Embed Size (px)
Citation preview

A
L
Hbntictnpd
detvtlhmhtc
0d
3
n Animal Model of Human Cytomegalovirus Infection
. Gao, S. Qian, L. Zeng, R. Wang, G. Wei, J. Fan, and S. Zheng
ABSTRACT
To develop a rat model that allowed in vivo progressive human cytomegalovirus (HCMV)infection, allogeneic liver transplantation was performed across a rat combination of DarkAgouti (DA) to Brown Norway (BN). AD169, a well-characterized laboratory strain ofHCMV, was used to establish a rat model of HCMV infection by injection of 0.4 mL (30.0logTCID50) supernate into the rat peritoneum. Histological and blood specimens wereobtained from animals sacrificed at predetermined timepoints. We performed immuno-histochemical staining in liver, heart, kidney, spleen, and lung for HCMV immediate-earlyantigen (IE), lower matrix protein (pp65) detection in peripheral blood leukocytes, andHCMV early antigen (EA) and late antigen (LA). We compared survival rates. Our resultsshowed positive HCMV IE and pp65 antigenemia detected in peripheral blood leukocytesin transplanted recipients from day 1 to day 30. Positive HCMV EA and LA staining cellswere only detected in sections 10 days after liver transplantation, namely, in hepatocytes,mononuclear cells, bile duct epithelial cells, and endothelial cells. Successful HCMVreplication was due to the combination of liver transplantation and cyclosporine (CsA)immunosuppression. Survival analysis showed no significant differences between theHCMV-infected group and HCMV-uninfected group. This new rat model of HCMV
infection may be helpful to understand immune system modulation of HCMV infection.vpb(
a(aNSACSCHmZ
Cg
AQ
UMAN CYTOMEGALOVIRUS (HCMV), a betaherpesvirus, represents the major infectious cause of
irth defects, as well as an important pathogen for immu-ocompromised individuals. Sources of infection includeransplacental transfer, transplantation of an organ from annfected donor, exposure to infected saliva or urine duringhildhood, and sexual activity in adulthood.1 The advent ofhe AIDS epidemic and the increasing numbers of immu-osuppressed organ transplant, blood dyscrasia, and canceratients have facilitated an explosive increase in the inci-ence of active HCMV clinical disease.2
Successful animal models of HCMV infection have beenifficult to develop, because of the extreme species-specificityxhibited by members of the CMV family. Current systemso evaluate antihuman CMV agents use either the humanirus with human cells cultured in vitro3,4 or animal CMV inheir natural host species.5 Each of these approaches hasimitations for extrapolation to the complexities of theuman condition. Recently, several attempts have beenade to develop models of HCMV infection in which
uman fetal material is implanted into laboratory animalso support viral infection in vivo. Allen et al6 used MRC-5
ells (derived from human fetal lung) that were infected in E041-1345/07/$–see front matteroi:10.1016/j.transproceed.2007.06.077
438
itro with HCMV, entrapped in agarose plugs and im-lanted intraperitoneally into mice. Some models haveeen based on severe combined immunodeficiency diseaseSCID) mice, in which conjoint implants of human fetal
From the Department of Hepatobiliary and Pancreatic Surgerynd Centre of Organ Transplantation, The First Affiliated HospitalL.G., R.W.), and Department of Respiratory, The Second Affili-ted Hospital (L.Z.), Nanchang University School of Medicine,anchang, China; Department of Hepatobiliary and Pancreaticurgery and Centre of Organ Transplantation, The Secondffiliated Hospital, Zhongshan Medicine College, Guangzhou,hina (S.Q.); and Department of Hepatobiliary and Pancreaticurgery and Centre of Organ Transplantation, Key Lab ofombined Multi-Organ Transplantation, Ministry of Publicealth (S.Z.), Department of Hematopathy (G.W.), and Depart-ent of Infectious Diseases (J.F.), The First Affiliated Hospital,hejiang University School of Medicine, Hangzhou, China.Supported by the National Nature Science Foundation ofhina (grant 30170899) and the National Basic Research Pro-ram of China (973 program, grant 2003CB515501).Address reprint requests to Shusen Zheng, PhD, MD, The First
ffiliated Hospital, Zhejiang University School of Medicine, 79inchun Road, Hangzhou, Zhejiang Provence 310003, China.
