Embed Size (px)
Citation preview

0014-2980/02/1212-3638$17.50+.50/0 © 2002 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
Adhesion of dendritic cells derived from CD34+
progenitors to resting human dermal microvascularendothelial cells is down-regulated uponmaturation and partially depends on CD11a–CD18,CD11b–CD18 and CD36
Van Anh Nguyen1, Susanne Ebner1, Christina Fürhapter1, Nikolaus Romani1, DieterKölle2, Peter Fritsch1 and Norbert Sepp1
1 Department of Dermatology and Venereology, University of Innsbruck, Innsbruck, Austria2 Department of Gynaecology and Obstetrics, University of Innsbruck, Innsbruck, Austria
DC are sentinels of the immune system. In order to reach the skin, bone-marrow-derived DCprecursors need to bind and migrate through microvascular endothelial cells. Binding of DCto primary endothelial cells of the skin has not been investigated. We therefore determinedadhesion of DC at different stages of development to human dermal microvascular endothe-lial cells (HDMEC). DC were derived from CD34+ progenitors in cord blood. To enhance DCmaturation, a defined cocktail of IL-1 g +IL-6+TNF- § +PGE2 was applied. Adhesion was quan-tified by fluorimetric and phase-contrast microscopical assays. Significantly more DC pre-cursors (tested on day 5 after isolation) than mature DC (spontaneously matured orcytokine-cocktail-matured and tested on day 13) bound to unstimulated HDMEC. In con-trast, the maturation stage of DC had no influence on their binding to human umbilical veinendothelial cells. Pretreatment of HDMEC with TNF- § and IFN- + resulted in an enhancedattachment of both DC precursors and mature DC. Mature DC lacked expression of CD31,CD36, CD45RA and CLA, and expressed lower levels of CD11a, CD11b and CD49d as com-pared with precursors tested on day 5. mAb against CD18, CD11a, CD11b, and CD36 mark-edly inhibited DC binding, whereas anti-CLA, anti-DC-SIGN, anti-CD29 and anti-CD49 mAbdid not. Our data support the hypothesis of immunosurveillance with selective recruitment ofblood DC precursors to resting and, more so, to inflamed skin. The data have potential rele-vance for anti-cancer immunotherapy strategies favoring the intracutaneous application ofmature DC.
Key words: Dendritic cell / Microvascular endothelial cell / Adhesion molecule / Immunosurveil-lance / Immunotherapy
Received 16/1/02Revised 12/9/02Accepted 14/10/02
[I 22839]
Abbreviations: EC: Endothelial cells HDMEC: Human der-mal microvascular EC HUVEC: Human umbilical vein endo-thelial cells
1 Introduction
DC are professional APC that are specialized for the initi-ation of primary T cell responses [1, 2]. They originatefrom stem cells in bone marrow and migrate as precur-sors through the bloodstream to nonlymphoid tissues,where they reside in an immature state such as Langer-hans cells in the epidermis and dermal DC in the dermis[3]. These immature DC are characterized by very highantigen-uptake and -processing activities, but low T cell
stimulatory capacity. After an antigenic challenge and inresponse to inflammatory stimuli, they leave the periph-ery via afferent lymph vessels to reach the lymph nodesor spleen T cell areas. At these sites, they efficiently pre-sent antigen to naive T cells and induce an antigen-specific T cell response. At the same time, the migratorycells undergo maturational changes that involve the lossof antigen-capturing capacity and the up-regulation ofMHC, adhesion and costimulatory molecules. Thus,mature DC are equipped with properties that arerequired to stimulate T cells [4–6].
Currently, it is possible to generate large quantities ofhuman DC either from monocytes in peripheral bloodupon culture with GM-CSF and IL-4 [7, 8], or from rareCD34+ progenitors in cord blood or bone marrow by
3638 Van A. Nguyen et al. Eur. J. Immunol. 2002. 32: 3638–3650

Fig. 1. Visualization of DC binding to confluent HDMECmonolayers grown on transwells. Calcein-AM-labeled DCappear in bright green; EC were visualized by biotinylatedanti-human CD31 antibodies and Texas-Red-conjugatedstreptavidin (red). Note also the integrity of the EC mono-layer.
applying GM-CSF and TNF- § [9]. Under the influence ofGM-CSF and TNF- § , CD34+ progenitors delineate amixed cell population that, at early time points, containstwo different subsets of DC. One precursor subset has aphenotype of CD1a+CD14– and are predetermined tohome to the epidermis, where they develop into Langer-hans cells that are characterized by the expression of E-Cadherin, Langerin/CD207 and Birbeck granules. Theprecursor subset with the opposite phenotype(CD1a–CD14+) gives rise to dermal and, more generally,interstitial DC that are characterized by the lack of E-Cadherin, Langerin and Birbeck granules [9].
Despite substantial information about the orchestrationof the traffic of DC from the peripheral tissues to second-ary lymphoid organs [4, 10, 11], the mechanisms under-lying the immigration of circulating DC precursors to theskin remain poorly understood. Before DC extravasatefrom the circulation to their tissue of terminal residence,they have to cross the endothelial lining of the blood ves-sels. In fact, the transmigration of DC is likely to occur asa multistep process, as has been proposed for other leu-kocytes [12].
The number of reports attempting to delineate the inter-actions between DC and endothelial cells (EC) is limited.It is important to emphasize that most functional studieshave been performed with fully differentiated monocyte-derived DC and large EC, i.e. human umbilical cord veinEC (HUVEC) [13–16]. However, none of these studies hasexamined the binding properties of undifferentiated DCprecursors to dermal microvessels. Therefore, we haveaddressed the question of how skin DC precursors inter-act with dermal microvascular EC and how this interac-tion is dependent on the developmental stages of DC. Inorder to establish an in vitro model that imitates in vivoskin pathophysiology, we have used, on the one hand,distinct developmental stages of DC grown fromenriched human cord blood CD34+ precursors and, onthe other hand, human dermal microvascular EC(HDMEC) isolated from human foreskins. Herein weexamine the critical adhesion molecules involved in thisprocess.
2 Results
2.1 Establishment of a DC adhesion assay
Since the fluorescence of calcein AM is dependent onthe metabolic activity of the cells to be labeled, the opti-mal concentration needed to be determined specificallyfor DC. To this end, different numbers of DC (0–1×105)were labeled with calcein AM at different concentrations(1–22 ? M) and immediately thereafter (i.e. without adhe-
sion onto EC) fluorescence was measured in the fluores-cence microtiter plate reader. In these preliminary experi-ments a concentration of 18 ? M was found optimal andwas used for all further experiments. Maximal sensitivityof the assay was achieved at a DC number of 1×105.Labeled DC (1–2×105) were therefore used for all furtherassays. Finally, the optimal conditions for DC binding toHDMEC were determined by allowing DC to adhere fordifferent periods of time (0.5, 1.5, 2.0 and 4.0 h). Time-course analyses were evaluated under the phase con-trast microscope. They revealed that DC bindingbecame clearly apparent after 30 min (20% binding) andreached a maximum of 32–33% at 1.5–2.0 h. No furtherincrease of DC binding was noted upon prolonged incu-bation times. In view of these results, all subsequentadhesion experiments were done with a standard 2 hadhesion period. Phase contrast analyses also con-firmed the integrity of the EC monolayers (Fig. 1).
