*
6 2 6
o o GO o

Inner cell mass and trophoblast tissues 297
Fig. 8. Two atypical implants obtained following transfer of maximal trophoblasticfragments to a recipient carrying intact blastocysts in the contralateral horn. (A) Lowmagnification to show relationship between them. (B) Detail of region of contact.

298 R. L. GARDNER AND M. H. JOHNSON
Fig. 9. Transverse sections of decidual swellings 3 days after transfer of maximaltrophoblastic fragments and intact blastocysts to opposite horns of the samerecipient. (A) Implanted maximal trophoblastic fragment consisting of only 25 cells,of which four can be identified in the figure. (B) The second implanted maximaltrophoblastic fragment consisting of approximately 36 cells of which 4-5 may be seenin the figure. (C) Implanted intact blastocyst in the contralateral horn.

Inner cell mass and trophoblast tissues 299
Table 6. Number of cells in intact 3\-day bias tocystsand dissected ICMs of PDE mice
Type of embryonicmaterial Number scored
Number of cells(mean ± S.E.)
BlastocystsDissected ICMs
1810
57-3 ±3-714-7 + 0-7
100pml
Fig. 10. A single isolated ICM from a 3^-day blastocyst approximately 2\ h after thebeginning of culture. It shows neither cavitation nor intracellular accumulation offluid (cf. Fig. 3B). The smaller fragment is residual trophoblastic debris left behindfollowing dissection of the ICM.
in the latter case). The nuclear diameter of these cells was much less than that offully differentiated giant cells. Their cytoplasmic detail was poor and the viabilityof some was questionable. No dividing cells were present. Indeed, the numberof cells did not exceed that expected in maximal fragments at the time of transfer(Table 6).
E M n 28

300 R. L. GARDNER AND M. H. JOHNSON
Fig. 11. Stages in the fusion of ICMs in vitro. Two pairs of 3^-day dissected ICMsthat had already fused were placed in contact at time zero. (A) +\ h, (B) + \\ h,(C) + 2 h, (D) + 4 | h after contact (darkground illumination). (From Gardner, 1971,with permission of F. Vieweg & Sohn.)
4. Observations on ICMs isolated from S\- and 4\-day blastocysts in culture
ICMs were very variable in shape after they had been scraped from the sheetof overlying trophoblast cells. They rapidly became spherical when cultured, butremained typically as solid balls of cells without developing a central cavity(Fig. 10). When placed in contact in pairs or small groups they almost invariablyaggregated to form unitary structures of spherical or near spherical appearance(Fig. 11). Such fusion consistently took place between pairs of 3^-day ICMs of

Inner cell mass and trophoblast tissues 301
100/mi
Fig. 12. Pair of 3^-day dissected ICMs which failed to fuse after 5 h culture. Intra-cellular accumulation of fluid, suggesting contamination by trophoblast cells, can beseen in both fragments.
the same or different genotype (Table 2, rows I and II). Fusion was also observedbetween pairs of 4^-day ICMs despite the endoderm having delaminated insome of these older embryos as discerned by phase-contrast microscopy (Table 2,rows V, VI and VII). In several instances pairs of ICMs differing by nearly 24 hin age behaved similarly (Table 2, row VIII).
Where fusion failed the fragments remained discrete but connected by a narrowwaist. Failure was often accompanied by the development of a small eccentriccavity or by accumulation of fluids within peripheral cells (Fig. 12). Both thesephenomena suggest that the ICM tissue was contaminated with trophoblastcells. This interpretation is in accordance with the finding that minimal ICMfragments which retain the cells overlying the ICM (see Fig. 2 B) did not aggregateunder similar conditions (Table 2, row IX). However, minimal ICM fragmentscontain more cells than dissected ICMs and hence fusion might simply dependon cell number rather than specifically on cell type. Cell number is probably notthe limiting factor because pairs of fused ICMs can fuse with other ICMs,

