Embed Size (px)
Citation preview

This article was downloaded by: [New York University]On: 11 November 2014, At: 22:15Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Invertebrate Reproduction & DevelopmentPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tinv20
Adhesive papillae of Ascidia malaca swimminglarvae: investigations on their sensory functionMARIO GIANGUZZA a , GIUSEPPE DOLCEMASCOLO a , UGO FASCIO b & FIORENZA DEBERNARDI ba Institute of Biology, University of Palermo , via Divisi, 83, Palermo, 90133, ItalyPhone: +39 (91) 617-1123 Fax: +39 (91) 617-1123 E-mail:b Department of Biology , University of Milano , via Celoria, 26, Milano, 20133, ItalyPhone: +39 (02) 2660-4490 Fax: +39 (02) 2660-4490Published online: 01 Dec 2010.
To cite this article: MARIO GIANGUZZA , GIUSEPPE DOLCEMASCOLO , UGO FASCIO & FIORENZA DE BERNARDI (1999)Adhesive papillae of Ascidia malaca swimming larvae: investigations on their sensory function, InvertebrateReproduction & Development, 35:3, 239-250, DOI: 10.1080/07924259.1999.9652390
To link to this article: http://dx.doi.org/10.1080/07924259.1999.9652390
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy, completeness, or suitabilityfor any purpose of the Content. Any opinions and views expressed in this publication are the opinionsand views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy ofthe Content should not be relied upon and should be independently verified with primary sources ofinformation. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands,costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial orsystematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution inany form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Invertebrate Reproduction and Development, 35:3 (1999) 239-250 Balaban, Philadelphia/Rehovot 0 168-8 170/99/$05.00 O 1999 Balaban
Adhesive papillae of Ascidia malaca swimming larvae: investigations on their sensory function
MARIO GIANGUZZA", GIUSEPPE DOLCEMASCOLO', UGO FASCI$ and FIORENZA DE BERNARDI~ 'Institute o f Biology, University ofpalenno, via Divisi, 83, 90133 Palermo, Italy
Tel. +39 (91) 61 7-1123; Fax +39 (91) 616-6750; email: biomec&[email protected] 2~epartment o f~ology , University ofMilano, via Celoria, 26,20133 Milano, Italy
Tel. +39 (02) 2660-4490; Fax +39 (02) 236-2726
Received 22 January 1999; Accepted 13 April 1999
Summary
Swimming larvae ofAscidia malaca bear, in the anterior region of cephalenteron, three adhesive papillae through which they can attach to a substratum prior to metamorphosis. The ultrastructural investigations described in the present work have mainly revealed the presence of three cell types constituting the papillae: (a) collocytes, (b) axial columnar cells, (c) sensory cells. Collocytes, whose ultrastructure is typical of secretory cells, form the adhesive secretion. The ultrastructura1 observations of the axial columnar cells are insufficient to explain their function. Probably this cell type plays a supporting role though some sensory function cannot be excluded due to the presence of long microvilli in the apical region. Sensory cells are characterized by the presence of a single cilium at the cell apex and an axonal prolongation at its base. Based on their ultrastructure, these cells are primary sensory neurons playing a sensory role in papillae. In order to highlight papillar innervations we carried out laser confocal microscopy investigations on Ascidia malaca swimming larvae marked with anti-Btubulin fluorescent antibody. In the larval cephalenteron we observed an extensive nervous network starting from the papillar base and converging into two papillar nerves extending to the visceral ganglion. In conclusion, besides confirming the adhesive role of Ascidia malaca papillae, our investigations have also revealed the sensory function played by these organs.
Key words: Ascidian larvae, ultrastructural study; adhesive papillae innervation, monoclonal antibody
Introduction role in the larva-substratum adhesion process. Prior to metamorphosis, the majority of ascidian Many morphological and ultrastructural investi-
swimming larvae attach to a substratum by means of gations agree that one of the functions of papillae is to peculiar organs of ectodermic origin located in the secrete an adhesive substance which enables larvae to anterior region of the cephalenteron. The organs, adhere to a substratum at the onset of metamorphosis known as palps or adhesive papillae, play an important (Kowalesb, 1866; Grave, 1934; Bemll, 1950; Cloney,
*Corresponding author.