2.2 Adherence of DC to resting, unstimulatedHDMEC is dependent on the stage ofdifferentiation and independent of thepresence of FCS
The study was designed to investigate the binding prop-erties of DC differentiated from human cord-blood-derived CD34+ hematopoietic progenitors to unstimu-lated HDMEC. Fluorimetric adhesion assays demon-strated that DC precursors tested on day 5 (day 5 DC)displayed a higher adherence to unstimulated HDMECwhen compared with mature (day 13) DC. Whereas amean of 37.8±9.7% day 5 DC precursors bound to un-
Eur. J. Immunol. 2002. 32: 3638–3650 Dendritic cell adhesion to dermal endothelial cells 3639

Fig. 2. Comparative adherence of DC precursors on day 5 (d5) and mature DC on day 13 (d13). Calcein-AM-labeled (A, C) andunlabeled (B) DC were co-incubated with endothelial monolayers for 2 h at 37°C. Adherent DC were then measured using a fluo-rescence microtiter plate reader (A, C) or counted manually under phase contrast microscopy (B). To enhance DC maturation, DCwere additionally exposed to the defined IL-1 g +IL-6+TNF- § +PGE2 cocktail from d11–d13 (C). Note that such cytokine-cocktail-matured DC failed to adhere to unstimulated HDMEC. Results are expressed as the percentage of added DC that adhered. Dataare from five independent experiments in (A), nine in (B) and five in (C). Differences between d5 and d13 were statistically signifi-cant at *p X 0.004, **p X 0.001, using Student’s t-test.
stimulated HDMEC, only a small percentage of sponta-neously matured day 13 DC (13.8±6.3%) were adherentto HDMEC (Fig. 2A). Results obtained from observationsunder the phase contrast microscope were consistentwith those obtained from fluorimetric adhesion assays(Fig. 2B).
During the daily routine inspections of DC cultures wenoted that spontaneously matured day 13 DC popula-tions were not homogeneously mature. Instead, at thistime-point two populations had developed, one withmedium-sized cells, corresponding to immature DC, andthe other with larger cells, corresponding to mature DC.These observations were confirmed in the FACS analy-ses: Day 13 populations contained both CD83+ cells (i.e.mature DC) and CD83– cells (i.e. immature DC). Oncytospins of day 13 DC this became apparent, too: asubstantial proportion of HLA-DR+ cells (i.e. DC) did notexpress the maturation marker DC-LAMP. In an attemptto generate a population of homogeneously mature DCwe used the defined maturation cocktail consisting of IL-1 g , IL-6, TNF- § and PGE2 for the last two days of culture.Such cytokine-cocktail-matured DC were indeed homo-geneously mature (see below) and they were even lessadhesive to HDMEC. Virtually no DC (2.5±0.87%)adhered to unstimulated HDMEC (Fig. 2C).
Adhesion properties of DC cultured in the absence ofFCS were identical to what we have observed in the FCS-containing system: again, significantly more immatureDC (74.6%±10.7%) than cytokine-cocktail-matured DC(14%±15.5) (Fig. 3) bound to HDMEC. Compared withDC cultured in FCS, FCS-free CD34+-derived DC demon-strated a generally increased binding capacity (Fig. 2, 3).
2.3 Adherence of CD34+-derived DC to resting,unstimulated HUVEC differs from that toHDMEC
When compared with HDMEC, fewer day 5 DC bound toHUVEC, and the adhesion of DC to HUVEC was inde-pendent of the stage of DC differentiation since a highpercentage of day 13 DC still bound to HUVEC. The per-centage of binding to HUVEC was similar in both popula-tions (46±6.5% versus 38.3±23.8%) (Fig. 3).
2.4 Pretreatment of EC monolayers withcytokines results in an enhanced attachmentof DC, irrespective of their state ofdifferentiation
Stimulation of endothelium with 100 U/ml TNF- § for 24 hor with 500 U/ml IFN- + for 72 h led to a significantincrease in the percentage of adhesion of both day 5 DCprecursors and day 13 cytokine-cocktail-matured DC.The attachment of day 5 DC to TNF- § - or IFN- + -stimulated HDMEC was enhanced by approximately50% when compared with unstimulated HDMEC(p X 0.001): Binding of day 5 DC to TNF- § -stimulatedHDMEC was 43.75±1.48% and that to IFN- + -stimulatedHDMEC was 43.50±5.52%. Binding of day 5 DC tounstimulated HDMEC, in contrast, was only27.25±1.25%. As described above, day 13 cytokine-cocktail-matured DC failed to adhere to unstimulatedHDMEC. However, when HDMEC were stimulated withTNF- § and IFN- + , the adherence of day 13 cytokine-cocktail-matured DC increased from 2.50±0.87% to16.75±1.79% and to 14.00±0.50%, respectively (Fig. 4).
3640 Van A. Nguyen et al. Eur. J. Immunol. 2002. 32: 3638–3650

Fig. 3. Different adhesion properties of CD34+-derived-DCcultured in FCS-free medium to HDMEC and HUVEC,assessed using fluorimetric adhesion assays. When com-pared with HDMEC, the adhesion of day 5 (d5) DC precur-sors to HUVEC was decreased and adhesion to HUVEC wasindependent of the stage of DC differentiation. Data are fromthree independent experiments.
Fig. 4. Effect of TNF- § - or INF- + -stimulated HDMEC on theadherence of DC precursors on day 5 (d5) and of cytokine-cocktail-matured day 13 (d13) DC. Confluent endothelialmonolayers were either left untreated (open columns), orprestimulated with INF- + (hatched columns) for 72 h or withTNF- § (closed columns) for 24 h. Results are expressed aspercentage of adherent cells and show mean values of oneexperiment carried out in duplicates. Similar values wereobtained in a replicate experiment. Note that the adherenceof both d5 DC precursors and terminally mature d13 DC wasenhanced in response to activation of the endothelium. Thedifferential adherence properties of the two DC populationpersisted, however. Differences were statistically significantat *p X 0.01, **p X 0.02, ***p X 0.04, using Student’s t-test,compared with DC attachment to untreated monolayers.
Fig. 5. Expression of DC-LAMP/CD208 and CD68 by cord-blood-derived DC. Cytospins of cord-blood-derived DC pre-cursors on day 5 (A, C), spontaneously matured (E) andcytokine-cocktail-derived DC (B, D, F) on day 13, wereimmunostained with mAb against DC-LAMP (A, B, E, F) orCD68 (C, D) and show red fluorescence. Counterstainingwith anti-HLA-DR–FITC appears as green fluorescence.Specimens were viewed on a conventional fluorescencemicroscope with a filter allowing the simultaneous visualiza-tion of both fluorescent colors. Note that developing DC onday 5 express absolutely no DC-LAMP (A) but display asome CD68 staining scattered all over the cytoplasm andapparently co-localizing with HLA-DR, therefore yielding yel-low fluorescence (C). Mature DC show a virtually identicalstaining pattern with both DC-LAMP (B) and CD68 (D): redfluorescence is largely confined to a perinuclear area; HLA-DR is expressed at the cell surface membrane (green). Thepopulation of spontaneously matured DC on day 13 stillcontains sizeable numbers of DC-LAMP–, i.e. immature, DC(E). In contrast, after treatment with a cytokine cocktail, DCare mature as indicated by their uniform expression of DC-LAMP (F). Magnification 390×. Bar=30 ? m.
2.5 Phenotype of DC — definition of maturationalstate by means of specific markers
Analyses on cytospins revealed that mature DCexpressed high levels of DC-LAMP, as discussed below;precursors on day 5 were totally negative for this marker
(Fig. 5A). CD68 was expressed to a low degree by day 5precursors (Fig. 5C); upon maturation expression levelsincreased and the staining was found in a perinuclearlocation, similar to DC-LAMP (Fig. 5B, D).
