Embed Size (px)
Citation preview

Marine Biology 53, 201-211 (1979) MARINE BIOLOGY �9 by Springer-Verlag 1979
Algal Chloroplasts in the Protoplasm of Three Species of Benthic Foraminifera: Taxonomic Affinity, Viability and Persistence
E, Lopez*
Institute of Ecology and Genetics; Ny Munkegade, DK-8000 Aarhus C, Denmark
Abstract
The ultrastructure and pigment content of algal chloroplasts (derived from Bacil- lariophyceae or Chrysophyceae) are described from 3 benthic species of brackish- water foraminiferans. Elphidiumwilliamsoni Haynes contains 4 x 106 chloroplasts mg -I , whereas the contents of Nonion germanicum (Ehrenberg) and E. excavatum (Terquem) are about 10% of this value. The two former contain chlorophylls a and c and fucoxan- thin, but these pigments were not detectable in the latter. E. williamsoni and N. ger- manicum had a net uptake of 14C-HCO~, proportional to their content of chlorophyll and number of chloroplasts, increasing linearly up to approximately 10 Klux. At light saturation the former assimilates 2.3 x 10-3 mg C mg -I h -I and the latter only about 20% of this value. Dark uptake was insignificant in all cases. Uptake could not be demonstrated in E. excavatum. The photosynthesis effected by these spe- cies is trivial in terms of the total benthic carbon fixation effected by the mi- croflora. The chloroplasts survived longer in foraminiferans kept in the dark than in light/dark adapted individuals. To keep a steady state population of chloro- plasts under light/dark conditions, E. williamsoni must eat at least 65 chloroplasts individual-1 h-l, whereas the minimum consumption rate in N. germanicum is 20.
I ntroduction
Associations with algae are common amongst foraminiferans, since the latter often exhibit delayed digestion ("food farming"); the algae survive in the pro- toplasm of the foraminiferans according to their resistance to digestion (Lee, 1974; Lee et al., 1974). In this respect, different types of associations have been demonstrated in experiments with algae-bearing members of the Nummuliti- dae and Soritidae. Thus, Heterostegina de- pressa lives solely on its symbiotic dia- toms (R~ttger, 1972; R~ttger and Berger, 1972; Schmalljohann and R~ttger, 1978), whereas Archaias angulatus and Sorites margi- nalis obtain less than 10% of their total carbon budget (Lee and Zucker, 1969; Lee and Bock, 1976) from their symbiotic green algae (Lee et al., 1974) and dino- flagellates (MHller-Merz and Lee, 1976). These differences may be correlated with
*Present address: Curriculum in Marine Sci- ences, University of North Carolina, 12-5 Venable Hall O45-A, Chapel Hill, North Caro- lina 27514, USA.
the survival of the algae in the foramini- ferans, since H. depressa needs no exter- nal food source, whereas primary produc- tion in A. angulatus is increased when fed isolated symbionts. This study describes a new type of "food farming" in fora- miniferans, of isolated algal chloro- plasts in the protoplasm of 3 benthic, brackish-water species, Elphidium william- soni Haynes, Nonion germanicum (Ehrenberg) and E. excavatum (Terquem) (cf. Hansen and Lykke-Andersen, 1976). The study has been concerned with the taxonomic affin- ity of the chloroplasts, and their pho- tosynthetic ability and persistence in the foraminiferans, in order to investi- gate whether this could constitute a "chloroplast symbiosis", a phenomenon hitherto described only for sacoglossan opisthobranchs (Trench, 1969, 1975; Tay- lor, 1970, 1974; Trench et al., 1973; Hinde and Smith, 1974).
MaWrialsandMcthods
The taxonomic affinity of the chloro- plasts was determined by means of their
0025-3162/79/0053/0201/S02.20

202 E. Lopez: Algal Chloroplasts in Foraminifera
ultrastructure, pigment composition and size. Photosynthetic ability was tested in fresh individuals of Eiphidium william- soni, Nonion germanicum and E. excavatum, as light-induced uptake of 14C-HC03; the persistence of the plastids was analyzed in starved foraminiferans adapted to al- ternating light and dark or to complete- ly dark conditions, as a decrease in photosynthetic carbon uptake, content of chlorophyll, and the number of chloro- plasts.
Collection and Isolation
The foraminiferans were isolated from surface sediments from two stations (depth O to 0.5 m and 6 m) in a Danish brackish water area, Limfjorden. Elphidium williamsoni and Nonion germanicum OCCUr to- gether at the shallowest station, where- as ~. germanicum and E. excavatum coexist at the deeper station, the density of each species being approximately 105 individ- uals m -2 .
The sediment was wet-sieved, and in- dividuals for the electron microscopic analysis were picked out from the 125 to 250 ~m sediment fraction and fixed imme- diately; in the remaining analyses and experiments, foraminiferans from 250 to 375 Lua sediment fraction were used. These individuals were washed several times in membrane-filtered sea water (HA-Millipore) to remove adhering algae. During this procedure, they were kept in the dark at temperatures below 15~ Samples of washed foraminiferans were examined under the fluorescence micro- scope; contamination was on an average I algae per 10 foraminiferans, and hence no corrections were made for adhering algae. All analyses and experiments were started within I week of collection, and less than 3 days after isolation of foraminiferans from sediment.
Ultras tructure
The method applied in the preparation of the foraminiferans for ultrastructural investigations with transmission elec- tron microscope was developed by Dr. C. F. Bardele, University of T~bingen, FRG (unpublished method).