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

240 M. Gianguzza et al. / 1RD 35 (1 999) 239-250
1961, 1977, 1978, 1979; Millar, 1971; Turon, 1991; Gianguzza and Dolcemascolo, 1994,1997). According to a recent classification scheme by Burighel and Cloney (1997), two types of adhesive papillae can be found in ascidian larvae: (a) eversible papillae whose shape is rapidly changed on secretion of the adhesive material; (b) non-eversible papillae whose structure is maintained on secretion of the adhesive material. To the former type belong adhesive papillae with a very complex ultrastructural and functional organization [Distaplia occidentalis (Cloney, 1977), Diplosoma listerium (Lane, 1973), Diplosoma macdonaldi (Cavey and Cloney, 1976), Euherdmania claviformis (Trason, 1957), Eudistoma ritteri (Levine, 1962)l. On the other hand, Ciona, Ascidia, Styela, Pyura larvae have papillae of the latter type, characterized by a relatively simple ultrastructural and morpho-functional organization (Millar, 1971; Anderson et al., 1976; Cloney and Cavey, 1982; Torrence, 1983; Cloney and Torrence, 1984; Turon, 1991; Gianguzza and Dolcemascolo, 1994, 1997).
In other studies reported in the literature a sensory function, besides secretion, is attributed to papillae. This thesis is supported by ultrastructural investi- gations evidencing the presence of sensory cells (primary sensory neurons) in papillae of many ascidian larvae (Cloney, 1977, 1978, 1979; Torrence and Cloney, 1983; Turon, 1991). The axons leaving the base of the sensory cells converge into a papillar nerve extending up to the visceral ganglion. Innervated papillae were found in larvae with a complex functional and ultrastructural organization as in Diplosoma listerianum (Ghobashy, 1967), Distaplia occidentalis (Cloney, 1 977), Aplidium constellatum, Diplosoma macdonaldi (Cloney, 1979; Torrence and Cloney, 1983), Polysyncraton lacazei and Diplosoma spongiforme (Turon, 199 1).
Other cells whose ultrastructure could be compatible with a sensory function have been described also in the adhesive papillae of Ascidia malaca larvae (Gianguua and Dolcemascolo, 1994). For these cells the sensory function has only been hypothesized since the sections showed no innervation between papillae and the visceral ganglion. Recent studies with monoclonal antibodies in the swimming larva of Ciona intestinalis (Takamura, 1998) have evidenced some innervation linking papillae with the visceral ganglion. The adhesive papillae innervation has been evidenced at the confocal laser microscope in swimming larvae of Phallusia mammillata marked by anti-P-tubulin fluorescent antibody (Sotgia et al., 1998).
The purpose of this work, which deepens our ultrastructural knowledge ofAscidia malaca swimming larvae, is to supply further data to demonstrate the sensory function of adhesive papillae in ascidian larvae. To this aim, to reveal nerve fibres and their connection between papillae and the visceral ganglion, swimming larvae of Ascidia malaca were also observed by confocal laser microscopy after staining with anti-P-tubulin fluorescent antibody.
Materials and Methods
Adult A. malaca specimens were taken from the Gulf of Palermo during July-November. They were transferred to an aquarium and maintained at 18-20°C. Fertilization occurred in Syracuse dishes containing eggs and spermatozoa withdryn from gonoducts of adult specimens. Embryos were left to develop up to the stage of swimming larva (6-8 h after hatching) and then utilized for optical and electron microscopy investigations.
Morphological investigations Light microscopy
A. malaca swimming larvae (6-8 h after hatching), vitally stained for 5 min with Nile blue sulphate (1 : 100.000 in seawater) and kept in culture at 1 8 " C, were observed and photographed with a Leitz Orthoplan microscope using an Ilfor FP4 plus film.
Scanning electron microscopy A. malaca swimming larvae were fixed in PAF
(picric acid-formaldehyde) 1200 mOsm, pH=7.5. The fixed larvae were dehydrated in a graded ethanol series, critical point dried and gold sputtered. They were then observed and photographed in a Cambridge Stereoscan S 250 Mk2 scanning electron microscope.
Transmission electron microscopy
A. malaca swimming larvae (6-8 h after hatching) were fixed in 2.5% glutaraldehyde in 0.2M phosphate buffer pH=7.5 and postfixed in 1% osmium tetroxide in the same buffer. The fixed material was dehydrated in a graded ethanol series and embedded in Epon 8 12 (Luf?, 196 1). The sections obtained with the,,Ultracut E (Reichert-Jung) microtome were contrasted with uranyl acetate and lead citrate (Reynolds, 1963) arrd photo- graphed with a Phillips EM 410 at 80 kV accelerating voltage using Kodak electron microscope film (Estar thick base 4489).
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

M. Gianguzza et al. / IRD 35 (1999) 239-250 24 1
Confocal laser scanning microscopy A. malaca swimming larvae (6-8 h after hatching)
were fixed at -20°C in methanol containing 1% formaldehyde (from a 37% formaldehyde solution) for 2 h or longer. They were gradually rehydrated to PBS (phosphate-buffered saline), extracted for 20 min with 0.25% Triton X-100 in PBS, rinsed in PBS and then incubated in anti-l)-tubulin (mouse) monoclonal N 357 (Amersham) diluted 150. After repeated rinsing in 0.1 % Tween-PBS, larvae were incubated with FITC- conjugated anti-mouse IgG diluted 150. They were then rinsed in PBS and mounted in Citifluor. The larvae were observed under a Leica TCSNT confocal laser scanning microscope equipped with an argon1 krypton laser. 40x and 10x objectives were used, and images were obtained from eight scans of laser beam.