DC-LAMP expression in spontaneously matured DC onday 13 was never found in all cells, but in varying propor-tions of the populations. For the adhesion assays, how-
Eur. J. Immunol. 2002. 32: 3638–3650 Dendritic cell adhesion to dermal endothelial cells 3641
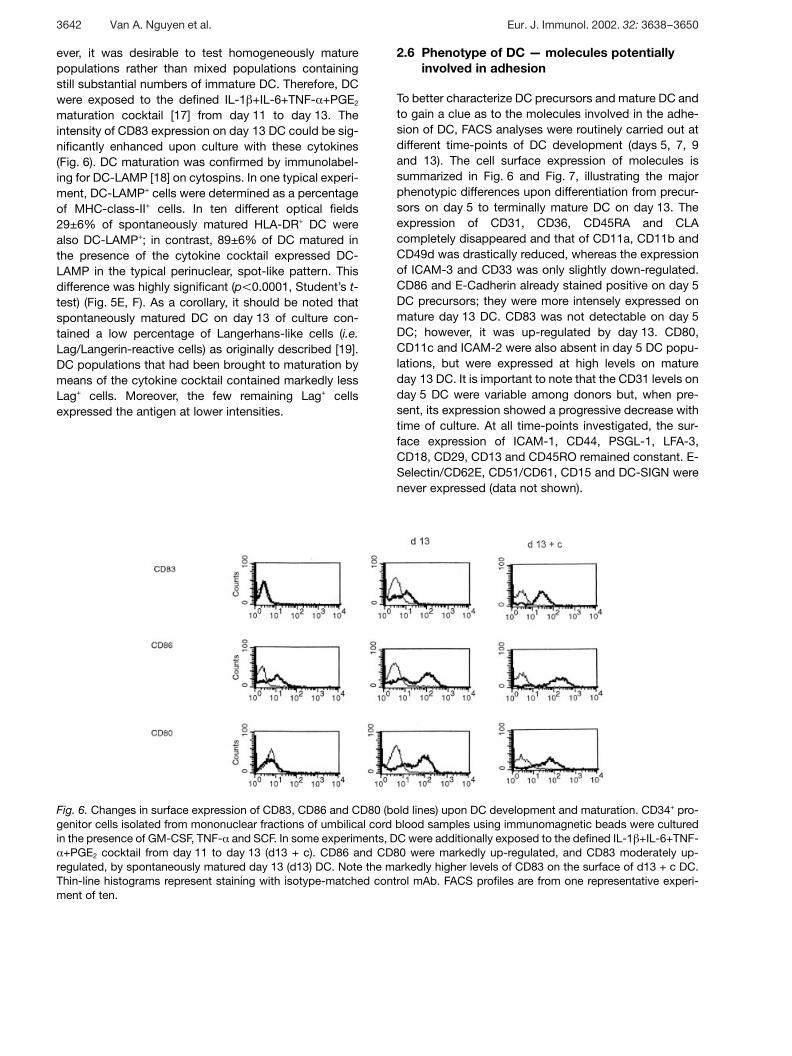
Fig. 6. Changes in surface expression of CD83, CD86 and CD80 (bold lines) upon DC development and maturation. CD34+ pro-genitor cells isolated from mononuclear fractions of umbilical cord blood samples using immunomagnetic beads were culturedin the presence of GM-CSF, TNF- § and SCF. In some experiments, DC were additionally exposed to the defined IL-1 g +IL-6+TNF-§ +PGE2 cocktail from day 11 to day 13 (d13 + c). CD86 and CD80 were markedly up-regulated, and CD83 moderately up-
regulated, by spontaneously matured day 13 (d13) DC. Note the markedly higher levels of CD83 on the surface of d13 + c DC.Thin-line histograms represent staining with isotype-matched control mAb. FACS profiles are from one representative experi-ment of ten.
ever, it was desirable to test homogeneously maturepopulations rather than mixed populations containingstill substantial numbers of immature DC. Therefore, DCwere exposed to the defined IL-1 g +IL-6+TNF- § +PGE2
maturation cocktail [17] from day 11 to day 13. Theintensity of CD83 expression on day 13 DC could be sig-nificantly enhanced upon culture with these cytokines(Fig. 6). DC maturation was confirmed by immunolabel-ing for DC-LAMP [18] on cytospins. In one typical experi-ment, DC-LAMP+ cells were determined as a percentageof MHC-class-II+ cells. In ten different optical fields29±6% of spontaneously matured HLA-DR+ DC werealso DC-LAMP+; in contrast, 89±6% of DC matured inthe presence of the cytokine cocktail expressed DC-LAMP in the typical perinuclear, spot-like pattern. Thisdifference was highly significant (p X 0.0001, Student’s t-test) (Fig. 5E, F). As a corollary, it should be noted thatspontaneously matured DC on day 13 of culture con-tained a low percentage of Langerhans-like cells (i.e.Lag/Langerin-reactive cells) as originally described [19].DC populations that had been brought to maturation bymeans of the cytokine cocktail contained markedly lessLag+ cells. Moreover, the few remaining Lag+ cellsexpressed the antigen at lower intensities.
2.6 Phenotype of DC — molecules potentiallyinvolved in adhesion
To better characterize DC precursors and mature DC andto gain a clue as to the molecules involved in the adhe-sion of DC, FACS analyses were routinely carried out atdifferent time-points of DC development (days 5, 7, 9and 13). The cell surface expression of molecules issummarized in Fig. 6 and Fig. 7, illustrating the majorphenotypic differences upon differentiation from precur-sors on day 5 to terminally mature DC on day 13. Theexpression of CD31, CD36, CD45RA and CLAcompletely disappeared and that of CD11a, CD11b andCD49d was drastically reduced, whereas the expressionof ICAM-3 and CD33 was only slightly down-regulated.CD86 and E-Cadherin already stained positive on day 5DC precursors; they were more intensely expressed onmature day 13 DC. CD83 was not detectable on day 5DC; however, it was up-regulated by day 13. CD80,CD11c and ICAM-2 were also absent in day 5 DC popu-lations, but were expressed at high levels on matureday 13 DC. It is important to note that the CD31 levels onday 5 DC were variable among donors but, when pre-sent, its expression showed a progressive decrease withtime of culture. At all time-points investigated, the sur-face expression of ICAM-1, CD44, PSGL-1, LFA-3,CD18, CD29, CD13 and CD45RO remained constant. E-Selectin/CD62E, CD51/CD61, CD15 and DC-SIGN werenever expressed (data not shown).
3642 Van A. Nguyen et al. Eur. J. Immunol. 2002. 32: 3638–3650
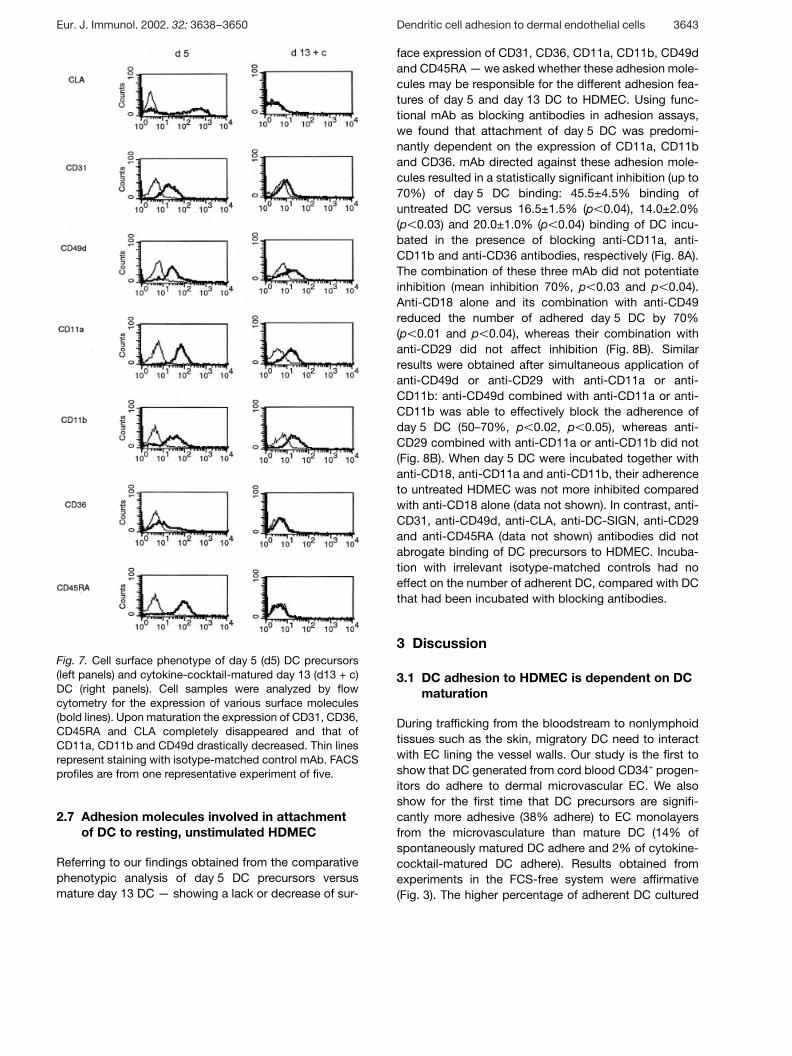
Fig. 7. Cell surface phenotype of day 5 (d5) DC precursors(left panels) and cytokine-cocktail-matured day 13 (d13 + c)DC (right panels). Cell samples were analyzed by flowcytometry for the expression of various surface molecules(bold lines). Upon maturation the expression of CD31, CD36,CD45RA and CLA completely disappeared and that ofCD11a, CD11b and CD49d drastically decreased. Thin linesrepresent staining with isotype-matched control mAb. FACSprofiles are from one representative experiment of five.
