Embed Size (px)
Citation preview

J. Comp. Path. 2001, Vol. 125, 292–303doi:10.1053/jcpa.2001.0508, available online at http://www.idealibrary.com on
An Experimental Model for Post-weaningMultisystemic Wasting Syndrome (PMWS) in
Growing Piglets
E. Albina, C. Truong, E. Hutet, P. Blanchard, R. Cariolet,R. L’Hospitalier, D. Mahe, C. Allee, H. Morvan∗, N. Amenna∗,
M. Le Dimna, F. Madec and A. Jestin
Agence Francaise de Securite Sanitaire des Aliments Ploufragan, Laboratoire d’Etudes et de Recherches Avicoles et
Porcines, BP 53, 22440 Ploufragan and ∗Laboratoire de Developpement et d’Analyses 22, BP 54, 22440, Ploufragan,
France
Summary
Post-weaning multisystemic wasting syndrome (PMWS) is a comparatively new disease of swine, andknown to occur in France since 1996. A porcine circovirus type 2 (PCV2) is found in the lesions of affectedpiglets. Six piglets aged 10–13 weeks were obtained from a French PMWS-affected farm. Two showingcharacteristic signs of PMWS (palor, weakness and emaciation) remained in poor condition and werefinally killed 6 and 9 days after their arrival in the experimental unit. Tissue homogenates from these twopiglets were used to reproduce mild PMWS in specific pathogen-free (SPF) piglets. This mild PMWSconsisted of pyrexia (up to 41·7°C) and growth retardation (up to 30% of weight reduction comparedwith controls) commencing 1 week after infection and lasting 3 weeks. In seven additional trials, pyrexia,growth retardation and lesions characteristic of PMWS were consistently produced in SPF and conventionalpiglets. However, only four of 55 inoculated SPF piglets (7·2%) showed severe wasting disease. One diedand the others had to be killed 3 to 4 weeks after inoculation. None of the inoculated animals developedantibodies to any common swine viruses or bacteria, but clear evidence of PCV2 seroconversion wasobtained. Our results therefore strongly suggest that PCV2 is the primary aetiological agent of PMWS.
2001 Harcourt Publishers Ltd
Introduction (Le Cann et al., 1997) and elsewhere (Nayar et al.,1997; Segales et al., 1997; Allan et al., 1998a; EllisIn France, post-weaning multisystemic wasting syn-et al., 1998). In vitro, this circovirus replicates to adrome (PMWS) of piglets was first reported in limited degree only, a fact that may account for1996, after which it spread relatively slowly. The difficulty in reproducing the disease by inoculation.
disease is characterized by an acute wasting syn- To our knowledge, the number of successful trans-drome, mainly affecting piglets aged 8 to 13 weeks mission experiments is limited to four, in which(Madec et al., 2000), the animals suddenly becoming the inoculum consisted of either material from fieldpale, lethargic, feverish and emaciated. Morbidity cases (Balasch et al., 1999; Ellis et al., 1999) or virusranges from 5 to 35% and mortality is often close grown in vitro (Allan et al., 1999, 2000; Ellis et al.,to 100%. The causal agent has not been identified 1999; Kennedy et al., 2000; Krakowka et al., 2000).with certainty, but a circovirus (PCV2) has regularly These experiments, however, were made in smallbeen found in association with the lesions in France numbers of animals, usually with inocula con-
taining PCV2 and another swine virus; moreover,there was no indication of reproducibility.Correspondence to: A. Jestin.
0021–9975/01/040292+12 $35.00 2001 Harcourt Publishers Ltd

Experimental Model for PMWS 293
This report describes the development of a re- the two piglets were homogenized separately. Aftercentrifugation of the two homogenates, the super-producible experimental model of PMWS.nates were collected and each was used for theintratracheal inoculation of two SPF piglets.
Materials and Methods After trial 1, all piglets were inoculated withtissue preparations as indicated in Table 1. In trialAnimals2, four SPF piglets were inoculated intratracheally
Six piglets (referred to as “farm piglets”) aged with 6 ml of lung preparation on day 0, four SPF10–13 weeks, from a conventional farm affected piglets being sham-inoculated with 6 ml of cellby PMWS, were brought to the experimental unit culture medium. In trials 3 and 4, the piglets allfor clinical monitoring. An attempt was first made received identical inocula derived from trial 1. Into transmit the disease from the farm piglets to five trials 5–9, the inoculum consisted of a filteredspecific pathogen-free (SPF) piglets aged 10 weeks (0·45 �m pore diameter) tissue preparation derivedby direct contact exposure. from trial 4. The presence of virus in the inocula
These SPF piglets and the SPF herd from which used for trials 3 and 4 was investigated directlythey were derived were known to be free from by immuno-electron microscopy with the PCV2African and classical swine fever, bovine viral diar- antiserum described in the next section and proteinrhoea, Border disease, Aujeszky’s disease, parvo- A-coated colloidal gold-labelled particles (10 nm;virus, porcine reproductive and respiratory Sigma, St Quentin Fallavier, France). The titre ofsyndrome virus (American and European geno- porcine circovirus in inocula for trials 5 to 9 wastypes), swine influenza, porcine circovirus type 1 104·5 TCID50/ml; the piglets were infected intra-and 2 (PCV1 and PCV2), transmissible gastro- tracheally with 105·2 TCID50 in a dose volume ofenteritis, porcine respiratory coronavirus, Myco- 5 ml and intramuscularly with 104·5 TCID50 inplasma hyopneumoniae, Pasteurella multocida, Bordetella 1 ml. It was demonstrated by specific polymerasebronchiseptica, Actinobacillus pleuropneumonia, Haemo- chain reaction (PCR) that only PCV type 2 (nophilus parasuis, Streptococcus suis, Treponema hyo- PCV type 1) was present in these inocula (data notdysenteriae (Cariolet et al., 1994). shown). All attempts to isolate bacterial pathogens
After the initial transmission experiment by from the tissues of the affected piglets were un-direct contact, seven groups of four to eight SPF successful.piglets aged 5–9 weeks and one group of eightconventional piglets aged 6–7 weeks were in-
Laboratory Investigationsoculated with different tissue preparations fromsick animals as described later. Control sham-in- Viral isolation was attempted on lymph nodes andoculated piglets were included in the different ex- lung tissue samples from the piglets of trials 1 toperimental trials. Each group of piglets was isolated 5. The cell cultures used included porcine alveolarin housing with air filtration and low air pressure. macrophages, peripheral blood mononuclear cellsClinical observations and rectal temperatures were (PBMCs), “immortalized” porcine lymphocytesrecorded daily and the piglets were bled and (L14 cell line; Kaeffer et al., 1990), primary porcineweighed weekly. Unless killed earlier for humane kidney cells, pig kidney cell line (PK) and monkeyreasons, the piglets were monitored for 35 days. At kidney cell line (MARC145; Kim et al., 1993).necropsy, any macroscopical lesions were examined These cell cultures were checked daily for anyand, when present, tissue samples were collected cytopathogenic effect. When this was absent afterfor laboratory investigation (see below). 7 days of culture, non-cytopathogenic replicating
virus was sought by an immunoperoxidase mono-layer assay (IPMA; Wensvoort et al., 1991), with
Inocula PCV2 antiserum consisting of serum from a con-valescent piglet. Briefly, monolayers of PK cellsFor the first experiment (“trial 1”), inocula were
prepared from lung tissues of two farm piglets were incubated with the inoculum for 1 h at 37°Cand then rinsed and treated with glucosamineshowing typical signs of PMWS. One of these two
piglets had grey-tan pulmonary lesions, but bacteria (300 mM) for 1 h at 37°C. After incubation for 36 hat 37°C, the cells were fixed with cold acetonewere not isolated from the lungs or from those of
their pen-mates. One of these two animals showed 80% for 45 min at −20°C. A convalescent pigletantiserum to PCV 2 was then added and left fora characteristic lesion of PMWS in a lymph node,
consisting of mononuclear cell depletion and multi- 45 min at 37°C. A secondary peroxidase-con-jugated rabbit anti-swine immunoglobulin was usednucleate giant cell infiltration. Lung tissues from

E. Albina et al.294
Tab
le1
Des
crip
tio
no
fth
ed
iffer
ent
tria
ls
Info
rmat
ion
Det
ails
oftr
ial
no.