The foraminiferans were fixed at room temperature (ca. 20~ and pH 6.6 in 2% glutaraldehyde (I h) and then in osmium tetroxide (2%, lh); they were decalci- fied at all stages of fixation with EGTA (Serva) incorporated into the cacodylate buffer. They were embedded in Epon 812, sectioned with a diamond knife on an LKB Ultrotome, stained with lead citrate and
uranyl acetate, and examined at 60 kV in a Philips 301 microscope.
Pigment composition and Chlorophyll Content
Two-dimensional thin-layer chromatogra- phy on sucrose plates (icing sugar with 10% potato starch) was used in the study of single pigments, in accordance with methods described by Jeffrey (1968). Ex- tractions were made with 90% acetone on samples of 500 to 1OOO individuals; the separated pigments were eluted in 90% acetone, and their absorption spectra were measured with a high-sensitivity spectrophotometer (Aminco DW 2a, UV-Vis). The pigments were identified by means of their absorption spectra, Rfvalues, and color.
Crude analyses of pigment content were made with extracts in 80 or 90% acetone on 300 to 500 specimens, and the absorption spectra of the extracts were measured with the Aminco spectrophoto- meter or a Unicam SP.800. The total amount of chlorophyll was calculated as described by Strickland and Parsons (1972).
Abundance and Size
Abundance and size of the chloroplasts were determined with an epifluorescence microscope, using the autofluorescence of pheophytin and chlorophyll (Fenchel and Straarup, 1971). Counts and measure- ments were made on samples of 100 crushed individuals on irgalan black- stained 0.2 ~m Nuclepore filters (Hobbie et al., 1977).
Photosynthetic Carbon Fixation
The photosynthetic ability of the chlo- roplasts was measured in samples con- taining 52 to 68 ~Ci 14C-HCO~ mmol-1, corresponding to a total concentration of 12.4 ~Ci per 1OO ml sea water (25 to 30~ S). The total CO 2 content was deter- mined by pH and alkalinity measurements (Strickland and Parsons, 1972). Each sample consisted of 20 individuals that were incubated in centrifuge tubes (dark controls were wrapped in aluminium foil), to which a small amount of autoclaved sediment (grain size 10 to 20 ~un) was added to provide a suitable physical substrate. Following incubation, the foraminiferans were collected on a plankton gauze (mesh size 63 ~m) in a Millipore filtering apparatus, and rinsed several times with filtered sea water to remove inorganic label and ad-

E. Lopez: Algal Chloroplasts in Foraminifera 203
hering sediment. They were then fixed in 5% neutral formaldehyde in filtered sea water, crushed, and counted in a liquid scintillation spectrometer, with Insta- gel as scintillation liquid. In all ex- periments, light intensity was measured with a Gossen luxmeter (I Klux =approxi- mately I. 11 J m -2 sec -I), and a Beckman Microbalance (LM-5OO) was used for de- termination of the ash-free dry weight (2.4 to 2.5 ~g individual-l), for which samples of 50 foraminiferans were dried at 80~ to constant weight and ashed for 4 h.
A series of experiments were con- ducted to determine the effect of light intensity on primary production. All ex- periments were made with incubations of foraminiferans in artificial light (At- las Super-Gro, 20 W) at 15~ for 4 or 8 h.
Primary production under natural con- ditions was measured in foraminiferans incubated in sequential 2 h periods for 24 h.
The rate of loss of 14C-photosynthate from Elphidium williamsoni and Nonion germani- cum was determined in control experi- ments at 7,000 lux and 15~ in which the foraminiferans were incubated in sea water containing 14C-HC03 for 8 h, then rinsed, and transferred to non-labelled medium. The loss rates were measured in samples taken O, 4, 8 and 12 h after transfer, giving an average loss rate of 0 to 3% uptake rate in both species.
survival of Chloroplasts
The persistence of chloroplasts in Elphi- diumwilliamsoni and Nonion germanicum was measured in individuals starved either in alternating light (3,500 lux) and dark (16 h light:8 h dark) or in com- plete darkness, at 15~ and 22~ S. Each day, adapted foraminiferans were incu- bated with 14C-HCO3 (10.3 ~Ci per 7OO ml sea water) for 8 h at 3,500 lux or as dark controls. The decrease in content of chlorophyll and number of chloro- plasts was measured every second day in similarly adapted individuals.
Results
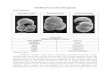
Several specimens of Eiphidiumwilliamsoni, Nonion germanicum and E. excavatum were exam- ined in the transmission electron micro- scope, and the only recognizable algal structures found were isolated algal chloroplasts, apparently of only one type. This type is illustrated in Fig. I for E. williamsoni. Fig. I: i shows the protoplasm in the proximal part of cham-
bers in the outer whorl of the foramini- feran. Chloroplasts are found throughout the protoplasm of all 3 species, but tend to be concentrated in the outermost parts.
No entire organelles have been ob- served in association with the chloro- plasts, but quite often tubular struc- tures, that could be part of algal or- ganelles, were observed in the lumen be- tween the chloroplast and the vacuolar membrane. These tubules are sometimes aggregated (Fig. I: 2, T).
Fig. I: 2 and 3 illustrate the chlo- roplasts in detail, and in the following they will be described in accordance with the chloroplast nomenclature of Dodge (1973).