2.7 Adhesion molecules involved in attachmentof DC to resting, unstimulated HDMEC
Referring to our findings obtained from the comparativephenotypic analysis of day 5 DC precursors versusmature day 13 DC — showing a lack or decrease of sur-
face expression of CD31, CD36, CD11a, CD11b, CD49dand CD45RA — we asked whether these adhesion mole-cules may be responsible for the different adhesion fea-tures of day 5 and day 13 DC to HDMEC. Using func-tional mAb as blocking antibodies in adhesion assays,we found that attachment of day 5 DC was predomi-nantly dependent on the expression of CD11a, CD11band CD36. mAb directed against these adhesion mole-cules resulted in a statistically significant inhibition (up to70%) of day 5 DC binding: 45.5±4.5% binding ofuntreated DC versus 16.5±1.5% (p X 0.04), 14.0±2.0%(p X 0.03) and 20.0±1.0% (p X 0.04) binding of DC incu-bated in the presence of blocking anti-CD11a, anti-CD11b and anti-CD36 antibodies, respectively (Fig. 8A).The combination of these three mAb did not potentiateinhibition (mean inhibition 70%, p X 0.03 and p X 0.04).Anti-CD18 alone and its combination with anti-CD49reduced the number of adhered day 5 DC by 70%(p X 0.01 and p X 0.04), whereas their combination withanti-CD29 did not affect inhibition (Fig. 8B). Similarresults were obtained after simultaneous application ofanti-CD49d or anti-CD29 with anti-CD11a or anti-CD11b: anti-CD49d combined with anti-CD11a or anti-CD11b was able to effectively block the adherence ofday 5 DC (50–70%, p X 0.02, p X 0.05), whereas anti-CD29 combined with anti-CD11a or anti-CD11b did not(Fig. 8B). When day 5 DC were incubated together withanti-CD18, anti-CD11a and anti-CD11b, their adherenceto untreated HDMEC was not more inhibited comparedwith anti-CD18 alone (data not shown). In contrast, anti-CD31, anti-CD49d, anti-CLA, anti-DC-SIGN, anti-CD29and anti-CD45RA (data not shown) antibodies did notabrogate binding of DC precursors to HDMEC. Incuba-tion with irrelevant isotype-matched controls had noeffect on the number of adherent DC, compared with DCthat had been incubated with blocking antibodies.
3 Discussion
3.1 DC adhesion to HDMEC is dependent on DCmaturation
During trafficking from the bloodstream to nonlymphoidtissues such as the skin, migratory DC need to interactwith EC lining the vessel walls. Our study is the first toshow that DC generated from cord blood CD34+ progen-itors do adhere to dermal microvascular EC. We alsoshow for the first time that DC precursors are signifi-cantly more adhesive (38% adhere) to EC monolayersfrom the microvasculature than mature DC (14% ofspontaneously matured DC adhere and 2% of cytokine-cocktail-matured DC adhere). Results obtained fromexperiments in the FCS-free system were affirmative(Fig. 3). The higher percentage of adherent DC cultured
Eur. J. Immunol. 2002. 32: 3638–3650 Dendritic cell adhesion to dermal endothelial cells 3643

Fig. 8. Molecules involved in adhesion of immature day 5 DC to resting HDMEC. Prior to the start of the adhesion assay, DC werepretreated with blocking antibodies recognizing CD31, CLA, CD11a, CD11b, CD36, a combination of antibodies to CD11a,CD11b, and/or CD36 (A), CD49d, or a combination of antibodies to CD18, CD11a, CD11b and CD29 or CD49d (B). All mAb wereused at a concentration of 10 ? g/ml and were additionally present throughout the assay. In (A), results are expressed as percent-ages of adherent cells and show mean values of one representative experiment carried out in duplicates. Similar values wereobtained in a replicate experiment. In (B), data are from three independent experiments. Differences were statistically significantat *p X 0.03, **p X 0.04, +p X 0.02, ++p X 0.03, +++p X 0.04, ++++p X 0.05 compared with controls, using Student’s t-test.
in the absence of FCS might be explained by the factthat fewer DC mature under FCS-free conditions. Flowcytometric analysis revealed a very similar phenotype ofthese FCS-free CD34+-derived DC to those recentlydescribed by Banchereau et al. [20] (data not shown). Inaccordance with previous studies the percentage of DCbound to HUVEC was low [13–15] and, more importantly,the adhesion to HUVEC did not depend on the matura-tion stage of DC (Fig. 3). Our findings underline the well-known differences between small-vessel and large-vessel EC [21].
Spontaneously matured DC populations on day 13 con-tained still abundant immature cells, regardless ofmaturation-promoting TNF- § present in the cultures. Wetherefore questioned whether these remaining immature(or not yet mature) DC were responsible for the residualbinding of 14% evident on day 13 in populations ofspontaneously matured DC. In order to render all DC fullymature on day 13 we treated DC with a defined cocktailconsisting of IL-1 g , IL-6, TNF- § and PGE2 for the lasttwo days of culture. Similar to what was recentlyreported by Jonuleit et al. [17] for monocyte-derived DC,CD34+-progenitor-derived DC also became homoge-neously mature as determined by their uniform expres-sion of CD83 and DC-LAMP (Fig. 5, 6). These DC exhib-ited virtually no attachment to resting HDMEC. We there-fore conclude that the low degree of adhesion observedin populations of spontaneously matured DC on day 13is likely due to “contaminating” DC precursors or imma-ture DC. At the single-cell level a terminally mature DCdoes not adhere to HDMEC.
3.2 DC adhesion to stimulated versusunstimulated HDMEC
Stimulation of HDMEC with TNF- § and IFN- + resulted ina significantly increased percentage of adherent DC(Fig. 4), consistent with the literature [13, 14]. In this con-text, it is noteworthy that cytokine-activated HDMECalso induced an enhanced attachment of cytokine-cocktail-matured DC. As mentioned above, such matureDC filed to adhere to unactivated HDMEC. Apparently,this effect might be caused by the elevated surfaceexpression of endothelial adhesion molecules suchas ICAM-1, VCAM-1 and E-Selectins involved inleukocyte–EC interactions, as shown for HUVEC [22–24]as well as for HDMEC [25].
