Embed Size (px)
Citation preview

An Introduction
to the Aquatic Insects
of North America
Third Edition
Edited by
Richard W. Merritt
Department of Entomology
Michigan Slate University
and
Kenneth W. Cummins
Ecosystem Research and Implementation DepartmentSouth Florida Water Management District
4&*
"i &
K E N D A L L / H U N T P U B L I S H I N G C O M P A N Y4 0 5 0 W e s t m a r k D r i v e O u b u q u e , I o w a 5 2 0 0 2

HABITAT, LIFE HISTORY, AND BEHAVIORAL
ADAPTATIONS OF AQUATIC INSECTS
J. Bruce WallaceUniversity of Georgia, Athens
N.H. AndersonOregon State University, Corvallis
INTRODUCTION
Aquatic insects have received considerable attention with-in the last decade. Book-length coverage of the subject matterin this chapter is found in: Resh and Rosenberg (eds.) 1984,The Ecology of Aquatic Insects; Ward 1992, Aquatic InsectEcology, and Williams and Feltmate 1992, Aquatic Insects.Since 1985 the Annual Review of Entomology has averagedabout an article per year on the ecology or behavior of aquaticinsects and the Journal of the North American BenthologicalSociety began publication in 1986 and many of the papers per-tain to aquatic insects.
Insects are very successful in the freshwater environment.This is demonstrated by their diversity and abundance, broaddistribution, and their ability to exploit most types of aquatichabitats. In this chapter, the adaptations that contribute to theirsuccess are considered. Examples are given of how somespecies have adapted to very restricted environments and thelife cycle is used to provide a framework for describing differ-ent ways that insects cope with the challenges presented byaquatic habitats.
Factors that influence utilization of a particular habitat canbe grouped into four broad categories: (1) physiological con-straints (e.g., oxygen acquisition, osmoregulation, temperatureeffects); (2) trophic considerations (e.g., food acquisition); (3)physical constraints (e.g., coping with habitat or habit as givenin the ecological tables); and (4) biotic interactions (e.g., pre-dation, competition). However, these categories are so interre-lated that detailed analysis of each factor is not appropriate.Respiration is covered in detail in Chapter 4, but it is alsoincluded here because activities related to obtaining oxygen arecentral to behavioral and morphological features associatedwith most other activities.
The traditional division of freshwater systems into stand-ing (lentic) and running (lotic) waters is useful for indicatingphysical and biological differences. Most insects are adaptedto either a lentic or a lotic habitat, but overlaps are commonsuch as in the floodplains of large rivers. For example, insectsinhabiting pools in streams have "lentic" respiratory adapta-tions, whereas those on wave-swept shores of lakes are similarto stream riffle inhabitants in both oxygen requirements and inclinging adaptations (clingers in ecological tables).
Despite their success in exploiting most types of aquaticenvironments, insects are only incompletely or secondarily
adapted for aquatic life. With very few exceptions, aquaticinsects are directly dependent on the terrestrial environment forpart of the life cycle. Even Hemiptera and Coleoptera withaquatic adults require access to surface air for respiration. Thisdependence on the terrestrial environment probably contributesto the prevalence of insects in shallow ponds and streams ascompared with deep rivers or lakes and, in part, to their virtualabsence from the open sea.
Williams and Feltmate (1992) stated that, "One of themost fascinating questions concerning the distribution ofinsects is: given the unparalleled success of insects in both ter-restrial and freshwater environments, why are they so poorlyrepresented in the sea?" While there are over 30,000 species infresh water, there are only several hundred that can be calledmarine. Cheng (1985) records 14 orders and 1400 species ofinsects in brackish and marine habitats, but only the genusHalobates (Hemiptera: Gerridae) in the open seas. One of themost widely accepted theories is that successful residentmarine invertebrates evolved long before aquatic insects andoccupy many of the same niches as do insects in fresh waters;thus, marine invertebrates have barred many insects frommarine habitats by competitive exclusion (e.g., Hynes 1984).Williams and Feltmate offer an interesting alternative view-point that since most marine insects are found in "bridginghabitats" (e.g., estuaries, salt marshes, the intertidal zone, man-grove swamps), perhaps "...we are currently looking not at theend of an evolutionary pathway leading freshwater insects intomarine habitats, but at early steps in the journey." However,this view indicates an extremely slow journey in the last sever-al hundred million years, and the fact remains that the onlytaxon reported for the open seas, Halobates, occurs in a habitat(surface film) occupied by few, if any, marine macroinverte-brates. Some Porifera, Cnidaria, and polychaetes, which aretypically considered to be marine, have representatives thathave also successfully colonized freshwater habitats (Thorpand Covich 1991).
ADAPTATION TO HABITAT
OsmoregulationAquatic insects need to maintain a proper internal salt and
water balance. Their body fluids usually contain a much high-
41

42 LIFE HISTORY
er salt concentration than does the surrounding water and watertends to pass into the hypertonic (higher osmotic pressure)hemolymph. The insect integument, especially the wax layerof the epicuticle, appears to be especially important in prevent-ing flooding of the tissues (Chapman 1982). Some freshwaterinsects take in large quantities of water during feeding. Theirfeces contain more water than the frass of terrestrial counter-parts since many aquatic insects excrete nitrogenous wastes asammonia, which is generally toxic unless diluted with largequantities of water (Chapman 1982). The production of ahypotonic urine (lower osmotic pressure, or more dilute thanthe body fluids) is an important osmoregulatory mechanism inaquatic insects, accomplished by specialized areas of thehindgut that reabsorb ions before wastes are eliminated.
In contrast, saltwater and terrestrial insects conserve waterby producing a rectal fluid that is hypertonic to the hemolymph(Stobbart and Shaw 1974). For example, the dipteran Ephydracinerea, which occurs in Great Salt Lake (salinity > 20%NaCl), maintains water and salt balance by drinking the salinemedium and excreting rectal fluid that is more than 20% salt.
Concentrations of freshwater ions vary tremendously.Many insects absorb salts directly from the surrounding waterby active transport across specialized regions of the body. Suchregions include the rectal gills of the mosquito, Culex pipiens;the rectal gills are larger in larvae reared in water with low ionicconcentrations, which increases the surface area available forabsorption of chloride ions (Wigglesworth 1938). Specializedareas of the integument, including the gills and anal papillae inlarvae of Ephemeroptera, Plecoptera, and Trichoptera, facilitateuptake of ions from the hypotonic external media (Wichard andKomnick 1973, 1974; Wichard et al. 1975; Wichard 1976,1978). In the mayfly Callibaetis sp. the number of cellsinvolved with chloride uptake decreases as the salinity of theenvironment increases, an adaptation to the increase in salinityof drying temporary ponds (Wichard and Hauss 1975; Wichardetal. 1975).
TemperatureVirtually all facets of life history and distribution of aquat-
ic insects are influenced by temperature (see Sweeney 1984,Table 4.1; Ward 1992). It is an important factor determiningthe distribution, diversity, and abundance patterns over eleva-tion gradients in lentic and lotic waters (Ward 1992), togetherwith flow regimes that are also very important (Statzner andHigler 1985; Statzner etal. 1988). Metabolism, growth, emer-gence, and reproduction are directly related to temperature,whereas food availability, both quantity and quality, may beindirectly related through associated microbial activity (Ander-son and Cummins 1979). The diel cycle of temperature alsomay be important: for example, the fecundity of Wyeomyiasmithii, the pitcherplant mosquito, was greater by a factor ofseven when exposed to fluctuating as opposed to constant tem-peratures (Bradshaw 1980).
A few insects occur in waters to almost 5CfC, while otherscan survive frozen in the substrates at the bottom of arcticponds where temperatures may be as low as -20 to -30°C. Incontrast, many taxa inhabiting Alaskan streams cannot surviveeven moderate freezing temperatures of -1°C; it appears thatthese species move away from the freezing zone or remain inareas that do not freeze (Irons et al, 1992). Chironomids andempidids (Diptera) constitute over 90% of the frozen individu-
als found in frozen habitats of Alaskan streams (Irons et al.1992).
The thermal death point of most freshwater invertebratesis between 30 and 40°C (Pennak 1978), so species such as thestratiomyiid fly, Hedriodiscus truquii, which tolerates temper-atures to 47°C in western North America (Stockner 1971), andthe ephydrid fly, Scatella thermarum, found at 47.7°C in Ice-landic hot springs (Tuxen 1944), have developed considerablethermal acclimation. In a general review of thermal springinsect fauna, Pritchard (1991) concluded that no insects liveabove 50°C, few above 40°C, and both temperature and unusu-al water chemistry may exclude certain species.
In contrast to the limited number of species found at hightemperatures, a diverse fauna exists where water is near freez-ing, and many lotic species can grow at winter temperatures.Hynes (1963) and Ross (1963) suggest that this is an adaptationto exploit the seasonal pulse of leaf input in the autumn. Theancestral aquatic habitat of many insects is postulated to becool streams; Hynes (1970a, 1970b) comments that the extanttaxa of Plecoptera, Ephemeroptera, Trichoptera, Corydalidae,and nematocerous Diptera occur in cool streams and these aresurvivors of primitive groups.
Chironomids overwintering as larvae in bottom mud arethe dominant taxon in arctic ponds that freeze to the substrate.Danks (1971b) indicates that overwintering is a distinct andimportant part of the life cycle. The required resistance to therigors of climate occurs in more than one instar. Larvae exhib-it physiological resistance to freezing and many are protectedby a winter cocoon. Danks suggests that robust cocoons areimportant because they can withstand the forces exerted in mudwhen it freezes and water expands to form ice.
Shallow lentic waters will generally reach higher summertemperatures than do streams of the same surface area, whichcan result in a greater algal food supply and faster insect growthrates. However, oxygen may become a limiting factor becauseO2 concentration is inversely proportional to temperature andhigh algal respiration during darkness may deplete the availableO2. Thus, a greater proportion of lentic than lotic species uti-lizes atmospheric O2 or has developed other more efficient res-piratory devices.
Life cycle adaptations have evolved that enable species toutilize favorable periods for growth, coupled with appropriatetiming for aerial existence. This may involve diapause or qui-escent periods in a resistant life stage for avoiding excessivelyhigh or low temperatures. Asynchronous larval cohorts andextended emergence periods occur in springs, where tempera-ture is uniform year around.
Sweeney and Vannote (1978) and Vannote and Sweeney(1980) suggest that an optimal thermal regime exists for a givenspecies and that deviations into warmer (southern) or cooler(northern) waters adversely affect fitness by decreasing bodysize and fecundity. Alteration of thermal regimes, for exampleby removal of riparian vegetation or by hypolimnetic releasefrom dams, will obviously affect insect life cycles or speciescomposition (Lehmkuhl 1972a; Ward 1976).
Lotic HabitatsSubstrates may range from bedrock and large boulders to
fine sediments and be interspersed with large woody debris ina relatively short reach, resulting in a large range of microhab-itats. Substratum particle size is influenced by several items,

LIFE HISTORY
including: geologic structure; the influence of past and presentgeomorphic processes (e.g., flowing water, glaciation, slope,etc.); and, length of time over which the processes occur. Thesein turn influence landform which exerts a major influence onvarious hydrologic characteristics of aquatic habitats (e.g.,Newbury 1984). In addition to influencing substratum particlesize, velocity of moving water exchanges water surrounding thebody and turbulence provides reaeration; thus, dissolved oxy-gen is rarely limiting to stream inhabitants. Local transport andstorage of inorganic and organic materials by the current maybe either detrimental (e.g., scouring action) or beneficial (as afood source).
Substratum characteristics are often perceived as a majorfactor contributing to the distribution of many invertebrates;however, Statzner et al. (1988) suggest that these may be lessimportant than velocity and various key three-dimensionalhydraulic characteristics (longitudinal, vertical, and lateral)associated with different substrates. Flow characteristicsimportant to stream ecology have been discussed by Vogel(1981), Newbury (1984), and Gordon et al. (1992). Theseworks are especially useful with respect to stream invertebrates,as the distribution of organisms and their food resources arestrongly influenced by local flow conditions.
Friction at the substratum surface results in a velocity gra-dient or a "boundary layer" adjacent to the substratum surface(Ambuhl 1959; Hynes 1970a; Statzner et al. 1988). Theboundary layer is regarded as the zone where a sharp velocityreduction prevails close to the substratum (Statzner et al. 1988).Boundary layer thickness is difficult to quantify because ofcomplex substratum profiles, but it generally extends for 1-4mm above the substratum surface.
Vogel (1981) pointed out that almost all of the informationon stream insects prior to the 1980s related to the boundarylayer is mainly anecdotal and few, if any, measurements of draghave been made "under reasonable or unreasonable conditions"and the absence of such measurements make conclusions withregard to shape adaptations of torrential fauna somewhat tenu-ous. Although the shape of dorsoventrally-flattened insects hasbeen proposed as a mechanism to reduce drag and allow themto stay attached in the boundary layer, Vogel (1981) suggeststhe idea may be an over simplification as a convex upper sur-face would generate lift. Statzner and Holm (1982, 1989)reached similar conclusions using Laser Doppler Anemometry.Flows in the vicinity of organisms are often characterized byReynolds number (a measure which relates to laminar and tur-bulent flows). Undisturbed flows immediately upstream of theorganism, the kinematic viscosity, and the shape of the organ-ism or its case determine the organism's Reynolds number andthe degree to which various physical forces influence the organ-ism (Vogel 1981). At low Reynolds number flows tend to belaminar and viscous shear stresses predominate. As theReynolds number increases there is a transition from laminar toturbulent flows and drag forces predominate. Furthermore, therelationship becomes much more complicated for individualorganisms since these forces vary as a function of size betweensmall and large individuals of the same species (Statzner et al.1988). Dorsoventrally-flattened larvae of water pennies(Coleoptera: Psephenidae) have been reported to reduce turbu-lence over the larva's body by active pumping of water throughlateral slots of the carapace, which may decrease drag forces onthe larva at high Reynolds numbers (Smith and Dartnall 1980;
McShaffrey and McCafferty 1987). However, McShaffrey andMcCafferty questioned the significance of the flattened body-shape of Psephenus herricki as an adaptation to fast currents,but rather as an adaptation to avoid the current since individu-als are often found in crevices and undersides of stones.
In contrast to a flattened body, a fusiform, streamlined,and less flattened shape may be a mechanism to avoid exces-sive lift, whereas flattening may be viewed as a mechanism ofmaintaining the body within the boundary layer and increasingthe area of contact with the surface substratum (Vogel 1981) oravoiding the current by occupying crevices. Based on hydro-dynamic studies of streamlined larvae of the mayfly, Cloeondipterum, Craig (1990) has proposed that streamliningenhances accelerative escape motion and "highly streamlinedcylindrical macroinvertebrates could be expected to have accel-erative motion in their behavioral repertoire". Considering thediverse array of physical forces acting on insects, it is not sur-prising that no one taxon would have the "perfect" collection ofadaptations to all forces as this would require incredibly differ-ent morphologies (Statzner et al. 1988, Statzner and Holm1989). Wilzbach et al. (1988) related body shape and drag tomodes of attachment and movement and to functional feedingmodes. Indeed, one wonders to what extent the diverse physi-cal forces influence the array of morphologies found amongaquatic insects.
The range of current velocities associated with varioussubstrates also increases habitat diversity, and various taxa areadapted for maintaining position at different velocities. Filter-feeding collectors exploit the current for gathering food withminimal energy expenditure. For example, certain filtering col-lectors exploit locations where flows converge over and aroundsubstrates, thus allowing the animals to exploit sites of greaterfood delivery (Smith-Cuffney and Wallace 1987; Wetmore etal. 1990). Such areas can be characterized by Froude number,a dimensionless parameter that combines both the depth atwhich flow is measured and velocity in one term (the ratio dfinertial force to the force of gravity; Chow 1959; Newbury1984). Flows are generally faster and the water column's depthis compressed as Froude number approaches or exceeds 1,whereas Froude numbers much less than 1 are found in deepriffles and pools. In a detailed analysis of the caddisfly,Brachycentrus occidentalis Banks, Wetmore et al. (1990) foundthat Froude number was a much better predictor of microhabi-tat than individual depth or velocity measurements.
Morphological and Behavioral Adaptations to Current. Ageneral flattening of the body and smooth, streamlined dorsumare typical of many rheophilic (current-loving) insects: e.g.,heptageniid mayflies, perlid stoneflies, and some psephenidbeetles. Many mayflies and stoneflies have legs that projectlaterally from the body, thereby reducing drag and simultane-ously increasing friction with the substrate. In some caddisflies(e.g., Glossosomatidae), the shape of the case rather than theinsect modifies turbulent flow to a laminar sublayer.
True hydraulic suckers are apparently found only in thelarvae of the dipteran family Blephariceridae (Fig. 22.11). :A V-shaped notch at the anterior edge of each of the six vefitralsuckers works as a valve out of which water is forced when thesucker is pressed to the substrate. The sucker operates as.&pis-ton with the aid of specialized muscles. In addition, a series ofsmall hooks and glands that secrete a sticky substance aid suck-er attachment (Brodsky 1980). Blepharicerids move in a