The chloroplasts consist of a central, relatively fine-grained body, the pyre- noid, which is surrounded by a coarser region containing the lamellae of thyla- koids, in which the chlorophylls are concentrated.
The pyrenoid is crossed by I lamella, consisting of 3 thylakoids (Fig. I: 2, PL). Sometines a faint membrane-like structure can be distinguished in the distal part of the pyrenoid (Fig. I: 3, M).
The lamellae in the coarse-grained part of the chloroplast consist of 3 thylakoids (L in Fig. I: 2 and 3). Thy- lakoids crossing from one lamella to an- other can be seen relatively frequently (Fig. I: 3, C). No grana (stacks of thy- lakoids) have been observed.
The entire chloroplast is surrounded by a girdle lamella, mostly consisting of 3 thylakoids (G in Fig. I: 2 and 3). At each end of the chloroplast, close to the girdle lamella, I or 2 electron- translucent areas (Fig. I: 2, D) often occur. These are probably the areas where the chloroplast deoxyribonucleic acid (DNA) is concentrated, a feature closely connected with the presence of a girdle lamella (Dodge, 1973).
The chloroplast is bounded by a dou- ble membrane, the chloroplast envelope (Fig. 1: 2, E); sometimes, situated to- wards the vacuolar lumen, fragments of a second double membrane can be seen (Fig. I: 2, S, between the arrows). This is probably part of the endoplasmic reticu- lum sheath which surrounds the chloro- plast in entire algae.
Occasionally, decaying chloroplasts are found in Eiphidium williamsoni (Fig. I: 4).
In Nonion germanicum and Elphidium excava- tum, the chloroplasts are of the same type as or very similar to the chloro- plasts in E. williamsoni, since they have a girdle lamella and the same type of pyrenoid. However, no detailed study of

Fig. i. Elphidium williamsoni; ultrastructure. 1: Chloroplasts in chambers in outermost whorl ix 3,000). 2, 3: Details of chloroplasts; C: interconnecting thylakoids between lamellae; D: DNA areas; E: chloroplast envelope; G: girdle lamella; L: lamella consisting of 3 thylakoids; M: membrane-like structure bounding pyrenoid; PL: lamella running through pyrenoid, consisting of 3 thylakoids; S: (presumably) part of endoplasmic reticulu~n sheath; T: tubules in vacuolar lumen (2 x 22,600; 3 x 27,500). 4: Decaying chloroplast in outermost part of chamber (x 12,OOO)

E. Lopez: Algal Chloroplasts in Foraminifera 205
the lamellae was possible in these two species, because many of these chloro- plasts were decaying at the time of fix- ation and because of problems in fixa- tion.
The absorption values of the total pigment content in the foraminiferans (Fig. 2) are expressed in units per mg ash-free dry weight of the foramini- ferans. Elphi~um williamsoni and Nonion ger- manicum have very similar absorption curves, that of the former being about 4 times greater than that of the latter (E. williamsoni: 5.6 ~g chlorophyll rag-l; N. germanicum: 1.6 ~g chlorophyll mg-1). E. excavatum achieves maximum absorption at a shorter wavelength than the other two species.
The two-dimensional thin-layer chro- matography separated 6 distinct pigments in Elphidiumwilliamsoni and Nonion germani- cum, but only 4 in E. excavatum (Fig. 3). Pigment Spot I had an absorption maximum at 425 nm in E. excavatum and was identi- fied as pheophytin c. In E. williamsoni and N. germanicum the same spot had maxi- mum absorption at 444 nm and was predom- inantly chlorophyll c (Smith and Beni- tez, 1955). Pigment Spot 2 has been identified as fucoxanthin (Jeffrey, 1968; Hallegraeff, 1977) (maximum ab- sorption 446 to 450 nm). Pigment Spot 3 was chlorophyll a (maximum absorption 412 to 420 nm; peak at 656 nm) and, like fucoxanthin, it was only found in E. wil- liamsoni and N. germanicum. The rest of the pigments (Spots 4 to 6) occurred in all 3 species; Pigment 4 was a xanthophyll with maximum absorption at 451 nm and a shoulder at 470 to 475 nm; this pigment is probably diatoxanthin or a mixture of diatoxanthin and diadinoxanthin. Pigment 5 was pheophytin a (maximum absorption
O.D.. mg -t
1.2
1.0
0.8
0,6
0.4
0.2
z.O0
% %
."% \ .." \
,\ /', '...\ , ~
. . . . . . - : . - . . - . . : . . : : . . . ~ 500 600 700 nm
Fig. 2. Elphidium williamsoni (dashed line), No- nion germanicum (continuous line) and E. excava- tum (dotted line). Absorption spectra of total pigment content. Values are absorbancy (O.D.)
409 nm; peak 662 nm), and Pigment Spot 6 contained I or more carotenes (maximum absorption 450 nm; small peaks at 425 and 475 nm).
Examination with the fluorescence mi- croscope revealed that the diameter of the chloroplasts in all three species is very similar (Elphidium williamsoni: 3. I +- I. 8 ~m; Nonion germanicum: 2.9 + I. 4 ~m; E. excavatum: 2.8 + I .O ~/n).