12
34
56
78
9
Hea
lthst
atus
ofpi
glet
sC
onve
ntio
nal
SPF
SPF
SPF
SPF
SPF
SPF
SPF
Con
vent
iona
lN
umbe
rof
pigl
ets
64∗
6∗6∗
8∗7∗
8∗8∗
8∗N
umbe
rof
cont
act-
expo
sed
54
43
00
00
0pi
glet
sW
eeks
ofag
eat
chal
leng
eN
och
alle
nge
96
55
77
66–
7In
ocul
um⊥
Lun
gti
ssue
Lym
phno
des
Lym
phno
des
Lym
phno
des
Lym
phno
des
Lym
phno
des
Lym
phno
des
Lym
phno
des
from
tria
l1
from
tria
l2
from
tria
l2
from
tria
l4
from
tria
l4
from
tria
l4
from
tria
l4
from
tria
l4
Filtr
atio
nof
inoc
ulum
⊥no
yes
yes
yes
yes
yes
yes
yes
(0·4
5�
m)
Vol
ume
ofin
ocul
um⊥
6m
l6
ml
5m
l5
ml
5m
l5
ml
5m
l5
ml
Rou
teof
infe
ctio
n⊥
intr
atra
chea
lin
trat
rach
eal
intr
atra
chea
lin
trat
rach
eal
intr
atra
chea
lin
trat
rach
eal
intr
atra
chea
lin
trat
rach
eal
++
++
++
1m
l1
ml
1m
l1
ml
1m
l1
ml
intr
amus
cula
rin
tram
uscu
lar
intr
amus
cula
rin
tram
uscu
lar
intr
amus
cula
rin
tram
uscu
lar
PC
V2
titr
ein
inoc
ulum
⊥N
DN
DN
D10
4·5
104·
510
4·5
104·
510
4·5
(TC
ID50
/ml)
∗E
qual
num
ber
ofun
infe
cted
cont
rol
pigl
ets
wer
eus
ed.
⊥,
No
entr
y.N
D,
not
dete
rmin
ed.

Experimental Model for PMWS 295
to develop the coloration in the presence of 3- from trials 1, 2, 3 and 9. In addition, nasal swabscollected from piglets in trial 6 were examinedamino-9-ethyl-carbazole. The antiserum used in
this reaction was collected from one contact SPF for the presence of bacteria. The detection ofantibodies to porcine circovirus was carried outpiglet in trial 2 (see Table 1), 4 weeks after the first
clinical signs. This serum, which was proved to with the IPMA test, already described for thedetection of non-cytopathogenic virus in cell cul-contain antibodies to PCV2 by immunoblotting
against recombinant PCV2 ORF2 protein (data ture (see above). The virus used in this test wasisolated from a sick piglet and passaged on PKnot shown), was used for IPMA and immuno-
electron microscopy. cells previously checked for freedom from con-tamination with PCV1. The virus used was iden-PCV2 titration was performed according to the
method of Kaerber (1931), titres being expressed tified as PCV2 by molecular sequencing(Mankertz et al., 2000). A peptide-based ELISAas TCID50/ml. To improve PCV2 replication in
PK cell cultures, treatment with glucosamine test for PCV2 antibodies was later used for theconfirmation of specific PCV2 seroconversion(300 mM) was applied as described by Tischer et
al. (1995). A PCR specific for PCV type 1 or type (Truong et al., 2001).2 was developed and used for the detection ofcircoviruses in lymph nodes of piglets from trial 4(Truong et al., 2001). Statistical Analysis
Histological examination was carried out on lungThe daily rectal temperatures in trial 2, the relativeand lymph node tissues after fixation with 10%daily weight gains and the absorbances in ELISAformaldehyde. Lung tissues were examined fromwere analysed by the non-parametric U-test ofthe piglets of trials 1 to 3 and lymph node tissuesMann and Whitney. The numbers of days whenfrom the piglets of trials 1, 2, 3, 5 and 9. Tissuethe rectal temperature was increased were analysedsections (8 �m) were stained with haemalum-eosin-by the Yates chi2 test.safranin and observed under a light microscope.
In-situ hybridization was performed essentially asdescribed by McNeilly et al. (1999). Briefly, 4-�msections of lymph nodes of piglets from trials 5 and Results9 were washed, heated to 40°C, deproteinized and
Case Study (Trial 1)then hybridized at 55°C with the PCV2 specificprobe 5′ CCGTTGTCCCTGAGATCTA 3′ con- All farm piglets remained clinically normal during
the experiment, except for two that already showedjugated at both ends with digoxigenin. After threewashes and one incubation at 40°C, a biotinylated PMWS signs before their introduction into the
experimental unit. In contrast, five SPF pigletsmouse monoclonal antibody antidigoxigenin (Ven-tana Medical, Strasbourg), a peroxidase-avidin placed in direct contact with these farm piglets
showed various disease signs including diarrhoeacomplex and 3-amino-9-ethyl-carbazole were ap-plied successively. The slides were finally coun- and pyrexia. All farm and SPF piglets were tested
for antibodies to M. hyopneumoniae, A. pleuro-terstained with haemalum and blueing reagent(Ventana). pneumoniae, CSFV, RPVs, and ADV at the end of
the experiment, with negative results. However,Serological tests were performed with eithercommercial or “in-house” diagnostic kits. Anti- the sera from four of the six farm piglets contained
either PRRSV or PRRSV antibodies. PRRSVbodies to classical swine fever virus (CSFV), ru-minant pestiviruses (RPVs), porcine reproductive could be isolated from the sera of SPF contact
piglets and a clear seroconversion in these pigletsand respiratory syndrome virus (PRRSV), Au-jeszky’s disease virus (ADV), porcine parvovirus against this virus was observed a few days later.
The PRRSV strain isolated was of the European(PPV), encephalo-myocarditis virus (EMCV),swine influenza H1N1 and H3N2, transmissible genotype and its pathogenicity was checked ex-
perimentally in vivo. It did not differ substantiallygastro-enteritis virus (TGEV), A. pleuropneumoniae,M. hyopneumoniae and Eperythrozoon suis were from other strains isolated during the epidemic
phase in France in 1992 (results not shown). In-checked in the sera of piglets from trials 1, 2, 4,5 and 6 as indicated in the section results. All terestingly, the two farm piglets with PMWS signs
remained negative for PRRSV and PRRSV anti-sera were tested before inoculation and at the endof the experiment, i.e. 5 weeks post-infection. bodies; both piglets were emaciated, weak, and
exhibited a weight loss of 200 to 400 g/day duringBacteriological examination was performed onlung tissues showing lesions at necropsy in piglets the 6 to 8 days spent in our institute. Both were
E. Albina et al.296
anaesthetized and killed for humane reasons. Grey-tan macroscopical lesions were observed at nec-ropsy in the lungs of one piglet. Analysis of the cellsub-populations in the lung washings from thispiglet showed a clear diminution of macrophages(28% of the whole population) and a markedlymphocyte infiltration (69% of the whole cell popu-lation). These lymphocytes were typed as CD8+cells (data not shown), suggesting the presence ofa viral infection. In addition, histological ex-amination of lymph nodes from this piglet showedlymphocyte infiltration and multinucleate giantcells. PCV (shown later by PCR to be of type 2)was isolated on PK cell culture from lung samplesof this piglet.
