Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMlSTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 267, No. 29, Issue of October 15, pp. 20728-20733,1992 Printed in U.S.A.
Alterations in Proglucagon Processing and Inhibition of Proglucagon Gene Expression in Transgenic Mice Which Contain A Chimeric Proglucagon-SV40 T Antigen Gene*
(Received for publication, May 11, 1992)
Patricia L. BrubakerS, Ying C. Lee, and Daniel J. DruckerP From the Departments of Physiology, Medicine, Clinical Biochemistry, and Genetics and the Banting and Best Diabetes Center, University of Toronto, Toronto, Ontario M5S 1A8, Canada
The proglucagon gene is expressed in A cells of the pancreas and L cells of the large and small intestine. Transgenic mice expressing SV40 large T antigen un- der the control of proglucagon regulatory sequences develop neuroendocrine carcinoma of the large intes- tine. To determine the consequences of coexpression of S V 4 0 large T antigen and proglucagon in different cell types, the levels of proglucagon mRNA transcripts and proglucagon-derived peptides were determined in tumor-bearing transgenic mice and in age- matched paired controls. Plasma levels of proglucagon- derived peptides (glicentin, oxyntomodulin, and glu- cagon, as determined by high pressure liquid chroma- tography and radioimmunoassay) were markedly ele- vated in association with tumor growth ( p < 0.001). Northern blot analysis demonstrated that the increased concentration of proglucagon-derived peptides was as- sociated with significant inhibition of the endogenous proglucagon gene in pancreas, and to a lesser extent, small intestine. Concomitantly, the concentrations of proglucagon-derived peptides fell to 1-1070 of control values in pancreas ( p < 0.001) and to 62% of control values in small intestine ( p < 0.001). Analysis of pro- glucagon-derived peptides in mice of different ages demonstrated that tumor growth was associated with a switch in the post-translational processing of proglu- cagon. Compared with normal mouse intestine, tumors contained increased proportions of glucagon and glu- cagon-like peptide- l(7-37) relative to glicentin, ox- yntomodulin, and glucagon-like peptide-l( 1-37). The results of these studies provide evidence for humorally- mediated tissue-specific inhibition of proglucagon gene expression.
The proglucagon gene is expressed in the A cells of the endocrine pancreas, L cells of the intestinal epithelium and selected neurons of the hypothalamus and brainstem (1-3). Tissue-specific post-translational processing of proglucagon generates a wide diversity of biologically active peptides (1,4, 5). Studies of glucagon biosynthesis in normal pancreatic
* These studies were supported by grants from the Medical Re- search Council of Canada and the Canadian Diabetes Association. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Diabetes Canada Scholar. To whom correspondence should be addressed Rm. 3366, Medical Sciences Bldg., University of Toronto, Toronto, Ontario M5S 1A8, Canada. Tel.: 416-978-2593; Fax: 416- 978-4940.
J Career Scientist of the Ontario Ministry of Health.
islets have demonstrated that proglucagon is cleaved first at the N-terminal end of the glucagon sequence to release the 30-amino acid fragment glicentin-related pancreatic peptide and subsequently at the C-terminal end of glucagon to gen- erate glucagon, small amounts of the 9-kDa peptide, and the major proglucagon fragment containing glucagon-like peptide (GLP)’-l and GLP-2 (Fig. 1) (6, 7). In the intestine, in contrast, it has been proposed that GLP-2 is generated first and that GLP-1 is subsequently cleaved, leaving glicentin and oxyntomodulin (7). GLP-1 then undergoes further N-terminal truncation to produce GLP-1(7-37) or GLP-1(7-36NH2) (1, 8), potent glucose-dependent stimulators of insulin biosyn- thesis and secretion (9-12).
We have recently established several lines of transgenic mice using 2000 base pairs of rat glucagon gene 5”flanking sequences fused to the coding sequence for SV40 large T antigen (GLUTag) (13). One of these lines, designated GLU- Tag-Y, expresses the transgene in pancreas, small and large intestine, and brain and develops invasive neuroendocrine carcinoma of the large intestine by 4-8 weeks of age. In contrast, expression of the transgene in the stomach and small bowel had no effect on the morphology of these tissues, whereas transgene expression in the endocrine pancreas pro- duced islet cell hyperplasia progressing to pancreatic endo- crine carcinoma in older mice. The effects of SV40 T antigen on proglucagon gene expression and on the post-translational processing of proglucagon have not been determined. We have therefore utilized the GLUTag-Y mouse model to study the consequences of oncogene expression on the regulation of endogenous proglucagon gene expression and the biosynthesis and secretion of the proglucagon-derived peptides.
MATERIALS AND METHODS
Animals-The transgenic mouse line was generated using a 2.1- kilobase rat glucagon gene EcoRI/AccI fragment ligated to a BglI/ BamHI fragment of SV40 T antigen coding sequence (GLUTag), as described previously (13). For examination of proglucagon-derived peptides in tissues and plasma, GLUTag-Y mice were studied at 4 ( n = 6) and 8 (n = 6) weeks of age, as were paired control littermates (n = 9). Plasma for HPLC was collected from two litters of 10-11-week- old wild-type mice (n = 8 and 9) and from individual 10-week-old transgenic mice ( n = 9), as well as from a litter of 10.5-week-old transgenic mice (n = 9). Northern blot analysis was carried out on tissues collected from a minimum of three mice a t day 1, day 10, and 6 weeks of age.
