Embed Size (px)
Citation preview

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD10.1146/annurev.cellbio.18.012502.105840
Annu. Rev. Cell Dev. Biol. 2002. 18:637–706doi: 10.1146/annurev.cellbio.18.012502.105840
Copyright c© 2002 by Annual Reviews. All rights reservedFirst published online as a Review in Advance on July 24, 2002
STRIATED MUSCLE CYTOARCHITECTURE:An Intricate Web of Form and Function
Kathleen A. Clark,3,4 Abigail S. McElhinny,1,4
Mary C. Beckerle,3 and Carol C. Gregorio1,21Departments of Cell Biology and Anatomy and2Molecular and Cellular Biology,University of Arizona, Tucson, Arizona 85724;3Huntsman Cancer Institute andDepartment of Biology, University of Utah, Salt Lake City, Utah 84112;e-mail: [email protected], K. A. Clark and A. S. McElhinny contributed equally to this work
Key Words sarcomere, cytoskeleton, Z-line, thick filament, thin filament
■ Abstract Striated muscle is an intricate, efficient, and precise machine that con-tains complex interconnected cytoskeletal networks critical for its contractile activity.The individual units of the sarcomere, the basic contractile unit of myofibrils, includethe thin, thick, titin, and nebulin filaments. These filament systems have been investi-gated intensely for some time, but the details of their functions, as well as how they areconnected to other cytoskeletal elements, are just beginning to be elucidated. Theseinvestigations have advanced significantly in recent years through the identification ofnovel sarcomeric and sarcomeric-associated proteins and their subsequent functionalanalyses in model systems. Mutations in these cytoskeletal components account for alarge percentage of human myopathies, and thus insight into the normal functions ofthese proteins has provided a much needed mechanistic understanding of these disor-ders. In this review, we highlight the components of striated muscle cytoarchitecturewith respect to their interactions, dynamics, links to signaling pathways, and functions.The exciting conclusion is that the striated muscle cytoskeleton, an exquisitely tuned,dynamic molecular machine, is capable of responding to subtle changes in cellularphysiology.
CONTENTS
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 638THE SARCOMERE: BASIC CONTRACTILEUNIT OF MYOFIBRILS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 642
THIN FILAMENTS AND I-BAND COMPONENTS . . . . . . . . . . . . . . . . . . . . . . . . . 643Actin: The Most Abundant Protein in Striated Muscle. . . . . . . . . . . . . . . . . . . . . . . 643Tropomyosin: The Regulatory Switch of the Thin Filaments. . . . . . . . . . . . . . . . . . 645Troponin C, I, and T (TnC, I, T): The MolecularLatch that Controls the TM Switch. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 646
CapZ and Tropomodulin: Thin Filament Capping Proteins. . . . . . . . . . . . . . . . . . . 648THICK FILAMENTS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 649
1081-0706/02/1115-0637$14.00 637

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
638 CLARK ET AL.
Myosin: Molecular Motor for Muscle Contraction. . . . . . . . . . . . . . . . . . . . . . . . . . 650Thick Filament-Associated Proteins. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 651
THE M-LINE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 654Myomesin and M-protein: Potential Filament-Linking Molecules. . . . . . . . . . . . . . 654MURF-1: A Potential Link Between SarcomereDegradation and Gene Expression. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 655
Muscle-Specific Calpain3/p94. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 656MM-Creatine Kinase. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 656
TITIN: THE IMMENSE, MULTIFUNCTIONALTHIRD FILAMENT SYSTEM . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 657Z-Disc Titin: A Key Contributor to Sarcomere Integrity. . . . . . . . . . . . . . . . . . . . . 657I-Band Titin: A Molecular Spring. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 658A-Band Titin: A Thick Filament Template?. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 660M-Line Titin: A Potential Signal Transducer. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 660Titin Homologues: Model Systems ProvideInsight into Titin Function. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 661
NEBULIN: A RULER FOR THIN FILAMENT ASSEMBLY? . . . . . . . . . . . . . . . . . 661THE Z-LINE: CYTOSKELETAL ANCHOR AND SIGNALTRANSDUCTION CENTER . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 663α-Actinin: Primary Cross-Linker of the Z-line. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 664α-Actinin-Associated Proteins. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 665Other Z-Line-Associated Proteins. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 668
INTERMEDIATE FILAMENT PROTEINS: LINKERS OFCYTOSKELETAL NETWORKS IN STRIATED MUSCLE. . . . . . . . . . . . . . . . . . . 670
MICROTUBULES: EMERGING ROLES IN STRIATEDMUSCLE MORPHOLOGY AND FUNCTION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 672
MEMBRANE LINKAGES TO THE CYTOSKELETON:ANCHORAGE, FORCE TRANSMISSION, ANDMECHANOSIGNAL TRANSDUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 673Costameres: Integrins, Spectrin, and the Dystroglycan Complex. . . . . . . . . . . . . . . 674The Myotendinous Junction: Site of ForceTransduction in Skeletal Muscle. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 677
Intercalated Discs: Terminal Anchors in Cardiac Muscle. . . . . . . . . . . . . . . . . . . . . 678Desmosomes: Intermediate Filaments and AssociatedProteins Form Links to the Membrane. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 679
Ankyrins: Organizer of Specialized Membrane Domains. . . . . . . . . . . . . . . . . . . . . 680COMMUNICATION BETWEEN THE SARCOMEREAND THE NUCLEUS: IMPLICATIONS FOR GENEREGULATION BY CYTOSKELETAL COMPONENTS . . . . . . . . . . . . . . . . . . . . . 681
CONCLUSIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 683
INTRODUCTION
Striated muscle contraction is a dramatic example of cell motility that is accom-plished through the workings of an interwoven network of specialized cytoskeletalarrays. This precise networking is critical for converting the molecular interactionsproduced by actin and myosin in each sarcomere into the efficient macroscopicmotion of contraction. Amazingly, although the contractile apparatus must be

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 639
maintained with almost crystalline order for its efficient function, it is not a passive,static framework. Instead, the components are in a requisite dynamic equilibriumwith constant coordinated alterations in protein synthesis, degradation, assembly,and maintenance. A striking example of this is human cardiac muscle, where thedynamic process of synthesizing and replacing contractile proteins occurs evenwhile force production is maintained at rates of 60–200 beats per min! It is clearthat the muscle cytoskeleton is under tight regulation and must have precise con-nections to numerous gene expression and signaling pathways.
Within the past decade, striated muscle has become a powerful model system forstudying cytoskeletal protein interactions and functions. In particular, it has beenvaluable for investigating direct linkages between cytoskeletal components andgene expression, the coordinate roles of actin filaments, intermediate filaments, andmicrotubules in allowing for efficient contractile activity, as well as myofibril at-tachment to the plasma membrane (sarcolemma), including links to ion channelsand extracellular matrix components. This has occurred through exciting advancesin imaging technology, molecular and genetic techniques including yeast-two hy-brid screens, gene expression profiling, and gene transfer techniques, as well asnovel model systems. These factors have been key for determining the functionalroles of previously identified myofibrillar components and for the continuing dis-covery of several novel proteins that are associated with the contractile apparatus.
Currently, a tremendous effort is focused on dissecting the pathogenesis of sev-eral striated muscle myopathies that result directly from mutations in contractileand associated proteins, highlighting their importance in normal muscle structureand activity. Deciphering the precise relationships among striated muscle com-ponents often reveals candidate molecules for myopathies (the genetic lesionsfor which had not been identified). To date, approximately 100 mutations havebeen identified in nine genes that all encode highly abundant sarcomeric compo-nents that result in familial hypertrophic cardiomyopathy (familial HCM or FHC).Because of the nature of these mutations, familial HCM is known as a “diseaseof the sarcomere” (Thierfelder et al. 1994). In contrast to familial HCM, thereis heterogeneity in the classes of proteins that are mutated in familial dilatedcardiomyopathy (DCM), which include membrane cytoskeletal proteins. Severalskeletal muscle myopathies such as nemaline myopathy and certain forms of mus-cular dystrophy (MD), including Limb Girdle, Duchenne, and Beckers MD, resultfrom mutations in sarcomeric or sarcolemmal components. How each mutationleads to the diverse clinical manifestations characteristic of these striated musclemyopathies remains obscure. However, efforts to understand both normal and myo-pathic muscle physiology have benefited greatly from the use of model systemssuch as cultured chick, rat, and mouse cardiac and skeletal myocytes, and geneticsystems that include rodents, rabbit,Caenorhabditis elegans, Drosophila, axolotl,and zebrafish. (For reviews on human myopathies, see Chien 2000, Gordon &Hoffman 2001, Marian & Roberts 2001, Tubridy et al. 2001, Seidman & Seidman2001, Towbin & Bates 2002) (Table 1).
In this review, we focus on the sarcomeric components (many of which areexpressed in a variety of other cell types), their functions, and how they are linked

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
640 CLARK ET AL.
TABLE 1 Vertebrate phenotypes and human myopathies associated with mutations and/orperturbed levels of sarcomeric and sarcomeric-associated proteins
Phenotypes in vertebratemodel systems Human myopathies
Thin filament/I band proteinsActin Lethality (cardiacα-actin: Kumar Actin and nemaline myopathies
et al. 1997) (skeletal muscleα-actin: Nowak et al.1999, Ilkovski et al. 2001);
dilated cardiomyopathy (cardiacα-actin: Olson et al. 1998);
hypertrophic cardiomyopathy (cardiacα-actin: Olson et al. 2000)
Tropomyosin Cardiac hypertrophy and lethality Hypertrophic cardiomyopathy(mouse) (α-TM: Prabhakar et al. 2001); (α-TM: Michele & Metzger 2000);
nemaline myopathy (mouse) nemaline myopathy (α-TM, β-TM:(slowα-TM: Corbett et al. 2001) Donner et al. 2002, Michele &
Metzger 2000);dilated cardiomyopathy (α-TM: Olsonet al. 2001)
Troponins Cardiomyopathies, myocyte disarray, Hypertrophic cardiomyopathy (cardiacpremature death (mouse) (TnT, TnI: TnT, TnI, TnC: Hoffman et al. 2001,Maass & Leinwand 2000, Dalloz Marian & Roberts 2001, Seidman &et al. 2001) Seidman 2001)
Tropomodulin Dilated cardiomyopathy (mouse) Unknown(E-Tmod: Sussman et al. 1998b)
CARP Levels of CARP↑ in cardiac Unknownhypertrophy (rat) (Aihara et al 2000)
Thick filamentand M linecomponentsMyosin Cardiomyopathies, embryonic lethality, Hypertrophic cardiomyopathy
myocyte disarray, sudden cardiac (MyHC, MLC: Marian & Robertsdeath, compromised exercise capacity 2001, Seidman & Seidman 2001)(mouse) (α andβ-MyHC);
myocyte disarray, cardiac hypertrophy(rabbit) (β-MyHC) (Maass & Leinwand2000, Dalloz et al. 2001);
skeletal muscle hypertrophy/growth andmuscled defects (mouse) (fast skeletalMyHC: Acakpo-Satchivi et al. 1997,Allen et al. 2001);
lethality (mouse) (α-MyHC: Joneset al. 1996)
MyBP-C Dilated cardiomyopathy (mouse) Hypertrophic cardiomyopathy(McConnell et al. 1999); (Marian & Roberts 2001,
hypertrophic cardiomyopathy and Seidman & Seidman 2001)sudden death,↓ exercise capacity(mouse) (Yang et al. 2001, Harriset al. 2002)
(Continued)

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 641
TABLE 1 (Continued)
Phenotypes in vertebratemodel systems Human myopathies
AMP-deaminase Unknown Skeletal muscle dysfunction andweakness (Morisaki et al. 1992)
P94/Calpain 3 Decreased grip strength and progressive Limb girdle muscular dystrophymuscular dystrophy (mouse) (Tagawa type 2A (Richard et al. 1995)et al. 2000, Richard et al. 2000)
Third and fourthfilament systemsTitin Cardiac dysfunction (zebrafish) (Xu Dilated cardiomyopathy (Gerull
et al. 2002); et al. 2002, Itoh-Satoh et al. 2002)muscular dystrophy with myositis(mouse) (Garvey et al. 2002)
Nebulin Unknown Nemaline myopathy (Pelin et al. 1999,Gurgel-Giannetti et al. 2001)
Nebulette Unknown Nonfamilial idiopathic dilatedcardiomyopathy (Arimura et al. 2000)
Z-line proteinsCRP3/MLP Dilated cardiomyopathy (mouse) Levels↓ in failing human myocardium
(Arber et al. 1997) (Zolk et al. 2000)ALP Right ventricular cardiomyopathy Unknown
(mouse) (Pashmforoush et al. 2001)Cypher Striated muscle failure (mouse) Unknown
(Zhou et al. 2001)Myotilin Unknown Limb girdle muscular dystrophy
type 1A (Hauser et al. 2000)Telethonin Unknown Limb girdle muscular dystrophy
type 2G (Moreira et al. 2000)Intermediatefilament proteinsDesmin Muscle degeneration and cardiomyopathy Desmin myopathy with corresponding
(mouse) (Li et al. 1997, Milner et al. cardiac and skeletal myopathies1999, Balogh et al. 2002) (Goebel 1995)
Emerin and Emery-Dreifuss muscular dystrophy Emery-Dreifuss (X-linked and ADLamins A and C (mouse) (Sullivan et al. 1999); forms) and limb girdle muscular
dilated cardiomyopathy (mouse) dystrophy type 1B;(Raharjo et al. 2001) dilated cardiomyopathy; lipodystrophy
(Emery 2000, Morris 2001)
MicrotubulesUnknown Aberrant levels and dynamics of
microtubules implicated in certaincardiomyopathies (Hein et al. 2000);
colchicine skeletal musclemyopathy (Fernandez et al. 2002)
Membrane-associatedproteinsIntegrins Muscular dystrophy (mouse) Congenital myopathy (Hayashi
(α7: Mayer et al. 1997) et al. 1998)
(Continued)

23 Sep 2002 10:29 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
642 CLARK ET AL.
TABLE 1 (Continued)
Phenotypes in vertebratemodel systems Human myopathies
Dystrophin Muscular dystrophy (mouse) Duchenne’s and Becker’s muscular(Sicinski et al. 1989) dystrophy (Hoffman et al. 1987);
X-linked dilated cardiomyopathy(Muntoni et al. 1993)
α, β, δ, andγ Muscular dystrophy (mouse) (δ: Duclos Limb girdle muscular dystrophiessarcoglycans et al. 1998;γ : Hack et al. 1998;β: Araishi (Gordon & Hoffman 2001)
et al. 1999;δ: Coral-Vazquez et al. 1999);Muscular dystrophy, dilated or hypertrophiccardiomyopathy (hamster) (δ: Nigro et al.1996, Sakamoto et al. 1997, Straub et al. 1998)
Plakoglobin/ Cardiac muscle rupture; lethality (mouse) Naxos disease (Norgett et al. 2000)Desmoplakin (Ruiz et al. 1996, Gallicano et al. 2001)
Plectin Lethality; epidermolysis bullosa simple EBS-MD (Gache et al. 1996)muscular dystrophy (EBS-MD) (mouse)(Andra et al. 1997)
to the distinct cytoskeletal assemblies, organelles and signaling components thatexist in striated muscle cells. Additionally, we discuss the roles of various myo-fibrillar protein isoforms in generating functional diversity in contractile prop-erties and human myopathies that result from perturbation of these cytoskeletalnetworks.
THE SARCOMERE: BASIC CONTRACTILEUNIT OF MYOFIBRILS
The characteristic striated appearance of myofibrils is readily observable by lightmicroscopy as alternating light and dark bands. This unique feature is a directresult of the precise alignment of the filament systems of the sarcomere, the ba-sic contractile unit of myofibrils. The light band is termed the I-band because itis isotropic in polarized light; the dark band is known as the A-band because itis anisotropic. The principle components of striated muscle sarcomeres includeparallel arrays of actin-containing thin filaments that span the I-band and overlapwith myosin-containing thick filaments in the A-band. The third filament sys-tem is made up of single molecules of titin (the largest vertebrate protein identi-fied to date), which span half sarcomeres. Another giant protein, nebulin, spansthe length of the actin filaments and forms the fourth filament system in skele-tal muscle. The Z-discs represent the lateral bounderies of the sarcomere wherethe thin, titin, and nebulin filaments are anchored. Z-discs also are implicatedin mechanosensation and signaling to the nucleus, which contribute to mainte-nance of muscle homeostasis. The key elements of the sarcomere are illustrated inFigure 1.

23 Sep 2002 10:29 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 643
THIN FILAMENTS AND I-BAND COMPONENTS
The actin (thin) filaments are anchored in the Z-disc, span the I-band, and extendtoward the middle of the sarcomere. In the A-band they interdigitate with the thickfilaments. Single molecules of the giant skeletal muscle protein nebulin, as well aspolymers of tropomyosin and the troponin complex, are present along the lengthof the actin filaments. The filaments are capped at the pointed and barbed endsby tropomodulin (Tmod) and CapZ, respectively. (Pointed and barbed ends referto the pattern observed by electron microscopy when polarized actin filamentsare decorated with myosin S1 fragments.) The I-band region of the sarcomerealso contains a portion of the vast, modular protein titin. I-band titin containselastic elements that act as molecular springs to maintain sarcomeric integrity.The size and elastic properties of I-band titin are dictated by alternative splicingof a single immense gene, which in turn contributes to the degree of musclestiffness. Thus the I-band region of the sarcomere has several essential functions.These include linking the region of active force generation, the A-band, withthe bordering Z-lines, as well as containing structures to act as springs for thereversible, mechanical stretch response required for efficient contractile activity(Figure 2).
Actin: The Most Abundant Protein in Striated Muscle
The ubiquitous actin molecule, implicated in diverse cellular functions such asmotility, cytokinesis, and contraction, is the main component of the thin fila-ments. Actin filaments form two twistedα helices that associate with the regu-latory proteins tropomyosin and the troponins. Early in myofibril assembly, thinfilament components associate with nascent Z lines to form the first identifiablestructures, I-Z-I complexes (Holtzer et al. 1997). The∼1 µm lengths of the ma-ture thin filaments in striated muscle are remarkably precise and are proposedto occur using several mechanisms including interactions with template proteins(e.g., nebulin), capping proteins (tropomodulin and CapZ), and other moleculesthat sequester monomeric actin, sever actin filaments, and/or otherwise promoteactin polymerization and depolymerization. For reviews on thin filament dynam-ics and length regulation see Littlefield & Fowler (1998) and Cooper & Schafer(2000).
The atomic structure of G-actin was obtained over a decade ago, generating amore complete structural model of the thin filaments (Kabsch et al. 1990; reviewedin Steinmetz et al. 1997). Actin monomers have four domains: two main domains ofthe molecule are further divided into two regions. A cleft between the two domainscontains the nucleotide-binding site and a divalent ion binding site, likely for Mg2+.Each half-helical turn of the thin filament is comprised of 7 actin monomers, whichinteract via their subdomains 3 and 4. Subdomain 1 binds to the myosin heads.Each myosin head has ATPase activity, which is activated upon its interactionwith actin. The motor activity of the myosin heads moves the thin filaments past

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
644 CLARK ET AL.
the thick filaments to generate force, resulting in muscle contraction (for reviewsee Huxley 2000). The actin-myosin interaction is tightly controlled in a Ca2+-dependent manner by the regulatory complex composed of tropomyosin and thetroponins (Weber & Murray 1973). Actin is terminally acetylated and also can bephosphorylated and methylated; these modifications influence actin dynamics andfunctional properties (Ampe & Vandekerckhove 1999). For example, mutations ina Drosophilaactin (ACT88F) that result in a loss of actin N-acetylation perturbflight muscle function, perhaps owing to disrupted interactions between myosinand the mutant actin (Schmitz et al. 2000).
Several actin isoforms exist, but their sequences and molecular structures areamazingly similar. The actin isoforms often are classified by their isoelectric pointsas α, β, andγ . Mammals and birds have six known actins, each encoded byseparate genes whose expression patterns are regulated developmentally and in atissue-specific manner. Two striated muscle-specific isoforms, cardiac and skeletalactins, are co-expressed at varying levels depending on the species, muscle fiber,and developmental stage. Vascular and visceral (enteric) actins are expressed insmooth muscle and also in striated muscle fibers transiently during development.Remarkably, the four actin isoforms found in muscle vary at only 10 of their375 amino acids (Vandekerckhove & Weber 1979)! Two non-muscle actins, thecytoplasmic isoforms, are co-expressed with other actin isoforms in many tissues.It is striking that highly similar actin isoforms are functionally non-equivalent(reviewed in Khaitlina 2001). For example, ectopic expression of humanβ-actinin Drosophilaindirect flight muscle (IFM), which differs from ACT88F actin byonly 15 amino acids, perturbed sarcomeric organization and flight ability (Braultet al. 1999). Furthermore, exogenous expression of non-muscle actin isoformsin rat cardiomyocytes induced gross alterations in morphology and contractileactivity (von Arx et al. 1995). The majority ofα-cardiac actin-null mice die soonafter birth, and although they can be rescued by expression of smooth muscleγ -actin, their hearts exhibit severe contractile dysfunction and hypertrophy (Kumaret al. 1997). These studies indicate that tight cellular control of actin isoformsexpression is critical for normal muscle structure and function.
Clinical investigations have revealed that specific mutations in striated muscleactin isoforms result in various forms of human muscle myopathies. For example,more than 20 different missense mutations in the skeletal muscleα-actin gene(ACTA1) are associated with two muscle diseases: actin myopathy and nemalinemyopathy, both of which are characterized by myofibrillar structural abnormali-ties and muscle weakness (Nowak et al. 1999, Ilkovski et al. 2001). Interestingly,missense mutations in regions of actin involved in attachment to Z-lines and inter-calated discs lead to familial DCM and heart failure, likely because of impairedforce transmission. On the other hand, missense mutations in regions of cardiacactin involved in its interactions with actin or myosin result in HCM (Olson et al.1998, 2000). Thus single amino acid substitutions in the actin molecule resultin distinct clinical manifestations, depending on the particular functional domainaffected.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 645
Tropomyosin: The Regulatory Switch of the Thin Filaments
Tropomyosin (TM) (Mr∼ 37 kDa) is constructed of twoα-helical chains arrangedas a coiled-coil rod that associate with actin filaments in virtually all eukaryoticcells. To accommodate the wide range of functions associated with actin filaments,TM exists as a large number of isoforms. In humans and rats, the TM family con-tains four separate genes,TPM1,2,3, and4, each encoding one to nine alternativelyspliced isoforms. In striated muscle cells, all TMs are the long form of 284 residuesand includeα-TM, encoded by TPM1;β-TM, encoded by TPM2; and the slowtwitch fiberα-TM, encoded by TPM3 (Perry 2001). TM forms homodimers orheterodimers to generate parallel coils that extend the length of actin filaments. Instriated muscle, all expressed TM dimers span 7 actin monomers. The binding ofTM along each of the two grooves of the thin filament is highly cooperative andoccurs in a head-to-tail manner, with an overlap of 8 to 11 amino acids (Wegner1979). In striated muscle, theα- andβ-forms of TM are∼87% identical, buttheir expression ratios differ based on muscle fiber type and stage of develop-ment. Perturbation of the expression ratios of mouse TM isoforms induces severecardiac abnormalities including contractile dysfunction (Muthuchamy et al. 1998,MacGowan et al. 2001). Recently, insights into how tropomyosin bends aroundactin in thin filaments (verifying predictions previously based on sequence analy-sis) were provided by the determination of the empirical structure of TM and thecrystal structure of an 81-residue fragment consisting of approximately one thirdof the molecule (Whitby & Phillips 2000, Brown et al. 2001).
A primary function of TM is to work together with the troponins in regulatingthe interaction of the thin and thick filaments. This complex regulatory mecha-nism involves a Ca2+- and troponin-mediated conformational shift in the positionof TM on the thin filaments (Parry & Squire 1973, Craig & Lehman 2001). In onewell-supported model, it is predicted that in relaxed muscle, TM blocks myosinhead-binding sites on the outer domain of the thin filaments. Upon Ca2+ bind-ing to troponin C, TM shifts its position (likely more than once) to expose theweak myosin-binding sites on actin. Myosin binding activates the myosin ATPaseand causes further translocation of TM on actin filaments, allowing strong, force-generating interactions between myosin and actin. Intriguingly, the positions ofTM on the thin filaments differ slightly depending on the TM and actin isoformsexpressed (e.g., Chandy et al. 1999, Lehman et al. 2000). Therefore, subtle vari-ations in the TM switch may contribute to the distinct contractile properties ofdifferent striated muscle types.
In addition to its critical regulatory role in contraction, TM also has a functionin stabilizing the thin filaments. Binding of TM to actin increases the stiffnessof the thin filaments and inhibits their fragmentation (Wegner 1982). TM alsostabilizes thin filaments by slowing depolymerization and polymerization at theirpointed ends (Weigt et al. 1990, Broschat 1990) where it interacts with the cappingprotein, tropomodulin. Together, tropomodulin and TM appear to play a pivotalrole in regulating thin filament lengths and stability.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
646 CLARK ET AL.
A myriad of genetic and clinical findings illustrate the importance of TM forkey physiological processes. For example, nematodes deficient in muscle TM iso-forms die or exhibit developmental and morphological abnormalities (Anyanfulet al. 2001). Likewise,α-TM null mice die during gestation (Rethinasamy et al.1998). Mexican axolotl mutants with reduced TM expression do not form cardiacmyofibrils (Zajdel et al. 1998), and flies with mutant IFM TM have abnormalmuscle fiber structure with severely perturbed power production and flight ability(Kreuz et al. 1996). Various mutations inα-TM cause severe cardiac hypertrophyand death in mice, whereas at least four mutations in the human TPMI gene areassociated with∼5% of human familial HCM cases (reviewed in Hernandez et al.2001). An interesting feature of humanα-TM-associated HCM is that althoughthe mutant TM protein is expressed in skeletal muscle, it does not produce a notedskeletal muscle myopathy. In contrast, a mutation in the humanTPM3gene, whichshares 90% sequence identity with TPM1, is associated with nemaline myopathy(Laing et al. 1995). Additionally, mutations that alter the surface charge ofα-TMare associated with DCM (Olson et al. 2001). Recent evidence suggests that thedistinct clinical presentations of the cardiac and skeletal muscle myopathies asso-ciated with different mutations inα-TM result from differential cellular changesin the ability of the mutant TM proteins to regulate muscle contraction and relax-ation in response to changing Ca2+ concentrations (Michelle et al. 2002). Overall,it is clear that proper TM expression is critical for development and muscle ul-trastructure and is an essential component of the regulatory machinery of musclecontraction.
Troponin C, I, and T (TnC, I, T): The MolecularLatch that Controls the TM Switch
Tns are a cooperative complex of three proteins (TnC, TnI, and TnT) that functionwith TM in modulating the interaction of myosin and actin during force generation.The Tn subunits are expressed as a number of isoforms whose expression patternsdiffer during development and among fiber types, and they contribute to the distinctcontractile properties of striated muscle (Westfall & Metzger 2001).
TnC (Mr∼ 18 kDa), the only Tn whose detailed structure is known, is a calmod-ulin-related protein containing EF-hand domains at each termini (Herzberg &James 1988). Current models indicate that binding of Ca2+ to the N-terminal reg-ulatory domain of TnC alters its interaction with TnI, which allosterically trans-mits the activation signal further along the thin filament regulatory complex (e.g.,McKay et al. 1997).
TnI (Mr∼ 20 kDa), the inhibitory subunit of the Tn complex, completely in-hibits actomyosin ATPase activity in vitro (Leavis & Gergely 1984); however, theCa2+-dependent regulation of this process in vivo requires the other Tns and TM.Remarkably, TnI changes affinity for its multiple binding partners based on thebinding of Ca2+ to TnC. This long-proposed idea that TnI functions as the molec-ular latch of the troponin complex is supported by structural data (Lehman et al.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 647
2001). In the off-state, TnI’s N-terminal region is bound to TnC and TnT, andits C-terminal region is tightly bound to actin. Upon Ca2+ binding to TnC duringthe on-state, TnI’s affinity for actin is reduced whereas its affinity for TnC andTnT is enhanced (e.g., Potter & Gergely 1974, Syska et al. 1976). Other molecularmechanisms also affect TnI conformations and binding affinities, which are key forregulating thin filament activation. For example, developmental events in cardiacmuscle such as the transition from slow skeletal to cardiac TnI expression, as wellas TnI’s phosphorylation byβ-adrenergic-activated protein kinase A (PKA), affectcardiac performance (Solaro & Van Eyk 1996, Perry 2001, Westfall & Metzger2001). Strikingly, loss of TnI function inC. eleganscauses aberrant muscle trem-bling and tearing, likely a consequence of an abnormal duration of force (McArdleet al. 1998).
The exact function of TnT (Mr∼ 30 kDa) is somewhat controversial. TnT isthought to anchor TnC and TnI dimers in the thin filament (e.g., Hitchcock 1975),thus functioning as the molecular organizer of the thin filament regulatory com-plex. TnT also is proposed to mediate the Ca2+ sensitivity of the actomyosin AT-Pase, as well as its activation and/or force development (Hernandez et al. 2001).A single TnT gene encodes several isoforms, including cardiac TnT, which con-tains multiple phosphorylation sites (Jin et al. 1992). The extended N terminusof TnT interacts with actin and with the overlapping regions of adjacent TMmolecules, whereas its highly conserved C-terminal region interacts with TnC andTnI (Stefancsik et al. 1998). It is likely that upon the transmission of the Ca2+
signal through the Tn complex, the interaction between TnT and TM weakens,allowing TM to shift and expose the myosin-binding sites on the thin filaments(Potter et al. 1995).
The influence of Tns on striated muscle contractility has received increasingattention due to a staggering amount of clinical evidence correlating Tn mutationswith changes in cardiac function. For example, several pathophysiological con-ditions known to compromise heart function are linked to TnI. In fact, release ofcTnI from the heart is used widely as a primary indicator of myocardial ischemia(Chapelle 1999). Ischemia and other cardiovascular conditions also are associatedwith alterations in the phosphorylation state of TnI (Zakhary et al. 1999). More-over, work in transgenic animals has shown that removal of 17 amino acids fromthe C-terminal end of cTnI is sufficient to produce myocardial stunning (an eventresulting from reduced perfusion) (Murphy et al. 2000). Interestingly, the thirdmost common gene responsible for human HCM is cTnT. Numerous cTnT mu-tations are associated with a particularly severe form of the disease characterizedby high incidence of sudden death despite only mild left ventricular hypertrophy(Watkins et al. 1995). Many of these cTnT mutations severely impair the regu-latory functions of the other troponins (Takahashi-Yanaga et al. 2001). Variousmissense and trucation mutations in the cardiac isoforms of TnI also are responsi-ble for many types of cardiomyopathy in rodents and humans. Recently, the firstmutation in human cardiac TnC associated with HCM was reported (Hoffman et al.2001). Together, mutations in Tns make up∼25% of total human HCM cases (e.g.,