44 LIFE HISTORY
"zigzag" fashion, releasing the anterior three suckers, liftingthe front portion of the body to a new position, and reattachingthe anterior suckers before releasing and moving the posteriorones to a new position. The larvae are commonly found onsmooth stones, and Hora (1930) attributes their absence fromcertain Indian streams to the presence of moss or roughenedstones that would interfere with normal sucker function.
Several aquatic insects have structures that simulate theaction of suckers. The enlarged gills of some mayflies (e.g.,Epeorus sp. and Rhithrogena sp.) function as a friction pad, andDrunella doddsi has a specialized abdominal structure for thesame purpose. Brodsky (1980) describes a "pushing-proleg" insome chironomids; it has a circlet of small spines that functionas a false sucker when pressed to the substrate. Mountainmidge larvae (Deuterophlebiidae) possibly use a similar mech-anism to attach their suckerlike prolegs.
Larval black flies (Simuliidae) use a combination of hooksand silk for attachment. The thoracic proleg resembles that ofchironomids and deuterophlebiids, described above, and the lastabdominal segment bears a circlet of hooks. The larva spreadsa web of silk on the substrate to which it attaches either the pro-leg or posterior hooks. The larva moves forward in an inch-worm-like manner, spins silk over the substrate, and attaches theproleg and then the posterior circlet of hooks to the silken web.
Silk is used for attachment by a number of caddisflies(e.g., Hydropsychidae, Philopotamidae, and Psychomyiidae),which build fixed nets and retreats. Some case-making caddis-flies (e.g., Brachycentrus sp.) also use silk for attaching theircases to the substrate in regions of fairly rapid flow, and free-living caddisflies may use "security threads" as they move overthe substrate in fast currents. The silk line is used in combina-tion with their large anal prolegs, which are employed as grap-ples. Many chironomid larvae construct fixed silken retreatsfor attachment, and black fly pupae are housed in silken casesthat are attached to the substrate. Other morphological adapta-tions to running water are given in Table 8A and are listed foreach genus in the ecological tables.
Despite the fact that unidirectional current is the basic fea-ture of streams, the majority of lotic insects have not adapted tostrong currents but instead have developed behavior patterns toavoid current. Very few lotic insects are strong swimmers,probably because of the energy expenditure required to swimagainst a current; downstream transport requires only a move-ment off the substrate to enter the current. Streamlined forms,such as the mayflies Baetis sp., Isonychia sp., and Ameletus sp.,are capable of short rapid bursts of swimming, but most loticinsects move by crawling or passive displacement. The benth-ic fauna chiefly occurs in cracks and crevices, between or underrocks and gravel, within the boundary layer on surfaces, or inother slack-water regions. Presumably, some portion of thebenthic community seeks refuge deeper in the substrates duringfloods; insects are difficult to find at such times, but normalpopulation levels are found soon after the flows subside(Williams 1984).
The hyporheic region is the area below the bed of a streamwhere interstitial water moves by percolation. In gravelly soilsor glacial outwash areas it may also extend laterally from thebanks. An extensive fauna occurs down to one meter in suchsubstrates (Williams and Hynes 1974; Williams 1984). Mostorders are represented, especially those taxa with slender flex-ible bodies or small organisms with hard protective exoskele-
tons. Stanford and Gaufin (1974) report that some stonefliesspend most of their larval period in this subterranean region ofa Montana River. They collected these larvae in wells over 4 mdeep, located 30-50 m from the river.
Drift. Downstream drift is a characteristic phenomenon ofinvertebrates in running waters. Despite the adaptations formaintaining their position in the current or avoiding it, occa-sional individuals could be expected to lose attachment or ori-entation and be transported downstream. However, the largenumbers of some taxa that drift indicate that this is more than apassive activity. Waters (1965) divided drift into three cate-gories: (1) catastrophic, resulting from physical disturbance ofthe bottom fauna, e.g., by floods, high temperatures, and pollu-tants; (2) behavioral, indicated by characteristic behavior pat-terns resulting in a consistent diel periodicity (usually at night);and (3) constant, the continual occurrence of low numbers ofmost species. Mayflies of the genus Baetis consistently exhib-it high behavioral drift rates with a night-active periodicity.Other mayflies, stoneflies, caddisflies, black flies, and theamphipod Gammarus sp. are frequently abundant in drift.Food resources may also influence drift as many more individ-uals of Baetis sp. (Ephemeroptera) drifted from an unfertilizedreach of the Kuparuk River in Alaska, compared to a down-stream fertilized reach where food was more abundant (Her-shey et al. 1993). However, Wilzbach (1990) reported non-concordance between drift and benthic feeding activity inBaetis. Drift is important to stream systems in the recoloniza-tion of denuded areas, as a dispersal mechanism, and particu-larly as a food source for visual predators. Many fish, espe-cially salmonids, select and defend territories best suited for theinterception of drift (Waters 1972).
Irrespective of its causes, drift results in a net downstreamdisplacement of some portion of the benthic population.Whether drift losses from upstream areas represent excess pro-duction or whether compensatory upstream movements arerequired is not known. Miiller (1954) proposed that upstreamflight of adults could be the mechanism to complete the "colo-nization cycle," and upstream migrations of some mayflies andamphipods in the slow water near shore have been recorded(e.g., Neave 1930; Minckley 1964; Hayden and Clifford 1974).Using stable isotopes, Hershey et al. (1993) estimated that one-third to one-half of the adult Baetis sp. population in theKuparuk River flew over 1.6 km upstream from their emer-gence sites. During three years of seasonal insecticide applica-tions to a headwater stream, colonization by early instars ofseveral taxa showed seasonal occurrences that closely paral-leled known life cycles and adult flight periods (Wallace et al.1991). However, under less adverse conditions the relativeimportance of upstream movement, or even its necessity formost taxa, remains an open question. For example, Wilzbachand Cummins (1989) showed that in the short term, recruitmentof immatures made it impossible to detect any 'populationdepletion due to drift. Several workers (Bishop and Hynes1969; Waters 1972; Miiller 1974; Williams 1981a; Brittain andEikeland 1988) have reviewed the extensive literature on thesignificance of drift to production biology, population dynam-ics, and life histories.
Unstable Substrates. Sandy substrates of rivers and streamsare poor habitats because the shifting nature of the bed affordsunsuitable attachment sites and poor food conditions. Anextreme example of this instability is the Amazon River, where
1
LIFE HISTORY 45
strong currents move bedload downstream as dunes of coarsesand reaching 8 m in height and up to 180 m in length, largelypreventing the establishment of a riverbed fauna (Sioli 1975).Despite substrate instability, some sandy streams are quite pro-ductive. Blackwater streams of the Southeast have extensiveareas of sand with an average standing stock, primarily smallChironomidae less than 3 mm in length, exceeding 18,000/m2.Though their biomass is small, rapid growth rates result in a sig-nificant annual production and an important food source forpredaceous invertebrates and fish (Benke et al. 1979).
The inhabitants of sandy or silty areas are mostlysprawlers or burrowers (Table 6B), with morphological adapta-tions to maintain position and to keep respiratory surfaces incontact with oxygenated water. The predaceous mayfliesPseudiron sp. and Analetris sp. have long, posterior-projectinglegs and claws that aid in anchoring the larvae as they faceupstream. Some mayflies (e.g., Caenidae, Tricorythidae, andBaetiscidae) have various structures for covering and protectinggills, and others (e.g., Ephemeridae, Behningiidae) have legsand mouthparts adapted for digging. The predaceous mayflyDolania sp. burrows rapidly in sandy substrates of Southeast-em streams. The larva utilizes its hairy body and legs to forma cavity underneath the body where the ventral abdominal gillsare in contact with oxygenated water.
Many dragonflies (e.g., Cordulegaster sp., Hagenius sp.,Macromiidae, and many Libellulidae) have flattened bodiesand long legs for sprawling on sandy and silty substrates. Theyare camouflaged by dull color patterns and hairy integumentsthat accumulate a coating of silt. The eyes, which cap the ante-riolateral comers of the head, are elevated over the surroundingdebris. Many gomphid larvae actually burrow into the sedi-ments using the flattened, wedge-shaped head, and fossorialtibiae. The genus Aphylla (Gomphidae) is somewhat unusualin that the last abdominal segment is upturned and elongate,allowing the larvae to respire through rectal gills while buriedfairly deep in mucky substrate.
Wood-Associated Insects. Wood debris provides a signifi-cant portion of the stable habitat for insects in small streamswhere water power is insufficient to transport it out of the chan-
nel. In addition to the insect component using wood primarilyas a substrate, a characteristic xylophilous fauna is associatedwith particular stages of degradation. These include: chirono-mid midges and scraping mayflies (Cinygma sp. and Ironodessp.) as early colonizers; the elmid beetle, Lara avara, and thecaddisfly, Hetemplectron califomicum, as gougers of firmwaterlogged wood; chironomids as tunnelers; and the tipulids,Lipsothrix spp., in wood in the latest stages of decomposition(Anderson et al. 1978, 1984; Dudley and Anderson 1982;Anderson 1989). Wood debris is most abundant in small forest-ed watersheds, but it is also an important habitat at the marginsand on point bars in larger streams with unstable beds. Cudneyand Wallace (1980) found that submerged wood in the CoastalPlain region of the Savannah River was the only substrate suit-able for net-spinning caddisflies, but that high standing cropsand production could be supported in a relatively small spacebecause the caddisflies were exploiting the food resource trans-ported to them by the current. Benke et al. (1979) reported thatsnags in a small Southeastern blackwater river were highlyproductive, not only for net-spinning caddisflies, but also forfilter-feeding Diptera and other typical "benthic" insects.Wood gouging habits of net-spinning caddisflies were blamed,at least in part, for the 1988 collapse of submerged timber pil-ings supporting a Maryland bridge (National TransportationSafety Board 1989). Even in the Amazon situation mentionedabove, large logs in the lee of dunes are heavily colonized bychironomid midges (Sioli 1975).
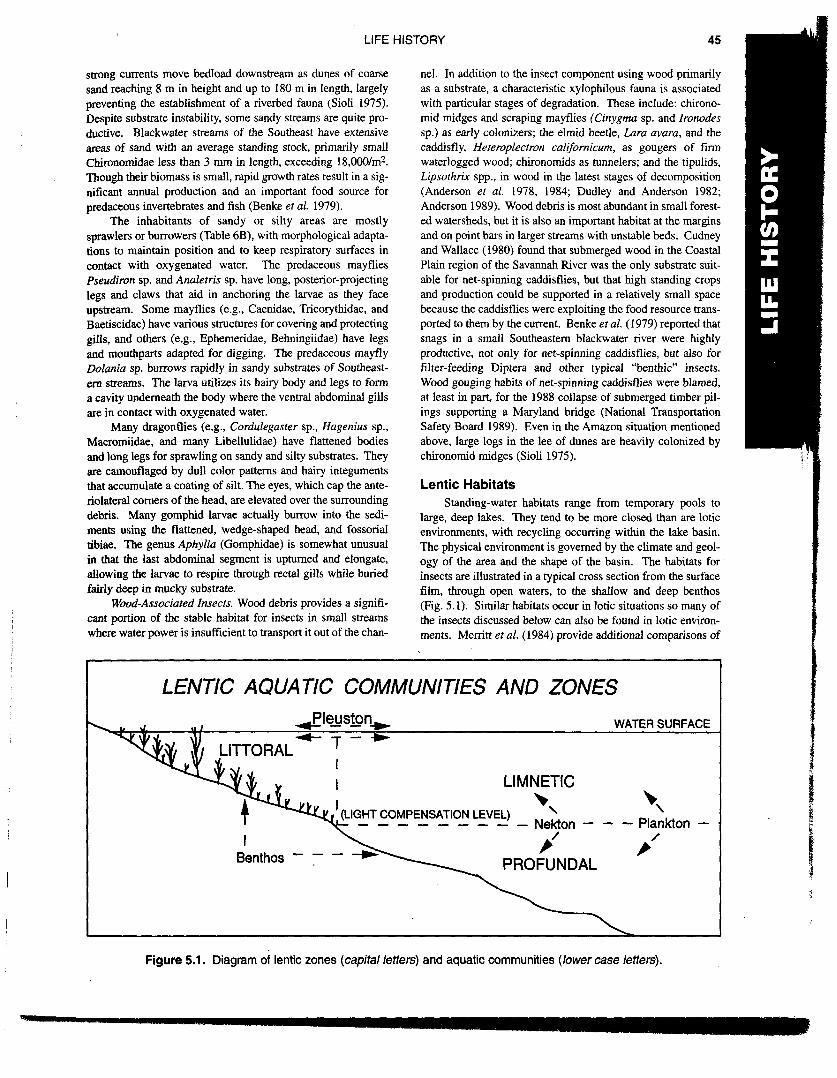
Lentic HabitatsStanding-water habitats range from temporary pools to
large, deep lakes. They tend to be more closed than are loticenvironments, with recycling occurring within the lake basin.The physical environment is governed by the climate and geol-ogy of the area and the shape of the basin. The habitats forinsects are illustrated in a typical cross section from the surfacefilm, through open waters, to the shallow and deep benthos(Fig. 5.1). Similar habitats occur in lotic situations so many ofthe insects discussed below can also be found in lotic environ-ments. Merritt et al. (1984) provide additional comparisons of
LENTIC AQUATIC COMMUNITIES AND ZONES
WATER SURFACE
LITTORAL T
LIMNETIC
' (LIGHT COMPENSATION LEVEL) N
— — Nekton —I
Benthos — — —PROFUNDAL
\— Plankton
/
Figure 5.1. Diagram of lentic zones (capital letters) and aquatic communities (lowercase letters).

