Embed Size (px)
Citation preview

1864 Journal of Lipid Research Volume 53, 2012
Copyright © 2012 by the American Society for Biochemistry and Molecular Biology, Inc.
This article is available online at http://www.jlr.org
deacetylases (HDACs) and histone acetyltransferases are enzymes that catalyze deacetylation and acetylation of the � -amino groups of lysine residues of proteins. Lysine acety-lation was fi rst discovered to occur in histones, and histone acetylation is important in controlling the structure and function of chromatin ( 1 ). A large number of studies have further shown the existence of acetylated nonhistone pro-teins, including transcription factors, hormone receptors, signal transducers, and chaperones. The reversible acety-lation of nonhistone proteins modulates a wide variety of key cellular processes, such as apoptosis, survival, and pro-liferation. Recently, several groups have reported that many metabolic enzymes are highly acetylated ( 2–5 ). Such enzymes are involved in glycolysis, fatty acid metabolism, gluconeogenesis, the TCA cycle, and the urea cycle, and the acetylation of these proteins regulates their activity so that they can respond to the metabolic demands of cells.
Adipose tissue, which consists of loose connective tissue composed of adipocytes, is an important meta-bolic organ that functions in energy homeostasis ( 6 ). Adipocytes regulate physiologic processes, including glucose metabolism, angiogenesis, the infl ammatory re-sponse, and reproductive functions through secreted adipokines. Adipocyte differentiation can contribute to the development of obesity via a positive energy balance (energy intake > energy expenditure) ( 7 ). Obesity leads to serious health problems all over the world. The coex-istence of obesity, type II diabetes, dyslipidemia, and hy-pertension, known as metabolic syndrome, constitutes an increased risk for the development of cardiovascular diseases ( 8 ). In adipocytes, fatty acids are stored as tri-glycerides, which serve as the fuel for maintaining en-ergy balance. Acetyl-CoA is the direct precursor in the synthesis of fatty acids, and all fatty acid carbons come from the acetyl group of acetyl-CoA. In addition, it has
Abstract Acetylation is one of the most crucial post-translational modifi cations that affect protein function. Protein lysine acetylation is catalyzed by acetyltransferases, and acetyl-CoA functions as the source of the acetyl group. Additionally, acetyl-CoA plays critical roles in maintaining the balance between carbohydrate metabolism and fatty acid synthesis. Here, we sought to determine whether lysine acetylation is an important process for adipocyte differenti-ation. Based on an analysis of the acetylome during adipo-genesis, various proteins displaying signifi cant quantitative changes were identifi ed by LC-MS/MS. Of these identifi ed proteins, we focused on malate dehydrogenase 1 (MDH1). The acetylation level of MDH1 was increased up to 6-fold at the late stage of adipogenesis. Moreover, overexpression of MDH1 in 3T3-L1 preadipocytes induced a signifi cant in-crease in the number of cells undergoing adipogenesis. The introduction of mutations to putative lysine acetyla-tion sites showed a signifi cant loss of the ability of cells to undergo adipogenic differentiation. Furthermore, the acet-ylation of MDH1 dramatically enhanced its enzymatic ac-tivity and subsequently increased the intracellular levels of NADPH. These results clearly suggest that adipogenic dif-ferentiation may be regulated by the acetylation of MDH1 and that the acetylation of MDH1 is one of the cross-talk mechanisms between adipogenesis and the intracellular en-ergy level. —Kim, E. Y., W. K. Kim, H. J. Kang, J. H. Kim, S. J. Chung, Y. S. Seo, S. G. Park, S. C. Lee, and K-H. Bae. Acety-lation of malate dehydrogenase 1 promotes adipogenic dif-ferentiation via activating its enzymatic activity. J. Lipid Res . 2012. 53: 1864–1876.
Supplementary key words acetyl-CoA • adipogenesis • obesity • protein acetylation
Protein lysine acetylation is a posttranslational modifi ca-tion that acts as a key regulator of cellular processes. Histone
This work was supported by grants from KRIBB, the Korea Research Council of Fundamental Science and Technology (NAP project), and the Research Program (grant nos. 2011-0027634, 2011-0027796 and 2011-0030028) of the Korea National Research Foundation.
Manuscript received 17 March 2012 and in revised form 10 June 2012.
Published, JLR Papers in Press, June 12, 2012 DOI 10.1194/jlr.M026567
Acetylation of malate dehydrogenase 1 promotes adipogenic differentiation via activating its enzymatic activity
Eun Young Kim , * ,† Won Kon Kim , † Hyo Jin Kang , § Jeong-Hoon Kim , † Sang J. Chung , § Yeon Soo Seo , * Sung Goo Park , † Sang Chul Lee , 1,† and Kwang-Hee Bae 1, †
Department of Biological Sciences,* KAIST , Daejeon 305-701, Republic of Korea ; Medical Proteomics Research Center, † and the BioNano Research Center, § KRIBB , Daejeon 305-806, Republic of Korea
Abbreviations: 2-DE, two-dimensional gel electrophoresis; HDAC, histone deacetylases; MDH, malate dehydrogenase.
1 To whom correspondence should be addressed. e-mail: [email protected] . (S.C.L.); [email protected] (K-H.B.)
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

Acetylation of MDH1 promotes adipogenesis 1865
FBS, and 10 � g/ml insulin. The medium was replenished every other day.
Oil-Red-O staining Cultured cells were washed twice with PBS and fi xed with 10%
formalin for 30 min at room temperature. The cells were then washed with distilled water and stained for 30 min at room tem-perature with 0.3% fi ltered Oil-Red-O solution in 60% isopropa-nol (Sigma, St. Louis, MO). The stained cells were washed with distilled water, and micrographs were obtained. To extract the incorporated Oil-Red-O dye, absolute isopropanol was added to the stained cell-culture dish, and the dish was shaken at room temperature for 30 min. Triplicate samples were read at 510 nm using a GeneQuant 1300 spectrophotometer (GE HealthCare, Uppsala, Sweden) ( 20, 21 ).
Sample preparation and 2D-electrophoresis The proteins were extracted from 3T3-L1 cells using a lysis
buffer (30 mM Tris [pH 8.5], 7 M urea, 2 M thiourea, and 4% 3-[(3-cholamidopropyl)-dimethylammonio propanesulfonate [CHAPS]). Protein samples to be separated on the same gel were pooled, and 2 volumes of cold acetone were added. After centrifugation, the precipitate was rehydrated using a rehydra-tion buffer (7 M urea, 2 M thiourea, 4% CHAPS, 2% ampholyte, and 20 mM DTT), and isoelectric focusing was conducted using a Multiphor II apparatus (GE HealthCare). After focusing, each strip (a 7 cm immobilized pH nonlinear strip [pH 3–11]) was equilibrated twice for 15 min in 2.5 ml of equilibrium buffer. The equilibrium buffer contains 50 mM Tris (pH 8.8), 6 M urea, and 30% glycerol. During the second equilibrium step, 260 mM iodoacetamide was added to the equilibrium buffer. The IPG strips were then loaded and separated on a 10% ac-rylamide SDS-PAGE gel using a Mini-Protean system (Bio-Rad, Hercules, CA).