Results
Papillar structure and organization Light microscope observations Fig. 1 shows an image of a swimming larva of A.
malaca (6-8 h after hatching) vitally stained with Nile blue sulphate. Three glandular organs of ectodermic origin are present in the anterior region of the cephalenteron. These organs, two being in the dorsal and the third in the ventral area, are known as palps or adhesive papillae. In the central region of the cephalenteron lies the cerebral vesicle with its two sensory organs, i.e., the otolith containing gravity receptors and the ocellus containing light receptors. At the back of the cerebral vesicle the visceral ganglion is located. It is composed of a group of neuronal and ependymal cells. The visceral ganglion receives sensory input and controls natatory movements of the larva. The tail, attached to the cephalenteron base, shows the notochord, an axial structure made of a row of cells. The notochordal cells have become rearranged to form a tubular epithelium that runs the whole length of the tail.
SEM observations SEM observations reveal the shape and arrange-
ment of the three adhesive papillae present in the anterior region of the cephalenteron. Papillae are cone- shaped (Fig. 2) and lie in the vertices of a triangular field. In the apical part the external tunic of papillae have numerous fenestrations of variable size (Fig. 3).
TEM observations Adhesive papillae consist of elongated cells,
forming the papillar body, and have a sort of hyaline cap covering the latter (Fig. 4). In the apex of the hyaline cap a medium electron-dense, finely granular substance is present that very likely corresponds to the adhesive substance secreted by the papilla (cf. Gianguza and Dolcemascolo, 1994, 1997).
Hyaline cap - The hyaline cap is an expansion of the matrix test at the distal end of each papilla. It is dome-shaped and is limited by a cuticular layer. Along its whole length it contains numerous microvilli that originate from axial columnar cells (Fig. 5). The far tips of some microvilli can be seen to protrude out of the hyaline cap apex through fenestrations of the cuticular layer that quite likely correpond to the cuticular fenestrations observed at SEM.
Papillar body - Three cell types constitute the papillar body and characterize its functions: (a) cells with a secretory activity or collocytes, (b) axial columnar cells, and (c) sensory cells.
(a) Secretory cells or collocytes: Their ultrastructure is typical of cells with a'secretory activity. These cells lie in the central or middle-lateral side of the papillar body (Fig. 4). While confirming previous data (cf. Gianguzza and Dolcemascolo, 1994, 1997), the ultrastructural investigations have shown the elongated shape of these cells and the presence of abundant RER vesicles in the cytoplasm. The apical part of the latter contains secretory granules and the Golgi complex is apparently actively engaged in synthesis (Figs. 4,6,10, 15).
@)'Axial columnar cells: In the mid-central position lie cells with an elongated shape. They run along the entire papilla, their basal part resting on a continuous laminar structure, the basal lamina (Fig. 6). Their cytoplasm contains (Figs. 6, 9) very elongated RER cisternae and a few Golgi profiles and secondary lysosomes. The nucleus lies at the cell base, the apical regicn of the cytoplasm contains abundant vesicles and mitochondria A peculiar characteristic of these cells is represented by long microvilli that, originating from the basal part of the membrane, run along the whole hyaline cap length (Fig. 8). Some observations have highlighted microvillar ends protruding out of the hyaline cup via fenestrations of the cuticle (Fig. 5). Fig. 7 is the cross section of an area of the basal lamina located below the axial columnar cells. The basal lamina is thick as result of the fusion of two layers. It consists of components from the papillary epidermis and components from underlying endodermal epi- thelium of the gut rudiment.
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

M. Gianguno et al. / IRD 35 (1999) 23S250
Fig. 1. Swimming larva of Ascidia malaca stained with blue Nile sulphate. At the anterior end of the cephalenteron (ce) three adhesive papillae, two dorsal (dp) and one ventral (vp) are present. Also are visible in the cerebral vesicle (cv) two sensory organs, the ocellus (oc) and the otholith (ot). The visceral ganglion (vg) is located at the back of the cerebral vesicle. In this micrograph the larval tunic and most of the attached test cells are missing. ta, tail; no, notochord. Bar: 1 pm. Fig. 2. SEM image of a swimming larva of A. malaca showing the three simple conic adhesive papillae (p) at the anterior end of the cephalenteron (ce), located in the vertices of a triangular field. Bar: 40 pm. Fig. 3. Papillae, at a higher magnification, showing fenestrations in the apical part of the tunic (arrowheads). Bar: 10 pm. Fig. 4. Ultrastructure of an adhesive papilla Each papilla consists of a mass of elongated cells on which rests the hyaline cap (hc). At the centre of the papillar body secretory cells (coll) with abundant RER vesicles (rer) and secretion granules (sg) are visible. At the margins sensory cells (s cell) with a single cilium at their apical ends are visible. In the apex of the hyaline cap (hc) a fine granular substance (*) can be noted. N, nucleus. Bar: 1 pm. Fig. 5. The hyaline cap (hc) appears ai an expansion of the larval tunic above the cells constituting the papillar body. Abundant and long microvilli (v) starting fiom the base of the cell membrane, extend for the whole cap length. Arrowheads indicate the places where the microvillar tips protrude from the hyaline cap. Bar: 1 bm.
