Embed Size (px)
Citation preview

Alteration in expression of smooth muscle a-actin associated with
transformation of rat 3Y1 cells
MITSUSE OKAMOTO-INOUE1, SHUNICHIRO TANIGUCHI1-*, HIROYUKI SADANO1,
TOYOKAZU KAWANO1, GENKI KIMUEA2, GIULIO GABBIANI3 and TSUNEO BABA1
^Department of Experimental Cell Research, and ^Department of Virology, Medical Institute of Bioregulation, Kyushu University,Fukuoka 812, Japan3Department of Pathology, Uniuersity of Geneva, 1211 Geneva 4, Switzerland
* Author for correspondence at Department of Experimental Cell Research, Medical Institute of Bioregulation,Kyushu University 69, Maidashi 3-1-1, Higashi-ku, Pukuoka 812, Japan
Summary
Expression of actin was examined in a cultured ratembryonic cell line 3Y1 and transformed cell linesthat originated from 3Y1. An a-actin in addition tocytoplasmic ft- and y-actins was detected in 3Y1 bytwo-dimensional gel electrophoresis. This a-actinwas hardly detected at all in the transformantsinduced by Rous sarcoma virus, v-H-ras oncogene oradenovirus type 12, while the a-actin was retained inthe transformed cell lines induced by JV-methyl-JV-nitro-JV-nitrosoguanidine or in SV40, which are celllines of relatively low malignancy. Western andNorthern blot analyses established that this a-actin
was a smooth muscle a-isofonn. An immunofluor-escence study revealed that smooth muscle a-actin in3Y1 cells is present in stress fibers. Thus, smoothmuscle a-actin is also a component of actin stressfibers, as /J- and y-actins are in 3Y1 cells. An alter-ation in the expression of this actin isoform may berelated to phenotypical changes accompanyingtransformation.
Key words: transformation, smooth muscle a-actin, 3Y1fibroblasts, actin stress fibers.
Introduction
Actin is a ubiquitous cytoskeletal protein in eukaryoticcells and highly conserved in evolution, probably involvedin the regulation of cellular motility, morphology andattachment to neighboring cells or to the matrix. To date,six isoforms of actins have been identified in mammaliancells (Vandekerckhove and Weber, 1978). It has beenwidely accepted that only /S- and y-actins are present innon-muscle cells in culture (Garrels and Gibson, 1976).However, cr-like actin has recently been detected in non-muscle cultured cells, such as chick embryo fibroblasts(Eubenstein and Spudlich, 1977; Witt et al. 1983), Rat-2and NTH 3T3 cell lines (Leavitt et al. 1985), and rat REF 52cells (Franza and Garrels, 1984). This olike actin hasbeennoted as a transformation-sensitive marker. This isoformof actin was not present in spontaneously transformedcells (Leavitt et al. 1985) and in transformed cells inducedby tumor viruses (Witt etal. 1983; Franza and Garrels,1984). Direct identification of the a-like actin, however,remains to be made, and its intracellular localization andfunctions are still unknown.
In this study, we noted that an olike actin_c*h also bedetected in a rat 3Y1 fibroblastic cell line and its ex-pression is altered in association with transformed and/ormalignant properties. Furthermore, we identified the a-like actin as a smooth muscle isoform and a component ofactin stress fibers.Journal of Cell Science 96, 631-637 (1990)Printed in Great Britain © The Company of Biologists Limited 1990
Materials and methods
Cell cultureA Fisher rat embryo fibroblastic cell line, 3Y1-B clone 1-6(referred to as 3Y1) (Kimura et al. 1976), and its transformed celllines Adl2-3Y1-W5 (Ad) (Okuda et al. 1983), SV-3Y1-66 (SV), SR-3Y1-1 (SR) (Zaitsu et al. 1988), HR-3Y1-3 (HR) (Shimura et al.1988), NG-3Y1-T15L (NG) (Ohno et al. 1987; Zaitsu et al. 1988)and /bs-SR-3Yl-202 (/bs-SR) (Taniguchi etal. 19866; Kawanoet al. 1987) were described previously (see Table 1 for transform-ing agents). Cells were cultured in Dulbecco's modified Eagle'smedium (GIBCO Laboratories, Grand Island, NY) supplementedwith 10% fetal bovine serum (GIBCO), 100 units ml"1 penicillinand 80/igml"1 kanamycin in moist air containing 10% CO2 at37 °C.
Preparation of proteins for two-dimensionalpolyacrylamide gel electrophoresisThe Triton-soluble fraction and the cytoskeletal fraction of cellu-lar proteins were prepared according to the method of Leavitt andKakunaga (1980). Subconfluent cells in a 60 mm Petri dish wererinsed with three changes of cold phosphate-buffered saline(PBS), followed by the addition of 0.5 ml of cold extraction solutionconsisting of 1% Triton X-100 (v/v), 10 mM Tris-HCl, pH7.8,10 mw MgCl2, 30 mM KC1, 10 mM 2-mercaptoethanol. Solublecellular proteins were extracted during incubation for 5min at4°C. The extraction solution was then collected and centrifuged at10 000 g to remove any cellular debris.