-mail: [email protected]© 2007 by Elsevier Inc. All rights reserved.360 Park Avenue South, New York, NY 10010-1710
Transplantation Proceedings, 39, 3438–3443 (2007)

tsfsoaivt
HoppatHtHcMtbmstrpnoti
MR
Ipftclhtw(pActutw4w4(plr
rpiest
C
HgTPbitmtVca
Hp
Hmidpwd0lmwadaAns
HI
HcsaHbcZSt
S
Awup
ANIMAL MODEL OF HCMV 3439
hymus and liver placed under the kidney capsule wereubsequently infected with HCMV,7 or fragments of humanetal retina implanted in the anterior chamber of the eye toupport HCMV growth.8 Laycock and colleagues9 devel-ped an in vivo model of HCMV retinal infection inthymic rats. This model appeared to be capable of sustain-ng active HCMV replication for at least 3 to 4 weeks. Liveirus was cultured from the implanted material at the end ofhis period.9
At the same time immunocompetent animal models ofCMV infection were also reported by direct injection
f virus. Dunkel et al10 established an animal model ofrogressive HCMV chorioretinal disease by injection of 105
laque forming units (pfu) into the rabbit vitreous. Vitritisnd focal areas of immune cellular infiltrates were seen inhe inner retinal layers on days 2 to 4 after inoculation.CMV was recovered from chorioretinal cell sonicates in
iters ranging from 104 to 105 pfu during peak disease.CMV antigens were detected focally by immunofluores-
ence in the retinal layers on days 2 and 4 after inoculation.oreover, HCMV pneumonitis was also demonstrated in
his model, in which 30% to 40% of animals had developedronchial and alveolar edema/congestion, interstitial ede-a/infiltrates, and vascular congestion. All these findings
upported the replication and dissemination of HCMV inhe rabbit. Not only can HCMV infect immunocompetentats, but also HCMV can vertically transmit from thelacenta of mice to infect their offspring in the centralervous system.11 On the basis of these successes in devel-ping animal models of HCMV infection, we approachedhe development of a progressive HCMV infection modeln rats.
ATERIALS AND METHODSats and Liver Transplantation
nbred Brown Norway (BN) and Dark Agouti (DA) rats wereurchased from Beijing Vital River Company in which all coreoundation colonies were introduced from Charles River Labora-ories (CRL). All animals in these experiments were handled inompliance with the UCLA Animal Research Committee Guide-ines for the Care and Use of Laboratory Animals. All rats wereoused in the vivarium, and fed rat chow and water ad libitum untilhe night prior to transplantation when food was withheld. All ratsere male; donor rats (DA) were 200 to 300 g and recipient rats
BN) were 200 to 250 g. Nonarterialized liver transplantations wereerformed using the technique described by Kamada and Calne.12
ll surgical procedures were performed at room temperature withlean but not sterile microsurgical equipment. Rats were anesthe-ized with diethyl-ether. Cold (4°C) heparinized 0.9% saline wassed for perfusion and preservation of the graft. After operationhe rats were kept in single cages and fed a standard diet. Animalsere randomly assigned to 1 of 6 treatment groups. Group A (n �0 BN rats) was inoculated with HCMV. Group B (n � 40 BN rats)as injected with 0.4 mL of saline intraperitoncally. Group C (n �0 BN rats) was inoculated with HCMV and received cyclosporineCsA; 1 mg/kg/d subcutaneously). Group D (n � 24 liver trans-lanted BN rats) was inoculated with HCMV. Group E (n � 46
iver transplanted BN rats) was inoculated with HCMV and
eceived CsA treatment. Group F (n � 42 liver transplanted BN eats) was injected with saline intraperitoneally 1 day after trans-lantation and received CsA treatment. HCMV was inoculated by
ntraperitoneal injection of 0.4 mL of supernate whose cytopathicffect (CPE) was 30.0 logTCID50. CsA was administered byubcutaneous injection (1 mg/kg/d) from transplantation to 30 dayshere after.