Peptide Analysis-Mice were anesthetized with CO, and exsan- guinated by cardiac puncture. Blood was collected into 50 pl of Trasylo1:EDTA (5000 KIU/ml:1.2 mg/ml), and the plasma was stored
The abbreviations used are: GLP, glucagon-like peptide; HPLC, high performance liquid chromatography; GLI, glucagon-like immu- noreactive; IRG, immunoreactive glucagon; IRI, immunoreactive in- sulin.
20728

Intestinal Proglucagon-producing Tumors 20729
Proglucagon
Glicentin MPGF
Oxyntomodulin GLP-1 (7-37) v
a
9 K
Glucagon
FIG. 1. Generalized scheme of proglucagon and proglu- cagon-derived peptides. GRPP, glicentin-related pancreatic pep- tide; MPGF, major proglucagon fragment. The sequences recognized by antisera K4023 (solid triangle), 04A (open triangle), and b5 (double triangle) are indicated.
at -70 "C prior to analysis. The entire small intestine and the upper half of the large intestine were cleaned, and the intestinal segments and pancreas were then homogenized twice in 5-7 ml of extraction medium (1 N HC1 containing 5% (v/v) formic acid, 1% (v/v) trifluo- roacetic acid, and 1% (w/v) NaCl (5, 14, 15)) at 4 "C. Peptides and small proteins were adsorbed from the extracts by passage twice through a cartridge of C18 silica (Waters Associates, Milford, MA). Adsorbed peptides were then eluted with 4 ml of 80% (v/v) isopro- panol containing 0.1% (v/v) trifluoroacetic acid. We have demon- strated previously a greater than 88% recovery of intact proglucagon- derived peptides from tissues using this methodology (14, 16).
Radioimmunoassays for glucagon-like immunoreactive (GLI) pep- tides, immunoreactive glucagon (IRG), and immunoreactive GLP-1 were carried out using three different antisera, respectively (Fig. 1): 1) Antiserum K4023 (Novo Alle, Bagsvaerd, Denmark) (17), which cross-reacts with the mid-sequence of glucagon and recognizes glu- cagon and both N- and C-terminally extended forms of glucagon (i.e. glicentin, oxyntomodulin, and 9-kDa glucagon), 2) antiserum 04A (Dr. R. H. Unger, Dallas, TX) (18), which cross-reacts with the free C-terminal end of glucagon, and recognizes glucagon and N-termi- nally modified forms of glucagon (i.e. 9-kDa glucagon), and 3) anti- serum b5 (a gift from Dr. S. Mojsov, New York) (1, 8), which cross- reacts with the C-terminal end of GLP-1 and recognizes GLP-1(1- 37) and GLP-l(7-37), but not C-terminally modified forms of GLP- 1 (i.e. GLP-1(7-36NHs) or the major proglucagon fragment which contains GLP-1 and GLP-2). Immunoreactive insulin (JRI) levels were determined by radioimmunoassay using Wright's antiserum (19), protein assays were carried out using the method of Lowry et al. (201, and blood glucose levels were determined using the glucose oxidase method (Glucose Autoanalyzer 2, Beckman Instruments, Fullerton, CA).
Tissue or plasma peptides were separated by HPLC using a Waters Associates Liquid Chromatography System and a C18 pBondapak column. The solvent system used for analysis of GLI and IRG peptides was a 45-min linear gradient of 25-62.5% (v/v) Solvent B (Solvent A, 1% (v/v) trifluoroacetic acid buffered to pH 2.5 with diethylamine; Solvent B, 80% (v/v) acetonitrile), as described previ- ously (5, 15). The flow rate was 1.5 ml/min and 0.3-min fractions were collected. Peptides contained in plasma were extracted and deproteinized before HPLC by passage twice through a cartridge of C18 silica. HPLC of GLP-1-containing peptides was carried out using a modification of the method of Mojsov et al. (8). In brief, peptides were separated on a C18 pBondapak column using a 30-min linear gradient of 45-68% (v/v) Solvent B (Solvent A, 0.085% (v/v) phos- phoric acid and 0.3% (v/v) triethylamine, pH 7.0; Solvent B, 40% (v/ v) solvent A in 60% (v/v) acetonitrile). The flow rate was 1.5 ml/min, and 1-min fractions were collected. All HPLC fractions were stored at -20 "C prior to radioimmunoassay.
mRNA Analysis-Total cellular RNA was isolated from freshly dissected tissues by homogenization in guanidium isothiocyanate, as described previously (2, 13). RNA was fractionated on a 1.3% (w/v) agarose-formaldehyde gel and the integrity of the RNA assessed by ethidium bromide staining. RNA was transferred onto a nylon mem- brane, fixed by UV irradiation, and prehybridized and hybridized in 1 X Denhardt's, 4 X SSC (1 X SSC is 0.15 M NaCl and 0.015 M
Con1 4 wk 8 wk Con1 4 wk 8 wk
n l & l z 240 D
4 Wk 8 wk
FIG. 2. Concentrations of GLI ( A ) , IRG ( B ) , IRI ( C ) , and glucose (D) in plasma from age-matched control mice (Cont) or from GLUTag-Y transgenic mice at 4 and 8 weeks of age. Due to limitations in plasma volume, IRG and IRI levels were determined in only two animals per age group; no standard error has been calculated for this data, and statistical differences were not assessed. *, p < 0.05; **, p < 0.01; ***,p < 0.001.
sodium citrate), 200 pg/ml salmon sperm DNA, and 40% (v/v) deion- ized formamide in 0.014 M Tris (hydroxymethyl)aminomethane, pH 7.4. Full-length rat proglucagon and proinsulin cDNAs were labeled to a specific activity of 5 X 10' cpm/pg with [m3*P]dATP by the random priming method, and hybridization with 1 X lo6 cpm/ml 3zP- labeled cDNA probe was carried out for 24 h a t 42 "C. The blot was then washed with 0.1 X SSC, 0.1% (w/v) sodium dodecyl sulfate a t 65 "C. Autoradiography was carried out using Kodak X-Omat film (Eastman Kodak, Rochester, NY) a t -70 "C.