Reproduction of the Disease (Trial 2)
This latter observation led us to prepare an in-oculum from each of the two sick (PRRS-negative)farm piglets, each inoculum being administeredintratracheally to two SPF animals, which wereimmediately placed in contact with a further fourSPF piglets. In this trial, four control piglets re-
56
40
Weeks post-infection
(b)
DW
G (
g/kg
of
body
wei
ght)
–1
35
30
25
20
15
10
543210
38.535
41.5
Days post-infection
(a)
Rec
tal t
empe
ratu
res
(°C
)
0
41.0
40.5
40.0
39.5
39.0
30252015105
* *
*
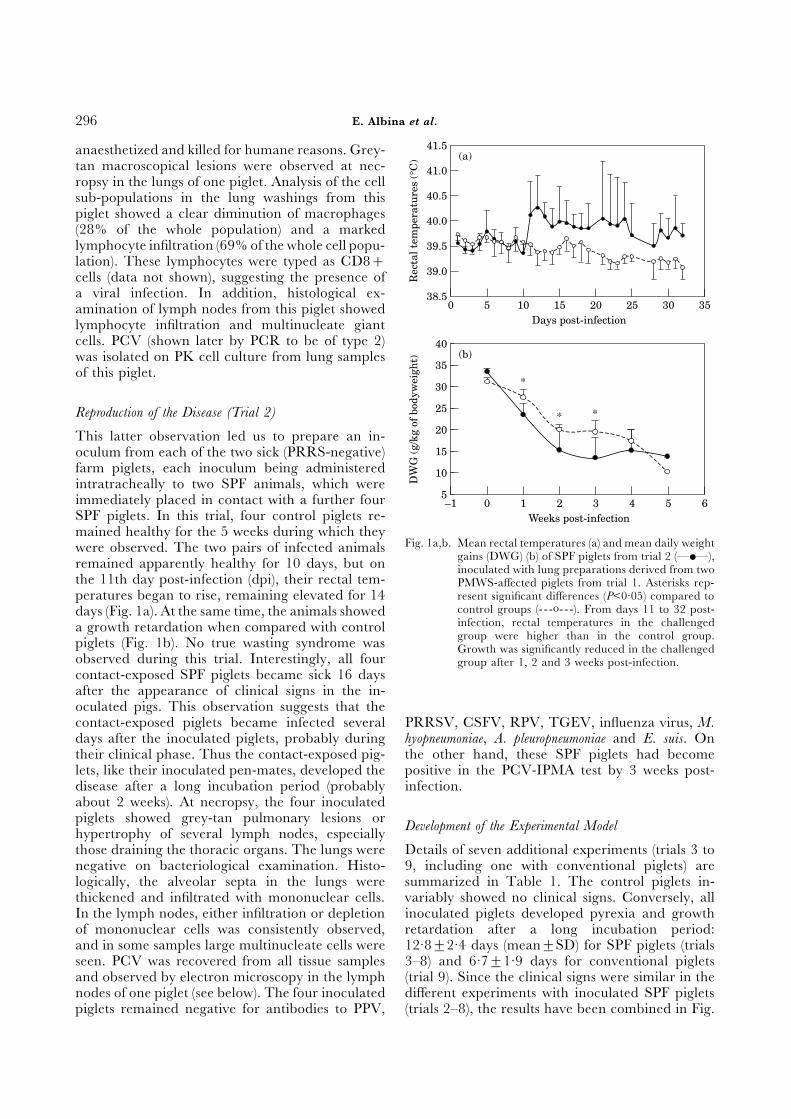
mained healthy for the 5 weeks during which theyFig. 1a,b. Mean rectal temperatures (a) and mean daily weightwere observed. The two pairs of infected animals
gains (DWG) (b) of SPF piglets from trial 2 (—•—),remained apparently healthy for 10 days, but on inoculated with lung preparations derived from twothe 11th day post-infection (dpi), their rectal tem- PMWS-affected piglets from trial 1. Asterisks rep-peratures began to rise, remaining elevated for 14 resent significant differences (P<0·05) compared to
control groups (- - -o- - -). From days 11 to 32 post-days (Fig. 1a). At the same time, the animals showedinfection, rectal temperatures in the challengeda growth retardation when compared with controlgroup were higher than in the control group.piglets (Fig. 1b). No true wasting syndrome was Growth was significantly reduced in the challenged
observed during this trial. Interestingly, all four group after 1, 2 and 3 weeks post-infection.contact-exposed SPF piglets became sick 16 daysafter the appearance of clinical signs in the in-oculated pigs. This observation suggests that thecontact-exposed piglets became infected several PRRSV, CSFV, RPV, TGEV, influenza virus, M.
hyopneumoniae, A. pleuropneumoniae and E. suis. Ondays after the inoculated piglets, probably duringtheir clinical phase. Thus the contact-exposed pig- the other hand, these SPF piglets had become
positive in the PCV-IPMA test by 3 weeks post-lets, like their inoculated pen-mates, developed thedisease after a long incubation period (probably infection.about 2 weeks). At necropsy, the four inoculatedpiglets showed grey-tan pulmonary lesions or
Development of the Experimental Modelhypertrophy of several lymph nodes, especiallythose draining the thoracic organs. The lungs were Details of seven additional experiments (trials 3 to
9, including one with conventional piglets) arenegative on bacteriological examination. Histo-logically, the alveolar septa in the lungs were summarized in Table 1. The control piglets in-
variably showed no clinical signs. Conversely, allthickened and infiltrated with mononuclear cells.In the lymph nodes, either infiltration or depletion inoculated piglets developed pyrexia and growth
retardation after a long incubation period:of mononuclear cells was consistently observed,and in some samples large multinucleate cells were 12·8±2·4 days (mean±SD) for SPF piglets (trials
3–8) and 6·7±1·9 days for conventional pigletsseen. PCV was recovered from all tissue samplesand observed by electron microscopy in the lymph (trial 9). Since the clinical signs were similar in the
different experiments with inoculated SPF pigletsnodes of one piglet (see below). The four inoculatedpiglets remained negative for antibodies to PPV, (trials 2–8), the results have been combined in Fig.