46 LIFE HISTORY
lotic and lentic ecosystems and nutritional resources availableto aquatic insects in each habitat.
Surface Film. The unique properties of the water surfaceconstitute the environment for the pleuston community. Waterstriders (Gerridae), whirligig beetles (Gyrinidae), mosquito lar-vae (Culicidae), and springtails (Collembola) are commonexamples. The surface film results from the attractive forcesamong water molecules. Within the body of water the forcesare equal on all sides, but at the surface the attraction is lessbetween air and water than on the other three sides. This resultsin a slight pull toward the center of the water mass and the sur-face acts as if it were a stretched elastic membrane. "To anorganism of small size, this air-water interface can be animpenetrable barrier, a surface on which to rest, or a ceilingfrom which to hang suspended" (Usinger 1956a).
Water striders and bugs of the related family Veliidae havepreapical claws that enable them to move about without break-ing the surface film and a velvety hydrofuge hair pile on theventer that is nonwettable. The skating motion of gerrids ismore accurately described as rowing. The weight causes aslight dimpling of the surface film (easily seen in the shadow:ast on the bottom of clear shallow waters). The long tarsi ofthe middle legs are pressed against the depression in the filmfor propulsion while the hind legs are held outstretched behindas rudders for steering.
The gerrids also exploit the surface film by detectingvibrations from surface ripples with sensors located between:he tarsal segments on the meso- and metathoracic legs. The•equence in which these sensors perceive an oncoming rippleells the insect how much to turn to face the disturbance, causedor example, by a potential prey. The strider turns by movingts rowing legs in opposite directions much as an oarsmanvould turn a boat. At this juncture, a combination of visual andibratory information allows the strider to choose betweenpproaching the disturbance or fleeing from it. If the former, itows forward, pausing between each stroke to evaluate furtheripple signals (Milne and Milne 1978). Some gerrids also useibrations of the surface film for communication in courtshipnd mating (Wilcox 1979).
Whirligig beetles have divided eyes; the lower part detects| vents under the water and the upper part detects events on or
oove the water surface. Glands keep the upper portion of thejdy greased to repel water, whereas the lower surface andwer eyes are wettable. Whirligig gyrations on the surface arefected by rapid propulsion with paddle-shaped swimminggs. The hindlegs are unique in that the segments can be fold-up during the forward stroke and spread apart like a fan for
•: powerful backward stroke. In addition to propulsion, the.imming activity causes a series of ripples or bow waves that,icn reflected back by obstacles, are detected by the antennaejching the surface film, allowing quick course correctionan in the dark (Milne and Milne 1978).
: Pleuston insects are potentially vulnerable to predation byual aquatic predators because they are silhouetted against
• sky; however, both gerrids and gyrinids seem to be avoidedfish, apparently because secretions produced by repugnato-glands make them distasteful (e.g., Blum 1981).
\ Hydrofuge structures are important adaptations for obtain-atmospheric air through the surface film. The openings ofninal spiracles have glands that discharge a waxy secretionthe cuticle; contact with the surface film repels the water
exposing the spiracles to the air. The waxy lining of the tra-cheae prevents water from running into them by capillarity.Some insects (e.g., Culex sp., Limonla sp., Stratiomys sp.) canhang from the surface film supported by a crown of semihy-drofuge hairs around the spiracles. The semihydrofuge hairwill lie in the water surface, half wetted and half exposed.When a stratiomyiid larva submerges, the crown is lifted off thewater and envelops an air bubble that is used as an air store.
Feeding adaptations associated with pleuston specializa-tion are exemplified by mosquito larvae. Though the mouth-parts are basically of the primitive chewing type, the larvaehave brushes arising from the labrum that sweep floating orsuspended material toward the mouth. Anopheles sp. larvae liebeneath the surface film supported by tufts of float hairs oneach segment. The larva rotates its head so that the mouthbrushes are uppermost and sets up a current that draws themicrobial-rich layer of water along the underside of the surfacefilm and into the mouth (Merritt et al. 1992).
Limnetic Zone. In open waters to the depth of photosyn-thetically-effective light penetration, a broad distinction ismade between nektonic and planktonic organisms: nekton areswimmers able to navigate at will (e.g., Coleoptera, Hemiptera,some Ephemeroptera), whereas plankton are floating organ-isms whose horizontal movements are largely dependent onwater currents.
The phantom midge Chaoborus sp. is the most commoninsect plankter; it is abundant in many eutrophic (nutrient-rich)ponds, lakes, and some large rivers. Chaoborus sp. exhibitsvertical migrations, occurring in benthic regions during the daybut migrating vertically into the water column at night. Thesemigrations are dependent on light and oxygen concentrations ofthe water (LaRow 1970). Larvae avoid predation by beingalmost transparent except for two crescent-shaped air-sacs orbuoyancy organs (Fig. 22.30); they lie horizontally in the water,slowly descending or rising by adjusting the volume of the air-sacs. Their prehensile antennae are used as accessory mouth-parts to impale zooplankton and deliver them to the mouth.
Many lentic insects are strong swimmers but relativelyfew are nektonic. They pass through the limnetic zone whensurfacing for emergence, but the vast majority of lentic insectsoccurs in shallow water with emergent plants. As mentionedpreviously, the scarcity of insects in limnetic areas may be aconsequence of the secondary adaptations for aquatic life.There are no resting supports in the limnetic zone so maintain-ing position requires continuous swimming or neutral buoyan-cy.
Littoral Zone. The littoral zone, the shallow region withlight penetration to the bottom, is typically occupied by macro-phytes (macroalgae and rooted vascular plants). It contains adiverse assemblage of insects with representatives of mostaquatic orders. Habitats include benthic and plant surfaces, thewater column, and the surface film (Table 6A); occupantsinclude burrowers, climbers, sprawlers, clingers, swimmers,and divers (Table 6B). Morphological and behavioral types inthe littoral zone or floodplain wetlands of larger rivers are sim-ilar to those in slow-moving or backwater regions of lotic habi-tats. The diversity and abundance of littoral species result inbiological factors (e.g., competition and predation) assumingimportance in shaping community structure.
The biomass and diversity of invertebrates associated withaquatic macrophytes in lentic or lotic habitats may exceed that
cSS'C;fl1'mbeattr;cuwhall
oriyhaTrigreflkpalofiph;chr;Ma.thearehos

LIFE HISTORY 47
of the fauna in the sediments at the same location. The impactof herbivorous insects on many living plants has previouslybeen considered to be low, and it has been suggested that aquat-ic macrophytes produce secondary plant substances that serveas chemical defenses against herbivores (Otto and Svensson1981b), or that they may be deficient in some essential aminoacids (Smimov 1962). However, the nitrogen content ofmacrophytes is similar to that of terrestrial plants and there isnow considerable evidence to suggest that herbivory on livingmacrophytes may be much more important than previously sus-pected (Lodge 1991; Newman 1991). Herbivore-chewers(shredders-herbivores), miners, and stem borers (see Chap. 6)feed on macrophytes. Floating leaf macrophytes such as waterlilies (Nuphar) can be heavily consumed by larvae and adultsof the chrysomelid, Galerucella sp. (Wallace and O'Hop 1985).Some aquatic herbivores include pests of economic importancesuch as the weevil Lissorhoptrus oryzophilus (^simplex) (Cur-culionidae) on rice (Isely and Schwardt 1934), and the caddis-fly Umnephilus lunatus (Limnephilidae) on watercress (Gower1967). Newman (1991) found that most herbivory on aquaticmacrophytes is usually by specialized oligophagous insectsbelonging to families and orders that are secondary invaders ofaquatic habitats and closely related to pest herbivores in terres-trial habitats (e.g., beetles such as Chrysomelidae and Cur-culionidae, aquatic Lepidoptera, and specialized Diptera),whereas herbivores from primarily aquatic groups were gener-alists and also detritivores.
Insects also use macrophytes as substrates for attachmentor a surface from which to graze periphyton, rather than direct-ly as food. For example, macrophytes in both lentic and lotichabitats often harbor a number of filter-feeding Ephemeroptera,Trichoptera, and Diptera. Many species found in weed beds aregreen in color and blend in with their surroundings. Dragon-flies of the family Aeshnidae often have contrasting bands ofpale and dark green or light brown that add to the effectivenessof the camouflage. Some species in most orders utilize macro-phytes as oviposition sites. Larvae of some Coleoptera, e.g., thechrysomelid Donacia sp., and Diptera such as the mosquitoMansonia sp. and the aquatic syrphid, Chrysogaster sp., rely onthe intracellular air spaces of aquatic plants for respiration andare thus limited in their distribution by that of their macrophytehost.
Profundal Zone. The number of taxa of aquatic insects thatoccur below ca. 10 m is limited, but the few species that dooccur there can be very abundant. This area includes the sub-littoral and profundal regions. The latter is the zone belowwhich light penetration is inadequate for plant growth. Thedeep water is a stable region because water movement is mini-mal and temperature varies only slightly between summer andwinter. Periodic depletion or absence of dissolved oxygen mayoccur, especially in eutrophic situations. Substrates are usual-ly soft or flocculent and offer little in the way of habitat diver-sity or cover. The inhabitants are mostly burrowers that feed onsuspended or sedimented materials and are capable of tolerat-ing low dissolved oxygen or even anaerobic conditions. Typi-cal deep-water insects are ephemerid mayflies (e.g., Hexageniasp., Ephemera sp.) and many genera of Chironomidae (e.g.,Chironomus, Heterotrissocladius, Tanytarsus). Predaceousdeep-water insects include Sialis sp. (Megaloptera) andChaoborus sp. (Diptera).
Dense populations of the midge genus Chironomus are
characteristic of profundal sediments. The larvae build U-shaped tubes with both openings at the mud-water interface.Body undulations cause a current of water, providing oxygenand paniculate food in the form of phytoplankton and finedetritus with its accompanying microbes to be drawn throughthe tube. Hemoglobin serves as an oxygen store during periodsof low dissolved oxygen, but the larvae become quiescentunder anaerobic conditions. The life cycle of profundal Chi-ronomus typically requires two years, compared with a year orless for the same species in shallow waters. This is due to theslow growth at cold temperatures, low food quality, and extend-ed periods of quiescence when the water is anoxic (Jonassonand Kristiansen 1967; Danks and Oliver 1972).
The profundal chironomid community has been usedextensively as an indicator of the nutrient conditions or pro-ductivity status of lakes. Saether (1980b) lists about twentygenera of chironomids that are characteristic of particular oxy-gen-nutrient-substrate combinations, which, in conjunctionwith other non-insect assemblages, can be used in lake typolo-gy classifications. Warwick (1980) documented over 2500years of land-use practices around Lake Ontario using the sub-fossil chironomid head capsules in a sediment core; changes inspecies composition of midges were associated with variousperiods of eutrophication, deforestation, sedimentation, andcontamination.
LIFE CYCLE ADAPTATIONS
Diverse life-history patterns have evolved to enablespecies to exploit foods that are seasonably available, to timeemergence for appropriate environmental conditions, to evadeunfavorable physical conditions (e.g., droughts, spates, andlethal temperature), and to minimize repressive biotic interac-tions such as competition and predation. Typically, a seasonalsuccession of species can be cataloged at a given location bydetermining the emergence and flight periods of adults or bystudying the larval growth periods. However, comparison of agiven species at different sites may indicate considerable flexi-bility in life histories. Some important adaptations include: (1)responses to temperature and oxygen levels; (2) use ofdaylength or temperature as environmental cues to synchronizelife stages; (3) resting stages to avoid unfavorable conditions;and (4) extended flight, oviposition, or hatching periods tospread the risk in coping with environmental conditions thatcannot be avoided entirely. Kohshima (1984) reported anextreme case of adaptation of a Diamesa sp. (Chironomidae)inhabiting glacial melt water channels above 5200 m in theHimalayan Mountains. The location of these channels, inwhich the univoltine species develop, is unpredictable fromyear to year. Koshima suggested the mated females, whichremain active at a temperature of -16°C, overwinter under snowand ice and are ready to oviposit when the new channels devel-op during the following summer.
All life stages of mayflies exhibit adaptations to wannerwaters compared with stoneflies. Egg development in mayflieshas higher thermal requirements and is more temperaturedependent than in stoneflies (Brittain 1990). A number ofmayflies have short life cycles whereas many stoneflies requiremore than one year to complete their development. AdultEphemeroptera mate in flight, which requires higher air tern-

48 LIFE HISTORY
peratures than for Plecoptera which generally remain in thevicinity of the larval habitat and mate on solid substratum (Brit-tain 1990). Such mating behavior may explain, in part, thegreater extension of plecopterans to arctic regions, althoughthis involves a trade off between disperal ability and reproduc-tive success in harsh environments. Brittain (1990) has sug-gested that plecopterans are relatively temperature independentand adapt to alternative food resources, whereas mayflies aremore thermally dependent and readily use diatoms, algae, etc.to maintain shorter life cycles in warmer waters. Thus, ple-copterans have a competitive advantage at cooler temperatureswhereas warmer thermal regimes favor mayflies (Brittain1990).
The duration of aquatic insect life cycles ranges from lessthan two weeks (e.g., some Baetidae, Tricorythidae, Culicidaeand Chironomidae) to several years (e.g., Elmidae and someOdonata), but in the north temperate zone an annual cycle ismost common. Examples of aquatic insect life cycles are givenin Table 5B. We have used the terms univoltine, bivoltine, tri-voltine, and multivoltine to indicate those taxa reported to com-plete their life cycles in a year or less; semivoltine for thosecompleting their life cycle in 2 years; and, merovoltine forthose that require more than two years from egg to adult.Although many taxa reported in Table 5B are univoltine, larvalgrowth and development may occur over a very short period,e.g., a few weeks or months. For example, a number of uni-voltine coleopterans require only a month or so to completetheir larval development.
Life histories of insects that spend all or a portion of theirlife history in marine environments are not included in Table5B. Cheng and Frank (1993) have provided a summary of cur-rent information on life history attributes and reproduction inmarine insects.
Many species of mayflies maintain univoltine life cyclesover broad latitudes in eastern North America. There is a ques-tion as to how voltinism and seasonal synchrony are maintainedacross broad latitudinal ranges and thermal regimes where tim-ing of adult emergence and oviposition may be restricted toone- or two-week periods. To examine this question, Newboldet al. (1994) used daily water temperatures and maximum andminimum threshold temperatures for larval development inorder to construct a temperature based model for severalmayfly species. The model incorporated periods of quiescenceassociated with both high and low temperatures for 11 riversystems ranging from the southeastern United States to Que-bec. The model simulations indicate that most mayfly devel-opment occurs in the spring and autumn between 34°N and50°N when thermal regimes are roughly similar at all latitudes(Newbold et al. 1994). At lower latitudes, there was a longerperiod of summer quiescence as the development rate droppedat higher temperatures. The model also predicted the earlieremergence observed at lower latitudes as well as a northwardtransition to a semivoltine life history for two of the. speciesexamined (Newbold et al. 1994).
Hynes (1970a) distinguished three main types of lifecycles for insects in a temperate stream: slow seasonal, fast sea-sonal, and nonseasonal cycles. In seasonal cycles, a distinctchange of larval size occurs with time, i.e., the progression ofgrowth by cohorts can be discerned by periodic sampling offield populations. In nonseasonal taxa, individuals of severalages are present at all times. The three types are illustrated by
three species of glossosomatid caddisflies, all collected fromone emergence trap (Fig. 5.2).
Slow-seasonal cycles are common in cool streams andtypified by many Plecoptera, some Ephemeroptera, and Tri-choptera. Eggs may hatch soon after deposition, and larvaegrow slowly, reaching maturity nearly a year later. In manyspecies, an extended hatching period results in recruitment overseveral months. Larvae grow during winter and most speciesreach the adult stage early in the year.
Fast-seasonal cycles are those in which growth is rapidafter a long egg or larval diapause or after one or more inter-mediate generations. The caddisfly Agapetus bifidus has anegg diapause of 8-9 months and a larval growth period of only2-3 months (Fig. 5.2). Various fast-seasonal cycles reach fullterm in spring, early and late summer, and fall. Two or morefast cycles may be exhibited by the same species when rapidgenerations succeed one another, as in the mayfly Baetis sp.and the black fly Simulium sp. Individuals that grow rapidly atwarm temperatures tend to be much smaller than those of theearlier, slow-growing generation (Hynes 1970b; Ross and Mer-ritt 1978; Sweeney 1978; Cudney and Wallace 1980; Merritt etal. 1982; Georgian and Wallace 1983; Freeman and Wallace1984).
In nonseasonal cycles, individuals of several stages or sizeclasses are present in all seasons. This may be because the lifecycle spans more than a year as in some large Plecoptera andMegaloptera, or because a series of overlapping generationsoccurs, or for other unexplained reasons. Chironomidae oftenexhibit nonseasonal patterns probably because two or morespecies have not been distinguished or because short life cyclesresult in overlapping cohorts.
Poorly synchronized life cycles would be expected tooccur in situations where limiting factors for growth or repro-duction (e.g., temperature, food, and moisture) are not season-ally dependent. Thus in the tropics the unvarying temperatureand light conditions could result in continuous periods ofgrowth and reproduction. It is somewhat surprising that aquat-ic insects in the southern hemisphere are predominantly of thenonseasonal type in contrast to the situation in the north tem-perate zone (Winterboum 1974, 1978; Hynes and Hynes 1975;Corbet 1978; Towns 1981). Hart (1985) suggested that flexibleand opportunistic life histories of southern hemisphere streamfauna may be a selective response to unpredictable, or to mildand uniform, environmental conditions.
Habitat SelectionOn a gross level, habitat selection is primarily the province
of the mated females as deposition of eggs determines wherethe larvae will initially occur. The chain of behavioral cuesleading to oviposition has rarely been elucidated but habitatselection may involve visual, tactile, and chemosensory cues.Coleoptera and Hemiptera detect water while in daytime flightapparently from the reflective surface so they are attracted toponds, swimming pools, and other shiny surfaces. For mostaquatic insects, oviposition occurs near the location from whichthe adult emerged, but dispersal flights are common especiallyin species from temporary habitats. Species that overwinter asadults (e.g., Coleoptera, Hemiptera) may have a fall dispersalflight to overwintering sites, and also a spring flight for ovipo-sition.
The widespread tropical dragonfly, Pantala flavescens