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

M. Gianguzza el al. / IRD 35 (1999) 239-250
Figs. 6-9. Ultrastructure of axial columnar cells (acc). Fig. 6. The cells extend for the whole papillar length and lie in the mid- central position. Fig. 9. Visible in the cytoplasm are scarce RER cisternae (rer), Golgi elements (gc) and secondary lysosomes The apical cytoplasm is highly vesicular with abundant mitochondria (m). Fig. 8. Long microvilli (v) starting from the cell membranes extend for the whole length of the hyaline cap (hc). The basal part of the cells rests on a continuous laminar structure, the basal lamina (bl). Fig. 7 shows the ultrastructure of the basal lamina from a cross section. N, nucleus. Bar: 1 prn.
(c) Sensory cells: These cells occupy a very apical end, of a single cilium whose initial part rests marginal position in the papilla (Fig. 4). Their ovoidal inside a sort of pocket of the cell membrane. A basal and elongated shape is enlarged in the basal area where body can be noted at the cilium base (Figs. 1 1, 12, 14). the nucleus lies. Mitochondria and rare RER elements Fig. 14 shows the cross section of a cilium and can be seen in the cytoplasm (Figs. 4, 10, 11). An highlights the 9x1 peripheral +1 central microtubules interesting feature of these cells is the presence, in the constituting the axoneme. The same figure shows how
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

M. Gianguzza et al. / IRD 35 (1999) 23S250
Figs. 10,ll. Longitudinal section of a papilla evidencing an ovoidal shaped sensory cell (s cell) located marginally. Mitochondria (m) and scarce RER cisternae (rer) are also visible. The nucleus (N) lies in the basal part of the cell. Bar: 1 pm. Figs. 12,13, 14. Ultrastructure of a cilium (c). A basal body (bb) is visible at its base. A cross section (Fig. 13) shows the 9 x 1 peripheral + 1 central microtubules constituting the axoneme. coll, collocyte; sg, secretion granule; hc, hyaline cap; ct, cuticle. Bar: 1 pm.
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

M. Gianguzza et al. / IRD 35 (1999) 239-250
Fig. 15. Longitudinal section of a papilla showing an axial columnar (acc), a collocyte (coll) and a sensory cell (s cell). The basal part of the sensory cell is prolonged into a long axon (ax) running between the ectoderm (ect) and the basal lamina (bl); ct, cuticle; tc, test cell. Bar: 1 pm.
the far end of the cilium, out of the cytoplasm pocket, sensory cells is shaped like a cone and is prolonged almost reaches the inner cuticular layer of the hyaline into a long axon running between the ectodermic cell cap. A further peculiarity of these cells in relation to layer of the cephalenteron and the basal lamina. their basal part can be seen in Fig. 15. The basal part of Abundant ribosomes, a few mitochondria and bundles
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

M. Gianguzza et al. / IRD 35 (1999) 239-250
Fig. 16. Swimming larva ofAscidia malaca stained by FITC-conjugated anti-P-tubulin antibody, observed by laser confocal scanning microscopy. Some bright fluorescence can be noted in the three adhesive papillae (p), in the visceral ganglion (vg), and in the neural tube of the tail (ta). Bar: 1 pm. Figs. 17, 18. Cephalenteron at a higher magnification. Fig. 17. The reconstruction made fiom eight confocal microscope images shows a papillar nerve (pn) into which converge the nervous branches coming fiom the two dorsal papillae (dp); the branches deriving from the ventral papilla (vp) join into the other papillar nerve. The two papillar nerves join with the visceral ganglion (vg). Fig. 18. An optical section taken at a greater depth show that before joining with the visceral ganglion, the two papillar nerves seem to converge for a short tract. Bar: 1Oym.