ell Culture and HCMV Propagation
uman embryo fibroblast (HF cells) and CMV strain AD169 wereifts of Professor Wang Mingli from AnHui Medical University.he virus was passaged many times before being received fromrofessor Wang. CMV AD169 was propagated in human fibro-lasts. When it was used, the virus had been passaged many times
n the HF cells for increasing the CPE. Virus TCID50 was tested inhe HF cells according to Gibson’s13 CPE terminal dilutionethod. The virus, after 1 year of incubation, whose CPE increased
o 30.0 logTCID50/mL, was selected as the test virus to reserve.irus was collected when CPE was �90%. Supernates were
larified by centrifugation at 1000g for 10 minutes at 4°C and storedt �80°C until use.
CMV Immediate-Early Antigen andp65 Antigenemia Assay
CMV immediate-early (IE) antigen and pp65 antigenemia wereeasured in polymorphonuclear leukocytes of peripheral blood by
mmunochemical staining and the immunoperoxidase technique onays 1, 6, 10, 20, and 30 after HCMV injection. All specimens wererocessed after 2 to 4 hours of collection. In summary, leukocytesere extracted from a 2 mL aliquot of heparinized blood with 6%extran solution. Contaminating erythrocytes were lysed with a.8% ammonium chloride solution. After washing and counting theeukocyte suspension was adjusted to 106 cells/mL. One hundred
icroliters of each sample was transferred to a glass slide, fixedith acetone, and incubated with the pp65 or IE monoclonalntibodies (Dako, Denmark). Bound antibodies were subsequentlyetected with a rabbit anti-mouse antibody and a goat anti-rabbitntibody conjugated to horseradish peroxidase (Dako, Denmark).ntigenemia positive results were quantified by counting theumber of pp65 or IE expressing cells per 50,000 leukocytes on thelide.
CMV Early Antigen and Late Antigenmmunohistochemical Staining
CMV early antigen (EA) and late antigen (LA) immunohisto-hemical staining was performed on paraffin-embedded tissueections of liver, heart, lung, spleen, and kidney on day, 1, 6, 10, 20,nd 30 after HCMV injection using Power Vision Two-Stepistostaining Reagent. This kit offers superior sensitivity without
ackground interference. Mouse anti-CMV antibodies (CMV-LA,lone QB1/06; CMV-EA, clone QB1/42) were purchased fromymed Laboratories Inc (South San Francisco, Calif, Unitedtates). All procedures were performed according to the manufac-urer’s instructions.
urvival Studies and Statistics
fter sacrificing animals at predetermined times, survival analysisas studied between HCMV-infected groups and control groupssing the Kaplan-Meier method. The results of HCMV IE andp65 antigenemia were expressed as mean values � standard
rrors. For statistical analysis, ANOVA followed by the least
sssc
RS
AAepgt.Hifst1gMd1
H
Pttatbii5
ptsTe3aal1aTcitdtaapow
H
PcdoogdsHcbhttsiHo
D
AsvHhbHMCtT
FsiH(
3440 GAO, QIAN, ZENG ET AL
ignificant difference (LSD) or Bonferroni/Dunn test (SPSS 11.5oftware) was performed, which compensated for unequal groupize and repeated measurement. P � .05 was considered statisti-ally significant.