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
648 CLARK ET AL.
Kimura et al. 1997, Sweeney et al. 1998, Tardiff et al. 1999). In conclusion, there isa complicated, yet extraordinary, picture emerging as to how the troponin subunitsinteract with each other and with the thin filaments to regulate contractile activity.
CapZ and Tropomodulin: Thin Filament Capping Proteins
The regulation of actin dynamics at each end of the thin filament is a key factorin their assembly, as well as in maintaining their uniform lengths. Proteins thatspecifically bind to and cap the thin filaments block filament elongation and short-ening. CapZ is a widely distributed, highly conserved barbed end capping proteininvolved in nucleation and stabilization of actin filaments. CapZ is a heterodimercontainingα (Mr∼ 36 kDa) andβ (Mr∼ 32 kDa) subunits, both of which are re-quired for capping. There are at least twoα and twoβ isoforms; theα isoformsbind differentially to actin (Casella & Torres 1994, Hug et al. 1992). Thus physi-ological differences in CapZ heterodimer subunits may contribute to variations inthin filament properties in different muscle types.
In striated muscle, CapZ localizes to Z-lines where it binds toα-actinin, likelyforming an anchoring complex for the thin filaments (Casella et al. 1987, Papa et al.1999). In support of this hypothesis, CapZ assembles early, before the thin filamentsattain their mature striated pattern, and inhibition of its capping activity in skeletalmyogenic cells delays the organization of the Z-lines and thin filaments (Schaferet al. 1995). These studies support the idea that CapZ organizes and aligns thebarbed ends of the thin filaments. Phosphatidylinositol 4,5-bisphosphate rapidlydissociates CapZ from the barbed ends of actin filaments (Schafer et al. 1996)and thus may play a role in regulating actin dynamics. CapZ also binds to S100,a Ca2+-binding protein in vertebrate muscle (Kilby et al. 1997); the functionalimplications of this interaction are unknown.
Tropomodulin (Tmod) (Mr∼ 40 kDa) caps the pointed ends of the thin filaments.Two Tmod isoforms have been identified in vertebrate muscle to date: erythrocyteTmod (E-Tmod:TMOD1) and skeletal Tmod (Sk-Tmod:TMOD4) (Fowler 1987,Almenar-Queralt et al. 1999, Cox & Zoghbi 2000). The Tmod isoforms are∼60%identical and are encoded by separate genes. The mechanisms that regulate theirdistinct temporal and spatial expression patterns remain to be to be determined (Chuet al. 2000, Cox et al. 2001). The N-terminal half of E-Tmod binds tropomyosin,and its primary actin-binding site resides within its C-terminal region (Babcock &Fowler 1994, Fowler & Conley 1999). When actin filaments are assembled in thepresence of tropomyosin, E-Tmod affinity for the pointed ends is greatly enhanced.Therefore, the full capping activity of E-Tmod is dependent on its interactions withboth tropomyosin and actin (Weber et al. 1994). The fact that E-Tmod contains twofunctionally distinct domains is highlighted by recent structural studies revealingthat its N-terminal half is elongated and highly flexible, whereas its C-terminal halfis globular. E-Tmod’s C-terminal half contains a tandem of leucine-rich repeats(LRR), a unique feature for actin-binding proteins (Kostyukova et al. 2000).
Chicken cardiac, slow-twitch skeletal and embryonic fast-twitch skeletal mus-cle fibers are enriched in E-Tmod, whereas adult fast-twitch skeletal muscle fibers

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 649
contain Sk-Tmod. In muscle fibers that co-express both isoforms, Sk-Tmod ispreferentially localized to thin filament pointed ends, and E-Tmod is localized tocostameres. Although the isoforms appear to have the same actin-capping and bind-ing properties to muscle isoforms of TM in vitro (Almenar-Queralt et al. 1999), iso-forms of E-Tmod have a higher affinity for nonmuscle TMs compared with muscleTMs in vitro; this maybe one mechanism responsible for its differential targeting(Greenfield & Fowler 2002). Additionally, the affinities for E-Tmod and Sk-Tmodfor the N-terminal end of nebulin are different: Sk-Tmod binds more tightly tonebulin modules M1-M2-M3 than E-Tmod does in vitro (McElhinny et al. 2001).This finding may partly explain the mechanistic differences in thin filament assem-bly, length and regulation that exist between skeletal and cardiac muscle (Gregorio1997).
Several functional studies emphasize the critical role of E-Tmod for properthin filament lengths, organization, and contractile activity. Specific disruption ofE-Tmod actin-capping activity in chick cardiac myocytes results in an elongationof the thin filaments from their pointed ends and a dramatic reduction in their beat-ing activity (Gregorio et al. 1995). Alternatively, disruption of E-Tmod interactionwith tropomyosin causes a loss of thin filaments in the same cell type (Mudryet al. 2001). These studies highlight the distinct functional domains of E-Tmod:its interaction with actin is required to maintain thin filament lengths, whereas itsinteraction with tropomyosin is necessary for thin filament stability. Other studiesunderscore the importance of regulated levels of Tmod in muscle structure. Over-expression of E-Tmod in rat and chick cardiomyocytes results in shortened thinfilaments and myofibril degeneration; decreased E-Tmod expression results in theformation of abnormally long myofibrils (Sussman et al. 1998a, Littlefield et al.2001). Transgenic mice that overexpress E-Tmod (TOT mice) in their myocardiumexhibit symptoms similar to DCM, including a loss of myofibril organization andimpaired contractile activity (Sussman et al. 1998b). Interestingly, overexpressionof theDrosophilahomologue of Tmod, Sanpodo, during development of IFM ir-reversibly arrests the elongation of thin filaments and affects flight ability. There-fore, a transient increase in the levels of Sanpodo in this system converts it froma dynamic to a permanent cap (Mardahl-Dumesnil & Fowler 2001). In contrast,GFP-E-Tmod rapidly binds to and dissociates from thin filament pointed ends inchick cardiac myocytes (Littlefield et al. 2001). The mechanisms responsible forthe dynamic versus static capping activity ot Tmod remain elusive. In conclusion,Tmod’s dynamics, expression levels, and its distinct sarcomeric associations arecritical for regulating actin assembly, which in turn modulate thin filament lengths.
THICK FILAMENTS
Toward the center of the sarcomere lies the A-band, which contains the bipolarthick filaments comprised of myosin and associated proteins. The pointed endsof the thin filaments interdigitate with the thick filaments within the A-band. Theglobular heads of the myosin molecules, also known as cross-bridges, extend from

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
650 CLARK ET AL.
the core of the thick filaments and cyclically interact with the thin filaments. Theregion of the thick filament containing the cross-bridges is known as the C-zone;the region of the filament containing only myosin tails that is anchored and alignedin the mid-line of the sarcomere, the M-line, is known as the bare zone. The C-terminal portion (Mr∼ 2 kDa) of titin also spans the A-band and M-line. Titinis widely believed to act as a molecular blueprint that dictates the sarcomericarchitecture of the thick filaments and M-line (Figure 2).
Myosin: Molecular Motor for Muscle Contraction
The thick filaments are composed mainly of hundreds of myosin molecules, themotor proteins of the sarcomere. Currently, the myosin superfamily is groupedinto 15 classes, and the conventional myosins, including muscle myosin, are classII members (reviewed in Sellers 2000). Muscle myosin contains two heavy chains(MHC) (Mr∼ 220 kDa) and four light chains (MLC) (Mr∼ 20 kDa). Two of thelight chains belong to the essential light chain (ELC) family, and the other twoare regulatory light chains (RLC); they may function to make fine adjustments tomyosin motor activity and add to the versatility of its kinetics. The entire myosinmolecule is often characterized into two functional regions: the head and the rod.The N-terminal region of each MHC and two light chains make up the myosinhead domain (the S-1 fragment) that forms the cross-bridges (reviewed in Milligan1996). The head domain, which forms the catalytic motor domain, is conservedin several organisms, as well as among other myosin classes (Sellers & Goodson1995), and contains the binding sites for actin and nucleotides (Rayment et al.1993). Upon hydrolysis of each ATP molecule, the head domain that interactswith actin undergoes a large angular rotation, resulting in a displacement of 100A. After completion of this power stroke, ADP is dissociated and the actomyosincomplex returns to the relaxed state. Each myosin head likely repeats this cycleseveral times in a single twitch (for a review on the detailed structure of the actin-myosin interaction, see Vale & Milligan 2000).
The C-terminal regions of the two MHC make up the elongated rod: The C-terminal end of the rod, also known as light meromyosin (LMM), contains coiled-coil domains involved in myosin polymerization. The other portion of the rod isreferred to as S-2 and connects the myosin heads to the thick filament core. Theprecise packing scheme of myosin in the thick filaments, as well as the structuralarrangements that occur during muscle relaxation and contraction, are currenttopics of intense investigation.
Importantly, because different striated muscles have varying power require-ments, myosin motors must exhibit a range of activity rates. This is accomplished,in part, by the existence of multiple striated muscle MHC and MLC isoforms andthe tight regulation of their expression (Weiss & Leinwand 1996, Reggiani et al.2000). The MHC isoforms combine with various MLC isoforms to form func-tionally distinct isomyosins. At least eight muscle MHC have been identified inmammals, each encoded by separate genes: MHCIIa, MHCIIx, and MHCIIb arefound in fast skeletal muscle fibers. Depending on the species, MHCI (also known

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 651
as MHCβ/slow) is expressed in the ventricle of heart and in slow skeletal muscle;MHCα is mainly expressed in the atria of heart muscle; MHCexoc is in extraocularmuscle fibers; and MHCemb and MHCneo are expressed in muscle at differentdevelopmental stages (e.g., Hughes et al. 1993: reviewed in Reggiani et al. 2000).In addition, slow and fast muscle fibers, as well as atrial and ventricular fibers,express various isoforms of ELC and RLC. Factors involved in the regulation ofmyosin isoform expression include thyroid hormone and load requirements (Schi-affino & Reggiani 1994, Canepari et al. 1998). Additionally, severalcisandtranselements that control skeletal muscle MHC gene expression have been identifiedin mouse (Allen et al. 2001). In stark contrast to vertebrates, many invertebrateshave only one MHC gene. InDrosophila, for example, tissue and stage-specificMHC forms are generated by alternative splicing of a single gene (Bernstein et al.1983, George et al. 1989). Interestingly, it appears that the functional diversity ofMHC isoforms in fly muscles is attained by sequence differences restricted to onlya few regions of the S1 molecule (mainly affecting sites involved in ATP bind-ing), compared with the more extensive sequence diversity that occurs throughoutthe vertebrate S1 molecule (Bernstein & Milligan 1997). This observation under-scores the importance of particular regions of myosin in tuning its power stroke,contributing to the diverse functional characteristics of different muscles.
In vitro ATPase and motility assays, along with animal model systems, havebeen utilized to determine the functional significance of myosin isoforms andthe physiological implications of myosin mutations. This is clinically significant,considering that many patients (∼30%) with HCM exhibit missense mutations inthe MHC head region or light chains (for reviews see Seidman & Seidman 2001,Marian & Roberts 2001). Many of these mutant myosins exhibit impaired ATPaseactivities, and the degree of impairment correlates with the severity of the disease(Cuda et al. 1997, Roopnarine & Leinwand 1998). In mice, a null mutation inthe cardiacα-MHC is lethal (Jones et al. 1996), and ectopic expression of theβ-(slow) isoform in adult ventricles causes significant changes in cardiac contrac-tility (Tardiff et al. 2000). Mutation of MHCIIβ in murine skeletal muscle causesa loss of body mass along with compensatory hypertrophy (Allen et al. 2001), andDrosophilaheterozygous for a MHC mutation display disrupted myofibril assem-bly in their IFMs (Mogami et al. 1986, Chun & Falkenthal 1988). These studiesindicate the critical importance of the regulation of the functionally distinct myosinisoform types and their level in muscle development, structure, and function.
Thick Filament-Associated Proteins
In addition to myosin, other proteins have been identified as components of thethick filaments and M-line region of the sarcomere. It is striking that many of theseproteins, as well as the giant protein titin, contain variable copies of∼80–100amino acid residue motifs belonging to the fibronectin (FN) type III superfamilyand the intermediate (I) set of the immunoglobulin (Ig) superfamily. Previously,these domains were characterized as components of extracellular matrix, adhesion,and cell surface receptor molecules, but it is now apparent that they have critical

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
652 CLARK ET AL.
roles in mediating sarcomeric protein interactions as well (reviewed in Kenny et al.1999). The thick filament-associated proteins appear to play more than a simpleaccessory structural role because they are implicated in thick filament assemblyand regulation of muscle contraction. For a review on the different mechanismsof thick filament assembly in vertebrates and invertebrates see Barral & Epstein(1999.)
MYOSIN BINDING PROTEINS C AND H (MyBP-C AND MyBP-H): MULTIFUNCTIONAL
THICK FILAMENT COMPONENTS MyBP-C (Mr∼ 140 kDa) and MyBP-H (previ-ously identified as 86 kDa protein although its actualMr is 58 kDa), also known asC- and H-protein, respectively, are myosin-binding proteins that contain severalFN and Ig domains. They are both distributed in the C-zone of the thick filament ina series of transverse stripes spaced 43 nm apart, although MyBP-H also is foundoutside this zone (Craig & Offer 1976, Bahler et al. 1985a, Bennett et al. 1986).Three embryonic isoforms of MyBP-C have been identified, and three isoformshave been characterized in adult muscle: skeletal fast, skeletal slow (previouslyidentified as MyBP-X), and cardiac, each of which is encoded by separate genes(e.g., Takano-Ohmuro et al. 1989, Vaughan et al. 1993a). Although the cardiacisoform is restricted to heart muscle, the slow and fast muscle isoforms can beco-expressed in various skeletal muscle tissues (Reinach et al. 1982, Dennis et al.1984). In contrast, only one isoform of MyBP-H has been identified that is exclu-sively expressed in the Purkinje fibers of heart, which lack MyBP-C, and in fastskeletal muscle fibers (Bahler et al. 1985b, Vaughan et al. 1993b, Alyonychevaet al. 1997). The functional implications of this restricted expression are unknown.
The unique biochemical properties of MyBP-C have implicated it in severalroles in muscle, while functional analyses of MyBP-H are cursory. First, bothMyBPs exhibit strong affinity for myosin; in fact, MyBP-C appears to contain twomyosin-binding sites. A myosin LMM-binding site is contained within MyBP-C’sC-terminal Ig motif, whereas its MyBP-C motif, a unique stretch of∼100 aminoacid residues at its N terminus, binds to myosin S2; this latter interaction is regu-lated by phosphorylation of MyBP-C (Okagaki et al. 1993, Gruen et al. 1999). TheC-terminal domains of MyBP-C also contain a titin-binding site, and currently it re-mains controversial as to which domains are required to target MyBP-C to the thickfilament (Gilbert et al. 1996, Gruen et al. 1999). Because MyBP-C interacts withboth the thick and titin filaments, it may function to link them together and/or alignthe thick filaments in the A-band. Although MyBP-H also contains a C-terminalmyosin-binding site, its binding to titin has not been reported. Second, both myosin-binding proteins appear to aid in the assembly of vertebrate muscle thick filamentsto their precise lengths of 1.6µm. MyBP-C reduces the critical concentrationfor myosin polymerization and the resulting filaments are longer and more uni-form in length than those polymerized in its absence (Koretz 1979, Davis 1988).Also, co-expression of MyBP-C (or -H) and myosin in nonmuscle cells resultsin the formation of cable-like copolymers (Seiler et al. 1996). Moreover, expres-sion of MyBP-C mutants lacking the C-terminal myosin-binding domain inhibits

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 653
myofibrillogenesis in chick skeletal muscle cultures (Gilbert et al. 1996). Fi-nally, MyBP-C and -H may be involved in regulating muscle contraction (forreview, see Winegrad 1999). For instance, they can inhibit actin-activated skele-tal muscle myosin ATPase and stimulate actin-activated cardiac muscle myosinATPase (Hartzell 1985). Interestingly, in cardiac muscle, the MyBP-C motif canbe phosphorylated at multiple sites by cAMP-dependent kinase (PKA) and aCa2+/calmodulin-dependent protein kinase (Gautel et al. 1995b); the extent ofits phosphorylation correlates with the rate of twitch relaxation and cross-bridgecycling (Hartzell 1984, Kunst et al. 2000). Thus MyBP-C may modulate the con-tractility of cardiac muscle and the assembly of thick filaments.
Recent animal models and clinical studies have emphasized the important rolesof MyBP-C. For example, left ventricle fibers from mice that ectopically expressan N-terminally truncated cardiac MyBP-C (still containing the myosin- and titin-binding sites and the N-terminal motif) exhibited an increased Ca2+ sensitivityof force development (Witt et al. 2001). Homozygous knock-in mice containingshortened MyBP-C myosin and titin-binding domains exhibit progressive DCMand disarrayed myofibrils (McConnell et al. 1999). Furthermore, mice expressingMyBP-C that totally lacked the myosin- and titin-binding domains had mild cardiachypertrophy, with some animals experiencing sudden death upon stress (Yang et al.2001). Mice lacking MyBP-C in heart are viable and fertile but exhibit profoundcardiac hypertrophy and impaired cardiac function (Harris et al. 2002). Notably,mutations in MyBP-C are responsible for∼20% of reported cases of familial HCMin humans. Most of the mutations affect the MyBP-C C-terminal region, whichcontains its myosin- and/or titin-binding sites (Marian & Roberts 2001). Thesestudies highlight the multifunctional roles of MyBP-C; however, much work isrequired to elucidate the physiological roles of MyBP-H.
ADENOSINE MONOPHOSPHATE DEAMINASE (AMP-DEAMINASE) AMP-deaminase isan allosteric enzyme involved in the regulation of adenosine metabolism and there-fore has been labeled a sensor of the cell’s changing energy requirements (Hisatomeet al. 1998). Interestingly, in myocytes, AMP-deaminase is not active in solubleform, but upon vigorous muscle contraction, a large portion binds to the myofibriland is activated (Rundell et al. 1992). In the sarcomere, AMP-deaminase localizesto the A-band, probably through its interaction with myosin heavy chain (Cooper& Trinick 1984). The enzyme also has been reported to bind titin, although the ex-act sites of interaction are unknown (Koretz et al. 1993). It appears that regulationof AMP-deaminase activity and levels is critical for muscle function. Patients withdeficient activity of the muscle isoform (AMPD1), due to truncations or mutations,develop skeletal muscle myopathies (Morisaki et al. 1992, Abe et al. 2000). On theother hand, patients with congestive heart failure or coronary artery disease whohave a mutant AMPD1 allele have a greater probability of survival than cardiacpatients with the normal AMPD1 locus (Anderson et al. 2000). Clearly, more in-sight into the regulatory mechanisms of muscle AMP-deaminase activity and itsclinical manifestations are required.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
654 CLARK ET AL.
THE M-LINE
Elegant electron microscopy studies in the 1960s and 1970s yielded ultrastruc-tural details of the middle of the sarcomere, the M-line region, proposed to be theanchoring site for the thick filaments (reviewed in Luther & Squire 1978). The ap-pearance of defined M-lines is considered to be the final step in myofibril assembly,as defined by restriction of thin filament length and the clear formation of two half-sarcomeres (Markwald 1973). The myosin filaments are cross-linked in the M-lineby transverse, electron-dense M-bridges. Most muscle types appear to have 3 to 5M-bridges plus up to 12 less dense lines. Additionally, other ultrastructural com-ponents may form M-filaments to link M-bridges, as well as secondary M-bridges(van der Ven et al. 1996); the components of these structures are unknown. Notably,the ultrastructure of the M-line regions varies with different muscle types and maybe dictated by the expression of differently spliced titin isoforms (Kolmerer et al.1996). Although the structure of the M-line is quite complex, it is surprising thatonly a small number of M-line constituents have been identified to date.
Myomesin and M-protein: PotentialFilament-Linking Molecules
Myomesin (Mr∼ 185 kDa) contains a unique N-terminal domain followed by 12repeating FN and Ig domains (Grove et al. 1984) and is detectable at the M-line region of all striated muscle fibers examined, arranged in an antiparallel,staggered fashion (Obermann et al. 1996). Myomesin contains a binding site forLMM within its N-terminal domain, but this is not sufficient for M-line targeting(Obermann et al. 1997, Auerbach et al. 1999). Instead, the neighboring Ig domainof myomesin is essential for its assembly into the M-line, suggesting that it alsointeracts with an unidentified sarcomeric component (Auerbach et al. 1999). Ad-ditionally, myomesin FN domains M4–M6 bind to titin M-line M4 domain, aninteraction inhibited by phosphorylation of myomesin Ser482 (Obermann et al.1997). Because myomesin binds titin and myosin, it may connect these filamentsystems, a role also proposed for MyBP-C and M-protein (see below). In this re-gard, it has been proposed that myomesin plays a role in integrating thick filamentsinto assembling sarcromeres (Ehler et al. 1999).
Several myomesin isoforms have been identified, and their expression patternsare highly regulated both spatially and temporally. In adult chicken, C-terminalsplicing events give rise to H-myomesin in heart, and S-myomesin in skeletal mus-cle (Agarkova et al. 2000). In embryonic hearts of vertebrates, the major isoform inearly development is EH-myomesin (embryonic heart myomesin), which is down-regulated around birth, and its sequence is identical to the previously identifiedskelemin sequence (Price 1987). EH-myomesin contains a serine and proline-richinsertion, the EH domain, which remarkably has elastic properties analogous tothe PEVK region of titin. Therefore, the M-line region of cardiac muscle may ex-hibit elasticity, at least in early development (Agarkova et al. 2000). Interestingly,

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 655
EH-myomesin appears to be re-expressed in cardiomyocytes isolated from mousestrains with dilated cardiomyopathy (Agarkova et al. 2000). However, future in-vestigations are required to establish the signficance of this finding.
Originally identified as a myosin-binding protein, M-protein (Mr ∼165 kDa)contains 12 Ig and FN domains in an order identical to myomesin (Masaki &Takaiti 1974, Vinkemeier et al. 1993). M-protein is restricted to the M-lines of fastmuscle fiber types and cardiac muscle, and is developmentally regulated (Groveet al. 1984, Noguchi et al. 1992). The cooperative interaction of two M-protein Igdomains, Mp2 and Mp3, is required for its interaction with LMM and is responsiblefor targeting M-protein to the M-line. The interaction of M-protein with myosin isrelatively weak in vitro and is regulated by protein kinase A (PKA) phosphorylationof a single serine residue (Ser76) in the adjacent Mp1 domain (Obermann et al.1998). The function of M-protein is not known. However, because it also interactswith titin (Nave et al. 1989), it may act in conjunction with MyBP-C and myomesinto anchor thick filament components to titin in fast skeletal and cardiac muscle.
MURF-1: A Potential Link Between SarcomereDegradation and Gene Expression
Yeast two-hybrid screens led to the identification of muscle-specific RING fingerprotein-1 (MURF-1) as a M-line-binding partner for titin (Centner et al. 2001).MURF-1 was also identified as striated muscle RING zinc finger (SMRZ) (Dai& Liew 2001). Subsequently, two highly similar proteins, MURF-2 and MURF-3were identified (Centner et al. 2001). Structurally, MURF family members con-tain an N-terminal zinc-finger RING domain, a MURF family conserved (MFC)domain, a B-box domain, leucine-rich coiled-coiled domains, and an acidic tail(Spencer et al. 2000, Centner et al. 2001). Thus the MURFs belong to the growingfamily of RING-B-box-coiled-coil (RBCC) proteins, molecules implicated in di-verse cellular functions including signaling, ubiquitination, and transcription (forreview, see Freemont 2000).
MURF family members homo- and hetero-oligomerize (Centner et al. 2001).However, MURF-1 (Mr∼ 40 kDa) appears to be the only MURF family memberthat binds directly to the titin M-line region. Its binding site is contained within thetitin Ig domains A168–169, located directly N terminal to the titin kinase domain.The titin-binding site on MURF-1 was mapped to its central region (Centner et al.2001). Recent studies have revealed important physiological roles for MURF-1in muscle. It is among a small subset of genes up-regulated in models of skeletalmuscle atrophy, and it has ubiquitin ligase activity (Bodine et al. 2001). MURF-1knock-out mice are resistant to skeletal muscle atrophy, consistent with the viewthat MURF-1 regulates muscle protein degradation (Bodine et al. 2001). In fact, afunction for MURF-1 interaction with the C-terminal region of titin in chick cardiacmyocytes was found to be the maintainence of M-line and thick filament structure(McElhinny et al. 2002). Intriguingly, MURF-1 is also a nuclear component thatinteracts with transcription factors and other nuclear components. Consequently,

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
656 CLARK ET AL.
MURF-1 may be a dynamic protein that links nuclear functions with myofibrilsignaling pathways, a function also proposed for calpain3/p94, cardiac ankyrinrepeat protein (CARP), and specific Z-line components.
Muscle-Specific Calpain3/p94
The titin M-line and I band regions contain a binding site for muscle-specificcalpain3/p94, a Ca2+-dependent intracellular cysteine protease with autolytic ac-tivity. At the M-line, p94 appears to bind to the titin intervening sequence 7 (IS7)(Sorimachi et al. 1995). This region of titin is present in heart but is missing insome fast twitch muscle fibers, suggesting that the presence of the p94-binding siteis regulated in different muscle types (Kolmerer et al. 1996). The titin-binding siteon p94 has been mapped to its IS2 domain, a unique sequence that also harbors anuclear localization signal (Sorimachi et al. 1995), suggesting that p94 may linknuclear functions with sarcomeric components.
Loss of function of p94 results in limb girdle muscular dystrophy type 2A(LGMD2A) (Richard et al. 1995). Studies with specific p94 mutants found inLGMD2A revealed that although some retained their autolytic activity and/orability to bind titin, all the mutants lost their proteolytic activity. This supports thehypothesis that the loss of p94 proteolytic activity is responsible for LGMD2A(Ono et al. 1998). Furthermore, in two different transgenic mouse models, bothexpressing inactive forms of p94, the mice exhibited significantly decreased gripstrength and a progressive muscular dystrophy (Tagawa et al. 2000, Richard et al.2000). Thus the proteolytic activity of p94 is critical for normal skeletal musclefunction. It remains to be determined whether titin is a substrate of p94.
MM-Creatine Kinase
Enzymes involved in metabolism interact with sarcomeric components at the M-line region. For instance, creatine kinase (CK) is an enzyme localized predom-inantly at cellular sites of energy production and consumption. In muscle cells,most of the muscle isoenzyme of CK (MM-CK) (Mr∼ 43 kDa) is soluble, but asmall portion (∼5–10%) is bound to the sarcomeric M-line region and, to a lesserextent, in the I-band (Turner & Walker 1987). Its interaction with the M-line ismediated through two pairs of lysine residues in the MM-CK N-terminal region(Hornemann et al. 2000). Creatine kinase functions to replenish ATP by trans-ferring the N-phosphoryl group from phosphocreatine to ADP (Kenyon & Reed1983). Thus this enzyme is likely utilized in the sarcomere to regenerate ATP, per-haps for the nearby actomyosin ATPase (see Ventura-Clapier et al. 1994). Trans-genic mice null for MM-CK lack skeletal muscle burst activity, but do not appearto have disarrayed myofibrils (van Deursen et al. 1993). The muscle isoform ofthe glycolytic enzymeβ-enolase also binds to the M-line region and to MM-CK(Foucault et al. 1999), indicating that the M-line region of the sarcomere mayanchor entire complexes of glycolytic enzymes, which could be critical in periodsof increased muscle activity. Future studies will reveal whether their enzymatic

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 657
activity is regulated by sarcomeric components, as has been reported for the en-zyme AMP-deaminase.
TITIN: THE IMMENSE, MULTIFUNCTIONALTHIRD FILAMENT SYSTEM
The third filament system of striated muscle consists of the huge, modular pro-tein titin. After actin and myosin, titin (also known as connectin) is the thirdmost abundant muscle protein and is the largest protein identified to date (Mr∼ 3–3.7 MDa) (Maruyama et al. 1977, Wang et al. 1979). The N-terminal ends oftitin, from adjacent sarcomeres, overlap in the Z-line. The molecules span theI- and A-bands, and their C-terminal ends overlap in the M-line, thus forminga continuous filament system in myofibrils. Several unique structural propertiesestablish titin as a crucial, multifunctional sarcomeric component. First, distinctstructural components in the titin I-band region have elastic properties. Thus titinappears to be a molecular spring that governs some aspects of myofibrillar stiffness.Second, the repeating motif structure of titin, its early assembly during myofibril-logenesis, and its interactions with several regulatory and sarcomeric components,make it a primary candidate for acting as a sarcomeric template and stabilizer.Third, the titin C-terminal region contains a Ser/Thr kinase domain whose func-tion remains elusive, but whose presence suggests that titin also is involved insignaling pathways. Rapid advances into the investigation of titin’s propertieshave resulted from the availability of its complete cDNA sequence, and now thegenomic sequence of the single gene that encodes it (Labeit & Kolmerer 1995b,Bang et al. 2001a). The human gene contains 363 exons, encoding 38,138 aminoacid residues (Mr ∼ 4.2 MDa) (Bang et al. 2001a)! Numerous splice isoformshave been identified, most located within I-band titin, that control the number andsize of the spring elements. Other splicing hot spots are included within Z-lineand M-line titin, both of which likely contribute to the layout of these sarcomericregions.
The majority (∼90%) of the titin molecule is made up of repeating modulardomains from the FNIII and the Ig superfamilies. About 10% of titin’s mass isorganized into 17 nonrepetitive sequences that are situated between the Ig and FNIIIrepeats. One interdomain insertion contains the kinase domain and the other 16have no significant similarity to each other or to other known proteins. Determiningthe functional properties of individual titin domains is presently the focus of intenseresearch (McElhinny et al. 2000, Sanger & Sanger 2001).
Z-Disc Titin: A Key Contributor to Sarcomere Integrity
The N terminus of titin, representing the first∼80 kDa of the protein, spans theentire Z-disc (Gregorio et al. 1998, Young et al. 1998). This region of titin has beendivided into three subdomains, based on their molecular features and functions.The extreme N-terminal region (residues 1–200) contains the first two Ig-repeats