of microtubules running parallel to the axoneme are resting on the basal lamina. The entire papilla is present in the cytoplasm. The axon is devoid of myelin. surrounded by the tunic on which a test cell can be Besides sensory cells, Fig. 15 also shows a collocyte observed. with secretory granules and an axial columnar cell
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014
M Gianguzza et al. / IRD 35 (1999) 23S250
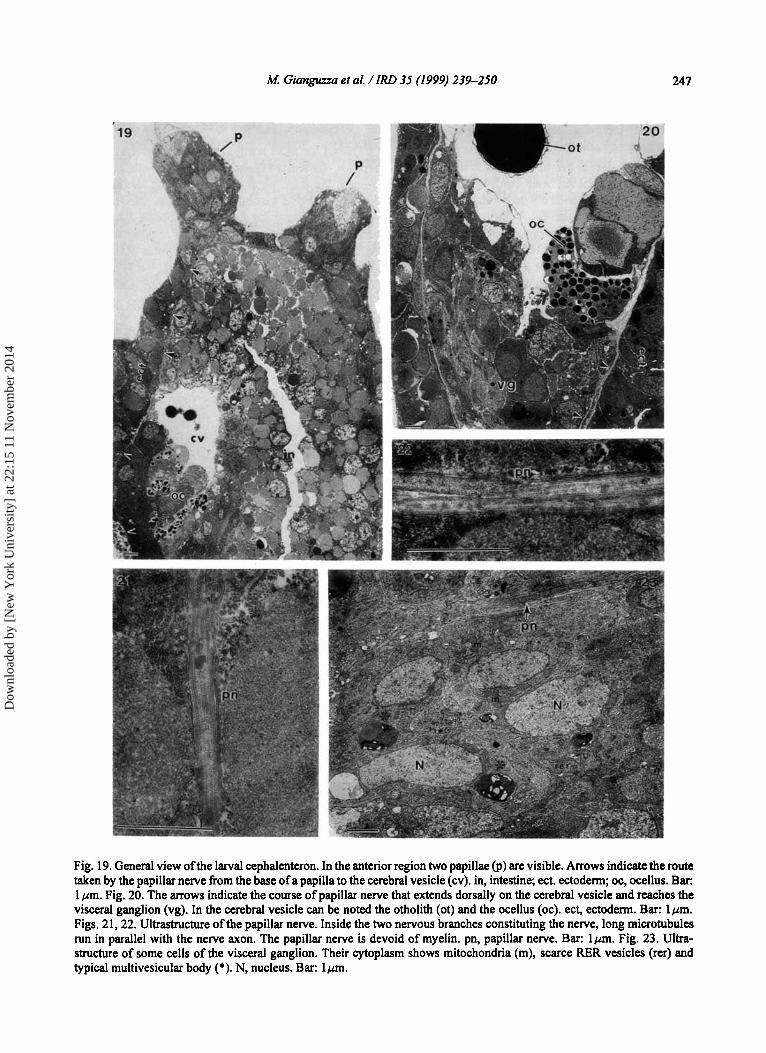
Fig. 19. General view of the larval cephalenteron. In the anterior region two papillae (p) are visible. Arrows indicate the route taken by the papillar nerve from the base of a papilla to the cerebral vesicle (cv). in, intestine; ect. ectoderm; oc, ocellus. Bar: 1 pm. Fig. 20. The arrows indicate the course of papillar nerve that extends dorsally on the cerebral vesicle and reaches the visceral ganglion (vg). In the cerebral vesicle can be noted the otholith (ot) and the ocellus (oc). ect, ectoderm. Bar: lpm. Figs. 21,22. Ultrastructure of the papillar nerve. Inside the two nervous branches constituting the nerve, long microtubules run in parallel with the nerve axon. The papillar nerve is devoid of myelin. pn, papillar nerve. Bar: lpm. Fig. 23. Ultra- structure of some cells of the visceral ganglion. Their cytoplasm shows mitochondria (m), scarce RER vesicles (rer) and typical multivesicular body (*). N, nucleus. Bar: lpm.
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

248 M. Gianguzza et al. / IRD 35 (1 999) 239-250
Papillar innervation Confocal microscope observations
In order to locate the innervation system arising from the papillar base, laser confocal microscopy investigations were carried out on swimming larvae of A. malaca treated with anti-P-tubulin antibody. Obser- vations have shown that fluorescence is strong in the adhesive papillae, in the visceral ganglion and in the neural tube (Figs. 16-1 8). The latter, located above the notochord, runs the whole length of the tail (Fig. 16). In papillae, marking by anti-P-tubulin antibody is stronger in cells sited centrally than in those at the margins (Figs. 17, 18). A higher magnification of the cephalenteron shows that at the papillar base the nerves are interconnected and proximally fused into two papillar nerves that reach the visceral ganglion. The reconstruction obtained from eight scanner images showed a dorsal nerve formed by branches deriving from the two dorsal papillae; a ventral nerve conveys the ventral papillar branches to the visceral ganglion (Fig. 17). In an optical section taken at a deeper level, the two nerves seem to h e into a short distal tract (Fig. 18).