Urea up to 9 M was added to the supernatant containing solublecellular proteins for two-dimensional gel electrophoresis. A cyto-skeletal fraction was prepared from the Triton-insoluble residue
631

Table 1. Alteration of expression of a-actin intransformed cell lines derived from rat 3Y1 cells
Cell line Transforming agent
3Y1SVAdSRHR/bj-SRNG
Simian virus 40Adenovirus type 12Rous avian sarcoma virusv-H-ros oncogeneRous avian sarcoma virus+v-/bs/V-methyl-W-nitro-iV-nitrosoguanidine
that remained on the Petri dish after the extraction of Triton-soluble proteins. The insoluble residue was collected from the dishby scraping with a rubber policeman and suspended in 1 ml of coldextraction solution, with the reduced concentration of Triton X-100 to 0.1 %. The cytoskeletal fraction was pelleted by centrifu-gation and dissolved in a lysis buffer described by OTarrell(1975). Total cellular proteins were prepared from cells in thesubconfluent stage. Cells were harvested with a rubber policemaninto a 1.5 ml microcentrifuge tube, rinsed with cold PBS andcentrifuged at 10 000 ̂ a t 4°C. The pellet was re-suspended in 1 mlof PBS, 50 /d of which was separated to measure the proteinconcentration with a BCA protein assay reagent (Pierce ChemicalCo. Rockford, IL) with bovine serum albumin (BSA) as standard.The cells were pelleted by centrifugation, then treated with 1 mlof ice-cold acetone, followed by centrifugation. After extractingthe cells in acetone three times, the pellet was dried, solubilizedeither in SDS sample buffer containing 80 mM Tris-HCl (pH 6.8),1 % SDS, 10 % glycerol and 1 °h Bromophenol Blue for one-dimensional SDS-PAGE or in OTarrell's lysis buffer for two-dimensional gel electrophoresis. Two-dimensional gel electro-phoresis was performed according to CFarrell (1975). The separ-ating gels in SDS-PAGE and the second dimension of electro-phoresis contained 10% acrylamide. After electrophoresis, gelswere used for Western blotting or stained with Coomassie Blue.Since the stain on the gel was too faint for analysis of solublecellular proteins, silver staining was subsequently carried outwith a silver staining kit (Wako Pure Chemical Industries, Ltd,Osaka, Japan).
Western blotting and immunostainingGels after two-dimensional polyacrylamide gel electrophoresis orSDS—PAGE were trimmed and equilibrated with 25 mM Tris,192 mM glycine, 20 % (v/v) methanol for 30 min at room tempera-ture. Proteins in the gels were electroblotted to nitrocellulose at0.8 mA cm"2 gel, for 1,5 h (Kyhse-Andersen, 1984) by Semi DryElectroblotter (Sartorius, Osaka, Japan). After the blotting, theremaining proteins in the gels were stained with silver to ensurethat the blotted area contained actin isoforms. The nitrocellulosesheets were gently shaken in 50 mM Tris-HCl, pH7.5, 150 mMNaCl, 0.1 % (v/v) Tween 20 (TBS-Tween 20), containing 1 % BSA,for 1 h. After a brief rinse in TBS-Tween 20, the nitrocellulosesheets were immersed in either anti-actin monoclonal antibody,which reacted with all actin isoforms (Amersham, Amersham,UK), or monoclonal antibody against smooth muscle o--actin(anti-asm) (Sigma Chemical Co., St Louis, MO) 1:1000 diluted in0.1% BSA-TBS-Tween 20 for 2h at room temperature, thenrinsed in three changes of TBS-Tween 20. Peroridase-conjugatedrabbit anti-mouse immunoglobulin (Organon Teknika Corp. WestChester, PA) 1:500 diluted in 0.1% BSA-TBS-Tween 20 wasreacted for l h as a secondary antibody. The nitrocellulose wasrinsed in four changes of TBS-Tween 20 and two changes ofdistilled water, then developed in 50 mM Tris-HCl, pH7.5,O^mgml"1 4-chloro-l-naphthol and 0.01% (v/v) H2O2. Thereaction was stopped by rinsing the nitrocellulose in distilledwater.