Experimental Model for PMWS 297
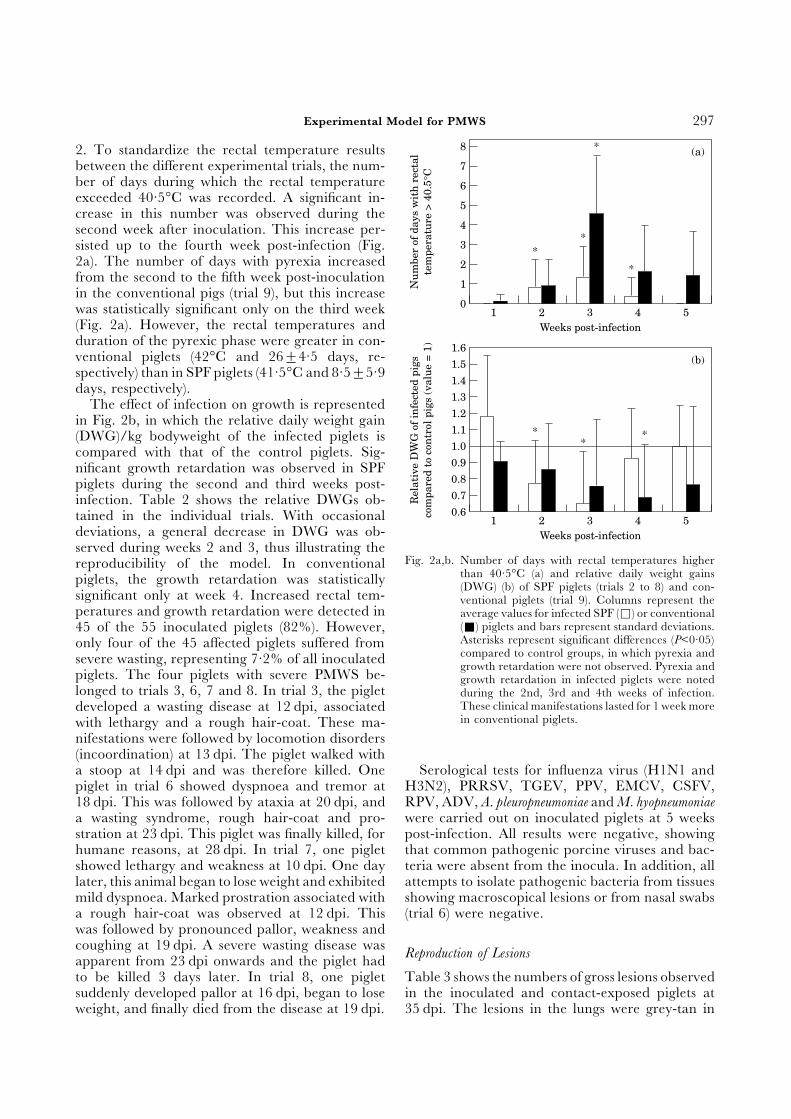
2. To standardize the rectal temperature resultsbetween the different experimental trials, the num-ber of days during which the rectal temperatureexceeded 40·5°C was recorded. A significant in-crease in this number was observed during thesecond week after inoculation. This increase per-sisted up to the fourth week post-infection (Fig.2a). The number of days with pyrexia increasedfrom the second to the fifth week post-inoculationin the conventional pigs (trial 9), but this increasewas statistically significant only on the third week(Fig. 2a). However, the rectal temperatures andduration of the pyrexic phase were greater in con-ventional piglets (42°C and 26±4·5 days, re-spectively) than in SPF piglets (41·5°C and 8·5±5·9days, respectively).
The effect of infection on growth is representedin Fig. 2b, in which the relative daily weight gain(DWG)/kg bodyweight of the infected piglets iscompared with that of the control piglets. Sig-nificant growth retardation was observed in SPFpiglets during the second and third weeks post-infection. Table 2 shows the relative DWGs ob-tained in the individual trials. With occasionaldeviations, a general decrease in DWG was ob-
0.6
1.6
Weeks post-infection
(b)
Rel
ativ
e D
WG
of
infe
cted
pig
sco
mpa
red
to c
ontr
ol p
igs
(val
ue
= 1)
1.5
1.4
1.3
1.2
1.1
1.0
0.9
0.8
0.7
1 2
*
3
*
4
*
5
0
8
Weeks post-infection
(a)
Nu
mbe
r of
day
s w
ith
rec
tal
tem
pera
ture
> 4
0.5°
C 7
6
5
4
3
2
1
1 2
*
3
*
*
4
*
5
served during weeks 2 and 3, thus illustrating theFig. 2a,b. Number of days with rectal temperatures higherreproducibility of the model. In conventional
than 40·5°C (a) and relative daily weight gainspiglets, the growth retardation was statistically (DWG) (b) of SPF piglets (trials 2 to 8) and con-significant only at week 4. Increased rectal tem- ventional piglets (trial 9). Columns represent theperatures and growth retardation were detected in average values for infected SPF (Φ) or conventional
(Ε) piglets and bars represent standard deviations.45 of the 55 inoculated piglets (82%). However,Asterisks represent significant differences (P<0·05)only four of the 45 affected piglets suffered fromcompared to control groups, in which pyrexia andsevere wasting, representing 7·2% of all inoculated growth retardation were not observed. Pyrexia and
piglets. The four piglets with severe PMWS be- growth retardation in infected piglets were notedlonged to trials 3, 6, 7 and 8. In trial 3, the piglet during the 2nd, 3rd and 4th weeks of infection.
These clinical manifestations lasted for 1 week moredeveloped a wasting disease at 12 dpi, associatedin conventional piglets.with lethargy and a rough hair-coat. These ma-
nifestations were followed by locomotion disorders(incoordination) at 13 dpi. The piglet walked witha stoop at 14 dpi and was therefore killed. One Serological tests for influenza virus (H1N1 and
H3N2), PRRSV, TGEV, PPV, EMCV, CSFV,piglet in trial 6 showed dyspnoea and tremor at18 dpi. This was followed by ataxia at 20 dpi, and RPV, ADV, A. pleuropneumoniae and M. hyopneumoniae
were carried out on inoculated piglets at 5 weeksa wasting syndrome, rough hair-coat and pro-stration at 23 dpi. This piglet was finally killed, for post-infection. All results were negative, showing
that common pathogenic porcine viruses and bac-humane reasons, at 28 dpi. In trial 7, one pigletshowed lethargy and weakness at 10 dpi. One day teria were absent from the inocula. In addition, all
attempts to isolate pathogenic bacteria from tissueslater, this animal began to lose weight and exhibitedmild dyspnoea. Marked prostration associated with showing macroscopical lesions or from nasal swabs
(trial 6) were negative.a rough hair-coat was observed at 12 dpi. Thiswas followed by pronounced pallor, weakness andcoughing at 19 dpi. A severe wasting disease was
Reproduction of Lesionsapparent from 23 dpi onwards and the piglet hadto be killed 3 days later. In trial 8, one piglet Table 3 shows the numbers of gross lesions observed
in the inoculated and contact-exposed piglets atsuddenly developed pallor at 16 dpi, began to loseweight, and finally died from the disease at 19 dpi. 35 dpi. The lesions in the lungs were grey-tan in
E. Albina et al.298
Table 2Relative mean daily weight gains (DWGs) of infected SPF piglets (trials 2 to 8) and infected conventional piglets
(trial 9) compared with the corresponding values in control piglets
Weeks post- DWG values∗ in trial no.infection
2 3 4 5 6 7 8 Mean values 9(n=4)† (n=6)† (n=6)† (n=8)† (n=7)† (n=8)† (n=8)† for SPF piglets (n=8)†
of trials 2–8(n=47)†
1 0·85‡ 1·28 0·92 1·05 0·98 1·13 2·14‡§ 1·19 0·912 0·76‡ 0·84 0·78‡ 0·92 0·59‡ 0·73‡ 0·88‡ 0·78‡ 0·653 0·69‡ 1·03 0·67‡ 0·68‡ 0·39‡ 0·70 0·59‡ 0·68‡ 0·754 0·86 0·99 1·11 0·98 1·2 ‡ 0·79‡ 0·93‡ 0·98 0·69‡5 1·34 0·98 0·87 1·16‡ 0·88‡ 0·95 0·88 1·01 0·76
∗Results represent ratios of the DWG value (g/kg bodyweight) of infected piglets to those of control piglets.† Number of piglets.‡ Significant differences (P<0·05) between infected and control group.§ This difference was ascribed to reduced feed intake (reason unknown) in controls, which remained otherwise clinically healthy.