ESULTSurvival Analysis
ctuarial survival analysis of the 112 transplanted rats usingNOVA followed by LSD revealed no significant differ-
nces in the nontransplanted group. Among the trans-lanted group, there was a significant difference betweenroup D and the other groups because severe acute rejec-ion developed in group D after liver transplantation (P �01). There was no statistical difference between the
CMV-infected group (group E) and the non-HCMV-nfected group (group F; Fig 1). Five animals which diedrom operative complications were excluded from furthertudy. Median survival time (expressed as standard devia-ion) was 208 � 75 days (95% confidence interval [CI] of45–271) in group A, 227 � 66 days (CI of 172–282) inroup B, and 224 � days (CI of 180—267) in group C.edian survival times in groups D, E, and F were 12 � 2
ays (CI of 10–14), 41 � 12 days (CI of 31–52), and 58 �5 days (CI of 46–70), respectively.
CMV IE and pp65 Antigenemia Assay
ositive HCMV IE and pp65 antigenemia were only de-ected in transplanted group E (inoculated with HCMV andreated with CsA; Fig 2). No positive HCMV IE or pp65ntigenemia was detected in other groups, including theransplanted group D that had been inoculated with HCMVut had not been treated with CsA. The maximal HCMV pos-
tive IE antigen level was detected at day 1 after HCMVnjection and the number of antigenemia positive cells per0,000 leukocytes was 4.75 � 2.50 (Fig 3). At this time, the
ig 1. Kaplan-Meier survival analysis after HCMV infectionhowed no statistical difference between HCMV-infected recip-ents (group E) and non-HCMV-infected recipients (group F) orCMV-injected rats (groups A, C) and non-HCMV-injected rats
ogroup B).
ercent of positive antigenemia was 100%. From then on,he value and the percent of positive antigenemia progres-ively decreased until the 20th day after HCMV injection.he number of IE antigen positive cells per 50,000 periph-ral blood leukocytes (PBL) increased to 4.0 � 0.91 at day0 and positive IE antigenemia was detected in all animalst the same time. A high incidence of HCMV IE positiventigenemia was observed after HCMV injection. Theowest positive IE antigenemia (60%) was detected at day0. In contrast, the highest value of HCMV pp65 positiventigenemia was detected at day 10 after HCMV injection.he number of antigenemia positive cells per 50,000 leuko-ytes was 3.0 � 1.79, which corresponded to the peakncidence of HCMV pp65 positive antigenemia (80%) athis time. At day 20, the value of HCMV pp65 antigenecreased to the lowest level, and began to increase afterhat. Interestingly, the incidence of HCMV pp65 positiventigenemia at every time correlated with the numbers ofntigenemia positive cells. A lower incidence of HCMVp65 positive antigenemia was detected compared with thatf HCMV IE positive antigenemia, though the differenceas not significantly significant.
CMV EA and LA Immunohistochemical Double Staining
ositive HCMV EA and LA immunohistochemical stainingells which were found in the cytoplasm or nuclei were onlyetected in transplanted liver sections in group E and nonef those positive cells were discovered in heart, lung, spleen,r kidney sections in the same or other groups (Fig 4). Inroup E, positive EA and LA staining cells were firstetected at 10 days after HCMV injection. Progressivelytrong or staining cells were observed after this time.CMV was demonstrated in the liver by immunohisto-
hemistry; it was found in hepatocytes, mononuclear cells,ile duct epithelial cells, and endothelial cells of both theepatic artery and the portal vein. In most cases, distribu-ion of positive cells was more often detected in portalracts and severely destroyed tissues. All animals that werehown to be positive for HCMV EA and LA in the liver bymmunohistochemistry were also shown to be positive for
CMV IE or pp65 in PBL by antigenemia assay, though thenset of positive cells was different.