RNA extraction and Northern blot hybridizationTotal cellular RNA was extracted by the guanidinium/cesiumchloride method (Maniatis etal. 1982). A DNA probe pRAoaA-
3'UT that is specific for smooth muscle a-actin (Clowes et al. 1988)and a probe specific for cardiac muscle a-actin (Hamada et al.1982) were radiolabelled with [o^PJdCTP using a DNA labellingkit (Nippon Gene, Tokyo, Japan). Northern blot analysis wasperformed according to the method of Leavitt et al. (1985) withslight modifications. Briefly, 5 //g of total RNA per one lane waselectrophoresed on 1% agarose gels containing 2.2 M formal-dehyde and transferred to nitrocellulose filters. The filters werehybridized with a radiolabelled DNA probe in 5xSSC (SSC is0.15M NaCl, 0 .015M sodium citrate, pH7.0), 5xDenhardt's sol-ution (Maniatis et al. 1982) 50 mM sodium phosphate buffer(pH6.5), 50% formamide and 250/igml"1 salmon sperm DNA at42°C, followed by sequential washing in 5xSSC, 0.2 % sodium N-lauroyl sarcosinate (SDSa) at 45°C, two changes of O.lxSSC,0.2% SDSa at 50°C, and 0.05xSSC, 0.2% SDSa at 55°C. Theniters were air-dried and exposed to Kodak XAR-5 film. Afterovernight exposure, the film was developed.
Immunofluorescence studyIndirect immunofluorescence staining of cytoskeletons was car-ried out basically as described by Leavitt et al. (1982a). The celllines were allowed to attach themselves to glass coverslips andwere cultured at least overnight. The cells were rinsed brieflywith divalent cation-free PBS, extracted for lmin at roomtemperature in aqueous 1 % (v/v) Triton X-100 containing 10 mMTris-HCl, pH 7.9, 30 mM KC1,10 mM MgCl2 and 10 mM 2-mercap-toethanol, then fixed in acetone for 10 min at room temperature.As for fixation of total proteins, 3Y1 cells were fixed in ice-coldmethanol for 20 min without pre-extraction. After air-drying,remaining cytoskeletons or whole cells on the coverslips wereincubated with 1:1000 dilutions of anti-a-sm or control mouseascites fluid (Cederlane Laboratories Ltd, Ontario, Canada) inPBS containing 0.1 % BSA at 4°C overnight. The coverslips wererinsed in three changes of PBS then incubated with FITC-conjugated rabbit anti-mouse IgG2a (ICN ImmunoBiologicals,Lusie, IL) diluted 1:1000. The coverslips were rinsed as describedabove and mounted in a 9:1 mixture of glycerol and PBScontaining 1 % 2-mercaptoethanol. The fluorescence of the cellswas examined with an Olympus microscope BHS-RFK with BH2-PC, DPlan Apo 20xUV and PCSplan 20xPL phase-contrast optics,and photographed on Kodak Tri-X 400 film.
Double immunostaining of the actin isoforms was performed asfollows. The cytoskeletons prepared on glass coverslips as de-scribed above were incubated with anti-cram (1:250) for 30 minand then with anti-actin monoclonal antibody (1:250), whichreacted with all actin isoforms, for 30 min. The coverslips werethen rinsed, followed by incubation with a mixture of FITC-conjugated rabbit anti-mouse IgG^a d: 250) and rhodamine-conjugated goat anti-mouse IgM (Tago, Inc., Burlingame, CA)(1:250). After rinsing, the coverslips were mounted and examinedwith a Zeiss Axiphot microscope.
Results
Detection of a-actin in 3Y1 cells by two-dimensionalpolyacrylamide gel electrophoresisTwo-dimensional gel electrophoretic patterns of actinextracted from 3Y1 cells were shown in Fig. 1. /S- and y-actins were easily recognized as doublets of proteinslocated in the area of Mr 43 000 and pi 5.2-5.3. A moreacidic and smaller spot of protein than /3-actin wasdetected both in the Triton-soluble fraction (Fig. 1A) andin the cytoskeletal fraction (Fig. IB). The pattern of thesespots was consistent with a-, (3- and y-actins reported byother groups (Garrels and Gibson, 1976; Witt et al. 1983;Franza and Garrets, 1984; Leavitt et al. 1985; Skalli et al.1986).
Identification of the a-actin as smooth muscle a-isoformand its alteration in expression in transformed cell linesThe three actin isoforms in 3Y1 cells were further ident-
632 M. Okamoto-Inoue et al.

IEF- 1
- -67
iQ: 7
2A B
1 ia is
-43
H
- 4 3
B
Fig. 1. A two-dimensional electrophoretic pattern of actinisoforms extracted from rat 3Y1 fibroblastic cells. Proteins inTriton-soluble fraction (A) and cytoskeletal fraction (B) wereprepared from cultured 3Y1 cells sequentially and applied totwo-dimensional gel electrophoresis as described in Materialsand methods. Silver-stained (A) or Coomassie Blue-stained gels(B). a, cr-actin; ji, /5-actin; y, y-actin. The positions of Mrmarkers are indicated (xlO~3) on the right, v, vimentin.