302 R. L. GARDNER AND M. H. JOHNSON
thereby yielding structures containing as many cells as entire blastocysts (Table 2,row III; Table 6; Fig. 11).
5. Transfer of fused pairs of 3\-day isolated ICMs to the uteri of recipient mice
A series of fused pairs of ICMs were transferred unilaterally, intact blastocystsbeing placed in the opposite horn of each recipient. The results show that of17 pairs transferred to females with pontamine blue sites in their control hornsat autopsy, none induced decidual changes (Table 3C). Also, in no case couldthe transferred tissue be identified in uterine flushings.
DISCUSSION
The trophoblast and ICM of the mouse blastocyst are very different. Theability to accumulate or pump fluid (Gamow & Daniel, 1970) and to induce thedecidual changes characteristic of implantation are properties peculiar to thetrophoblast. Though the trophoblastic fragments can form vesicles, as docultured fragments of rabbit blastocysts (Daniel, 1961,1963; Klinger, Kosseff&Plotnick, 1971), they do not aggregate with similar fragments or vesicles. Like-wise, mouse blastocysts cannot be induced to fuse together under conditions thatfavour fusion of cleaving eggs or morulae in mice (Mintz, 1965; Mulnard, 1971).This may be related to the presence of specialised junctions between trophoblastcells (Enders & Schlafke, 1965).
Isolated ICMs resemble cleaving eggs in the ease with which they fuse together,as also in their inability to accumulate fluid and evoke decidual reactions (Kirby,1970). The ICMs were fused in pairs before transfer to the uterus to ensure theycontained a similar number of cells to trophoblastic fragments. Hence theirfailure to induce decidual changes could have been due to reduction in viabilityduring the culture period necessary for fusion. Cole & Paul (1965) have arguedthat presumptive ICM cells show poor survival in culture. However, this ex-planation is inappropriate here because cultured ICMs transferred into thecavity of blastocysts or trophoblastic vesicles can contribute extensively to theresulting (chimaeric) offspring or give rise to morphologically normal foetusesrespectively (Gardner, 1971).
The cells overlying the ICM cannot be studied directly with present techniquesbecause they are either removed together with the ICM (in minimal ICM frag-ments - see Fig. 2B) or destroyed when the ICM is scraped from the blastocyst(Fig. 2C). Nevertheless, failure of minimal ICM fragments to fuse togetherwhile isolated ICMs do, indicates that these cells differ from those of the ICM.Whether they are identical to those of the rest of the trophectoderm with whichthey are continuous is uncertain at present.
The absence of embryo, amnion, allantois, yolk sac and Reichert's membranefrom implants developed from pure trophoblast accords with the conclusions ofhistological studies which attribute an ICM origin to these structures (Snell &