ISCUSSION
lthough the HCMV laboratory strains have species–pecificity, they have been propagated and refrigerated initro for many years. Experiments have demonstrated thatCMV can infect primary differentiated macrophages,14
uman brain capillary endothelial cells,15 human peripherallood mononuclear cells,16 and human neural cells.17
CMV also infects a wide variety of cells and tissues.18
oreover, HCMV and more importantly other animalMVs lack nucleotide homology, differing in their replica-
ion times and their responses to pharmacologic agents.19
hus, there is an urgent need to develop an animal model
f HCMV infection.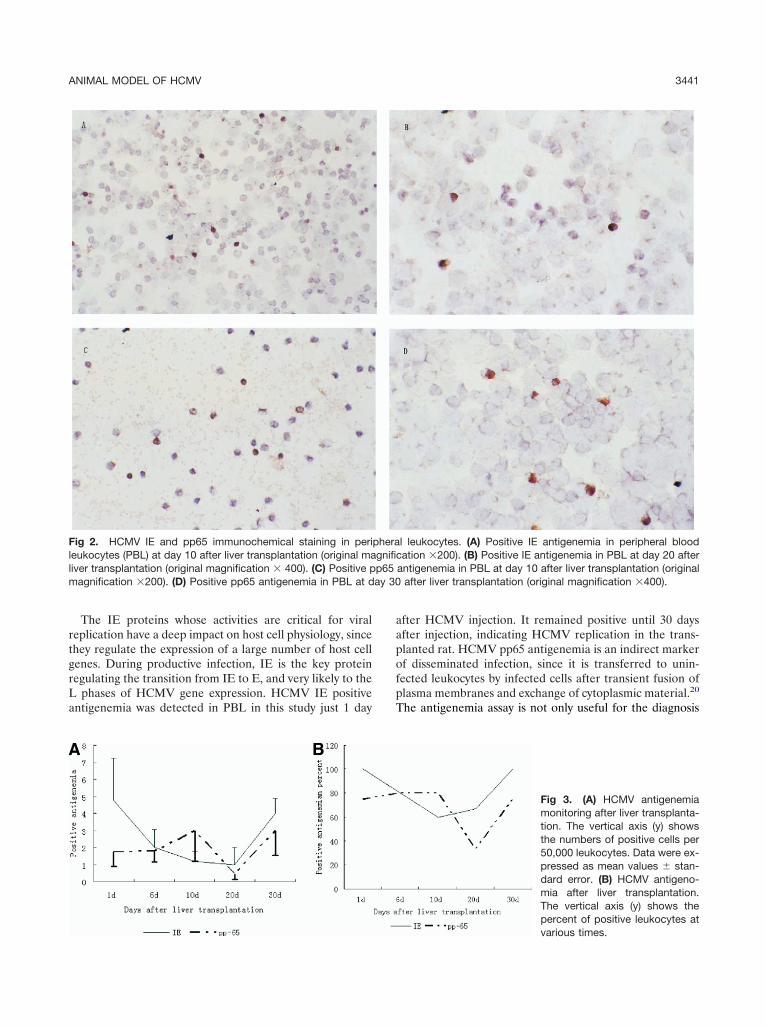
rtgrLa
aapofpT
Fllm ay 30
ANIMAL MODEL OF HCMV 3441
The IE proteins whose activities are critical for viraleplication have a deep impact on host cell physiology, sincehey regulate the expression of a large number of host cellenes. During productive infection, IE is the key proteinegulating the transition from IE to E, and very likely to the
phases of HCMV gene expression. HCMV IE positiventigenemia was detected in PBL in this study just 1 day
ig 2. HCMV IE and pp65 immunochemical staining in perieukocytes (PBL) at day 10 after liver transplantation (original maiver transplantation (original magnification � 400). (C) Positive p
agnification �200). (D) Positive pp65 antigenemia in PBL at d
fter HCMV injection. It remained positive until 30 daysfter injection, indicating HCMV replication in the trans-lanted rat. HCMV pp65 antigenemia is an indirect markerf disseminated infection, since it is transferred to unin-ected leukocytes by infected cells after transient fusion oflasma membranes and exchange of cytoplasmic material.20
he antigenemia assay is not only useful for the diagnosis
l leukocytes. (A) Positive IE antigenemia in peripheral bloodation �200). (B) Positive IE antigenemia in PBL at day 20 after
antigenemia in PBL at day 10 after liver transplantation (originalafter liver transplantation (original magnification �400).