Data Analysis-All data are expressed as mean & S.E. Values for blood glucose and for tissue and plasma GLI, IRG, GLP-1, and IRI were found to be identical for control mice a t 4 and 8 weeks of age. Therefore, data for control mice a t 4 and 8 weeks of age were pooled for statistical analysis. Statistical differences between samples of n 2 3 were assessed by analysis of variance using a Statistical Analysis System package for personal computers (SAS Institute Inc., Cary, NC). Significance was assumed a t p 5 0.05.
RESULTS
To determine if tumor growth in the large bowel was associated with an increase in the plasma levels of proglu- cagon-derived peptides, the concentrations of GLI and IRG were determined in whole plasma from GLUTag-Y mice and age-matched controls (Fig. 2). At 4 and 8 weeks of age, GLI concentrations were increased by 20- and 42-fold ( p < 0.001), respectively, relative to controls, and those of IRG were increased by 4- and 20-fold, respectively. In contrast to the changes in GLI and IRG levels, plasma concentrations of IRI in transgenic mice were comparable with those of control mice at 4 weeks of age and decreased to 15% of control levels by 8 weeks of age. Similarly, blood glucose levels in GLUTag-Y mice were similar to control values at 4 weeks of age but decreased at 8 weeks of age, to 63% of control values ( p < 0.01). Plasma samples were examined by HPLC to determine the circulating forms of GLI and IRG in GLUTag-Y mice (Fig. 3). In control mice, plasma contained predominantly oxyntomodulin (59% of total peak GLI), with lesser peaks of glicentin and glucagon (25 and 16% of total peak GLI, re- spectively). Several other unidentified peaks of GLI were also detected. In plasma pooled from two separate groups of 10- 10.5-week-old GLUTag-Y mice, the proportion of total peak GLI constituted by glicentin was increased relative to controls ( to 65% of the total peak GLI, as compared with 25% in controls). In contrast, the relative proportions of total peak GLI represented by oxyntomodulin and glucagon were de- creased to 28 and 8%, respectively, as compared with 59 and

Intestinal Proglucagon-producing Tumors
1" k b 100
50
n 60 70 80 90 100 110
600
400
200
0
600
400
200
0 60 70 80 90 100 110
600
400 200
0 60 70 SO 90 100 110
Fraction #
FIG. 3. HPLC separation of GLI (solid squares) and IRG (open squares) peptides contained in 1 ml of plasma from 17 control mice (A), 9 GLUTag-Y mice at 10 weeks of age ( B ) , and 9 GLUTag-Y mice at 10.5 weeks of age (C) . The peaks are identified as glicentin ( a ) , oxyntomodulin ( b ) , and glucagon (c), based upon immunoreactivity and similar analyses of rat plasma, panc- reatic, and intestinal extracts (15). The elution position of synthetic glucagon is indicated by the arrowhead.
16%, respectively, in control mice. To establish whether SV40 T antigen gene expression and/
or increased circulating concentrations of proglucagon-de- rived peptides had any effect on expression of the endogenous proglucagon gene, proglucagon mRNA transcript levels were examined in extracts of the pancreas and the gastrointestinal tract. At day 1 and day 10 postnatally, transgenic mice and control mice were found to have comparable levels of proglu- cagon mRNA transcripts in the stomach, pancreas, and intes- tine (Fig. 4), although the levels of SV40 T antigen mRNA transcripts are reduced in the pancreas of day 10 mice (13). By 6 weeks of age, pancreatic proglucagon mRNA transcript levels were found to be reduced to almost undetectable levels in transgenic mice. Proinsulin mRNA transcript levels in transgenic mice were also reduced but were readily detectable. Proglucagon mRNA transcript levels in the small intestine were also diminished by 6 weeks of age in transgenic mice. In contrast to the changes seen in pancreas and small intestine, expression of the endogenous proglucagon gene in the large intestine of GLUTag-Y mice increased markedly a t 6 weeks of age. These changes occurred concomitantly with tumor growth in the large intestine, as determined by histogical examination of these tissues (13).