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
658 CLARK ET AL.
(Z1 and Z2) and binds telethonin/Tcap (Gregorio et al. 1998, Mues et al. 1998).Residues 450–750 contain a 45-amino acid repeat known as the titin-repeat (Zr).The number of repeats varies among different muscle types, implicating titin as adeterminant of Z-line architecture (Peckham et al. 1997, Sorimachi et al. 1997). Infact, the Z-repeats interact with the C terminus ofα-actinin, suggesting a model inwhich the number of repeats regulates the number ofα-actinin cross-links in the Z-line (Ohtsuka et al. 1997, Sorimachi et al. 1997, Young et al. 1998). The N terminusof α-actinin contains a stretch of sequence similar to the Z-repeat of titin, whichmay prevent their association. This inhibition is relieved by phosphatidylinositol4,5 bisphosphate, (PI4,5P2), which provides an attractive regulatory mechanism(Young & Gautel 2000). Just distal to the Z-repeats lies the third Z-domain of titin,which contains anotherα-actinin-binding site that resides within the spectrin-repeats in the rod domain (Young et al. 1998).
All functional studies to date indicate that Z-disc titin is critical for sarcom-eric stability in cultured myocytes. Disruption of myofibrils has been reportedas a result of expression of (a) the first 362 amino acids of titin (zeugmatin)(Turnacioglu et al. 1997); (b) the entire Z-disc region of titin (Peckham et al. 1997);(c) a C-terminal truncated fragment ofα-actinin (missing its titin-binding site)(Schultheiss et al. 1992); (d) telethonin or the Z1–Z2 domains of titin(Gregorio et al. 1998); and (e) single Z repeats (Ayoob et al. 2000). Clinically,two mutations in DCM patients were found in titin’s Z-line region; both decreasedtitin’s affinity for telethonin/Tcap orα-actinin in yeast two-hybrid assays (Itoh-Satoh et al. 2002). These studies indicate that the association of titin filaments withZ-disc components is critical for the maintenance of myofibril structure.
I-Band Titin: A Molecular Spring
In addition to the active force generated by the actomyosin-ATPase, myofibrilsproduce an independent passive force. When unactivated myofibrils are stretchedbeyond or shortened below their resting (slack) length, passive force maintainsthe overlap of the thin and thick filaments. This intrinsic property of muscle isspecifically attributed to the I-band region of titin, which contains spring-likeelements that contribute to the degree of myofibril stiffness.
The I-band region of titin is composed of tandem stretches of Ig modules withunique sequences inserted in between. One insert is the PEVK domain, so namedbecause∼70% of it is made up of proline, glutamine, valine, and lysine residues(Labeit & Kolmerer 1995b). The structure of the PEVK domain has been describedas a random coil, but it may actually assume a wide range of elastic conformations(Li et al. 2001). The I-band region of titin also contains the N2A linker element,made of four Ig domains and a 106-residue insert. Another element, N2B, isspecific to heart muscle and contains several Ig domains plus unique sequences.Innovative biophysical studies indicate that the titin molecule behaves as the “sumof its parts.” That is, upon physiological stretching of the sarcomere, the distincttitin I-band spring elements appear to be recruited in a sequential order: The Ig

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 659
domains elongate (but do not unfold) at lower forces, whereas at higher forces andgreater stretch, the PEVK region unravels (Linke 2000,Wang et al. 2001, Granzier& Labeit 2002). In cardiac muscle, the third spring element, the N2B region,extends upon still higher levels of stretch (Linke et al. 1999, Trombitas et al.1999). Recent molecular characterization of titin transcripts from various muscletypes has revealed that exon-skipping in the titin I-band region is the basis for theelastic diversity of different vertebrate muscles (Freiburg et al. 2000, Cazorla et al.2000). The story becomes more remarkable with the recent discovery of 50 novelI-band exons and three unique I-band titin isoforms in striated muscle (novex1–3).Novex-3 is surprisingly small for a titin isoform (∼700 kDa). It interacts in theI-band with the giant protein obscurin, which contains several signaling domains:the complex therefore may include an elastic signaling system that links Z-linesto I-bands (Bang et al. 2001a).
A puzzling issue is whether the titin I-band region is “free” like a spring orwhether it interacts with the thin filaments. Previously, it was proposed that titinis laterally associated with the thin filaments, albeit weakly (Maruyama et al.1987, Funatsu et al. 1993). Several in vitro binding assays with native titin, itsfragments, or recombinant titin modules have supported this hypothesis, but therelative affinities of these interactions vary (e.g., Soteriou et al. 1993, Jin 1995).Electron microscopy studies involving the selective removal of the thin filamentsin myofibrils suggest that the I-band region of titin is independent of the thinfilaments, except possibly near the Z-line (Trombitas et al. 1997). The issue becameeven more complex when it was determined that exogenous expression of titinN2B specifically disrupts thin filaments in chick cardiac myocytes, suggesting thatcardiac titin directly or indirectly stabilizes the thin filaments (Linke et al. 1999).Recent studies have found that the PEVK region of the N2B form of cardiac titin,but not the PEVK region of skeletal muscle titin, interacts under physiologicalconditions with actin filaments and inhibits actin filament sliding in vitro (Kulkeet al. 2001a, Yamasaki et al. 2001). Interestingly, this interaction is inhibited bythe vertebrate Ca2+-binding protein, S100/A1, providing a possible mechanismthat could free the thin filaments from titin before contraction or stretch (Yamasakiet al. 2001). Recently, a recombinant PEVK fragment was also reported to bind tonebulin; the interaction was abolished by Ca2+/calmodulin (Gutierrez-Cruz et al.2001). These studies support the exciting hypothesis that different thin filamentcomponents may interact with different titin isoforms, perhaps further modulatingtitin’s elastic properties.
The critical physiological significance of the elastic region of titin is emphasizedfrom genetic and clinical studies. Titin mutations in humans, including mutationsin the I-band region, result in familial DCM (Gerull et al. 2002, Itoh-Satoh et al.2002). In zebrafish, a novel genetic model system for the titin field, a mutation inthe N2B region also causes a functional disorder resembling human DCM (Xu et al.2002). Skeletal muscles from Duchenne MD patients exhibit titin mRNA down-regulation, and a specific N2A mutation is associated with muscular dystrophywith myositis (mdm) in mice (Tkatchenko et al. 2001, Garvey et al. 2002).

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
660 CLARK ET AL.
A-Band Titin: A Thick Filament Template?
The A-band region of titin interacts with the thick filament components MyBP-C,AMP-deaminase, and the tail region of myosin (Soteriou et al. 1993, Houmeidaet al. 1995) whose binding sites may reside within its FNIII and Ig domains (Labeit& Kolmerer 1995b). One type of super-repeat lies within the D-zone of titin, theregion located near the A/I junction: Five FNIII and two Ig domains are repeatedseven times. C-zone titin contains seven FNIII domains and four Ig domains,which are repeated 11 times. Because the 11 domain super-repeats correspondto the 11 43-nm repeats of the thick filament C-zone, these repeats may specifythe number and location of myosin and MyBP-C molecules (Whiting et al. 1989,Houmeida et al. 1995). In fact, titin’s MyBP-C binding sites have been mapped tothe first Ig domain within these super-repeats (Freiburg & Gautel 1996). Recently,recombinant FNIII domains of A-band titin were found to bind to the myosin headdomain and to influence the actomyosin interaction. Thus titin FNIII domains mayposition the myosin heads close to the thick filament backbone (Muhle-Goll et al.2001). Functional evidence also indicates an organizational and structural rolefor titin in the A-band. For instance, ablation of titin expression using antisensetechniques in rat cardiomyocytes disrupted myosin incorporation into myofibrils(Person et al. 2000). Furthermore, a titin homozygous knock-out in a myofibroblastcell line resulted in a lack of thick filament formation and a subsequent impairmentof myofibril assembly (van der Ven et al. 2000a). These studies support a proposedrole for titin as template for the assembly of thick filaments.
M-Line Titin: A Potential Signal Transducer
Approximately 250 kDa of the C-terminal region of titin lies within the M-lineregion and is subjected to considerable splicing. Thus the titin C-terminal regionmay be a template for the M-line ultrastructure that differs among fiber types(Kolmerer et al. 1996b). Interestingly, the titin M-line region contains a putativephosphorylation site and a catalytically active serine/threonine (Ser/Thr)-kinasedomain (Labeit & Kolmerer 1995b). This domain is highly homologous to theSer/Thr-kinase domains contained in myosin light chain kinase (MLCK) and insome titin homologues (e.g., Benian et al. 1989, Ayme-Southgate et al. 1991,Labeit et al. 1992; see below). Clues into the regulation of the titin kinase wereobtained when its crystal structure was determined (Mayans et al. 1998). One ofthe kinase domain’s tyrosine residues occludes its active site; thus the kinase isauto-inhibitory. Activation occurs upon phosphorylation of this tyrosine and thebinding of Ca2+/calmodulin to a portion of the kinase tail. Titin kinase was pro-posed to phosphorylate the Z-line protein telethonin, an event hypothesized tobe involved in myofibrillogenesis (Mayans et al. 1998). The feasibility of howthe Z-disc and M-line regions of a sarcomere interact during myofibrillogenesis isunknown. Additionally, the titin kinase may phosphorylate myosin to regulate mus-cle contraction. Although recombinant twitchin kinase (see below) was reportedto phosphorylate myosin light chains from mollusk, evidence for vertebrate titin

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 661
performing the same function is lacking (Heierhorst et al. 1995). Another proteinthat may be involved in specific titin kinase-based pathways is MURF-1, whichinteracts with the Ig domains flanking the kinase domain (Centner et al. 2001).
Titin Homologues: Model Systems ProvideInsight into Titin Function
Genetic studies using insect and nematode model systems have led to a greaterunderstanding of the roles of “mini-titin” homologues in muscle.Drosophilatitin(Mr 2 MDa), D-titin, is only∼54% the size of human soleus muscle titin and ismost homologous to the N-terminal half of its human counterpart. D-titin containstwo PEVK domains but lacks the A-band and M-line domains characteristic ofvertebrate titin. Functional studies indicate D-titin is required for myoblast fusionand sarcomere assembly and structure (Zhang et al. 2000, Machado & Andrew2000). Surprisingly, D-titin also appears to have a role in chromosomal structureand in mitosis, perhaps because of its extensible properties (Machado & Andrew2000).Drosophilakettin, originally reported as a Z-disc-associated mini-titin, islikely an alternatively spliced variant of D-titin (Kolmerer et al. 2000, Machado& Andrew 2000). Kettin is required for fly development, normal muscle function,and flight activity, and may reinforce the Z-disc anchoring of the thin filamentsbecause it binds to actin andα-actinin (e.g., van Straaten et al. 1999, Kolmereret al. 2000, Hakeda et al. 2000).Drosophilaprojectin is most homologous to theC. elegansmini-titin, twitchin (UNC-22), which interacts with the thick filaments,functions in sarcomere assembly, and also may regulate myosin activity (Benianet al. 1996a). Interestingly, depending on the isoform and muscle fiber, projectinlocalizes to the I-band or the A-band in flies, which may reflect its potential multi-functionality in different muscle types (Vigoreaux et al. 1991). Both twitchin andprojectin contain a kinase domain and FNIII/Ig domains in their C termini but lackPEVK domains. Consequently, in invertebrates, D-titin and kettin (or homologousproteins inC. elegans) may span the Z-disc and I-band regions, and act togetherwith projectin or twitchin in the A-bands, to function as a single vertebrate titin-like molecule (Machado & Andrew 2000, Kulke et al. 2001b). Indeed, isoformsof projectin and kettin may work together to modulate the high passive stiffnessof IFM (Kulke et al. 2001b). Lastly, the sequence of another putativeDrosophilamini-titin gene,stretchin-MLCK(Mr∼ 1 MDa) recently was reported. The proteinis predicted to contain two PEVK regions, 32 Ig domains, two FNIII domains,and a kinase domain, based on the particular isoform (Champagne et al. 2000).Stretchin-MLCK is similar toC. elegansunc-89 and human obscurin MLCK.
NEBULIN: A RULER FOR THIN FILAMENT ASSEMBLY?
Nebulin is another giant protein that forms the fourth filament system in vertebrateskeletal muscles (Mr 600–900 kDa). The C-terminal end of nebulin is partiallyinserted into the Z-lines, whereas its N-terminal end extends to the pointed ends of

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
662 CLARK ET AL.
the thin filaments (Millevoi et al. 1998, McElhinny et al. 2001). Nebulin’s unique,inextensible structural properties have made it a prime candidate to act as a mole-cular ruler for specifying the precise lengths of the thin filaments. For example,different sizes of nebulin isoforms, generated by alternative splicing events froma single gene, correlate with the different thin filament lengths present in variousmuscle fibers and during muscle development (Kruger et al. 1991). Nebulin ap-pears to assemble early during myofibrillogenesis, before actin filaments attaintheir mature lengths and organization, consistent with its role as a thin filamenttemplate (Ojima et al. 1999). Analysis of the nebulin 20.8 kb cDNA sequencealso supports its proposed role as a thin filament template (Labeit & Kolmerer1995a). Specifically, the vast majority of the nebulin molecule (∼97%) contains185 modular motifs, each∼35-amino acid residues in size. Although a single,central tyrosine residue is the only amino acid strictly conserved among all themotifs, groups of modules are classified into distinct subfamilies. The central 154modules (M9–M162) make up 22 super-repeats of 7 modules each: These super-repeats precisely correspond to the periodicity of the 7 actin monomers per 1TM/Tn regulatory complex. The nebulin extreme C-terminal end contains an SH3domain and multiple phosphorylation motifs; its N-terminal end contains uniquemodules M1–M8 and an 8-kDa acidic domain at the pointed ends of the thin fila-ments. This overall architecture appears to be highly conserved among vertebrates(Herrera et al. 2000). Giant proteins similar to vertebrate nebulin also have beenidentified in chordates and earthworms, suggesting that nebulin homologues mayexist throughout the animal kingdom (Royuela et al. 1997, Fock & Hinssen 1999).
Owing to the giant size of native nebulin and its susceptibility to degradation,most information concerning its properties has come from biochemical studiesusing proteolytic nebulin fragments or synthetic or recombinant modules. Nebulinmodules from the central super-repeat region of the molecule (1–15 modules inlength) bind to actin, although the relative affinities of these interactions vary.Some modules also promote actin nucleation and filament bundling, as well asstability to actin filaments (e.g., Jin & Wang 1991, Pfuhl et al. 1994). It is likelythat a single nebulin module interacts with each actin monomer, whereas eachnebulin super-repeat interacts with one thin filament regulatory complex (Labeit& Kolmerer 1995a). In support of this idea, certain nebulin modules also appear tobind to tropomyosin and the troponins (Wang 1996). Additionally, the extreme N-terminal nebulin modules M1–M3 interact with the thin filament capping protein,Tmod (McElhinny et al. 2001). Therefore, it is plausible that the nebulin moleculeworks together with capping proteins in order to specify and maintain the preciselengths of the thin filaments.
Intriguingly, nebulin may have additional, more complex functions in the sar-comere. Modules from nebulin predicted to span the A-I junction bind to actin,myosin, and calmodulin (Jin & Wang 1991, Wang et al. 1996, Root & Wang2001). Importantly, modules from this region of nebulin also inhibit actomyosinATPase activity, as well as sliding velocities of actin filaments over myosin, in aCa2+/calmodulin-dependent manner in vitro (Root & Wang 2001). Thus nebulin

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 663
may comprise another system that regulates the actin-myosin interaction, an ideasupported by recent studies indicating that nebulin may have multiple binding siteson the thin filament, analogous to tropomyosin (Lukoyanova et al. 2002). More-over, initial clues into the functional properties of the C-terminal region of nebulinhave recently been elucidated: Its SH3 domain interacts with myopalladin, andmodules M160–170 from the peripheral region of the Z-line interact with the in-termediate filament protein, desmin (Bang et al. 2001b). The interaction of nebulinwith desmin likely contributes to a lateral linkage system to maintain the alignmentof Z-lines from one myofibril to the next.
A perplexing issue arises from the finding that nebulin is not detected in car-diac muscle. Although cardiac thin filament lengths are broader in their distributionthan their skeletal muscle counterparts, their lengths nevertheless must be highlyregulated (Robinson & Winegrad 1979). A cardiac-specific petite nebulin, neb-ulette (Mr∼ 107 kDa), contains 22 nebulin-related modules, a C-terminal regionthat is virtually identical to nebulin’s, and a unique N-terminal end. It binds toactin, tropomyosin, and myopalladin and is critical for myofibril assembly, thinfilament organization, and contractile activity of cardiac myocytes (Moncman &Wang 2002, Bang et al. 2001b). Another nebulin-related protein (N-RAP), is foundin intercalated discs in cardiac muscle (Luo et al. 1997). Based on their small sizesand distribution patterns, it is unlikely that nebulette and N-RAP act as moleculartemplates for thin filament length specification. Surprisingly, full-length nebulinmRNA transcripts have been detected in heart muscle, but at lower levels than inskeletal muscle (S. Kazmierski, P. B. Antin, S. Labeit & C. C. Gregorio, unpub-lished data). Determination of the functional significance of this finding awaitsfurther study. Finally, it is possible that cardiac tissue utilizes molecular mech-anisms other than nebulin for thin filament length specification and regulation(Littlefield & Fowler 1998). Further studies on nebulin and nebulin-like proteinsare expected to address these issues, as well as to shed light on specific muscledisorders. In particular, nebulin mutations are associated with nemaline myopa-thy (e.g., Gurgel-Giannetti et al. 2001), and nebulin levels are reduced in someforms of muscular dystrophy (Wood et al. 1987). In cardiac muscle, a polymor-phism in the human nebulette gene is associated with nonfamilial idiopathic DCM(Arimura et al. 2000). The mechanisms to explain the essential roles of nebulinand nebulin-like proteins await further study.
THE Z-LINE: CYTOSKELETAL ANCHOR AND SIGNALTRANSDUCTION CENTER
The Z-line defines the lateral boundaries of the sarcomere and constitutes an an-choring site for the thin, titin, and nebulin filaments. Because of this anchoringproperty, Z-lines are the primary conduits of the force generated by contraction.Z-lines of adjacent myofibrils are aligned, providing a means to coordinate con-tractions between individual myofibrils to a focal point where the Z-line is linked

23 Sep 2002 10:30 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
664 CLARK ET AL.
to the muscle membrane. Abundant intermediate filament proteins are presentat the Z-line periphery and probably form the link between adjacent myofibrils.Interestingly, during muscle assembly, precursor Z-lines and associated proteins(also referred to as Z-bodies or I-Z-I structures) are the first identifiable structuresobserved, suggesting that they furnish the initial cue for the polarized organizationof filament systems in nascent sarcomeres.
Just eight years ago,α-actinin was the only well-characterized Z-line protein;however, the rapid advances in molecular biology have revealed that the Z-line isactually a complex protein network (reviewed in Faulkner et al. 2001) (Figure 3).Interestingly, numerous signal transduction components are constituents of theZ-line, facilitating its role as a biomechanical sensor that can respond to changesin tension at the sarcolemma. Another recent and intriguing observation is that asurprisingly large number of Z-line associated proteins have a dynamic distributionin muscle and may shuttle between the Z-line and other subcellular locations(e.g., the nucleus) to transmit signals. Thus the Z-line is not simply the structuralborder of the sarcomere but likely plays an important role in signaling and musclehomeostasis.
α-Actinin: Primary Cross-Linker of the Z-line
A major component of the Z-line isα-actinin (Mr 97 kDa), a member of the spectrinsuperfamily that functions as an actin cross-linking protein (Maruyama & Ebashi1965; reviewed in Blanchard et al. 1989).α-Actinin contains three major domains:a globular N-terminal actin-binding domain, a central rod domain composed of fourspectrin-repeats, and a C-terminal domain that bears similarity to calmodulin. Therecently solved crystal structure of theα-actinin rod domain provides a mechanisticunderstanding of howα-actinin functions to cross-link actin filaments (Djinovic-Carugo et al. 1999). The rod domains ofα-actinin monomers interact to establishantiparallel dimers that are capable of cross-linking actin and titin filaments fromneighboring sarcomeres. The width of a Z-line can vary from∼ 30 nm (fish skele-tal muscle) to∼ 160 nm (vertebrate cardiac muscle) (Franzini-Armstrong 1973,Goldstein et al. 1979), suggesting that the configuration and/or number of theα-actinin cross-links may differ between muscles.
In higher vertebrates, there are fourα-actinin genes that appear to have someoverlapping function.α-Actinin-1 and -4 are nonmuscle isoforms involved in actinfilament bundling and attachment to membranes (Honda et al. 1998).α-Actinin-2and -3 are both expressed in skeletal muscle and localize to the Z-line (North &Beggs 1996). However, they are not co-expressed in all striated muscle becauseα-actinin-3 is absent from cardiac muscle. Especially intriguing is the observationthat∼ 18% of the human population is deficient inACTN-3, with no observablephenotype, strongly suggesting thatα-actinin isoforms can compensate for theloss of another family member (North et al. 1999).
Although α-actinin is a prominent component of the Z-disc, genetic studiessuggest that, at least in invertebrates, it is not absolutely essential for the properassembly of the contractile machinery. For example, inDrosophila (which hasonly oneα-actinin gene) embryos deficient inα-actinin form morphologically

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 665
normal muscles that retain some capacity to contract, albeit poorly (Fyrberg et al.1998). These mutant embryos die soon after hatching and exhibit defects in Z-line organization and muscle attachment to the tendon matrix. Thusα-actinin isnot absolutely required for initial sarcomere assembly in this system and insteadappears to be critical for stabilizing the muscle cytoskeleton once contractionbegins. This observation has led to the hypothesis that other proteins cooperatewith α-actinin to cross-link and/or anchor thin filaments at the Z-line.
In the past decade, several new Z-line components have been discovered (forreviews see Chien 2000, Faulkner et al. 2001). Many of these Z-line-associatedproteins directly bindα-actinin. The modular nature ofα-actinin allows for multi-ple protein interactions to occur simultaneously. From this perspective,α-actininmay act as part of a nexus for the assembly of the Z-line complex. However, asnoted above, Z-line assembly in at least one system can occur in the absence ofα-actinin; this may be a reflection of the multiplicity of stabilizing protein-proteininteractions at the Z-disc that allows some function to be maintained when an in-dividual constituent is absent. Clearly,α-actinin itself has the capacity to associatewith numerous protein partners at the Z-disc. In addition to actin, the N-terminalglobular domain of ofα-actinin binds cysteine-rich protein (CRP) family members(Pomies et al. 1997) and zyxin (Crawford et al. 1992). The central four spectrinrepeats ofα-actinin bind a variety of Z-line proteins such asα-actinin associatedLIM protein (ALP) (Xia et al. 1997), FATZ (filamin,α-actinin- and telethonin-associated Z-line protein), calsarcin-2/myozenin (Faulkner et al. 2000, Frey et al.2000, Takada et al. 2001), myotilin (Salmikargas et al. 1999), and titin (Younget al. 1998). Other interactions have been mapped to the C terminus ofα-actininincluding cypher/ZASP/oracle (Zhou et al. 1999, Faulkner et al. 1999, Passier et al.2000), myopalladin (Bang et al. 2001b), and titin (through a different titin domain)(Ohtsuka et al. 1997, Sorimachi et al. 1997). Together, these protein interactionsmay provide tensile integrity to the Z-line and also serve as additional dockingsites for other Z-line-associated proteins.
α-Actinin-Associated Proteins
MUSCLE LIM PROTEIN (MLP) MLP (Mr 23 kDa) is a member of the cysteine-richprotein (CRP) family, a group of evolutionarily conserved cytoskeletal proteinsprominently expressed in a variety of muscle tissues (Louis et al. 1997). All fam-ily members are enriched withα-actinin at sites of actin filament anchorage (Louiset al. 1997). MLP is the striated muscle-specific CRP present in both embryonicand adult muscle, where it localizes at the periphery of the Z-line (Arber et al.1997), and at sites of membrane/cytoskeletal linkage such as the intercalated disc(Ehler et al. 2001, Flick & Konieczny 2000). MLP is comprised of two tandemlyarrayed LIM domain/glycine-rich modules (protein interacton domains), and uti-lizes the first glycine-rich region to associate directly withα-actinin (Harper et al.2000). In addition toα-actinin, MLP can bind several other cytoskeletal proteinsincluding zyxin (Sadler et al. 1992),β-spectrin (Flick & Konieczny 2000), andnebulin-related anchoring protein (N-RAP) (Ehler et al. 2001), reinforcing the ideathat it acts as a scaffold or linker protein. Functional elimination of MLP in the

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
666 CLARK ET AL.
mouse results in DCM, associated with a severely disorganized cardiac contractileapparatus, suggesting that MLP plays a primary role in stabilizing Z-disc structure(Arber et al. 1997). A more recent analysis of MLP−/− cardiomyocytes foundmorphological defects at costameres and intercalated discs, membrane attachmentsites for the contractile apparatus (Ehler et al. 2001). Although speculative, MLPmay contribute to muscle cytoskeleton integrity by linking the contractile appa-ratus and the muscle membrane at multiple sites through its associations withα-actinin (at the Z-line),β-spectrin (at costameres), and N-RAP (at intercalateddiscs) (Flick & Konieczny 2000, Ehler et al. 2001).
In addition to its contribution to the mechanical integrity of muscle cytoar-chitecture, MLP has also been implicated in communication with the nuclearcompartment. MLPs in vertebrates and inDrosophilacan be found in the nucleus(Arber et al. 1994, Stronach et al. 1996). Moreover, the translocation of MLP to thenucleus may be regulated by hypertrophic signals (Ecarnot-Laubriet et al. 2000).Expression of MLP in cultured myocytes appears to enhance muscle differentia-tion (Arber et al. 1994); this could reflect a nuclear function for MLP or a role instabilization of the developing contractile apparatus. The ability of MLP to interactin vitro with the muscle transcription factor, MyoD, has led to some speculationabout whether MLP plays a direct role in transcriptional regulation (Kong et al.1997); however, additional experiments will be required to confirm this possibilityin vivo.
THE ALP-ENIGMA FAMILY The ALP-enigma family is a newly emerging groupof cytoskeletal proteins defined by an N-terminal PDZ domain (a protein-proteininteraction module) and one or three C-terminal LIM domains. Several familymembers are expressed in striated muscle and localize to the Z-line, includingenigma (Mr∼ 55 kDa) (Guy et al. 1999), ALP (α-actinin associated LIM protein)(Mr∼ 36 and 40 kDa) (Xia et al. 1997, Pomies et al. 1999), and cypher/ZASP/oracle(Mr∼ 77kDa) (Zhou et al. 1999, Faulkner et al. 1999, Passier et al. 2000). Z-linetargeting of these proteins is thought to take place, in part, by a direct interac-tion between the N-terminal PDZ domain andα-actinin. Enigma seems to be theexception; its PDZ domain binds tropomyosin, and this interaction appears es-sential for enigma’s localization to the Z-line (Guy et al. 1999). The C-terminalLIM domain(s) serves as a docking site for other proteins; thus one function ofenigma proteins may be to anchor signaling molecules at the Z-line. In supportof this hypothesis, enigma and cypher bind to protein kinase C isoforms throughtheir LIM-domains (Zhou et al. 1999, Kuroda et al. 1996). Enigma also binds theinsulin receptor and Ret/ptc (Durick et al. 1996, Wu & Gill 1994), and cypher islinked to the phosphatase calcineurin by binding to members of the FATZ family(Frey & Olson 2002).
Both cypher and ALP have been genetically ablated in the mouse, allowing fora dissection of their functions in vivo (Zhou et al. 2001, Pashmforoush et al. 2001,Jo et al. 2001). These analyses indicate a role for ALP and cypher in stabilizing themuscle cytoskeleton, suggesting that this could be a general property of enigma

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 667
family members. Cypher−/− mice exhibit disrupted Z-lines and die from stri-ated muscle failure, including ventricular dilation, respiratory distress, and severeskeletal muscle weakness (Zhou et al. 2001). The skeletal muscle defect is postna-tal, indicating that cypher is not necessary for the assembly and organization of theZ-line, but instead stabilizes this structure once muscle contraction commences.ALP-deficient mice present a mild cardiomyopathy whose pathology is similarto a human cardiomyopathy caused by mutations in plakoglobin, a protein thatlinks cellular adhesion molecules to the actin cytoskeleton (Pashmforoush et al.2001). In addition to its Z-line localization, ALP is enriched with plakoglobin andα-actinin at cardiac muscle intercalated discs, suggesting that the primary cellulardefect in the ALP−/−mice is at this membrane linkage. Overexpression of ALPin rat neonatal cardiomyocytes potentiates myofibrillogenesis, and biochemicalstudies nicely demonstrate that ALP enhances the actin cross-linking propertyof α-actinin (Pashmforoush et al. 2001). Together these observations support amodel by which ALP, and perhaps other enigma family members, promotesα-actinin’s cross-linking activity at sites of actin filament anchorage (e.g., Z-linesand intercalated discs).
THE FATZ FAMILY FATZ (Mr 32 kDa) is an acronym for filamin-,α-actinin- andtelethonin-binding protein of the Z-disc (Faulkner et al. 2000), indicating itsrepertoire of binding partners. FATZ originally was identified as an EST highlyexpressed in skeletal muscle (Faulkner et al. 2000). Two other laboratories indepen-dently identified FATZ by yeast two-hybrid analysis; one discovered calsarcin-2as a calcineurin-binding protein (Frey et al. 2000). The other identified myozeninas anα-actinin- and filamin-binding protein (Takada et al. 2001). The protein canbe divided into three domains: the N- and C- terminal domains areα-helical in na-ture and separated by a glycine-rich central domain (Faulkner et al. 2000, Takadaet al. 2001). These domains led to the identification of two other family members(calsarcin-1 and calsarcin-3) that also display a high degree of sequence similaritywithin these defined regions (Frey et al. 2000, Frey & Olsen 2000); therefore,FATZ is the founding member of a new protein family.
FATZ association withα-actinin, filamin, and telethonin may direct and/ortether FATZ to the Z-line. In turn, FATZ is proposed to anchor calcineurin to theZ-line, and therefore may participate in regulating calcineurin activity (Frey et al.2000). Calcineurin signaling is an important step in initiating the hypertrophicresponse of muscle. Calcineurin can dephosphorylate and activate NF-AT3, aZ-line-associated transcription factor (reviewed in Olson et al. 2000, Olson &Williams 2000). Intriguingly, calcineurin,α-actinin, and FATZ can form a ternarycomplex, with FATZ acting as the bridge betweenα-actinin and calcineurin (Freyet al. 2000). This may provide region-specific regulation of calcineurin signaling;the Z-line may act as a “sink” for NF-AT until calcineurin becomes activated anddephosphorylates NF-AT. Recently, FATZ has been shown to interact with yetanother Z-line component, cypher (Frey & Olson 2002), which may link differentsignaling networks at the Z-line.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
668 CLARK ET AL.
MYOPALLADIN Myopalladin (Mr∼ 145 kDa) and the related protein, palladin, areIg-domain-containing proteins enriched at sites of actin filament anchorage (Parastet al. 2000, Bang et al. 2001b). The C-terminal domain of myopalladin that is con-served in palladin is responsible for its association withα-actinin. Myopalladininteracts with the SH3 domain of both nebulin and nebulette through its proline-richrepeat (between Ig-2 and Ig-3). Additionally, the N-terminal region of myopalladininteracts within the I-band with cardiac ankyrin repeat protein (CARP), a nuclearprotein involved in control of gene expression (Bang et al. 2000b). This associ-ation may have important functional significance because overexpression of theN-terminal region of myopalladin disrupts both Z-line organization and overall sar-comere structure in chick cardiac myocytes. This intriguing observation suggestsyet another connection between myofibrillar organization and gene expression.Linking the subcellular distribution of CARP to its functions may reveal a novelmechanism for regulating myofibrillogenesis.
Other Z-Line-Associated Proteins
FILAMIN Three distinct filamins (α, β, γ ) (Mr∼ 300 kDa) have been identifiedthat share a similar molecular structure: an N-terminal actin-binding head followedby 24 Ig-like domains.γ -Filamin (also known as ABP-280) is the muscle-specificfilamin isoform and differs from the two other proteins by insertion of a novelsequence in the twentieth Ig-repeat. This novel sequence may represent the Z-disctargeting sequence inγ -filamin because the corresponding region inα-filamin can-not localize to the Z-disc (van der Ven et al. 2000b). Filamin is capable of bindingto a number of other muscle proteins, includingδ- andγ -sarcoglycan (Thompsonet al. 2000), myotilin (a novel protein with Ig-repeats) (van der Ven et al. 2000b),β1D-integrin (Loo et al. 1998), and the FATZ-family of Z-line proteins (Faulkneret al. 2000, Takada et al. 2001). Based on these protein interactions, filamin maybe a critical link between the membrane and the sarcomeric cytoskeleton. Overex-pression of the Ig repeats of myotilin that bind filamin disrupts myofibril assemblyin C2C12 myogenic cells (van der Ven et al. 2000b), and different filamin iso-forms appear to be critical for specific aspects of myofibrillogenesis (van der Flieret al. 2002). Moreover, several filamin binding partners are mutated or implicatedin different forms of LGMD, suggesting they act together to stabilize the musclecytoskeleton (e.g., Hauser et al. 2000, Nigro et al. 1996). Filamin may have addi-tional signaling roles at the Z-line, as it binds to SHIP-2, an inositol polyphosphate5-phosphatase known to regulate insulin signaling (Dyson et al. 2001).
TELETHONIN/T-CAP Telethonin, (Mr 19 kDa; also known as titin cap or Tcap) wasindependently identified by several groups (Valle et al. 1997, Mues et al. 1998,Gregorio et al. 1998). Telethonin can bind to FATZ (Faulkner et al. 2000) andthe N-terminal Z1-Z2 domains of titin (Gregorio et al. 1998). Overexpression ofeither telethonin or the Z1–Z2 region of titin in cardiac myocytes disrupts Z-linestructure and sarcomere organization, suggesting that this interaction is critical