TEM observations Ultrastructural investigation ofthe cephalenteron of
A. malaca larvae have evidenced a nervous connection between the base of the adhesive papilla and the visceral ganglion, thus confirming our confocal microscope observations. Fig. 19 shows a general view of two papillae sited in the anterior region of the larval cephalenteron. In the same micrograph can be followed the course of papillar nerve (arrows). The papillar nerve begins at the base of papilla, extends to the cerebral vesicle and continues its course dorsally between the ectoderm and the cerebral vesicle. The primitive intestinal cavity can be noted on the side of the cerebral vesicle. Fig. 20 shows the area where the distal end of the papillar nerve extends to the visceral ganglion. In the cerebral vesicle the otholith, the receptor organ for gravity detection, and the ocellus, the photoreceptor organ, can be noted. Figs. 2 1 and 22 show ultra-thin sections of the papillar nerve. This is made of two or more axons, with microtubules inside running parallel to the long axis. In Fig. 23 some cells of the visceral ganglion are shown. These cells do not present a highly differentiated ultrastructure. In their cytoplasm mitochondria, rare RER vesicles and typical multivesicular bodies can be noted. The arrow indicates the papillar nerve connected to the visceral ganglion.
Discussion
One of the most peculiar features of ascidians are papillae, glandular organs of ectodermic origin. In A. malaca, the three papillae of swimming larvae, two dorsally and one ventrally located, are sited in the vertices of a triangular field. Like those of Phallusia mammillata (Sotgia et al., 1998) and of other solitary ascidians, they are simple coniform papillae. Rarely, as in Eurherdmania claviformis (Trason, 1957) and in numerous species of the Didemnum genus (Lafargue, 1976), only two papillae are present, located along a sagittal plane.
It is agreed that the papillae secrete an adhesive substance by means of which swimming larvae can adhere to a substratum and thus start their colonization by becoming adult sessile organisms (Cloney, 1977, 1978,1979; Millar, 197 1 ; Turon, 199 1 ; Gianguzza and Dolcemascolo, 1994, 1997; Burighel and Cloney, 1997). According to other authors, molgulids are devoid of adhesive papillae (Berrill, 1950; Grave, 1926; Cloney, 1961; Torrence and Cloney, 1981; Torrence, 1983). In these ascidian larvae the anterior region of the cephalenteron tunic becomes sticky and allows larvae to fix to a substratum to start meta- morphosis.
Besides adhesion, papillae of ascidian larvae have been attributed a sensory function. According to Grave (1934) and Grave and Riley (1935) in Botrillus, for instance, papillae are exclusively sensory organs. The sensory function attributed to ascidian adhesive papillae is mainly the result of ultrastructural investi- gations. This function is established in eversible adhesive papillae with a very complex ultrastructural and morpho-functional organization. In Distaplia occidentalis, for instance, Cloney (1977, 1978, 1979) has described papillae made of six cell types each with a distinct ultrastructure and function. Secretion of the adhesive material is performed by peculiar cells known as "collocytes", whereas the sensory function is possessed by "anchor" cells and basal cells. The presence of microvilli and cilia in the above cells enables them to carry out the sensory function through recognition of certain stimuli (chemical andlor mechanical) coming from the substratum and the adhesion process is triggered by secretion of the adhesive material. Papillae with a sensory function have been found in larvae of Diplosoma macdonaldi (Torrence and Cloney, 1983), and more recently in Polysyncraton lacazei, Diplosoma spongiforme and Ecteinascidia turbinata (Turon, 199 1). In larvae of D. macdonaldi papillae are organs with a remarkable
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

M. Giangwza et al. / IRD 35 (1999) 239-250 249
structural and functional complexity: not only are papillae everted at the same time, thus proving that this function is controlled by a synchronous process, but also the substratum to which they adhere is tested and chosen according to its features (Cloney, 1978, 1982; Torrence and Cloney, 1983).
Papillae with a simpler ultrastructural and functional organization have been described in larvae of some species in the families Cionidae, Ascididae, Styelidae and Pyuridae (Millar, 1971 ; Cloney, 196 1, 1977, 1978, 1979; Gianguzza and Dolcemascolo, 1994, 1997). In the classification made by Burighel and Cloney (1 997), they were considered to be simple, coniform papillae, not eversible and not consistently endowed with a sensory function. Papillae of A. malaca larvae, which have already been described by Gianguzza and Dolcemascolo (1 994, 1997), belong to this type and are made of two cell types with a distinct ultrastructure and function. The cell type known as "A" has the typical ultrastructure of secretory cells and its function is to form the adhesive material. The function of the second cell type, known as "B", is uncertain based on ultrastructural information alone. A sensory h c t i o n for these cells was hypothesized based on the presence of long microvilli although no axonal prolongations have been observed on these cells. This hypothesis is not longer credible.
The ultrastructural investigations herewith described carried out on the adhesive papillae of A. malaca have confirmed the data reported in the literature which attribute a sensory function, besides secretion, to the adhesive papillae of ascidian larvae. Moreover, the observations have more clearly revealed the presence and position of the cells constituting the papillar body and characterizing its functions. Three cell types were essentially found: secretory cells or collocytes, axial columnar cells and sensory cells.