To determine whether the decreases in proglucagon mRNA transcript levels in pancreas and small intestine were associ- ated with alterations in tissue proglucagon biosynthesis, the levels of the proglucagon-derived peptides were determined. In parallel with the reduction in proglucagon mRNA trans- cript levels in the pancreas, the concentrations of GLI, IRG, and GLP-1 in the pancreas were all significantly reduced by 4 weeks of age ( p < 0.001) and had markedly decreased to 0.5, 0.4, and 10.3% of control levels by 8 weeks ( p < 0.001; Fig. 5). In contrast, pancreatic insulin concentrations rose significantly at 4 weeks ( p < 0.001) but returned to control
PAN SB LB WT TG WT TG WT TG
FIG. 4. A , Northern blot analysis of proglucagon ( G ) mRNA tran- scripts in stomach (ST), pancreas ( P A N ) , whole intestine (ZNT), small bowel ( S B ) , and large bowel ( L B ) from transgenic (TG) and wild-type ( WT) mice at 1 ( D l ) and 10 (DIO) days of age. Each lane contains 10 pg of total cellular RNA. The blot shown is representative of at least three separate analyses. B, Northern blot analysis of proglucagon (G) and proinsulin ( I ) mRNA transcripts from pancreas (PAN) , small bowel ( S B ) , and large bowel (LB) from wild-type ( WT) and transgenic (TG) mice at 6 weeks of age. Each lane contains 10 pg of total cellular RNA except the LB-TG lane, which contains 1 pg of total cellular RNA. The blot shown is representative of at least three separate analyses. Accurate densitometric analysis of the rela- tive amounts of proglucagon mRNA transcripts was not possible due to the barely detectable autoradiographic signal in the PAN- and SB- TG lanes.
CONT 4 W K 8 W K 300 1B
CONT 4'fK 8 W K
CONT 4 WK 8 W K CONT 4 W K 8 W K
FIG. 5. Concentrationsof GLI (A), IRG ( B ) , GLP-1 (C), and IRI (0) in extracts of pancreas from age-matched control mice (CONT) or from GLUTag-Y transgenic mice at 4 and 8 weeks of age. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
levels by 8 weeks of age. The concentrations of proglucagon- derived peptides in the small intestine were also reduced to 62% of control values by 8 weeks of age ( p < 0.001; Fig. 6).
Analysis of the proglucagon-derived peptide content of the large intestine demonstrated that the increment in proglu- cagon mRNA transcript levels with tumor growth was asso- ciated with significant and progressive increases in the con- centrations of GLI, IRG, and GLP-1 to 950% ( p < 0.001), 5400% ( p < 0.001), and 1100% ( p < 0.01) of control values, respectively, by 8 weeks of age (Fig. 7). Thus, the increment in IRG during tumor growth was found to be much greater than that in GLI. This was confirmed by analysis of the ratio
Intestinal Proglucagon-producing Tumors 20731
31 2 0 1 CONT 4 WK 8 WK
0.5 ~:~~ 0.0 CONT
4 WK 8 WK
FIG. 6. Concentrations of GLI ( A ) , IRG ( B ) , and GLP-1 (C) in extracts of small intestine from age-matched control mice (CONT) or from GLUTag-Y transgenic mice at 4 and 8 weeks of age. *, p < 0.05; **, p < 0.01; ***, P < 0.001.