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 669
for sarcomeric structure. Therefore, telethonin may act as a “bolt” to anchor titinwithin Z lines (Gregorio et al. 1998). Moreover, telethonin may be phosphorylatedby the titin kinase region (located at the M-line) (Mayans et al. 1998). Mutationsin telethonin are responsible for LGMD type 2G (Moreira et al. 2000), provid-ing additional evidence for telethonin’s role in stabilizing the muscle sarcomere.Intriguingly, a direct interaction of cardiac muscle telethonin with the potassium(IKS) channelβ-subunit minK was recently reported. This interaction suggestsa T-tubule-myofibril linking system that may contribute to a stretch-dependentregulation of K+ flux (Furukawa et al. 2001).
OBSCURIN An interesting yet perplexing story recently has arisen from work inlaboratories studying a newly characterized Z-line titin-binding protein. This giantmolecule was named obscurin because its complexity, large size (Mr∼ 700–800 kDa), and low abundance hindered its characterization (Young et al. 2001).It is restricted to striated muscle and has 57 Ig/FN3 domains as well as signalingdomains (Bang et al. 2001a). Obscurin is homologous to theC. elegansproteinUnc-89, a giant protein implicated in the regulation of M-line assembly and struc-ture (Benian et al. 1996b). Obscurin, however, appears to localize to the Z-lineregion as well as to M-lines. Specifically, the titin Z-line repeats Z9–Z10 bind tothe obscurin Ig domains 48–49. Obscurin domains 48–49 localized at the Z-linein early development of chicken embryos, but at later stages of development, theylocalized at the M-line (Young et al. 2001). In another study, these obscurin do-mains were found in the Z-line region, likely via their interaction with a novel titinisoform, novex 3; thedistance from the Z-line varied with sarcomere length, sug-gesting that the novex-3 titin/obscurin complex formed an elastic Z-disc to I-bandlinking system. In this study, obscurin was also detectable at the M-line (Banget al. 2001a). Thus in different muscle fibers or developmental stages, obscurinlocalization and the signaling pathways it participates in may change. Additionalclues may be obtained soon because another obscurin-related protein has beenfound, human obscurin-MLCK, which contains two Ser/Thr kinase domains. Ithas been proposed that the obscurin-MLCK gene may encode several proteins inaddition to obscurin (Russell et al. 2002).
MURF-3 MURF-3 is the founding member of the muscle-specific Ring Finger pro-teins (MURFs), a new family of muscle cytoskeletal proteins. MURF-3 in skeletalmuscle cells localizes to the Z-line, but it is not known with which sarcomericcomponent(s) it interacts (Spencer et al. 2000). Interestingly, MURF-3 also bindsto and stabilizes microtubules in the presence of depolymerizing agents. Antisenseablation of MURF-3 expression in C2 skeletal muscle cells severely disrupted theinitiation of myogenesis (Spencer et al. 2000), indicating a critical role in muscledifferentiation. The interaction of MURF-3 with many binding partners (MURF-1,MURF-2, Z-line protein(s), and microtubules) indicates that it also may functionto link multiple cytoskeletal assemblies together including the Z-line, the M-lineregion of titin, and microtubules.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
670 CLARK ET AL.
INTERMEDIATE FILAMENT PROTEINS: LINKERS OFCYTOSKELETAL NETWORKS IN STRIATED MUSCLE
Intermediate filaments and their associated proteins tether together distinct cy-toskeletal assemblies and organelles (e.g., nuclei and mitochondria) in striatedmuscle, a key factor for generating force and maintaining mechanical stability.Structurally, the intermediate filament proteins are composed of a centralα-helicalrod domain flanked by N-terminal head and C-terminal tail regions. The proteinsassociate both side to side and head to tail to form homo- or hetero-polymerswhose expression patterns are highly regulated spatially and temporally. The de-tailed molecular interactions between intermediate filaments and other cytoskeletalstructures are only beginning to be elucidated.
Desmin (Mr∼ 52 kDa), the predominant intermediate filament protein of stri-ated muscle (Lazarides 1980), is a component of the Z-line region, costameres, themyotendinous junction, and intercalated discs. In mature muscle, desmin filamentslaterally interlink Z-discs, perhaps via their interaction with nebulin (Bang et al.2001b) and integrate the myofibrils with the sarcolemma, nuclei, mitochondria and,possibly, microtubules. Previously, desmin was proposed to have a critical role inearly myogenesis because ablation of its expression in mouse embryoid bodiesinhibited skeletal and smooth muscle development (Weitzer et al. 1995). Surpris-ingly, however, desmin null mice are viable and fertile, and the expression levels ofother known muscle intermediate filament proteins are not up-regulated (Li et al.1997, Carlsson et al. 1999). Nevertheless, the mice develop a postnatal, multi-system disorder characterized by muscle architecture disruption and degeneration.This phenotype involves a reduction of lateral myofibril alignment, increased myo-fibrillar mobility, perturbation of myofibril anchorage to the sarcolemma, interca-lated disc alterations, abnormal mitochondrial shape and positioning, and cardiacmuscle degeneration and necrosis (Li et al. 1997, Milner et al. 1999, Balogh et al.2002, Shah et al 2002). In one study utilizingdesminnull mice, the severity of mus-cle damage correlated with the degree of muscle usage, and it was hypothesizedthat the myofibrils were more susceptible to damage upon contraction (Li et al.1997). Paradoxically, however, a second study reported thatdesmin-null skeletalmuscles are actually less vulnerable to mechanical injury, perhaps because theabsence of a functional intermediate filament system decreased the production ofisometric stress (Sam et al. 2000). Therefore, the exact functional consequences ofablating desmin in muscle await further clarification. Clinically, several missenseand truncation mutations in the human gene lead to desmin myopathy, character-ized by the presence of intracytoplasmic desmin aggregates, myofibril fragility,and cardiac and skeletal muscle myopathies (Goebel 1995, Carlsson & Thornell2001). Furthermore, mutations inαβ-crystallin, a molecular chaperone involvedin desmin filament assembly, are implicated in certain muscle myopathies (Vicartet al. 1998). In conclusion, although desmin is not absolutely essential for musclefunction, it clearly contributes to maintaining the integrity and alignment of matureand regenerating myofibrils.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 671
Less is known about other intermediate filament proteins in striated muscle in-cluding vimentin, nestin, synemin, and paranemin. Vimentin (Mr∼ 55 kDa) is thepredominant intermediate filament protein expressed during striated muscle devel-opment and is lost upon maturation (Granger & Lazarides 1979). When expressed,it appears to have a distribution pattern similar to that of desmin. Vimentin nullmice have no obvious phenotype, even though no compensatory expression of otherintermediate filament proteins is detectable (Colucci-Guyon et al. 1994). Similar-ily, disruption of vimentin filaments by expression of a dominant-negative desminsubunit in transgenic mice did not reveal an obvious phenotype (Pieper et al. 1995).Although no known human myopathies are associated with vimentin mutations, itis highly expressed in regenerating fibers in many neuromuscular disorders (Misraet al. 1992). Nestin (Mr∼ 240 kDa) is expressed in myotendinous and neuromus-cular junctions, colocalizes with vimentin in developing and regenerating musclefibers, and is expressed with desmin at low levels in mature muscle (Sejersen &Lendahl 1993, Zimmerman et al. 1994). Like desmin and vimentin, nestin ex-pression is detected in regenerating muscle fibers in human myopathies, includingDuchenne’s muscular dystrophy (DMD) and myositis (Sjoberg et al. 1994). Syne-min (Mr∼ 230 kDa) and paranemin (Mr∼ 178 kDa), originally characterized asproteins that interact with desmin and vimentin, are members of the intermediatefilament superfamily with unusually large tail domains. Synemin and paraneminare both transiently expressed during striated muscle development, and paraneminexpression becomes restricted to adult heart, whereas synemin is expressed in alladult muscle (Price & Lazarides 1983, Bilak et al. 1998). Synemin’s rod domain in-teracts with desmin and vimentin, and its tail domain bindsα-actinin and vinculin.Thus synemin heteropolymeric filaments may directly or indirectly link adjacentmyofibrils, as well as link the contractile apparatus to costameres (Bellin et al.2001). In summary, vimentin and nestin are expressed during striated muscle de-velopment and are upregulated during fiber regeneration (e.g., in DMD), indicatingthey may participate in the assembly of muscle cytoarchitechture. Further inves-tigation of the functional characteristics of synemin and paranemin is required.
Exciting progress in the intermediate filament protein field recently has beenattained by the identification of other members of this superfamily that are enrichedin striated muscle. Two novel proteins, desmuslin and syncoilin, were identifiedas binding partners ofα-dystrobrevin, a peripheral protein of the sarcolemmaldystrophin-glycoprotein complex (DGC), and both also colocalize with desminat the level of Z-lines (Mizuno et al. 2001, Newey et al. 2001, Poon et al. 2002).Future functional studies on these proteins are expected to yield valuable insightinto the establishment and maintenance of myofibril integrity because they ap-pear to directly link the intermediate filament system with the DGC. In addi-tion to these novel proteins, cytokeratins also are expressed in muscle. Althoughinitially reported to be suppressed during development, they recently have beendetected in adult muscle. Cytokeratins appear to be concentrated near the sar-colemma at costameres in both normal anddesmin-null skeletal muscle. Theseintermediate filament proteins are hypothesized to cooperate with desmin and its

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
672 CLARK ET AL.
associated molecules in organizing and stabilizing the myoplasm and the sar-colemma (Kosmehl et al. 1990; J. O’Neill, submitted).
Lastly, specialized nuclear intermediate filament proteins and their associatedproteins have been the subjects of recent intense investigation because of theirunexpected contributions to muscle cytoarchitecture and function. The nuclearlamins are the main components of a meshwork underlying the inner nuclearmembrane and likely act as attachment sites for intranuclear proteins and chromo-somes and may stabilize the nuclear envelope in most cell types. Multiple laminisoforms are expressed in muscle and are referred to as type-A (lamins A andC) and type-B (lamins B1 and B2). Emerin (Mr∼ 34 kDa) is a widely expressedprotein that binds to lamins A and C (Bione et al. 1994). Strikingly, an X-linkedform of Emery-Dreifuss muscular dystrophy (X-EDMD) results from the absenceof emerin, and an autosomal-dominant form (AD-EDMD) is caused by variousmissense mutations in lamins A and C (see Morris 2001 and citations within).These disorders are characterized by muscle weakness and conduction defects incardiac muscle with sudden heart failure (Emery 2000). Type-A lamin mutationsalso cause DCM and LGMD type-1B (Morris 2001). Although the mechanisms bywhich mutations in these genes result in muscle myopathies remain elusive, oneidea is that mechanical stress in muscle damages the weakened nuclear envelope(Morris 2001). Emerin may have other roles in muscle because it also is present inintercalated discs (Cartegni et al. 1997). Genetic models are expected to provideinsight into these issues because an insertional mutation in theDrosophilalamingene Dm0 results in impaired locomotion, and mice lacking type-A lamins developa disorder reminiscent of EDMD (Lenz-Bohme et al. 1997, Sullivan et al. 1999).The study of novel proteins that interact with emerin and lamins should aid in dis-cerning these questions. In this regard, Myne-1, a myocyte-specific inner nuclearmembrane protein expressed early in muscle differentiation, interacts with laminA/C and may be a mediator of muscular myopathies (Mislow et al. 2002).
MICROTUBULES: EMERGING ROLES IN STRIATEDMUSCLE MORPHOLOGY AND FUNCTION
Microtubules are involved in numerous cellular processes including intracellulartransport, positioning of organelles, cell motility, and mitosis. In contrast to theother cytoskeletal filament systems, relatively little is known about the roles ofmicrotubules in striated muscle. However, mounting evidence implicates them instriated muscle differentiation, morphology, and contractile activity. Microtubulesare polymers of heterodimericα andβ tubulin subunits (Mr∼ 55 kDa each), ofwhich there are several isoforms whose expression patterns are highly regulated.They comprise a diverse array of both stable and transient filaments, dependingon the functional requirements of the particular cell type.
In skeletal muscle, microtubules are positioned between myofibrils at the levelof the A-I junction, as well as associated with the sarcolemma, the Golgi complex,and nuclei. This suggests that microtubules participate in mechanical integration ofvarious organelles in striated muscle. Interestingly, the predominance of transverse,

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 673
longitudinal, or oblique orientations of microtubules varies among different muscletypes (Kano et al. 1991, Boudriau et al. 1993), perhaps correlating with the dif-fering functional requirements of muscle. Disruption of microtubule dynamics bytreatment of skeletal myocytes with depolymerizing or stabilizing agents perturbsevents that occur during myogenesis, particularly myoblast fusion and differentia-tion (Antin et al. 1981, Saitoh et al. 1988). Although the mechanisms responsiblefor the disruption of myogenesis remain unclear, insight may be provided by thediscovery of novel muscle microtubule-associated proteins. In particular, MAP4and MURF-3 (which colocalizes with both microtubules and sarcomeric Z-lines),have critical roles in myotube formation, perhaps acting as microtubule stabilizingand signaling components (Mangan & Olmsted 1996, Spencer et al. 2000). Addi-tionally, the interaction of muscle M-cadherin with skeletal muscle microtubulesis essential for myoblast fusion, perhaps by regulating events that occur at the my-ocyte membrane (Kaufmann et al. 1999). Interestingly, perturbation of microtubuledynamics in postmitotic skeletal myoblasts affects their morphology and disruptsmyofibrillogenesis: Their A-bands contain thick filaments interdigitated with longmicrotubules; their I-bands contain microtubules but lack thin filaments (Antinet al. 1981). The clinical importance of regulated microtubule dynamics also ishighlighted by the observation that some cancer patients treated with colchicinedevelop “colchicine myopathy,” a disorder characterized by muscle weakness andmyofibril disarray (e.g., Fernandez et al. 2002).
In heart muscle, disruption of microtubules perturbs contractile function, andmicrotubule levels significantly increase in pressure-overload models of cardiachypertrophy (e.g., Klein 1983, Tsutsui et al. 1993; see Hein et al. 2000 for review).How microtubules are linked to contractile activity is unclear, but disruption ofcardiac microtubule arrays perturbs Ca2+ signaling, cellular stiffness and viscosity,myofibrillogenesis, gene expression, and transport of certain sarcomeric mRNAtranscripts (Perhonen et al. 1998, Takahashi et al. 1998, Kerfant et al. 2001).Investigation into the function of microtubules in these cellular processes maybe challenging because it appears that not all microtubule arrays are the same.In this regard, new muscle isoforms of tubulin have been identified (e.g., Stanchiet al. 2000). Additionally, different posttranslationally modified populations ofmicrotubules are present during cardiac development, and a specialized subsetof microtubules that are resistant to depolymerization are sufficient for normalcontractile activity (Webster & Patrick 2000). Although many questions remainconcerning striated muscle microtubules, it is clear that these cytoskeletal elementshave surprisingly diverse functions.
MEMBRANE LINKAGES TO THE CYTOSKELETON:ANCHORAGE, FORCE TRANSMISSION, ANDMECHANOSIGNAL TRANSDUCTION
To effectively transmit force, the contractile cytoskeleton must be tethered to thesarcolemma, the cellular membrane enveloping the myofibrils. Striated musclecontains specialized membrane linkages known as costameres that anchor Z-lines

23 Sep 2002 10:33 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
674 CLARK ET AL.
and M-lines to the membrane. Costameric structures are also present parallel tothe long axis of the muscle, but their frequency is highly variable. Skeletal musclemyofibrils terminate at the myotendinous junction, a second type of membranelinkage that bears some structural and molecular similarities to the costameres.In contrast, the physiology of the heart dictates a very different terminal anchor-age system. Cardiac myocytes form end-to-end attachments known as intercalateddiscs that serve both as anchorage sites for the cytoskeleton and to electricallycouple adjacent myocytes. Each type of membrane linkage (i.e., costamere, my-otendinous junction, and intercalated disc) is described in more detail below, withan emphasis on their unique properties (Figure 4). This section also includes a dis-cussion of a newly appreciated function of the muscle cytoskeleton in organizingion channels and other membrane-bound receptors.
Costameres: Integrins, Spectrin, andthe Dystroglycan Complex
Striated muscle is surrounded by a basement membrane rich in extracellular matrix(ECM) proteins (e.g., collagen, laminin). This matrix acts as an adherent substra-tum and provides structural stability to the cell. The contractile apparatus attachesto the basement membrane at periodic membrane-associated plaques (costameres)that co-distribute with the Z-line (Pardo et al. 1983b) and the M-line (Porter et al.1992). Costameres coordinately transduce contractile force from the Z-line to thebasement membrane, where the force is transmitted laterally to the muscle termini(Danowski et al. 1992). In addition, costameres are organizational points for themembrane cytoskeleton, which maintains the structural integrity of the membraneduring contraction. These features are a result of at least three different cytoskeletalnetworks (integrins/focal adhesion complexes, the dystroglycan complex, and thespectrin-based cytoskeleton) acting in concert to promote the stable linkage of themyofibril to the muscle membrane.
Integrins are heterodimeric transmembrane proteins that mediate attachmentof the actin cytoskeleton to the ECM. The large extracellular domain of the in-tegrin subunits directly binds ECM components (Adams & Watt 1993), whereasthe cytoplasmic domains interact with the actin cytoskeleton through associa-tions with other proteins such as talin (Mr∼ 225 kDa), vinculin (Mr 116 kDa)andα-actinin (reviewed in Calderwood et al. 2000). In vertebrates, there are 18α (Mr∼ 150 kDa) and 8β (Mr∼ 90 kDa) subunits that can heterodimerize into atleast 24 distinct isoforms, each possessing unique properties (for a recent reviewsee Bouvard et al. 2001); thus ECM binding and the repertoire of cytoplasmic-associated proteins is dictated by the heterodimer isotype.β1 integrin is enrichedat costameres (Bozyczko et al. 1989, Belkin et al. 1986) along with vinculin (Pardoet al. 1983a) and talin (Belkin et al. 1986). Functional studies of theβ1 integrinsubunit indicate a critical role for integrins in maintaining the organization of sar-comeres in cardiomyocytes (Fassler et al. 1996) andDrosophilamuscle (Volk et al.1990).

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 675
In addition to cytoskeletal attachment, integrins have a well-established role assignaling molecules (Giancotti & Ruoslahti 1999). This dual function allows themto sense mechanical stress and activate intracellular signal transduction pathways,leading to changes in gene transcription and cytoskeletal reorganization (reviewedin Shyy & Chien 1997). In striated muscle, their prominent location at costameresmakes them ideal candidates for acting as biomechanical sensors that respond tochanges in load, an idea supported by recent evidence. For instance, overexpressionof β1 integrin in cardiomyocytes induces hypertrophic gene expression, whereasexpression of an inhibiting form ofβ1 integrin blocks hypertrophic responses invitro (Pham et al. 2000). Furthermore, focal adhesion kinase (FAK;Mr∼ 125 kDa),an important integrin effector, is enriched with integrins at the myotendinous junc-tion (Baker et al. 1994) and co-distributes with costameric integrins at the Z-line(Kovacic-Milivojevic et al. 2001). FAK is phosphorylated (and thus activated) bymechanical stretch (Kovacic-Milivojevic et al. 2001) and expression of a dominantform of FAK abrogates the normal hypertrophic response in cultured cardiomy-ocytes (Kovacic-Milivojevic et al. 2001, Pham et al. 2000). These data implicateFAK and integrins in functioning as a sensory/signaling complex in muscle. In-triguingly, the integrin response to mechanical load changes may involve morethan simply an initiation of chemical signals. Because integrins are physicallyconnected to the cytoskeleton, they may also induce conformational changes invarious Z-line components. A clear example of this behavior was elegantly demon-strated for force-dependent clustering of focal adhesion proteins upon cytoskeletaldeformation (Sawada & Sheetz 2002). Although speculative, this may provide anadditional mechanism by which Z-line-associated signaling molecules are acti-vated (e.g., FAK) or triggered to translocate to the nucleus (e.g., NF-AT3).
Dystrophin (Mr 427 kDa) is the core component of a multimeric protein com-plex known as the dystroglycan complex (DGC) that includes theα/β-dystroglycantransmembrane receptor (which binds laminin) and the sarcoglycans (reviewed inWinder 2001). Mutations in dystrophin are responsible for DMD, a fatal disordercharacterized by a severe muscle weakness and degeneration (Hoffman et al. 1987).Themdxmouse also results from a dystrophin mutation (Sicinski et al. 1989), andprovides a model system in which to study dystrophin function and treatmentsfor this debilitating disorder. Dystrophin mediates the critical connection betweenthe surrounding extracellular matrix, the muscle membrane, and the underlyingmembrane cytoskeleton by linkingβ-dystroglycan to costameric actin (see Figure4). In the absence of dystrophin, costameric actin is no longer tightly associatedwith the muscle membrane (Rybakova et al. 2000); this results in a weakening ofthe sarcolemma, making it susceptible to damage during contraction (Beam 1988,Petrof et al. 1993). Muscle biopsies from DMD patients andmdx mice have adramatic reduction of the sarcoglycans and certain other DGC components at thesarcolemma, indicating a critical role for dystrophin in maintenance of the DGCat the muscle membrane (Ervasti et al. 1990, Ohlendieck & Campbell 1991). Inaddition, dystrophin is important for the organization of other costameric com-ponents such as vinculin and spectrin (Williams & Bloch 1999). Together, these

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
676 CLARK ET AL.
results demonstrate a fundamental role for dystrophin in organizing the musclemembrane cytoskeleton and its associated transmembrane components.
Dystrophin may execute some of these cellular activities in conjunction withintegrins. Theα7β1 integrin complex is up-regulated in both DMD patients andmdxmice (Hodges et al.1997), indicating a possible mode of compensation byintegrins. Furthermore, enhanced expression ofα7β1 integrin inmdxmice partiallyameliorates the dystrophic phenotype (Burkin et al. 2001). These observationsstrongly indicate that integrins and the DGC cooperate in maintaining the integrityof the muscle cytoskeleton.
Other components of the DGC also have critical roles in stabilizing the musclecytoskeleton. The sarcoglycans (α, β, δ, andγ ) are a set of single-pass trans-membrane proteins that associate withβ-dystroglycan (reviewed in Gordon &Hoffman 2001). Individual sarcoglycans are novel proteins, but interestingly theydo bear some similarity to each other in that they are all small glycosylated proteinswith extensive extracellular domains. Although their function is unknown, geneticlesions in each of the four sarcoglycans can produce different forms of LimbGirdle MD (Gordon & Hoffman 2001). Similar to DMD, mutations in any of thesarcoglycans destabilize the DGC, resulting in a reduction of certain DGC com-ponents at the muscle membrane, including the other sarcoglycans. The BIO14.6CM hamster is a model for cardiomyopathy and results from a spurious mutationin δ-sarcoglycan (Nigro et al. 1997, Sakamoto et al. 1997). Recently, this animalmodel system has been used to test high-efficiency transfer techniques of saro-clycan genes, which restore cardiac function. Thus this approach could lead totherapeutic strategies for treating various muscle myopathies (Ikeda et al. 2002,Kawada et al. 2002).
Additional components of the DGC includeα-dystrobrevin and the syntrophins.These molecules may function as adaptor molecules that link other proteins tothe DGC.α-Dystrobrevin and the syntrophins are especially intriguing as theyprovide a connection between dystrophin and nitric oxide signaling. Syntrophindirectly binds bothα-dystrobrevin and dystrophin, and its C-terminal PDZ-domainassociates with neuronal-type nitric oxide synthase (nNOS) (Blake et al. 2002).Muscle samples from DMD patients andmdxmice show a loss of nNOS from mus-cle membranes and a subsequent reduction in nNOS catalytic activity normallypresent at the sarcolemma (Brenman et al. 1995). nNOS also is displaced from thesarcolemma in mice that lack functionalα-dystrobrevin orα-syntrophin (Gradyet al. 1999, Kameya et al. 1999). Because nitric oxide synthesized from endothelialcells is a major vasodilator, muscle nNOS may regulate activity-dependent bloodflow in muscle. The reduction in NOS activity potentially contributes to the dys-trophic phenotype seen in loss of dystrophin and in theα-dystrobrevin null mice.This story grows even more complex, however, with the reports thatα-dystrobrevinalso binds the intermediate filament proteins syncoilin and desmuslin (Mizuno et al.2001, Newey et al. 2001). Thus the phenotype that results from mutations in dys-trophin, as well as its associated proteins, is probably the effect of perturbationof several critical biological processes. This idea reflects the intricacies of thisimportant protein complex.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 677
Finally, although spectrin is prominently enriched in costameres (Porter et al.1992), its function in this junction has not been explored extensively. Spectrinis best known for organizing the membrane cytoskeleton of erythrocytes and,in conjunction with ankyrin, also plays a prominent role in anchoring integralmembrane proteins to the membrane cytoskeleton (reviewed in Thomas 2001). Instriated muscle,α/β spectrin heterodimers are found at Z-line costameres, whereasβ homodimers are enriched at M-lines and the longitudinal strands (Porter et al.1997). Both populations of spectrin are complexed with ankyrinG, which maylimit the spectrin cytoskeleton to the sacrolemma by binding the Na,K-ATPase(Williams et al. 2001). InC. elegans, loss of spectrin revealed its requirement forstabilizing the contractile apparatus (Hammarlund et al. 2000, Moorthy et al. 2000).Interestingly, this occurred in the absence of any noticeable membrane damage,indicating that spectrin’s involvement was not in maintaining membrane integrity,but instead it may play a more direct role in anchoring the contractile apparatusto the muscle membrane. In vertebrates,β-spectrin can bind to MLP, which, inturn, binds toα-actinin and is associated with the Z-line (Flick & Konieczny 2000,Louis et al. 1997). This provides an attractive model whereby spectrin acts as alink between the muscle membrane and the Z-line though MLP.
The Myotendinous Junction: Site of ForceTransduction in Skeletal Muscle
The myotendinous junction (MTJ) is a unique structure in skeletal muscle thatconstitutes the termination point of the myofibrils. The MTJ is specially reinforcedbecause it is the focal point for much of the force transmitted during contraction.At the muscle termini, the cell membrane is highly invaginated and interdigitatedwith the surrounding tendon matrix, increasing the surface area of the MTJ andthereby reducing stress on the membrane. The thin filaments terminate at a densesubplasmalemmal layer that appears to mediate thin filament attachment to eachother and to the muscle membrane (reviewed in Tidball 1991). The dystrophin-based membrane/cytoskeletal linkage system and focal-adhesion-like adhesionplaques containing integrins, vinculin, and talin are found at the MTJ, suggestingthat both complexes participate in connecting the terminal thin filaments to themuscle membrane.
Two reports analyzing the MTJ in themdxmouse (which lacks dystrophin) havereached different conclusions concerning the effect of the absence of dystrophinon the MTJ. One study reported that the thin filaments remain associated with theMTJ in themdxmouse, although some necrosis occurs at the junction (Miosgeet al. 1999). The other study found that MTJs ofmdxmice do not associate betweenthe thin filaments and the membrane (Tidball & Law 1991). The reasons for thesedifferences are not clear, thus the role of dystrophin at the MTJ remains elusive.
Theα7β1 integrin complex is present throughout the muscle membrane but isenriched at the MTJ (Bao et al. 1993). Loss of theα7 subunit produces a formof congenital myopathy in both mice and humans (Hayashi et al. 1998, Mayeret al. 1997). Ultrastructural studies of mice with a targeted disruption of theα7