ified by immunoblotting with anti-actin antibody thatreacts with all isoforms of actin (Fig. 2B). When anti-asmwas used in immunoblotting, only the most acidic spot wasstained but /?- or y-actins were not (Fig. 2C). Among thetransformed cell lines derived from 3Y1 (Fig. 2, D-R), NGcells showed a large spot, and SV cells contained a smallbut significant amount of smooth muscle a-actin as com-pared with 3Y1 cells. Though the a-actin in the SV cellswas either not, or only scarcely, visible in Coomassie Blue-stained gels and immunoblotting with anti-actin antibodycommon to all actin isoforms, a distinct spot was detectedby anti-asm. Thus, immunoblotting with anti-cvsm wasthe most sensitive of these three methods for detecting thea-actin. On the other hand, smooth muscle a-actin was notdetected in the rest of the transformants, such as HR, SRand Ad.
As the specific reaction of anti-asm with the a-actin in3Y1 cells was confirmed (Fig. 2), total cellular proteinswere separated by SDS-PAGE, and then the amount ofsmooth muscle cv-actin was compared among the cell linesby Western blot analysis (Fig. 3). The stained proteinsremaining on the gel after the blotting indicated thatalmost the same amount of protein was applied in eachlane (Fig. 3A). Smooth muscle a--actin was detected in3Y1, NG and SV, but not in SR, HR, /bs-SR or Ad. Theseresults are summarized in Table 1.
Northern blot analysis of smooth muscle a-actinDetection of RNA transcripts specific for smooth muscle a-actin by Northern blot analysis is shown in Fig. 4. RNA
M
K
N
Q
O
R
Fig. 2. Identification of n--actin and alteration in its expressionin transformed cell lines originated from 3Y1. Total cellularproteins from cultured cells were separated by two-dimensionalgel electrophoresis, and then stained with Coomassie Blue (theleft row: A, D, G, J, M, P) or electroblotted. Western blots werestained either with anti-actin antibody that reacts with allactin isoforms (the middle row: B, E, H, K, N, Q) or with anti-asm (the right row: C, F, I, L, O, R). Parental 3Y1 (A, B, C);NG (D, E, F); SV (G, H, I); SR (J, K, L); HR (M, N, 0) and Ad(P, Q, R).
bands of smooth muscle cv-actin at 1.7 kb (103 bases) weredetected in 3Y1, NG and SV cells, and the density wasconsistent with the protein levels detected by Western blotanalysis (Fig. 4A). The amount of intact RNA applied wasalmost comparable, as shown by direct staining of 28 S and18 S rRNA (Fig. 4B). We also carried out a Northern blotanalysis with a specific probe for cardiac cv-actin but noband was detected in any cell line examined (Fig. 4C).
Immunofluorescence study of smooth muscle a-actin in3Y1 and transformed cell linesSince cv-actin was detected electrophoretically in the cyto-skeletal fraction, it was speculated that a-actin is incor-porated into microfilaments as well as /3- and y-actins. Toexamine the distribution of a'-actin, immunofluorescencestaining was carried out in whole cell preparations of 3Y1cells fixed in methanol (Fig. 5A) and acetone-fixed cyto-skeletons of Triton X-100-extracted 3Y1 cells (Fig. 5B).Stress fibers containing smooth muscle a-actin were visu-
Smooth muscle a-actin in 3Y1 cells 633

1 2 3 4 5 6 7
M1 2 3 4 5 6 7
* -94
-43-30-20
-94-43-30-20
B
Fig. 3. SDS-PAGE and Western blot analysis on total proteinextracts from cell lines using antd-ffsm. The gel was stainedafter blotting (A). Blots were immunostained with anti-ffsm(B). Lane 1, 3Y1; lane 2, NG; lane 3, SV; lane 4, fos-SR; lane 5,HR; lane 6, SR; and lane 7, Ad. Forty /ig protein waselectrophoresed in each lane. Smooth muscle o^actin(arrowhead) was detected in 3Y1. NG and SV. The positions ofMr markers are indicated (xlO ) on the right.
alized in cytoplasm of 3Y1 cells prepared by both methods.Faint, ground glass-like staining in the cytoplasm inwhole cell preparations appeared to correspond to soluble,unpolymerized a--actin. Although 3Y1 is a cloned cell line,the development of cr-actm stress fibers was uneven amongthe cells; some cells were intensely stained, while othercells showed little staining. Such heterogeneity of actinstress fibers was not observed when the cells were stainedwith anti-actin monoclonal antibody, which binds to allisoforms of actin. Staining in the cytoplasmic area of 3Y1cells with anti-o-sm was much the same as seen withmonoclonal antibody reacting with all actin isoforms. Inthe perinuclear area, however, a slight difference instaining between those two antibodies was detected. Boththe significance and cause of this phenomenon remain tobe examined. The intensity of the fluorescence wasstronger when stained with the anti-actin monoclonalantibody than when stained with anti-a-sm. This mayreflect the difference in the amount of the isoforms. Thesefindings indicate that there is no selectivity among theisoforms in the incorporation into most of the stress fibersin the cytoplasm, except for the perinuclear area.