Cells with an ultrastructure typical of secretory cells lie in a median position. Ultrastructurally and functionally this cell type matches the collocytes described by Cloney (1977) in Distaplia occidentalis and the secretory cells described by Turon (1991) in Ecteinascidia turbinata. In order to utilize the same terms, as "collocytes" we also include the above cell type found in the adhesive papillae of A. malaca.
The second cell type present in adhesive papillae of A. malaca are axial columnar cells. They lie in the mid- central papillar region and extend for the whole papillar length. Their base rests on a continuous laminar structure, the basal lamina. No axonal prolongation has been observed in the sections. These cells could have a supporting role, as suggested by
Cloney (1977) for cells with a similar ultrastructure found in papillae of D. occidentalis. Our ultrastructural investigations have shown the presence in the apical part of the cells of long microvilli extending for the whole length of the hyaline cap and beyond it with their far ends, through fenestrations of the tunic. Also in Phallusia mammillata larva at the onset of meta- morphosis, we observed cells located in the papillar centre that present actin-containing microvilli protruding out of the tunic fenestrations (Sotgia et al., 1998). This arrangement undoubtedly facilitates the perception of stimuli coming from the substratum, and thus the hypothesis of a sensory function for these cells cannot be ruled out.
The thud cell type lies in a definitely marginal area of the papilla. The apical part of the cells bears a cilium originating from a sort of pocket of the cell membrane. The far end of the cilium extends to touch the internal cuticle of the hyalin cap. The main feature of these cells is their basal part prolonging into a long axon. The presence of a cilium in the apical region and of an axonic process at their base indicate that these cells possesses a sensory function. The present ultra- structural investigations have also shown that the axons starting from the basal regions of numerous cells fuse into the papillar nerve, a bundle connecting the papillar base with the visceral ganglion. Ultrastructurally these cells quite strongly resemble the "anchor" cells and basal cells described by Torrence and Cloney (1 983) in Diplosoma macdonaldi, by Cloney (1977, 1979) in Distaplia occidentalis and by Turon (1 99 1) in Ecteinascidia turbinata and Polysincraton lacazei. These authors have classified this cell type as primary sensory neurons. Sensory cells with axons forming the papillar nerve have also been shown in larval papillae of Boltenia villosa, Styela plicata (Cloney and Torrence, unpublished observations) and Ciona intestinalis (Torrence, 1 983).
The confocal microscopy observations made by use of anti-p-tubulin antibodies have shown numerous nervous branches that, starting from the papillar base, become interconnected and fused into two papillar nerves. These last seem to converge into a single short nerve that extends to visceral ganglion. The nervous network observed in A. malaca is similar to that found in Phallusia mammillata (Sotgia et al., 1998); how- ever, in the latter, the nerves of two dorsal papillae join at a short distance from their bases, while the nerve of the ventral papilla fuses just before the brain vesicle. By means of monoclonal antibodies Takamura (1998) has recently highlighted two main axon bundles linking papillae with the visceral ganglion directly or
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014

250 M Gianguzza et al. / IRD 35 (1999) 239-250
indirectly. This type of innervation is probably present in papillae of many ascidians, and by ultrastructural investigations and confocal microscopy, the route taken by the nervous network in the larval cephalen- teron could be observed.
In conclusion, the research presented in this work, besides confirming the secretory function of some cells, has also demonstrated the presence of primary sensory cells in the papilla of A. malaca.
Acknowledgements
This research was supported by MURST (60%) grants.
References
Anderson, D.T., White,B.M. and Egan, E.A., The larval development and metamorphosis of the ascidiabs W r a praeputialis (Heller) and Pyura pachydermatina (Herdman) (Pleurogona, family Pyuridae). Proc. Linn. Soc. N.S.W., 100 (1976) 205-217.
Berrill, N.J., The Tunicata with an account of the British species. Ray Society, London, 1950.
Burighel, P. and Cloney, R.A., Urochordata. Ascidiacea In: Microscopic Anatomy of Invertebrates, Vol. 15, Hemi- chordata, Chaetognatha and the Invertebrates Chordates. Wiley-Liss, 1997, pp. 221-347.
Cavey, M.J. and Cloney, R.A., Ultrastructure and differ- entiation of ascidian muscle; I. Caudal musculature of the larva of Diplosoma macdonaldi. Cell Tiss. Res., 174 (1976) 289-313.
Cloney, R.A., Observations on the mechanism of tail resorption in ascidians. Amer. Zool., 1 (1961) 67-87.
Cloney, R.A., Larval adhesive organs and metamorphosis in ascidians. I. Fine structure of the everting papillae of Distaplia occidentalis. Cell Tiss. Res., 183 (1977) 423- 444.