Two-dimensional gel electrophoresis Western blot analysis
Sixty micrograms of total protein was used for the detection of lysine acetylated proteins during adipogenic differentiation. The samples were separated in the first and second dimension following previously described protocols ( 22, 23 ) and transferred to an Immunoblot PVDF membrane (Millipore). Membranes were blocked with 5% skim milk for 1 h and probed with a rabbit anti-acetyl-lysine antibody (ICP0380 Immunechem) diluted 1:2,000 in TBS with 5% skim milk for 4 h at 4°C as previously described ( 21 ).
Protein digestion, peptide enrichment, and MS analysis The separated proteins in SDS-PAGE gels were visualized by
Coomassie brilliant blue G-250 staining, and spots of interest were detected. Gel pieces were washed twice with 150 � l of 100 mM ammonium bicarbonate (pH 8.2) and 70% v/v acetonitrile and dried at 37°C for 20 min. Trypsin in 50 mM ammonium bicar-bonate (20 � g/ � l) was added to each gel piece, and gel pieces were incubated at 37°C for 2 h. Peptides were then extracted using a mixture containing 20 � l of 0.1% v/v trifl uoroacetic acid and 70% acetonitrile. The peptides were purifi ed using ZipTips TM (Millipore) according to the manufacturer’s instructions before LC-MS/MS spectrometric analysis. The peptides were analyzed using a Synapt High Defi nition mass spectrometer (Waters, Manchester, UK) as previously described ( 24, 25 ).
Construction of retroviral vectors and transduction To construct 3T3-L1 cells that stably express a FLAG-tagged
wild-type or mutant MDH1 protein, a retroviral infection system
been reported that the adipogenic differentiation could be differentially regulated by HDAC inhibitors ( 9–11 ). These facts led us to hypothesize that the acetylation of proteins plays a key role in controlling adipogenesis.
Malate dehydrogenase (MDH) catalyzes the intercon-version of malate and oxaloacetate in the mitochondrial membrane using the coenzyme NAD + /NADH ( 12 ). In eu-karyotic cells, two MDH proteins exist and are classifi ed according to their subcellular localization as MDH1 or MDH2 ( 13 ). In the mitochondrial matrix, MDH2 oxidizes malate to oxaloacetate through the citric acid cycle. The cytosolic form of the protein (MDH1) participates in the malate/aspartate shuttle. The shuttle allows for the overall balancing of the import and export of nitrogen, oxaloace-tate, and the � -ketoglutarate intermediate in the cytosol and mitochondrial matrix ( 14 ). The malate required for antiport exchange can be exported by citrate, which accu-mulates in large amounts, instead of isocitrate. Accumu-lated citrate is transported from the mitochondria to the cytosol. Then, oxaloacetate and acetyl-CoA are synthesized in the cytosol. Cytosolic oxaloacetate is then converted to malate by MDH1, and acetyl-CoA is used for fatty acid syn-thesis. In mammals, MDH1 is highly expressed in many tissues, and its different roles are determined by its expres-sion levels ( 15–17 ).
In this study, we attempted to determine whether lysine acetylation is an important process for adipocyte differen-tiation. Based on an analysis of the acetylome during adi-pogenesis, a number of proteins displaying signifi cant quantitative changes were identifi ed using LC-MS/MS. Among these proteins, we focused on MDH1. The acetyla-tion level of MDH1 is signifi cantly increased during adipo-genic differentiation. Interestingly, the overexpression of MDH1 in 3T3-L1 preadipocytes induces enhanced adipo-genesis in these cells. The introduction of mutations to putative lysine acetylation sites in MDH1 reduces the ability of the enzyme to promote adipogenic differentia-tion. Additionally, the acetylation of MDH1 signifi cantly increases its enzymatic activity. These data suggest that the lysine acetylation of MDH1 is involved in the regulation of adipogenic differentiation.
MATERIALS AND METHODS
Cell culture and adipogenic differentiation The preadipocyte cell line, 3T3-L1, which is derived from
mouse embryonic fi broblasts, was purchased from ATCC. The cells were cultured in growth medium (high glucose DMEM con-taining a 1% antibiotic-antimycotic solution and 10% bovine calf serum; Gibco-Invitrogen, Carlsbad, CA) at 37°C in a humidifi ed atmosphere with 5% CO 2 . GP2-293 packaging cells were grown in DMEM containing a 1% antibiotic-antimycotic solution and 10% FBS. 3T3-L1 cells were induced to differentiate into mature adi-pocytes as described in our previous reports ( 18–21 ). Confl uent 3T3-L1 cells were incubated in differentiation medium com-posed of DMEM, 10% FBS, and MDI (a differentiation cocktail of 0.5 mM 3-isobutyl-1-methylxanthine, 1 � M dexamethasone, and 10 � g/ml insulin [Sigma, US]). After 48 h, the medium was changed to a maintenance medium composed of DMEM, 10%
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

1866 Journal of Lipid Research Volume 53, 2012
Activity assay of MDH1 Cell pellets were lysed with NP-40 buffer supplemented with a
protease inhibitor (Roche, Indianapolis, IN) and a KDAC inhib-itor (10 µM trichostatin A, 10 mM nicotinamide, and 50 mM bu-tyric acid; Sigma, US) ( 5 ). For immunoprecipitation, extracted proteins were incubated with anti-FLAG M2 agarose beads (Sigma) at 4°C overnight. The immunoprecipitated beads were washed 10 times with NP-40 buffer, and then the FLAG fusion proteins were eluted using a 3×FLAG peptide (Sigma). The con-centration of the eluted proteins was measured using a Bradford assay and Coomassie blue G-250 staining. Purifi ed human MDH1 that was overexpressed in 3T3-L1 cells was incubated with 0.2 mM oxaloacetate and 1 mM NADH in PBS (pH 7.5). The activity of MDH1 was determined by measuring the decrease in the fl uores-cence of NADH (Ex. 350 nm; Em. 470 nm). The reaction was monitored 15 times at 1-min intervals on a Victor TM X3 Multilabel plate reader (PerkinElmer, Waltham, MA).
Analysis of intracellular NADPH levels Commercial NADP/NADPH assay kits (Abcam, Cambridge, UK)
were used to detect NADPH and total NADP. The cells were washed three times with ice-cold PBS and placed in NADP/NADPH extraction buffer. The washed cells were extracted by two freeze (20 min on dry-ice)/thaw (10 min at room tempera-ture) cycles, and then the cell lysates were dissolved in buffer using gentle sonication. The concentration of the eluted proteins was measured using a Bradford assay (Bio-Rad). To calculate the NADP/NADPH ratio, total NADP (NADPt) and NADPH-only were prepared, and an NADPH standard was made using a serial dilution of NADPH. The NADPH reaction was monitored on a microplate reader (Bio-Rad) for 1 to 4 h at 450 nm.