LIFE HISTORY 49
SLOW SEASONALAnagapetus bernea
FAST SEASONALAgapetus blfidus
NON-SEASONALGlossosoma penftum
AUG 31 50%
SEP 25 50% -
OCT 24- 50o/NOV 4 50/oh
DEC 2 50%
JAN 2 50%
FEB 5 50%
MAR 7-21 50%
APR 2-26 50%
MAY 7-25 50%
JUN 11-30 50%
JUL 8-28 50%
AUG II 50%
n = 31
(Diapausing'Eggs)
n = 98
n = 52
n = 63
r\=76
n= 50
n=69I 2 3 4 5 P
Adultsi
n = 55
n= 109
n=2
Adults
I 2 3 4 5 P
V
I 2 3 4 5 P
n= HO
n = ISO
n= 61
n = 67
n= 118
Figure 5.2. Age distribution of three glossosomatid caddisflies, illustrating life cycles. Field dataare expressed as percentage composition per month for each instar. There are five larval instars;P = prepupa + pupa; n = number per sample. Flight period of adults is also indicated. (Data fromAnderson and Bourne [1974].)
(Libeliulidae), an obligate migrant that breeds in temporaryponds, exhibits long-distance dispersal in search of ovipositionsites. The adults fly upward at emergence and are then trans-ported by the wind to places where rain will later fall. Windmay transport the adults over 1400 km across the Indian Oceantoward the cold front produced by monsoons (Corbet 1963).
Caddisflies from temporary streams (Hespemphylax sp.,Grammotaulius sp.) and ponds (Limnephilus indivisus) (Table5A) deposit eggs in damp channels or depressions before thehabitat fills with autumnal rains or snow. A similar selection ofhabitat before it is suitable for the larva occurs with snowpool
mosquitos (Aedes sp.), which deposit diapausing eggs indepressions, and hatching occurs the following spring wheneggs are wetted by the melting snow.
Males are responsible for habitat selection in manyodonates; they establish territories over ponds or streams,depending on the species, and actively court females to ovipositwithin that locale. A male in tandem flight may even remaincoupled while the female is ovipositing below the surface.Belostomatid bugs, which incubate eggs on the back of themale (Table 5A), are another instance of male influence onhabitat selection. Movement by the male through swimming or

50 LIFE HISTORY
Table 5A. Summary of data for oviposition and egg stage of selected aquatic insects.
EPHEMEROPTERABaetidae
Baetis spp. 1 day (includingsubimago)
extended (spring-fall) Shallow riffles, under-side of stones
v folds wings along abdomenwalks into water and selects site
Callibaetisfloridanus
EphemeridaeHexagenia spp.
LeptophlebiidaeLeptophlebia
cupida
EphemerellidaeEphemerella
ignita
HeptageniidaeEpeorus pleurialis
ODONATALestidae
Lestes congener
Archilestesgrandis
AeshnidaeAnax imperator
CorduliidaeTetragoneuria
spp.
5-7 days (includingsubimago)
2-4 days (includingsubimago)
2 days (includingsubimago)
2 days (includingsubimago)
3 days (includingsubimago)
3wk
-
11- 14 days
1 wk
entire year Lentic, often temporaryhabitats
May-Sept Lakes, large rivers
mid May-early July Mainstream of slow-moving rivers
late June-Sept Turbulent streams, inmoss
Mar-May Streams, fast water
Aug Temporary ponds; onlyin dry stems of Scirpussp., 5-30 cm abovewater
mid June-late July Lentic; in branches orpetioles up to 13 mabove water
early June-early Aug Ponds; warmest area
mid May- July (ca. 1 Lotic, ca. 10 or moremo at a site) (individu- feet from shore
V lands on water and extrudesmilky mass; dies on site
V V plummet to water and extrude 2egg packets; on contact with water,eggs separate and sink. Some 9 9can take off and repeat
9 dips abdomen in water, releases afew eggs at a time; completed in 5min; flies up or down stream,daytime only
9 flies upstream with extruded eggball; contact with water releasesball, and eggs then separate
9 touches water several times,washes eggs off in batches
C? + 9 in tandem; eggs insertedsingly in incisions 2 cm apart
C? + 9 in tandem; oviposition dur-ing daytime. Endophytic oviposi-tion; 2 eggs per incision in petiolesbut about 10 in pithy stems
9 alights on floating plants and de-posits eggs endophytically; eggmay penetrate leaf; 1 egg per inci-sion
9 makes 2 passes to select a site;on 3rd pass she drags abdomen
als live 2 wks) through water trailing egg stringuntil it adheres to solid object.Several 9 9 add to egg-stringaggregations on same day

LIFE HISTORY 51
Table 5A. Continued
contiguous rows of yel-low, spherical eggs, forma flat semicircular plate
viscous, reddish brownegg mass on venter ofabdomen; mass is re-leased and eggs separatewhen abdomen toucheswater
eggs ellipsoid,.16-.19X.28-.32 mm;sticky surface results ineggs adhering in clumps
ovoid, .23 X .12 mm;eggs anchored by peg-like structures thatspring out after wetting
greenish egg ball; eggshave polar anchoringmechanism to attach tosubstrate
cream colored; ellipsoid,.08-.09X.13-.15 mm;adhere firmly to substrate
elongate, 1.2 mm long;soft and greyish whendeposited; cuticle thenhardens and darkens
white with dark anteriorend; 1.9X .3mm
elongate cylinder,.2 X .4 mm, with ante-rior bladelike projectionto anchor egg in leaf
oval, brown eggs,.7 X .4 mm, within astring of gelatinous ma-trix that swells to 3.5nun wide X 1 1 cm long
B. rhodani, winter gen-eration up to 2500; sum-mer generation up to1200
450-500
2260-7684;*= 4000for average-size 9 9(24-25 mm)
x (mean-sized 9 ), 2959;* Fecundity, F = 2.02 L3-04
eggs per mass, 156-603;x = 322
2000-6000. x = 4260(dissected 9 9 )
87-297 (x=205) ma-ture oocytes; no. of eggclutches per 9 not deter-mined
up to 149/9; 70-180(Smith and Pritchard1956)
no data; can maturesuccessive batches ofeggs
up to 1000 per string.Egg-string aggregationaverages 250 thousandeggs; exceptionally largemass contained over1 million
B. rhodani, start ofhatching 17 wks @3°C, 1 wk @ 22°C.Hatching interval 34days @ 3°C, 3 days @22°C.
Hatch in 5- 10 sec(ovoviviparbus)
1 1-26 days; 2 wk @summer temp
Hatching started in10-14 days @20°C;50% emerge on day 1,hatching continues for43 days
112-392 days @8.2°C;56-294 days® 15.8°C.In stream, 10% hatch by100 days, 90% by 300days
Estimated 7 mo in field@ 11-14°C
Prediapause develop-ment for 1 wk; 3 modiapause; 6-7 wk post-diapause; synchronoushatching (1 wk,in May)
15 days; eggs hatchwithout wetting
Direct development; atfield temp., 22-26 days;in lab, range of 28-51days @ 15-20°C
Individual strings in lab,hatching began @ 2 wk,50% complete by 3 wk,continues for 7 wk
Hatching period is vari-able for species and site.Delayed hatch of someeggs for 40 wk
Adapted for temporaryhabitats. 9 9 mate, andembryonic developmentcomplete before oviposi-tion
Both low temp, and lowD.O. will delay incuba-tion (Fremling 1967)
Latitude effect on ovipo-sition period and fecun-dity. 1072-2065 eggsper 9 in Penn.
Extended hatching pe-riod. Temp. > 14.5°Cdelays hatching; eggdiapause in some popu-lations
Extended hatching pe-riod, with peak of smalllarvae in Feb
Diapause occurs undersnow cover, post-dia-pause embryogenesistriggered by snowmelt.Hatching threshold is5°C
At emergence, prolarva"jumps" to water, use ofdeciduous petioles foregg site possible becauseeggs do not overwinter
Long oviposition periodresults in many larvalsize classes overwinter-ing; thus, synchrony re-quired in final instar
Hatching of eggs withinlarge aggregations is re-tarded and thousandsfail to hatch
Europe 1050, 1910North America 661
Florida 288, 4049
Midwest 1303, 1871,1309'
Alberta 663, 3921Pennsylvania
England, Lake 1052District
Kentucky 2716
Saskatchewan 3549
Oklahoma 306
Southern England 698, 701, 703
Michigan 2163
'Fecundity of many mayflies conforms to power law: F = alA where L = body length, and a and 6 are constants (Elliott 1972; Clifford and Boerger 1974).

52 LIFE HISTORY
Table 5A. Continued
PLECOPTERANemouridae
Nemouratrispinosa
Amphinemuranigritta
PerlodidaeHydroperla
cmsbyi
PerlidaeParagnetina
media
HEMIPTERAGerridae
Gerris spp.
CorixidaeRamphocorixa
acuminata
t
2-3 wk (feeding required) mid June-early July Lotic, midstream Extruded as mass; flying 9 dipsabdomen into water; mass"explodes" when jelly expands
as above June As above As above (?)
2-5 days Feb-Mar Lotic, head of riffle 9 alights on water, extrudes eggmass. Mass separates and eggssink
< 1 day June (synchronous Lotic Extruded as massemergence)
several mo (adults spring and summer Pools and running water 9 glues eggs to floating objects oroverwinter) at water's edge, at or below water-
line; deposited in parallel rows
— early spring-fall; some Ponds, stock watering Preferentially on crayfish (Camba-eggs overwinter holes rus sp.); also on other smooth sur-
faces. Several 9 9 oviposit on onecrayfish
NotonectidaeNotonecta
undulata
BelostomatidaeAbedus herberti
MEGALOPTERACorydalidae
Orohermescrepusculus
SialidaeSialis rotunda
(Buenoasp.= 16 days; early spring through Lentic, widespreadBare 1926) summer (clear pools to slimy
ponds)
— nonseasonal Warm streams andponds
few days July-early Sept Above lotic waters;bridges, trees, rocks
1 day late Apr-June On vegetation or other
Eggs glued to submerged plantsand other objects; irregular spacing
Starting at apex of wings and mov-ing forward, 9 deposits a solidmass of eggs on dorsum of C?
9 deposits rows of eggs; she mayadd 3-4 smaller tiers on top of thebase layer
9 secretes adhesive, then depositsobjects overhanging len-tic waters
upright rows of eggs forming atight mass

LIFE HISTORY 53
Table 5A. Continued
sticky coating attacheseggs to substratum
114-833/batch;x =514
Immediate development@ 10°C. Incubationminimum of 3 wk, 80%by 6 wk; continues to 12wk
In field, hatching frommid July to mid Sept.
Ontario 1574
brown, oval, triangularcross section. Gelatinouscoating glues eggs tosubstratum
—
elongate, cylindrical,3 X 1 mm; white, turn-ing amber brown beforehatching
elongate, oval, .9 X .4mm, with apical nipple.Egg attached by elasticpedestal and disk withstrong glue
elongate, oval, 1.7 X. 6mm; with small tubularmicropyle; rough sur-face with hexagonalsculpturing
oval with rounded top;yellow, darkening to tan.Egg swells during devel-opment from 3.1 X 1.7mm to 5.0 X 2.0 mm
oblong mass, 30 X 25mm. Eggs, 1.0 X. 5mm, with micropylarprojection; greyish yel-low, becoming reddish
90-188/batch;x-=12I
172-330 eggs/mass;up to 3 masses/ 9 ;442-1418, x = 787(dissected 9 9 )
3-7 batches; 1207-2929egg/ 9 (dissected 9 9)
—
up to 22 mature eggs inabdomen at one time;successive batches arematured
one 9 produced 4 massesin 13 mo, total = 344eggs
1000- 1700 eggs/massx = 1500
4-mo embryonic dia-pause (germ disk stage)
7 mo egg diapause toSept-Oct
At 20°C, 32 days; con-tinues for 58 days
2 wk incubation
5-10 day incubation
5- 14 day incubation
Incubation in lab @18°C=21-23 days
In field, 26-63 days,x =43 days; in lab(20°C), 25days
In lab, hatching began Ontario@ 12°C in Sept.; contin-ued to Nov with temp,decrease from 12 to 8°C
Synchronous hatching Texaswith decreasing temp,from 25 to 19°C
Partial hatch in fall, re- Ontariomainder in spring. Someparthenogenesis
<3 of Rhagadotarsus Kansascalls 9 with wave pat-terns; defends oviposi-tion territory (Wilcox 1979)
Continuous reproduction Kansasand asynchronous incu-bation. Synchrony ofoviposition induced byfreezing or recoloniza-tion. Oviposition oncrayfish gives protection,aeration, and solid, non-silted, attachment sur-face
Continuous oviposition Kansas,in summer and overlap- New Yorkping broods. Some noto-nectids insert eggs intoplants
Cf aerates eggs by raising Arizonaand lowering wings.Encumbered Cf C? occurthroughout the year
Synchronous hatching Oregonwithin a mass
1574
2953
1573
1857
1484
1857
3715
1114
cylindrical, rounded attop with micropylar tu-bercle; .7 X .3 mm;white when laid, turningdark brown before hatching
300-500 eggs/mass;9 may deposit a second,smaller mass
8-12 days Synchronous hatchingwithin a mass, usuallyat night. Larvae dropfrom egg mass into thewater
Oregon 137,138

54
Table 5A. Continued
LIFE HISTORY
TRICHOPTERAGlossosomatidaeAgapetusfitscipes
Agapetus bifidus
few days Apr-Oct
July-early Aug
Chalk streams, slowflow over clean gravel
Streams, in crevices oncobble
V swims underwater and ovipositson rock, then places "capstone" ofsmall gravel on egg mass beforecement is dry; submerged for15-20 min
T probably swims to substrate
HydropsychidaeHydropsyche sp.Cheumatopsyche sp.
few days May-Sept Large rivers, on sub-merged objects
V swims underwater, deposits eggmass on firm surface. Masses con-centrated 3-5 ft below surface
LimnephilidaeLimnephilusindivisus
several wk (ovarial early falldiapause over summer)
Dry basin of temporarypool under log or to other protected site
Limnephiluslunatus
variable: 2-3 wk in fall; fall> 3 mo in spring
Chalk streams, attachedto watercress above water. Mass absorbs water
and swells to 10 mm in diameter
Clistomniamagnifies
CalamoceratidaeHettroplearonspp.
2 wk (in lab)
few days? (pupaecontain mature eggs)
luly-Aug
early summer
Lentic, attached to sub-merged logs or plants, orloose in littoral benthos
Small streams; looselyattached masses nearwaterline; also in dropzones not attached
In lab, V observed to enter waterfor 5 min and attach egg mass.May also oviposit on surface andegg mass sinks to substrate
Some 9 V may oviposit underwater,but site of eggs suggests that mostoviposit at the waterline
LeptoceridaeCemclea spp. < I day spring, or spring and
mid-late summerLentic and lode
until it absorbs water, then sinksand adheres to submerged objects
ChathamiidaePhilanisusplebeius
fewwk summer-fall Marine; in starfish(Patiriclla) in tidepools through pores on aboral surface of
starfish to deposit eggs in coelom