Cloney, R.A., Ascidian metamorphosis: review and analysis. In: Settlement and Metamorphosis of Marine Inver- tebrate Larvae, F.S. Chia and M. Rice (eds.), Elsevier1 North-Holland Biomedical Press, New York, 1978, pp. 255-282.
Cloney, R.A., Larval adhesive organs and metamorphosis in ascidians. 11. The mechanism of eversion of the papillae of Distaplia occidentalis. Cell Tiss. Res., 200 (1979) 453473.
Cloney, R.A., Ascidian larvae and the events of metamor- phosis. Amer. Zool., 22 (1982) 817-826.
Cloney, R.A. and Cavey, M.J., Ascidian larval tunic: extraembryonic structures influence morphogenesis. Cell Tiss. Res., 222 (1 982) 547-562.
Cloney, R.A. and Torrence, S.A., Ascidian larvae: structure and settlement. In: Marine Biodeterioration: an inter- disciplinary study, J.D. Costlow and C. Tipper (eds.), Naval Institute Press, Anapolis, 1984, pp. 14 1-148.
Giangu~a , M. and Dol&mascolo, G., Adhesive papillae in the swimming larva ofAscidia malaca: an ultrastructural
investigation. Eur. Arch. Biol., 105 (1994) 51-62. Gianguzza, M. and Dolcemascolo, G., Adhesive organs in
the swimming larvae of Ascidia maloco: an ultra- structural and cytochemical study. Anim. Biol., 6 (1997) 53-60.
Ghobashy, A.F.A.A., Larval behavior and metamorphosis in the ascidian Diplosoma Iisterianum (Milne Edwards). PhD Thesis, University of Wales, 1967.
Grave, C., Molgula citrina (Alder and Hancock). Activities and structure of the free-swimming larva. J. Morph., 42 (1926) 453-467.
Grave, C., The Botryllus type of ascidian larva Carnegie Inst. Wash. Publ., 435 (1934) 143-156.
Grave, C., The larva of Siyela (Cynthia) partita: Structure activities and duration of life. J. Morph., 75 (1944) 173- 191.
Grave, C. and Riley, G., Development of the sense organs of the larva of Botryllus schlosseri. J. Morphol., 57 (1935) 185-211.
Kowalesky, A., Entwicklungsgeschichte der einfachen Ascidien. Mem. Acad. Sci. St. Petersb., 7(10) (1866) 1-19.
Lafargue, F., Rbvision taxonomique des Didemnidae des c6tes de France (ascidies composbes). Les espkes de Banyuls-sur-mer. Genre Didemnum. 2e partic: larves A dew ventouses. Ann. Inst. O h o g r . , Paris, 52 (2) (1976) 259-28 1.
Lane, D.J.W., Attachment of the larva of the ascidian Diplosoma listerium. Mar. Biol., 21 (1973) 47-58.
Levine, E.P., Studies on the structure, reproduction, development and accumulation of metals in the colonial ascidian Eudistoma ritteri Van Name, 1945. J. Morphol., 1 11 (1962) 105-138.
Luft, J:H., Improvements in epoxy resin embedding methods. J. Bioph. Biochem. Cytol., 9 (1961) 409427.
Millar, R.H., The biology of ascidians. Adv. Mar. Biol., 9 (1971) 1-100.
Reynolds, E.S., The use of lead citrate at hi'gh pH as an electron opaque stain in electron microscopy. J. Cell Biol., 17 (1963) 208-212.
Sotgia, C., Fascio, U., Melone , G. and De Bernardi, F., Adhesive papillae of Phallusia mammillata larvae: morphology and innervation. Zool. Sci., 15 (1998) 363- 370.
Takamura, K., Nervous network in larvae of the ascidian Ciona intestinalis. Dev. Genes Evol., 208 (1998) 1-8.
Torrence, S.A., Ascidian larval pervous system: anatomy, ultrastructure and metamorphosis. Doctoral Thesis, University of Washington, 1983.
Torrence, S.A. and'cloney, R.A., Rhythmic contractions of the anpullar epidermis during metamorphosis of the ascidian Molgula occidentalis. Cell Tiss. Res., 216 (1981) 293-3 12.
Torrence, S.A. and Cloney, R.A., Ascidian larval nervous system: primary sensory neurons in adhesive papillae. Zoomorphology, 102 (1983) 1 1 1-123.
Trason, W.B., Larval structure and development of the oozooid in the ascidian Euherdmania claviformis. J. Morph., 100 (1957) 509-545.
Turon, X., Morphology of the adhesive papillae of some ascidian larvae. Cah. Biol. Mar., 32 (1991) 295-309.
Dow
nloa
ded
by [
New
Yor
k U
nive
rsity
] at
22:
15 1
1 N
ovem
ber
2014





