Inner cell mass and trophoblast tissues 303
Stevens, 1966). The similarity of all but 3 of the 61 trophoblastic implantsexamined 4-8 days after transfer attests to the homogeneity of the tissue trans-ferred. The very few giant cells evident in the majority of implants are almostcertainly of embryonic rather than maternal origin (Fawcett, Wislocki & Waldo,1947; Snell & Stevens, 1966; Gwatkin, 1966). Whether the absence of giant cellsfrom 19 implants was because they did not develop or were too advanced inpycnosis to be identified remains conjectural. No qualitative differences werenoticed between implants developed from maximal and equatorial trophoblasticfragments.
There was no evidence that the trophoblast had proliferated normally andsubsequently dispersed from the implantation chamber. The low number oftrophoblast cells and absence of mitotic figures at all the stages examined leadsto the inescapable conclusion that the trophoblastic fragments did not proliferatein utero. This might be because the mass of trophoblast tissue, though able toinduce a decidual response, is insufficient to enable subsequent proliferation.This explanation is ruled out by the fact that partial and half blastocysts con-taining as few trophoblast cells as trophoblastic fragments can give rise tonormal conceptuses and young at term (Gardner, 1972).
A further possibility might be that the cells overlying the ICM are requiredfor normal development of the trophoblast. The notion of regional differentiationof trophoblast is implicit in this explanation. Although the ectoplacental conedevelops over the ICM in the early egg-cylinder (Snell & Stevens, 1966), 're-constituted' blastocysts specifically lacking embryonic polar trophoblast candevelop normally (Gardner, 1971, and unpublished observations).
The remaining and perhaps most obvious difference between trophoblasticfragments that fail to produce significant trophoblastic development and intact,partial or 'reconstituted' blastocysts that do, is that the latter all contain someICM tissue. Hence it is concluded that ICM tissue is required to permit normalproliferation of trophoblast of the 3^-day mouse blastocyst. Abundant tropho-blast is found in some implants developed from partial blastocysts that do notcontain embryos (Gardner, 1972), so proliferation presumably does not dependon development of a definitive embryo. Also, dependence on the ICM is prob-ably limited to an early stage of trophoblast development since ectoplacenta-cones from 6- to 7^-day egg-cylinders can proliferate following transferectopically (Grobstein, 1950; Billington, 1965; Simmons & Russell, 1966; Clarke,1969) or to the uteri of cyclic or pseudopregnant mice (Kirby, 1965, 1970;Kirby & Cowell, 1968).
If the proliferation of trophoblast does indeed depend initially on the ICMone would expect to find derivatives of the latter tissue in implants with abund-ant trophoblast. This expectation was fulfilled in nearly all implanted partial and'reconstituted' blastocysts that did not develop foetuses (Gardner, 1972). Theexceptions may have been more apparent than real since they all showed signsof cellular degeneration. One of the three atypical implants developing from

304 R. L. GARDNER AND M. H. JOHNSON
implanted trophoblast in the present experiments carried a foetus, and thesecond is believed to have acquired its trophoblast from the first. The thirdshowed a continuous Reichert's membrane enclosing some cells (Fig. 7). Thismembrane has been found by elegant immunofluorescence studies to be a secre-tion of the distal endoderm (Pierce, Midgley, Sri Ram & Feldman, 1962;Midgley & Pierce, 1963) and hence presumably an ICM derivative (Snell &Stevens, 1966).
There are few published reports containing detailed descriptions of uterineimplants lacking embryos. Tarkowski (1962) found that rat eggs that had under-gone blastulation in the oviduct of the mouse showed dispersal of their ICMsand developed poorly after implanting in the rat uterus. In extreme cases noembryo had formed, but profuse trophoblast surrounded a Reichert's membranelined with a single layer of cells (Tarkowski, 1962, plate 4, fig. V). The innercells are presumably those of the distal endoderm. A similar condition wasfound in another implant following transfer of rat eggs to the oviduct of the rat(Tarkowski, 1962, plate 4, fig. U).
The relevance of descriptions of 20 mouse implantation chambers in whichspontaneous embryonic death had occurred is limited both by the uncertaintyregarding when development went awry and by the fact that the material wasobtained late in pregnancy (Droogleever Fortuyn, 1920). Hence the absence ofall ICM derivatives from five chambers that only contained clusters of giant cellsdoes not preclude their being formerly present.
Failure of the trophoblast of mouse blastocysts to grow ectopically in recipi-ents specifically immunized against the donor strain while similar ectoplacentalcones succeed has been considered evidence that the former express antigens andthe latter do not (Simmons & Russell, 1966). Present considerations raise thepossibility that it is the ICM of the blastocyst that is susceptible to damage orimmunological attack and that failure of trophoblast development dependssecondarily on this factor (Gardner, Johnson & Edwards, 1972).
Critical appraisal of the relationship between the trophoblast and ICM inrodent embryos transferred to ectopic sites is complicated by factors such asabnormal morphogenesis, haemorrhage and degenerative changes. Consequentlymany authors fail to specify derivatives other than trophoblast or embryoproper (e.g. Jollie, 1961). The general conclusion from numerous studies is thatproliferation of the trophoblast is favoured and embryonic development poorand infrequent, especially if oviducal stages rather than blastocysts are trans-ferred (Billington, Graham & McLaren, 1968; Kirby, 1970). Nevertheless,Reichert's membrane is found frequently in grafts lacking embryos. Even whenabsent as a discrete membrane it may indeed be present as a homogeneousmatrix with cells embedded in it (e.g. Fawcett, 1950, figs. 6 and 7; Whitten, 1958,fig. 2; McLaren & Tarkowski, 1963, fig. 4; and author's unpublished observa-tions). Such 'abortive yolk sacs' closely resemble the undifferendatedmurine teratocarcinoma embedded in neoplastic hyalin with which they