Among the transformed cell lines, the NG cells showedabundant and minute fibers throughout the cytoplasm(Fig. 5D), but large bundles of a--actin were hardly everorganized in these cells. There was some variation ofexpression among the cells in NG cells, as seen in the 3Y1cells. Stress fibers containing a--actin were also shown insome of the SV cells (Fig. 5E), although the number of thecells expressing o<-actin was fewer than those seen in NGor 3Y1. In SR cells, ff-actin was not detected (Fig. 5F).These results are compatible with the results of theWestern and Northern blot analyses.
-1.7kb
Fig. 4. Northern blot analysis on expression of smooth musclea--actin. Autoradiogram of Northern blots of total RNA(5/ig/lane) from 3Y1 (lane 1) and its tranformants (lanes 2-7,the same order as in Fig. 3) after hybridization with 32P-labelled pRAoaA-3'UT (A). The same amount of each samplewas subjected to electrophoresis in another agarose gel followedby staining with O^/igml"1 ethidium bromide (B). The blot inC was hybridized with a specific probe for cardiac muscle a-actin.
Discussion
In this study we report that avactin, in addition to /S- and y-cytoplasmic actins, was expressed in rat 3Y1 cells. Theexpression of this a^actin was suppressed or decreased intransformed cell lines such as SV, SR, HR and Ad. Thesame phenomenon has been reported in chick embryofibroblasts (Witt etal. 1983), rat REF52 (Franza andGarrels, 1984), mouse 3T3 and Rat-2 cells (Leavitt et al.1986); the <*-actin found in these systems was suggested tobe a smooth muscle-specific isoform on the basis of mo-bility in isoelectric focusing (Franza and Garrels, 1984)and smaller length of mRNA (1.7 kb) than that of /S- or y-isoforms (2.2kb) (Leavitt etal. 1985). However, a directidentification of this transformation-sensitive actin hasnot yet been made. In this study we demonstrated that theoactin in 3Y1 cells is a smooth muscle isoform by Western
634 M. Okamoto-Inoue et al.
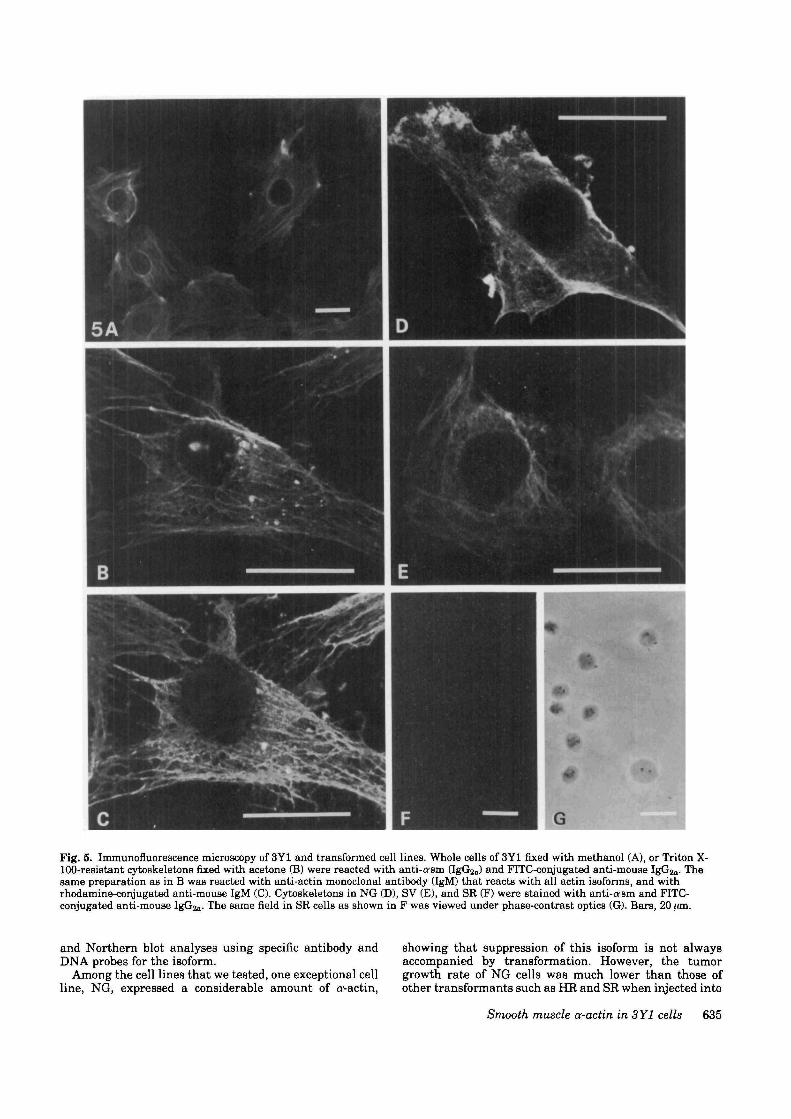
Fig. 5. Immunofluorescence microscopy of 3Y1 and transformed cell lines. Whole cells of 3Y1 fixed with methanol (A), or Triton X-100-resistant cytoskeletons fixed with acetone (B) were reacted with anti-cram (IgGua) and FITC-conjugated anti-mouse IgGsjn- Thesame preparation as in B was reacted with anti-actin monoclonal antibody (IgM) that reacts with all actin isoforms, and withrhodamine-conjugated anti-mouse IgM (C). Cytoskeletons in NG (D), SV (E), and SR (F) were stained with anti-asm and FITC-conjugated anti-mouse IgG2a- The same field in SR cells as shown in F was viewed under phase-contrast optics (G). Bars, 20 ;an.