Fig 3. (A) HCMV antigenemiamonitoring after liver transplanta-tion. The vertical axis (y) showsthe numbers of positive cells per50,000 leukocytes. Data were ex-pressed as mean values � stan-dard error. (B) HCMV antigeno-mia after liver transplantation.The vertical axis (y) shows thepercent of positive leukocytes at
pheragnificp65
various times.

ataHctcawbpss
bclpchpbOvvtsw
dpl
diaHvafpvnifi
fdWrTu
R
FhoospPlHdr2im�
3442 GAO, QIAN, ZENG ET AL
nd monitoring of HCMV infections, but also helpful forhe detection of treatment failures. In general, high levels ofntigenemia are detected in patients with disseminatedCMV disease, whereas low levels of antigenemia mostly
orrelate with asymptomatic infections. Moreover, the an-igenemia test is now considered to be more sensitive thanell cultures.21 Our study showed persistently positive pp65ntigenemia in PBL after HCMV injection, though its levelsere a little lower compared with threshold values that haveeen established in prospective studies to determine whichatients were at a greater risk of developing HCMV-relatedymptoms and may mostly benefit from timely initiation ofpecific therapy.
The primary localization of viral antigens determinedy immunohistopathological staining changed during theourse of infection. On day 1 after HCMV injection,eukocytes in PBL were positive for HCMV. On day 6,ositive antigenemia for HCMV was decreased in leuko-ytes. On day 10, the liver parenchyma was infected; theepatocytes had become mildly positive for the virus. IEositive antigenemia values in PBL decreased continuously,ut pp65 positive antigenemia levels increased to a peak.n day 20, the hepatocytes as well as bile duct cells and
ascular endothelia became intensively positive for theirus, but both IE and pp65 positive antigenemia decreasedo their lowest values. After this, all positive antigen expres-ion for HCMV in rats increased slightly. HCMV proteins
ig 4. (A) HCMV EA immuno-istochemical staining of rat livern day 10. (B) HCMV EA stainingf rat liver on day 20. Positivetaining of hepatocytes in cyto-lasm or nuclei was detected. (C)ositive HCMV LA staining of rat
iver on day 10. (D) PositiveCMV LA staining of rat liver onay 30. (E) HCMV LA staining ofat bile duct epithelial cells on day0. (F) Negative HCMV EA stain-
ng of rat liver on day 6. (Originalagnifications �150; A,C,F;600 B,D,E.)
hich represent various phases of the replication cycle were T
etected not only in PBL but also in liver sections. Inarticular, positive cells were found in the most damaged
iver structures.Our study showed that successful HCMV replication was
ue to a combination of liver transplantation and CsAmmunosuppression. Just liver transplantation or CsAlone was not sufficient for successful HCMV replication inCMV incubated rats. Immunologic injury, immune acti-
ation, and immunosuppression may be involved in virusctivation and replication, the mechanisms of which needurther research. Extensive research has focused on theossible relationship between CMV infection and the de-elopment of chronic allograft dysfunction. Features of thisewly developed animal model are mimicked by HCMV
nfection after solid organ transplantation, and are useful forurther study to better understand the role of HCMV infectionn the pathogenesis of chronic allograft dysfunction.
In conclusion, our study demonstrated that HCMV in-ected and replicated within immunocompromised rats butid not infect and replicate within immunocompetent rats.e demonstrated infected cells which expressed proteins
epresentative of all phases of the HCMV replication cycle.his model may be helpful for further research to betternderstand immune system modulation of HCMV infection.