Inner cell mass and twphoblast tissues 305
are almost certainly homologous (Pierce et al. 1962; Midgley & Pierce,1963).
A particularly illuminating study, in which all host kidneys were examinedafter transfer of 3^-day mouse blastocysts, is reported by Johnson in an appendixto this paper. Only two grafts definitely lacked ICM derivatives and these, likemost uterine implants developed from trophoblastic fragments, contained justa few giant cells. Such grafts are likely to have been overlooked in studies wherehost organs showing macroscopic 'takes' were selected for examination.
Any explanation of the relationship deduced between trophoblast and ICMmust account for the failure of proliferation of implanted trophoblastic fragmentson the one hand, but development of some giant cells in most of them on theother. Available data are embraced by the following hypothesis. The muraltrophoblast of the mouse blastocyst which gives rise to the primary giant cells(Dickson, 1963) can do so autonomously, or has already received the appropriatestimulus before it is separated from the ICM. It thus provides the few giant cellsfound in trophoblastic implants. The development of the ectoplacental cone(and hence the multitude of secondary giant cells derived from it, Snell &Stevens, 1966) is specifically dependent on the presence of ICM tissue in theblastocyst or early egg-cylinder. The role of the ICM might be to promotedivision or inhibit giant transformation of the overlying trophoblast cells. Thenotion of an inductive interaction is attractive because the ectoplacental coneinvariably develops over the ICM, and because the latter may attain its finalmesometrial position by migrating round the inner surface of the trophoblastwall (Kirby, Potts & Wilson, 1967; Jenkinson & Wilson, 1970; Gardner, 1971).However, it is also possible that the ectoplacental cone is a derivative of theICM (Duval, 1892).
Finally, the morphological similarity between early implants of pure tropho-blast and those of homozygous lethal yellow embryos in utero has not escapednotice (fig. 9 A, B; Eaton & Green, 1963). It is of considerable interest to discoverwhether this resemblance is more than superficial.
I wish to thank Professor C. R. Austin, Dr R. G. Edwards, Dr C. F. Graham, Dr M. H.Johnson and Dr M. I. Sherman for advice and discussion, and Mrs S. C. Barton, Mrs W. J.Gardner and Mrs W. Redmond for help. This work was supported by the Medical ResearchCouncil, the World Health Organisation and the Ford Foundation.