RESULTS
Analysis of acetylome changes during adipogenesis Many groups have reported that the adipogenic differ-
entiation process could be differentially regulated de-pending on the type of HDAC inhibitors, suggesting that acetylation may be intimately associated with adipogenesis ( 9–11 ). As an initial step, 1D SDS-PAGE was conducted to determine whether acetylation patterns changed during adipogenesis ( Fig. 1A ). Whole cell proteins from 3T3-L1 cells derived from 0, 2, and 12 days after the induction of differentiation were used for analysis. It was confi rmed that the level of aP2 was increased after adipogenic stimu-lation ( Fig. 1B ). The expression level of histone H3 was unchanged during differentiation, and it was thus used as a loading control. Our results clearly demonstrated that the acetylation pattern and protein expression were signifi cantly altered during the differentiation of 3T3-L1 preadipocytes into adipocytes. Next, we performed two-dimensional gel electrophoresis (2-DE). The expression pattern of proteins with isoelectric points ranging between pH 3 and 11 was analyzed. The overall changes in the acet-ylation pattern (acetylome) were also investigated by West-ern blot analysis using an anti-acetyl-lysine antibody after 2-DE ( Fig. 1C ). As shown, the levels of lysine-acetylated proteins were generally increased after adipogenic dif-ferentiation. A total of 69 spots showing changes in acet-ylation during adipogenesis were detected by image
was used. For the expression of MDH1, DNA encoding the FLAG-tagged MDH1 was inserted into 3T3-L1 cells using the pRetroX-IRES-ZsGreen1 vector (Clontech). For virus produc-tion, GP2-293 cell lines were transfected using Lipofectamine 2000 (Gibco-Invitrogen). The details of the transfection and transduction methods have been described in our previous re-ports ( 18–21, 26 ). Infected cells were selected using a FACSAria cell sorter (BD Biosciences, San Jose, CA) and further main-tained in growth medium. The ectopic expression of MDH1 was confi rmed by Western blot analysis.
Introduction of mutations at putative acetylation sites FLAG-tagged MDH1 was mutated using the EZchange TM
Site-Directed Mutagenesis kit (Enzynomics, KR). The putative acetylation sites in MDH1 were mutated to arginine or gluta-mine. Arginine is charged and abolishes acetylation, whereas glutamine is uncharged and may mimic the acetylated lysine. The sites mutated in this study were based on those reported by Zhao et al. (K118, K121, and K298) ( 5 ). We introduced the mutation into each site individually or at all three sites of MDH1.
Construction and purifi cation of mutant MDH1 proteins in Escherichia coli
Sequences from the wild-type and mutant MDH1 gene were amplifi ed using the forward primer 5 ′ - gggaattc catatg atgtctgaac-caatcagagtc-3 ′ containing an Nde I site (in bold) and the reverse primer 5 ′ -ccg ctcgag ttaggcagaggaaagaaattcaaa-3 ′ containing a Xho I site (in bold) and inserted into the corresponding sites of the pET28a plasmid (Novagen, Madison, WI). Escherichia coli Rosetta DE3 cells (Novagen) carrying the respective expression plasmids were grown at 37°C in LB medium until the optical density at A 600 reached 0.6–0.8. The expression of MDH1 and MDH1 mutants was induced by adding 1 mM IPTG at 18°C for 18 h. The cells were harvested, washed with lysis buffer (50 mM Tris [pH 7.5], 250 mM NaCl, 5% glycerol, and 1 mM � -mercaptoethanol), and lysed by ultrasonication. After centrifugation (29,820 g for 30 min), the supernatants were incubated with a cobalt affi nity resin (TALON ® , Clontech) on a rocker for 1 h at 4°C and washed with lysis buffer supplemented with 10 mM imidazole. The proteins were eluted from the metal affi nity resin using a lysis buffer sup-plemented with 100 mM imidazole. The proteins were concen-trated to 5 mg/ml and stored at � 80°C.
Immunoblotting and immunoprecipitation The cells were washed three times with ice-cold PBS and
were harvested in ice-cold 7 M urea lysis buffer containing a protease inhibitor cocktail (Roche). The protein concentra-tions were measured using a Bradford assay (Bio-Rad). Immuno-blotting and immunoprecipitation were performed as described previously. The anti-MDH1 antibody was obtained from Santa Cruz, and the acetyl-lysine antibody was purchased from Im-muneChem Pharmaceuticals Inc. Anti-FLAG and anti- � -tubulin antibodies were purchased from Sigma. Anti-enolase1 and anti-histone H3 antibodies were obtained from Cell Signaling (Beverley, MA). The secondary antibodies were purchased from Abcam. The membranes were visualized using the Super-Signal West Pico Chemiluminescent Substrate kit (Pierce, Rockford, IL).
For immunoprecipitation, anti-acetyl-lysine antibody agarose beads (ImmuneChem Pharmaceuticals Inc.) were added to 200 � g of the lysates. The mixture was incubated overnight at 4°C. The immunoprecipitates were recovered by centrifugation at 2,500 g , washed fi ve times with NP-40 lysis buffer, and analyzed as de-scribed above.