and Northern blot analyses using specific antibody andDNA probes for the isoform.
Among the cell lines that we tested, one exceptional cellline, NG, expressed a considerable amount of oactin,
showing that suppression of this isoform is not alwaysaccompanied by transformation. However, the tumorgrowth rate of NG cells was much lower than those ofother transformants such as HR and SR when injected into
Smooth muscle a-actin in 3Y1 cells 635

syngeneic F344 rats, and the NG tumor cells did notmetastasize (data not shown), suggesting that NG cellsbehave like benign tumor cells in vivo. These facts indicatethat a'-actin is not the only protein responsible for thebiological behavior of the transformed cells. Other el-ements including actin-associated proteins may alsochange throughout the process of transformation. Poorlydeveloped bundles of a'-actin seen in NG cells suggest thatthe organization of actin fibers may be impaired byqualitative and/or quantitative alteration of actin-associ-ated proteins. Thus, irrespective of a--actin expression, thecytoskeletal system may not function properly, leadingthese cells to lose their normal phenotypes. Whether theabundant expression of smooth muscle a'-actin is respon-sible for the low-grade malignancy of NG cells remains tobe examined. Another transformant expressing a'-actinwas SV, though the amount was less than that in 3Y1. Aprevious study showed that SV cells do not grow in softagar, i.e. they retain anchorage dependency (Zaitsu et al.1988). Therefore, SV cells seem also to be of a lower gradeof malignancy than the rest of the transformed cell lines.
oActin was suppressed in clone HE-3Y1-3 (Figs 2, 3 and4), while a small amount was detected in another indepen-dent transformant induced by v-H-ros oncogene and thetumor growth rate of the latter clone in animals was lowerthan HR-3Y1-3 (data not shown). Thus, a shutdown ordecrease in the expression of a<-actin may be related to cellmalignancy in the 3Y1 system. In mouse B16 melanomacell lines of denned metastatic potential, we noted asimilar phenomenon: the suppression and/or decrease inexpression of a newly found /J-actin was inversely relatedto the metastatic potential (Taniguchi et al. 1986a, 1989;Sadano et al. 1988).
Since alterations in other proteins have also beendescribed in transformed cells (Leavitt et al. 19826; Matsu-mura et al. 1983; Franza and Garrels, 1984), we examinedother proteins on two-dimensional gels. We found thattropomyosin-1 was suppressed in SR and Ad, and tropo-myosin-5 seemed to be increased in SR (data not shown).These findings were consistent with a report by Matsu-mura et al. (1983).
Although /S-actin is widely used as a standardization inNorthern blot analysis, we used staining intensity of 18 Sand 28 S rRNA instead of/3-actin because the expression of/S- or y-actin is sometimes altered due to transformation.We noted that the j3-actin was transcribed in a largeramount in NG than in SR and Ad cells (data not shown).There is other evidence of an alteration in actin synthesisconcomitant with the expression of oncogenes after stimu-lation by serum (Greenberg and Ziff, 1984; Lau andNathans, 1985; Masibay et al. 1988), or by growth factors(Elder et al. 1984).
The specific function of o--actin found in 3Y1 cellsremains to be clarified. There is a report on functionalisoactin sorting in the motile cytoplasm of microvascularpericytes (DeNofrio et al. 1989). In our system of 3Y1, a'-actin was incorporated into actin stress fibers as well asother isoforms (Fig. 5). The heterogeneous staining of cellsafter immunostaining with anti-a-sm indicates that theexpression of a'-actin alters with cellular conditions suchas cell-cycle phases and attachment. Further experimentsincluding gene transfer experiments and biochemicalanalysis of interactions of a--actin with other actin-associ-ated proteins are under way. These experiments will helpto clarify the physiological functions of a'-actin and eluci-date whether the suppression of this o--actin is responsiblefor malignant properties of the cells. Thus, a--actin can
possibly become a clue in elucidating the relation betweentransformation and/or malignancy of cells and the alter-ation of cytoskeletal proteins.