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
678 CLARK ET AL.
integrin gene demonstrated a critical role for integrins in maintaining the linkbetween the myofilaments and the muscle membrane at the MTJ. By nine monthsof age, the MTJ in theα7-/- mice lose their high degree of membrane foldingconcomitant with retraction of the myofilaments from the membrane (Miosgeet al. 1999). Both vinculin and talin are enriched at the MTJ withα7 integrin andmay link the integrins and the thin filaments (Shear & Bloch 1985, Tidball et al.1986). However, it is not clear if a talin/vinculin-based linkage to the thin filamentswould be strong enough to withstand the forces of the MTJ (Tidball 1991). N-RAP(Mr∼ 133 kDa), a nebulin-related-anchoring protein, is found exclusively at theMTJ in skeletal muscle and intercalated discs in cardiac muscle and may facilitatethe association of thin filaments to these membrane junctions (Luo et al. 1997).The C terminus of N-RAP has significant sequence similarity to nebulin’s superrepeats and can bind actin, whereas the very N terminus encodes a LIM domain(Luo et al. 1997, 1999). N-RAP interacts with talin through its LIM domain andmay associate with vinculin at its C terminus (Luo et al. 1999). The potentialfor multiple interactions between integrins, vinculin, N-RAP, talin, and actin atthe MTJ may provide a stable, mechanical link between the membrane and theactin-based cytoskeleton.
Striated muscles inDrosophila melanogasteralso terminate at a MTJ that dis-plays many similarities to their vertebrate counterpart. Elimination of integrinfunction does not prevent MTJ formation, but the muscle cells detach upon con-traction, indicating a critical role for integrins in stabilizing this junction (reviewedin Brown et al. 2000). Studies inDrosophilaalso have indicated a role for twointegrin-associated proteins in stabilizing the attachment of actin to integrins atthe MTJ. Mutations in either integrin-linked kinase (ILK;Mr∼ 50 kDa) or PINCH(particularly interesting new cysteine histidine protein;Mr∼ 31 kDa), which areenriched at the fly MTJ (Zervas et al. 2001; K.A. Clark & M.C. Beckerle, un-published observations), result in the destabilization of the actin filaments at thejunction. Neither molecule has been shown to directly bind actin, so the mechanismthat stabilizes the MTJ is unknown.
Intercalated Discs: Terminal Anchors in Cardiac Muscle
Intercalated discs of cardiomyocytes contain three main junctional complexes: gapjunctions, which chemically couple neighboring cardiomyocytes; fascia adherens,which connect the actin cytoskeleton to the membrane; and desmosomes, which at-tach intermediate filaments to the muscle termini. N-cadherin (Mr∼ 135 kDa) is theprimary receptor at fascia adherens that links the membrane to the contractile ap-paratus (Volk & Geiger 1986). Cadherins are single-pass transmembrane proteinsthat mediate homotypic interactions between adjacent cells. Their cytoplasmic tailsserve as a scaffold to assemble a multiprotein linkage to the actin cytoskeleton. TheC-terminal 25 amino acids bind either plakoglobin (Mr∼ 82 kDa; also known asγ -catenin) orβ-catenin (Mr∼ 88 kDa). Plakoglobin andβ-catenin fulfill the samerole at facia adherins as a bridging factor between the cadherins (Mr∼ 102 kDa),the main linkage between the actin cytoskeleton and the cadherin complex.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 679
α-Catenin can mediate a link to actin directly or indirectly via interactions withthe actin binding proteinsα-actinin and vinculin (Steinberg & McNutt 1999).
Disruption of N-cadherin function with inhibitory antibodies (Soler & Knudsen1994), or by expression of a dominant-negative N-cadherin in cultured cardiomyo-cytes (Hertig et al. 1996), dissociates the intercalated discs between cells andalso disrupts myofibrillar organization. Mutations in metavinculin, a cardiac andsmooth muscle-specific vinculin isoform, are associated with DCM in humans(Olson et al. 2002), and cardiomyocytes that harbor metavinculin mutations dis-play perturbed intercalated disc structure (Maeda et al. 1997, Olson et al. 2002).Together, these studies indicate the importance of the cadherin/catenin complex inboth cell adhesion and anchorage of the contractile cytoskeleton to the intercalateddisc.
Other components of the intercalated disc are implicated in stabilizing theattachments of the myofibrils to the muscle termini. N-RAP is found at the in-tercalated disc and could play a similar role at this junction as at the MTJ (Luoet al. 1997). Functional elimination of the muscle LIM protein, MLP, in the mousedisrupts intercalated disc morphology, and this defect is probably responsible, inpart, for the development of DCM in these animals (Arber et al. 1997, Ehler et al.2001). Interestingly, MLP and N-RAP can associate in vitro, and thus probablyparticipate together in stabilizing the intercalated disc.
Desmosomes: Intermediate Filaments and AssociatedProteins Form Links to the Membrane
Desmosomes are specialized cadherin-based adherens junctions that link the in-termediate filament system (e.g., desmin) to the membrane (reviewed in Greenet al. 1998). These junctions form a three-dimensional scaffolding that is espe-cially prominent in tissues subjected to mechanical stress, such as heart and skin,implicating intermediate filaments and associated proteins in the maintenance ofstructural integrity. The desmososmal complex contains three main componentssimilar to the cadherin complex at fascia adherins: desmosomal cadherins anchoredat the cell membrane (which differ from classical cadherins by their attachmentto intermediate filaments, rather than actin filaments); a bridging protein, eitherplakoglobin or plakophilin (Mr∼ 86 kDa) that associates with the cytoplasmic tailof the cadherins; and desmoplakin (Mr∼ 300 kDa). Desmoplakin is a member ofthe plakin family, molecules that connect cytoskeletal elements to each other andto junctional complexes in many cell types (see Leung et al. 2002 for review).
Although desmosomes have been investigated mainly in epithelium, there isaccumulating evidence for their importance in maintaining the cellular integrityof cardiomyocytes. For example, cardiomyocytes that lack plakoglobin are devoidof structurally normal desmosomes, and plakoglobin−/− mice die from cardiacrupture (Ruiz et al. 1996). Also, a C-terminal truncation of human plakoglobinresults in Naxos disease, a rare condition characterized by cardiac arrythmia andheart failure (Norgett et al. 2000, McKay et al. 2000). Desmoplakin-deficientembryos die early in development, but rescue of this extraembryonic requirement

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
680 CLARK ET AL.
for the protein results in a cardiac defect similar to that exhibited by plakoglobin−/−mice (Gallicano et al. 2001). Both plakoglobin and desmoplakin are expressedin skeletal muscle and likely participate in intermediate filament anchorage in thattissue as well. This issue is complex because intermediate filaments may associatewith integrin-based adhesion plaques via interactions with other proteins, includingvinculin.
Finally, it is important to note that other proteins are important for cytoskeletallinkages to membranes, to each other, and to organelles. A prime example is thegiant protein plectin (Mr∼ 500 kDa), another member of the plakin family ex-pressed in a wide variety of cell types (Foisner & Wiche 1991). Plectin appearsto be a versatile cross-linker because it interacts with intermediate filaments, actinfilaments, microtubules, and integrins (see Leung et al. 2002 for review). In striatedmuscle, plectin colocalizes with desmin at the periphery of Z-discs, in intercalateddiscs, myotendinous junctions, and costameres (Wiche et al. 1983, Hijikata et al.1999). Strikingly, the association of plectin with Z-discs appears to be a prereq-uisite for the formation of the intermyofibrillar desmin cytoskeleton in culturedhuman skeletal muscle cells (Schroder et al. 2000). Plectin also may aid desminin positioning muscle mitochondria and linking them to myofibrils (Reipert et al.1999, Milner et al. 2000). Plectin null mice die 2–3 days after birth, exhibitinga phenotype reminiscent of human epidermolysis bullosa simplex (EBS)-MD, ahereditary skin blistering disorder with MD caused by plectin gene mutations(Gache et al. 1996, Andra et al. 1997). The plectin−/− mice exhibit disruptedmyofibrils and sarcolemmae and a disintegration of their intercalated discs (Andraet al. 1997). These data demonstrate that plectin has a role in maintaining theintegrity of skeletal and heart muscle architecture.
Ankyrins: Organizer of Specialized Membrane Domains
Another field that has made significant progress in the last few years is elucidat-ing the role that ankyrins have in organizing the sarcolemma in relation to thecontractile apparatus. Ankyrins are a family of peripheral membrane proteins thatinteract with a diverse range of integral proteins, including ion channels and pumpsand cell adhesion molecules, and immobilize them to the cortical spectrin-basedmembrane skeleton. All three ankyrin genes (Ank1–3) are expressed in cardiac andskeletal muscle. These genes give rise to a wide array of protein isoforms rangingfrom 17 to 440 kDa as a result of alternative mRNA processing. Based on theirlocalizations, ankyrins could play a role in promoting costamere-sarcolemmal as-sociations, as well as establishing or maintaining organization of the sarcolemmawith respect to the contractile apparatus. For example, ankyrinG (Ank 3gene prod-uct) may play a role in establishing and maintaining localized concentrations ofvoltage-gated sodium channels at the post-synaptic neuromuscular junction andsarcolemma (Wood & Slater 1998, Kordeli et al. 1998). AnkyrinG also participatesin the spectrin complex that anchors the Na,K-ATPase at costameres in skele-tal muscle (Williams et al. 2001). The localization of ankyrins to triads (Flucheret al. 1990), the junctions between T tubules and the sarcoplasmic reticulum (SR),

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 681
suggests other roles for ankyrins in excitation-contraction coupling beyond control-ling sodium channel distribution. This is evident in recent studies of the ankyrinB
(Ank2) knock-out mouse (Tuvia et al. 1999). Cardiomyocytes derived from thesemice exhibit reduced contraction rates and mislocalization of the IP3 and ryanodinereceptors within the SR. Transfection of ankyrinB but not ankyrinG rescues thesecells, demonstrating that these highly homologous ankyrin molecules have non-overlapping functions. Sodium channel densities are also affected in cardiomyo-cytes from the ankyrinB mouse, and intriguingly these channels exhibit changesin their inactivation kinetics (Chauhan et al. 2000). Furthermore, small unconven-tional Ank1 isoforms are concentrated in the SR at sites surrounding the Z- andM-lines of myofibrils, suggesting that these ankyrins link the SR to the contrac-tile apparatus (Zhou et al. 1997). These data suggest that ankyrin and its ligandsplay a significant role in maintaining specialized membrane domains in striatedmuscle.
COMMUNICATION BETWEEN THE SARCOMEREAND THE NUCLEUS: IMPLICATIONS FOR GENEREGULATION BY CYTOSKELETAL COMPONENTS
An emerging concept in cell biology involves the recognition that cytoskeleton-associated proteins often display dynamic distributions within cells and can partic-ipate in signal transduction cascades. Several sarcomere-associated proteins havebeen reported to reside in the nucleus, where they could participate in the responseof the cell to changes in the structure or function of the contractile machinery. Thesemolecules may serve as molecular messengers that enable muscle cells to mount ef-ficient physiological response to muscle stress, load requirements, and/or stretch.For example, the transcription factor NF-AT3 exhibits dual localization at theZ-line and nucleus of skeletal muscle. Its nuclear translocation is activity de-pendent in mature fibers and also is developmentally regulated (Liu et al. 2001).NF-AT3 is probably tethered to the Z-line though its interaction with calcineurin, aCa2+-dependent phosphatase that can also dephosphorylate NF-AT3; this activatesNF-AT3 and allows it to enter the nucleus (Frey et al. 2000, Olson et al. 2000).NF-AT transcription factors and calcineurin are well-characterized mediators ofhypertrophy in both skeletal and cardiac muscle (reviewed in Olson et al. 2000),indicating that this sequestering mechanism is likely to be critical for normal cellfunction.
Another sarcomeric component, originally identified as human C-193 (Chuet al. 1995), rodent cardiac ankyrin repeat protein (Zou et al. 1997) and cardiacadriamycin-responsive protein (CARP) (Jeyaseelan et al. 1997), localizes to theI-band and to the the nucleus. Evidence for CARP (Mr∼ 37 kDa) as a gene reg-ulator includes its interaction with the ubiquitous transcription factor YB-1, andits negative effect on the expression of specific cardiac genes (Jeyaseelan et al.1997, Zou et al. 1997). CARP mRNA levels are down-regulated during heart de-velopment but are induced dramatically early in hypertrophy (Baumeister et al.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
682 CLARK ET AL.
1997, Aihara et al. 2000). Additionally, although CARP levels are relatively low innormal skeletal muscle, they greatly increase upon denervation (Baumeister et al.1997). These data implicate CARP as a component of both muscle developmentaland stress-related pathways. Intriguingly, CARP also is a myopalladin-bindingpartner. In fact, the interaction of CARP with myopalladin appears to be criticalfor sarcomeric integrity in chick cardiac myocytes (Bang et al. 2001b). Recently,the human Arpp protein was characterized as a homologue of mouse Ankrd, aprotein very similar to CARP that is involved in the mechanical stretch response inskeletal muscle (Kemp et al. 2000, Pallavicini et al. 2001, Moriyama et al. 2001).Thus CARP may be the founding member of a gene family involved in myofibrilstructure, gene regulation, and myocyte stress response pathways.
Two titin-interacting proteins, the muscle-specific calpain p94 and MURF-1,also may participate in linking nuclear and sarcomeric functions. p94 has twotitin-binding sites, one within the I band and another in the M-line, as well asa nuclear localization sequence.URF-1 binds to titin’s M-line region and is alsoa nuclear protein that interacts with glucocorticoid modulatory element bindingprotein-1 (GMEB-1) (McElhinny et al. 2002), a transcription factor whose activityis regulated by cellular glucocorticoid levels (e.g., Zeng et al. 1998, Theriault et al.1999). Based on their unique properties including their interactions with titin, bothp94 and MURF-1 are hypothesized to be involved in coordinating stress responsepathways within striated muscle.
Other nuclear-shuttling proteins may participate in gene regulation indirectly bypost-transcriptional mechanisms (raver1) or by affecting nuclear structure (myo-podin). Raver1 encodes an RNA-binding protein (Huttelmaier et al. 2001) thatassociates with vinculin andα-actinin in a ternary complex and colocalizes withthese proteins at costameres and other sites of actin filament anchorage. Raver1 isnuclear in undifferentiated myoblasts and becomes associated with costameres interminally differentiated C2C12 skeletal myogenic cells (Huttelmaier et al. 2001).In addition to the cytoskeletal proteins mentioned above, raver1 is complexed withother RNA-binding proteins and may be part of a tissue-specific processing ofmRNAs important for striated muscle function. Similar to raver1, the novel actin-binding protein myopodin exhibits a dynamic localization coincident with differentstages of muscle differentiation (Weins et al. 2001). Its dynamic localization prob-ably reflects a signaling role because myopodin can translocate from the Z-lineto the nucleus upon stress. Overexpression of myopodin in undifferentiated cellsresults in the formation of nuclear actin bundles that also contain myopodin. Thepurpose of this bundling property is not clear, but recent experiments have demon-strated critical and unexpected functions for actin in nuclear transport (Hofmannet al. 2001) and in modulating properties of the nuclear endoskeleton (Wasser &Chia 2000).
These examples illustrate several potential mechanisms by which communi-cation between the contractile machinery and the nucleus may be mediated bysarcomeric constituents. Such a link would be expected to be critical for signalingrequired for maintenance of muscle homeostasis as well as for muscle plasticity.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 683
The sarcomere-association of molecules with nuclear roles may provide a mech-anism for the cell to couple cytoarchitectural integrity to nuclear response. Theability of the muscle cell to respond to mechanical stress and other physiologicalsignals is clearly essential for optimal muscle function. It is intriguing that the sar-comere may have a mechanism for signaling and feedback, not unlike that whichexists for organelles such as mitochondria and the endoplasmic reticulum.
CONCLUSIONS
Diverse cytoskeletal networks and their associated regulatory molecules are es-sential for efficient muscle contraction, as well as for most motile processes innonmuscle cells. In this review, we have discussed numerous examples from manymodel systems that demonstrate the exquisite molecular and developmental reg-ulation involved in creating the cytoskeletal assemblies of striated muscle. To-gether, these intricate molecular complexes ultimately define the differentiatedmorphology (e.g., different sarcomeric layouts) and function of distinct muscletypes. These studies also demonstrate the power of striated muscle as a modelsystem for investigating more general mechanisms that integrate cytoskeletal net-works in many eukaryotic cells since numerous structural, regulatory, and sig-naling molecules expressed in muscle also are expressed in nonmuscle cells. Italso is exciting to witness the emergence and growth of new areas of researchthat have sprung from the muscle field, including the study of signaling moleculesand pathways linking cytoskeletal architecture to gene expression and the cellularstress response. Finally, the realization that functional mutations in most (if notall) sarcomeric and many cytoskeletal components lead to various myopathies em-phasizes their importance in normal muscle function. It also continues to demon-strate that striated myocytes are intricately balanced units that depend on the in-terworkings of all their cytoskeletal networks for proper function. In summary,investigating the properties of striated muscle provides refreshing and intrigu-ing insights into cytoskeletal interactions and dynamics, signaling pathways, andfunctions.
ACKNOWLEDGMENTS
We are particularly thankful to A. Geach for preparing the figures; ElisabethEhler, Robert Bloch, Velia Fowler, Siegfried Labeit, and Ju Chen for criticalreading of the review; and K. Vaughan and S. Lambert for helpful discussions.A.S. McElhinny and K.A. Clark are postdoctoral fellows of the Desert Moun-tain and Western States Affiliates of the American Heart Association, respectively.This work was supported by the National Institutes of Health (HL57461, HL03985,HL63926 to C.C. Gregorio and HL60591 to M.C. Beckerle) and the HuntsmanCancer Foundation (M.C. Beckerle). Because of the breadth of the subject matterdiscussed in this review and the accompanying space limitations, we were unableto mention all of the relevant studies and cite all original references.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
684 CLARK ET AL.
The Annual Review of Cell and Developmental Biologyis online athttp://cellbio.annualreviews.org
LITERATURE CITED
Abe M, Higuchi I, Morisaki H, Morisaki T,Osame M. 2000. Myoadenylate deaminasedeficiency with progressive muscle weak-ness and atrophy caused by new missensemutations in AMPD1 gene: case reportin a Japanese patient.Neuromusc. Disord.10:472–77
Adams JC, Watt FM. 1993. Regulation of de-velopment and differentiation by the extra-cellular matrix.Development117:1183–98
Acakpo-Satchivi LJ, Edelmann W, Sartorius C,Lu BD, Wahr PA, et al. 1997. Growth andmuscle defects in mice lacking adult myosinheavy chain genes.J. Cell Biol.139:1219–29
Agarkova I, Auerbach D, Ehler E, Perriard JC.2000. A novel marker for vertebrate em-bryonic heart, the EH-myomesin isoform.J.Biol. Chem.275:10256–64
Aihara Y, Kurabayashi M, Saito Y, Ohyama Y,Tanaka T, et al. 2000. Cardiac ankyrin repeatprotein is a novel marker of cardiac hyper-trophy: role of M-CAT element within thepromoter.Hypertension36:48–53
Allen DL, Sartorius CA, Sycuro LK, LeinwandLA. 2001. Different pathways regulate ex-pression of the skeletal myosin heavy chaingenes.J. Biol. Chem.276:43524–33
Almenar-Queralt A, Lee A, Conley CA, Ribasde Pouplana L, Fowler VM. 1999. Identi-fication of a novel tropomodulin isoform,skeletal tropomodulin, that caps actin fila-ment pointed ends in fast skeletal muscle.J.Biol. Chem.274:28466–75
Alyonycheva T, Cohen-Gould L, Siewert C,Fischman DA, Mikawa T. 1997. Skeletalmuscle-specific myosin binding protein-H isexpressed in Purkinje fibers of the cardiacconduction system.Circ. Res.80:665–72
Ampe C, Vandekerckhove J. 1999. Actin. InGuidebook to the Cytoskeletal and MotorProteins, ed. T Kreis, R Vale, pp. 11–14. Ox-ford, UK: Sambrook & Tooze
Anderson JL, Habashi J, Carlquist JF, Muh-lestein JB, Horne BD, et al. 2000. A com-mon variant of the AMPD1 gene predictsimproved cardiovascular survival in patientswith coronary artery disease.J. Am. Coll.Cardiol. 36:1248–52
Andra K, Lassmann H, Bittner R, Shorny S,Fassler R, et al. 1997. Targeted inactivationof plectin reveals essential function in main-taining the integrity of skin, muscle, and heartcytoarchitecture.Genes Dev.11:3143–56
Antin PB, Forry-Schaudies S, Friedman TM,Tapscott SJ, Holtzer H. 1981. Taxol inducespostmitotic myoblasts to assemble interdig-itating microtubule-myosin arrays that ex-clude actin filaments.J. Cell Biol.90:300–8
Anyanful A, Sakube Y, Takuwa K, KagawaH. 2001. The third and fourth tropomyosinisoforms ofCaenorhabditis elegansare ex-pressed in the pharynx and intestines and areessential for development and morphology.J. Mol. Biol.313:525–37
Araishi K, Sasaoka T, Imamura M, Noguchi S,Hama H, et al. 1999. Loss of the sarcoglycancomplex and sarcospan leads to musculardystrophy in beta-sarcoglycan-deficientmice.Hum. Mol. Genet.8:1589–98
Arber S, Halder G, Caroni P. 1994. Muscle LIMprotein, a novel essential regulator of myo-genesis, promotes myogenic differentiation.Cell 79:221–31
Arber S, Hunter JJ, Ross J Jr, Hongo M, SansigG, et al. 1997. MLP-deficient mice exhibit adisruption of cardiac cytoarchitectural orga-nization, dilated cardiomyopathy, and heartfailure.Cell 88:393–403
Arimura T, Nakamura T, Hiroi S, Satoh M,Takahashi M, et al. 2000. Characterization ofthe humannebulettegene: a polymorphismin an actin-binding motif is associated withnonfamilial idiopathic dilated cardiomyopa-thy. Hum. Genet.107:440–51

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 685
Auerbach D, Bantle S, Keller S, Hinderling V,Leu M, et al. 1999. Different domains of theM-band protein myomesin are involved inmyosin binding and M-band targeting.Mol.Biol. Cell 10:1297–308
Ayme-Southgate A, Vigoreaux J, BenianG, Pardue ML. 1991.Drosophila has atwitchin/titin-related gene that appears to en-code projectin.Proc. Natl. Acad. Sci. USA88:7973–77
Ayoob JC, Turnacioglu KK, Mittal B, SangerJM, Sanger JW. 2000. Targeting of cardiacmuscle titin fragments to the Z-bands anddense bodies of living muscle and non-mus-cle cells.Cell. Motil. Cytoskeleton45:67–82
Babcock G, Fowler VM. 1994. Isoform spe-cific interaction of tropomodulin with skele-tal muscle and erythrocyte tropomyosins.J.Biol. Chem.269:27510–18
Bahler M, Eppenberger HM, Wallimann T.1985a. Novel thick filament protein ofchicken pectoralis muscle: the 86 kDa pro-tein. I. Purification and characterization.J.Mol. Biol. 186:381–91
Bahler M, Wallimann T, Eppenberger HM.1985b. Myofibrillar M-band proteins repre-sent constituents of native thick filaments,frayed filaments and bare zone assemblages.J. Muscle Res. Cell Motil.6:783–800
Baker LP, Daggett DF, Peng HB. 1994. Con-centration of pp125 focal adhesion kinase(FAK) at the myotendinous junction.J. CellSci.107:1485–97
Balogh J, Merisckay M, Li Z, Paulin D, ArnerA. 2002. Hearts from mice lacking desminhave a myopathy with impaired active forcegeneration and unaltered wall compliance.Cardiovasc. Res.53:439–50
Bang ML, Centner T, Fornoff F, Geach AJ, Got-thardt M, et al. 2001a. The complete genesequence of titin, expression of an unusualapproximately 700-kDa titin isoform, and itsinteraction with obscurin identify a novelZ-line to I-band linking system.Circ. Res.89:1065–72
Bang ML, Mudry RE, McElhinny AS, Trom-bitas K, Geach AJ, et al. 2001b. Myopal-ladin, a novel 145-kilodalton sarcomeric pro-
tein with multiple roles in Z-disc and I-bandprotein assemblies.J. Cell Biol.153:413–27
Bao ZZ, Lakonishok M, Kaufman S, HorwitzAF. 1993. Alpha 7 beta 1 integrin is a compo-nent of the myotendinous junction on skeletalmuscle.J. Cell Sci.106:579–89
Barral JM, Epstein HF. 1999. Protein machinesand self assembly in muscle organization.BioEssays21:813–23
Baumeister A, Arber S, Caroni P. 1997. Ac-cumulation of muscle ankyrin repeat proteintranscript reveals local activation of primarymyotube endcompartments during musclemorphogenesis.J. Cell Biol.139:1231–42
Beam KG. 1988. Duchenne muscular dystro-phy. Localizing the gene product.Nature333:798–99
Belkin AM, Zhidkova NI, Koteliansky VE.1986. Localization of talin in skeletal andcardiac muscles.FEBS Lett.200:32–36
Bellin RM, Huiatt TW, Critchley DR, RobsonRM. 2001. Synemin may function to directlylink muscle cell intermediate filaments toboth myofibrillar Z-lines and costameres.J.Biol. Chem.276:32330–37
Benian GM, Kiff JE, Neckelmann N, MoermanDG, Waterston RH. 1989. Sequence of anunusually large protein implicated in regula-tion of myosin activity inC. elegans. Nature342:45–50
Benian GM, Tang X, Tinley TL. 1996a.Twitchin and related giant Ig superfamilymembers ofC. elegansand other inverte-brates.Adv. Biophys.33:183–98
Benian GM, Tinley TL, Tang X, BorodovskyM. 1996b. TheCaenorhabditis elegansgeneunc-89, required fpr muscle M-line assem-bly, encodes a giant modular protein com-posed of Ig and signal transduction domains.J. Cell Biol.132:835–48
Bennett P, Craig R, Starr R, Offer G. 1986.The ultrastructural location of C-protein, X-protein and H-protein in rabbit muscle.J.Muscle Res. Cell Motil.7:550–67
Bernstein SI, Milligan RA. 1997. Fine tuning amolecular motor: the location of alternativedomains in theDrosophilamyosin head.J.Mol. Biol. 271:1–6

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
686 CLARK ET AL.
Bernstein SI, Mogami K, Donady JJ, Emer-son CP Jr. 1983.Drosophilamuscle myosinheavy chain encoded by a single genein a cluster of muscle mutations.Nature302:393–97
Bilak SR, Sernett SW, Bilak MM, Bellin RM,Stromer MH, et al. 1998. Properties of thenovel intermediate filament protein syneminand its identification in mammalian muscle.Arch. Biochem. Biophys.355:63–76
Bione S, Maestrini E, Rivella S, Mancini M,Regis S, et al. 1994. Identification of anovel X-linked gene responsible for Emery-Dreifuss muscular dystrophy.Nat. Genet.8:323–27
Blake DJ, Weir A, Newey SE, Davies KE.2002. Function and genetics of dystrophinand dystrophin-related proteins in muscle.Physiol. Rev.82:291–329
Blanchard A, Ohanian V, Critchley D. 1989.The structure and function of alpha-actinin.J. Muscle Res. Cell Motil.10:280–89
Bodine SC, Latres E, Baumhueter S, Lai VK,Nunez L, et al. 2001. Identification of ubiq-uitin ligases required for skeletal muscle at-rophy.Science294:1704–8
Boudriau S, Vincent M, Cote CH, RogersPA. 1993. Cytoskeletal structure of skele-tal muscle: identification of an intricate exo-sarcomeric microtubule lattice in slow- andfast-twitch muscle fibers.J. Histochem. Cy-tochem.41:1013–21
Bouvard D, Brakebusch C, Gustafsson E, As-zodi A, Bengtsson T, et al. 2001. Functionalconsequences of integrin gene mutations inmice.Circ. Res.89:211–23
Bozyczko D, Decker C, Muschler J, HorwitzAF. 1989. Integrin on developing and adultskeletal muscle.Exp. Cell Res.183:72–91
Brault V, Reedy MC, Sauder U, Kammerer RA,Aebi U, Schoenenberger C. 1999. Substitu-tion of flight muscle-specific actin by human(beta)-cytoplasmic actin in the indirect flightmuscle ofDrosophila. J. Cell Sci.112:3627–39
Brenman JE, Chao DS, Xia H, Aldape K, BredtDS. 1995. Nitric oxide synthase complexedwith dystrophin and absent from skeletal
muscle sarcolemma in Duchenne musculardystrophy.Cell 82:743–52
Broschat KO. 1990. Tropomyosin prevents de-polymerization of actin filaments from thepointed end.J. Biol. Chem.265:21323–29
Brown JH, Kim KH, Jun G, Greenfield NJ,Dominguez R, et al. 2001. Deciphering thedesign of the tropomyosin molecule.Proc.Natl. Acad. Sci. USA98:8496–501
Brown NH, Gregory SL, Martin-Bermudo ZD.2000. Integrins as mediators of morphogen-esis inDrosophila. Dev. Biol.223:1–16
Burkin DJ, Wallace GQ, Nicol KJ, Kaufman DJ,Kaufman SJ. 2001. Enhanced expression ofthe alpha 7 beta 1 integrin reduces musculardystrophy and restores viability in dystrophicmice.J. Cell Biol.152:1207–18
Calderwood DA, Shattil SJ, Ginsberg MH.2000. Integrins and actin filaments: recipro-cal regulation of cell adhesion and signaling.J. Biol. Chem.275:22607–10
Canepari M, Cappelli V, Pellegrino MA, Za-nardi MC, Reggiani C. 1998. Thyroid hor-mone regulation of MHC isoform compo-sition and myofibrillar ATPase activity inrat skeletal muscles.Arch. Physiol. Biochem.106:308–15
Carlsson L, Li Z, Paulin D, Thornell LE. 1999.Nestin is expressed during developmentand in myotendinous and neuromuscularjunctions in wild type and desmin knock-outmice.Exp. Cell Res.251:213–23
Carlsson L, Thornell LE. 2001. Desmin-relatedmyopathies in mice and man.Acta Physiol.Scand.171:341–48
Cartegni L, di Barletta MR, Barresi R, Squar-zoni S, Sabatelli P, et al. 1997. Heart-specificlocalization of emerin: new insights intoEmery-Dreifuss muscular dystrophy.Hum.Mol. Genet.6:2257–64
Casella JF, Craig SW, Maack DJ, Brown AE.1987. Cap Z(36/32), a barbed end actin-capping protein, is a component of the Z-lineof skeletal muscle.J. Cell Biol.105:371–79
Casella JF, Torres MA. 1994. Interaction ofCap Z with actin. The NH2-terminal do-mains of the alpha-1 and beta-subunitsare not required for actin capping, and