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

Acetylation of MDH1 promotes adipogenesis 1867
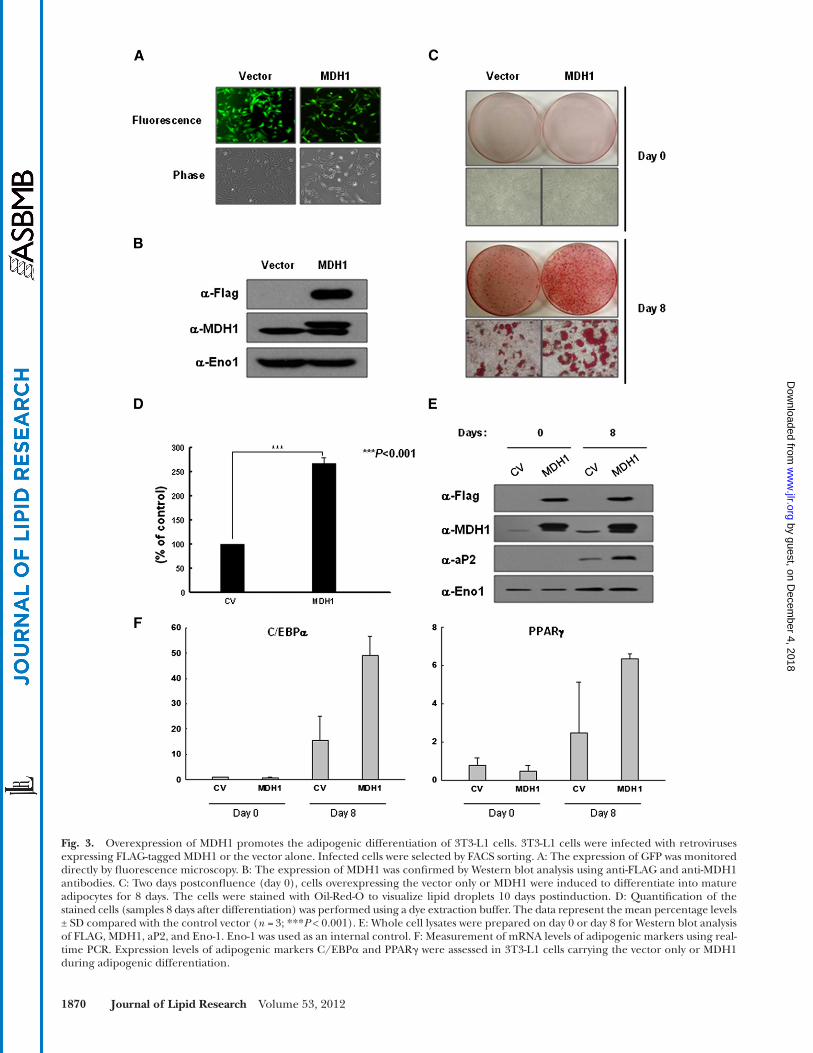
in adipogenic differentiation. Next, to clarify the role of MDH1 acetylation in adipogenesis, 3T3-L1 cells were in-fected with the FLAG-tagged, full-length human MDH1 protein using a retroviral infection system (IRES-GFP). A control vector was used as the negative control. In-fected cells were isolated by a FACS sorter, and most of the cells were GFP positive by fl uorescence microscopy ( Fig. 3A ). The overexpression of MDH1 proteins was confi rmed by Western blot analysis using anti-FLAG and anti-MDH1 antibodies ( Fig. 3B ). Infected cells were in-duced to differentiate into mature adipocytes, and after 8 days, lipid accumulation was assessed by Oil-Red-O staining of control vector- and MDH1-expressing cells ( Fig. 3C ). The results clearly showed that the overex-pression of MDH1 signifi cantly increased lipid accumu-lation when compared with the control vector ( Fig. 3C, D ). Consistently, the level of aP2 in the MDH1-infected cells was higher than that in the control vector-expressing cells ( Fig. 3E ). In addition, the expression levels of adi-pogenic markers, such as C/EBP � and PPAR � , were also increased upon ectopic expression of MDH1.
Effect of the mutation(s) of putative acetylation sites in MDH1 on adipogenic differentiation
To examine the effect of MDH1 acetylation on adipo-genesis, putative acetylation sites in MDH1 were mutated to arginine, which is charged but blocks acetylation, or to glutamine, which is uncharged and may mimic the acetylated lysine ( 2, 4, 5, 27 ). K118, K121, and K298 were
analysis and visual confi rmation; these spots were identifi ed by LC-MS/MS analysis ( Table 1 ). As expected, a variety of metabolic enzymes, including those involved in glycolysis, fatty acid metabolism, and the TCA cycle, were acetylated during adipogenesis.
To validate the results of the acetylome analysis, we as-sessed the expression and acetylation level of several of the identifi ed proteins in 3T3-L1 cells using Western blot and immunoprecipitation analyses, respectively ( Fig. 2A ). MDH1, one of the identifi ed candidates, showed an ap-proximately 2-fold increase in expression during adipo-genesis. The acetylation level of MDH1 was also dramatically enhanced (6-fold) during adipogenic differentiation ( Fig. 2B ). MDH2 also demonstrated this enhanced acety-lation pattern, but enolase showed no changes in expres-sion or acetylation level during adipogenesis. Generally, the Western blot and immunoprecipitation results corre-lated well with the 2-DE acetylome data (data not shown).
Effect of the ectopic expression of MDH1 on adipogenic differentiation in 3T3-L1 preadipocytes
To examine if the acetylation of MDH1 occurred during adipogenesis, we assessed the expression and acetyla-tion levels of MDH1 in other differentiation processes, such as myogenesis and osteogenesis. There was no sig-nifi cant change in expression and acetylation levels during myogenesis and osteogenesis (data not shown). These results suggest that the acetylation of MDH1 occurred
Fig. 1. Acetylome analysis during the adipogenic differentiation of 3T3-L1 cells. A: Patterns of protein expression during adipogenic differentiation. Cell lysates were loaded and visualized by Coomassie blue G-250 staining, and the acetylome pattern was confi rmed by Western blot analysis using an anti-acetyl-lysine antibody. IB, immunoblotting. B: Whole cell lysates were prepared on days 0, 2, and 12 for Western blot analysis of aP2, H3, and Eno-1. H3 and Eno-1 were used as internal controls. C: Screening of acetylated proteins by 2-DE Western blot analysis. Whole cell lysates were applied to a fi rst dimension of a pH 3-11 non-linear IPG strip (7 cm) and a second dimension of 12.5% SDS-PAGE visualized by Coomassie blue G-250 staining. The acetylome was identifi ed by Western blot analysis using an anti-acetyl-lysine antibody (anti-AcK). The circles mark spots that show signifi cant changes in acetylation levels during adipogenesis. The indicated spot numbers in Table 1 correspond to the spots shown in Fig. 1 .