Our finding that DC precursors adhere to dermal micro-vascular EC and that this attachment is increased inresponse to inflammatory cytokines supports thehypothesis of immunosurveillance as previously empha-sized by Robert et al. [26]. Using an in vivo imaging sys-tem, they have found that blood DC precursors constitu-tively tether and roll via P-Selectin glycoprotein ligand(PSGL)-1 on P- and E-Selectins — expressed at low lev-els on capillary EC and up-regulated by inflammatorystimuli — which contributes to the rapid recruitment ofDC to sites of cutaneous inflammation, resulting in anenhancement of local antigen capture and presentation.Independent confirmation has recently been provided byFerrero et al. [27] showing that immature DC preferen-tially invaded the extracellular matrix and reverse-transmigrated across endothelial monolayers using theurokinase plasminogen activator receptor (uPAR).
3644 Van A. Nguyen et al. Eur. J. Immunol. 2002. 32: 3638–3650

3.3 Molecules involved in the adhesion of DC
To examine the specific adhesion molecules operative inDC binding to HDMEC, we carried out a detailed pheno-typic characterization of both day 5 DC precursors andmature day 13 DC with and without application of thematuration-inducing cytokine cocktail. Remarkably,these FACS analyses demonstrated that mature DClacked the expression of CD31, CD36, CD45RA andCLA, and expressed CD11a, CD11b and CD49d at verylow levels. Therefore, we chose to study the role of theseparticular molecules by means of blocking antibodies.
DC-SIGN is an adhesion molecule that is specificallyexpressed on monocyte-derived DC, dermal DC, and DCfrom peripheral blood. It mediates contact of monocyte-derived DC to T cells via ICAM-3 [28] and to umbilicalcord EC via ICAM-2 [29]. Interestingly, neither DC precur-sors on day 5 nor mature DC on day 13 expressed thismolecule on their surface. Monocyte-derived DC thatwere used as controls were brightly DC-SIGN+. Blockingwith anti-DC-SIGN antibodies, therefore, did not haveany influence on the binding properties of CD34+-derivedDC to HDMEC (data not shown).
Anti-CD11a, anti-CD11b or anti-CD18 antibodies mark-edly impeded adhesion of DC precursors to HDMEC(Fig. 8A, B). These g 2-integrins apparently interact withICAM-1, which is constitutively expressed on restingHDMEC and is up-regulated by inflammatory stimuli.This would explain the increased number of DC adherentto TNF- § - and IFN- + -stimulated HDMEC. Our results arein accordance with reports published by Brown et al. [13]and D’Amico et al. [14], showing that the CD11a andCD11b expression was necessary for the binding ofmonocyte-derived DC to HUVEC. Obviously, the g 1-integrins did not affect adherence of DC to restingHDMEC as both CD49d and CD29 failed to block bind-ing of DC (Fig. 8A, B).
The simultaneous application of anti- g 2-integrins withanti-CD49d reproduced an inhibition of day 5 DC bindingto HDMEC to a level of anti- g 2-integrins alone (Fig. 8B).Interestingly, when DC were incubated together withantibodies against g 2-integrins and CD29, the blockingeffect of anti- g 2-integrins was abrogated (Fig. 8B). Thisphenomenon might be due to a cross-linking of cellscaused by the binding of anti-CD29 antibodies to inte-grins on the surface on HDMEC, because flow cyto-metric analysis showed a more than tenfold higherexpression of CD29 on HDMEC compared with that ofCD49d (data not shown).
In spite of some residual expression of CD11a andCD11b, mature DC did not adhere to resting HDMEC.
Probably the expression of CD11 molecules on DC andof their binding partners on HDMEC is too low to allow afirm binding that would resist the necessary rinsing stepsfor the adhesion assay. It is also tempting to speculatethat the activation state of these integrins is modulatedupon DC maturation so that in spite of low level expres-sion on mature DC they cannot bind to appropriateligands on HDMEC because of their inactive state.
Blocking-studies with anti-CD36 antibodies have not yetbeen described. Our data suggest that CD36 — a mole-cule known to be essential in the binding of EC to throm-bospondin [30] and to Plasmodium-falciparum-infectederythrocytes [31] — might play a novel and essential rolein the binding of DC to microvascular EC. In this respect,it is of note that only HDMEC are known to express CD36on their surface, in contrast to HUVEC, which do not butare used in all other published DC–EC adherence stud-ies.
Blocking-antibodies to CLA, unexpectedly, did not influ-ence the DC–HDMEC binding, although this moleculewas differentially expressed on precursors and matureDC. From this, one might assume that CLA is moreresponsible for the rolling and tethering of DC on the sur-face of HDMEC rather than for the tight attachment toHDMEC [26]. Flow cytometric analysis of bound andunbound DC yielded a similar percentage of CLA expres-sion of both fractions, indicating that the CLA+ and CLA–
DC subsets did not exhibit a distinct binding behavior toHDMEC (data not shown).
It is of note that simultaneous application of antibodiesto CD11a, CD11b, CD18 and/or CD36 did not producean additive inhibition (Fig. 8A). We therefore hypothesizethat CD11a, CD11b and CD36 alone can not be solelyresponsible for the attachment of DC precursorsbecause antibodies directed against these adhesionmolecules never reduced the binding to 0% as observedfor mature DC. It is likely that other, yet-unknown mole-cule(s) [32] are involved in adhesion of DC precursors toHDMEC.
3.4 Relevance for DC-based immunotherapy
Apart from their relevance to the mechanisms of replen-ishing the pool of tissue-residing immature DC, our find-ings might have potential relevance in the developmentof immunotherapy strategies that use antigen-chargedDC. The optimal immunization strategy is not yet clear.The maturation stage of DC and the route of administra-tion both appear to be critical. Mature DC are preferableto immature ones in immunization strategies for a num-ber of reasons [33]. Our findings provide yet another
Eur. J. Immunol. 2002. 32: 3638–3650 Dendritic cell adhesion to dermal endothelial cells 3645

important reason for using mature DC, especially whenapplied intracutaneously. If immature DC are injected,they would display a high tendency to adhere to micro-vascular EC of blood vessels, which may distract manyof them from entering lymph vessels and thus from gen-erating immunity in the lymph nodes. The failure ofmature DC to adhere to microvascular blood EC, in turn,would guarantee their unimpaired entry into the lymphat-ics and migration to the lymph nodes [34], where theythen activate antigen-specific T cells. Because intracuta-neously injected DC interact with EC lining lymph ves-sels during their migration from the skin to afferent lymphnodes [34, 35], it would be also interesting to examinewhether the adhesion properties and mechanisms ofCD34+-derived DC with HDMEC are the same for purifieddermal lymphatic EC [36]. Further detailed investigationsare required to confirm these hypotheses.
4 Materials and methods
4.1 Generation of CD34+-derived DC
DC were generated from CD34+ progenitor cells enrichedfrom mononuclear fractions of umbilical cord blood samplesusing immunomagnetic beads (Miltenyi Biotech, BergischGladbach, Germany) according to the manufacturer’s proto-col. The mean CD34+ cell purity was shown to be 85%(range 70–99%) by flow cytometry. The isolated cells werecryopreserved in 10% DMSO (Merck, Darmstadt, Germany).After thawing, cultures of CD34+ cells were established inculture flasks (Greiner, Frickenhausen, Germany) by seedingat 4×104 cells/ml in RPMI 1640 medium supplemented with10% heat-inactivated FCS, 2 mM L-glutamine (SigmaChemicals, St. Louis, MO), 100 U/ml penicillin, 100 ? g/mlstreptomycin (Irvine Scientific, Santa Ana, CA), 10 mMHepes (Biochrom KG, Berlin, Germany), rhGM-CSF (100 ng/ml, 200 U/ml; LeukomaxTM, Novartis, Basel, Switzerland),rhTNF- § (2.5 ng/ml, 50 U/ml, specific activity 1×108 U/mg;generously provided by Dr. G. R. Adolf, Bender, Vienna, Aus-tria), and rhSCF (25 ng/ml, specific activity 5×105 U/mg;PeproTech, London, UK) as described previously [9, 19]. Tomaintain optimal culture conditions, changes of media withfresh GM-CSF and TNF- § were performed at day 4 andday 10. For most experiments, cells were harvested atdays 5 (“DC precursors”) and 13 (“spontaneously maturedDC”).