We acknowledge the helpful comments of Mr B. T. Quinn andthe late Dr T. Kakunaga and thank Ms R. Morohashi and Ms K.Fukuda for technical assistance.
This work was supported in part by a grant-in-aid for CancerResearch from the Ministry of Education, Science and Culture,and from the Ministry of Health and Welfare, Japan, and by theSwiss National Science Foundation, Grant no. 3, 108-0.88.
References
CLOWES, A. W., CLOWES, M. M., KOCHKB, 0 , ROPHAZ, P., CHAPONNIER, C.AND GABBIANI, G. (1988). Arterial smooth muscle cells in vivo:relationship between actin isoform expression and mitogenesis andtheir modulation by heparin. J. Cell Biol. 107, 1939-1946.
DENOFTUO, D., HOOCK, T. C. AND HERMAN, I. M. (1989). Functionalsorting of actin isoforms in microvascular pericytes. J. Cell Biol. 109,191-202.
ELDER, P. K., SCHMIDT, L. J., ONO, T. AND GETZ, M. J. (1984). Specificstimulation of actin gene transcription by epidermal growth factor andcycloheximide Proc. natn. Acad. Sci. U.S.A. 81, 7476-7480.
FRANZA, B. R. JR AND GARRELS, J. I. (1984). Transformation-sensitiveproteins of REF52 cells detected by computer-analyzed two-dimensional gel electrophoreais. Cancer Cells 1, 137-146.
GARRELS, J. I. AND GIBSON, W. (1976). Identification and characterizationof multiple forms of actin Cell 9, 793-805.
GREENBERO, M. E. AND ZIFF, E. B. (1984). Stimulation of 3T3 cellsinduces transcription of the c-fos proto-oncogene. Nature 311, 433—438.
HAMADA, H., PETRINO, M. G. AND KAKUNAGA, T. (1982). Molecularstructure and evolutionary origin of human cardiac muscle actin gene.Proc. natn. Acad. Sci. U.S.A. 79, 5901-5905
KAWANO, T., TANIGUCHI, S., NAKAMATSU, K., SADANO, H. AND BABA, T.(1987). Malignant progression of a transformed rat cell line by transferof the v-fos oncogene. Biochem. biophys. Res. Comnwn. 149, 173-179.
KIMURA, G., ITAGAKI, A. AND SUMMERS, J. (1975). Rat cell line 3Y1 andits virogenic polyoma- and SV40-transformed derivatives. Int.J. Cancer 15, 694-706.
KYHSB-ANDERSEN, J. (1984). Electroblotting of multiple gels: a simpleapparatus without buffer tank for rapid transfer of proteins frompolyacrylamide to nitrocellulose. J. biochem. biophya. Meth. 10,203-209.
LAU, L F. AND NATHANS, D. (1985). Identification of a set of genesexpressed during the Go/Gi transition of cultured mouse cells. EMBOJ. 4, 3145-3151.
LEAVITT, J., BUSHAR, G., KAKUNAGA, T , HAMADA, H., HIRAKAWA, T.,GOLDMAN, D. AND MERRIL, C. (1982a). Variations in expression ofmutant /J-actin accompanying incremental increases in humanfibroblast tumorigemcity. Cell 28, 259-268.
LBAVITT, J., GOLDMAN, D., MERRIL, C. AND KAKUNAGA, T. (19826).Changes in gene expression accompanying chemically-inducedmalignant transformation of human fibroblasts. Carcinogenesis 3,61-70.
LEAVTTT, J., GUNNING, P., KEDES, L. AND JARTWALLA, R. (1985). Smoothmuscle a'-actin is a transformation-sensitive marker for mouse NIH3T3 and Rat-2 cells. Nature 316, 840-842.
LKAVITT, J. AND KAKUNAGA, T. (1980). Expression of a variant form ofactin and additional polypeptide changes following chemical-inducedm vitro neoplastic transformation of human flbroblasts. J. biol. Chem.265,1650-1661.
MANIATIS, T., FRTTSCH, E. F. AND SAMBROOK, J. (1982). Isolation of totalcellular RNA. In Molecular Cloning. A Laboratory Manual, pp.194-196. New York: Cold Spring Harbor Laboratory Press.
MASIBAY, A. S., QASBA, P. K., SENGUPTA, D. N., DAMEWOOD, G. P. ANDSTREEVALSAN, T. (1988). Cell-cycle-specific and serum-dependentexpression of gamma-actin mRNA in Swiss mouse 3T3 cells. Molec.cell. Biol. 8, 2288-2294.