.- 30001 Q
2 P
2000
CL 1 0 o o I
. 0
- C O M
4t
3-n 4 WK 0 WK
* 4t -r
CONT 4 WK 8 WK
E' .- 80-c
2 60-
2 40-
c 20-
- Q
P 0
0 P Y
*
i 5 11-I I I I 1 0 0
CONT 4 WK 8 WK
FIG. 7. Concentrations of GLI ( A ) , IRG ( B ) , and GLP-1 (C) in extracts of large intestine from age-matched control mice (CONT) or from GLUTag-Y transgenic mice at 4 and 8 weeks of age. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
of GLI to IRG peptides (an index of glicentin plus oxynto- modulin to glucagon), which was 42 & 8 in control large intestine but decreased to 7.0 & 0.4 and 5.4 & 0.7 ( p C 0.001) in the 4- and 8-week-old tumors, respectively.
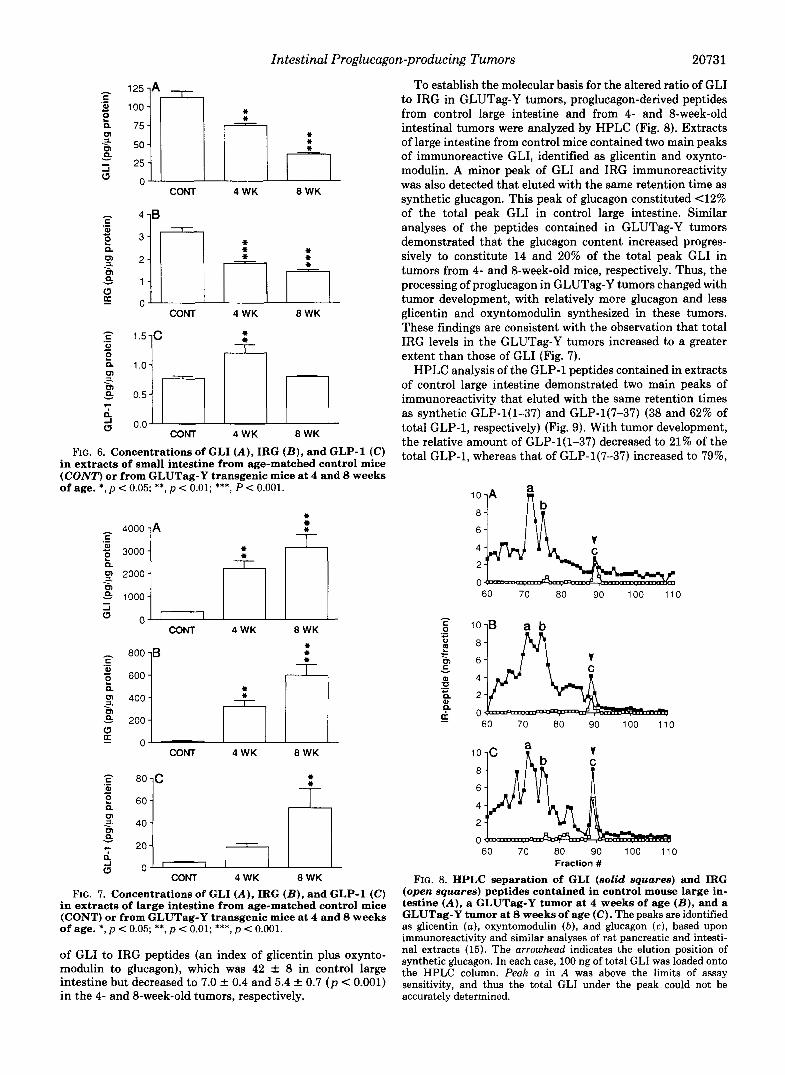
To establish the molecular basis for the altered ratio of GLI to IRG in GLUTag-Y tumors, proglucagon-derived peptides from control large intestine and from 4- and 8-week-old intestinal tumors were analyzed by HPLC (Fig. 8). Extracts of large intestine from control mice contained two main peaks of immunoreactive GLI, identified as glicentin and oxynto- modulin. A minor peak of GLI and IRG immunoreactivity was also detected that eluted with the same retention time as synthetic glucagon. This peak of glucagon constituted 4 2 % of the total peak GLI in control large intestine. Similar analyses of the peptides contained in GLUTag-Y tumors demonstrated that the glucagon content increased progres- sively to constitute 14 and 20% of the total peak GLI in tumors from 4- and 8-week-old mice, respectively. Thus, the processing of proglucagon in GLUTag-Y tumors changed with tumor development, with relatively more glucagon and less glicentin and oxyntomodulin synthesized in these tumors. These findings are consistent with the observation that total IRG levels in the GLUTag-Y tumors increased to a greater extent than those of GLI (Fig. 7).
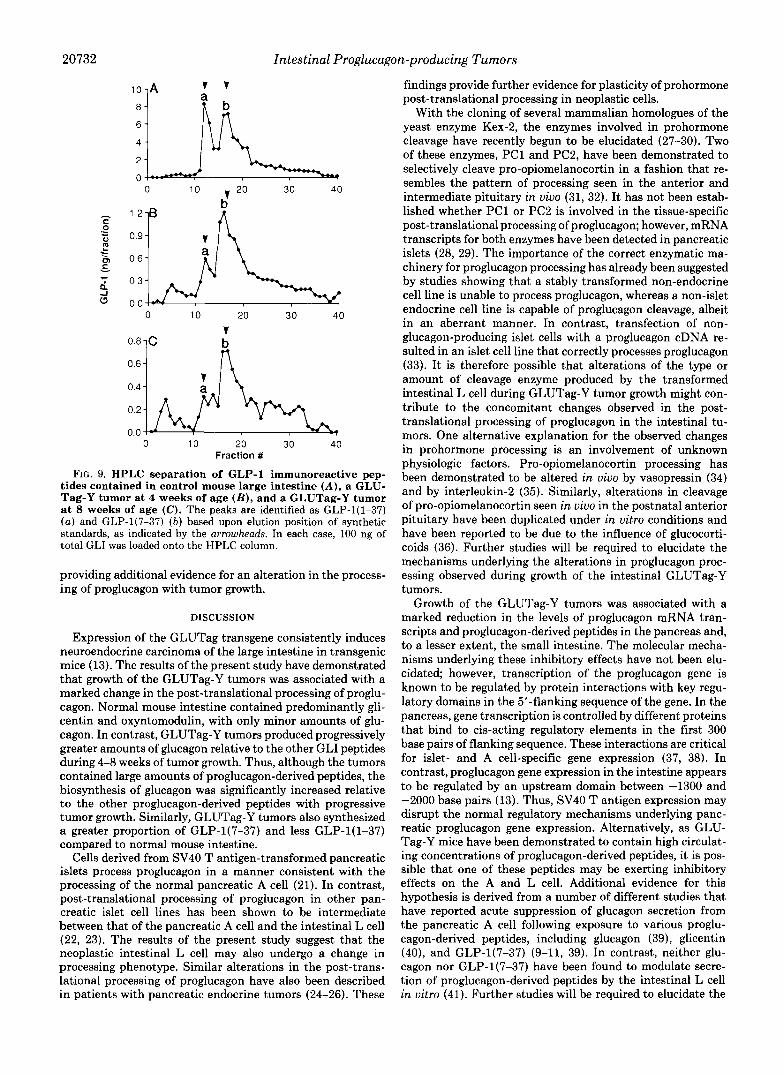
HPLC analysis of the GLP-1 peptides contained in extracts of control large intestine demonstrated two main peaks of immunoreactivity that eluted with the same retention times as synthetic GLP-l(1-37) and GLP-1(7-37) (38 and 62% of total GLP-1, respectively) (Fig. 9). With tumor development, the relative amount of GLP-l(1-37) decreased to 21% of the total GLP-1, whereas that of GLP-1(7-37) increased to 79%,
60 70 80 90 100 110
60 70 80 90 100 110
60 70 80 90 100 110 Fraction #
FIG. 8. HPLC separation of GLI (solid squares) and IRG (open squares) peptides contained in control mouse large in- testine ( A ) , a GLUTag-Y tumor at 4 weeks of age ( B ) , and a GLUTag-Y tumor at 8 weeks of age (C). The peaks are identified as glicentin ( a ) , oxyntomodulin ( b ) , and glucagon (c), based upon immunoreactivity and similar analyses of rat pancreatic and intesti- nal extracts (15). The arrowhead indicates the elution position of synthetic glucagon. In each case, 100 ng of total GLI was loaded onto the HPLC column. Peak a in A was above the limits of assay sensitivity, and thus the total GLI under the peak could not be accurately determined.