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 687
alpha-1-beta and alpha-2-beta heterodimersbind differentially to actin.J. Biol. Chem.269:6992–98
Cazorla O, Freiburg A, Helmes M, Centner T,McNabb M, et al. 2000. Differential expres-sion of cardiac titin isoforms and modulationof cellular stiffness.Circ. Res.86:59–67
Centner T, Yano J, Kimura E, McElhinny AS,Pelin K, et al. 2001. Identification of mus-cle specific ring finger proteins as potentialregulators of the titin kinase domain.J. Mol.Biol. 306:717–26
Champagne MB, Edwards KA, Erick-son HP, Kiehart DP. 2000.Drosophilastretchin-MLCK is a novel member of theTitin/Myosin light chain kinase family.J.Mol. Biol. 300:759–77
Chandy IK, Lo JC, Ludescher RD. 1999. Dif-ferential mobility of skeletal and cardiactropomyosin on the surface of F-actin.Bio-chemistry38:9286–94
Chapelle JP. 1999. Cardiac troponin I and tro-ponin T: recent players in the field of myocar-dial markers.Clin. Chem. Lab. Med.37:11–20
Chauhan VS, Tuvia S, Buhusi M, Bennett V,Grant AO. 2000. Abnormal cardiac Na(+)channel properties and QT heart rate adapta-tion in neonatal ankyrin(B) knockout mice.Circ. Res.86:441–47
Chien KR. 2000. Genomic circuits and the in-tegrative biology of cardiac diseases.Nature407:227–32
Chu W, Burns DK, Swerlick RA, Presky DH.1995. Identification and characterization ofa novel cytokine-inducible nuclear proteinfrom human endothelial cells.J. Biol. Chem.270:10236–45
Chu X, Thompson D, Yee LJ, Sung LA. 2000.Genomic organization of mouse and hu-man erythrocyte tropomodulin genes encod-ing the pointed end capping protein for theactin filaments.Gene256:271–81
Chun M, Falkenthal S. 1988. Ifm(2)2 is amyosin heavy chain allele that disrupts my-ofibrillar assembly only in the indirect flightmuscle ofDrosophila melanogaster. J. CellBiol. 107:2613–21
Cole HA, Perry SV. 1975. The phosphorylationof troponin I from cardiac muscle.Biochem.J. 149:525–33
Colucci-Guyon E, Portier MM, Dunia I, PaulinD, Pournin S, Babinet C. 1994. Mice lackingvimentin develop and reproduce without anobvious phenotype.Cell 79:679–94
Cooper J, Trinick J. 1984. Binding and locationof AMP deaminase in rabbit psoas musclemyofibrils.J. Mol. Biol.177:137–52
Cooper JA, Schafer DA. 2000. Control of actinassembly and disassembly at filament ends.Curr. Opin. Cell Biol.12:97–103
Coral-Vazquez R, Cohn RD, Moore SA, HillJA, Weiss RM, et al. 1999. Disruption of thesarcoglycan-sarcospan complex in vascularsmooth muscle: a novel mechanism for car-diomyopathy and muscular dystrophy.Cell98:465–74
Corbett MA, Robinson CS, Dunglison GF,Yang N, Joya JE, et al. 2001. A mutationin alpha-tropomyosin (slow) affects musclestrength, maturation and hypertrophy in amouse model for nemaline myopathy.Hum.Mol. Genet.10:317–28
Cox PR, Siddique T, Zoghbi HY. 2001. Ge-nomic organization of tropomodulins 2 and 4and unusual intergenic and intraexonic splic-ing of YL-1 and tropomodulin 4.BioMed.Cent. Genomics2:7
Cox PR, Zoghbi HY. 2000. Sequencing, expres-sion analysis, and mapping of three uniquehuman tropomodulin genes and their mouseorthologs.Genomics63:97–107
Craig R, Lehman W. 2001. Crossbridge andtropomyosin positions observed in native, in-teracting thick and thin filaments.J. Mol.Biol. 311:1027–36
Craig R, Offer G. 1976. The location of C-protein in rabbit skeletal muscle.Proc. R.Soc. London Ser. B192:451–61
Crawford AW, Michelsen JW, Beckerle MC.1992. An interaction between zyxin andalpha-actinin.J. Cell Biol.116:1381–93
Cuda G, Fananapazir L, Epstein ND, Sellers JR.1997. The in vitro motility activity of beta-cardiac myosin depends on the nature of thebeta-myosin heavy chain gene mutation in

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
688 CLARK ET AL.
hypertrophic cardiomyopathy.J. Muscle Res.Cell Motil. 18:275–83
Dai KS, Liew CC. 2001. A novel human striatedmuscle RING zinc finger protein, SMRZ, in-teracts with SMT3b via its RING domain.J.Biol. Chem.276:23992–99
Dalloz F, Osinska H, Robbins J. 2001. Manip-ulating the contractile apparatus: geneticallydefined animal models of cardiovascular dis-ease.J. Mol. Cell Cardiol.33:9–25
Danowski BA, Imanaka-Yoshida K, Sanger JM,Sanger JW. 1992. Costameres are sites offorce transmission to the substratum in adultrat cardiomyocytes.J. Cell Biol.118:1411–20
Davis JS. 1988. Interaction of C-protein withpH 8.0 synthetic thick filaments preparedfrom the myosin of vertebrate skeletal mus-cle.J. Muscle Res. Cell Motil.9:174–83
Dennis JE, Shimizu T, Reinach FC, FischmanDA. 1984. Localization of C-protein iso-forms in chicken skeletal muscle: ultrastruc-tural detection using monoclonal antibodies.J. Cell Biol.98:1514–22
Djinovic-Carugo K, Young P, Gautel M, SarasteM. 1999. Structure of the alpha-actinin rod:molecular basis for cross-linking of actin fil-aments.Cell 98:537–46
Donner K, Ollikainen M, Ridanpaa M, Chris-ten HJ, Goebel HH, et al. 2002. Mutations inthe beta-tropomyosin (TPM2) gene—a rarecause of nemaline myopathy.Neuromuscul.Disord.12:151–58
Duclos F, Straub V, Moore SA, Venzke DP,Hrstka RF, et al. 1998. Progressive muscu-lar dystrophy in alpha-sarcoglycan-deficientmice.J. Cell Biol.142:1461–71
Durick K, Wu RY, Gill GN, Taylor SS. 1996.Mitogenic signaling by Ret/ptc2 requires as-sociation with enigma via a LIM domain.J.Biol. Chem.271:12691–94
Dyson JM, O’Malley CJ, Becanovic J, Mun-day AD, Berndt MC, et al. 2001. TheSH2-containing inositol polyphosphate 5-phosphatase, SHIP-2, binds filamin and reg-ulates submembraneous actin.J. Cell Biol.155:1065–79
Ecarnot-Laubriet A, De Luca K, Vandroux D,
Moisant M, Bernard C, et al. 2000. Downreg-ulation and nuclear relocation of MLP duringthe progression of right ventricular hypertro-phy induced by chronic pressure overload.J.Mol. Cell Cardiol.32:2385–95
Ehler E, Horowits R, Zuppinger C, Price RL,Perriard E, et al. 2001. Alterations at the in-tercalated disk associated with the absence ofmuscle LIM protein.J. Cell Biol.153:763–72
Ehler E, Rothen BM, Hammerle SP, KomiyamaM, Perriard JC. 1999. Myofibrillogenesis inthe developing chicken heart: assembly of Z-disk, M-line and the thick filaments.J. CellSci.112:1529–39
Emery AE. 2000. Emery-Dreifuss musculardystrophy—a 40-year retrospective.Neuro-muscul. Disord.10:228–32
Ervasti JM, Ohlendieck K, Kahl SD, Gaver MG,Campbell KP. 1990. Deficiency of a glyco-protein component of the dystrophin comp-lex in dystrophic muscle.Nature345:315–19
Fassler R, Rohwedel J, Maltsev V, Bloch W,Lentini S, et al. 1996. Differentiation and in-tegrity of cardiac muscle cells are impairedin the absence of beta 1 integrin.J. Cell Sci.109:2989–99
Faulkner G, Lanfranchi G, Valle G. 2001. Tele-thonin and other new proteins of the Z-discof skeletal muscle.IUBMB Life51:275–82
Faulkner G, Pallavicini A, Comelli A, SalamonM, Bortoletto G, et al. 2000. FATZ, a filamin-,actinin-, and telethonin-binding protein ofthe Z- disc of skeletal muscle.J. Biol. Chem.275:41234–42
Faulkner G, Pallavicini A, Formentin E,Comelli A, Ievolella C, et al. 1999. ZASP: anew Z-band alternatively spliced PDZ-motifprotein.J. Cell Biol.146:465–75
Fernandez C, Figarella-Branger D, Alla P, HarleJR, Pellissier JF. 2002. Colchicine myopa-thy: a vacuolar myopathy with selective typeI muscle fiber involvement. An immunohis-tochemical and electron microscopic studyof two cases.Acta Neuropathol.103:100–6
Flick MJ, Konieczny SF. 2000. The muscle reg-ulatory and structural protein MLP is a cy-toskeletal binding partner of betaI-spectrin.J. Cell Sci.113:1553–64

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 689
Flucher BE, Morton ME, Froehner SC, DanielsMP. 1990. Localization of the alpha 1 andalpha 2 subunits of the dihydropyridine re-ceptor and ankyrin in skeletal muscle triads.Neuron5:339–51
Fock U, Hinssen H. 1999. Identification and lo-calisation of nebulin as a thin filament com-ponent of invertebrate chordate muscles.J.Comp. Physiol. B169:555–60
Foisner R, Wiche G. 1991. Intermediatefilament-associated proteins.Curr. Opin.Cell Biol. 3:75–81
Foucault G, Vacher M, Merkulova T, Keller A,Arrio-Dupont M. 1999. Presence of enolasein the M-band of skeletal muscle and pos-sible indirect interaction with the cytosolicmuscle isoform of creatine kinase.Biochem.J. 338:115–21
Fowler VM. 1987. Identification and purifi-cation of a novelMr 43,000 tropomyosin-binding protein from human erythrocytemembranes.J. Biol. Chem.262:12792–800
Fowler VM, Conley C. 1999. Tropomodulin.In Guidebook to the Cytoskeletal and MotorProteins, ed. T Kreis, R Vale, pp. 154–59.Oxford, UK: Sambrook & Tooze
Franzini-Armstrong C. 1973. The structure ofa simple Z line.J. Cell Biol.58:630–42
Freemont PS. 2000. RING for destruction?Curr. Biol. 10:R84–87
Freiburg A, Gautel M. 1996. A molecular mapof the interactions between titin and myosin-binding protein C. Implications for sarcom-eric assembly in familial hypertrophic car-diomyopathy.Eur. J. Biochem.235:317–23
Freiburg A, Trombitas K, Hell W, Cazorla O,Fougerousse F, et al. 2000. Series of exon-skipping events in the elastic spring regionof titin as the structural basis for myofibrillarelastic diversity.Circ. Res.86:1114–21
Frey N, Olson EN. 2000. Making matters worsefor a broken heart.J. Clin. Invest.106:1437–39
Frey N, Olson EN. 2002. Calsarcin-3, a novelskeletal muscle-specific member of the cal-sarcin family, interacts with multiple Z-discproteins.J. Biol. Chem.277:13998–4004
Frey N, Richardson JA, Olson EN. 2000.
Calsarcins, a novel family of sarcomericcalcineurin-binding proteins.Proc. Natl.Acad. Sci. USA97:14632–37
Funatsu T, Kono E, Higuchi H, Kimura S, Ishi-wata S, et al. 1993. Elastic filaments in situ incardiac muscle: deep-etch replica analysis incombination with selective removal of actinand myosin filaments.J. Cell Biol.120:711–24
Furukawa T, Ono Y, Tsuchiya H, Katayama Y,Bang ML, et al. 2001. Specific interactionof the potassium channel beta-subunit minKwith the sarcomeric protein T-cap suggestsa T-tubule-myofibril linking system.J. Mol.Biol. 313:775–84
Fyrberg C, Ketchum A, Ball E, Fyrberg E.1998. Characterization of lethalDrosophilamelanogasteralpha-actinin mutants.Bio-chem. Genet.36:299–310
Gache Y, Chavanas S, Lacour JP, Wiche G,Owaribe K, et al. 1996. Defective expressionof plectin/HD1 in epidermolysis bullosa sim-plex with muscular dystrophy.J. Clin. Invest.97:2289–98
Gallicano GI, Bauer C, Fuchs E. 2001. Rescu-ing desmoplakin function in extra-embryonicectoderm reveals the importance of this pro-tein in embryonic heart, neuroepithelium,skin and vasculature.Development128:929–41
Garvey SM, Rajan C, Lerner AP, Frankel WN,Cox GA. 2002. The muscular dystrophy withmyositis (mdm) mouse mutation disrupts askeletal muscle-specific domain of titin.Ge-nomics79:146–49
Gautel M, Zuffardi O, Freiburg A, LabeitS. 1995b. Phosphorylation switches specificfor the cardiac isoform of myosin bindingprotein-C: a modulator of cardiac contrac-tion?EMBO J.14:1952–60
George EL, Ober MB, Emerson CP Jr.1989. Functional domains of theDrosophilamelanogastermuscle myosin heavy-chaingene are encoded by alternatively spliced ex-ons.Mol. Cell Biol.9:2957–74
Gerull B, Gramlich M, Atherton J, McNabbM, Trombitas K, et al. 2002. Mutations ofTTN, encoding the giant muscle filament

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
690 CLARK ET AL.
titin, cause familial dilated cardiomyopathy.Nat. Genet.30:201–4
Giancotti FG, Ruoslahti E. 1999. Integrin sig-naling.Science285:1028–32
Gilbert R, Kelly MG, Mikawa T, Fischman DA.1996. The carboxyl terminus of myosin bind-ing protein C (MyBP-C, C-protein) speci-fies incorporation into the A-band of striatedmuscle.J. Cell Sci.109:101–11
Goebel HH. 1995. Desmin-related neuromus-cular disorders.Muscle Nerve18:1306–20
Goldstein MA, Schroeter JP, Sass RL. 1979.The Z lattice in canine cardiac muscle.J. CellBiol. 83:187–204
Gordon ES, Hoffman EP. 2001. The ABCsof limb-girdle muscular dystrophy: alpha-sarcoglycanopathy, Bethlem myopathy, cal-painopathy and more.Curr. Opin. Neurol.14:567–73
Grady RM, Grange RW, Lau KS, MaimoneMM, Nichol MC, et al. 1999. Role foralpha-dystrobrevin in the pathogenesis ofdystrophin-dependent muscular dystrophies.Nat. Cell Biol.1:215–20
Granger BL, Lazarides E. 1979. Desmin andvimentin coexist at the periphery of the my-ofibril Z disc.Cell 18:1053–63
Granzier G, Labeit S. 2002. Cardiac titin: anadjustable multi-functional spring.J. Phys-iol. In press
Green KJ, Kowalczyk AP, Bornslaeger EA,Palka HL, Norvell SM. 1998. Desmosomes:integrators of mechanical integrity in tissues.Biol. Bull. 194:374–76; discussion 6–7
Greenfield NJ, Fowler VM. 2002. Tropomyosinrequires an intact N-terminal coiled coilto interact with tropomodulin.Biophys. J.82:2580–91
Gregorio CC. 1997. Models of thin filament as-sembly.Cell Struct. Funct.22:191–96
Gregorio CC, Trombitas K, Centner T,Kolmerer B, Stier G, et al. 1998. The NH2
terminus of titin spans the Z-disc: its inter-action with a novel 19-kD ligand (T-cap) isrequired for sarcomeric integrity.J. Cell Biol.143:1013–27
Gregorio CC, Weber A, Bondad M, Pennise CR,Fowler VM. 1995. Requirement of pointed-
end capping by tropomodulin to maintainactin filament length in embryonic chick car-diac myocytes.Nature377:83–86
Grove BK, Kurer V, Lehner C, Doetschman TC,Perriard JC, Eppenberger HM. 1984. A new185,000-dalton skeletal muscle protein de-tected by monoclonal antibodies.J. Cell Biol.98:518–24
Gruen M, Prinz H, Gautel M. 1999. cAPK-phosphorylation controls the interaction ofthe regulatory domain of cardiac myosinbinding protein C with myosin-S2 in an on-off fashion.FEBS Lett.453:254–59
Gurgel-Giannetti J, Reed U, Bang ML, PelinK, Donner K, et al. 2001. Nebulin expressionin patients with nemaline myopathy.Neuro-muscul. Disord.11:154–62
Gutierrez-Cruz G, Van Heerden AH, Wang K.2001. Modular motif, structural folds andaffinity profiles of the PEVK segment of hu-man fetal skeletal muscle titin.J. Biol. Chem.276:7442–49
Guy PM, Kenny DA, Gill GN. 1999. The PDZdomain of the LIM protein enigma binds tobeta-tropomyosin.Mol. Biol. Cell 10:1973–84
Hack AA, Ly CT, Jiang F, Clendenin CJ,Sigrist KS, et al. 1998. Gamma-sarcoglycandeficiency leads to muscle membrane defectsand apoptosis independent of dystrophin.J.Cell Biol. 142:1279–87
Hakeda S, Endo S, Saigo K. 2000. Require-ments of Kettin, a giant muscle protein highlyconserved in overall structure in evolution,for normal muscle function, viability, andflight activity of Drosophila. J. Cell Biol.148:101–14
Hammarlund M, Davis WS, Jorgensen EM.2000. Mutations in beta-spectrin disrupt axonoutgrowth and sarcomere structure.J. CellBiol. 149:931–42
Harper BD, Beckerle MC, Pomies P. 2000.Fine mapping of the alpha-actinin bindingsite within cysteine-rich protein.Biochem. J.350:269–74
Harris SP, Bartley CR, Hacker TA, McDonaldKS, Douglas PS, et al. 2002. Hypertrophiccardiomyopathy in cardiac myosin binding

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 691
protein-C knockout mice.Circ. Res.90:594–601
Hartzell HC. 1984. Phosphorylation of C-protein in intact amphibian cardiac muscle.Correlation between 32P incorporation andtwitch relaxation.J. Gen. Physiol.83:563–88
Hartzell HC. 1985. Effects of phosphorylatedand unphosphorylated C-protein on cardiacactomyosin ATPase.J. Mol. Biol. 186:185–95
Hauser MA, Horrigan SK, Salmikangas P, To-rian UM, Viles KD, et al. 2000. Myotilin ismutated in limb girdle muscular dystrophy1A. Hum. Mol. Genet.9:2141–47
Hayashi YK, Chou FL, Engvall E, Ogawa M,Matsuda C, et al. 1998. Mutations in the inte-grin alpha7 gene cause congenital myopathy.Nat. Genet.19:94–97
Heierhorst J, Probst WC, Kohanski RA, BukuA, Weiss KR. 1995. Phosphorylation ofmyosin regulatory light chains by the mol-luscan twitchin kinase.Eur. J. Biochem.233:426–31
Hein S, Kostin S, Heling A, Maeno Y, SchaperJ. 2000. The role of the cytoskeleton in heartfailure.Cardiovasc. Res.45:273–78
Hernandez OM, Housmans PR, Potter JD.2001. Invited review: pathophysiology ofcardiac muscle contraction and relaxation asa result of alterations in thin filament regula-tion. J. Appl. Physiol.90:1125–36
Herrera AH, Elzey B, Law DJ, Horowits R.2000. Terminal regions of mouse nebulin:sequence analysis and complementary local-ization with N-RAP.Cell Motil. Cytoskeleton45:211–22
Hertig CM, Eppenberger-Eberhardt M, Koch S,Eppenberger HM. 1996. N-cadherin in adultrat cardiomyocytes in culture. I. Functionalrole of N-cadherin and impairment of cell-cell contact by a truncated N-cadherin mu-tant.J. Cell Sci.109:1–10
Herzberg O, James MN. 1988. Refined crystalstructure of troponin C from turkey skele-tal muscle at 2.0A resolution.J. Mol. Biol.203:761–79
Hijikata T, Murakami T, Imamura M, Fujimaki
N, Ishikawa H. 1999. Plectin is a linker ofintermediate filaments to Z-discs in skeletalmuscle fibers.J. Cell Sci.112:867–76
Hisatome I, Morisaki T, Kamma H, SugamaT, Morisaki H, et al. 1998. Control of AMPdeaminase 1 binding to myosin heavy chain.Am. J. Physiol. Cell Physiol.275:C870–C81
Hitchcock SE. 1975. Regulation of muscle con-traction: bindings of troponin and its com-ponents to actin and tropomyosin.Eur. J.Biochem.52:255–63
Hodges BL, Hayashi YK, Nonaka I, Wang W,Arahata K, Kaufman SJ. 1997. Altered ex-pression of the alpha7beta1 integrin in humanand murine muscular dystrophies.J. Cell Sci.110:2873–81
Hoffman EP, Brown RH Jr, Kunkel LM.1987. Dystrophin: the protein product of theDuchenne muscular dystrophy locus.Cell51:919–28
Hofmann W, Reichart B, Ewald A, Muller E,Schmitt I, et al. 2001. Cofactor requirementsfor nuclear export of Rev response element(RRE)- and constitutive transport element(CTE)-containing retroviral RNAs. An unex-pected role for actin.J. Cell Biol.152:895–910
Holtzer H, Hijikata T, Lin ZX, Zhang ZQ,Holtzer S, et al. 1997. Independent assemblyof 1.6µm long bipolar MHC filaments andI-Z-I bodies.Cell Struct. Funct.22:83–93
Honda K, Yamada T, Endo R, Ino Y, Gotoh M,et al. 1998. Actinin-4, a novel actin-bundlingprotein associated with cell motility and can-cer invasion.J. Cell Biol.140:1383–93
Hornemann T, Stolz M, Wallimann T. 2000.Isoenzyme-specific interaction of muscle-type creatine kinase with the sarcomeric M-line is mediated by NH(2)-terminal lysinecharge-clamps.J. Cell Biol.149:1225–34
Houmeida A, Holt J, Tskhovrebova L, TrinickJ. 1995. Studies of the interaction betweentitin and myosin.J. Cell Biol. 131:1471–81
Hug C, Miller TM, Torres MA, Casella JF,Cooper JA. 1992. Identification and charac-terization of an actin-binding site of CapZ.J.Cell Biol. 116:923–31

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
692 CLARK ET AL.
Hughes SM, Cho M, Karsch-Mizrachi I, TravisM, Silberstein L, et al. 1993. Three slowmyosin heavy chains sequentially expressedin developing mammalian skeletal muscle.Dev. Biol.158:183–99
Huttelmaier S, Illenberger S, Grosheva I, Rudi-ger M, Singer RH, Jockusch BM. 2001.Raver1, a dual compartment protein, is a lig-and for PTB/hnRNPI and microfilament at-tachment proteins.J. Cell Biol.155:775–86
Huxley AF. 2000. Cross-bridge action: presentviews, prospects, and unknowns.J. Biomech.33:1189–95
Ikeda Y, Gu Y, Iwanaga Y, Hoshijima M, OhSS, et al. 2002. Restoration of deficient mem-brane proteins in the cardiomyopathic ham-ster by in vivo cardiac gene transfer.Circu-lation 105:502–8
Ilkovski B, Cooper ST, Nowak K, Ryan MM,Yang N, et al. 2001. Nemaline myopathycaused by mutations in the muscle alpha-skeletal-actin gene.Am. J. Hum. Genet.68:1333–43
Itoh-Satoh M, Hayashi T, Nishi H, Koga Y,Arimura T, et al. 2002. Titin mutations as themolecular basis for dilated cardiomyopathy.Biochem. Biophys. Res. Commun.291:385–93
Jeyaseelan R, Poizat C, Baker RK, Abdishoo S,Isterabadi LB, et al. 1997. A novel cardiac-restricted target for doxorubicin. CARP, a nu-clear modulator of gene expression in cardiacprogenitor cells and cardiomyocytes.J. Biol.Chem.272:22800–8
Jin J-P. 1995. Cloned rat cardiac titin class I andclass II motifs.J. Biol. Chem.270:6908–16
Jin J-P, Huang QQ, Yeh HI, Lin JJ. 1992. Com-plete nucleotide sequence and structural or-ganization of rat cardiac troponin T gene. Asingle gene generates embryonic and adultisoforms via developmentally regulated al-ternative splicing.J. Mol. Biol.227:1269–76
Jin J-P, Wang K. 1991. Nebulin as a giantactin-binding template protein in skeletalmuscle sarcomere. Interaction of actin andcloned human nebulin fragments.FEBS Lett.281:93–96
Jo K, Rutten B, Bunn RC, Bredt DS. 2001.
Actinin-associated LIM protein-deficientmice maintain normal development andstructure of skeletal muscle.Mol. Cell Biol.21:1682–87
Jones WK, Grupp IL, Doetschman T, GruppG, Osinska H, et al. 1996. Ablation of themurine alpha myosin heavy chain gene leadsto dosage effects and functional deficits inthe heart.J. Clin. Invest.98:1906–17
Kabsch W, Mannherz HG, Suck D, Pai EF,Holmes KC. 1990. Atomic structure of theactin:DNase I complex.Nature347:37–44
Kameya S, Miyagoe Y, Nonaka I, Ikemoto T,Endo M, et al. 1999. alpha1-syntrophin genedisruption results in the absence of neuronal-type nitric-oxide synthase at the sarcolemmabut does not induce muscle degeneration.J.Biol. Chem.274:2193–200
Kano Y, Fujimaki N, Ishikawa H. 1991. Thedistribution and arrangement of microtubulesin mammalian skeletal muscle fibers.CellStruct. Funct.16:251–61
Kaufmann U, Kirsch J, Irintchev A, Wernig A,Starzinski-Powitz A. 1999. The M-cadherincatenin complex interacts with microtubulesin skeletal muscle cells: implications for thefusion of myoblasts.J. Cell Sci.112:55–68
Kawada T, Nakazawa M, Nakauchi S, Ya-mazaki K, Shimamoto R, et al. 2002. Rescueof hereditary form of dilated cardiomyopa-thy by rAAV-mediated somatic gene ther-apy: amelioration of morphological findings,sarcolemmal permeability, cardiac perfor-mances, and the prognosis of TO-2 hamsters.Proc. Natl. Acad. Sci. USA99:901–6
Kemp TJ, Sadusky TJ, Saltisi F, Carey N,Moss J, et al. 2000. Identification of Ankrd2,a novel skeletal muscle gene coding fora stretch-responsive ankyrin-repeat protein.Genomics66:229–41
Kenny PA, Liston EM, Higgins DG. 1999.Molecular evolution of immunoglobulin andfibronectin domains in titin and related mus-cle proteins.Gene232:11–23
Kenyon GL, Reed GH. 1983. Creatine kinase:structure-activity relationships.Adv. Enzy-mol. Relat. Areas Mol. Biol.54:367–426
Kerfant BG, Vassort G, Gomez AM. 2001.

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 693
Microtubule disruption by colchicine re-versibly enhances calcium signaling in intactrat cardiac myocytes.Circ. Res.88:E59–65
Khaitlina SY. 2001. Functional specificity ofactin isoforms.Int. Rev. Cytol.202:35–98
Kilby PM, Van Eldik LJ, Roberts GC. 1997.Identification of the binding site on S100Bprotein for the actin capping protein CapZ.Protein Sci.6:2494–503
Kimura A, Harada H, Park JE, Nishi H, SatohM, et al. 1997. Mutations in the cardiac tro-ponin I gene associated with hypertrophiccardiomyopathy.Nat. Genet.16:379–82
Klein I. 1983. Colchicine stimulates the rate ofcontraction of heart cells in culture.Cardio-vasc. Res.17:459–65
Kolmerer B, Clayton J, Benes V, Allen T,Ferguson C, et al. 2000. Sequence and ex-pression of the kettin gene inDrosophilamelanogasterandCaenorhabditis elegans.J.Mol. Biol. 296:435–48
Kolmerer B, Olivieri N, Witt CC, HerrmannBG, Labeit S. 1996. Genomic organization ofthe M line titin and its tissue-specific expres-sion in two distinct isoforms.J. Mol. Biol.256:556–63
Kong Y, Flick MJ, Kudla AJ, Konieczny SF.1997. Muscle LIM protein promotes myo-genesis by enhancing the activity of MyoD.Mol. Cell Biol.17:4750–60
Kordeli E, Ludosky MA, Deprette C, FrappierT, Cartaud J. 1998. AnkyrinG is associatedwith the postsynaptic membrane and the sar-coplasmic reticulum in the skeletal musclefiber.J. Cell Sci.111:2197–207
Koretz JF. 1979. Effects of C-protein on syn-thetic myosin filament structure.Biophys. J.27:433–46
Koretz JF, Irving TC, Wang K. 1993. Fila-mentous aggregates of native titin and bind-ing of C-protein and AMP-deaminase.Arch.Biochem. Biophys.304:305–9
Kosmehl H, Langbein L, Katenkamp D. 1990.Transient cytokeratin expression in skeletalmuscle during murine embryogenesis.Anat.Anz.171:39–44
Kostyukova A, Maeda K, Yamauchi E, KriegerI, Maeda Y. 2000. Domain structure of
tropomodulin: distinct properties of the N-terminal and C-terminal halves.Eur. J.Biochem.267:6470–75
Kovacic-Milivojevic B, Roediger F, AlmeidaEA, Damsky CH, Gardner DG, Ilic D. 2001.Focal adhesion kinase and p130Cas mediateboth sarcomeric organization and activationof genes associated with cardiac myocyte hy-pertrophy.Mol. Biol. Cell12:2290–307
Kreuz AJ, Simcox A, Maughan D. 1996. Al-terations in flight muscle ultrastructure andfunction inDrosophilatropomyosin mutants.J. Cell Biol.135:673–87
Kruger M, Wright J, Wang K. 1991. Nebulin asa length regulator of thin filaments of verte-brate skeletal muscles: correlation of thin fil-ament length, nebulin size, and epitope pro-file. J. Cell Biol.115:97–107
Kulke M, Fujita-Becker S, Rostkova E, NeagoeC, Labeit D, et al. 2001a. Interaction betweenPEVK-titin and actin filaments: origin of aviscous force component in cardiac myofib-rils. Circ. Res.89:874–81
Kulke M, Neagoe C, Kolmerer B, MinajevaA, Hinssen H, et al. 2001b. Kettin, a majorsource of myofibrillar stiffness inDrosophilaindirect flight muscle.J. Cell Biol.154:1045–57
Kumar A, Crawford K, Close L, Madison M,Lorenz J, et al. 1997. Rescue of cardiac alpha-actin-deficient mice by enteric smooth mus-cle gamma-actin.Proc. Natl. Acad. Sci. USA94:4406–11
Kunst G, Kress KR, Gruen M, Uttenweiler D,Gautel M, Fink RH. 2000. Myosin bind-ing protein C, a phosphorylation-dependentforce regulator in muscle that controls the at-tachment of myosin heads by its interactionwith myosin S2.Circ. Res.86:51–58
Kuroda S, Tokunaga C, Kiyohara Y, Higuchi O,Konishi H, et al. 1996. Protein-protein inter-action of zinc finger LIM domains with pro-tein kinase C.J. Biol. Chem.271:31029–32
Labeit S, Gautel M, Lakey A, Trinick J. 1992.Towards a molecular understanding of titin.EMBO J.11:1711–16
Labeit S, Kolmerer B. 1995a. The completeprimary structure of human nebulin and its