colour and consisted of non-purulent pneumonia, weeks after the occurrence of disease in the in-oculated pen-mates. In trial 3, three of four contact-essentially limited to the cranial lobes. Histo-
logically, the alveolar septa were infiltrated with exposed piglets became sick, while in the fourthtrial, all three were affected. In all, 90% of themononuclear cells. In lymph nodes, gross lesions
consisted of hyperplasia and discoloration. Thor- contact-exposed piglets in trials 2–4 developed dis-ease manifestations resembling those of the in-acic lymph nodes were often enlarged, at least one
piglet in each of trials 1–5 and 7 being affected. oculated piglets.Other lymph nodes (e.g. pharyngeal, testicular,inguinal and mesenteric) were occasionally en-
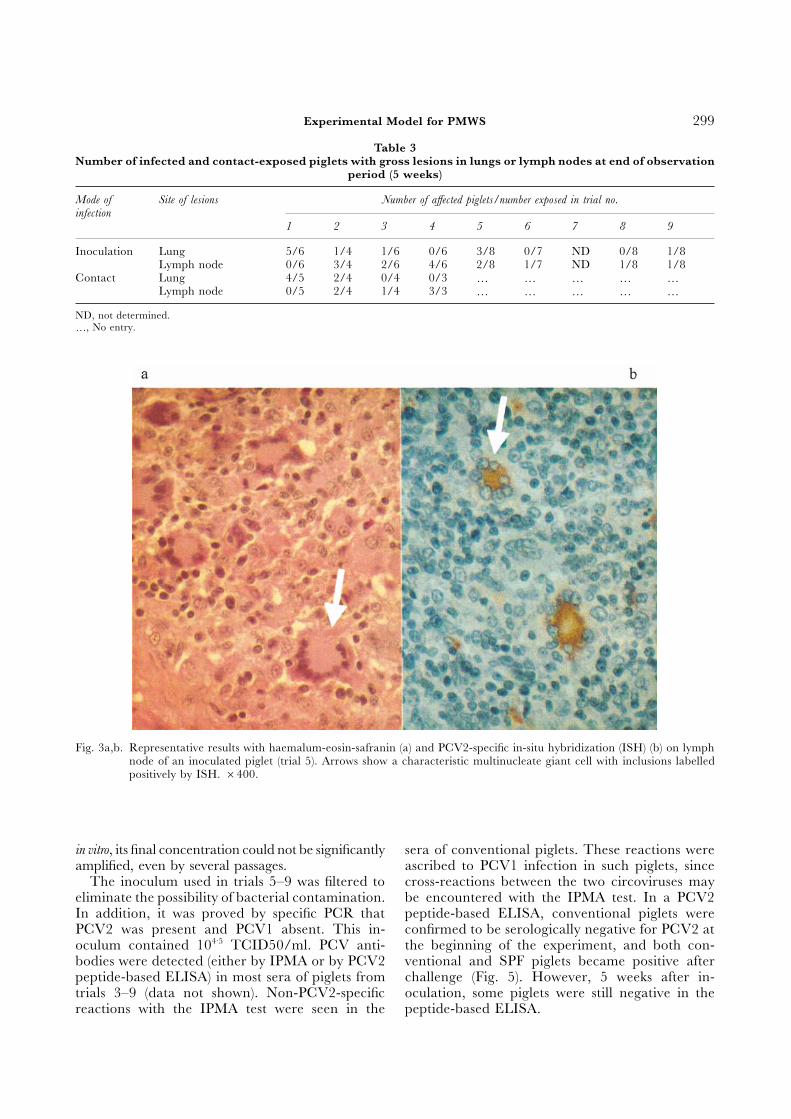
Laboratory Investigations on the Aetiological Agentlarged. The lesions were characterized histo-logically by infiltration of mononuclear cells and Small viral particles resembling porcine circovirusmultinucleate giant cells which stained positively in were observed by immuno-electron microscopy inPCV2-specific in-situ hybridization (Fig. 3). These the lymph node homogenate of one piglet from trialmultinucleate giant cells were seen in the lymph 2 (Fig. 4). In trials 2, 3 and 5, viral isolation wasnodes of piglets from trials 1 (three animals), 2 attempted only from tissues of sick piglets showing(two), 3 (one), 5 (one) and 9 (one). In trial 2, the lesions: a non-cytopathogenic virus was isolated frommultinucleate giant cells were seen in two contact- one inoculated piglet in each trial. Similarly, a non-exposed piglets. In piglets from trials 2, 3, 5 and cytopathogenic virus was recovered from the lung of9, some lymph nodes also showed a severe depletion one contact-exposed piglet in trial 2. Viral isolationof lymphocytes, suggesting a late stage in the patho- was consistently obtained in macrophage/monocytegenic process induced by PCV infection. Three cultures derived from lung washings, or from isolatedpiglets that developed severe wasting syndrome had PBMCs, in primary kidney cells or in the PK celllymph node (thoracic or mesenteric) enlargement line. In trial 4, PCV was isolated on PK cells fromand one had pneumonia. In these piglets, histo- a pool of lymph nodes (mesenteric, gastric, thoraciclogical lesions were characterized by infiltration of and tracheal) from six inoculated piglets; the isolatemononuclear cells. The piglet from trial 8 that died was confirmed to be of type 2 by type-specific PCR.from the disease was pale, with haemorrhages in Virus was also isolated from the tissue homogenatethe stomach and severe congestion of the intestine; that had served to inoculate the piglets in trials 5 toother organs were normal. 9. In addition, the virus was titrated on the PK cell
line by IPMA. Viral replication did not lead to theformation of true plaques. Consequently, only isolated
Transmission of the Disease infected cells were observed, and the virus did notseem to spread easily from one cell to another. ThisTransmission of the disease was investigated by
contact experiments. In the second trial, all four suggested that the capacity of the virus to leave thecells was poor; therefore, although the virus replicatedcontact-exposed SPF piglets became affected 2
Experimental Model for PMWS 299
Table 3Number of infected and contact-exposed piglets with gross lesions in lungs or lymph nodes at end of observation
period (5 weeks)
Mode of Site of lesions Number of affected piglets/number exposed in trial no.infection
1 2 3 4 5 6 7 8 9
Inoculation Lung 5/6 1/4 1/6 0/6 3/8 0/7 ND 0/8 1/8Lymph node 0/6 3/4 2/6 4/6 2/8 1/7 ND 1/8 1/8
Contact Lung 4/5 2/4 0/4 0/3 ⊥ ⊥ ⊥ ⊥ ⊥Lymph node 0/5 2/4 1/4 3/3 ⊥ ⊥ ⊥ ⊥ ⊥
ND, not determined.⊥, No entry.
Fig. 3a,b. Representative results with haemalum-eosin-safranin (a) and PCV2-specific in-situ hybridization (ISH) (b) on lymphnode of an inoculated piglet (trial 5). Arrows show a characteristic multinucleate giant cell with inclusions labelledpositively by ISH. ×400.
in vitro, its final concentration could not be significantly sera of conventional piglets. These reactions wereascribed to PCV1 infection in such piglets, sinceamplified, even by several passages.
The inoculum used in trials 5–9 was filtered to cross-reactions between the two circoviruses maybe encountered with the IPMA test. In a PCV2eliminate the possibility of bacterial contamination.
In addition, it was proved by specific PCR that peptide-based ELISA, conventional piglets wereconfirmed to be serologically negative for PCV2 atPCV2 was present and PCV1 absent. This in-
oculum contained 104·5 TCID50/ml. PCV anti- the beginning of the experiment, and both con-ventional and SPF piglets became positive afterbodies were detected (either by IPMA or by PCV2
peptide-based ELISA) in most sera of piglets from challenge (Fig. 5). However, 5 weeks after in-oculation, some piglets were still negative in thetrials 3–9 (data not shown). Non-PCV2-specific
reactions with the IPMA test were seen in the peptide-based ELISA.