MATSUMURA, F., LIN, J. J.-C, YAMASHIRO-MATSUMURA, S-, THOMAS, G. P.AND TOPP, W. C. (1983). Differential expression of tropomyosin formsin the micro-filaments isolated from normal and transformed ratcultured cells. J. biol. Chem. 258, 13 954-13964.
OTARRELL, P. H. (1976). High resolution two-dimensional electrophoresisof proteins. J. biol. Chem 260, 4007-4021.
OHNO, K., ZAITSU, H. AND KIMURA, G. (1987). Maintenance ofpostconfluence stationary cell density by transient increase anddecrease in cell number upon medium renewals in rat 3Y1 fibroblasts.Fukuoka Acta Med. 78, 569-577.
OKUDA, A., KAJIWARA, Y. AND KIMURA, G. (1983). Difference in growth
636 M. Okamoto-Inoue et al.

factor requirements of rat 3Y1 cells among growth in mass culture,clonal growth in low density culture, and stimulation to enter S phasein resting culture. In Vitro 19, 376-384.
RUBBNSTEIN, P. A. AND SPUDICH, J. A. (1977). Actin microheterogeneityin chick embryo fibroblasts. Proc. natn. Acad. Sci. U.S-A. 74, 120-123.
SADANO, H., TANIOUCHI, S., KAKUNAQA, T. AND BABA, T. (1988). cDNAcloning and sequence of a new type of actin in mouse B16 melanoma.J. biol Chem. 263, 15868-15 871.
SHIMURA, H., OHTSU, M., MATSUZAKI, A., MITSUDOMI, T., ONODEEA, K.AND KIMUEA, G. (1988). Selective cytotoxicity of phoapholipids anddiacylglycerols to rat 3Y1 fibroblasts transformed by adenovinis type12 or its E1A gene. Cancer Res. 48, 578-583.
SKALLI, 0., ROPEAZ, P., THZECIAK, A., BKNZONANA, G., GnXKSSKN, D. ANDGABBLANI, G. (1986). A monoclonal antibody against (^smooth muscleactin: a new probe for smooth muscle differentiation. J. Cell Biol. 103,2787-2796.
TANIGUCHI, S., KAWANO, T., KAKUNAGA, T. AND BABA, T. (1986a).Differences in expression of a variant actin between low and highmetastatic B16 melanoma. J. biol. Chem. 281, 6100-6106.
TANIOUCHI, S., KAWANO, T., MITSUDOMI, T., KIMUHA, G. AND BABA, T.(19866). fos Oncogene transfer to a transformed rat flbroblast cell line
enhances spontaneous lung metastasis in rat. Jap. J. Cancer Res.(Gann)n, 1193-1197.
TANIGUCHI, S , SADANO, H., KAKUNAGA, T. AND BABA, T. (1989). Alteredexpression of a third actin accompanying malignant progression inmouse B16 melanoma cells. Jap. J. Cancer Res. (Gann) 80, 31-40.
VANDEKEROKHOVB, J. AND WKBKR, K. (1978). At least six different actinsare expressed in a higher mammal: an analysis based on the aminoacid sequence of the amino-terminal tryptic peptide. J. molec. Biol.128, 783-802.
WITT, D. P., BROWN, D. J. AND GORDON, J. A. (1983). Transformation-sensitive isoactin in passaged chick embryo fibroblasts transformed byRons sarcoma virus. J. Cell Biol. 96, 1766-1771.
ZAITSU, H., TANAKA, H., MITSUDOMI, T., MATSUZAKI, A., OHTSU, M. ANDKIMURA, G. (1988). Differences in proliferation properties amongsublines of rat 3Y1 fibroblasts transformed by various agents in vitro.Biomed. Res. 9, 181-197.
(Received 8 March 1990 - Accepted 14 May 1990)
Smooth muscle a-actin in 3Y1 cells 637






![Transient production of [alpha]-smooth muscle actin by ...blaulab.stanford.edu/pdfs/Springer-2002-Transient production.pdf · Differentiation in Culture and Following Intramuscular](https://img.dokumen.tips/doc/110x75/5f0926fc7e708231d42579bf/transient-production-of-alpha-smooth-muscle-actin-by-productionpdf-differentiation.jpg)










![Review Actin-targeting natural products: structures ... · actin-binding proteins actively break or ‘sever’ actin filaments [e.g. actin-depolymerizing factor (ADF) and cofilin]](https://img.dokumen.tips/doc/110x75/5f0f85bd7e708231d44494d0/review-actin-targeting-natural-products-structures-actin-binding-proteins-actively.jpg)