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
694 CLARK ET AL.
correlation to muscle structure.J. Mol. Biol.248:308–15
Labeit S, Kolmerer B. 1995b. Titins, giant pro-teins in charge of muscle ultrastructure andelasticity.Science270:293–96
Laing NG, Wilton SD, Akkari PA, Dorosz S,Boundy K, et al. 1995. A mutation in thealpha tropomyosin gene TPM3 associatedwith autosomal dominant nemaline myo-pathy NEM1.Nat. Genet.10:249
Lazarides E. 1980. Desmin and intermediate fil-aments in muscle cells.Results Probl. CellDiffer. 11:124–31
Leavis PC, Gergely J. 1984. Thin filamentproteins and thin filament-linked regulationof vertebrate muscle contraction.Crit. Rev.Biochem.16:235–305
Lehman W, Hatch V, Korman V, Rosol M,Thomas L, et al. 2000. Tropomyosin andactin isoforms modulate the localization oftropomyosin strands on actin filaments.J.Mol. Biol. 302:593–606
Lehman W, Rosol M, Tobacman LS, CraigR. 2001. Troponin organization on relaxedand activated thin filaments revealed by elec-tron microscopy and three-dimensional re-construction.J. Mol. Biol.307:739–44
Lenz-Bohme B, Wismar J, Fuchs S, ReifegersteR, Buchner E, et al. 1997. Insertional muta-tion of the Drosophila nuclear lamin Dm0gene results in defective nuclear envelopes,clustering of nuclear pore complexes, and ac-cumulation of annulate lamellae.J. Cell Biol.137:1001–16
Leung CL, Green KJ, Liem RK. 2002. Plakins:a family of versatile cytolinker proteins.Trends Cell Biol.12:37–45
Li H, Oberhauser AF, Redick SD, Carrion-Vazquez M, Erickson HP, Fernandez JM.2001. Multiple conformations of PEVKproteins detected by single-molecule tech-niques. Proc. Natl. Acad. Sci. USA98:10682–86
Li Z, Mericskay M, Agbulut O, Butler-BrowneG, Carlsson L, et al. 1997. Desmin isessential for the tensile strength and in-tegrity of myofibrils but not for myogeniccommitment, differentiation, and fusion of
skeletal muscle.J. Cell Biol. 139:129–44
Linke WA. 2000. Stretching molecular springs:elasticity of titin filaments in vertebrate stri-ated muscle.Histol. Histopathol.15:799–811
Linke WA, Rudy DE, Centner T, Gautel M, WittC, et al. 1999. I-band titin in cardiac muscle isa three-element molecular spring and is criti-cal for maintaining thin filament structure.J.Cell Biol. 146:631–44
Littlefield R, Almenar-Queralt A, Fowler VM.2001. Actin dynamics at pointed ends regu-lates thin filament length in striated muscle.Nat. Cell Biol.3:544–51
Littlefield R, Fowler VM. 1998. Defining actinfilament length in striated muscle: rulers andcaps or dynamic stability?Annu. Rev. CellDev. Biol.14:487–525
Liu YW, Cseresnyes Z, Randall WR, Schnei-der MF. 2001. Activity-dependent nucleartranslocation and intranuclear distribution ofNFATc in adult skeletal muscle fibers.J. CellBiol. 155:27–39
Loo DT, Kanner SB, Aruffo A. 1998. Filaminbinds to the cytoplasmic domain of the beta1-integrin. Identification of amino acids re-sponsible for this interaction.J. Biol. Chem.273:23304–12
Louis HA, Pino JD, Schmeichel KL, PomiesP, Beckerle MC. 1997. Comparison of threemembers of the cysteine-rich protein familyreveals functional conservation and divergentpatterns of gene expression.J. Biol. Chem.272:27484–91
Lukoyanova N, VanLoock MS, Orlova A,Galkin VE, Wang K, Egelman EH. 2002.Each actin subunit has three nebulin bindingsites. Implications for steric blocking.Curr.Biol. 12:383–88
Luo G, Herrera AH, Horowits R. 1999. Molec-ular interactions of N-RAP, a nebulin-relatedprotein of striated muscle myotendon junc-tions and intercalated disks.Biochemistry38:6135–43
Luo G, Zhang JQ, Nguyen TP, Herrera AH, Pa-terson B, Horowits R. 1997. Complete cDNAsequence and tissue localization of N-RAP, a

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 695
novel nebulin-related protein of striated mus-cle.Cell Motil. Cytoskeleton38:75–90
Luther P, Squire J. 1978. Three-dimensionalstructure of the vertebrate muscle M-region.J. Mol. Biol.125:313–24
MacGowan GA, Du C, Wieczorek DF, KoretskyAP. 2001. Compensatory changes in Ca(2+)and myocardial O(2) consumption in beta-tropomyosin transgenic hearts.Am. J. Phys-iol. Heart Circ. Physiol.281:H2539–H48
Machado C, Andrew DJ. 2000. D-titin: a giantprotein with dual roles in chromosomes andmuscles.J. Cell Biol.151:639–52
Maeda M, Holder E, Lowes B, Valent S, BiesRD. 1997. Dilated cardiomyopathy associ-ated with deficiency of the cytoskeletal pro-tein metavinculin.Circulation95:17–20
Mangan ME, Olmsted JB. 1996. A muscle-specific variant of microtubule-associatedprotein 4 (MAP4) is required in myogene-sis.Development122:771–81
Mardahl-Dumesnil M, Fowler VM. 2001. Thinfilaments elongate from their pointed endsduring myofibril assembly inDrosophilain-direct flight muscle.J. Cell Biol.155:1043–53
Marian AJ, Roberts R. 2001. The molecular ge-netic basis for hypertrophic cardiomyopathy.J. Mol. Cell Cardiol.33:655–70
Markwald RR. 1973. Distribution and relation-ship of precursor Z material to organizingmyofibrillar bundles in embryonic rat andhamster ventricular myocytes.J. Mol. Cell.Cardiol. 5:341–50
Maruyama K, Ebashi S. 1965. Alpha-actinin, anew structural protein from striated muscle.II. Action on actin.J. Biochem.58:13–19
Maruyama K, Hu DH, Suzuki T, Kimura S.1987. Binding of actin filaments to connectin.J. Biochem.101:1339–46
Maruyama K, Matsubara R, Natori Y, Nono-mura S, Kimura S, et al. 1977. Connectin,an elastic protein of muscle.J. Biochem.82:317–37
Masaki T, Takaiti O. 1974. M-protein.J.Biochem.75:367–80
Mayans O, van der Ven PFM, Wilm M, MuesA, Young P, et al. 1998. Structural basis
for activation of the titin kinase domainduring myofibrillogenesis.Nature395:863–69
Mayer U, Saher G, Fassler R, Bornemann A,Echtermeyer F, et al. 1997. Absence of inte-grin alpha 7 causes a novel form of musculardystrophy.Nat. Genet.17:318–23
McConnell BK, Jones KA, Fatkin D, Ar-royo LH, Lee RT, et al. 1999. Dilatedcardiomyopathy in homozygous myosin-binding protein-C mutant mice.J. Clin. In-vest.104:1235–44
McElhinny AS, Kolmerer B, Fowler VM,Labeit S, Gregorio CC. 2001. The N-terminalend of nebulin interacts with tropomodulin atthe pointed ends of the thin filaments.J. Biol.Chem.276:583–92
McElhinny AS, Labeit S, Gregorio CC. 2000.Probing the functional roles of titin ligands incardiac myofibril assembly and maintenance.Adv. Exp. Med. Biol.481:67–86
McElhinny AS, Sorimachi H, Labeit S, Grego-rio CC. 2002. Dual roles of MURF-1 in theregulation of sarcomeric M-line structure andin the potential modulation of gene expres-sion via its interaction with titin and GMEB-1. J. Cell Biol.157:125–36
McKay RT, Tripet BP, Hodges RS, SykesBD. 1997. Interaction of the second bind-ing region of troponin I with the regulatorydomain of skeletal muscle troponin C asdetermined by NMR spectroscopy.J. Biol.Chem.272:28494–500
Michele DE, Coutu P, Metzger JM. 2002.Divergent abnormal muscle relaxation byhypertrophic cardiomyopathy and nemalinemyopathy mutant tropomyosins.Physiol.Genom.103–11
Michele DE, Metzger JM. 2000. Physiologi-cal consequences of tropomyosin mutationsassociated with cardiac and skeletal my-opathies.J. Mol. Med.78:543–53
Millevoi S, Trombitas K, Kolmerer B, KostinS, Schaper J, et al. 1998. Characterization ofnebulette and nebulin and emerging conceptsof their roles for vertebrate Z-discs.J. Mol.Biol. 282:111–23
Milligan RA. 1996. Protein-protein interactions

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
696 CLARK ET AL.
in the rigor actomyosin complex.Proc. Natl.Acad. Sci. USA93:21–26
Milner DJ, Mavroidis M, Weisleder N,Capetanaki Y. 2000. Desmin cytoskeletonlinked to muscle mitochondrial distribu-tion and respiratory function.J. Cell Biol.150:1283–98
Milner DJ, Taffet GE, Wang X, Pham T, TamuraT, et al. 1999. The absence of desmin leads tocardiomyocyte hypertrophy and cardiac dila-tion with compromised systolic function.J.Mol. Cell. Cardiol.31:2063–76
Miosge N, Klenczar C, Herken R, Willem M,Mayer U. 1999. Organization of the myo-tendinous junction is dependent on the pres-ence of alpha7beta1 integrin.Lab. Invest.79:1591–99
Mislow JM, Kim MS, Davis DB, McNally EM.2002. Myne-1, a spectrin repeat transmem-brane protein of the myocyte inner nuclearmembrane, interacts with lamin A/C.J. CellSci.115:61–70
Misra AK, Menon NK, Mishra SK. 1992. Ab-normal distribution of desmin and vimentinin myofibers in adult onset myotubular myo-pathy.Muscle Nerve15:1246–52
Mizuno Y, Thompson TG, Guyon JR, LidovHG, Brosius M, et al. 2001. Desmuslin, anintermediate filament protein that interactswith alpha-dystrobrevin and desmin.Proc.Natl. Acad. Sci. USA98:6156–61
Mogami K, O’Donnell PT, Bernstein SI, WrightTR, Emerson CP Jr. 1986. Mutations of theDrosophila myosin heavy-chain gene: ef-fects on transcription, myosin accumulation,and muscle function.Proc. Natl. Acad. Sci.USA83:1393–97
Moncman CL, Wang K. 2002. Targeted disrup-tion of nebulette protein expression alters car-diac myofibril assembly and function.Exp.Cell Res.273:204–18
Moorthy S, Chen L, Bennett V. 2000.Caenorhabditis elegansbeta-G spectrin isdispensable for establishment of epithelialpolarity, but essential for muscular andneuronal function.J. Cell Biol. 149:915–30
Moreira ES, Wiltshire TJ, Faulkner G, Nil-
foroushan A, Vainzof M, et al. 2000. Limb-girdle muscular dystrophy type 2G is causedby mutations in the gene encoding thesarcomeric protein telethonin.Nat. Genet.24:163–66
Morisaki T, Gross M, Morisaki H, Pongratz D,Zollner N, Holmes EW. 1992. Molecular ba-sis of AMP deaminase deficiency in skeletalmuscle.Proc. Natl. Acad. Sci. USA89:6457–61
Moriyama M, Tsukamoto Y, Fujiwara M,Kondo G, Nakada C, et al. 2001. Identifi-cation of a novel human ankyrin-repeatedprotein homologous to CARP.Biochem. Bio-phys. Res. Commun.285:715–23
Morris GE. 2001. The role of the nuclear enve-lope in Emery-Dreifuss muscular dystrophy.Trends Mol. Med.7:572–77
Mudry RE, Bondad M, Perry CN, FowlerVM, Gregorio CC. 2001. The interactionof tropomyosin with tropomodulin preventsdepolymerization of thin filaments cardiacmuscle.Mol. Biol. Cell12:S292a
Mues A, van der Ven PFM, Young P, Furst DO,Gautel M. 1998. Two immunoglobulin-likedomains of the Z-disc portion of titin inter-act in a conformation-dependent way withtelethonin.FEBS Lett.428:111–14
Muhle-Goll C, Habeck M, Cazorla O, NilgesM, Labeit S, Granzier H. 2001. Structural andfunctional studies of titin’s fn3 modules re-veal conserved surface patterns and bindingto myosin S1—a possible role in the Frank-Starling mechanism of the heart.J. Mol. Biol.313:431–47
Muntoni F, Cau M, Ganau A, Congiu R, ArvediG, et al. 1993. Brief report: deletion of thedystrophin muscle-promoter region associ-ated with X-linked dilated cardiomyopathyN. Engl. J. Med.329:921–25
Murphy AM, Kogler H, Georgakopoulos D,McDonough JL, Kass DA, et al. 2000. Trans-genic mouse model of stunned myocardium.Science287:488–91
Muthuchamy M, Boivin GP, Grupp IL, Wiec-zorek DF. 1998. Beta-tropomyosin overex-pression induces severe cardiac abnormali-ties.J. Mol. Cell. Cardiol.30:1545–47

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 697
Nadal-Ginard B, Smith CW, Patton JG, Breit-bart RE. 1991. Alternative splicing is an ef-ficient mechanism for the generation of pro-tein diversity: contractile protein genes as amodel system.Adv. Enzyme Regul.31:261–86
Nave R, Furst DO, Weber K. 1989. Visualiza-tion of the polarity of isolated titin molecules:a single globular head on a long thin rod asthe M band anchoring domain?J. Cell Biol.109:2177–87
Newey SE, Howman EV, Ponting CP, BensonMA, Nawrotzki R, et al. 2001. Syncoilin,a novel member of the intermediate fila-ment superfamily that interacts with alpha-dystrobrevin in skeletal muscle.J. Biol.Chem.276:6645–55
Nigro V, de Sa Moreira E, Piluso G, Vain-zof M, Belsito A, et al. 1996. Autosomalrecessive limb-girdle muscular dystrophy,LGMD2F, is caused by a mutation in thedelta-sarcoglycan gene.Nat. Genet.14:195–98
Noguchi J, Yanagisawa M, Imamura M, Ka-suya Y, Sakurai T, et al. 1992. Completeprimary structure and tissue expression ofchicken pectoralis M-protein.J. Biol. Chem.267:20302–10
Norgett EE, Hatsell SJ, Carvajal-Huerta L,Cabezas JC, Common J, et al. 2000. Re-cessive mutation in desmoplakin disruptsdesmoplakin-intermediate filament interac-tions and causes dilated cardiomyopathy,woolly hair and keratoderma.Hum. Mol.Genet.9:2761–66
North KN, Beggs AH. 1996. Deficiency ofa skeletal muscle isoform of alpha-actinin(alpha-actinin-3) in merosin-positive con-genital muscular dystrophy.Neuromuscul.Disord.6:229–35
North KN, Yang N, Wattanasirichaigoon D,Mills M, Easteal S, Beggs AH. 1999. Acommon nonsense mutation results in alpha-actinin-3 deficiency in the general popula-tion. Nat. Genet.21:353–54
Nowak KJ, Wattanasirichaigoon D, Goebel HH,Wilce M, Pelin K, et al. 1999. Mutations inthe skeletal muscle alpha-actin gene in pa-
tients with actin myopathy and nemaline my-opathy.Nat. Genet.23:208–12
Obermann WMJ, Gautel M, Steiner F, van derVen PFM, Weber K, Furst DO. 1996. Thestructure of the sarcomeric M band: local-ization of defined domains of myomesin, Mprotein, and the 250 kD carboxy terminal re-gion of titin by immunoelectron microscopy.J. Cell Biol.134:1441–53
Obermann WMJ, Gautel M, Weber K, FurstDO. 1997. Molecular structure of the sar-comeric M band: mapping of titin and myosinbinding domains in myomesin and the identi-fication of a potential regulatory phosphory-lation site in myomesin.EMBO J.16:211–20
Obermann WMJ, van der Ven PFM, SteinerF, Weber K, Furst DO. 1998. Mapping ofa myosin-binding domain and a regulatoryphosphorylation site in M-protein, a struc-tural protein of the sarcomeric M band.Mol.Biol. Cell 9:829–40
Ohlendieck K, Campbell KP. 1991. Dystrophin-associated proteins are greatly reduced inskeletal muscle from mdx mice.J. Cell Biol.115:1685–94
Ohtsuka H, Yajima H, Maruyama K, KimuraS. 1997. The N-terminal Z repeat 5 of con-nectin/titin binds to the C-terminal region ofalpha-actinin.Biochem. Biophys. Res. Com-mun.235:1–3
Ojima K, Lin ZX, Zhang ZQ, Hijikata T,Holtzer S, et al. 1999. Initiation and matura-tion of I-Z-I bodies in the growth tips of trans-fected myotubes.J. Cell Sci.112:4101–12
Okagaki T, Weber FE, Fischman DA, VaughanKT, Mikawa T, Reinach FC. 1993. The majormyosin-binding domain of skeletal muscleMyBP-C (C protein) resides in the COOH-terminal, immunoglobulin C2 motif.J. CellBiol. 123:619–26
Olson EN, Williams RS. 2000. Calcineurinsignaling and muscle remodeling.Cell101:689–92
Olson TM, Doan TP, Kishimoto NY, WhitbyFG, Ackerman MJ, Fananapazir L. 2000. In-herited and de novo mutations in the cardiacactin gene cause hypertrophic cardiomyopa-thy. J. Mol. Cell. Cardiol.32:1687–94

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
698 CLARK ET AL.
Olson TM, Illenberger S, Kishimoto NY, Hut-telmaier S, Keating MT, Jockusch BM. 2002.Metavinculin mutations alter actin interac-tion in dilated cardiomyopathy.Circulation105:431–37
Olson TM, Kishimoto NY, Whitby FG, MichelsVV. 2001. Mutations that alter the surfacecharge of alpha-tropomyosin are associatedwith dilated cardiomyopathy.J. Mol. Cell.Cardiol. 33:723–32
Olson TM, Michels VV, Thibodeau SN, TaiYS, Keating MT. 1998. Actin mutations indilated cardiomyopathy, a heritable form ofheart failure.Science280:750–52
Ono Y, Shimada H, Sorimachi H, Richard I,Saido TC, et al. 1998. Functional defectsof a muscle-specific calpain, p94, caused bymutations associated with limb-girdle mus-cular dystrophy type 2A.J. Biol. Chem.273:17073–78
Pallavicini A, Kojic S, Bean C, Vainzof M, Sala-mon M, et al. 2001. Characterization of hu-man skeletal muscle Ankrd2.Biochem. Bio-phys. Res. Commun.285:378–86
Papa I, Astier C, Kwiatek O, Raynaud F, Bon-nal C, et al. 1999. Alpha actinin-CapZ, ananchoring complex for thin filaments in Z-line. J. Muscle Res. Cell Motil.20:187–97
Parast MM, Otey CA. 2000. Characterizationof palladin, a novel protein localized tostress fibers and cell adhesions.J. Cell Biol.150:643–56
Pardo JV, Siliciano JD, Craig SW. 1983a. Vin-culin is a component of an extensive net-work of myofibril-sarcolemma attachmentregions in cardiac muscle fibers.J. Cell Biol.97:1081–88
Pardo JV, Siliciano JD, Craig SW. 1983b.A vinculin-containing cortical lattice inskeletal muscle: transverse lattice elements(“costameres”) mark sites of attachment be-tween myofibrils and sarcolemma.Proc.Natl. Acad. Sci. USA80:1008–12
Parry DA, Squire JM. 1973. Structural role oftropomyosin in muscle regulation: analysisof the X-ray diffraction patterns from relaxedand contracting muscles.J. Mol. Biol.75:33-55
Pashmforoush M, Pomies P, Peterson KL,Kubalak S, Ross J Jr, et al. 2001. Adult micedeficient in actinin-associated LIM-domainprotein reveal a developmental pathway forright ventricular cardiomyopathy.Nat. Med.7:591–97
Passier R, Richardson JA, Olson EN. 2000. Or-acle, a novel PDZ-LIM domain protein ex-pressed in heart and skeletal muscle.Mech.Dev.92:277–84
Peckham M, Young P, Gautel M. 1997.Constitutive and variable regions of Z-disk titin/connectin in myofibril formation:a dominant-negative screen.Cell Struct.Funct.22:95–101
Pelin K, Hilpela P, Donner K, Sewry C, AkkariPA, et al. 1999. Mutations in the nebulin geneassociated with autosomal recessive nema-line myopathy.Proc. Natl. Acad. Sci. USA96:2305–10
Perhonen M, Sharp WW, Russell B. 1998. Mi-crotubules are needed for dispersal of alpha-myosin heavy chain mRNA in rat neona-tal cardiac myocytes.J. Mol. Cell Cardiol.30:1713–22
Perry SV. 2001. Vertebrate tropomyosin: dis-tribution, properties and function.J. MuscleRes. Cell Motil.22:5–49
Person V, Kostin S, Suzuki K, Labeit S,Schaper J. 2000. Antisense oligonucleotideexperiments elucidate the essential role oftitin in sarcomerogenesis in adult rat car-diomyocytes in long-term culture.J. Cell Sci.113(21):3851–59
Petrof BJ, Shrager JB, Stedman HH, Kelly AM,Sweeney HL. 1993. Dystrophin protects thesarcolemma from stresses developed duringmuscle contraction.Proc. Natl. Acad. Sci.USA90:3710–14
Pfuhl M, Winder SJ, Pastore A. 1994. Nebu-lin, a helical actin binding protein.EMBO J.13:1782–89
Pham CG, Harpf AE, Keller RS, Vu HT, ShaiSY, et al. 2000. Striated muscle-specificbeta(1D)-integrin and FAK are involved incardiac myocyte hypertrophic response path-way. Am. J. Physiol. Heart Circ. Physiol.279:H2916–H26

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 699
Pieper FR, Raats JM, Schaart G, Dunia I, vander Kemp A, et al. 1995. Disruption of vi-mentin intermediate filaments in transgenicmice by expression of a dominant-negativemutant desmin subunit.Eur. J. Cell Biol.68:355–68
Pittenger MF, Kazzaz JA, Helfman DM.1994. Functional properties of non-muscletropomyosin isoforms.Curr. Opin. Cell Biol.6:96–104
Pomies P, Louis HA, Beckerle MC. 1997.CRP1, a LIM domain protein implicated inmuscle differentiation, interacts with alpha-actinin.J. Cell Biol.139:157–68
Pomies P, Macalma T, Beckerle MC. 1999. Pur-ification and characterization of an alpha-actinin-binding PDZ-LIM protein that is up-regulated during muscle differentiation.J.Biol. Chem.274:29242–50
Poon E, Howman EV, Newey SE, Davies KE.2002. Association of syncoilin and desmin:linking intermediate filament proteins to thedystrophin-associated protein complex.J.Biol. Chem.277:3433–39
Porter GA, Dmytrenko GM, Winkelmann JC,Bloch RJ. 1992. Dystrophin colocalizes withbeta-spectrin in distinct subsarcolemmal do-mains in mammalian skeletal muscle.J. CellBiol. 117:997–1005
Porter GA, Scher MG, Resneck WG, Porter NC,Fowler VM, Bloch RJ. 1997. Two popula-tions of beta-spectrin in rat skeletal muscle.Cell Motil. Cytoskelet.37:7–19
Potter JD, Gergely J. 1974. Troponin, tropo-myosin, and actin interactions in the Ca2+
regulation of muscle contraction.Biochem-istry 13:2697–703
Potter JD, Sheng Z, Pan BS, Zhao J. 1995. Adirect regulatory role for troponin T and adual role for troponin C in the Ca2+ regu-lation of muscle contraction.J. Biol. Chem.270:2557–62
Prabhakar R, Boivin GP, Grupp IL, Hoit B,Arteaga G, et al. 2001. A familial hyper-trophic cardiomyopathy alpha-tropomyosinmutation causes severe cardiac hypertrophyand death in mice.J. Mol. Cell Cardiol.33:1815–28
Price MG. 1987. Skelemins: cytoskeletal pro-teins located at the periphery of M-discsin mammalian striated muscle.J. Cell Biol.104:1325–36
Price MG, Lazarides E. 1983. Expressionof intermediate filament-associated proteinsparanemin and synemin in chicken develop-ment.J. Cell Biol.97:1860–74
Raharjo WH, Enarson P, Sullivan T, StewartCL, Burke B. 2001. Nuclear envelope de-fects associated with LMNA mutations causedilated cardiomyopathy and Emery-Dreifussmuscular dystrophy.J. Cell Sci.114:4447–57
Rayment I, Holden HM, Whittaker M, YohnCB, Lorenz M, et al. 1993. Structure of theactin-myosin complex and its implicationsfor muscle contraction.Science261:58–65
Reggiani C, Bottinelli R, Stienen GJ. 2000. Sar-comeric myosin isoforms: fine tuning of amolecular motor.News Physiol. Sci.15:26–33
Reinach FC, Masaki T, Shafiz S, Obinata T, Fis-chman DA. 1982. Isoforms of C-protein inadult chicken skeletal muscle: detection withmonoclonal antibodies.J. Cell Biol. 95:78–84
Reipert S, Steinbock F, Fischer I, BittnerRE, Zeold A, Wiche G. 1999. Associationof mitochondria with plectin and desminintermediate filaments in striated muscle.Exp. Cell Res.252:479–91
Rethinasamy P, Muthuchamy M, Hewett T,Boivin G, Wolska BM, et al. 1998. Molec-ular and physiological effects of alpha-tropomyosin ablation in the mouse.Circ. Res.82:116–23
Richard I, Broux O, Allamand V, FougerousseF, Chiannilkulchai N, et al. 1995. Muta-tions in the proteolytic enzyme calpain 3cause limb-girdle muscular dystrophy type2A. Cell 81:27–40
Richard I, Roudaut C, Marchand S, Baghdigu-ian S, Herasse M, et al. 2000. Loss of calpain3 proteolytic activity leads to muscular dys-trophy and to apoptosis-associated I kappa Balpha/nuclear factor kappa B pathway pertur-bation in mice.J. Cell Biol.151:1583–90

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
700 CLARK ET AL.
Robinson TF, Winegrad S. 1979. The measure-ment and dynamic implications of thin fil-ament lengths in heart muscle.J. Physiol.286:617–19
Roopnarine O, Leinwand LA. 1998. Functionalanalysis of myosin mutations that causefamilial hypertrophic cardiomyopathy.Bio-phys. J.75:3023–30
Root DD, Wang K. 2001. High-affinity actin-binding nebulin fragments influence the ac-toS1 complex.Biochemistry40:1171–86
Royuela M, Fraile B, Paniagua R. 1997.Nebulin-like protein in the earthwormEise-nia foetida. Immunocytochemical electronmicroscopic study and western blot analy-sis of several muscle cell types.Eur. J. CellBiol. 73:276–80
Ruiz P, Brinkmann V, Ledermann B, BehrendM, Grund C, et al. 1996. Targeted mutationof plakoglobin in mice reveals essential func-tions of desmosomes in the embryonic heart.J. Cell Biol.135:215–25
Rundell KW, Tullson PC, Terjung RL. 1992.Altered kinetics of AMP deaminase bymyosin binding.Am. J. Physiol. Cell Physiol.263:C294–C99
Russell MW, Raeker MO, Korytkowski KA,Sonneman KJ. 2002. Identification, tissueexpression and chromosomal localization ofhuman Obscurin-MLCK, a member of thetitin and Dbl families of myosin light chainkinases.Gene282:237–46
Rybakova IN, Patel JR, Ervasti JM. 2000. Thedystrophin complex forms a mechanicallystrong link between the sarcolemma andcostameric actin.J. Cell Biol.150:1209–14
Sadler I, Crawford AW, Michelsen JW, Beck-erle MC. 1992. Zyxin and cCRP: two inter-active LIM domain proteins associated withthe cytoskeleton.J. Cell Biol.119:1573–87
Saitoh O, Arai T, Obinata T. 1988. Distributionof microtubules and other cytoskeletal fila-ments during myotube elongation as revealedby fluorescence microscopy.Cell Tissue Res.252:263–73
Sakamoto A, Ono K, Abe M, Jasmin G, EkiT, et al. 1997. Both hypertrophic and dilatedcardiomyopathies are caused by mutation of
the same gene, delta-sarcoglycan, in hamster:an animal model of disrupted dystrophin-associated glycoprotein complex.Proc. Natl.Acad. Sci. USA94:13873–78
Salmikangas P, Mykkanen OM, Gronholm M,Heiska L, Kere J, Carpen O. 1999. Myotilin,a novel sarcomeric protein with two Ig-likedomains, is encoded by a candidate gene forlimb-girdle muscular dystrophy.Hum. Mol.Genet.8:1329–36
Sanger JW, Sanger JM. 2001. Fishing out pro-teins that bind to titin.J. Cell Biol.154:21–24
Sawada Y, Sheetz MP. 2002. Force transduc-tion by triton cytoskeletons.J. Cell Biol.156:609–15
Schafer DA, Hug C, Cooper JA. 1995. Inhibi-tion of CapZ during myofibrillogenesis al-ters assembly of actin filaments.J. Cell Biol.128:61–70
Schafer DA, Jennings PB, Cooper JA. 1996.Dynamics of capping protein and actin as-sembly in vitro: uncapping barbed endsby polyphosphoinositides.J. Cell Biol.135:169–79
Schiaffino S, Reggiani C. 1994. Myosin iso-forms in mammalian skeletal muscle.J. Appl.Physiol.77:493–501
Schmitz S, Clayton J, Nongthomba U, Prinz H,Veigel C, et al. 2000.Drosophila ACT88Findirect flight muscle-specific actin is notN-terminally acetylated: a mutation in N-terminal processing affects actin function.J.Mol. Biol. 295:1201–10
Schroder R, Furst DO, Klasen C, Reimann J,Herrmann H, van der Ven PFM. 2000. Asso-ciation of plectin with Z-discs is a prerequi-site for the formation of the intermyofibrillardesmin cytoskeleton.Lab. Invest.80:455–64
Schultheiss T, Choi J, Lin ZX, Dilullo C,Cohen-Gould L, et al. 1992. A sarcomericalpha-actinin truncated at the carboxyl endinduces the breakdown of stress fibers inPtK2 cells and the formation of nemaline-likebodies and breakdown of myofibrils in my-otubes.Proc. Natl. Acad. Sci. USA82:9282–86
Seidman JG, Seidman C. 2001. The geneticbasis for cardiomyopathy: from mutation