306 R. L. GARDNER AND M. H. JOHNSON
APPENDIX
Relationship between inner cell mass derivatives andtrophoblast proliferation in ectopic pregnancy
SUMMARY
Forty-two 3^-day mouse embryos were transferred to ectopic sites in the kidneys of malemice. Microscopic investigation of all transfers revealed that trophoblast proliferation hadoccurred only when inner cell mass derivatives were present. In the absence of inner cell massderivatives, non-proliferated giant cells were present. This data is taken as evidence compatiblewith the hypothesis that the inner cell mass is essential for trophoblast proliferation.
The results achieved by the use of elegant microsurgical techniques have ledGardner to formulate the hypothesis that the inner cell mass of the 3^-daymouse embryo is essential for development of normal proliferated trophoblast(see the first part of this paper). In the presence of the inner cell mass, normalectoplacental cone tissue with its peripheral secondary giant cells develops. Inthe absence of an inner cell mass, the mural trophoblast of the 3^-day blastocystmerely differentiates into primary giant cells without prior division.
This hypothesis should also apply to ectopically transferred 3^-day embryos,but appears at first sight to be refuted by the reports emphasizing substantialtrophoblast proliferation with little or no embryonic development in suchconditions (Fawcett et al. 1947; Runner, 1947; Fawcett, 1950; Kirby, 1960,1962). In fact, careful examination of these reports in most cases reveals a de-scription of some derivatives of inner cell mass such as an abortive yolk sac(Fawcett, 1950) or some more complex, but imperfect structure of proven innercell mass origin (Snell & Stevens, 1966). With the hypothesis of Gardner in mind,careful histological analysis was made of 42 mouse embryos (3^-day, C3H orC 5 7 B L ) transplanted ectopically to the kidneys of male mice of homologousstrains. The transfer technique has been described elsewhere (Johnson &Dharmawardena, 1972). Seven days after transfer, all kidneys were removed,fixed and serial sections were cut and examined regardless of whether the embryoappeared to have 'taken' macroscopically. Each embryo found was classifiedinto one of the five categories described in Table 7. The classification of group 4transplants (Fig. 13) as Reichert's membrane plus endoderm-like nuclei was madeby others (Fawcett, 1950; Stevens, 1968) and has been discussed in detail byGardner in the first part of this paper.
The data show that, with one exception, trophoblastic proliferation and thepresence of secondary type giant cells were detected only where some evidenceof inner cell mass derivatives existed. For the exceptional transplant, an area ofnecrosis was present in the core of the ectoplacental cone tissue normally occupiedby the embryo. For two transplants, in which inner cell mass derivatives werenot present, small nodules of primary giant cells were detected (Fig. 14). No

Inner cell mass and trophoblast tissues 307
Table 7
No. ofClassification of transplant embryos
1. Embryo showing normal organization at centre 6of proliferating ectoplacental cone withnumerous peripheral secondary-type giant cells.
2. Embryo showing minor abnormalities or areas 10of necrosis at centre of proliferating ectoplacentalcone with numerous peripheral secondary-typegiant cells.
3. Area of necrotic cells (probably embryo) 1surrounded by proliferating ectoplacental conewith numerous peripheral secondary-type giantcells.
4. Compact nodule of Reichert's membrane and 23distal endoderm surrounded by proliferating ecto-placental cone with numerous peripheral secondary-type giant cells (Fig. 13).
5. A few non-proliferated, primary-type giant cells 2only. No proliferating ectoplacental cone or innercell mass derivatives (Fig. 14).
proliferating trophoblast of the type seen in the ectoplacental cone of uterineor other ectopic transplants was present, suggesting that the giant cells wereprimary rather than secondary. The number of cells in each nodule, 9 and 21,were of the order expected for primary trophoblast after transfer of the 30-40mural trophoblast cells in the 3^-day embryo (Gardner, above). The giant cellswere not surrounded by any extensive necrotic or scar tissue, thereby negatingthe possibility that they were the residuum of a proliferated transplant thatsubsequently died. In fact, the two clusters of giant cells bore a remarkableresemblance, both histological and in terms of cell numbers, to those occurringin utero after transfer of pure trophoblastic vesicles (Gardner, above). Presum-ably the inner cell masses of these two embryos had been damaged on transfer,resulting in the absence both of inner cell mass derivatives, and, according to thehypothesis of Gardner, of a stimulus to the proliferation of ectoplacentaltrophoblast. The trophectoderm of the transplanted blastocyst therefore merelydifferentiated into primary giant cells without prior division.
Examination of the literature on ectopic transfers has revealed only one studyin which ectopic transfer sites have been examined histologically regardless ofwhether a macroscopic haemorrhage was present (Stevens, 1968). In that studyalso, non-proliferated primary trophoblastic giant cells only evidently occurredin the absence of any inner cell mass derivatives. Careful analysis of ectopictransfers in other studies would be worth while. The absence of such analyses in

308 R. L. GARDNER AND M. H. JOHNSON
Fig. 13. Nodule of Reichert's membrane and distal endoderm nuclei embeddedin proliferating trophoblast. x 720.
ectopic transfers of blastocysts to recipients preimmunized against donor histo-compatibility antigens may have led to misleading conclusions about the distri-bution of these antigens on the early embryo. The failure to detect macroscopichaemorrhage in these studies could have been due to failure of trophoblast toproliferate secondary to immunological destruction of the inner cell mass rather