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

1868 Journal of Lipid Research Volume 53, 2012
TABLE 1. The list of proteins showing differential acetylation pattern during adipogenesis
Spot No. Accession Number Protein Name Function Moscot Score
1 gi|7106439 Tubulin, � -5 Cytoskeleton 1,8952 gi|55408 Vimentin Cytoskeleton 5693 gi|2078001 Vimentin Cytoskeleton 1,1774 gi|31982755 Vimentin Cytoskeleton 1,4355 gi|13384620 Heterogeneous nuclear ribonucleoprotein K Translation 6406 gi|76779273 Hspd1 protein Molecular chaperone 8007 gi|7305075 ras GTPase-activating protein binding protein Nucleotide binding 2968 gi|4504445 hnRNP A1 Translation 5879 gi|226443091 hnRNP A0 Translation 29710 gi|3329498 hnRNP A2/B1 Translation 77411 gi|3329498 hnRNP A2/B1 Translation 96812 gi|51769129 Similar to hnRNP A3, isoform 1 Translation 82713 gi|51769129 Similar to hnRNP A3, isoform 1 Translation 83514 gi|51769129 Similar to hnRNP A3, isoform 1 Translation 28815 gi|3329498 hnRNP A2/B1 Translation 16116 gi|31982186 Malate dehydrogenase 2 (MDH2) Glycolysis 117617 gi|387397 Epidermal keratin subunit I Protein binding 39518 gi|18700004 Acetyl-CoA acyltransferase 1 Fatty acid metabolism 121119 gi|28201978 Pyruvate dehydrogenase complex, component X TCA cycle 39120 gi|28201978 Pyruvate dehydrogenase complex, component X TCA cycle 41821 gi|10946928 hnRNP H1 Translation 69122 gi|70794816 Hypothetical protein LOC433182 Glycolysis 166323 gi|70794816 Hypothetical protein LOC433182 Glycolysis 227224 gi|70794816 Hypothetical protein LOC433182 Glycolysis 185825 gi|70794816 Hypothetical protein LOC433182 Glycolysis 41626 gi|6680618 Medium-chain acyl-CoA dehydrogenase Fatty acid metabolism 76427 gi|21450129 Acetyl-Co A acetyltransferase 1 precursor Fatty acid metabolism 66928 gi|6680618 Medium-chain acyl-CoA dehydrogenase Fatty acid metabolism 62229 gi|21450129 Acetyl-Co A acetyltransferase 1 precursor Fatty acid metabolism 79730 gi|21704100 Mitochondrial trifunctional protein, � subunit Glycolysis 51131 gi|114326546 Bisphosphoglycerate mutase 1 Glycolysis 117632 gi|34328230 Adenylate kinase 2, isoform b ATP binding 32633 gi|2851390 Triosephosphate isomerase Glycolysis 83234 gi|2851390 Triosephosphate isomerase Glycolysis 78935 gi|114326546 Bisphosphoglycerate mutase 1 Glycolysis 69836 gi|9789726 Septin-7 GTP binding 19537 gi|162287370 Lamin A, isoform A Protein binding 98538 gi|162287370 Lamin A, isoform A Protein binding 112739 gi|162287370 Lamin A, isoform A Protein binding 160340 gi|6753620 DEAD/H box polypeptide 3, X-linked Protein binding 55941 gi|126116585 Keratin complex 2, basic, gene 1 Structural activity 19942 gi|20072624 hnRNP L Translation 49843 gi|20072624 hnRNP L Translation 31844 gi|20072624 hnRNP L Translation 124045 gi|21313308 hnRNP M, isoform a Translation 60946 gi|21313308 hnRNP M, isoform a Translation 41547 gi|20982845 Pigpen Nucleic acid binding 111148 gi|21313308 hnRNP M, isoform a Translation 51249 gi|6680748 ATP synthase subunit � , mitochondrial precursor Metabolism 39350 gi|9789726 Septin-7 GTP binding 46251 gi|9789726 Septin-7 GTP binding 54952 gi|556301 Elongation factor Translation 24853 gi|6753022 Adenylate kinase 3-like 1 ATP/GTP binding 61554 gi|31982861 Carbonic anhydrase III Metabolism 105755 gi|31982186 Malate dehydrogenase 2 (MDH2) Glycolysis 80356 gi|31982186 Malate dehydrogenase 2 (MDH2) Glycolysis 31357 gi|32130423 Long-chain specifi c acyl-CoA dehydrogenase Fatty acid metabolism 54358 gi|21450129 Acetyl-Co A acetyltransferase 1 precursor Fatty acid metabolism 74359 gi|6680618 Medium-chain acyl-CoA dehydrogenase Fatty acid metabolism 74760 gi|21450129 Acetyl-Co A acetyltransferase 1 precursor Fatty acid metabolism 75861 gi|29126205 Acetyl-Co A acyltransferase 2 Fatty acid metabolism 68862 gi|21450129 Acetyl-Co A acetyltransferase 1 precursor Fatty acid metabolism 79463 gi|6671539 Fructose-bisphosphate aldolase A Glycolysis 35764 gi|127798626 Acetyl-CoA acetyltransferase 2 Fatty acid metabolism 46665 gi|31982186 Malate dehydrogenase 2 (MDH2) Glycolysis 51666 gi|6680690 Peroxiredoxin 3 Antioxidant protein 26767 gi|6680690 Peroxiredoxin 3 Antioxidant protein 48268 gi|6679299 Prohibitin Antioxidant protein 70369 gi|387129 Malate dehydrogenase 1 (MDH1) Glycolysis/TCA cycle 512
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

Acetylation of MDH1 promotes adipogenesis 1869
using the E. coli expression system. Mutant MDH1s have similar activity to wild-type MDH1, indicating that the introduction of the mutation has no effect on enzymatic activity (data not shown).
Activity assay of MDH1 An immunoblot analysis showed that the acetylation
level of MDH1 increased during adipogenesis ( Fig. 5A ). Generally, acetylation modulates enzymatic function by al-tering enzymatic stability, catalytic activity, or protein-protein interactions. To explore the direct effect of acetylation on MDH1 activity, we purifi ed the ectopically expressed wild-type MDH1 during adipogenesis (day 0 and day 8). MDH1 activity was increased by more than approximately 50% 8 days after differentiation when compared with the ac-tivity at day 0 ( Fig. 5B ). We then investigated the activity of the ectopically expressed wild-type and mutant MDH1 proteins after immunopurifi cation. MDH1-3KR and MDH1-K118R showed signifi cantly reduced enzymatic activity when compared with the wild-type and KQ mutant (Fig. 5C, D). Furthermore, the acetylation level of MDH1-3KR and MDH1-K118R considerably decreased when compared with wild-type MDH1 ( Fig. 5E ). These results clearly indicate that the acetylation level of MDH1 is enhanced during adipogenesis and that the enhanced acetylation of MDH1 increases its enzymatic activity.
Analysis of intracellular levels of NADPH after the introduction of MDH1 or mutant MDH1
Intracellular NADPH is an important source of re-ducing power for fatty acid synthesis. As mentioned above, MDH1 participates in the malate/aspartate shuttle that contributes the supply of acetyl-CoA and NADPH for fatty acid synthesis in the cytoplasm. To elucidate the mechanism of action of acetylated MDH1 on adipogenic differentiation, the NADP/NADPH ra-tio was assessed in cells 8 days after differentiation.