I O - A 0 -
6 -
4 -
v v
Intestinal Proglucagon-producing Tumors
0 10 20 30 40
0 10 20 30 40 V
0 10 20 30 40 Fraction #
FIG. 9. HPLC separation of GLP-1 immunoreactive pep- tides contained in control mouse large intestine ( A ) , a GLU- Tag-Y tumor at 4 weeks of age ( B ) , and a GLUTag-Y tumor a t 8 weeks of age (C). The peaks are identified as GLP-l(1-37) ( a ) and GLP-1(7-37) ( b ) based upon elution position of synthetic standards, as indicated by the arrowheads. In each case, 100 ng of total GLI was loaded onto the HPLC column.
providing additional evidence for an alteration in the process- ing of proglucagon with tumor growth.
DISCUSSION
Expression of the GLUTag transgene consistently induces neuroendocrine carcinoma of the large intestine in transgenic mice (13). The results of the present study have demonstrated that growth of the GLUTag-Y tumors was associated with a marked change in the post-translational processing of proglu- cagon. Normal mouse intestine contained predominantly gli- centin and oxyntomodulin, with only minor amounts of glu- cagon. In contrast, GLUTag-Y tumors produced progressively greater amounts of glucagon relative to the other GLI peptides during 4-8 weeks of tumor growth. Thus, although the tumors contained large amounts of proglucagon-derived peptides, the biosynthesis of glucagon was significantly increased relative to the other proglucagon-derived peptides with progressive tumor growth. Similarly, GLUTag-Y tumors also synthesized a greater proportion of GLP-1(7-37) and less GLP-l(1-37) compared to normal mouse intestine.
Cells derived from SV40 T antigen-transformed pancreatic islets process proglucagon in a manner consistent with the processing of the normal pancreatic A cell (21). In contrast, post-translational processing of proglucagon in other pan- creatic islet cell lines has been shown to be intermediate between that of the pancreatic A cell and the intestinal L cell (22, 23). The results of the present study suggest that the neoplastic intestinal L cell may also undergo a change in processing phenotype. Similar alterations in the post-trans- lational processing of proglucagon have also been described in patients with pancreatic endocrine tumors (24-26). These
findings provide further evidence for plasticity of prohormone post-translational processing in neoplastic cells.
With the cloning of several mammalian homologues of the yeast enzyme Kex-2, the enzymes involved in prohormone cleavage have recently begun to be elucidated (27-30). Two of these enzymes, PC1 and PC2, have been demonstrated to selectively cleave pro-opiomelanocortin in a fashion that re- sembles the pattern of processing seen in the anterior and intermediate pituitary in vivo (31, 32). It has not been estab- lished whether PC1 or PC2 is involved in the tissue-specific post-translational processing of proglucagon; however, mRNA transcripts for both enzymes have been detected in pancreatic islets (28, 29). The importance of the correct enzymatic ma- chinery for proglucagon processing has already been suggested by studies showing that a stably transformed non-endocrine cell line is unable to process proglucagon, whereas a non-islet endocrine cell line is capable of proglucagon cleavage, albeit in an aberrant manner. In contrast, transfection of non- glucagon-producing islet cells with a proglucagon cDNA re- sulted in an islet cell line that correctly processes proglucagon (33). It is therefore possible that alterations of the type or amount of cleavage enzyme produced by the transformed intestinal L cell during GLUTag-Y tumor growth might con- tribute to the concomitant changes observed in the post- translational processing of proglucagon in the intestinal tu- mors. One alternative explanation for the observed changes in prohormone processing is an involvement of unknown physiologic factors. Pro-opiomelanocortin processing has been demonstrated to be altered i n vivo by vasopressin (34) and by interleukin-2 (35). Similarly, alterations in cleavage of pro-opiomelanocortin seen i n vivo in the postnatal anterior pituitary have been duplicated under i n vitro conditions and have been reported to be due to the influence of glucocorti- coids (36). Further studies will be required to elucidate the mechanisms underlying the alterations in proglucagon proc- essing observed during growth of the intestinal GLUTag-Y tumors.
Growth of the GLUTag-Y tumors was associated with a marked reduction in the levels of proglucagon mRNA tran- scripts and proglucagon-derived peptides in the pancreas and, to a lesser extent, the small intestine. The molecular mecha- nisms underlying these inhibitory effects have not been elu- cidated; however, transcription of the proglucagon gene is known to be regulated by protein interactions with key regu- latory domains in the 5’-flanking sequence of the gene. In the pancreas, gene transcription is controlled by different proteins that bind to cis-acting regulatory elements in the first 300 base pairs of flanking sequence. These interactions are critical for islet- and A cell-specific gene expression (37, 38). In contrast, proglucagon gene expression in the intestine appears to be regulated by an upstream domain between -1300 and -2000 base pairs (13). Thus, SV40 T antigen expression may disrupt the normal regulatory mechanisms underlying panc- reatic proglucagon gene expression. Alternatively, as GLU- Tag-Y mice have been demonstrated to contain high circulat- ing concentrations of proglucagon-derived peptides, it is pos- sible that one of these peptides may be exerting inhibitory effects on the A and L cell. Additional evidence for this hypothesis is derived from a number of different studies that have reported acute suppression of glucagon secretion from the pancreatic A cell following exposure to various proglu- cagon-derived peptides, including glucagon (39), glicentin (40), and GLP-l(7-37) (9-11, 39). In contrast, neither glu- cagon nor GLP-l(7-37) have been found to modulate secre- tion of proglucagon-derived peptides by the intestinal L cell i n vitro (41). Further studies will be required to elucidate the

Intestinal Proglucagon-producing Tumors 20733
mechanism by which endogenous proglucagon gene expres- sion is inhibited in the GLUTag-Y mice.