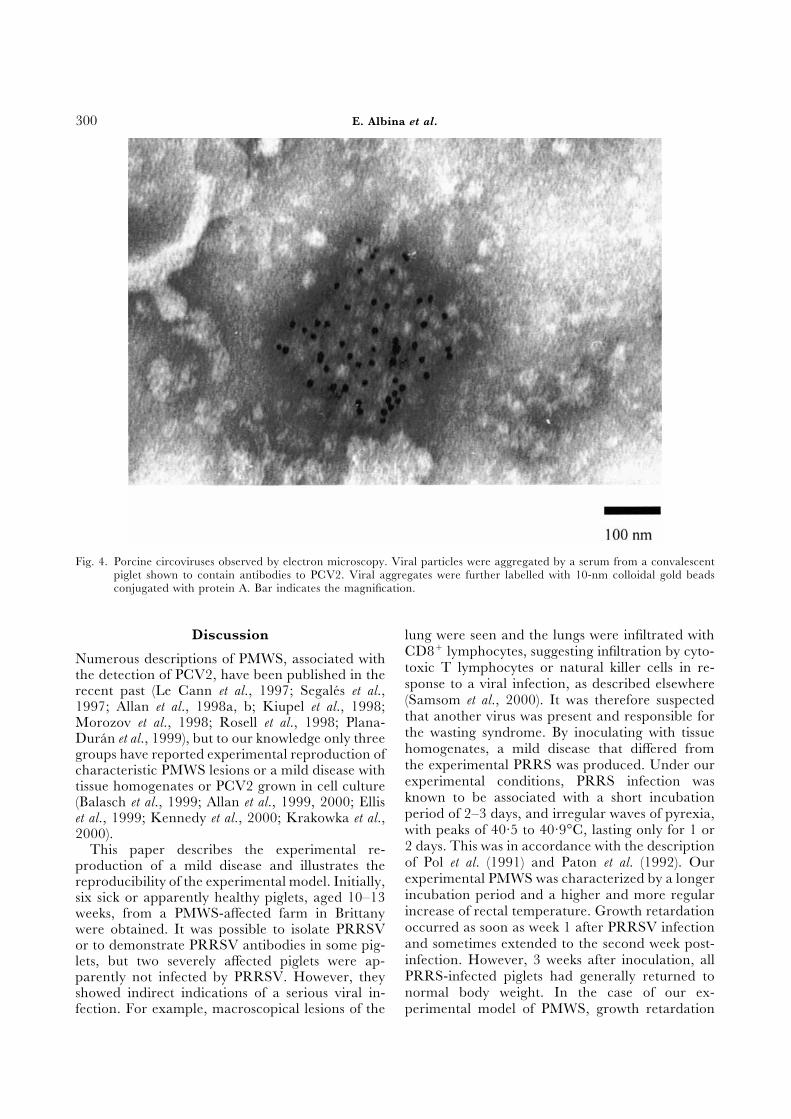
E. Albina et al.300
Fig. 4. Porcine circoviruses observed by electron microscopy. Viral particles were aggregated by a serum from a convalescentpiglet shown to contain antibodies to PCV2. Viral aggregates were further labelled with 10-nm colloidal gold beadsconjugated with protein A. Bar indicates the magnification.
Discussion lung were seen and the lungs were infiltrated withCD8+ lymphocytes, suggesting infiltration by cyto-Numerous descriptions of PMWS, associated withtoxic T lymphocytes or natural killer cells in re-the detection of PCV2, have been published in thesponse to a viral infection, as described elsewhererecent past (Le Cann et al., 1997; Segales et al.,(Samsom et al., 2000). It was therefore suspected1997; Allan et al., 1998a, b; Kiupel et al., 1998;that another virus was present and responsible forMorozov et al., 1998; Rosell et al., 1998; Plana-the wasting syndrome. By inoculating with tissueDuran et al., 1999), but to our knowledge only threehomogenates, a mild disease that differed fromgroups have reported experimental reproduction ofthe experimental PRRS was produced. Under ourcharacteristic PMWS lesions or a mild disease withexperimental conditions, PRRS infection wastissue homogenates or PCV2 grown in cell cultureknown to be associated with a short incubation(Balasch et al., 1999; Allan et al., 1999, 2000; Ellisperiod of 2–3 days, and irregular waves of pyrexia,et al., 1999; Kennedy et al., 2000; Krakowka et al.,with peaks of 40·5 to 40·9°C, lasting only for 1 or2000).2 days. This was in accordance with the descriptionThis paper describes the experimental re-of Pol et al. (1991) and Paton et al. (1992). Ourproduction of a mild disease and illustrates theexperimental PMWS was characterized by a longerreproducibility of the experimental model. Initially,incubation period and a higher and more regularsix sick or apparently healthy piglets, aged 10–13increase of rectal temperature. Growth retardationweeks, from a PMWS-affected farm in Brittanyoccurred as soon as week 1 after PRRSV infectionwere obtained. It was possible to isolate PRRSVand sometimes extended to the second week post-or to demonstrate PRRSV antibodies in some pig-infection. However, 3 weeks after inoculation, alllets, but two severely affected piglets were ap-PRRS-infected piglets had generally returned toparently not infected by PRRSV. However, theynormal body weight. In the case of our ex-showed indirect indications of a serious viral in-
fection. For example, macroscopical lesions of the perimental model of PMWS, growth retardation
Experimental Model for PMWS 301
When these inocula were used under our ex-perimental conditions, a disease was consistentlyinduced after a long incubation period (>12 dayson average), regardless of the health status of thepigs (SPF or conventional). After this incubationperiod, an increased rectal temperature was ob-served for 11 consecutive days and growth re-tardation for 2 to 3 weeks. This mild form of thedisease was reproduced in 82% of the inoculatedpiglets and in 90% of contact-exposed piglets. How-ever, a characteristic severe wasting syndrome wasinduced in only four inoculated piglets (7·2%): thiscan be compared with the incidence of PMWSobserved in well-managed farms, where only 5 to10% of the pigs showed a severe wasting syndrome(Madec et al., 2000). The comparatively low in-cidence of severe PMWS in well-managed farmsand in the present study can be attributed to therelative lack of secondary infections. Allan et al.(1999, 2000), Kennedy et al. (2000) and Krakowkaet al. (2000), demonstrated experimentally that asecondary agent such as PPV or PRRSV, whenco-injected with PCV2, increased the number ofpigs developing PMWS. Such results might explainwhy, in poor farming conditions, secondary in-fections may sometimes increase piglet mortalitydue to PMWS by up to 35% (Madec et al., 2000).In the present study, disease was transmitted to
% o
f po
siti
ve p
igs
in E
LIS
A
0
70
Weeks post-infection
(b)60
50
40
30
20
10
1 2 3 4 5
Mea
n a
bsor
ban
ce v
alu
es in
EL
ISA
0.000
3.500
Weeks post-infection
(a)3.000
2.500
2.000
1.500
1.000
0.500
1 2 3 4 5
90% of the direct contact-exposed piglets; it cannotFig. 5a,b. (a) Absorbance values of pig sera in the PCV2 be excluded, however, that good management pro-
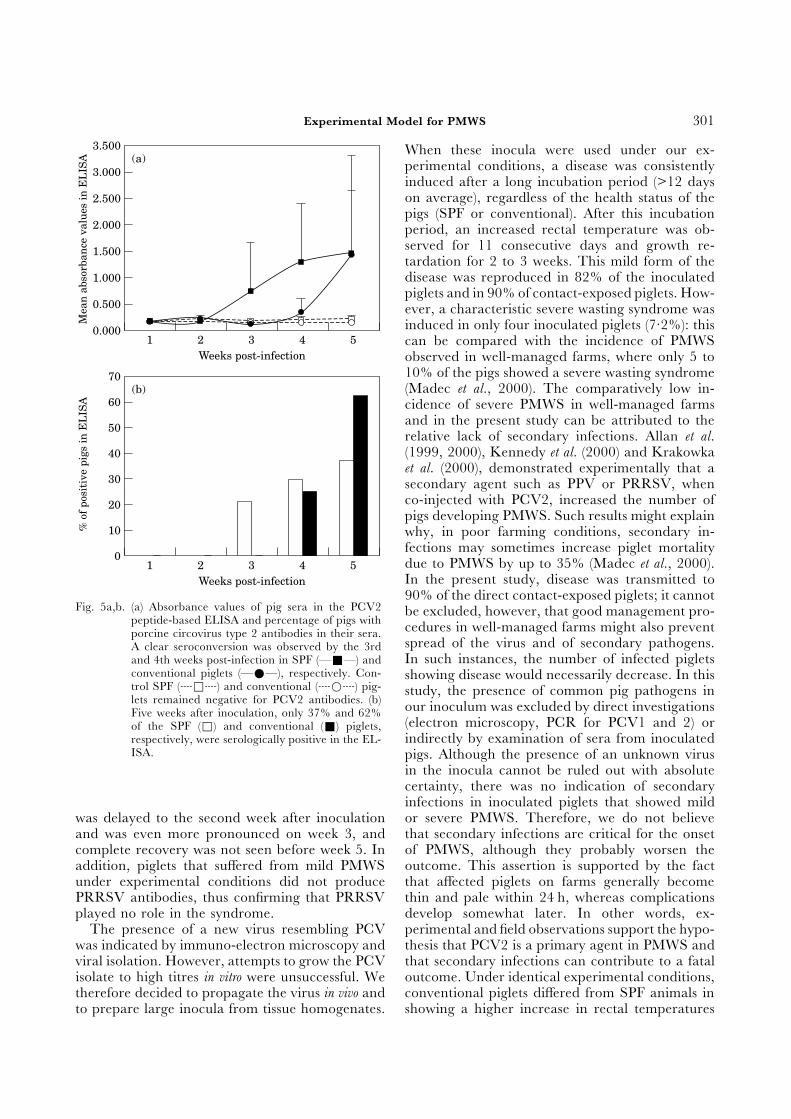
peptide-based ELISA and percentage of pigs with cedures in well-managed farms might also preventporcine circovirus type 2 antibodies in their sera.spread of the virus and of secondary pathogens.A clear seroconversion was observed by the 3rdIn such instances, the number of infected pigletsand 4th weeks post-infection in SPF (—Ε—) and
conventional piglets (—Χ—), respectively. Con- showing disease would necessarily decrease. In thistrol SPF (hΦh) and conventional (hΒh) pig- study, the presence of common pig pathogens inlets remained negative for PCV2 antibodies. (b) our inoculum was excluded by direct investigationsFive weeks after inoculation, only 37% and 62%
(electron microscopy, PCR for PCV1 and 2) orof the SPF (Φ) and conventional (Ε) piglets,indirectly by examination of sera from inoculatedrespectively, were serologically positive in the EL-
ISA. pigs. Although the presence of an unknown virusin the inocula cannot be ruled out with absolutecertainty, there was no indication of secondaryinfections in inoculated piglets that showed mildor severe PMWS. Therefore, we do not believewas delayed to the second week after inoculation
and was even more pronounced on week 3, and that secondary infections are critical for the onsetof PMWS, although they probably worsen thecomplete recovery was not seen before week 5. In