LIFE HISTORY 55
Table 5A. Continued
round, .2-.25 mm indiameter, creamy white;deposited in single-lay-ered, compact mass
eggs/mass= 12-94,x = 27. In lab, 12 9 9deposited 70 masses
Direct development. At12°C, hatching starts@ 1 mo, and continuesfor 3 wk
In cold springbrook,eggs hatched from earlyMay-Get
Southern England 86
mass, firm matrix,I .2X 1.6 mm. Eggsround, .2-3 mm in di-ameter
concentric rows ofclosely packed eggs(Badcock 1953)
dome-shaped mass,opaque yellow when de-posited. When swollen,jelly is colorless witheggs in rows. Eggsbluntly elliptical,'.4 X .3 mm
spherical colorless mass,up to 3 cm in diameter,takes 8 hr to achievefull size. Eggs greenish,arranged in lines
egg mass spherical,7-15 mm, coaled withsilt, jelly very fluid; eggsyellowish, arranged in
mass, dark green, ca. 1mm sphere when depos-ited. Eggs spherical,green yolk, transparentchorion; eggs swell from.1 mm to .2 mm indiameter by time ofhatching
eggs spherical, .4 mm,yellow-yellowish gray;single or in smallclumps in coelomiccavity of host
30-100 eggs/mass
Hydropsyche, dissected9 9 =331-465 eggs(Fremling 1960a). 820deposited in 50 min by a9 (Badcock 1953)
270-636/mass
200-300 eggs per mass;9 may deposit a secondsmaller mass
132-348 eggs per mass,x = 209. Dissected 9 9contain 300-400 eggs
100-300 eggs per mass
dissected 9 9 contain upto 400 mature eggs,x = ca. 160. Variablenumbers in starfish:mostly < 10, but up to112
7 mo to hatching
In lab, 8-11 days
Larvae hatch in few wk,but remain in matrixuntil flooded
Duration of egg stage(room temp.) averages17 days
2-2-1/2 wk @ 16°C; lar-vae continue to emergefrom a mass for 1 wk
At 20°C, hatching @12 days; 15 days®15°C; larvae remain inmatrix 2-9 days
In lab, 1-3 wk
Eggs overwinter inobligatory diapause
Oregon 92
At least 5 wk incubation@ 16-18°C
Emergence from eggmass dependent on timeof flooding; may be infall or the followingspring
Fall oviposition synchro-nized by decreasingduration of ovarial dia-pause as summer pro-gresses
Field oviposition periodextended by long flightperiod. May be 2-3 moovarial diapause inspring
Short, synchronous ovi-position period. Eggsabove waterline require100% humidity; larvaeremain in mass until in-undated, then allemerge in ca. 1 min
Direct embryonicdevelopment
Extended incubationand hatching periods aseggs were found in star-fish throughout the year
England, Iowa 145,1302
Ontario 4381,4395
Southern England 1461
Oregon, BritishColumbia
Pennsylvania
Oregon
Kentucky
89,4482
3034
R. Wisseman(unpublished)
3291
New Zealand 4487

56
Table SA. Continued
LIFE HISTORY
COLEOPTERAGyrinidaeDineutus spp. May-Aug Ponds, on underside of
Potamogeton sp. leavesDeposited in clusters; each eggglued separately to leaf
HaliplidaePeltodytes sp.,
Haliplus sp.
DytiscidaeAgabus erichsoni
May-early July
May-June
Lenb'c, in beds of Chamsp. and Nitella sp.
Temporary woodlandpools
Peltodytes, eggs glued to macro-phytes or algae.Haliplus, 9 chews hole inhollow stems and depositsseveral eggs within
Eggs deposited in clumps of 2-3among root fibers or mosson bottom of pond
Colymbetessculptilis
late Mar-April Temporary woodlandpools
Eggs firmly attached to submergedvegetation or to edge of rearingcontainer
HydrophilidaeHydrophilus
triangular!*early-mid summer Eutrophic ponds, with
some vegetation9 spins a silken egg case; ellipso-idal shape with elongate "mast."Construction takes > 1 hr
PsephenidaePsephenus falli
ElmidaeStenelmissatineata
(mature ovariesat emergence)
early May-mid Aug
May-Aug
In riffles, under rocks
Lotic, in riffles, on sidesand bottom of rocks
V crawls down a rock and remainssubmerged for life (1-3 days)
Submerged 9 selects depressions orcracks on rocks; deposits group ofeggs usually touching each other;each egg pressed against surfacefor 10-20 sec to glue it down
LEP1DOPTERAPyralidae
Nymphula sp. 1 day July-Aug Lentic, underside offloating Potamogetonsp.leaves
v generally does not enter waterbut extends tip of abdomen to at-tach egg mass on underside nearmargin of leaf. Oviposition occursat night
HYMENOPTERAAgriotypidae
Agriotypus sp. few days May-July Lentic or lotic; in casesof goerid or odontoceridcaddisflies
V crawls down a support into waterand searches for a host. Eggs onlydeposited on prepupa or pupa. 9may stay underwater for severalhr, enveloped in air bubble

LIFE HISTORY 57
Table 5A. Continued
white, elongated ellips-oid, 1.9 X.6 mm, clus-ter arranged diagonally@ 45° angle from mid-rib
Peltodytes, oval, withprojecting plug, .5 X .3mm, yellowish brown;Haliplus, oval, .4 X .2mm, whitish
short oval; pale creambecoming light brownwith age; 1.7 X 1.1 mm
elongate oval, somewhatkidney-shaped; pale yel-low with smooth cho-rion; 1.8 X .7 mm
egg case is yellow, turnsbrown; eggs, elongate el-lipsoid, 4.4 X 1 mm;bright yellow
spherical, lemon yelloweggs, deposited in com-pact, single-layeredmass
oblong: whitish-yellow;.55-.62 mm long
7-40 eggs per cluster(Wilson 1923b), 12-17eggs per 9 per day(Istock 1966)
30-40 eggs within a wk
9 9 contain 14-31 eggsat one time; ovaries con-tinue to develop eggs
10-130 eggs per case; 9probably matures morethan 1 batch
ca. 500 eggs per 9 ; sev-eral may oviposit to-gether, forming massesof over 2000 eggs
5-6 days
8-10 days® 21 °C
8-9 mo; some embry-onic development beforepond dries up, then dia-pause; hatching occursthe following spring
6 days @ 19°C; longerin field as oviposition oc-curs at < 14°C
16-17 days @ 23°C
6-10days@22-25°C
Synchronous hatchingof clusters, extendedoviposition period of thepopulation
Iowa, Michigan
9 9 live over 1 yr, so sev- Michiganeral batches of eggs arematured
Eggs from dry pond bot- Ontariotorn were chilled for 3mo @ 0°C, then floodedand larvae emergedwithin a few hr
Apparently a short incu- Ontariobation and hatching pe-riod, as Ist-instar larvaeonly found for 3 wk inApril
Egg case floats and eggs Iowado not hatch if caseturns over; mast as-sumed to aid in stabiliz-ing the case
1917,4464
1710,2515
1955
1955
4463
Apparently synchronoushatching within a mass,but extended ovipositionperiod
Protracted oviposition Kentuckyperiod; adults live un-derwater for > 1 yr
Southern California 2821
4328
elliptical eggs, .45 X .6mm; light grey or whi-tish; about 20 eggs/
9 of N. badiusalis laid441 eggs in one night
6-11 days Direct development ofeggs; synchronoushatching within a mass
Michigan 264
elongate, .9 X .2 mm;tapered to a stalk whichis inserted into thehost's integument
In lab, 5-8 days Several eggs may be de-posited on one host butonly one larva can de-velop per host
Japan, France 649,1475

58 ,, t
Table 5A. Continued
LIFE HISTORY
DIPTERATipulidaeTipula sacra < 1 day June-July Lentic, in soil or algae
mats near shore9 9 emerge during the day; mateand begin ovipositing immediately
Lipsothrixnigrilinea
PtychopteridaePtychopteralenis
< 12 hr
< 1 day
Mar-Aug peak inMay-June
late May-June
In saturated wood instreams
Lentic, stagnant water
Q searches for suitable site on woodnear waterline with ovipositor. De-posits egg ca. 1 mm deep in softwood or cracks; then moves to makeanother insertion
Mating and oviposition occurshortly after emergence. Eggsoccur loose on substrate, so proba-bly scattered at pond surface andsink to substrate
SimuliidaeSimulium spp. variable; blood meal spring and summer;
may be required for egg multivoltinematuration
Lotic; various sites (wet-ted vegetation, damfaces, debris, etc.)
Variable even within a species;may oviposit in flight, butmore commonly on solid surface inmasses or strings, at or belowwaterline
CulicidaeAedes aegypti variable; blood meal re- nonseasonal
quired for egg developmentartificial containers: cis-terns, cans, old tires
Eggs deposited singly, at or nearwaterline
Culex pipiens variable; blood meal re- spring-late autumn Lentic; small catch-quired, except in autogen- ments and pools withous strains; some overwinter high organic contentas nulliparous 9 9
9 lands on water and deposits eggsin raftlike masses. Oviposition usu-ally at night
ChironomidaeChiwnomusplumosus
2-5 days mid May, July-Sept Lentic; on water or onflotsam
9 flies over water (sometimes sev-eral mi); extrudes egg mass be-tween hind tibiae and deposits iton first surface that she touches
TabanidaeTabanus atratus I w k June-Oct On plants, near or over
water9 faces head downward while de-positing egg mass on vertical por-tion of plant
EphydridaeDichaela sp.(= Notiphila)(Mathis 1979a)
SciomyzidaeSepedon spp.
5-15 days
4-24 days
throughout summer Marshy areas with ac-cumulation of decayingvegetation
Lentic; on emergentvegetation, from 5 cm to> I m above water
9 scatters eggs along shore or onfloating detritus. Eggs hot glued tosubstrate but many in crevices
9 in head downward position, de-posits eggs in vertical row

LIFE HISTORY 59
Table 5A. Continued
shining black, elongate,convex on one side; 1.0X .4 mm; posterior fila-ment uncoils when wet-ted as anchoring device
cream colored, elongate,smooth; no anchoringdevice
dissected 9 9 , x = 925,range, 500-1600 eggs
dissected 9 9 , x = 185,range, 106-380 eggs
In lab, few days; infield, < 1 mo
About3wk@ 16°C
Direct development of Albertaeggs; hatching periodfrom early JulyrmidAug
Direct development, but Oregonextended hatching pe-riod because of longflight period
3199,3206
979
elongate oval; whitishyellow; longitudinal re-ticulations on chorion;.8-.9 mm long
dissected 9 9 contain530-806 eggs
In field, 14-20 days Egg maturation occursduring pharate adultstage
Alberta 1790
oval to triangular .25 X.14 X.13 mm; whitish,turning brown as theymature.
300-600 eggs per 9 .Eggs may occur inlarge aggregations(72,000/ft2)
5 days @ 23°C Successive generationsin summer; overwinteroften as diapausing eggs
Ontario 853
elongate oval
cylindrical, tapered
egg mass is dark brown,tear-shaped; swells to 25X 5 mm. Eggs, creamcolored, oval, .5 X .2
average about 140 eggswhen fed on humans;may be 2 or more eggcycles
100-400 eggs per mass;9 lays 2-4 masses
Highly variable; embry-onic development com-pleted in 2-4 days afterflooding
1-3 days
Direct development inwater but eggs with-stand desiccation for atleast 1 yr
First batch of eggs maymature without a blood
eggs per mass: X =1676, range, 1154-2014
3 days @ 24°C; 14 days@9°C
meal. Size of latermasses depends on bloodmeals. Several genera-tions per yr
Egg mass floats and lar-vae remain in it for 1day after hatching. 2generations per yr
Southeastern U.S.
Holarctic
1618
1618
Wisconsin 1725
egg mass is subconical,oval at base, with 4-5tiers of eggs, 5-25 mmX 2-10 mm. Eggswhite when laid, thendarken
egg ellipsoidal, convexon venter; longitudinallyridged; white; .9 X .3
eggs lie horizontaltouching preceding one.Egg elongate withcoarse, longitudinalstriations; white, becom-ing colored during de-velopment
500-800 eggs per mass 4-12 days Florida 2018
Up to 25 eggs per row;9 probably deposits sev-eral rows
1-2 days @ 21-25T
3-5 days
Eggs float when marshfloods and have plastronfor underwater respira-tion
Ohio, Montana
USA
1009
2871

60 • LIFE HISTORY
Table 5B. Summary of data providing information on life histories and vctfnism of selected aquatic insects.
EPHEMEROPTERAAmeletidaeAmeletus spp.
BaetidaeBaetis spp.Baetis pygmaeusBaetis spp.Baetis quilleriCallibaetis floridanusCloeon trianguliferHetervcloeon curiosum
IsonychiidaeIsonychia bicolorIsonychia spp.
PseudironidaePseudiron centralis
HeptageniidaeCinygmula ramaleyiCinygmula reticulataLeucmcuta sp.Nixesp.Rhithwgena semicolorataStenacron interpunctatum
Stenonema modestum
Stenonema vicariumEphemerellidae
Ephemerella (2 spp.)Ephemerella ignitaEurylophella funeralis
Eurylophella dorisC!)Eurylophella temporalisDrunella (2 spp.)Dntnella doddsi
Serratella sp.Tricorythidae
Tricorythodes atratusTricorythodes allectus (?)
Tricorythodes dimorphusTricorythodes minutus
CaenidaeCaenis spp.Caenis arnica
BaetiscidaeBaetisca rogersi
LeptophlebiidaeParaleptophlebia (2 spp.)Paraleptophlebia (4 spp.)Paraleptophlebia volitansHabrophlebia vibrans
BehningiidaeDolania americana
univoltineunivoltine
uni- to multivoltinemultivoltine (2-3/yr)multivoltine(~ 10/yr)tmultivoltine (18+/yr)tbivoltinemultivoltine (3/yr)bivoltine
bivoltinebi- or trivoltine(?)
univolrine
univoltineunivoltineunivoltineunivoltineunivoltineunivoltine to3 generations in 2 yrsbivoltinebivoltineunivoltine
univoltineunivoltinesemivoltine (2 yr)univoltineunivoltineunivoltineunivoltineunivoltineunivoltineunivoltine
bivoltinemultivoltine (4 to 5 fromMay to late Oct.)multivoltine (18+/yr)tunivoltine
uni- to multivoltinebivoltine
univoltine
univoltineunivoltineunivoltinesemivoltinesemivoltinesemivoltine
semivoltine(ca. 10 mon. as egg)
Mic
V/ficloticIt/lie\>KKfemic
lotic
k*K
lolicloticMicMicloiic
loticl(Xiclixic
loticliXicloilcloticlolicIdlelolicloticlotic
loticlotic
lolicloiic
lentlc
lotic
loiicloticloticloticloticlotic
lotic
QuebecNorth Carolina
N. America & EuropeQuebecGeorgiaArizonaVirginiaPennsylvaniaVirginia
PennsylvaniaGeorgia
Alberta
AlbertaOregonNorth CarolinaNorth CarolinaEuropeIndiana
VirginiaSouth CarolinaMinnesota
OregonEuropeOntarioPennsylvaniaGeorgiaSouth CarolinaOregonAlbertaOregonNorth Carolina
MinnesotaGeorgia
ArizonaAlberta
N. America & EuropeVirginia
Florida
OregonOntarioSouth CarolinaQuebecNorth CarolinaOntario
South Carolina
24141877
658227519421199,146963239232156
39171942
3750
324123121877187716482591
215537282193
1637164813873922194237281637324116371877
15351942
1199,1469193
658632
3094
231313873728227418771387
1616
1 based on larval growth rates obtained from in situ measurements.