As compared with the relative tumor content of glicentin, oxyntomodulin, and glucagon, the levels of glicentin in plasma from GLUTag-Y mice were found to be disproportionately increased relative to oxyntomodulin and glucagon. This dis- crepancy between the molecular forms of proglucagon-derived peptides contained in the tumor and those circulating in the plasma may relate to the metabolic degradation of the secreted peptides in the circulation. In rats, the half-life of circulating glucagon is 1.9-3.2 min (42,43), as compared with the longer half-lives of 6.4 min for oxyntomodulin (43) and 6.9 min for 9-kDa glucagon (42). Although the half-life of glicentin is not known, it might be expected to have a longer half-life than that of the peptides contained within its sequence (glucagon, oxyntomodulin, and the 9-kDa peptide). Indeed, the half-life of a crude small bowel GLI extract in dogs has been reported to be 15.9 min (44). Thus, glicentin levels might be predicted to be disproportionately elevated in plasma due to a relatively slower disappearance rate. Similar discrepancies between plasma and tumor proglucagon-derived peptides have been reported for patients with glucagonomas (24, 25, 45).
Interestingly, the proglucagon-derived peptides released by GLUTag-Y tumors into the circulation did not appear to exert significant effects on either insulin or blood glucose levels. GLP-1(7-37) has been found to be a potent stimulator of insulin biosynthesis and secretion (9-12), and yet panc- reatic and plasma insulin levels were not elevated in the GLUTag-Y mice. Similarly, although glucagon would be ex- pected to exert significant hyperglycemic activity, blood glu- cose levels were observed to fall in the 8-week-old mice. Some of these findings may relate to our observations that tumor development was associated with loss of body weight and eventually death (13), presumably due, at least in part, to intestinal obstruction and subsequently reduced nutrient in- take. As the effects of GLP-l(7-37) on the pancreatic B cell are dependent upon concomitant hyperglycemia (46, 47), the fall in blood glucose levels may have thereby precluded the insulinotropic effects of this peptide. Alternatively, the recep- tors for both glucagon and GLP-1(7-37) are known to undergo autologous desensitization (48,49), and thus, elevated plasma levels of these hormones may ultimately lead to some loss of biologic activity.
In conclusion, the post-translational processing of proglu- cagon in intestinal GLUTag-Y tumors undergoes a progres- sive change away from that of the normal intestinal L cell, resulting in increased glucagon synthesis relative to glicentin and oxyntomodulin. Tumor growth was associated with marked inhibition of endogenous proglucagon gene expression in pancreas and, to a lesser extent, small intestine. These findings suggest that the tumor proglucagon-derived peptides may participate in a novel negative feedback loop, inhibiting the expression of the endogenous mouse proglucagon gene in the pancreas and small intestine.
REFERENCES
1. Mojsov, S., Heinrich, G., Wilson, I. B., Ravazzola, M., Orci, L., and Habener, J. F. (1986) J. Biol. Chem. 2 6 1 , 11880-11889
2. Drucker, D. J., and Asa, S. (1988) J. Bid. Chem. 2 6 3 , 13475-13478 3. Drucker, D. J., and Brubaker, P. L. (1989) Proc. Natl. Acad. Sci. U. S. A.
4. Kervran, A,, Blache, P., and Bataille, D. (1987) Endocrinology 121 , 704- 86,3953-3957
5. Lui, E. Y., Asa, S. L., Drucker, D. J., Lee, Y. C., and Brubaker, P. L. (1990) 713
6. Patzelt, C., and Schiltz, E. (1984) Proc. Natl. Acad. Sci. U. S. A. 8 1 , 5007- Endocrinology 126,110-117
5011 7. Bataille, D., Blache, P., Mercier, F., Jarrousse, C., Kervran, A., Dufour, M.,
Mangeat, P., Dubrasquet, M., Mallat, A,, Lotersztajn, S., Pavoine, C., and Pecker, F. (1988) Ann. N. Y. Acad. Sci. 527,168-185
8. Mojsov, S., Kopczynski, M. G., and Habener, J. F. (1990) J. Biol. Chen.
9. Mojsov, S., Weir, G. C., and Habener, J. F. (1987) J. Clin. Inuest. 79 , 616- 265,8001-8008
619 10. Kreymann, B., Ghatei, M. A,, Williams, G., and Bloom, S. R. (1987) Lancet
5 .1 mn-1x04 11. Holst, J. J., Orskov, C., Vagn Nielsen, O., and Schwartz, T. W. (1987)
12. Fehmann, H.-C., and Habener, J. F. (1992) Endocrinology 130 , 159-166 13. Lee, Y. C., Asa, S. L., and Drucker, D. J. (1992) J. Bid. Chem. 267,10705-
- 7 "" ""
FEBS Lett. 211 , 169-174
1 nmx 14. Bibaker , P. L., and Vranic, M. (1987) Endocrinology 120,1976-1985 15. Brubaker, P. L., So, D. C. Y., and Drucker, D. J. (1989) Endocrinology 124 ,
16. Brubaker, P. L.(1988) Endocrinology 123,220-226 17. Heding, L. G.(1971) Diabetologia 7, 10-19 18. Faloona, G. R., and Unger, R. H. (1974) in Methods of Hormone Radio-
immunoassay (Jaffe, B. M., and Behrman, H. R., eds) pp. 317-330,
19. Drucker. D. J.. CamDos. R.. Revnolds. R.. Stobie. K.. and Brubaker. P. L. Academic Press, New York
3003-3009
20.