chosen for mutation to arginine or glutamine. 3T3-L1 cells were then infected with these mutants of MDH1 using a retroviral system (IRES-GFP). The control vector was used as a negative control, and wild-type MDH1 was used as a positive control. Infected cells were isolated using a FACS sorter. Most of the infected cells were GFP positive by fl uorescence microscopy ( Fig. 4A ). The over-expression of the mutated MDH1 proteins was con-fi rmed by Western blot analysis using anti-FLAG and anti-MDH1 antibodies ( Fig. 4B ). Infected cells were cultured in differentiation medium for 8 days, and lipid accumulation was assessed by Oil-Red-O staining. Among the MDH1 mutants, the 3KR (in which the three puta-tive acetylation lysine residues were replaced with argi-nine) and K118R mutants showed a signifi cantly lower level of Oil-Red-O staining than the cells infected with wild-type MDH1 ( Fig. 4C, D ). These results clearly dem-onstrate that the mutation of the acetylation site(s) in-fl uences adipogenic differentiation, indicating that the acetylation of MDH1 is intimately involved in control-ling adipogenic differentiation. We confi rmed the con-tinuous overexpression of MDH1 until the late stage of adipogenic differentiation. In addition, we found that differences in adipogenic differentiation correlated well with the expression level of aP2 ( Fig. 4E, F ). The acety-lation levels of 3KR and K118R were considerably lower than those of wild-type MDH1 ( Fig. 4G, H ). Next, we measured the mRNA level of adipogenic markers, such as adiponectin, resistin, C/EBP � , and PPAR � , using real-time PCR. As expected, the expression levels of these adipo-genic markers were signifi cantly increased by introducing the MDH1. Additionally, 3T3-L1 cells carrying 3KR or K118R showed the reduced expression levels of adipo-genic markers, compared with wild-type MDH1 ( Fig. 4I ). To rule out the possibility that the mutation of MDH1 af-fects its enzymatic activity, we expressed and purifi ed the wild-type and mutant MDH1 proteins (3KR and K118R)
Fig. 2. Expression and acetylation level of MDH1 during 3T3-L1 adipocyte differentiation. A: The expression and acetylation levels of endogenous MDH1 and MDH2 were examined by IB and immunoprecipitation (IP). Enolase-1 (Eno-1) indicates equal amounts of each protein. B: Quantifi cation of the expression and acetylation levels of MDH1 and MDH2. The quantifi cation was calculated based on the intensity of (A). The data represent the mean percentage levels ± SD compared with day 0 ( n = 3; ** P < 0.01, *** P < 0.001). The black bar indicates the expression level of MDH1, and the gray bar indicates the acetylation level of MDH1.
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from
1870 Journal of Lipid Research Volume 53, 2012
Fig. 3. Overexpression of MDH1 promotes the adipogenic differentiation of 3T3-L1 cells. 3T3-L1 cells were infected with retroviruses expressing FLAG-tagged MDH1 or the vector alone. Infected cells were selected by FACS sorting. A: The expression of GFP was monitored directly by fl uorescence microscopy. B: The expression of MDH1 was confi rmed by Western blot analysis using anti-FLAG and anti-MDH1 antibodies. C: Two days postconfl uence (day 0), cells overexpressing the vector only or MDH1 were induced to differentiate into mature adipocytes for 8 days. The cells were stained with Oil-Red-O to visualize lipid droplets 10 days postinduction. D: Quantifi cation of the stained cells (samples 8 days after differentiation) was performed using a dye extraction buffer. The data represent the mean percentage levels ± SD compared with the control vector ( n = 3; *** P < 0.001). E: Whole cell lysates were prepared on day 0 or day 8 for Western blot analysis of FLAG, MDH1, aP2, and Eno-1. Eno-1 was used as an internal control. F: Measurement of mRNA levels of adipogenic markers using real-time PCR. Expression levels of adipogenic markers C/EBP � and PPAR � were assessed in 3T3-L1 cells carrying the vector only or MDH1 during adipogenic differentiation.
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

Acetylation of MDH1 promotes adipogenesis 1871
Fig. 4. Effects of putative acetylation site(s) mutation of MDH1 on the adipogenic differentiation of 3T3-L1 cells. A: 3T3-L1 cells were infected with retroviruses containing the vector alone, MDH1-3K mutants (3KQ and 3KR), or MDH1-K118 mutants (K118Q and K118R). The infected cells were enriched by FACS sorting and confi rmed by fl uorescence microscopy. B: Mutants of MDH1 were confi rmed by Western blot analysis using anti-FLAG, anti-MDH1, and anti- � -tubulin antibodies. � -tubulin was used as a loading control. C: Cells carrying the mutant MDH1 were induced to differentiate into adipocytes for 8 days. The cells were stained with Oil-Red-O. D: Quantifi cation of the stained cells was performed using a dye extraction buffer. The data represent the mean percentage levels ± SD compared with the control vector (n = 3; * P < 0.05, ** P < 0.01, and *** P < 0.001). E and F: Analysis of MDH1 expression by Western blot during adipocyte differenti-ation. G and H: The acetylation level of MDH1 or mutant MDH1 was assessed. Ectopically expressed MDH1 or mutant MDH1 was enriched by immunoprecipitation using an anti-FLAG antibody. Western blot analysis was then performed using an anti-acetyl-lysine antibody. I: Measurement of mRNA levels of adipogenic markers using real-time PCR. Expression levels of several adipogenic markers were assessed in 3T3-L1 cells carrying wild-type MDH1 or mutant MDH1 (3KR or K118R) during adipogenic differentiation. Adiponec-tin, resistin, C/EBP � , and PPAR � were selected as positive adipogenic markers. For comparison, data were normalized with 18s RNA. The data represent the mean percentage levels ± SD compared with day 0 control vector (n = 3).
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

1872 Journal of Lipid Research Volume 53, 2012
vector ( Fig. 5F, G ). These results strongly indicate that there is a relationship between the NADP/NADPH ra-tio and the enzymatic activity of MDH1, which is regu-lated by acetylation during adipogenesis.
Fig. 4. Continued.
The NADP/NADPH ratio was considerably decreased when wild-type MDH1 was overexpressed ( Fig. 5F, G ). In contrast, MDH1-3KR and MDH1-K118R showed a roughly similar ratio of NADP/NADPH to the control
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

Acetylation of MDH1 promotes adipogenesis 1873
suggested that acetylation plays a key role in metabolic reg-ulation. Protein acetylation is catalyzed by acetyltransferases that use acetyl-CoA as the acetylation donor. It has been shown that changes in the availability of acetyl-CoA can di-rectly affect the acetylation status of critical substrates ( 5 ). In cells, acetyl-CoA enters the TCA cycle or is used to syn-thesize fatty acids depending on the energy requirements of the cell. In the case of fatty acid synthesis, acetyl-CoA is con-verted to malonyl-CoA and donates two carbon units ( 28 ). Therefore, it is speculated that there are interconnections between adipogenesis and protein lysine acetylation.
DISCUSSION
Recent studies have suggested the existence of nonhis-tone protein acetylation. More than 2,000 acetylated proteins have been identifi ed by mass spectrometric analysis in mammalian cells ( 2–5 ). The acetylated proteins are highly conserved in human and mouse tissues. The prefer-entially acetylated proteins participate in intermediate met-abolic processes, such as glycolysis, gluconeogenesis, the TCA cycle, the urea cycle, fatty acid metabolism, nitrogen metabolism, and glycogen metabolism. Thus, it has been
Fig. 5. Effects of MDH1 acetylation on its enzymatic activity and the intracellular levels of NADPH. 3T3-L1 cells were infected with retro-viruses expressing FLAG-tagged wild-type MDH1, mutant MDH1, or the vector only. Immunoprecipitated FLAG-tagged samples were eluted by a 3×FLAG peptide. A: Overexpressed MDH1 (0 and 8 days after differentiation) was immunopurifi ed, and its acetylation status was determined using an anti-AcK antibody. B: The activity of ectopically expressed wild-type MDH1 from 3T3-L1 cells was assessed. C and D: The activity of mutant MDH1 proteins was assessed after immunopurifi cation using an anti-MDH1 antibody. E: The amount of immu-nopurifi ed MDH1 and its acetylation level were analyzed by Western blot analysis. F and G: The mutated samples were prepared on day 8 after differentiation for the NADP total (NADP t ) and NADPH-only assays. NADP/NADPH ratio is calculated as [NADP t -NADPH]/NADPH. The data represent the mean percentage levels ± SD compared with the control vector (n = 3; * P < 0.05, ** P < 0.01, and *** P < 0.001).