Elphidium williamsoni contains the largest number of chloroplasts (4.0 _+ 2.0) x 106 mg-1~ corresponding to (9.7 _+ 4.9) x 10 3 individual -I. This is about 8 times the average number in the other two species (Nonion germanicum: (5.2 _+ 1.6) x 105 mg-1 (1.3 _+ 0.4) x 10 3 individ- ual-l; E. excavatum: (5.0 -+ 2.9) x 105 tug-1 (1.2 + 0.7) x 10 3 individual-l). The number of chloroplasts in E. williamsoni and N. germanicum was the same in individ- uals collected in summer and in winter, and therefore a steady state population of chloroplasts independent upon season is assumed to exist in these foramini- ferans.
In the study of photosynthetic carbon fixation at different light intensities, the rates of light fixation in Elphidium williamsoni always differed significantly from the corresponding dark controls -- (9.4 + 3.4) x 10-5 mg C mg -I ash-free dry weight h -I, which is on an average
g
.E_ "10
I.L
.0
0.5
~ 0
3 0
Rf votues, Second dimension
6 C Z ~ . 1 . 0
o.s ~ s
0.5 | 1.0
2@ | ,'~1 0.5 1.0
i ] i , , t t , t I
1 .0
5 ~
0.g
�9 ~ I , , , , 0,5 , , , , t,O
Fig. 3. (A) Elphidium williamsoni, (B) Nonion germanicum, (C) E. excavatum. Two-dimensional thin-layer chromatography of pigments. Pigment spots = i: chlorophyll c and pheophytin c, light-green; 2: fucoxanthin, orange; 3: chloro- phyll a, blue-green; 4: xanthophyil, unidenti- fied, light-orange; 5: pheophytin a, yellowish-
per mg ash-free dry weight (80% acetone extracts) green; 6: carotenes, yellow

206 E. Lopez: Algal Chloroplasts in Foraminifera
4% of the total light uptake at satura- tion level. The light values for Nonion germanicum were also significantly dif- ferent from dark values, except at the lowest light intensities -- (1.6 • 0.6) x 10 -4 mg C mg -I ash-free dry weight h -I, i.e., 25% of total light fixation at saturation level. There was no differ- ence between light and dark uptake in E. excavatum -- (5.3 • 2.3) x 10-5 mg C mg -I ash-free dry weight h-1.
In Fig. 4 the results of these ex- periments are given as net light fixa- tion. The lines have been calculated from the mean values at light intensi- ties less than 8,000 lux. The slopes of the lines indicate that Elphidiumwilliam- soni on an average takes up inorganic carbon at a rate about 5 times that in Nonion germanicum, and that the chloro- plasts in E. excavatum are inactive with respect to primary production.
The variation in primary production during 24 h was measured in Eiphidium wil-
liamsoni and Nonion germanicum in natural light. The production rates are plotted in Fig. 5, together with the variation in light intensity and temperature. The experiments were made on a cloudy day with relatively small variation in tem- perature. Both species reached light saturation level a few hours after dawn
xlO -4 mgC. mg~lhrs -1
28 I
24
20 O
16
12
8
0 - . --
2 4 6 8 10 12 1& klux
Fig. 4. Elphidium williamsoni (circles), Nonion germanicum (triangles) and E. excavatum (squares). Primary production with artificial light source as a function of light intensity, in mg carbon per mg ash-free dry weight of fora- miniferans per hour; dark controls subtracted. Values are means and ranges; for each species, 2 to 4 measurements were made at each light in- tensity investigated
and remained at this level for most of the remaining day.
In Fig. 6 the mean values of primary production from these experiments in natural light are given as a function of light intensity. This figure also shows the constructed light saturation curves for the two species. The saturation lev- els of these curves were calculated from the results of samples taken around mid- day. Thus, primary production rates at light saturation are 2.3 x 10-3 mg C mg -I ash-free dry weight h -I for Elphidium wil- liamsoni and 5.0 x 10-4 mg C mg-1 ash- free dry weight h-1 for Nonion germanicum; the latter rate is about 20% of the pro- duction rate of E. williamsoni. The under- saturated parts of these curves are the regression lines calculated from the re- sults of the experiments in artificial light. The data suggest that the chloro- plasts in the two species have the same
26
2 \ ~e/e'-'-" i / ~ 1 6
E
' 8
x
0
I , , , m J l l ' , , , u , , , ,
~0 I ~C
30 �9 22
10
I I l ~ l [ I t l l [ l ( $ 1 1 1 [ I
& 6 8 10 12 1A 16 16 ZO HOURS
Fig. 5. El~idi~ willi~soni (circles) and No- nion germanicum (triangles). Variation in prima- ry production throu~out a 24 h period in natu- ral light; values are means and ranges of prima- ry production in mg carbon per mg ash-free dry weight of foraminiferans per hour; dark controls s~tracted. For both species, i to 2 measure- ments were made d~ing each period. Variations in mean values of light intensity (squares) and temperature (crosses) are also shown

E. Lopez: Algal Chloroplasts in Foraminifera 207
Fig. 6. Elphidium williamsoni (circles) and No- nion germanicum (triangles). Primary production in natural light (data from Fig. 5) as a func- tion of light intensity (mean values), and con- structed light-saturation curves. Saturation levels of light-saturation curves were calcu- lated from primary production rates around mid- day in natural light (cf. Fig. 5); undersatu- rated part of the curves (light intensities up
to ca. i0,OOO lux) represents regression lines from Fig. 4. Values are in mg carbon per mg ash- free dry weight of foraminiferans per hour; dark
controls subtracted
x l 0 -/. mgC.mg-.lhrs-1
24-
20
16
12
8
@
,~ z~ ~
klux
10 20 30
10 3
'T �9 i i -
x
I
z 0 s
X
!