4.2 Enhancement of DC maturation by a definedcytokine cocktail (“cytokine-cocktail-matured DC”)
Maturation of DC was enhanced by adding a cocktail ofinflammatory cytokines (IL-1 g at 2 ng/ml, IL-6 at 1000 U/ml,and TNF- § at 10 ng/ml final concentrations) and PGE2 at1 ? g/ml to the cultures from day 11 to day 13. This cocktailhas been described to lead to a reproducible terminal matu-
ration of the vast majority of DC in a given population [17].IL-1 g (specific activity 1×107 U/mg) and IL-6 (specific activ-ity 1×107 U/mg) were purchased from PeproTech. TNF- §(specific activity 6×107 U/mg) was generously provided byDr. G. R. Adolf. PGE2 (Prostin ETM) was from Pharmacia-Upjohn, Peapack, NJ.
4.3 Generation of CD34+-derived DC under FCS-freeconditions
CD34+ cells were purified from human cord blood asdescribed above and cultured in a special DC medium(CellGro DCTM from CellGenix Corporation, Freiburg, Ger-many) containing 1% autologous serum, 1% non-essentialamino acids (Gibco, Paisley, Scotland), 2 mM L-glutamine(Sigma Chemicals), 100 U/ml penicillin, 100 ? g/ml strepto-mycin (Irvine Scientific), and 10 mM Hepes (Biochrom KG).Cytokines and maturation stimuli were identical to FCS-containing cultures (see above).
4.4 Preparation of HDMEC and HUVEC
HDMEC were isolated from surgically removed normal fore-skins obtained from newborns, and children up toseven years old, according to a modification of a previouslydescribed standard technique [37]. Briefly, foreskins thathad been stored less than 15 h at 4°C were first washed in70% ethanol and then rinsed in Hank’s buffered salt solution(HBSS; Gibco, Paisley, Scotland). Thereafter, skin fragmentswere cut into small pieces and, instead of trypsinization,incubated overnight in 1.2 U dispase II (Boehringer-Mannheim, Mannheim, Germany) per ml of Puck’s solution(Sigma Chemicals) at 4°C to separate the epidermis from thedermis. For the HDMEC isolation only the dermal layer wasused. Cells were plated in tissue culture flasks (Greiner) thathad been coated with 1% gelatin (Sigma Chemicals), andcultured in EC basal medium (EBM; Clonetics Corp., Wal-kersville, MD), supplemented with 10% normal humanserum, 5 ng/ml epidermal growth factor (Clonetics Corp.),2 mM L-glutamine, 1 ? g/ml hydrocortisone acetate,5×10–5 M dibutyryl cyclic adenosine monophosphate (SigmaChemicals), 100 U/ml penicillin, 100 ? g/ml streptomycin,and 250 ? g/ml amphotericin B (all purchased from IrvineScientific). The resulting cultures were consistently purewithout contaminating fibroblasts, as assessed by morpho-logical and immunohistochemical criteria.
HUVEC, kindly provided by Dr. G. Wick (Innsbruck, Austria),were isolated and cultured as described earlier [38].
4.5 Quantitative adhesion assay of unlabeled DC toHDMEC using phase contrast microscopy
Adhesion assays were performed with day 5 DC precursorsand day 13 mature DC. In co-culture experiments, HDMEC
3646 Van A. Nguyen et al. Eur. J. Immunol. 2002. 32: 3638–3650

were seeded in 6-well plates and allowed to grow to conflu-ence for 2–3 days of culture. In some experiments, EC werestimulated with 100 U/ml TNF- § or 500 U/ml IFN- + (Pepro-Tech) for 24 h and 72 h, respectively. Resting or stimulatedendothelial monolayers were washed three times withserum-free RPMI 1640 immediately before addition of unla-beled DC (2×105 cells in complete medium) to each well.After an incubation of 2 h at 37°C, non-adherent DC wereremoved by 3–5 gentle washes with serum-free, warm RPMI1640. Plates were checked by phase contrast microscopy toensure that there was no disruption of the endothelial mono-layers. Adherent DC on endothelial surfaces were fixed with2.5% glutaraldehyde and counted by means of a calibratedocular grid under the phase contrast microscope. Using 25×objective lenses, 15 randomly chosen fields were enumer-ated, and mean values and standard deviations were deter-mined. From these values the total number of DC bound tothe surface of one well of a 6-well plate was calculated.
4.6 Quantitative adhesion assay of calcein-AM-labeledDC to HDMEC
To quantify adherent DC, a fluorimetric adhesion assay (Vy-brantTM cell adhesion assay kit, Molecular Probes, Eugene,OR) was used [39, 40]. Before labeling with the fluorogenicdye, calcein AM, DC were washed twice with PBS, resus-pended in RPMI plus 10% FCS but without phenol red andadjusted to 1×106 cells/ml. DC were then labeled at a finalconcentration of 18 ? M for 30 min at 37°C and 5% CO2.Labeling was stopped by adding prewarmed (37°C) RPMIwithout phenol red and serum, and DC were washed twiceby centrifugation at 296×g for 8 min at 4°C and resuspendedto 1×106 cells/ml. Prior to the onset of the adhesion assay,endothelial monolayers grown in 96-well plates (Packard,Canberra Company, Dreieich, Germany) were washed 2–3times with RPMI plus 10% FCS but without phenol red.Labeled DC (1–2×105) were added to each well and co-incubated for 2 h at 37°C. After the 2 h incubation period,unbound DC were removed by gentle washing (3–5 times)with RPMI plus 10% FCS but without phenol red. Finally,each well was scanned by a fluorescence microtiter platereader (Fluoreskan Ascent, Labsystems, Helsinki, Finland) tomeasure the fluorescent signal emitted by the bound DCusing excitation and emission wavelengths of 485 nm and538 nm, respectively. The percentage of cell adhesion wascalculated using the following formula: adhesion % = [(fluo-rescence of adherent DC – background fluorescence)/(fluo-rescence of total DC deposited – background fluores-cence)×100].
4.7 Adhesion assays using monoclonal blockingantibodies
DC (1×106 DC/ml of RPMI plus 10% FCS but without phenolred) were incubated with blocking mAb recognizing CD31,CD36, CD18, CD29, CD11a, CD11b, CD49d (purchased
from Ancell, Bayport, MN), CLA (PharMingen, San Diego,CA) and/or DC-SIGN/CD209 (a generous gift from Dr. Y. vanKooyk, Nijmegen, Netherlands). All mAb were used at a finalconcentration of 10 ? g/ml. Appropriate isotype-matchedIgG served as negative controls in all experiments.
4.8 Flow cytometric analysis of surface molecules onCD34+-derived DC
The panel of mAb listed in Table 1 was used to determine thesurface expression of molecules on day 5 DC precursors,spontaneously matured DC and cytokine-cocktail-maturedday 13 DC. Cells were washed in PBS with 0.5% BSA(Boehringer Ingelheim, Germany), counted and aliquotted at100,000 cells/tube for antibody staining. Unconjugated orFITC-/PE-conjugated antibodies at a concentration of 1 ? g/ml were incubated on ice with the cells. After 30 min incuba-tion, cells were washed in PBS with 0.5% BSA. For indirectfluorescent staining, cells were further incubated with FITC-labeled F(ab)2 fragments of sheep anti-mouse IgG for30 min, washed as above and resuspended in PBS with0.5% BSA. Negative controls were stained with isotype-matched IgG. Surface marker expression was examined in aFACScalibur flow cytometer (Becton Dickinson, MountainView, CA) measuring a total of 5,000–10,000 events per ali-quot.