LIFE HISTORY 61
Table SB. Continued
EphemeridaeHexagenia bilineataHexagenia mundaHexagenia limbataEphemera simulans
PolymitarcyidaeEphoron albumEphoron leukon
ODONATAAnisopteraGomphidae
Lanthus vemalisAeshnidae
Anaxjunius
AnaxjuniusBoyeria vinosa
CorduliidaeNeurocordulia molesta
LibelluUdaeCelithemis fasciataCelithemis elisaEpitheca (2 spp.)Epitheca (2 spp.)Epitheca cynosuraLadona deplanataUbellula (3 spp.)Perithemis tenera
ZygopteraLestidaeArchilestes grandisLestes (3 spp.)Lestes (2 spp.)
CoenagrionidaeEnallagma hageni
& E. aspersionTelebasis salvaPLECOPTERAPteronarcyidae
Pteronarcys dorsataPteronarcys proteus
Pteronarcys scotti
PeltoperlidaeTallaperla maria
TaeniopterygidaeOemopteryx contortaStmphopteryx limataTaenionema atlanticumTaeniopteryx nivalis
NemouridaeAmphinemura (4 spp.)Nemaura trispinosaOslrocerca albidipennisProstoia besametsaProstoia similisShipsa rotundaSayedina carolinesisSoyedina valliculariaZapada cinaipesZapada columbiana
univoltine loticunivoltine loticsemivoltine, with variable cohorts lenticsemivoltine lentic
univoltine loticunivoltine lotic
semivoltine lotic
univoltine (with 3 & 11- loticmonth populations)uni- & semivoltine lenticunivoltine loticsemivoltine lotic
univoltine lotic
univoltine lenticunivoltine lenticunivoltine lenticuni- & semivoltine lenticuni- & semivoltine lenticunivoltine lenticunivoltine lenticunivoltine lentic
univoltine lenticunivoltine lenticunivoltine lentic
univoltine lenticuni- & bivoltineunivoltine lentic
univoltine loticmerovoltine (3-4 yrs) loticmerovoltine (3 yis) lotic
semivoltine loticsemivoltine lotic
semivoltine lotic
univoltine loticunivoltine loticunivoltine loticunivoltine lotic
univoltine loticunivoltine loticunivoltine loticunivoltine loticunivoltine loticunivoltine loticunivoltine loticunivoltine loticunivoltine loticsemivoltine lotic
midwestSouth CarolinaManitobaManitoba
ManitobaVirginia
North Carolina
Ontario
IndianaVirginiaSouth Carolina
Georgia
South CarolinaIndianaSouth CarolinaIndianaTennesseeSouth CarolinaIndianaIndiana
North CarolinaSaskatchewanNorth Carolina
North Carolina
Arizona
VirginiaMassachuchettsVirginia & WestVirginia
GeorgiaSouth Carolina
TennesseeNorth Carolina
TennesseeTennesseeQuebecOntario
Ontario, QuebecOntarioQuebecNew MexicoMinnesotaOntarioPennsylvaniaQuebecOregonAlberta
1302'372816561656
13863738
1877
4050
451613393728
258
25445162544516249425445164516
191335491913
1914
3489
229218074340
26111235
10802949
2875287515863626
1574, 2414157424141622219215743926241420792824

62 '
Table SB. Continued
LIFE HISTORY
LeuctridaeLeuctra (2 spp.)Zealeuctra (2 spp.)
CapniidaeAllocapnia (4 spp.)Allocapnia nivicolaAllocapnia rickeriCapnia vemalisParacapnia angulata
PerlidaeAcmneuria evolutaAcmneuria lycoriasAgnetina capitata
Beloneuria (2 spp.)Calineuria califomica
Claassenia sabulosa
Eccoptura xanthenesNeoperla clymeneParagnetina media
Perlesta placida
PerlodidaeClioperia clio
Hehpicus (2 spp.)Hydmperla crosbyiIsogenoides olivaceusIsoperla (4 spp.)IsoperlafulvaIsoperla signata
ChloroperiidaeHaploperla brevisSweltsa onkosSweltsa (2 spp.)Triznaka signala
HEMIPTERAHydrometridae
Hydrometra martini
GerridaeGerris agenticollisGerris canaliculatusGerris remigisUmnoporus notabilis
BelostomatidaeAbedus herbtiBelostoma malkiniLethocerus americanus
Lethocerus maximus
PleidaeNeoplea striola
NepktaeNepa apiculataRanatra montezuma
uni- & semivoltineuni- & semivoltine
univoltineunivoltineunivoltineunivoltineunivoltine
univoltinemerovoltine (3 yts)semivoltinesemi- and merovoltine(3yrs)semivoltine ?merovoltine (3 yrs ?)semivoltinesemi- merovoltine(3yrs)semivoltineunivoltinesemivoltinesemivoltinemerovoltine (3 yrs)univoltineunivoltine
univoltine
univoltine ?univoltineunivoltineunivoltineunivoltineunivoltine
univoltinesemivoltinesemivoltineunivoltine
multivoltine (3-4/yr)
univoltinetrivoltine?bi- and univoltineunivoltine
multivoltine (4+/yr)multivoltine (5- 6/yr)multivoltine ? (33degg to adultduring summer)multivoltine(5- 6/yr)
univoltine
univoltinetrivoltine
loticlotic
loticloticloticloticlotic
loticloticloticlotic
loticloticloticloticloticloticloticloticloticloticloticlotic
lotic
loticloticloticloticloticlotic
loticloticloticlotic
lentic
lenticlenticloticlentic
loticlenticlentic
lentic
lentic
lenticlentic
OntarioTexas
QuebecQuebecMinnesotaQuebecOntario
OklahomaMichiganOntarioNew York
North CarolinaOregonCaliforniaColoradoNew MexicoKentuckyTexasKentuckyMichiganOntarioTexasGeorgia
KentuckyOntarioArkansasSoutheastern U.S.TexasMinnesotaOntarioNew MexicoWisconsin
OklahomaOntarioNorth CarolinaNew Mexico
Michigan &Massachusetts
IllinoisVirginiaNew YorkBritish Columbia
ArizonaTrinidadMichigan
Trinidad
Illinois
IllinoisArizona
15743731
15861574219215861583
1103104215732751
187720793652133116224641193946164915733732258
271915731158381129532192157316222026
1103157318771622
3780
21613463251132
14697823246
782
2635
26373487

LIFE HISTORY 63
Table SB. Continued
NaucoridaeCryphocricos hungerfordi
CorixidaeGraptocorixa serrulate
Hesperocorixa interrupta
Trichocorua reticulataNotonectidae
Notonecta hoffmanniNotonecta undulata
MEGALOPTERASialidaeSialis (2 spp.)
Sialis aequalisSialis dreisbachi
bivoltine
multivoltineH7/yr)tunivoltine
bi- and trivoltine
univoltinebivoltine?
univoltine &a few semivoltineunivoltineunivoltine
lotic
lotic
lentic& loticlentic
loticlentic
lentic&loticloticlotic
Texas
Arizona
Virginia
California
CaliforniaCalifornia
Oregon
South CarolinaMinnesota
3679
1469
347
1132
26334026
138
37282193
CorydalidaeCorydalus comutus
Nigronia senicomis
NEUROPTERASisyridae
Climacia areolarisTRICHOPTERAPhilopotamidae
Chimarra (2 spp.)Chimarra (2 spp.)Chimarra aterrimaDolophilodes distinctus
Wormaldia moestus
PolycentropodidaeNeureclipsis bimaculatusNeureclipsis crepuscularisPolycentropus maculatusPolycentropus ceiaralis
PsychomyiidaeLype diversa
HydropsychidaeArctopsyche grandis
Arctopsyche irrorataCheumatopsyche pasellaCheumatopsyche pettiti
Diplectrona modestaHydropsyche slossonaeHydropsyche (2 spp.)
Hydropsyche (2 spp.)Hydropsyche (2 spp.)Hydropsyche (5 spp.)Hydropsyche (3 spp.)Hydropsyche (2 spp.)Hydropsyche (2 spp.)Macrvstemum Carolina
univoltinesemivoltinesemi-&merovoltine (3 yrs)
trivoltine
bivoltinebivoltinebivoltinebi -& some trivoltine (?)bivoltinebivoltineunivoltine
bivoltinebivoltineunivoltinebivoltine
univoltine
univoltinesemivoltineunivoltinebivoltineunivoltineunivoltineunivoltineunivoltineunivoltinesome bivoltine?univoltineunivoltine .uni- & bivoltinetrivoltinetrivoltinebi- & trivoltineunivoltine
Macrostemum zebratumParapsyche apicalis
bivoltineunivoltineunivoltine
Parapsyche cardis univoltinet based on larval growth rates obtained from in situ measurements.
loticloticlotic
lentic
loticloticloticloticloticloticlotic
loticloticloticlotic
lotic
loticloticloticloticlotic
loticloticlotic
loticloticloticloticloticloticlotic
loticloticloticlotic
TexasWest VirginiaMichigan
Ohio
VirginiaArkansasQuebecGeorgiajt CarolinaNorth CarolinaNorth CarolinaArkansas
AlbertaGeorgiaNorth CarolinaArkansas
North Carolina
IdahoMontanaGeorgia, N. CarolinaGeorgiaMinnesotaQuebecGeorgia, N. CarolinaVirginiaMinnesota
Georgia, N. CarolinaQuebecOntarioVirginiaGeorgiaOntarioGeorgiaSouth CarolinaVirginiaNorth CarolinaQuebecGeorgia
443,364834163107
450
3019386275925718781878386
33367751878386
1878
778162825716282420275925744582420
25727592418301912992419775372830191878,34452414257

64 LIFE HISTORY
Table SB. Continued
RhyacophilidaeRhyacophila (5 spp.)Rhyacophila (2 spp.)Rhyacophila vao
GlossosomatidaeGlossosoma intermediumGlossosoma nigriorGlossosoma pentiumAgapetus sp.Agapetus illiniAgapetus occidenlisAnagapetus bemea
HydroptilidaeDibusa angata
PhryganeidaeBanksiola crotchi
BrachycentridaeBrachycentrus spinaeBrachycentrus (2 spp.)Brachycentrus (2 spp.)Micrasema kluane
LepidostomatidaeLepidostoma bryantiLepidostoma (2 spp.)Lepidostoma (2 spp.)
LimnephilidaeClistoronia magnifcaDesmona bethulaDicosmoecus gilvipesEocosmoecus (2 spp)Goerita semataLimnephilus indivisusOligophlebodes zeltiOnocosmoecus unicolorPhilocasca demita
Pycnopsyche (3 spp.)Pycnopsyche (2 spp.)Sphagnophylax meiops
UenoidaeNeophylax consimilisNeophylax mitchelliNeophylax (5 spp.)Neothremma alicia
SericostomatidaeAgarodes libalisFattigia pele
MolannidaeMolannaflavicomis
HelicopsychidaeHelicopsyche borealis
Helicopsyche limnellaHelicopsyche mexicana
CalamoceratidaeHeteroplectroncalifomicum
univoltineunivoltinesemivoltine
bivoltinebivoltinebivoltineunivoltineunivoltineunivoltineunivoltine
bivoltine
univoltine
univoltineunivoltineunivoltineunivoltine
univoltineunivoltineunivoltine
univoldneunivoltinesemivoltinesemivoltinesemivoltineunivoltineunivoltineunivoltineunivoltine
univoltineunivoltineunivoltine
univoltineunivoltineunivoltinesemivoltine(2yrs)
univoltinesemivoltine(2 yis-someunivoltine?)
univoltine
univoltine?univoltinebivoltinemultivoltine(~7/yr)
semivoltine(2yrs)
loticloticlotic
loticloticloticloticloticloticlotic
lotic
lentic
loticloticloticlotic
loticloticlotic
lenticloticloticloticloticlenticloticloticterrestrial(saturatedleaf litter)loticloticlentic
loticloticloticlotic
loticlotic
lotic
loticloticloticlotic
lotic
South CarolinaOntarioAlberta
MinnesotaNorth CarolinaOregonNorth CarolinaArkansasOregonOregon
Kentucky
British Columbia
North CarolinaMontanaMinnesotaMinnesota
MinnesotaOregonNorth Carolina
British ColumbiaCaliforniaMontanaPacific NorthwestNorth CarolinaOntarioAlbertaOregonOregon
QuebecVirginiaN.W.T. Canada
North CarolinaNorth CarolinaOntarioAlberta
VirginiaNorth Carolina
Alberta
OntarioCaliforniaArkansasArizona
Oregon
24672480920
2194137592137538613751375
3305
4483
3444163021942194
219431121878
4482110116294398115533382961451484
241534154477
137518782172961
34151878
3337
444011563861469
93

LIFE HISTORY 65
Table 5B. Continued
LeptoceridaeCeraclea transversaCeraclea excisaMystacides interjectaNectopsyche albidaOecetis immobilisOecetis inconspicua
Triaenodes injustaYlodes frontalis
LEPIDOPTERAPyralidae
Eoparargyractis pleviePetrophila canadensisPetrophila confusalis
NoctuidaeBellura gortynoides
COLEOPTERAAdephagaGyrinidae
Dineutus (2 spp.)Haliplidae
Haliplus immaculicollis &Peltodytes (2 spp.)
DytiscidaeAcilius (3 spp.)
Agabus spp.CybisterfimbriolatusColymbetes spp.
Copelatus glyphicusCoptotomus spp.Dytiscus (4 spp.)Graphoderus (6 spp.)
Hydaticus (2 spp.)
Ilybius spp.Laccophilus maculosusMatus bicarinatus
Rhantus binotatus
Thermonectus ornaticollisNoteridae
Suphisellus puncticollis
HydiophilidaeTropistemus ellipticus
Psephenus montanus
Sciltidae (= Helodidae)Scirtes tibialis
univoltineunivoltineunivoltineunivoltineunivoltineunivoltine?univoltineunivoltineunivoltine
univoltinebivoltinemultivoltine(2-3/yr)univoltine
bivoltine
univoltine
uni- & bivoltine(?)
primarily univoltinefew bivoltineprimarily univoltineunivoltineunivoltineunivoltineunivoltineunivoltineunivoltineprimarily univoltinefew bivoltine
primarily univoltinefew bivoltineprimarily univoltinebivoltineprimarily univoltinesome bivoltineprimarily univoltinefew bivoltineunivoltine
univoltine
bi- & trivoltine
semivoUine (2 yrs)some univoltine?
univoltine
loticloticloticlenticloticlenticloticloticlotic
lenticloticlotic& lenticlotic
lentic
lentic
lentic
lentic
lenticlenticlenticlenticlenticlenticlenticlentic
lentic
lenticlenticlentic
lentic
lentic
lentic
lotic
lotic
lentic
KentuckyAlbertaAlbertaIndianaAlbertaBritish ColumbiaAlbertaAlbertaAlberta
New HampshireNew YorkCalifornia
Montana
Indiana
Michigan
Michigan
Wisconsin
WisconsinWisconsinOntarioWisconsinWisconsinWisconsinWisconsinWisconsin
Wisconsin
WisconsinWisconsinWisconsin
Wisconsin
Wisconsin
Wisconsin
Arizona
Arizona @elev.=2,576 ma.s.l.
Wisconsin
1744
1744
174617431745
1743
2175

66 >
Table SB. Continued
LIFE HISTORY
ElmidaeAncymnyx variegataDubiraphia quadrinotataLara avara
Macrvnychus glabratusOptioservus fasliditusPromoresia elegansStenelmis crenata
Stenelmis nr. bicarinataStenelmis sexlineata
PtilodactylidaeAnckytarsus bicolor
ChrysomelidaeDonaria cinereaGalerucella nymphaeae
G. nymphaeaeCurculionidae
Lissorhopterus simplex(=oryzophilus)
DIPTERANymphomyiidae
Nymphpmyia spp.= (Palaeodipteron)Deuterophlebiidae
Deuterophlebia (4 spp.)
BlephariceridaeBlepharicera williamsae
TipulidaeAntocha saxicolaDicranota spp.Dicranota bimaculataLipsothrix sylviaLipsothrix (2 spp.)
Tipula sacra
Tipula abdominalisPtychopteridae
Ptychopteru lenis lenisCulicidaeAedes taeniorhynchus
Culex nigripalpus
Culex resluransCbaobohdae
Chaoborus americanusC. trivittatusC. punctipennis
CeratopogoniidaeCulicoides variipennisSphaeromiini &PaJpomyiini
univoltineunivoltinemerovoltine(4 -6+ yrs)univoltinesemivoltinesemivoltinemerovoltine(3 yrs)univoltine ?uni- and semivoltine
semivoltine(2-3 yrs)
uni- and semivoltinemultlvoltine (8+/yr)t
bivoltine
univoltine(?) withmultiple cohorts (egg toadult in 36 days) -depending on flooding
primarily bivoltine,some trivoltine?
primarily univoltinesome bi- & trivoltine
univoltine
bivoltineunivoltineunivoltineunivoltinesemi- & merovoltine<3yr)semivoltine withsome univoltineunivoltine
univoltine
multivoltine (6 -15days: egg to adult)multivoltine (8 -10generations/yr)multivoltine
univoltinesemivoltine (2 yrs)bivoltine
multivoltine (3/yr ?)univoltine
loticloticlotic
loticloticloticlotic
loticlotic
lotic
lenticlotic &lenticlentic
lentic
lotic
lotic
lotic
loticloticloticloticlotic
lentic
lotic
lentic
salt marsh
lentic& containerslentic
lenticlenticlentic
lenticlentic
South CarolinaSouth CarolinaOregon
South CarolinaOntarioOntarioQuebec
OntarioKentucky
Quebec
SwedenGeorgia
Sweden
Arkansas
372837283824
3728395639562328
39564328
2329
29944190
2997
1916
Eastern N. Amer.
Western N. America
North Carolina
OntarioMinnesotaEnglandVirginiaOregon
Alberta
Virginia
Alberta
Florida
Florida
Ontario
British ColumbiaBritish ColumbiaNorth Carolina
New YorkNorth Carolina
731
728
1375
1326219310543426981
3199
3415
1790
2841
2840
4442
114711471010
2787383
t based on larval growth rales obtained from in situ measurements.