addition, piglets that suffered from mild PMWS outcome. This assertion is supported by the factthat affected piglets on farms generally becomeunder experimental conditions did not produce
PRRSV antibodies, thus confirming that PRRSV thin and pale within 24 h, whereas complicationsdevelop somewhat later. In other words, ex-played no role in the syndrome.
The presence of a new virus resembling PCV perimental and field observations support the hypo-thesis that PCV2 is a primary agent in PMWS andwas indicated by immuno-electron microscopy and
viral isolation. However, attempts to grow the PCV that secondary infections can contribute to a fataloutcome. Under identical experimental conditions,isolate to high titres in vitro were unsuccessful. We
therefore decided to propagate the virus in vivo and conventional piglets differed from SPF animals inshowing a higher increase in rectal temperaturesto prepare large inocula from tissue homogenates.

E. Albina et al.302
and greater growth retardation. It is possible that Referencesthe genetic and infectious background of these Allan, G., Kennedy, S., McNeilly, F., Foster, J., Ellis,animals affected their immune reactivity and J., Krakowka, S., Meehan, B. and Adair, B. (1999).thereby inclined them to develop a more pro- Experimental reproduction of severe wasting disease
by co-infection of pigs with porcine circovirus andnounced wasting syndrome after circovirus in-porcine parvovirus. Journal of Comparative Pathology,fection. Such a hypothesis was recently investigated121, 1–11.by Krakowka et al. (2001), who showed that ac-
Allan, G., McNeilly, F., Ellis, J., Krakowka, S., Meehan,tivation of the immune system could play a role inB., McNair, I., Walker, I. and Kennedy, S. (2000).the production of wasting disease. Experimental infection of colostrum deprived piglets
In the papers of Allan et al. (1999) and Kennedy with porcine circovirus 2 (PCV2) and porcine re-et al. (2000), PPV in association with PCV2 was productive and respiratory syndrome virus (PRRSV)particularly efficient in reproducing PMWS. Thus, potentiates PCV2 replication. Archives of Virology, 145,
2421–2429.only one of four piglets infected with PCV2 aloneAllan, G., McNeilly, F., Kennedy, S., Daft, B., Clarke,developed clinical signs, as compared with all of
E., Ellis, J., Haines, D., Meehan, B. and Adair, B.four dually infected piglets. In the present study,(1998a). Isolation of porcine circovirus-like virusesPPV was absent from the inocula and none of the from pigs with a wasting disease in the USA and
piglets developed antibody to this virus. Despite the Europe. Journal of Veterinary Diagnostic Investigation, 10,absence of PPV, more than 80% of the inoculated 3–10.piglets developed clinical signs. The possible Allan, G., Meehan, B., Todd, D., Kennedy, S., McNe-
illy, F. and Ellis, J. (1998b). Novel porcine circovirusesreasons for these discrepancies include the presencefrom pigs with wasting disease syndromes. Veterinaryof an additional unknown agent in the tissue homo-Record, 142, 467–468.genate, the strain pathogenicity, the inoculum titre
Balasch, M., Segales, J., Rosell, C., Domingo, M., Man-and the route of administration. The first of these kertz, A., Urniza, A. and Plana-Duran, J. (1999).possibilities seems unlikely, because recent evidence Experimental inoculation of conventional pigs with(to be published) indicated that pre-immunization tissue homogenates from pigs with post-weaningagainst PCV2 alone protected against infection by multisystemic wasting syndrome. Journal of Comparative
Pathology, 121, 139–148.the inoculum used in the present study. TheseCariolet, R., Marie, P., Moreau, G. and Robert, H.results confirmed that PCV2 was the primary agent
(1994). Summary of the different methods for pro-in our experimental model of PMWS. In the studyducing, maintaining and benefitting from piglets ofby Allan et al. (1999), each infected piglet received high health status. Journees Recherche Porcine en France,
102·4 TCID50 of PCV2 intranasally, whereas in 26, 1–12.our trials 5–9, each piglet received 105·2 TCID50 Ellis, J., Hassard, L., Clark, E., Harding, J., Allan, G.,and 104·5 TCID50 by the intratracheal and intra- Willson, P., Strokappe, J., Martin, K., McNeilly, F.,
Meehan, B., Todd, D. and Haines, D. (1998). Isolationmuscular routes, respectively. In a study by Balaschof circovirus from lesions of pigs with postweaninget al. (1999), a mild disease was reproduced in fivemultisystemic wasting syndrome. Canadian Veterinaryof eight piglets (62%) by intranasal infection withJournal, 39, 44–51.10 ml of a tissue homogenate (105·8 TCID50/ml). Ellis, J., Krakowka, S., Lairmore, M., Haines, D., Bra-
In contrast, Ellis et al. (1999), using an inoculum tanich, A., Clark, E., Allan, G., Konoby, C., Hassard,with a titre of only 102 TCID50, induced histo- L., Meehan, B., Martin, K., Harding, J., Kennedy,logical lesions in piglets but failed to induce clinical S. and McNeilly, F. (1999). Reproduction of lesions
of postweaning multisystemic wasting syndrome indisease.gnotobiotic piglets. Journal of Veterinary Diagnostic In-In conclusion, it would seem that the severity ofvestigations, 11, 3–14.PCV2-induced disease is dose-dependent. In this
Kaeffer, B., Bottreau, E., Phan-Than, L., Olivier, M.study, the failure of PCV2 to produce high viral and Salmon, H. (1990). Histocompatible miniaturetitres in vitro was overcome by propagation in vivo. boar model: selection of transformed cell lines of B andInoculum prepared by this means and checked to T lineages producing retrovirus. International Journal ofensure the absence of PCV1 and other common Cancer, 46, 481–488.