EFERENCES
1. Meyers J: Cytomegalovirus infection after organ allografting.he Herpesviruses 4:201, 1985

ip
r
pc
ga
aA
mf4
i7
o1
fD
hc
h
C
i
bH
nt
nl
rR
T
lte
cb
ANIMAL MODEL OF HCMV 3443
2. Faber DW, Wiley CA, Lynn GB, et al: Role of HIV and CMVn the pathogenesis of retinitis and retinal vasculopathy in AIDSatients. Invest Ophthalmol Vis Sci 33:2345, 19923. Hai R, Chu A, Li H, et al: Infection of human cytomegalovi-
us in cultured human gingival tissue. Virol J 3:84, 20064. Sinzger C, Bissinger AL, Viebahn R, et al: Hepatocytes are
ermissive for human cytomegalovirus infection in human liver cellulture and in vivo. J Infect Dis 180:976, 1999
5. Loh HS, Mohd-Lila MA, Abdul-Rahman SO, et al: Patho-enesis and vertical transmission of a transplacental rat cytomeg-lovirus. Virol J 3:42, 2006
6. Allen LB, Li SX, Arnett G, et al: Novel method for evaluatingntiviral drugs against human cytomegalovirus in mice. Antimicrobgents Chemother 36:206, 19927. Kern ER, Bidanset DJ, Hartline CB, et al: Oral activity of aethylenecyclopropane analog, cyclopropavir, in animal models
or cytomegalovirus infections. Antimicrob Agents Chemother8:4745, 20048. DiLoreto DJ, Epstein LG, Lazar ES, et al: Cytomegalovirus
nfection of human retinal tissue: an in vivo model. Lab Invest1:141, 19949. Laycock KA, Fenoglio ED, Hook KK, et al: An in vivo model
f human cytomegalovirus retinal infection. Am J Ophthalmol24:181, 199710. Dunkel EC, de Freitas D, Scheer DI, et al: A rabbit model
or human cytomegalovirus-induced chorioretinal disease. J Infectis 168:336, 199311. Tang JL, Wang ML, Qiu JJ, et al: Building a mouse model
allmarking the congenital human cytomegalovirus infection in
entral nervous system. Arch Virol 147:1189, 2002 M12. Kamada N, Calne RY: A surgical experience with fiveundred thirty liver transplants in the rat. Surgery 93:64, 198313. Gibson W: Structural and nonstructural proteins of strain
oluburn cytomegalovirus. Virology 111:516, 198114. Ibanez CE, Schrier R, Ghazal P, et al: HCMV productively
nfects primary differentiated macrophages. J Virol 65:658, 199115. Lathey JL, Wiley CA, Verity MA, et al: Cultured human
rain capillary endothelial cells are permissive for infection byCMV. Virology 176:266, 199016. Soderberg C, Larsson S, Bergstedt-Lindqvist S, et al: Defi-
ition of a subset of human peripheral blood mononuclear cellshat are permissive to HCMV infection. J Virol 67:3166, 1993
17. McCarth M, Resnick L, Taub F, et al: Infection of humaneural cell aggregate cultures with a clinical isolate of cytomega-
ovirus. J Neuropathol Exp Neural 50:441, 199118. Plachter B, Sinzger C, Jahn G: Cell types involved in
eplication and distribution of human cytomegalovirus. Adv Viruses 46:195, 199619. Mocarski ES Jr: Biology and replication of cytomegalovirus.
ransfus Med Rev 2:229, 198820. Gerna G, Percivalle E, Baldanti F, et al: Human cytomega-
ovirus replicates abortively in polymorphonuclear leukocytes afterransfer from infected endothelial cells via transient microfusionvents. J Virol 74:5629, 2000
21. St George K, Boyd MJ, Lipson SM, et al: A multisite trialomparing two cytomegalovirus (CMV) pp65 antigenemia test kits,iotest CMV brite and Bartels/Argene CMV antigenemia. J Clin
icrobiol 38:1430, 2000




