4.9 Immunohistochemical studies of CD34+-derived DCand HDMEC on transwells and cytospinpreparations
HDMEC were grown to confluence on transwells (Costar)coated with 1% gelatin (Sigma Chemicals). Transwells bear-ing confluent endothelial monolayers with adherent calcein-AM-labeled DC were fixed with 90% ethanol for 10 min andthen incubated with biotinylated anti-human CD31 mAb(Ancell, Bayport, MN) followed by Texas-Red-conjugatedstreptavidin (Amersham-Pharmacia, Amersham, UK). Cellswere mounted in Vectashield (Vector, Burlingame, CA) andviewed on a fluorescence microscope. Binding of mousemAb on acetone-fixed cytospins was visualized by a biotiny-lated anti-mouse-Ig (Amersham-Pharmacia) followed byTexas-Red-conjugated streptavidin. After blocking of resid-ual binding sites with an excess of mouse + -globulin(100 ? g/ml) DC were counterstained with an FITC-conjugated anti-HLA-DR mAb (BD Immunocytometry Sys-tems, San Jose, CA). DC-LAMP/CD208 [18] (mouse IgG1)was a kind gift of Dr. Serge Lebecque, Laboratory for Immu-nological Research, Schering-Plough, Dardilly, France. mAbLag [41] (mouse IgG1), an anti-human-Birbeck-granule-associated Langerin/CD207 was donated by Dr. KozoYoneda, Dermatology, Kyoto University, Kyoto, Japan. Anti-CD68 (clone KP1, mouse IgG1) was purchased from Dako,Glostrup, Denmark.
Eur. J. Immunol. 2002. 32: 3638–3650 Dendritic cell adhesion to dermal endothelial cells 3647

Table 1. mAb used for flow cytometry and antibody blocking studies
Acknowledgements: We thank all midwives from theDepartment of Obstetrics and Gynecology, Innsbruck, andDr. Talasz from the Department of Biochemistry, Innsbruck,for providing umbilical cord samples and the fluorescencemicrotiter plate reader, respectively. We are also grateful toHella Stössel for immunohistochemical stainings on cyto-spins. We thank Dr. Yvette van Kooyk, Nijmegen, The Neth-erlands, for critical reading and helpful discussions. S. Ebneris a member of KMT (Kompetenzzentrum Medizin Tirol). Thiswork was supported by the Austrian National Bank, Jubi-laeumsfonds project No 7881.
References
1 Cella, M., Sallusto, F. and Lanzavecchia, A., Origin, maturationand antigen presenting function of dendritic cells. Curr. Opin.Immunol. 1997. 9: 10–16.
2 Banchereau, J. and Steinman, R. M., Dendritic cells and thecontrol of immunity. Nature 1998. 392: 245–252.
3 Katz, S. I., Tamaki, K. and Sachs, D. H., Epidermal Langerhanscells are derived from cells originating in bone marrow. Nature1979. 282: 324–326.
4 Austyn, J. M., New insights into the mobilization and phagocyticactivity of dendritic cells. J. Exp. Med. 1996. 183: 1287–1292.
3648 Van A. Nguyen et al. Eur. J. Immunol. 2002. 32: 3638–3650

5 Sallusto, F., Cella, M., Danieli, C. and Lanzavecchia, A., Den-dritic cells use macropinocytosis and the mannose receptor toconcentrate macromolecules in the major histocompatibilitycomplex class II compartment: Downregulation by cytokines andbacterial products. J. Exp. Med. 1995. 182: 389–400.
6 Inaba, K., Turley, S., Yamaide, F., Iyoda, T., Mahnke, K., Inaba,M., Pack, M., Subklewe, M., Sauter, B., Sheff, D., Albert, M.,Bhardwaj, N., Mellman, I. and Steinman, R. M., Efficient pre-sentation of phagocytosed cellular fragments on the major histo-compatibility complex class II products of dendritic cells. J. Exp.Med. 1998. 188: 2163–2173.
7 Romani, N., Gruner, S., Brang, D., Kämpgen, E., Lenz, A.,Trockenbacher, B., Konwalinka, G., Fritsch, P. O., Steinman,R. M. and Schuler, G., Proliferating dendritic cell progenitors inhuman blood. J. Exp. Med. 1994. 180: 83–93.
8 Sallusto, F. and Lanzavecchia, A., Efficient presentation of solu-ble antigen by cultured human dendritic cells is maintained bygranulocyte/macrophage colony-stimulating factor plus interleu-kin 4 and downregulated by tumor necrosis factor a. J. Exp. Med.1994. 179: 1109–1118.
9 Caux, C., Massacrier, C., Vanbervliet, B., Dubois, B., Durand,I., Cella, M., Lanzavecchia, A. and Banchereau, J., CD34+
hematopoietic progenitors from human cord blood differentiatealong two independent dendritic cell pathways in response togranulocyte-macrophage colony-stimulating factor plus tumornecrosis factor § . 2. Functional analysis. Blood 1997. 90:1458–1470.
10 McWilliam, A. S., Nelson, D., Thomas, J. A. and Holt, P. G.,Rapid dendritic cell recruitment is a hallmark of the acute inflam-matory response at mucosal surfaces. J. Exp. Med. 1994. 179:1331–1336.
11 Xia, W., Pinto, C. E. and Kradin, R. L., The antigen-presentingactivities of Ia+ dendritic cells shift dynamically from lung tolymph node after an airway challenge with soluble antigen. J.Exp. Med. 1995. 181: 1275–1283.
12 Springer, T. A., Traffic signals for lymphocyte recirculation andleukocyte emigration: the multistep paradigm. Cell 1994. 76:301–314.
13 Brown, K. A., Bedford, P., Macey, M., McCarthy, D. A., Leroy,F., Vora, A. J., Stagg, A. J., Dumonde, D. C. and Knight, S. C.,Human blood dendritic cells: Binding to vascular endotheliumand expression of adhesion molecules. Clin. Exp. Immunol. 1997.107: 601–607.
14 D’Amico, G., Bianchi, G., Bernasconi, S., Bersani, L., Pie-monti, L., Sozzani, S., Mantovani, A. and Allavena, P., Adhe-sion, transendothelial migration, and reverse transmigration of invitro cultured dendritic cells. Blood 1998. 92: 207–214.
15 Srinivas, U., Larsson, M., Lundblad, A. and Forsum, U., E-selectin involvement in in vitro adhesion of blood dendritic cellsto human umbilical cord endothelial cells. Scand. J. Immunol.1993. 38: 273–278.
16 Ferrero, E., Bondanza, A., Leone, B. E., Manici, S., Poggi, A.and Zocchi, M. R., CD14+CD34+ peripheral blood mononuclearcells migrate across endothelium and give rise to immunostimu-latory dendritic cells. J. Immunol. 1998. 160: 2675–2683.
17 Jonuleit, H., Kühn, U., Müller, G., Steinbrink, K., Paragnik, L.,Schmitt, E., Knop, J. and Enk, A. H., Pro-inflammatory cyto-kines and prostaglandins induce maturation of potent immuno-stimulatory dendritic cells under fetal calf serum-free conditions.Eur. J. Immunol. 1997. 27: 3135–3142.