z 10 2
: (
( , n t - " Z
O O
10
y ~ 7.6 �9 102e -'001x
0
\ 0 o ~ y = 1./. - 103e - '024x
r 2=. 96 O O
\ o " o
~ y = 7 . 6 " 10 e -010x A ~ t r2='43
2 - Y = &9 - 10 e - ' 0 4 5 • r 2 = . 9 3
| 20 /-0 60 80 100 120 140
I 1 I t P I I I I I I I I I I I
HOURS
10, 6,
- 5-" I
&
2 •
@
6
&
I >
2 Z I
u 3
~n 1
o .6 (23
0
x
.2
y= 7.8 .10-3e --001x r2=.33
�9 �9 ~ O = 7.6 - 10 -3 e -'007x
~ r2_- 99
~ A y = 4.4.10-3e --007x r 2 =1.00
A " - &
~ , y = 4.4.10-3e -'016x r2=1.00
20 40 60 60 100 120 140 f i I i i I I I i I i I I I I I
y = 8.7.103e--003x
y = 9.&.103e-.010x O r2='95
"•A y = 1.5-103e -'010x ~ & r2=1, 00
y =1,3.103e ".019x r2=.88
20 40 60 60,5 100 120 140 I ] "1 I I I I I I I I i i I * i
HOURS
Fig. 7. Elphidium williamsoni and Nonion germanicum. Survival of chloroplasts in starved individuals of E. williamsoni (dark-adapted = filled circles; light/dark adapted = open circles) and N. germanicum (dark-adapted = filled triangles; light/dark adapted = open triangles) as a function of adaptation time. (A) Photosynthetic carbon fixation (mean and range, duplicate measurements) as counts min -I 20
individuals -i h -I. (B) Content of chlorophyll, 90% acetone extracts, as ug total chlorophyll per individual. (C) Number of chloroplasts per individual. Equations and correlation coefficients (r 2) are indicated for both adaptation series of each species in all 3 types of measurements

208 E. Lopez: Algal Chloroplasts in Foraminifera
Table i. Elphidium williamsoni and Nonion ger- manicum. Turnover time (TT, in hours) and ini- tial loss rate (ILR, in chloroplasts individ- ual -i h -l) of chloroplasts in foraminiferans starved either in alternating light and dark (L/D) or in complete darkness (D). Calcula- tions are based on measurements (cf. Fig. 7) of photosynthetic carbon fixation (A), chloro- phyll content (B) and number of chloroplasts (C), using the exponent of the equations and the assumed steady state population of chloro- plasts in the foraminiferans
Species and Turnover time and initial light condition loss rate
A B C TT ILR TT ILR TT ILR
E. williamsoni
D 830 iO 13OO iO L/D 45 230 155 65
N. germanicum
D IO5 15 150 iO L/D 25 60 6O 20
380 30 105 95
i00 15 55 25
saturation light intensity (ca. 10,0OO lux) .
Chloroplasts in Elphidium williamsoni and Nonion germanicum (Fig. 7) survive for a longer time in individuals adapted to continuous darkness than those kept in alternating light and dark. The regres- sion lines were calculated assuming an exponential loss (least-squares fit) of chloroplasts. The photosynthetic carbon- fixation experiments were terminated be- fore light-fixation values equalled dark controls. In each adaptation series, photosynthetic ability (measured in counts per minute), chlorophyll content, and number of chloroplasts decrease at comparable rates, indicating that the photosynthetic efficiency of the chloro- plasts, measured in carbon fixed per mg chlorophyll and per chloroplast, does not differ between fresh and starved in- dividuals. Using the average number of chloroplasts given in the previous sec- tion, the average turnover times and initial loss rates of chloroplasts were calculated (Table I).
Discussion and Conclusions
The isolated chloroplasts in Eiphidium williamsoni, Nonion germanicum and E. excavatum
the pyrenoid and the presence of a gir- dle lamella, the latter being present only in 5 classes of Chromophyta: Chlo- romonadophyceae, Xanthophyceae, Chryso- phyceae, Bacillariophyceae and Phaeophy- ceae (Dodge, 1973). The 3 thylakoids per lamella, the absence of grana, and the interconnecting thylakoids between la- mellae are also characters common to these classes. The pyrenoid is of the simple internal type with one lamella running through it (Dodge, 1973), a type known from several species of Bacillario- phyceae. In many of these species there is a reduction in the number of thyla- koids in the lamella of the pyrenoid (Drum and Pankratz, 1964), but in Nitz- schia sp. (Drum and Pankratz, 1964), Nitz- schia palea (Drum et al., 1966), and Melo- sira varians (Crawford, 1973) this lamella consists of 3 thylakoids. In a member of Chrysophyceae, Ankylonoton luteum (Van der Veer, 1970), the same type of pyrenoid, without reduction in number of thyla- koids, is present. The investigated mem- bers of the remaining 3 classes either do not contain pyrenolds, or these are of a different type than that described here (Dodge, 1973).
The chlorophylls a and c are typical for the algal classes Chrysophyceae, Bacillariophyceae, Phaeophyceae and Di- nophyceae, whereas the most abundant xanthophyll, which was identified as fucoxanthin, is not present in the lat- ter (Egle, 1960).