Kaerber, G. (1931). Beitrag zur kollektiven Behandlungviruses was consistently capable of producingpharmakologischer. Reihenversuche. Archiv fuer Phar-PMWS in SPF piglets. Frozen inoculum now avail-makologie und Experimentelle Pathologie, 162, 480–483.able is sufficient to enable the disease to be re-
Kennedy, S., Moffett, D., McNeilly, F., Meehan, B.,produced in approximately 700 piglets. It should Ellis, J., Krakowka, S. and Allan, G. M. (2000).therefore be of value in further studies on patho- Reproduction of lesions of postweaning multisystemicgenicity, immunity and vaccination in relation to wasting syndrome by infection of conventional pigs
with porcine circovirus type 2 alone or in combinationPCV2.

Experimental Model for PMWS 303
with porcine parvovirus. Journal of Comparative Path- microscopy of infected macrophages. Veterinary Micro-ology, 122, 9–24. biology, 33, 195–201.
Kim, H. S., Kwang, J., Yoon, I. J., Joo, H. S. and Plana-Duran, J., Balasch, M., Segales, J., Rosell, C.,Frey, M. L. (1993). Enhanced replication of porcine Rodrıguez-Arrioja, G., Domingo, M., Folch, J., San-reproductive and respiratory syndrome (PRRS) in chez, A. and Mankertz, A. (1999). Post-weaning mul-a homogeneous subpopulation of MA104 cell line. tisystemic wasting syndrome in Spain. Veterinary Record,Archives of Virology, 133, 477–483. 145, 3.
Kiupel, M., Stevenson, G., Mittal, S., Clark, E. and Pol, J. M. A., Van Dijk, J. E., Wensvoort, G. andHaines, D. (1998). Circovirus-like viral associated Terpstra, C. (1991). Pathological, ultrastructural anddisease in weaned pigs in Indiana. Veterinary Pathology, immunohistochemical changes caused by Lelystad35, 303–307. virus in experimentally induced infections of mystery
Krakowka, S., Ellis, J., McNeilly, F., Ringler, S., Rings, swine disease (synonym: porcine epidemic abortionD. and Allan, G. (2001). Activation of the immune and respiratory syndrome [PEARS]). Veterinary Quar-system is the pivotal event in the production of wasting terly, 13, 137–143.disease in pigs infected with porcine circovirus-2 Rosell, C., Segales, J., Plana-Duran, J., Balasch, M.,(PCV-2). Veterinary Pathology, 38, 31–42. Rodrıguez-Arrioja, G., Kennedy, S., Allan, G.,
Krakowka, S., Ellis, J., Meehan, B., Kennedy, S., McNe- McNeilly, F., Latimer, K. and Domingo, M. (1998).illy, F. and Allan, G. (2000). Viral wasting syndrome Pathological, immunohistochemical and in-situ hy-of swine: experimental reproduction of postweaning bridization studies of natural cases of postweaningmultisystemic wasting syndrome in gnotobiotic swine multisystemic wasting syndrome (PMWS) in pigs.by coinfection with porcine circovirus 2 and porcine Journal of Comparative Pathology, 120, 59–78.parvovirus. Veterinary Pathology, 37, 254–263. Samsom, J. N., de Bruin, T. G. M., Voermans, J. J. M.,
Le Cann, P., Albina, E., Madec, F., Cariolet, R. and Meulenberg, J. J. M., Pol, J. M. A. and Bianchi, A.Jestin, A. (1997). Piglet wasting disease. Veterinary Re- T. J. (2000). Changes of leukocyte phenotype andcord, 141, 660. function in the broncho-alveolar lavage fluid of pigs
Madec, F., Eveno, E., Morvan, P., Hamon, L., Blanch- infected with porcine reproductive and respiratoryard, P., Cariolet, R., Amena, N., Morvan, H., Truong, syndrome virus: a role for CD8+ cells. Journal ofC., Mahe, D., Albina, E. and Jestin, A. (2000). Post- General Virology, 81, 497–505.weaning multisystemic wasting syndrome (PMWS) in Segales, J., Sitjar, M., Domingo, M., Dee, S., Del Pozo,pigs in France: clinical observations from follow-up M. and Noval, R. (1997). First report of post-weaningstudies on affected farms. Livestock Production Sciences, multisystemic wasting syndrome in pigs in Spain.63, 223–233. Veterinary Record, 41, 600–601.
Mankertz, A., Domingo, M., Folch, J. M., Le Cann, P., Tischer, I., Bode, L., Peters, D., Pociuli, S. and Ger-Jestin, A., Segales, J., Chmielewicz, B., Plana-Duran, mann, B. (1995). Distribution of antibodies to porcineJ. and Soike, D. (2000). Characterisation of PCV-2 circovirus in swine populations of different breedingisolates from Spain, Germany and France. Virus Re- farms. Archives of Virology, 140, 737–743.search, 66, 65–77. Truong, C., Mahe, D., Blanchard, P., Le Dimna, M.,
McNeilly, F., Kennedy, S., Moffett, D., Meehan, B. M., Eveno, E., Morvan, P., Madec, F., Jestin, A. andFoster, J. C., Clarke, E. G., Ellis, J. A., Haines, D. M., Albina, E. (2001). Identification of an immunorelevantAdair, B. M. and Allan, G. M. (1999). A comparison of ORF2 epitope from porcine circovirus type 2 asin situ hybridization and immunohistochemistry for a serological marker for experimental and naturalthe detection of a new porcine circovirus in formalin- infection. Archives of Virology, 146, 1197–1211.fixed tissues from pigs with post-weaning multi- Wensvoort, G., Terpstra, C., Pol, J. M. A., ter Laak, E.systemic wasting syndrome (PMWS). Journal of Vir- A., Bloemraad, M., de Kluyver, E. P., Kragten, C.,ological Methods, 80, 123–128. van Buiten, L., den Besten, A., Wagenaar, F., Boe-
Morozov, I., Sirinarumitr, T., Sorden, S., Halbur, P., khuijsen, J. M., Moonen, P. L., Zetstra, T., De Boer,Morgan, M. and Yoon, K. (1998). Detection of a novel E. A., Tibben, H. J., De Jong, M. F., Van’t Veld, P.,strain of porcine circovirus in pigs with postweaning Groenland, G. J. R., Van Gennep, J. A., Voets, M.multisystemic wasting syndrome. Journal of Clinical T., Verheijden, J. H. M. and Braamskamp, J. (1991).Microbiology, 36, 2535–2541. Mystery swine disease in The Netherlands: the isol-
Nayar, G., Hamel, A. and Lin, L. (1997). Detection and ation of Lelystad virus. Veterinary Quarterly, 13, 121–characterization of porcine circoviruses associated 130.with postweaning multisystemic wasting syndrome inpigs. Canadian Veterinary Journal, 38, 385–386.
Paton, D. J., Brown, I. H., Scott, A. C., Done, S. H.
�Received, April 17th, 2000
Accepted, June 16th, 2001�and Edwards, S. (1992). Isolation of a Lelystad virus-like agent from British pigs and scanning electron





