Inner cell mass and trophoblast tissues 309
Fig. 14. Non-proliferated trophoblastic giant cells embedded in kidney tissue.xl80.
than to a primary immunological destruction of the trophoblast (Gardner etal. 1972).
In conclusion, the findings that trophoblastic proliferation is invariablyassociated with the presence of inner cell mass derivatives and that a failure oftrophoblastic proliferation is associated with the absence of inner cell mass

310 R. L. GARDNER AND M. H. JOHNSON
derivatives are not in conflict with the hypothesis of Gardner (above) and indeedoffer positive support for it.
I wish to acknowledge the stimulation provided by discussion with Richard Gardner andthe technical assistance of Vinitha Dharmawardena. The work was done whilst the author wasin receipt of an M.R.C. Junior Fellowship, and was supported by a grant to Professor C. R.Austin from the Ford Foundation.
REFERENCESBILLINGTON, W. D. (1965). The invasiveness of transplanted mouse trophoblast and the
influence of immunological factors. /. Reprod. Fert. 10, 343-352.BILLINGTON, W. D., GRAHAM, C. F. & MCLAREN, A. (1968). Extra-uterine development of
mouse blastocysts cultured in vitro from early cleavage stages. / . Embryol. exp. Morph. 20,391-400.
CLARKE, A. G. (1969). Factors affecting the growth of trophoblast transplanted to the testis./ . Reprod. Fert. 18, 539-541.
COLE, R. J., EDWARDS, R. G. & PAUL, J. (1965). Cytodifferentiation in cell colonies and cellstrains derived from cleaving ova and blastocysts of the rabbit. Expl Cell Res. 37, 501-504.
COLE, R. J. & PAUL, J. (1965). Properties of cultured preimplantation mouse and rabbitembryos, and cell strains derived from them. In Preimplantation Stages of Pregnancy(ed. G. E. W. Wolstenholme & M.O'Connor), pp. 82-112. London: Churchill.
DALCQ, A. M. (1957). Introduction to General Embryology. Oxford University Press.DANIEL, J. C. JR. (1961). Reconstitution of rabbit blastocysts from fragments in vitro. Am.
Zool 1, 444.DANIEL, J. C. JR. (1963). Some kinetics of blastocyst formation as studied by the process of
reconstitution. / . exp. Zool. 154, 231-237.DICKSON, A. D. (1963). Trophoblastic giant cell transformation of mouse blastocysts. J.
Reprod. Fert. 6, 465-466.DROOGLEEVER FORTUYN, A. B. (1920). The involution of the placenta in the mouse after the
death of the embryo. Archs Bioi, Paris 30, 323-355.DUVAL, M. (1892). Le Placenta des Rongeurs. (Extrait du Journal de l'Anatomie et de la
Physiologie Annees 1889-1892) (ed. F. Alcan). Paris: Acienne Librarie Germer Bailliereet Cie.
EATON, G. J. & GREEN, M. M. (1963). Giant cell differentiation and lethality of homozygousyellow mouse embryos. Genetica 34, 155-161.
ENDERS, G. J. & SCHLAFKE, S. J. (1965). The fine structure of the blastocyst: some comparativestudies. In Preimplantation Stages of Pregnancy (ed. G. E. W. Wolstenholme & M. O'Con-nor), pp. 29-54. London: Churchill.
FAWCETT, D. W. (1950). The development of mouse ova under the capsule of the kidney.Anat. Rec. 108, 71-91.
FAWCETT, D. W., WISLOCKI, G. B. & WALDO, C. M. (1947). The development of mouse ovain the anterior chamber of the eye and in the abdominal cavity. Am. J. Anat. 81, 413-443.
FINN, C. A. (1965). Oestrogen and the decidual cell reaction of implantation in mice. / .Endocr. 32, 223-229.
FINN, C. A. & MCLAREN, A. (1967). A study of the early stages of implantation in mice.J. Reprod. Fert. 13, 259-267.
GAMOW, E. & DANIEL, J. C. JR. (1970). Fluid transport in the rabbit blastocyst. Wilhelm RouxArch. EntwMech. Org. 164, 261-278.
GARDNER, R. L. (1968). Mouse chimaeras obtained by the injection of cells into the blastocyst.Nature, Lond. 220, 596-597.
GARDNER, R. L. (1971). Manipulations on the blastocyst. In Advances in the Biosciences, no. 6:Schering Symposium on Intrinsic and Extrinsic Factors in Early Mammalian Development(ed. G. Raspe), pp. 279-296. Oxford: Pergamon.