The ultrastructure and pigment compo- sition therefore suggest that the chlo- roplasts originate from Chrysophyceae or Bacillariophyceae. However, at this lev- el it is impossible to know how many al- gal species are donors of chloroplasts to the foraminiferans or to determine whether the chloroplasts of the three species of Foraminifera studied are de- rived from the same algae. All three species are deposit-feeders, feeding on sediment particles that they collect from their surroundings with their pseu- dopods. Because they have not been able to catch phytoflagellates, motile dia- toms and ciliates, when observed under dissection microscope, they are probably unable to capture motile food organisms. Therefore it is assumed that they obtain their supply of chloroplasts from ses- sile algae living on the surface of sed- iment particles.
Symbiotic algae belonging to either Chrysophyceae or Bacillariophyceae have been described in association with other
are typical eukaryotic organelles (Dodge, species of foraminiferans: Amphistegina Io- 1973) of algal origin, since they pos- bifera, "Cycloclypeus" tuberculatus, Operculina sess a pyrenoid (Griffiths, 1970). The ammonoides and Heterostegina depressa (Leu- most important characteristics for fur- tenegger, 1977; Schmalljohann and R~tt- ther identification are the structure of ger, 1978), but none of these algae con-

E. Lopez: Algal Chloroplasts in Foraminifera 209
tain chloroplasts identical to those in Elphidium williamsoni, Nonion germanicum and E. excava rum.
Thin-layer chromatography revealed that the observed difference in absorp- tion spectrum on crude pigment extracts between Elphidium excavatum and the other two species is caused by a lack in the former of the photosynthetically essen- tial pigments, chlorophylls a, c and fucoxanthin. In accordance with this, no light-induced uptake of 14C-HCO~ by the chloroplasts in E. excavatum could be de- tected, which means that they are prob- ably merely food items being digested by the foraminiferan. As opposed to this, E. williamsoni and Nonion germanicum pos- sessed a photosynthetic ability propor- tional to their content of chlorophyll and number of chloroplasts. Therefore, the association between the algal chlo- roplasts and these two species might be "chloroplast symbiosis", a term defined by Hinde and Smith (1974) as "the situ- ation where an animal takes up and re- tains free, undamaged chloroplasts under natural conditions, shows active photo- synthesis for at least several days after removal from its host plant, and utilizes products of photosynthesis re- leased from the chloroplasts", although in this study there is no evidence pre- sented for transfer of photosynthetic products from chloroplasts to hosts (see also Trench, 1975).
Total daily production by foramini- ferans at the shallow station can be estimated using the results of the pri- mary production experiments in natural light as approximately 3 x 10 -2 mg C mg-1 ash-free dry weight day -I in Elphi- dium williamsoni and 6.5 x 10-3 in Nonion germanicum. Assuming that the thickness of the photic zone in the sediment (Fenchel and Straarup, 1971) is 2 mm (the density of the 2 species being 4.2 x 104 and 1.6 x 104 individuals m -2, respectively) then, at this station, the total primary production by the chloroplasts in the foraminiferans would be 3.26 mg C m -2 day -I. At this station, the total prima- ry production of algae at the same time of the year has been estimated as ap- proximately 680 mg C m -2 day-1 (unpub- lished results), which means that about 0.5% of this production is achieved by the chloroplasts in the foraminiferans. This is in contrast to estimates in a tropical environment, where symbiotic algae in foraminiferans accounted for most of the primary production (Sournia, 1976).
The turnover times of chloroplasts estimated for Elphidium williamsoni and No- nion germanicum are within the range of previous estimates of survival of symbi-
otic chloroplasts in sacoglossan opis- thobranchs (Trench et al., 1969; Greene, 1970; Hinde and Smith, 1972, 1974, 1975), that can vary from a few days to several months, the longevity of the association probably reflecting the overall impor- tance of these chloroplasts in the ener- gy budget of their hosts (Hinde and Smith, 1975). The higher loss rates (ILR in Table I) of chloroplasts in light/dark than in dark are in accor- dance with measurements of survival in sacoglossans, and are probably caused by a faster depletion in the light of cer- tain photosynthetically essential com- pounds (Hinde and Smith, 1975), whose synthesis is controlled by the algal nuclei (Trench et al., 1973). Furthermore, the different loss rates indicate that the foraminiferans require a different input of chloroplasts to maintain a steady state in light and dark. Under natural conditions, the loss rates in the uppermost part of the illuminated zone of the sediment are probably higher than those measured in these experiments, since light intensity is usually much higher than 3,500 lux. This means that the consumption rates of the chloroplast donors in the foraminiferans are presum- ably higher at the sediment surface than the estimated loss rates.
The results of the primary production experiments with the foraminiferans rep- resent the total light-induced uptake of 14C-HCO~, assuming that the respiratory loss during the uptake period was negli- gible (control experiment gave an aver- age total loss rate of O to 3% of uptake rate).
The presence of symbiotic dinoflagel- lates in reef-building corals have been demonstrated to accelerate calcification in the light (Goreau, 1959, 1963; Goreau and Goreau, 1959; Vandermeulen and Mus- catine, 1974), and in the reef-coral Po- cillopora damicornis the 14C built into the skeleton was 9 to 12% of total uptake over a 24 h period (Muscatine and Cerni- chiari, 1969). In accordance with this, light-enhancement of calcification has been demonstrated in laboratory experi- ments with the symbiont-bearing fora- miniferans Archaias angulatus (Lee and Zucker, 1969) and Amphistegina lessoni (Mul- ler, 1978); in the latter, the deposi- tion of carbon into CaCO 3 was estimated to approximate 10% of the total uptake. However, in field experiments with Archa- ias angulatus and Sorites marginalis, Lee and Bock (1976) found no evidence of light- enhanced calcification; this may have been due to inadequate incubation time.