LIFE HISTORY 67
Table SB. Continued
SimuliidaePmsimulium
mvOum/fuscumSimuliumjenningsi
ChironomidaeChironomus ripariusChironomus (2 spp.)
Chimnomus plumosusChironomus tenuistylus
Cladotantarsus mancusCorynoneura sp.
Cricotopus bicinctusCricotopus triannulatusCricotopus trifasciaCricotopus sp.
Diamesa incallida
Diamesa nivoriundaDicrotendipes sp.
Einfeldia synchronaEutiefferiella brevinervis
Eukiefferiella devonicaGlyptotendipes paripesHeteniella sp.
Micropsectra sp.
Orthocladius obumbratus
Pagastia sp.Parachaetocladius
abnobaeusParatendipes albimanusPseudorthocladius sp.Rheosmittia sp.Rheotanytarsus exiquusRobackia demeijereiXylotopus par
TabanidaeChrysops (several spp.)
Tabanus attratus
Tabanus jumipennisAthericidae
Athervc lanthaEmpididae
several species
Hemerodromia empiformisEphydridae
Dichaeta caudata
Discocerina obscurella
univoltine
multivoltine(3-6 /yr )
univoltinemerovoltine(7 yrs?)bivoltinemerovoltine(3 yrs)bivoltinemultivoltine(7-8/yr)ttrivoltinebivoltinebivoltinemultivoltine(~30/yr)tmultivoltine(8-10/yr)tbivoltinemultivoltine(-30/yr)t
univoltinemultivoltine(4/yr)trivoltineunivoltinemultivoltine(2-3/yr)multivoltine(2/yr)bivoltine
bivoltineunivoltine
univoltineunivoltinebivoltinebivoltineunivoltine?univoltine
primarily univoltine
univoltine
univoltine
univoltine
univoltine?
bivoltine?
multivoltine(4-6/yr)multivoltine(2-5/yr)
lotic
lotic
lenticlentic
lenticlentic
loticlotic
loticloticloticlotic
lotic
loticlotic
lenticlotic
loticlenticlotic
lotic
lotic
loticlotic
loticloticloticloticloticlotic
lotic& lenticlotic& lenticlentic
lotic
lotic &lentic
lotic
lentic& riparianlentic& riparian
Michigan
West Virginia
AlbertaAlaska
WisconsinWisconsin
IndianaNorth Carolina
IndianaIndianaIndianaArizona
Germany
IndianaArizona
OntarioIndiana
IndianaAlbertaNorth Carolina
North Carolina
Indiana
IndianaOntario
MichiganNorth CarolinaAlbertaIndianaAlbertaMichigan
Florida
Florida
Florida
Quebec
Quebec
Quebec
Ohio
Ohio
2677
4147
3252501
1725502
2771873
2772772771469
2932
2771469
837277
11132521873
1873
277
2771387
42291873374911137492059
2018
2018
2018
2276
1575
1575
1009
1256
t based on larval growth rates obtained from in situ measurements.

LIFE HISTORY
Table SB. Continued
SciomyzidaeElgiva mfa
Pherbellia (13 spp.)
Sepedon spp.Tetanocera loewi
Tetanocera (3 spp.)
ScathophagidaeCordilura (Achaelelld) (2 spp.)
Cordilura (1 1 spp.)
Orthacheta hirtipes
MuscidaeUmnophora riparia
multivoltine(3-8?/yr)primarily multivoltine(2 - 9 /yr) with someunivoltineprimarily multivoltineunivoltine
trivoltine
bivoltine
bivoltine &univoltinebivoltine
bivoltine
lentic(littoral)lentic(littoral)
lentic (littoral)lentic(littoral)lentic(littoral)
(emergentzones)(emergentzones)(emergentzones)
lotic
Northern U.S. & Canada
Nearctic
North AmericaNew York
Northern U.S.
Virginia & WestVirginiaEastern United States
Virginia& West Virginia
England
2135
398
2871271
4046
2872
4188
2873
2681
overland crawling to nearby pools will determine the site inwhich the larvae develop.
In general, taxa with specialized habitat requirementsmight be expected to exhibit the greatest degree of habitatselection by the ovipositing female. Insect parasitism (which israre in aquatic insects) requires habitat specialization. ThePalaearctic wasp, Agriotypus sp., crawls into the waterenveloped by a film of air and seeks out a mature larva or pupaof the caddisflies Silo sp. or Goera sp. (Goeridae). An egg isthen deposited within the caddisfly case so that the Agriotypussp. larva can develop as a parasite of the pupal stage or the cad-disfly (Clausen 1931).
Oviposition and EggsThe diversity of oviposition strategies is illustrated in
Table 5A. Hinton (1981) provides a comprehensive treatmentof insect eggs. Much of the life cycle can be deduced from thetuning and habits of oviposition and the location of the eggs,but this aspect of natural history frequently receives less atten-tion than it deserves. Elliott and Humpesch (1980) point outthat information on fecundity, oviposition behavior, and hatch-ing is essential for: (1) interpretation of life cycles; (2) identifi-cation of larval cohorts; (3) study of spatial patterns and move-ments; (4) construction of life tables; and (5) estimation ofgrowth rates, mortality, and production.
Oviposition behavior has been studied extensively inmosquitoes because disease transmission usually involves anoviposition cycle before pathogen transmission occurs at thenext blood meal. Bentley and Day (1989) reviewed the inte-gration of physical and chemical cues involved in the searchbehavior. Females use vision to identify habitat and site char-acteristics. Then short-range cues become important; forexample, temperature and chemical signals received by contactchemoreceptors (see also Chap. 24).
The number of eggs varies greatly among taxa and amongindividuals of a species. Within the Ephemeroptera, Dolaniasp. deposits about 100 eggs (Peters and Peters 1977), whereasthe fecundity of Ecdyonurus sp. is 5000-8000 eggs per female(Elliott and Humpesch 1980). The number of eggs produced byBaetis rhodani ranges from 600 to 2400 per female (Elliott andHumpesch 1980). The within-species variation in fecundity isrelated to size of the fpmale, which in turn is associated withgrowth conditions of the larvae. In taxa in which feeding byadults is required for egg maturation, fecundity will depend notonly on food availability but also on appropriate ovipositionconditions, because eggs may be resorbed if conditions areunsuitable for oviposition.
The eggs of some insects mature in the pupa or last larvalstage (e.g., Megaloptera, Ephemeroptera, some Plecoptera) andare ready to be laid soon after emergence and mating. At theother extreme (e.g., Odonata, Hemiptera, some Plecoptera, andColeoptera), adults emerge with undeveloped ovaries, require afeeding period before oviposition (or overwintering), anddeposit eggs over an extended period or in discontinuousclutches. Potential fecundity can be estimated by dissection ofnewly emerged females if eggs mature all at one time. Howev-er, when egg maturation is a continuing process or when suc-cessive clutches are produced, total egg production depends onthe length of life of individual adults.
The egg is potentially a vulnerable stage because it lacksmobility. However, in many instances the egg is the most resis-tant stage of the life cycle and spans periods of cold, heat,drought, or food shortages. The hatching period may extendfor several months, which spreads the risk over a range of envi-ronmental conditions. Extended hatching may be due toextended oviposition periods, irregular rates of development ofindividual embryos, or to irregular breaking of diapause.

LIFE HISTORY 69
Larval Growth and FeedingDifferences in duration of the larval stage and in growth
patterns provide for much of the variation in life histories. Theexamples presented below illustrate food acquisition strategiesand larval growth patterns that adapt species to particular envi-ronments.
Though the fauna of temporary habitats is quite limited,most orders of aquatic insects are represented and these exhib-it extreme seasonal regulation of life cycles (Williams 1987).Wiggins et al. (1980) describe several life-cycle patterns fortemporary ponds, depending on the timing of adult colonizationand kind of resistant stage. The prime requisite for all speciesis rapid larval development during the wet phase. Manymosquitoes are temporary-habitat breeders; under optimumconditions some species can complete their growth in five days.Rapid growth implies adaptation to warm waters as even thespecies that emerge early in snow-melt pools (e.g., limnephilidcaddisflies) will be exposed to high temperatures when watervolume diminishes. The timing of recruitment in temporarypools is correlated with feeding behavior. The earliest larvaeare detritivores, either shredders (e.g., limnephilid caddisflies)or fine-particle feeders (e.g., siphlonurid mayflies, Aedes sp.mosquitoes, Chironomidae), then algal feeders (e.g., Cal-libaetis sp. mayflies, haliplid beetles) and finally predators(e.g., Odonata, Hemiptera, dytiscid and gyrinid beetles) arerecruited coinciding with abundant prey resources.
Fast development is characteristic of temporary inhabi-tants of water, but larval diapause or quiescence may extend thelife cycle to several years. Corydalidae (Megaloptera) larvae inintermittent streams burrow into the streambed when surfacewater dries up; growth only occurs during the wet cycle so lar-val duration may be 3-4 years, depending on the annual dura-tion of stream flow. The extreme example of tolerance ofdrought is the chironomid, Polypedilum vanderplanki, fromephemeral pools in Africa. The larvae can withstand completedehydration and exist in a state of suspended metabolism, orcryptobiosis. The dehydrated larvae can survive immersion inliquid helium and heating to over 100°C (Hinton 1960). In theirnatural habitat, they can survive several years in sun-baked mudand rehydrate when wetted (McLachlan and Cantrell 1980).They are normally the first invaders of small pools and caninhabit the shallowest and most ephemeral pools with virtuallyno competitors.
Many stream insects are adapted to a narrow range of cooltemperatures (cold stenothermy). Hynes (1970b) attributeswinter growth not only to use of leaf detritus as a food base, butalso suggests that this growth pattern may have been selectedfor because predation by fish would be less at low tempera-tures. Larval diapause to avoid high summer temperaturesoccurs in some early-instar Plecoptera (e.g., Capniidae, Tae-niopterygidae) and in mature larvae or prepupae of some lim-nephilid and uenoid caddisflies (e.g., Dicosmoecus sp., Pyc-nopsyche sp., Neophylax sp.).
Functional feeding groups may facilitate temporal andspatial partitioning of resources. Thus, in addition to shredders,the winter-growing stream species include scrapers (e.g., hep-tageniid mayflies, glossosomatid caddisflies), filter-feeders (fil-tering collectors, e.g., Simuliidae), and deposit feeders (gather-ing collectors, e.g., several mayflies, some Chironomidae).Within a group, coexistence may be based on differentialresponses to food and temperature. For example, three coex-
isting species of the caddisfly genus Pycnopsyche are all shred-ders and winter growers, but the food resource is partitioned bythe species having different timing of rapid growth intervalsand also some differences in microhabitat preferences (Mackay1972).
Elaborate and specialized feeding adaptations occur in thefilter-feeding Ephemeroptera, Trichoptera, and Diptera (Wal-lace and Merritt 1980). The adaptations include specializedanatomical structures (e.g., leg setae in Isonychia spp. mayfliesand Brachycentrus spp. caddisflies, mouth brushes inmosquitoes, head fans in Simulium spp.), and silk nets in manycaddisflies and some chironomid midges. Habitat partitioningwithin the filter-feeding guild occurs along the water velocitygradient. Also, there is some selectivity in feeding habits.Georgian and Wallace (1981) demonstrated that hydropsychidcaddisflies that build large-meshed nets filter larger volumes ofwater than do those species with small nets and mesh size; theformer nets will select for animal or algal foods rather than fordetritus.
Black fly larvae attach the posterior end of the body to thesubstratum with the dorsal surface facing upstream and thebody rotated 90 - 180 degrees longitudinally. Although thecylindrical body shape reduces drag, lateral deviation (yaw)created by the flowing water generates vortices which passthrough the cephalic fans (Craig and Galloway 1986, Currieand Craig 1986). In general, larvae that inhabit fast-flowingwater have smaller cephalic fans in proportion to the size of thehead, whereas larvae inhabiting slower currents have relativelylarger fans (Currie and Craig 1986). Black fly larvae show noselectivity with respect to food quality and will readily ingestinorganic as well as organic materials. Their food includesdetritus, bacteria, diatoms, and animal fragments, and particlesizes of ingested material range from colloidal to 350 urn.Because they do not select for food quality, the type of foodavailable will affect growth rates (Wotton 1994). Carlsson etal. (1977) found that black fly larvae at lake outfalls occurredin very dense aggregations and had exceptionally high growthrates associated with the availability of colloidal-sized organicmaterial washed into the river at ice melt.
The above examples are primarily winter-growing lotictaxa. Most stream species respond to increasing temperaturesin the spring by increasing their rate of growth. Multivoltinestream taxa include Baetis sp., Glossosoma sp., somehydropsychid caddisflies, and several black fly species.Though typical life cycles are univoltine, the duration and tim-ing are more indeterminate than is usually suggested. Pritchard(1978) cites examples of cohort splitting in which individualsof the same cohort may have 1-, 2-, or even 3-year life cycles,depending on food availability and environmental conditions.
Aquatic insect predators include large conspicuousspecies with relatively long life cycles compared with those oftheir prey. Some predaceous beetles belonging to the Adepha-ga are notable exceptions in that larval development occurs in afew months, or less, and adults are long-lived. Predators havea range of morphological specializations and behavior patterns.Some are ambush or "sit-and-wait" predators, whereas othersactively pursue their prey. The Hemiptera and some Coleoptera(e.g., Dytiscidae, Gyrinidae) are aquatic predators both as lar-vae and adults, whereas all others (e.g., Odonata, Megaloptera,some Diptera) are aquatic only as larvae.
Some of the most specialized predators are larval Odona-