Inner cell mass and trophoblast tissues 311GARDNER, R. L. (1972). Microsurgical approaches to the study of early mammalian develop-
ment. In the Seventh Harold C. Mack Symposium: Birth Defects and Fetal Development,Endocrine and Metabolic Factors. (In the Press.)
GARDNER, R. L., JOHNSON, M. H. & EDWARDS, R. G. (1972). Are H-2 antigens expressed inthe preimplantation blastocyst? In Proceedings of the Second International Symposium onthe Immunology of Reproduction, Varna, Bulgaria. (In the Press.)
GRAHAM, C. F. (1971). The design of the mouse blastocyst. In Control Mechanisms of Growthand Differentiation. Symposium Society for Experimental Biology, no. 25 (ed. D. D. Davis& M. Balls), pp. 371-378. Cambridge University Press.
GROBSTEIN, C. (1950). Production of intra-ocular haemorrhage by mouse trophoblast. / . exp.Zool. 114, 359-373.
GWATKIN, R. B. L. (1966). Amino acid requirements for attachment and outgrowth of mouseblastocysts in vitro. J. Cell Physiol. 68, 335-345.
JENKINSON, E. J. & WILSON, I. B. (1970). In vitro support system for the study of blastocystdifferentation in the mouse. Nature, Lond. 228, 776-778.
JOHNSON, M. H. & DHARMAWARDENA, V. (1972). Effect of preimmunisation with spermatozoaon survival of ectopically transferred mouse blastocysts. Fert. Steril. 23, 353-356.
JOLLIE, W. P. (1961). The incidence of experimentally produced abdominal implantations inthe rat. Anat. Rec. 141, 159-167.
KIRBY, D. R. S. (1960). Development of mouse eggs beneath the kidney capsule. Nature,Lond. 187, 707-708.
KIRBY, D. R. S. (1962). The influence of the uterine environment on the development ofmouse eggs. /. Embryol. exp. Morph. 10, 496-506.
KIRBY, D. R. S. (1965). The 'invasiveness' of the trophoblast. In The Early Concept us, Normaland Abnormal (ed. W. W. Park), pp. 68-73. Dundee: University of St Andrews.
KIRBY, D. R. S. (1970). Immunological aspects of implantation. In Ovo-Implantation. HumanGonadotrophins and Prolactin (ed. P. O. Hubinont, F. Leroy, C. Robyn & P. Leleux),pp. 86-97. Basel: S. Karger.
KIRBY, D. R. S. & COWELL, T. P. (1968). Trophoblast/host interactions. In Epithelial-Mesenchyme Interactions (ed. R. Fleischmajer & R. E. Billingham), pp. 64-77. Baltimore:Williams & Wilkins.
KIRBY, D. R. S., POTTS, D. M. & WILSON, I. B. (1967). On the orientation of the implantingblastocyst, /. Embryol. exp. Morph. 17, 527-532.
KLINGER, H. P., KOSSEFF, A. L. & PLOTNICK, F. (1971). Sex chromatin formation andRNA and protein synthesis during preimplantation development of the rabbit. In Advancesin the Biosciences, no.6: Schering Symposium on Intrinsic and Extrinsic Factors in EarlyMammalian Development (ed. G. Raspe), pp. 207-224. Oxford: Pergamon Press.
LIN, T. P. (1969). Microsurgery of the inner cell mass of mouse blastocysts. Nature, Lond. 222,480-481.
MCLAREN, A. & MICHIE, D. (1954). Transmigration of unborn mice. Nature, Lond. 173, 630.MCLAREN, A. & TARKOWSKI, A. K. (1963). Implantation of mouse eggs in the peritoneal
cavity. /. Reprod. Fert. 6, 384-392.MIDGLEY, A. R. & PIERCE, G. B. (1963). Immunohistochemical analysis of basement mem-
branes of the mouse. Am. J. Path. 43, 929-943.MINTZ, B. (1965). Experimental genetic mosaicism in the mouse. In Preimplantation Stages of
Pregnancy (ed. G. E. W. Wolstenholme & M. O'Connor), pp. 194-207. London: Churchill.MULNARD, J. G. (1965). Studies of regulation of mouse ova in vitro. In Preimplantation Stages
of Pregnancy (ed. G. E. W. Wolstenholme & M. O'Connor), pp. 123-138. London:Churchill.
MULNARD, J. G. (1971). Manipulation of cleaving mammalian embryos with special referenceto a time-lapse cinematographic analysis of centrifuged and fused mouse eggs. In Advancesin the Biosciences, no. 6: Schering Symposium on Intrinsic and Extrinsic Factors in EarlyMammalian Development (ed. G. Raspe), pp. 255-277. Braunschweig: Pergamon Press.
NOYES, R. W. (1959). Trophoblast: problems of invasion and transport. Ann. N.Y. Acad. Sci.80, 54-61.