From literature values on respiration of other foraminifer species -- Ammonia beccarii tepida (Bradshaw, 1961), Allogromia

210 E. Lopez: Algal Chloroplasts in Foraminifera
laticollaris, Rosalina leei and Spiroloculina hyalina (Lee and Muller, 1973) -- and oth- er types of meiofauna of similar size (Lasserre, 1976), tentative estimates of the respiration of the foraminiferans can be calculated as (0.3 to 1.0) x 10 -3 mg C mg -I ash-free dry weight h -I. As- suming that 10% of the carbon uptake is
deposited as CaC03 in the shells, then the carbon gain through photosynthesis at maximum respiratory rate at light saturation level will be approximately 200% of the respiratory loss in Elphidium williamsoni and 45% of that in Nonion germa- nicum. However, the actual importance for the foraminiferans of the chloroplasts' photosynthetic activity depends upon the amount of photosynthate released to the hosts. In chloroplast-bearing sacoglos- sans, 20 to 50% of the photosynthetical- ly fixed carbon is transferred from
chloroplasts to hosts (Trench, 1975). If similar proportions of photosynthate are transferred to the foraminiferans then the photosynthetic activity of the chloroplasts could account for 40 to 100% of respiration in E. williamsoni and 10 to 20% in N. germanicum.
Acknowledgements. My gratitude is due to Drs. C.F. Bardele and J. Theilade for their kind ad- vice and help in connection with the ultrastruc- tural study. I am grateful to Dr. B. Deutch for help in pigment analyses and for providing the energy conversion data for the light measure- ments. I am also grateful to Ms. T. Kelstrup, Ms. A. SElling and Ms. K. Andersen for assis- tance in preparations. Dr. H.J. Hansen kindly read the manuscript. I am especially thankful to Professor T. Fenchel and Dr. G. Lopez for valu- able discussions and encouragement during this study. The spectrophotometers were provided by the Institute of Medical Biochemistry, Aarhus, and Department of Plant Physiology, Aarhus. The study was supported by a grant from the Danish Natural Science Research Council.
Literature Cited
Bradshaw, J.S.: Laboratory experiments on the ecology of Foraminifera. Contr. Cushman Fdn foramin. Res. 12, 87-106 (1961)
Crawford, R.M.: The protoplasmic ultrastructure of the vegetative cell of Melosira varians C.A. Agardh. J. Phycol. 9, 50-61 (1973)
Dodge, J.D.: The fine structure of algal cells, 261 pp. London: Academic Press 1973
Drum, R.W. and H.S. Pankratz: Pyrenoids, raphes, and other fine structure in diatoms. Am. J. Bot. 51, 405-418 (1964)
- - and E.F. Stoermer: Electron microscopy of diatom cells. In: Diatomeenschalen im elek- tronenmikroskopischen Bild, Vol. 6. pp 3-24, plates 514-613. Hrsgb. von J.-G. Helmcke and W. Krieger. Lehre: J. Cramer 1966
Egle, K.: Menge und Verh~ltnis der Pigmente. In: Encyclopedia of plant physiology, Vol. 5(i). The assimilation of carbon dioxide, pp 444-496. Ed. by W. Ruhland. Berlin: Springer-Verlag 1960
Fenchel, T. and B.J. Straarup: Vertical distri- bution of photosynthetic pigments and the penetration of light in marine sediments. Oikos 22, 172-182 (1971)
Goreau, T.F.: The physiology of skeleton forma- tion in corals. I. A method for measuring the rate of calcium deposition by corals under different conditions. Biol. Bull. mar. biol. Lab., Woods Hole 116, 59-75 (1959)
- Calcium Carbonate deposition by coralline algae and corals in relation to their roles as reef-builders. Ann. N.Y. Acad. Sci. 109, 127-167 (1963)
- and N.I. Goreau: The physiology of skeleton formation in corals. II. Calcium deposition by hermatypic corals under various conditions in the reef. Biol. Bull. mar. biol. Lab., Woods Hole 117, 239-250 (1959)
Greene, R.W.: Symbiosis in sacoglossan opistho- branchs: functional capacity of symbiotic chloroplasts. Mar. Biol. 7, 138-142 (1970)
Griffiths, D.J.: The pyrenoid. Bot. Rev. 36, 29- 58 (1970)
Hallegraeff, G.M.: Pigment diversity in freshwa- ter phytoplankton. I. A comparison of spectro- photometric and paper chromatographic methods. Int. Revue ges. Hydrobiol. 61, 149-168 (1977)
Hansen, H.J. and A.-L. Lykke-Andersen: Wall structure and classification of fossil and recent elphidiid and nonionid Foraminifera. Fossils Strata 10, 1-37 (1976)
Hinde, R. and D.C. Smith: Persistence of func- tional chloroplasts in Elysia viridis (Opis- thobranchia, Sacoglossa). Nature New Biol. 239 (88), 30-31 (1972)