21.
22.
23.
(1991) Endocrinohgy'l28, 354-400' '
Bwl. Chem. 193,265-275
Hanahan, D. (1990) Diabetes 39,406-414
nology 119,2833-2839
, ,
Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J.
Powers, A. C., Efrat, S., Mojsov, S., Spector, D., Hahener, J. F., and
Philippe, J., Mojsov, S., Drucker, D. J., and Habener, J. F. (1986) Endocri-
Drucker, D. J., Philippe, J., and Mojsov, S. (1988) Endocrinology 123 , 1 Xfil -1 Xfi7
24. Fukushima, H., Yamaguchi, K., and Uzawa, H. (1981) Endocrinol. Jpn. 28 ,
25. Holst, J. J.(1983) Diabetologia 2 4 , 359-365 26. Alarcbn. C.. Kervran. A,. Bataille. D.. and Valverde. I. (1991) Horm. Metab.
_"_ 575-582
, . Res. 23,290-292
, . . . .
27. Smeekens, S. P., and Steiner, D. F. (1990) J. Biol. Chem. 265,2997-3000 28. Seidah, N. G., Gaspar, L., Mion, P., Marcinkiewicz, M., Mbikay, M., and
29. Smeekens, S. P., Avruch, A. S., LaMendola, J., Chan, S. J., and Steiner, D. Chretien, M. (1990) DNA Cell Biol. 9 , 415-424
30. Seidah, N. G., Marcinkiewicz, M., Benjannet, S., Gaspar, L., Beaubien, G., F. (1991) Proc. Natl. Acad. Sci. U. S. A. 8 8 , 340-344
Mattei,,M. G., Lazure, C., Mbikay, M., and Chretien, M. (1991) Mol. Endocrtnol. 5 , 111-122
31. Benjannet, S., Rondeau, N., Day, R., Chretien, M., and Seidah, N. G. (1991) Proc. Natl. Acad. Sci. U. S. A. 8 8 , 3564-3568
32. Thomas, L., Leduc, R., Thorne, B. A., Smeekens, S. P., Steiner, D. F., and Thomas, G. (1991) Proc. Natl. Acad. Sci. U. S. A. 8 8 , 5297-5301
33. Drucker, D. J., Mojsov, S., and Habener, J. F. (1986) J. Biol. Chem. 261 , qfillli-CIfiA.?
34. Canny, B. J., Smith, A. I., Clements, J. A., and Funder, J. W. (1988)
35. Zakarian, S., Eleazar, M. S., and Silvers, W. K. (1989) Nature 339 , 553-
_l-l
Neuroendocrinology 4 8 , 603-610
36. Noel, G., and Mains, R. E. (1991) Endocrinology 1 2 9 , 1317-1325 37. Drucker, D. J., Philippe, J., Jepeal, L., and Habener, J. F. (1987) J. Biol.
38. PhiliDDe. J.. Drucker. D. J.. KneDel. W.. Jeoeal. L.. Misulovin. Z.. and
556
Chem. 262,15659-15665
H&ener,J. F. (1988) Md'Cell . Bioi. 8,'487?-4888 '
and Yamashita, K. (1989) Endocrinology 124,1768-1773
I , ~~
39. Kawai, K., Suzuki, S., Obashi, S., Mukai, H., Ohmori, H., Murayama, Y.,
40. Ohneda, A., Kobayashi, T., and Nihei, J. (1986) Dtabetologia 29 , 397-401 41. Brubaker. P. L.(1991) Endocrinoloev 128.3175-3182 42. O'Connor, K. J., and'lazarus, N. R: (1976) Biochem. J. 156,265-277 43. Kervran, A., Dubrasquet, M., Blache, P., Martinez, J., and Bataille, D. 44. Tanaka, R., Matsuyama, T., Shima, K., Sawazaki, N., Tarui, S., and
45. Drucker, D. J., Asa, S., Silverherg, J., and Brubaker, P. L. (1990) Cancer
46. Schmidt, W. E., Siege], E. G., and Creutzfeldt, W. (1985) Diabetologia 2 8 ,
47. Komatsu, R., Matsuyama, T., Namba, M., Watanabe, N., Itoh, H., Kono,
48. Murphy, G. J., Hruhy, V. J., Trived, D., Wakelam, J. O., and Houslay, M.
49. Fehmann, H. C., and Habener, J. F. (1991) Endocrinology 128,2880-2888
(1990) Regul. Pept. 31,41-52
Kumahara, Y. (1979) Endocrinol. Jpn. 26,59-63
6 5 , 1762-1770
704-707
N., and Tarui, S. (1989) Diabetes 3 8 , 902-905
D. (1987) Biochem. J. 2 4 3 , 39-46





