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

1874 Journal of Lipid Research Volume 53, 2012
conditions, the expression level of MDH1 was increased approximately 2-fold during adipocyte differentiation, and its acetylation level showed up to a 6-fold increase. This result indicates that the acetylation of MDH1 may play a crucial role in adipogenic differentiation. The ec-topic expression of MDH1 in 3T3-L1 preadipocytes was associated with the induction of adipogenesis. Under our experimental conditions, approximately 12 days are needed for the cells to fully differentiate into adipocytes after the initiation of differentiation. However, the overexpression of MDH1 induced accelerated differentiation of the cells such that differentiation was completed only 8 days after differentiation was initiated. Several recent reports have identifi ed possible acetylation sites in MDH1 by mass spec-trometric analysis of mammalian cells ( 2, 4, 5 ). We thus further investigated the role of the acetylation of MDH1 in adipogenesis using MDH1 mutants in which putative MDH1 acetylation sites were replaced by arginine or gluta-mine. In previous studies, substituting glutamine for a lysine residue was found to mimic the acetylated form ( 39 ), whereas substituting arginine was found to mimic the deacetylated form of Foxo1 and MDH2 ( 5, 32, 40–42 ). Three acetylated lysine residues (K118, K121, and K298) were identifi ed in MDH1 ( 5 ). Among the mutant MDH1 proteins that we constructed, only MDH1-3KR and K118R
Although there is evidence to support the acetylation of histone and nonhistone proteins during adipogenesis ( 29–31 ), extensive studies have not been performed. In the present study, we conducted an acetylome analysis to identify the changes in the acetylation level of nonhis-tone proteins during adipocyte differentiation. The pattern of acetylated proteins is signifi cantly different be-tween preadipocytes and mature adipocytes (supplemen-tary Fig. IE ). A total of 69 spots were detected by 2-DE Western blot analysis and identifi ed by LC-MS/MS analysis ( Table 1 ). Most of the identifi ed acetylated proteins were involved in metabolic pathways, such as gly-colysis, the TCA cycle, and fatty acid synthesis. In con-junction with earlier reports, these results suggest that acetylation is an extensive regulator of adipogenic differ-entiation ( 32–34 ). A role for acetylation in lipid metabo-lism can be inferred by the acetyl-CoA synthetase ( 30, 35 ), the bifunctional enzyme, enoyl CoA hydrolase/3-hy-droxyl CoA dehydrogenase ( 2, 5, 36 ), SIRT3 ( 31 ), and long-chain acyl CoA dehydrogenase ( 37 ).
MDH1, one of the identifi ed proteins that showed an altered acetylation pattern, is known to be a regulator that is required for calorie restriction ( 38 ). Thus, we focused on MDH1, which catalyzes the conversion of oxaloacetate into malate in the cytoplasm ( 13 ). Under our experimental
Fig. 6. The proposed mechanism of action of MDH1 acetylation during adipogenic differentiation. 1: Glucose enters the glycolytic path-way to generate pyruvate. Pyruvate is converted to acetyl-CoA in mitochondria. 2: The citrate is transported to the cytoplasm by the citrate shuttle. 3: The cytoplasmic citrate disintegrates into acetyl-CoA and oxaloacetate in the cytoplasm. This reaction provides the cytoplasmic acetyl-CoA used for fatty acid synthesis. Cytoplasmic oxaloacetate is converted to malate by MDH1, and the malate then rejoins the glyco-lytic pathway in a pyruvate form. The citrate shuttle produces two NADPH molecules per cycle. 4: The acetylation of MDH1 increases its enzymatic activity. 5: Activated MDH1 promotes the citrate shuttle that provides acetyl-CoA and NADPH for fatty acid synthesis. It is sug-gested that the acetylation of MDH1 occurs only when the cytoplasmic level of acetyl-CoA is increased above a certain level as a result of exposure to an excess energy source. Therefore, the acetylation of MDH1 functions as the cross-talk mechanism between the intracellular energy level and adipogenesis.
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

Acetylation of MDH1 promotes adipogenesis 1875
and Drs. Yoo-Sook Cho, Jeong-Ki Min, Doo Byoung Oh, and Seung-Wook Chi for carefully reading the manuscript and providing insightful comments.
REFERENCES
1 . Gershey , E. L. , G. Vidali , and V. G. Allfrey . 1968 . Chemical studies of histone acetylation. The occurrence of � - N -acetyllysine in the f2a1 histone. J. Biol. Chem. 243 : 5018 – 5022 .
2 . Choudhary , C. , C. Kumar , F. Gnad , M. L. Nielsen , M. Rehman , T. C. Walther , J. V. Olsen , and M. Mann . 2009 . Lysine acetylation tar-gets protein complexes and co-regulates major cellular functions. Science . 325 : 834 – 840 .
3 . Kim , S. C. , R. Sprung , Y. Chen , Y. Xu , H. Ball , J. Pei , T. Cheng , Y. Kho , L. Xiao , N. V. Grishin , et al . 2006 . Substrate and functional diversity of lysine acetylation revealed by a proteomics survey. Mol. Cell . 23 : 607 – 618 .
4 . Wang , Q. , Y. Zhang , C. Yang , H. Xiong , Y. Lin , J. Yo , H. Li , L. Xie , W. Zhao , Y. Yao , et al . 2010 . Acetylatioin of metabolic enzymes coordinates carbon source utilization and metabolic fl ux. Science . 327 : 1004 – 1007 .
5 . Zhao , S. , W. Xu , W. Jiang , W. Yu , Y. Lin , T. Zhang , J. Yai , L. Zhou , Y. Zeng , H. Li , et al . 2010 . Regulation of cellular metabolism by protein lysine acetylation. Science . 327 : 1000 – 1004 .
6 . Rosen , E. D. , C. J. Walkey , P. Puigserver , and B. M. Spiegelman . 2000 . Transcriptional regulation of adipogenesis. Genes Dev. 14 : 1293 – 1307 .
7 . Hausman , D. B. , M. DiGirolamo , T. J. Bartness , G. J. Hausman , and R. J. Martin . 2001 . The biology of white adipocyte proliferation. Obes. Rev. 2 : 239 – 254 .
8 . Wilson , P. W. , R. B. D’Aqostino , H. Parise , L. Sullivan , and J. B. Meiqs . 2005 . Metabolic syndrome as a precursor of cardiovascular disease and type 2 diabetes mellitus. Circulation . 112 : 3066 – 3072 .