- -"Chloroplast symbiosis" and the extent to which it occurs in Sacoglossa (Gastropoda: Mollusca). Biol. J. Linn. Soc. 6, 349-356 (1974)
- -The role of photosynthesis in the nutrition of the mollusc Elysia viridis. Biol. J. Linn. Soc. 7, 161-171 (1975)
Hobble, J.E., R.J. Daley and S. Jaspar: Use of Nuclepore filters for counting bacteria by fluorescent microscopy. Appl. envirl Micro- biol. 33, 1225-1228 (1977)
Jeffrey, S.W.: Quantitative thin-layer chroma- tography of chlorophylls and carotenes from marine algae. Biochim. biophys. Acta 162, 271-285 (1968)
Lasserre, P.: Metabolic activities of benthic microfauna and meiofauna. Recent advances and review of suitable methods of analysis. In: Benthic boundary layer, pp 95-142. Ed. by J.N. McCave. New York: Plenum 1976
Lee, J. J.: Towards understanding the niche of Foraminifera. In: Foraminifera, Vol. i. pp 207-260. Ed. by R.H. Hedley and C.G. Adams. London: Academic Press 1974
- and W.D. Bock: The importance of feeding in two species of soritid Foraminifera with al-

E. Lopez: Algal Chloroplasts in Foraminifera 211
gal symbionts. Bull. mar. Sci. 26, 530-537 (1976)
-, L.J. Crockett, J. Hagen and R.J. Stone: The taxonomic identity and physiological ecology of Chlamydomonas hedleyi sp. nov., algal flagellate symbiont from the foraminifer Ar- chaias angulatus. Br. phycol. J. 9, 407-422 (1974)
- and W.A. Muller: Trophic dynamics and niches of salt marsh Foraminifera. Am. Zool. 13, 215-223 (1973)
- and W. Zucker: Algal flagellate symbiosis in the foraminifer Archaias. J. Protozool. 16, 71-81 (1969)
Leutenegger, S.: Ultrastructure de foraminif@res perfor@s et imperfor~s ainsi que de leur sym- biotes. Cah. Micropal~ont. (NRS, Paris) 3, 1-51, plates 1-54 (1977)
Muller, P.H.: 14C fixation and loss in a fora- miniferal-algal symbiont system. J. foraml. Res. 8, 35-41 (1978)
M~ller-Merz, E. and J.J. Lee: Symbiosis in the larger foraminiferan Sorites marginalis (with notes on Archaias spp.). J. Protozool. 23, 390-396 (1976)
Muscatine, L. and E. Cernichiari: Assimilation of photosynthetic products of zooxanthellae by a reef coral. Biol. Bull. mar. biol. Lab., Woods Hole 137, 506-523 (1969)
R~ttger, R.: Die Kultur von Heterostegina de- pressa (Foraminifera: Nummulitidae). Mar. Biol. 15, 150-159 (1972)
- and W.H. Berger: Benthic Foraminifera: mor- phology and growth in clone cultures of Hete- rostegina depressa. Mar. Biol. 15, 89-94 (1972)
Schmalljohann, R. and R. R6ttger: The ultra- structure and taxonomic identity of the sym- biotic algae of Heterostegina depressa (Fora- minifera: Nummulitidae). J. mar. biol. Ass. U.K. 58, 227-237 (1978)
Smith, J.H. and A. Benitez: Chlorophylls: analy- sis in plant materials. In: Moderne Methoden der Pflanzenanalyse. Vol. 4, pp 142-196. Hrsg. von K. Paeck and M.V. Tracey. Berlin: Springer'Verlag 1955
Sournia, A.: Primary production of sands in the lagoon of an atoll and the role of foramini- feran symbionts. Mar. Biol. 37, 29-32 (1976)
Strickland, J.D.H. and T.R. Parsons: A practical handbook of seawater analysis, 2nd ed. Bull. Fish. Res. Bd Can. 167, 1-310 (1972)
Taylor, D.L.: Chloroplasts as symbiotic organ- elles. Int. Rev. Cytol. 27, 29-64 (1970)
- Chloroplast endosymbiosis. Port. Acta biol. (S~r. A) 14, 385-396 (1974)
Trench, R.K.: Chloroplasts as functional endo- symbionts in the mollusc Tridachia crispata (B~rgh), (Opisthobranchia, Sacoglossa). Na- ture, Lond. 222, 1071-1072 (1969) Of "leaves that crawl": Functional chloro- plasts in animal cells. Symp. Soc. exp. Biol. 29, 229-265 (1975)
-, J.E. Boyle and D.C. Smith: The association between chloroplasts of Codium fragile and the mollusc Elysia viridis II. Chloroplast ultrastructure and photosynthetic carbon fixation in E. viridis. Proc. R. Soc. (Ser. B) 184, 63-81 (1973)
-, R.W. Greene and B.G. Bystrom: Chloroplasts as functional organelles in animal tissues. J. Cell Biol. 42, 404-417 (1969)
Vandermeulen, J.H. and L. Muscatine: Influence of symbiotic algae on calcification in reef corals: critique and progress report. In: Symbiosis in the sea, Vol. 2. pp 1-19. Ed. by W. Vernberg. Columbia: University of South Carolina Press (The Belle W. Baruch Library in Marine Science) 1974
Van der Veer, J.: Ankylonoton luteum (Chryso- phyta), a new species from the Tamar estuary, Cornwall. Acta bot. neerl. 19, 616-636 (1970)
Date of final manuscript acceptance: May 4, 1979. Communicated by T.M. Fenchel, Aarhus