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 701
identification to mechanistic paradigms.Cell104:557–67
Seiler SH, Fischman DA, Leinwand LA. 1996.Modulation of myosin filament organizationby C-protein family members.Mol. Biol. Cell7:113–27
Sejersen T, Lendahl U. 1993. Transient expres-sion of the intermediate filament nestin dur-ing skeletal muscle development.J. Cell Sci.106:1291–300
Sellers JR. 2000. Myosins: a diverse superfam-ily. Biochim. Biophys. Acta1496:3–22
Sellers JR, Goodson HV. 1995. Motor proteins2: myosin.Protein Profile2:1323–423
Shah SB, Su FC, Jordan K, Milner DJ, Friden J,et al. 2002. Evidence for increased myofib-rillar mobility in desmin-null mouse skeletalmuscle.J. Exp. Biol.205:321–25
Shear CR, Bloch RJ. 1985. Vinculin in subsar-colemmal densities in chicken skeletal mus-cle: localization and relationship to intracel-lular and extracellular structures.J. Cell Biol.101:240–56
Sheng Z, Strauss WL, Francois JM, Potter JD.1990. Evidence that both Ca(2+)-specificsites of skeletal muscle TnC are required forfull activity. J. Biol. Chem.265:21554–60
Shyy JY, Chien S. 1997. Role of integrins incellular responses to mechanical stress andadhesion.Curr. Opin. Cell Biol.9:707–13
Sicinski P, Geng Y, Ryder-Cook AS, BarnardEA, Darlison MG, Barnard PJ. 1989. Themolecular basis of muscular dystrophy inthe mdx mouse: a point mutation.Science244:1578–80
Sjoberg G, Edstrom L, Lendahl U, SejersenT. 1994. Myofibers from Duchenne/Beckermuscular dystrophy and myositis express theintermediate filament nestin.J. Neuropathol.Exp. Neurol.53:416–23
Solaro RJ, Van Eyk J. 1996. Altered interactionsamong thin filament proteins modulate car-diac function.J. Mol. Cell Cardiol.28:217–30
Soler AP, Knudsen KA. 1994. N-cadherin in-volvement in cardiac myocyte interactionand myofibrillogenesis.Dev. Biol.162:9–17
Sorimachi H, Freiburg A, Kolmerer B, Ishiura
S, Stier G, et al. 1997. Tissue-specific expres-sion and alpha-actinin binding properties ofthe Z-disc titin: implications for the natureof vertebrate Z-discs.J. Mol. Biol.270:688–95
Sorimachi H, Tsukahara T, Okada-Ban M,Sugita H, Ishiura S, Suzuki K. 1995. Identifi-cation of a third ubiquitous calpain species—chicken muscle expresses four distinct cal-pains.Biochim. Biophys. Acta1261:381–93
Soteriou A, Gamage M, Trinick J. 1993. A sur-vey of interactions made by the giant proteintitin. J. Cell Sci.14:119–23
Spencer JA, Eliazer S, Ilaria RL Jr, Richard-son JA, Olson EN. 2000. Regulation ofmicrotubule dynamics and myogenic differ-entiation by MURF, a striated muscle RING-finger protein.J. Cell Biol.150:771–84
Stanchi F, Corso V, Scannapieco P, IevolellaC, Negrisolo E, et al. 2000. TUBA8: A newtissue-specific isoform of alpha-tubulin thatis highly conserved in human and mouse.Biochem. Biophys. Res. Commun.270:1111–18
Steinberg MS, McNutt PM. 1999. Cadherinsand their connections: adhesion junctionshave broader functions.Curr. Opin. Cell Biol.11:554–60
Steinmetz MO, Stoffler D, Hoenger A, BremerA, Aebi U. 1997. Actin: from cell biology toatomic detail.J. Struct. Biol.119:295–320
Straub V, Duclos F, Venzke DP, Lee JC, Cut-shall S, et al. 1998. Molecular pathogen-esis of muscle degeneration in the delta-sarcoglycan-deficient hamster.Am. J. Pathol.153:1623–30
Stronach BE, Renfranz PJ, Lilly B, BeckerleMC. 1999. Muscle LIM proteins are asso-ciated with muscle sarcomeres and requiredMEF2 for their expression duringDroso-phila myogenesis.Mol. Biol. Cell 10:2329–42
Stronach BE, Siegrist SE, Beckerle MC.1996. Two muscle-specific LIM proteins inDrosophila. J. Cell Biol.134:1179–95
Sullivan T, Escalante-Alcalde D, Bhatt H, An-ver M, Bhat N, et al. 1999. Loss of A-type lamin expression compromises nuclear

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
702 CLARK ET AL.
envelope integrity leading to muscular dys-trophy.J. Cell Biol.147:913–20
Sussman MA, Baque S, Uhm CS, Daniels MP,Price RL, et al. 1998a. Altered expression oftropomodulin in cardiomyocytes disrupts thesarcomeric structure of myofibrils.Circ. Res.82:94–105
Sussman MA, Welch S, Cambon N, Klevit-sky R, Hewett TE, et al. 1998b. Myofibrildegeneration caused by tropomodulin over-expression leads to dilated cardiomyopathyin juvenile mice.J. Clin. Invest.101:51–61
Sweeney HL, Feng HS, Yang Z, Watkins H.1998. Functional analyses of troponin T mu-tations that cause hypertrophic cardiomyopa-thy: insights into disease pathogenesis andtroponin function.Proc. Natl. Acad. Sci. USA95:14406–10
Syska H, Wilkinson JM, Grand RJ, Perry SV.1976. The relationship between biologicalactivity and primary structure of troponinI from white skeletal muscle of the rabbit.Biochem. J.153:375–87
Tagawa K, Taya C, Hayashi Y, NakagawaM, Ono Y, et al. 2000. Myopathy pheno-type of transgenic mice expressing activesite-mutated inactive p94 skeletal muscle-specific calpain, the gene product responsiblefor limb girdle muscular dystrophy type 2A.Hum. Mol. Genet.9:1393–402
Takada F, Vander Woude DL, Tong HQ,Thompson TG, Watkins SC, et al. 2001.Myozenin: an alpha-actinin- and gamma-filamin-binding protein of skeletal muscle Zlines. Proc. Natl. Acad. Sci. USA98:1595–600
Takahashi M, Tsutsui H, Tagawa H, Igarashi-Saito K, Imanaka-Yoshida K, Takeshita A.1998. Microtubules are involved in earlyhypertrophic responses of myocardium dur-ing pressure overload.Am. J. Physiol. HeartCirc. Physiol.275:H341–H48
Takahashi-Yanaga F, Ohtsuki I, Morimoto S.2001. Effects of troponin T mutations in fa-milial hypertrophic cardiomyopathy on reg-ulatory functions of other troponin subunits.J. Biochem.130:127–31
Takano-Ohmuro H, Goldfine SM, Kojima T,Obinata T, Fischman DA. 1989. Size andcharge heterogeneity of C-protein isoformsin avian skeletal muscle. Expression of sixdifferent isoforms in chicken muscle.J. Mus-cle Res. Cell Motil.10:369–78
Tardiff JC, Hewett TE, Factor SM, VikstromKL, Robbins J, Leinwand LA. 2000. Expres-sion of the beta (slow)-isoform of MHC in theadult mouse heart causes dominant-negativefunctional effects.Am. J. Physiol. Heart Circ.Physiol.278:H412–H19
Tardiff JC, Hewett TE, Palmer BM, Olsson C,Factor SM, et al. 1999. Cardiac troponin Tmutations result in allele-specific phenotypesin a mouse model for hypertrophic cardio-myopathy.J. Clin. Invest.104:469–81
Theriault JR, Charette SJ, Lambert H, LandryJ. 1999. Cloning and characterization ofhGMEB1, a novel glucocorticoid modula-tory element binding protein.FEBS Lett.452:170–76
Thierfelder L, Watkins H, MacRae C, Lamas R,McKenna W, et al. 1994. Beta-tropomyosinand cardiac troponin T mutations cause fa-milial hypertrophic cardiomyopathy: a dis-ease of the sarcomere.Cell 77:701–12
Thomas GH. 2001. Spectrin: the ghost in themachine.BioEssays23:152–60
Thompson TG, Chan YM, Hack AA, BrosiusM, Rajala M, et al. 2000. Filamin 2 (FLN2): amuscle-specific sarcoglycan interacting pro-tein.J. Cell Biol.148:115–26
Tidball JG. 1991. Force transmission acrossmuscle cell membranes.J. Biomech.24:43–52
Tidball JG, Law DJ. 1991. Dystrophin is re-quired for normal thin filament-membraneassociations at myotendinous junctions.Am.J. Pathol.138:17–21
Tidball JG, O’Halloran T, Burridge K. 1986.Talin at myotendinous junctions.J. Cell Biol.103:1465–72
Tkatchenko AV, Pietu G, Cros N, Gannoun-Zaki L, Auffray C, et al. 2001. Identificationof altered gene expression in skeletal musclesfrom Duchenne muscular dystrophy patients.Neuromuscul. Disord.11:269–77

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 703
Towbin JA, Bowles NE. 2002. The failing heart.Nature415:227–33
Tripet B, Van Eyk JE, Hodges RS. 1997. Map-ping of a second actin-tropomyosin and asecond troponin C binding site within the Cterminus of troponin I, and their importancein the Ca2+-dependent regulation of musclecontraction.J. Mol. Biol.271:728–50
Trombitas K, Freiburg A, Centner T, Labeit S,Granzier H. 1999. Molecular dissection ofN2B cardiac titin’s extensibility.Biophys. J.77:3189–96
Trombitas K, Greaser ML, Pollack GH. 1997.Interaction between titin and thin filamentsin intact cardiac muscle.J. Muscle Res. CellMotil. 18:345–51
Tsutsui H, Ishihara K, Cooper G. 1993. Cy-toskeletal role in the contractile dysfunc-tion of hypertrophied myocardium.Science260:682–87
Tubridy N, Fontaine B, Eymard B. 2001. Con-genital myopathies and congenital musculardystrophies.Curr. Opin. Neurol.14:575–82
Turnacioglu KK, Mittal B, Dabiri GA, SangerJM, Sanger JW. 1997. An N-terminal frag-ment of titin coupled to green fluorescent pro-tein localizes to the Z-bands in living musclecells: overexpression leads to myofibril dis-assembly.Mol. Biol. Cell8:705–17
Turner DM, Walker JB. 1987. Enhanced abil-ity of skeletal muscle containing cyclocrea-tine phosphate to sustain ATP levels duringischemia following beta-adrenergic stimula-tion. J. Biol. Chem.262:6605–9
Tuvia S, Buhusi M, Davis L, Reedy M, BennettV. 1999 Ankyrin-B is required for intracellu-lar sorting of structurally diverse Ca2+ hom-eostasis proteinsJ. Cell Biol.147:995–1008
Vale RD, Milligan RA. 2000. The way thingsmove: looking under the hood of molecularmotor proteins.Science288:88–95
Valle G, Faulkner G, De Antoni A, PacchioniB, Pallavicini A, et al. 1997. Telethonin, anovel sarcomeric protein of heart and skele-tal muscle.FEBS Lett.415:163–68
Vandekerckhove J, Weber K. 1979. The com-plete amino acid sequence of actins frombovine aorta, bovine heart, bovine fast skele-
tal muscle, and rabbit slow skeletal muscle.A protein-chemical analysis of muscle actindifferentiation.Differentiation14:123–33
van der Flier A, Kuikman I, Kramer D, GeertsD, Kreft M, et al. 2002. Different splice vari-ants of filamin-B affect myogenesis, subcel-lular distribution, and determine binding tointegrin β subunits.J. Cell Biol. 156:361–76
van der Ven PFM, Bartsch JW, Gautel M,Jockusch H, Furst DO. 2000a. A functionalknock-out of titin results in defective myofib-ril assembly.J. Cell Sci.113:1405–14
van der Ven PFM, Obermann WM, Weber K,Furst DO. 1996. Myomesin, M-protein andthe structure of the sarcomeric M-band.Adv.Biophys.33:91–99
van der Ven PFM, Wiesner S, Salmikangas P,Auerbach D, Himmel M, et al. 2000b. Indi-cations for a novel muscular dystrophy path-way. gamma-filamin, the muscle-specific fil-amin isoform, interacts with myotilin.J. CellBiol. 151:235–48
van Deursen J, Heerschap A, Oerlemans F,Ruitenbeek W, Jap P, et al. 1993. Skeletalmuscles of mice deficient in muscle creatinekinase lack burst activity.Cell 74:621–31
van Straaten M, Goulding D, Kolmerer B,Labeit S, Clayton J, et al. 1999. Associationof kettin with actin in the Z-disc of insectflight muscle.J. Mol. Biol.285:1549–62
Vaughan KT, Weber FE, Einheber S, FischmanDA. 1993a. Molecular cloning of chickenmyosin-binding protein (MyBP) H (86-kDaprotein) reveals extensive homology withMyBP-C (C-protein) with conserved im-munoglobulin C2 and fibronectin type IIImotifs.J. Biol. Chem.268:3670–76
Vaughan KT, Weber FE, Ried T, Ward DC,Reinach FC, Fischman DA. 1993b. Humanmyosin-binding protein H (MyBP-H): com-plete primary sequence, genomic organi-zation, and chromosomal localization.Ge-nomics16:34–40
Ventura-Clapier R, Veksler V, Hoerter JA. 1994.Myofibrillar creatine kinase and cardiac con-traction.Mol. Cell. Biochem.133/134:125–44

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
704 CLARK ET AL.
Vicart P, Caron A, Guicheney P, Li Z, Pre-vost MC, et al. 1998. A missense mu-tation in the alphaB-crystallin chaperonegene causes a desmin-related myopathy.Nat.Genet.20:92–95
Vigoreaux JO, Saide JD, Pardue ML. 1991.Structurally different Drosophila striatedmuscles utilize distinct variants of Z-band-associated proteins.J. Muscle Res. CellMotil. 12:340–54
Vinkemeier U, Obermann W, Weber K, FurstDO. 1993. The globular head domain of titinextends into the center of the sarcomericM band. cDNA cloning, epitope mappingand immunoelectron microscopy of two titin-associated proteins.J. Cell Sci.106:319–30
Volk T, Fessler LI, Fessler JH. 1990. A role forintegrin in the formation of sarcomeric cyto-architecture.Cell 63:525–36
Volk T, Geiger B. 1986. A-CAM: a 135-kD re-ceptor of intercellular adherens junctions. II.Antibody-mediated modulation of junctionformation.J. Cell Biol.103:1451–64
von Arx P, Bantle S, Soldati T, Perriard JC.1995. Dominant-negative effect of cytoplas-mic actin isoproteins on cardiomyocyte cyto-architecture and function.J. Cell Biol.131:1759–73
Wang K. 1996. Titin/connectin and nebulin: gi-ant protein rulers of muscle structure andfunction.Adv. Biophys.33:123–34
Wang K, Forbes JG, Jin AJ. 2001. Singlemolecule measurements of titin elasticity.Prog. Biophys. Mol. Biol.77:1–44
Wang K, Knipfer M, Huang QQ, van HeerdenA, Hsu LC, et al. 1996. Human skeletal mus-cle nebulin sequence encodes a blueprint forthin filament architecture. Sequence motifsand affinity profiles of tandem repeats andterminal SH3.J. Biol. Chem.271:4304–14
Wang K, McClure J, Tu A. 1979. Titin: majormyofibrillar components of striated muscle.Proc. Natl. Acad. Sci. USA76:3698–702
Wasser M, Chia W. 2000. The EAST protein ofDrosophila controls an expandable nuclearendoskeleton.Nat. Cell Biol.2:268–75
Watkins H, McKenna WJ, Thierfelder L, SukHJ, Anan R, et al. 1995. Mutations in the
genes for cardiac troponin T and alpha-tropomyosin in hypertrophic cardiomyo-pathy.N. Engl. J. Med.332:1058–64
Weber A, Murray JM. 1973. Molecular controlmechanisms in muscle contraction.Physiol.Rev.3:612–73
Weber A, Pennise CC, Babcock GG, FowlerVM. 1994. Tropomodulin caps the pointedends of actin filaments.J. Cell Biol. 127:1627–35
Webster DR, Patrick DL. 2000. Beating rateof isolated neonatal cardiomyocytes is reg-ulated by the stable microtubule subset.Am.J. Physiol. Heart Circ. Physiol.278:H1653–H61
Wegner A. 1979. Equilibrium of the actin-tropomyosin interaction.J. Mol. Biol. 131:839–53
Wegner A. 1982. Kinetic analysis of actin as-sembly suggests that tropomyosin inhibitsspontaneous fragmentation of actin fila-ments.J. Mol. Biol.161:217–27
Weigt C, Schoepper B, Wegner A. 1990.Tropomyosin-troponin complex stabilizesthe pointed ends of actin filaments againstpolymerization and depolymerization.FEBSLett.260:266–68
Weins A, Schwarz K, Faul C, Barisoni L,Linke WA, Mundel P. 2001. Differentiation-and stress-dependent nuclear cytoplasmicredistribution of myopodin, a novel actin-bundling protein.J. Cell Biol.155:393–404
Weiss A, Leinwand LA. 1996. The mammalianmyosin heavy chain gene family.Annu. Rev.Cell Dev. Biol.12:417–39
Weitzer G, Milner DJ, Kim JU, Bradley A,Capetanaki Y. 1995. Cytoskeletal control ofmyogenesis: a desmin null mutation blocksthe myogenic pathway during embryonicstem cell differentiation.Dev. Biol.172:422–39
Westfall MV, Metzger JM. 2001. Troponin Iisoforms and chimeras: tuning the molecularswitch of cardiac contraction.News Physiol.Sci.16:278–81
Whitby FG, Phillips GN Jr. 2000. Crystal struc-ture of tropomyosin at 7 Angstroms resolu-tion. Proteins38:49–59

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
STRIATED MUSCLE CYTOSKELETON 705
Whiting A, Wardale J, Trinick J. 1989. Doestitin regulate the length of muscle thick fila-ments?J. Mol. Biol.205:263–68
Wiche G, Krepler R, Artlieb U, Pytela R, DenkH. 1983. Occurrence and immunolocaliza-tion of plectin in tissues.J. Cell Biol.97:887–901
Williams MW, Bloch RJ. 1999. Extensive butcoordinated reorganization of the membraneskeleton in myofibers of dystrophic (mdx)mice.J. Cell Biol.144:1259–70
Williams MW, Resneck WG, Kaysser T, Ur-sitti JA, Birkenmeier CS, et al. 2001. Na,K-ATPase in skeletal muscle: two populationsof beta-spectrin control localization in thesarcolemma but not partitioning between thesarcolemma and the transverse tubules.J.Cell Sci.114:751–62
Winder SJ. 2001. The complexities of dystro-glycan.Trends Biochem. Sci.26:118–24
Winegrad S. 1999. Cardiac myosin binding pro-tein C.Circ. Res.84:1117–26
Witt CC, Gerull B, Davies MJ, Centner T,Linke WA, Thierfelder L. 2001. Hypercon-tractile properties of cardiac muscle fibers ina knock-in mouse model of cardiac myosin-binding protein-C.J. Biol. Chem.276:5353–59
Wood DS, Zeviani M, Prelle A, Bonilla E,Salviati G, et al. 1987. Is nebulin the defectivegene product in Duchenne muscular dystro-phy?N. Engl. J. Med.316:107–8
Wood SJ, Slater CR. 1998. beta-Spectrin iscolocalized with both voltage-gated sodiumchannels and ankyrinG at the adult rat neu-romuscular junctionJ. Cell Biol. 140:675–84
Wu RY, Gill GN. 1994. LIM domain recog-nition of a tyrosine-containing tight turn.J.Biol. Chem.269:25085–90
Xia H, Winokur ST, Kuo WL, Altherr MR,Bredt DS. 1997. Actinin-associated LIM pro-tein: identification of a domain interactionbetween PDZ and spectrin-like repeat mo-tifs. J. Cell Biol.139:507–15
Xu X, Meiler SE, Zhong TP, Mohideen M,Crossley DA, et al. 2002. Cardiomyopathy inzebrafish due to mutation in an alternatively
spliced exon of titin.Nat. Genet.30:205–9
Yamasaki R, Berri M, Wu Y, Trombitas K, Mc-Nabb M, et al. 2001. Titin-actin interaction inmouse myocardium: passive tension modu-lation and its regulation by calcium/S100A1.Biophys. J.81:2297–313
Yang Q, Osinska H, Klevitsky R, Robbins J.2001. Phenotypic deficits in mice expressinga myosin binding protein C lacking the titinand myosin binding domains.J. Mol. Cell.Cardiol. 33:1649–58
Young P, Ehler E, Gautel M. 2001. Obscurin, agiant sarcomeric Rho guanine nucleotide ex-change factor protein involved in sarcomereassembly.J. Cell Biol.154:123–36
Young P, Ferguson C, Banuelos S, Gautel M.1998. Molecular structure of the sarcomericZ-disk: two types of titin interactions leadto an asymmetrical sorting of alpha-actinin.EMBO J.17:1614–24
Young P, Gautel M. 2000. The interaction oftitin and alpha-actinin is controlled by aphospholipid-regulated intramolecular pseu-doligand mechanism.EMBO J. 19:6331–40
Zajdel RW, McLean MD, Lemanski SL,Muthuchamy M, Wieczorek DF, et al. 1998.Ectopic expression of tropomyosin promotesmyofibrillogenesis in mutant axolotl hearts.Dev. Dyn.213:412–20
Zakhary DR, Moravec CS, Stewart RW, BondM. 1999. Protein kinase A (PKA)-dependenttroponin-I phosphorylation and PKA regu-latory subunits are decreased in human di-lated cardiomyopathy.Circulation 99:505–10
Zeng H, Jackson DA, Oshima H, Simons SSJr. 1998. Cloning and characterization of anovel binding factor (GMEB-2) of the gluco-corticoid modulatory element.J. Biol. Chem.273:17756–62
Zervas CG, Gregory SL, Brown NH. 2001.Drosophilaintegrin-linked kinase is requiredat sites of integrin adhesion to link the cy-toskeleton to the plasma membrane.J. CellBiol. 152:1007–18
Zhang Y, Featherstone D, Davis W, Rushton E,

6 Sep 2002 14:46 AR AR170-CB18-22.tex AR170-CB18-22.SGM LaTeX2e(2002/01/18)P1: IBD
706 CLARK ET AL.
Broadie K. 2000.Drosophila D-titin is re-quired for myoblast fusion and skeletal mus-cle striation.J. Cell Sci.113:3103–15
Zhou D, Birkenmeier CS, Williams MW,Sharp JJ, Barker JE, Bloch RJ. 1997. Small,membrane-bound, alternatively splicedforms of ankyrin 1 associated with the sarco-plasmic reticulum of mammalian skeletalmuscle.J. Cell Biol.136:621–31
Zhou Q, Chu PH, Huang C, Cheng CF, MartoneME, et al. 2001. Ablation of Cypher, a PDZ-LIM domain Z-line protein, causes a severeform of congenital myopathy.J. Cell Biol.155:605–12
Zhou Q, Ruiz-Lozano P, Martone ME, ChenJ. 1999. Cypher, a striated muscle-restrictedPDZ and LIM domain-containing protein,
binds to alpha-actinin-2 and protein kinaseC. J. Biol. Chem.274:19807–13
Zimmerman L, Parr B, Lendahl U, Cunning-ham M, McKay R, et al. 1994. Indepen-dent regulatory elements in the nestin genedirect transgene expression to neural stemcells or muscle precursors.Neuron12:11–24
Zolk O, Caroni P, Bohm M. 2000. Decreased ex-pression of the cardiac LIM domain proteinMLP in chronic human heart failure.Circu-lation 101:2674–77
Zou Y, Evans S, Chen J, Kuo HC, Harvey RP,Chien KR. 1997. CARP, a cardiac ankyrinrepeat protein, is downstream in the Nkx2-5 homeobox gene pathway.Development124:793–804

21 Sep 2002 14:56 AR AR170-22-COLOR.tex AR170-22-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Figure 1 A schematic overview of cytoskeletal linkages in striated muscle (modifiedfrom Carlsson & Thornell, 2001). The sarcomeres contain four filament systems: actin-thin, myosin-thick, titin, and nebulin filaments. The borders of individual sarcomeresare the Z-lines, which are precisely aligned and laterally associated with intermediatefilament proteins (such as desmin) and other cytoskeletal proteins (such as plectin). Theintermediate filaments and associated proteins also may link the peripheral myofibrilsto costameres at the sarcolemma (the muscle membrane), to mitochondria, and to thenuclear membrane. Although many of the detailed interactions are not yet known, theselinkages are responsible for the mechanical integration and stability of myofibrils, or-ganelles, and membrane components for effective force transmission. The microtubulesystem is not depicted in the schematic because it is unclear how they are arranged instriated muscle; however, they may be linked to myofibrils and intermediate proteinsthrough proteins such as plakin family members.
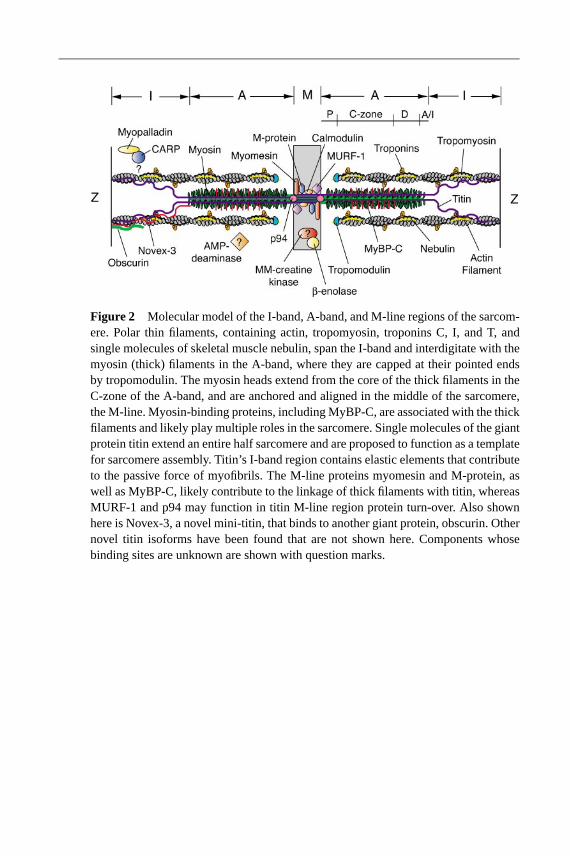
21 Sep 2002 14:56 AR AR170-22-COLOR.tex AR170-22-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Figure 2 Molecular model of the I-band, A-band, and M-line regions of the sarcom-ere. Polar thin filaments, containing actin, tropomyosin, troponins C, I, and T, andsingle molecules of skeletal muscle nebulin, span the I-band and interdigitate with themyosin (thick) filaments in the A-band, where they are capped at their pointed endsby tropomodulin. The myosin heads extend from the core of the thick filaments in theC-zone of the A-band, and are anchored and aligned in the middle of the sarcomere,the M-line. Myosin-binding proteins, including MyBP-C, are associated with the thickfilaments and likely play multiple roles in the sarcomere. Single molecules of the giantprotein titin extend an entire half sarcomere and are proposed to function as a templatefor sarcomere assembly. Titin’s I-band region contains elastic elements that contributeto the passive force of myofibrils. The M-line proteins myomesin and M-protein, aswell as MyBP-C, likely contribute to the linkage of thick filaments with titin, whereasMURF-1 and p94 may function in titin M-line region protein turn-over. Also shownhere is Novex-3, a novel mini-titin, that binds to another giant protein, obscurin. Othernovel titin isoforms have been found that are not shown here. Components whosebinding sites are unknown are shown with question marks.
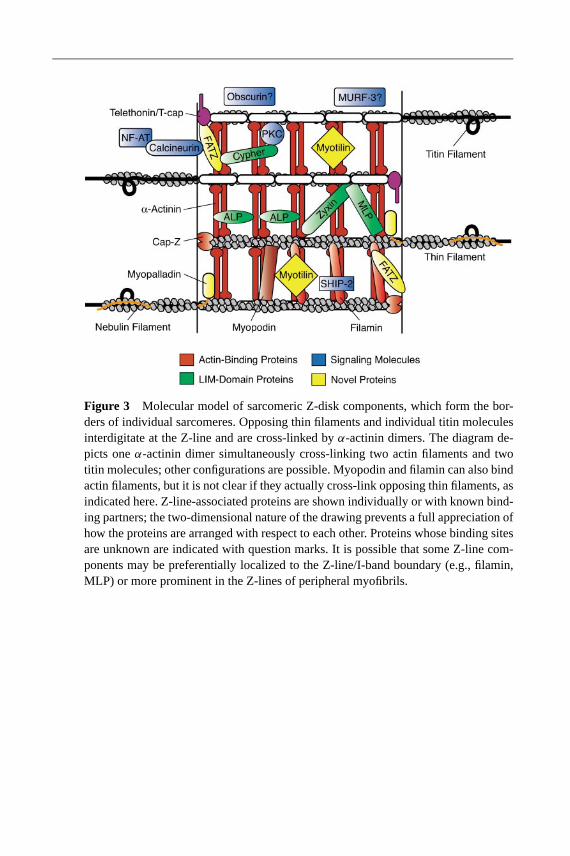
21 Sep 2002 14:56 AR AR170-22-COLOR.tex AR170-22-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Figure 3 Molecular model of sarcomeric Z-disk components, which form the bor-ders of individual sarcomeres. Opposing thin filaments and individual titin moleculesinterdigitate at the Z-line and are cross-linked byα-actinin dimers. The diagram de-picts oneα-actinin dimer simultaneously cross-linking two actin filaments and twotitin molecules; other configurations are possible. Myopodin and filamin can also bindactin filaments, but it is not clear if they actually cross-link opposing thin filaments, asindicated here. Z-line-associated proteins are shown individually or with known bind-ing partners; the two-dimensional nature of the drawing prevents a full appreciation ofhow the proteins are arranged with respect to each other. Proteins whose binding sitesare unknown are indicated with question marks. It is possible that some Z-line com-ponents may be preferentially localized to the Z-line/I-band boundary (e.g., filamin,MLP) or more prominent in the Z-lines of peripheral myofibrils.

21 Sep 2002 14:56 AR AR170-22-COLOR.tex AR170-22-COLOR.SGM LaTeX2e(2002/01/18)P1: GDL
Figure 4 A schematic model of the cytoskeletal filament linkages at the sarcolemma of stri-ated muscle. Four major cytoskeletal/membrane junctions are depicted: (a) cadherin-basedlinkages to actin and intermediate filaments (desmin); (b) integrin-based focal adhesions;(c) dystroglycan complex (DGC); and (d) spectrin-based membrane cytoskeleton. Thecadherin-based fascia adheren at the intercalated disc couples neighboring cardiomyocytes(through homotypic interactions) and tethers the contractile apparatus to the muscle termini.Desmosomes are a second cadherin-based junction that anchor desmin filaments at the in-tercalated disc. Connections between intermediate filament proteins and the membrane mayoccur through a plectin/αβ-crystallin complex or via an association with DGC via dystro-brevin. Integrin-based focal adhesions and the DGC act as transmembrane receptors for ECMcomponents (e.g., laminin) and link the extracellular surface with the actin cytoskeleton. Inte-grins associate with talin,α-actinin, vinculin and N-RAP to form a strong mechanical link toactin filaments. Integrins could directly interact withα-actinin and/or other components notdepicted here to mediate a connection with actin. The DGC consists of the transmembranecomplexα/β-dystroglycan, dystrophin, the sarcoglycans, and other components not depictedhere. Spectrin is enriched at costameres, and is an important component of the membranecytoskeleton. It is linked to the membrane through ankyrin and probably the Na,K-ATPasetransmembrane protein. Spectrin may have an additional role in anchoring the contractileapparatus to the membrane though an interaction with MLP. Importantly, all of these link-age complexes can bind to the submembraneous actin (γ -actin) and are probably interlinkedthrough this association as well as other unknown interactions.





