312 R. L. GARDNER AND M. H. JOHNSON
PIERCE, G. B., MIDGLEY, A. R., SRI RAM, J. & FELDMAN, J. D. (1962). Parietal yolk saccarcinoma: clue to the histogenesis of Reichert's membrane of the mouse embryo. Am.J. Path. 41, 549-566.
PSYCHOYOS, A. (1961). Permeabilitie capillaire et decidualisation uterine. C. r. hebd. Seanc.A cad. Sci. Paris 252, 1515.
RUNNER, M. N. (1947). Development of mouse eggs in the anterior chamber of the eye. Anat.Rec. 98, 1-17.
RUNNER, M. N. (1951). Differentiation of intrinsic and maternal factors governing intrauterinesurvival of mammalian young. J. exp. Zool. 116, 1-20.
SEIDEL, F. (1960). Die Entwicklungsfahigkeiten isolierter Furchungszellen aus dem Ei desKaninchens, Oryctolagus cuniculus. Wilhelm Roux Arch. EntwMech. Org. 152, 43-130.
SIMMONS, R. L. & RUSSELL, P. S. (1966). The histocompatibility antigens of fertilized mouseeggs and trophoblast. Ann. N.Y. Acad. Sci. 129, 35-45.
SNELL, G. D. & STEVENS, L. C. (1966). Early embryology. In Biology of the Laboratory Mouse,2nd ed. (ed. E. L. Green), pp. 205-245. New York: McGraw-Hill.
STEVENS, L. C. (1968). The development of teratomas from intratesticular grafts of tubalmouse eggs. / . Embryol. exp. Morph. 20, 329-341.
TARKOWSKI, A. K. (1962). Inter-specific transfers of eggs between rat and mouse. / . Embryol.exp. Morph. 10, 476-495.
TARKOWSKI, A. K. (1966). An air-drying method for chromosome preparations from mouseeggs. Cytogenetics 5, 394-400.
TARKOWSKI, A. K. & WROBLEWSKA, J. (1967). Development of blastomeres of mouse eggsisolated at the 4- and 8-cell stage. /. Embryol. exp. Morph. 18, 155-180.
WHITTEN, W. K. (1958). Endocrine studies on delayed implantation in lactating mice. Roleof the pituitary in implantation. / . Endocr. 16, 435-440.
(Manuscript received 22 March 1972)





