70 LIFE HISTORY
ta; the hinged prehensile labium, or mask, is unique to thisorder. The mask is projected forward by elevated blood pres-sure induced by abdominal muscles. Prey are impaled byhooks or setae on the labial palpi. The food is then returned tothe mouth when the labium is folded back by adductor muscles.Prey perception involves receptor organs on the antennae andtarsi as well as use of the eyes. Sight is more important in laterinstars and in climbing species that live on vegetation than inbottom-sprawling or burrowing forms (Corbet 1963). Inmature larvae of visual hunters, such as Anax sp., prey captureinvolves a highly integrated binocular vision, resulting fromstimulation of certain ommatidia in each eye that enables thedistance of the prey to be accurately judged. Feeding behaviorof odonates is influenced by factors such as degree of hunger,time since the last molt, and the density, size, and movement ofpotential prey. As larvae grow, individual prey items becomelarger and more varied because larger larvae can also consumesmall prey (Corbet 1980).
Preoral digestion is a feature of hemipteran and some bee-tle predators. Salivary secretions are injected to immobilizeprey and to liquefy tissues with hydrolytic enzymes. InHemiptera the mouthparts are modified into a 3- or 4-segment-ed beak that is used to pierce the prey and suck the fluids.Dytiscid beetle larvae have chewing mouthparts, but their longsickle-shaped mandibles are grooved for fluid feeding. Larvaeof some hydrophilid beetles (e.g., Tropisternus sp. andHydrophilus sp.) are unusual in that the prey is held out of thewater. In this position, the prey juices flow down the mandiblesand into the oral opening rather than being lost into the sur-rounding water (Wilson 1923b).
Territoriality and intraspecific competition were shown byMacan (1977) to be important factors affecting growth rate andlife cycle of the damselfly Pyrrhosoma sp. In years when thelarvae were abundant and prey populations were low, two sizeclasses existed at the end of the summer. Macan attributed thisto cohort splitting; larvae in superior feeding territories grewrapidly whereas those occupying poorer feeding sites grewslowly and would either require an extra year or die of starva-tion. Furthermore, Macan suggests that fish predation wasselective for the larger specimens, which led to vacancies in thesuperior feeding sites, and that these were then readily filled bysmaller larvae. Thus, predation by fish did not greatly reducethe numbers of damselflies reaching maturity because elimina-tion of large larvae allowed smaller ones to exploit the foodresource.
The prevalence of insect predators suggests that predationmay be a dominant biotic factor influencing aquatic insects.Anti-predator defenses of aquatic insects include: refugiaafforded by the substratum; camouflage including coloration;reduced movement and feeding in the presence of predators;potential detection of predators by chemical cues; and variousdefensive repertoires used by potential prey (Williams and Felt-mate 1992). Defensive repertoires used by aquatic insectsinclude: a) reflex bleeding (Benfield 1974, Moore and Williams1990) and death feigning in stoneflies; b) deimatic behavior, asdisplayed by some mayfly prey which display scorpionlikethreat postures, or (c) accelerative drift in the presence ofpredators (Peckarsky 1980; Peckarsky and Dodson 1980).
Other defenses found in aquatic insects include morpho-logical and chemical defenses. For example, hairy bodies thatcan be effective against other invertebrates such as Hydra (Her-
shey and Dodson 1987). Some chemicals promote rapid escapeand return to shore in some semiaquatic beetles and veliid bugs.Glands at the tips of their abdomens discharge secretions thatlower the surface tension of water at the rear of the animal sothat the animal is being drawn forward by the normal surfacetension of water in front (Jenkins I960; Linsenmair and Jander1963). This mechanism propels the small insects forward atspeeds of 0.4 - 0.75 m/s, and was coined "Entspannungss-chwimmen" or expansion swimming by Linsenmair and Jander(1963). There is evidence that some mayflies can detect preyby chemoreception (Peckarsky 1980; Kohler and McPeek1989); however, in lotic systems with unidirectional or chaoticflow paths the use of chemical cues may be limited (Dodds1990). Chemical cues may be most effective only in direct con-tact with predators as this provides precise information aboutthe identity (predator or non-predator) of the intruder; and itavoids the high cost of premature escape in lotic environments(Ode and Wissinger 1993).
Evaluating the impact of predation on aquatic inverte-brates, especially in lotic habitats, has been somewhat difficult.Cooper et al. (1990) have identified several problems associat-ed with assessing the impact of predators in aquatic ecosys-tems. These problems involve primarily prey exchange (immi-gration/emigration) among reaches and may have an over-whelming influence on the perceived effects of predators. Insome cases predator/prey relationships have been assessed incages. Mesh size of cages exerts a strong influence onexchange rates and assessment of impacts of predators. Smallmeshed cages curtail exhange rates and indicate greater preda-tor effects. At larger spatial scales, drift into and out of studyreaches may influence results, and reduced prey mobilityincreases the probability of a significant predator impact onprey populations. Cooper et al. (1990) also suggested that loticecosystems may display a lower impact of predators on com-munity structure than that of lentic systems since fast-flowingstreams exhibit a much greater exchange rate among habitatsthan lentic habitats. Sih and Wooster (1994) have published arecent paper that examines some of the assumptions of theabove model of Cooper et al. 1990 that should be consulted byall students interested in predator-prey relationships.
Metamorphosis and EclosionMolting during the larval stages results in a larger insect
of essentially the same body form, whereas the final molt pro-duces a more complete change associated with the developmentof wings and other adult structures. These are considered twodistinct types of physiological differentiation, and Chapman(1982) suggests that the term metamorphosis be restricted tothe latter. Molting and metamorphosis are mediated by hor-mones. There is no fundamental physiological differencebetween the metamorphosis of hemimetabolous andholometabolous insects; the difference is a matter of degreerather than kind. The pupal instar may be regarded as theequivalent of the last larval instar of hemimetabolous insects(Gillott 1980).
The primary morphological difference between hemi-metabolous (Exopterygota) and holometabolous (Endopterygo-ta) insects is the external development of the wings in the for-mer and the delayed eversion of wings in the latter (Hinton1963). The gradual development of adult structures is apparentin hemimetabolous insects by the progressive development of

LIFE HISTORY 71
external wing pads and rudiments of the genitalia. Changesalso occur in the last larval instar of the Holometabola, butthese are internal and major differentiation is constrained bylack of space. During the pupal instar, the wings are evaginat-ed to outside the body; this makes room for development of theindirect flight muscles and the reproductive system. The pupalinstar is usually of short duration. In the strict sense, the insectbecomes an adult immediately after the apolysis (separation) ofthe pupal cuticle and the formation of the adult epicuticle towhich the musculature is now attached (Hinton 197la). Thus,in most instances, locomotion and mandibular chewing areactivities, not of the pupa, but of the pharate adult that isenclosed within the pupal exuviae.
Control of molting involves the interaction of a moltinghormone (ecdysone) and a juvenile hormone. The latter exertsan influence on development only in the presence of the former.When the concentration of juvenile hormone in the blood ishigh, the next molt will be larval-larval. At intermediate con-centrations, a larval-pupal molt occurs, and when there is littleor no circulating juvenile hormone (due to inactivity of the cor-pora allata of the brain) an adult insect will emerge at the nextmolt (Gillott 1980).
The final event in metamorphosis is eclosion, or emer-gence, which is the escape of the adult from the cuticle of thepupa or last larval instar. See Chapter 9 for a further discussionof aquatic insect metamorphosis.
The development of a pupal stage has permitted the greatdivergence of larval and adult forms in the Holometabola. Lar-vae exploit environments and food resources that result ingrowth, whereas the activities of the adult center on dispersaland reproduction. If the number of species is considered to bea measure of biological success, complete (holometabolous)metamorphosis is a prime contributor to the success of insects.Hinton (1977) states that this is a focal point of insect evolutionbecause about 88% of the extant insect species belong to theEndopterygota.
Metamorphosis in hemimetabolous aquatic insects differsin detail among the various taxa but the behaviors involved aremostly associated with switching from an aquatic to a terrestri-al mode of life (especially respiration and flight) and overcom-ing the abiotic and biotic hazards during an especially vulnera-ble stage. Data from Corbet (1963) for Odonata illustrate bothmorphological and behavioral changes occurring during meta-morphosis and eclosion.
The onset of metamorphosis in some dragonflies can bedetected several weeks before emergence when the facets of theadult compound eye begin to migrate to the top of the head.Respiration rate also increases prior to metamorphosis associ-ated with increased metabolic requirements. Behavioralchanges characteristically involve movement to shallow wateror up the stems of plants towards the surface. In Anax impera-tor, histolysis of the labium occurs and feeding stops severaldays before emergence.
Nearly all Odonata have a diurnal rhythm of emergence.The timing is presumably to restrict emergence to an intervalwhen weather conditions are favorable and when predation isleast likely to occur. The major predators are birds or matureadult dragonflies that hunt by sight, so night is the safest timeto leave the water. In the tropics, most of the large dragonfliesemerge after dusk; they eclose, expand their wings, and arethen ready to fly before sunrise. In the temperate regions or at
high altitudes where nocturnal temperatures are low, odonatestend to emerge in early morning or during the daytime. Mor-tality during emergence of Anax imperator may amount to 16%of the annual population (Corbet 1963). The individuals areimmobile and defenseless for several hours during eclosion andwhile the wings expand and harden. Cold and wind increasemortality by prolonging ecdysis or by postponing emergenceand thereby exposing more individuals to predation. Over-crowding is common in species that have mass emergences;competition for emergence supports is so intense that the firstlarva to climb a support may be used as a platform by othersthat follow. Although overcrowding is an ecological disadvan-tage of mass emergence, this type of synchronization is appar-ently adaptive, perhaps because it satiates predators and affordsthe appropriate synchronization of males and females.
Adaptations of pupae pertain to respiration, protection,and emergence from the water. These factors also operate onthe pharate adult, so it is convenient to discuss the two stagestogether. This is usually a quiescent stage, but mosquito pupae(tumblers) are relatively active swimmers, and Hinton (1958a)demonstrated that some black fly pupae both feed and spin acocoon. Pupation occurs out of water in Megaloptera, Neu-roptera, most Coleoptera, and many Diptera. Larvae constructa chamber in the soil (e.g., Megaloptera, Coleoptera) for meta-morphosis, and eclosion may occur within the chamber(Coleoptera) or after the pharate adult has worked its way to thesoil surface (Megaloptera).
Many nematocerous Diptera have aquatic pupae, some ofwhich are active swimmers (e.g., Culicidae, Dixidae, Chaobori-dae, some Chironomidae) throughout the pupal stage, whereasothers (e.g., most Chironomidae, some Tipulidae) only swim tothe surface for emergence. In taxa in which the pupa is gluedto a substrate (e.g., Blephariceridae, Deuterophlebiidae,Simuliidae), the adult emerges underwater and rises to the sur-face enveloped in a gas bubble. These adults expand theirwings and are capable of flying immediately after reaching thesurface. Species with swimming pupae will emerge using theexuviae as a platform at the water surface. All Chironomidaeemerge in this manner, and the cast skins remain trapped in thesurface film for some time. Collections of exuviae are usefulfor taxonomic purposes and also as a rapid method of samplingthe entire chironomid fauna of a water body (Wartinbee andCoffman 1976; also see Chap. 26).
Respiration in most aquatic Diptera pupae differs fromthat of the larvae in which the terminal abdominal spiracles aregenerally the most important. The pupae have respiratoryhorns or other extensions of the prothoracic spiracles. Depend-ing on the species, these may be used for breathing at the sur-face, in the water column, or even to pierce the tissues of sub-merged plants.
The case-making caddisflies illustrate several morpholog-ical and behavioral adaptations in the pupal stage. The maturelarva selects a protected site, such as under a stone or in acrevice in wood, and attaches the case with silk. Then the caseis shortened, and the ends are closed with mineral or detritalparticles and a silk mesh that allows for water flow. Metamor-phosis occurs within the case. The pupa (pharate adult) isactive within the case, maintaining a flow of water for respira-tory purposes by undulating the body. The pupa has elongatebristles on the labrum and on anal processes that are used forremoving debris from the silk grating at either end of the case.

72 LIFE HISTORY
The back-and-forth movement within the case is effected by thedorsal hook-plates on the abdomen. The pharate adult has largemandibles used for cutting an exit hole at emergence. It thenswims to the surface or crawls to shore where eclosion occurs.In some limnephilid caddisflies, such as the genus Pycnopsy-che, the larvae undergo a prepupal inactive period. For exam-ple, P. guttifer attaches its case to a log or rock and remainssequestered for most of the summer. The digestive tract atro-phies, but the legs remain functional so that under conditions ofunfavorable habitat changes (e.g., water level) the prepupa canmove to a new location.
Sweden indicate that both populations display Type II sur-vivorship curves, with constant mortality (Fig. 5.6). However,the Georgia population has much faster growth and shorter gen-eration time, as well as much greater mortality, indicated by thesteep slope of the survivorship curve. There were also manyother differences between these populations with respect toclutch size, investment in reproduction, larval densities, andterminal weights of larvae and adults. These differences sug-gest that these populations are subject to very different selec-tion pressures and gene flow, and possibly could be considereddifferent species (Otto and Wallace 1989).
SURVIVORSHIP
Mortality among most aquatic insects is usually very high;a very small percentage of eggs deposited reach the adult stage.For example, Willis and Hendricks (1992) found that onlyabout 0.5% of the eggs deposited by a caddisfly, Hydropsycheslossonae, survived to be adults of the ensuing generation.Sources of mortality include both abiotic (e.g., physical distur-bances such as floods and drought) and biotic factors (e.g., pre-dation, disease, parasitism, etc.). Within a given stream reach,downstream drift may also contribute to significant losses (orgains) of individuals through time (e.g., Hershey el al. 1993).
Survivorship curves for several aquatic insects are shownin Figures 5.3-5.6. Several of these include only larval stages(Glossosoma, Brachycentrus, and Tallaperla) and were fit byleast squares regression beginning with peak abundances offirst instar larvae in field samples. These best fit a Type IIexponential model of mortality, representing constant deathsthrough time. However, first instars are generally undersam-pled (e.g., Benke 1984, Cummins and Wilzbach 1988). If theycould be followed in sufficient detail, high mortalities in earlyinstars would shift the curves toward a Type III curve (highermortality in earlier larval stages). For example Willis and Hen-dricks (1992) detected no egg mortality in Hydropsyche slos-sonae, exceptionally high mortality (92.5%) among newlyhatched larvae, due in part to sibling cannibalism (Fig. 5.5).Cummins and Wilzbach (1988) have proposed that pathogenicmortality in the first two instars might be a major cause of mor-tality in many cases. There was constant mortality of H. slos-sonae during larval instars II through V, followed by high mor-tality in the pupal stage. This study indicates that when fol-lowed in sufficient detail, Type in survivorship curves may becommon in aquatic insects. Following larval mortality fromfield samples does not give a true picture of mortality over theentire egg to adult cycle.
Eggs, larvae, pupae, and adults of the water lily leaf bee-tle, Gallerucella nymphaeae, occur on the upper surfaces ofleaves, and this offers a unique situation in that all stages canbe sampled readily. Total number of eggs deposited, larvaereaching each instar, and pupae can be calculated per m2 of leafsurface area (Otto and Wallace 1989). Comparisons of popula-tions of G. nymphaeae from Georgia (U.S.A.) and southern
CONCLUDING COMMENTS
The critical importance of systematics to basic life-historystudies has been emphasized on numerous occasions (e.g.,Wiggins 1966; Ross 1967a; Waters 1979a, 1979b). Excellentprogress has been made for several aquatic insect groups inNorth America (e.g., Edmunds et al. 1976; Wiggins 1977;Stewart and Stark 1988) as well as more general works onimmatures (e.g., Stehr 1987, 1991); however, eggs and allinstars of immatures cannot always be identified. Studies ofaquatic insect life histories require that individual investigatorsdevelop reliable methodologies for separating early instars ofclosely related species (e.g., Mackay 1978). It is unfortunatethat life histories of so many aquatic insect species remainunknown. In some quarters, the mistaken impression still per-sists that such efforts are unfashionable and of little value.Ecosystem-level studies are often directed to studies of the pro-cessing of organic matter by various groups of animals, andinsects are frequently the most abundant group considered.The integration of production, feeding habit, and bioenergeticdata can yield a much better understanding of the role of indi-vidual species in ecosystems. Benke (1979), Benke et al.(1979), and Waters (1979a,b) have emphasized that a knowl-edge of basic life histories is mandatory for reasonable esti-mates of production. Waters has pointed out that voltinism andlength of aquatic life (=CPI, or Cohort Production Interval[Benke 1979]) are two of the most important life history fea-tures influencing secondary production estimates. The "once-per-month" sampling program of most studies is not adequatefor many estimates of aquatic insect secondary production, andsampling schedules need to be tailored to the life histories ofthe organisms being studied (Cummins 1975; Waters 1979a).
Both temperature and food may influence life-history pat-terns; however, data on the potential combined effects of tem-perature and food quality on life histories (e.g., Anderson andCummins 1979) of various groups of aquatic insects are toomeager for any broad generalizations as yet. All students areencouraged to develop an appreciation for the importance ofsystematics, life histories, secondary production, and bioener-getics as interconnecting links toward the basic understandingof the structure and function of aquatic communities.
I•*.

LIFE HISTORY 73
5.3LI
Glossosoma nigrior
(summer cohort larvae)
5.4 Brachycentrus spinae —L ' Tallaperla mafia —
50 100 150Time in days from hatching
1 o.. 5.5 Hydropsyche slossonae
200 100 200 300 400 500Time in days from hatching
10..,5-6 Gallerucella nymphaeae
9-
600
Sweden (*)
Georgia (A)
100 200 300 400Time in days from opposition
10 20 30 40Time in days from oviposition
50
Figure 5.3. Survivorship curves for the summer cohort ofthe caddisfly Glossosoma nigrior (larval instars I [L I]through V [L V] based on instream sampling) (data fromGeorgian and Wallace [1983]).Figure 5.4. Survivorship curves for larval stages of thecaddisfly Brachycentrus spinae (larval instars I [L I]through V [L V]) (data from Ross and Wallace [1981]),and the stonefly Tallaperla maria larval instars I (L I)through final instar (L F) (data from O'Hop et al. [1984]).
Figure 5.5. Survivorship curve for the caddisflyHydropsyche slossonae based on egg, larval, pupal, andadult stages. Note the high mortality during the first lar-val instar (L I), followed by little mortality in later larvalstages (L II - L V), and high mortality in pupal to adultstages (data from Willis and Hendricks 1992).Figure 5.6. Survivorship curve for egg (E), three larvalinstars (L I - L III), and pupal stage (P) of the chrysomelidbeetle, Gallerucella nymphaeae, from Georgia (U.S.A.)and Sweden. Note the much shorter life cycle andgreater mortality in the Georgia population (data fromOtto and Wallace 1989). The equations for the lines areas follows: Georgia, y = 10.1 - 0.23x, r2 = 0.98; and, Swe-den, y = 8.4 - 0.07x, r2 = 0.95.





