18 De Saint-Vis, B., Vincent, J., Vandenabeele, S., Vanbervliet,B., Pin, J. J., Aı̈t-Yahia, S., Patel, S., Mattei, M. G., Banche-reau, J., Zurawski, S., Davoust, J., Caux, C. and Lebecque, S.,
A novel lysosome-associated membrane glycoprotein, DC-LAMP, induced upon DC maturation, is transiently expressed inMHC class II compartment. Immunity 1998. 9: 325–336.
19 Caux, C., Vanbervliet, B., Massacrier, C., Dezutter-Dambuyant, C., De Saint-Vis, B., Jacquet, C., Yoneda, K.,Imamura, S., Schmitt, D. and Banchereau, J., CD34+ hemato-poietic progenitors from human cord blood differentiate alongtwo independent dendritic cell pathways in response to GM-CSF+TNF § . J. Exp. Med. 1996. 184: 695–706.
20 Banchereau, J., Palucka, A. K., Dhodapkar, M., Burkeholder,S., Taquet, N., Rolland, A., Taquet, S., Coquery, S., Witt-kowski, K. M., Bhardwaj, N., Pineiro, L., Steinman, R. M. andFay, J., Immune and clinical responses in patients with meta-static melanoma to CD34+ progenitor-derived dendritic cell vac-cine. Cancer Res. 2001. 61: 6451–6458.
21 Swerlick, R. A., Lee, K. H., Wick, T. M. and Lawley, T. J.,Human dermal microvascular endothelial but not human umbili-cal vein endothelial cells express CD36 in vivo and in vitro. J.Immunol. 1992. 148: 78–83.
22 Cronstein, B. N. and Weissmann, G., The adhesion moleculesof inflammation. Arthritis Rheum. 1993. 36: 147–157.
23 Pober, J. S., Gimbrone, M. A., Jr., Lapierre, L. A., Mendrick, D.L., Fiers, W., Rothlein, R. and Springer, T. A., Overlapping pat-terns of activation of human endothelial cells by interleukin 1,tumor necrosis factor, and immune interferon. J. Immunol. 1986.137: 1893–1896.
24 Griffiths, C. E. M., Vorhees, J. J. and Nickoloff, B. J., Charac-terization of intercellular adhesion molecule-1 and HLA-DRexpression in normal and inflamed skin: Modulation by recombi-nant gamma interferon and tumor necrosis factor. J. Am. Acad.Dermatol. 1989. 20: 617–629.
25 Swerlick, R. A., Garcia-Gonzalez, E., Kubota, Y., Xu, Y. andLawley, T. J., Studies of the modulation of MHC antigen and celladhesion molecule expression on human dermal microvascularendothelial cells. J. Invest. Dermatol. 1991. 97: 190–196.
26 Robert, C., Fuhlbrigge, R. C., Kieffer, J. D., Ayehunie, S.,Hynes, R. O., Cheng, G. Y., Grabbe, S., Von Andrian, U. H. andKupper, T. S., Interaction of dendritic cells with skin endothelium:A new perspective on immunosurveillance. J. Exp. Med. 1999.189: 627–635.
27 Ferrero, E., Vettoretto, K., Bondanza, A., Villa, A., Resnati, M.,Poggi, A. and Zocchi, M. R., uPA/uPAR system is active inimmature dendritic cells derived from CD14+CD34+ precursorsand is down-regulated upon maturation. J. Immunol. 2000. 164:712–718.
28 Steinman, R. M., DC-SIGN: A guide to some mysteries of den-dritic cells. Cell 2000. 100: 491–494.
29 Geijtenbeek, T. B., Krooshoop, D. J., Bleijs, D. A., Van Vliet, S.J., van Duijnhoven, G. C., Grabovsky, V., Alon, R., Figdor, C. G.and Van Kooyk, Y., DC-SIGN-ICAM-2 interaction mediates den-dritic cell trafficking. Nat. Immunol. 2000. 1: 353–357.
30 Lawler, J., Weinstein, R. and Hynes, R. O., Cell attachment tothrombospondin: the role of ARG-GLY-ASP, calcium, and integrinreceptors. J. Cell Biol. 1988. 107: 2351–2361.
31 Oquendo, P., Hundt, E., Lawler, J. and Seed, B., CD36 directlymediates cytoadherence of Plasmodium falciparum parasitizederythrocytes. Cell 1989. 58: 95–101.
32 Shikano, S., Bonkobara, M., Zukas, P. K. and Ariizumi, K.,Molecular cloning of a dendritic cell-associated transmembraneprotein, DC-HIL, that promotes RGD-dependent adhesion ofendothelial cells through recognition of heparan sulfate proteo-glycans. J. Biol. Chem. 2001. 276: 8125–8134.
Eur. J. Immunol. 2002. 32: 3638–3650 Dendritic cell adhesion to dermal endothelial cells 3649

33 Steinman, R. M. and Dhodapkar, M., Active immunizationagainst cancer with dendritic cells: The near future. Int. J. Cancer2001. 94: 459–473.
34 Lukas, M., Stössel, H., Hefel, L., Imamura, S., Fritsch, P.,Sepp, N. T., Schuler, G. and Romani, N., Human cutaneousdendritic cells migrate through dermal lymphatic vessels in a skinorgan culture model. J. Invest. Dermatol. 1996. 106: 1293–1299.
35 Weinlich, G., Heine, M., Stössel, H., Zanella, M., Stoitzner, P.,Ortner, U., Smolle, J., Koch, F., Sepp, N. T., Schuler, G. andRomani, N., Entry into afferent lymphatics and maturation in situof migrating cutaneous dendritic cells. J. Invest. Dermatol. 1998.110: 441–448.
36 Kriehuber, E., Breiteneder-Geleff, S., Groeger, M., Soleiman,A., Schoppmann, S. F., Stingl, G., Kerjaschki, D. and Maurer,D., Isolation and characterization of dermal lymphatic and bloodendothelial cells reveal stable and functionally specialized cell lin-eages. J. Exp. Med. 2001. 194: 797–808.
37 Sepp, N. T., Cornelius, L. A., Romani, N., Li, L. J., Caughman,S. W., Lawley, T. J. and Swerlick, R. A., Polarized expressionand basic fibroblast growth factor-induced down-regulation ofthe alpha 6 beta 4 integrin complex on human microvascularendothelial cells. J. Invest. Dermatol. 1995. 104: 266–270.
38 Sgonc, R., Gruschwitz, M. S., Boeck, G., Sepp, N., Gruber, J.and Wick, G., Endothelial cell apoptosis in systemic sclerosis isinduced by antibody-dependent cell-mediated cytotoxicity viaCD95. Arthritis Rheum. 2000. 43: 2550–2562.
39 Braut-Boucher, F., Pichon, J., Rat, P., Adolphe, M., Aubery, M.and Font, J., A non-isotopic, highly sensitive, fluorimetric, cell-cell adhesion microplate assay using calcein AM-labeled lym-phocytes. J. Immunol. Methods 1995. 178: 41–51.
40 Akeson, A. L. and Woods, C. W., A fluorometric assay for thequantitation of cell adherence to endothelial cells. J. Immunol.Methods 1993. 163: 181–185.
41 Valladeau, J., Ravel, O., Dezutter-Dambuyant, C., Moore, K.,Kleijmeer, M., Liu, Y., Duvert-Frances, V., Vincent, C., Schmitt,D., Davoust, J., Caux, C., Lebecque, S. and Saeland, S., Lan-gerin, a novel C-type lectin specific to Langerhans cells, is anendocytic receptor that induces the formation of Birbeck gran-ules. Immunity 2000. 12: 71–81.
Correspondence: Van Anh Nguyen, Department of Derma-tology and Venereology, University of Innsbruck, Anichs-trasse 35, Innsbruck, A-6020 AustriaFax: +43-512-504-3017e-mail: van.nguyen — uibk.ac.at
3650 Van A. Nguyen et al. Eur. J. Immunol. 2002. 32: 3638–3650