9 . Xu , Y. , K. E. Hammerick , A. W. James , A. L. Carre , P. Leucht , A. J. Giaccia , and M. T. Longaker . 2009 . Inhibition of histone deacetylase activity in reduced oxygen environment enhances the osteogenesis of mouse adipose-derived stromal cells. Tissue Eng. Part A . 15 : 3697 – 3707 .
10 . Lagace , D. C. , and M. W. Nachtiqal . 2004 . Inhibition of histone deacetylase activity by valproic acid blocks adipogenesis. J. Biol. Chem. 279 : 18851 – 18860 .
11 . Yoo , E. J. , J. J. Chung , S. S. Choe , K. H. Kim , and J. B. Kim . 2006 . Down-regulation of histone deacetylases stimulates adipocyte dif-ferentiation. J. Biol. Chem. 281 : 6608 – 6615 .
12 . Webb , L. E. , L. E. Webb , E. J. Hill , and L. J. Banaszak . 1973 . Conformation of nicotinamide adenine dinucleotide bound to cy-toplasmic malate dehydrogenase. Biochemistry 12 : 5101 – 5109 .
13 . Minarik , P. , N. Tomaskova , M. Kollarova , and M. Antalik . 2002 . Malate dehydrogenases-structure and function. Gen. Physiol. Biophys. 21 : 257 – 265 .
14 . Musrati , R. A. , M. Kollárová , and D. Mikulásová . 1998 . Malate de-hydrogenase: distribution, function and properties. Gen. Physiol. Biophys. 17 : 193 – 210 .
15 . Joh , T. , H. Takeshima , T. Tsuzuki , C. Setoyama , K. Shimada , S. Tanase , S. Kuramitsu , H. Kaqamiyama , and Y. Morino . 1987 . Cloning and sequence analysis of cDNAs encoding mammalian cytosolic malate dehydrogenase. Comparison of the amino acid sequences of mammalian and bacterial malate dehydrogenase. J. Biol. Chem. 262 : 15127 – 15131 .
16 . Lo , A. S. , C. T. Liew , S. M. Nqai , S. K. Tsui , K. P. Fung , C. Y. Lee , and M. M. Waye . 2005 . Developmental regulation and cellular distribution of human cytosolic malate dehydrogenase (MDH1). J. Cell. Biochem. 94 : 763 – 773 .
17 . Tanaka , T. , J. Inazawa , and Y. Nakamura . 1996 . Molecular cloning and mapping of a human cDNA for cytosolic malate dehydroge-nase (MDH1). Genomics . 32 : 128 – 130 .
18 . Jung , H. , W. K. Kim , D. H. Kim , Y. S. Cho , S. J. Kim , S. G. Park , B. C. Park , H. M. Lim , K-H. Bae , and S. C. Lee . 2009 . Involvement of PTP-RQ in differentiation during adipogenesis of human mesenchymal stem cells. Biochem. Biophys. Res. Commun. 383 : 252 – 257 .
19 . Kim , W. K. , C. Y. Lee , M. S. Kang , M. H. Kim , Y. H. Ryu , K-H. Bae , S. J. Shin , S. C. Lee , and Y. Ko . 2008 . Effects of leptin on lipid metabolism and gene expression of differentiation-associated
signifi cantly suppressed the adipogenic differentiation of 3T3-L1 cells. These results show that a specifi c site (or sites) of MDH1 is acetylated during adipogenic differenti-ation and that the acetylation of MDH1 directly regulates the lipid metabolic pathway. To determine the direct ef-fect of acetylation on enzymatic activity, we fi rst confi rmed the increased acetylation level of MDH1 during adipo-genic differentiation. Consistent with endogenous MDH1 during adipogenesis, the acetylation level of ectopic MDH1 showed up to a 50% increase during adipogenesis, and this correlated with a dramatic increase in enzymatic ac-tivity. In contrast, KR mutants of MDH1 showed a de-creased acetylation level and reduced enzymatic activity when compared with wild-type MDH1. Remarkably, the acetylation of the lysine 118 residue alone can control MDH1 activity during adipogenesis. Additionally, the acet-ylation of MDH1 regulates the intracellular ratio of NADP/NADPH during fatty acid synthesis. Adipose tissue uses glucose to produce triglyceride, which is the major form of fat ( 43 ). Glucose enters the glycolytic pathway to generate pyruvate and ATP. Pyruvate is converted to acetyl-CoA, which is used as citrate with oxaloacetate in the mitochon-dria. In adipose tissue, citrate is transported to the cyto-plasm by the citrate shuttle when its concentration is high in the mitochondria. The cytoplasmic citrate disintegrates into acetyl-CoA and oxaloacetate. This reaction provides cytoplasmic acetyl-CoA that is used for fatty acid synthesis. Cytoplasmic oxaloacetate is converted to malate by MDH1, and the malate then rejoins the glycolytic pathway in a py-ruvate form. NADPH is necessary for fatty acid synthesis as a reducing equivalent. Thus, we propose that MDH1 is in-volved in the citrate shuttle that produces two NADPH molecules per one cycle ( 44, 45 ) and that the activity of MDH1 is modulated by Lys-118 acetylation during adipo-genesis. The pentose phosphate pathway also generates NADPH from glucose in most species. It has been reported that the majority of NADPH production in horse adipose tissue is due to the specifi c activity of glucose-6-phosphate dehydrogenase and 6-phospho-gluconate dehydrogenase ( 46 ). In contrast, the generation of NADPH from the malate cycle is higher than its generation from the pen-tose phosphate pathway in adipose tissue ( 47 ). Addition-ally, we speculated that adipogenesis as well as lipogenesis may be up-regulated by NADPH-dependent transcrip-tional regulators after increased intracellular NADPH level ( 48 ). In summary, depending on the cellular energy re-quirements, acetyl-CoA enters the TCA cycle or is used to synthesize fatty acids. When the cellular energy level is suf-fi cient, excess acetyl-CoA is used to acetylate MDH1. The acetylation of MDH1 increases its enzymatic activity, and activated MDH1 supports acetyl-CoA and NADPH in fatty acid synthesis by accelerating the citrate shuttle ( Fig. 6 ). In agreement with earlier reports, the present results indi-cate that the acetylation of MDH1 functions as a cross-talk mechanism between adipogenesis and the intracellular energy level.
The authors thank Professors Sayeon Cho and Byoung Chul Park for their continuous encouragement and helpful advice
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from

1876 Journal of Lipid Research Volume 53, 2012
growth factors and transcription factors during differentiation and maturation of 3T3–L1 preadipocytes. Endocr. J. 55 : 827 – 837 .
20 . Kim , W. K. , H. Jung , D. H. Kim , E. Y. Kim , J. W. Chung , Y. S. Cho , S. G. Park , B. C. Park , K-H. Bae , and S. C. Lee . 2009 . Regulation of adipocyte differentiation by LAR tyrosine phosphatase in human mesenchymal stem cells and 3T3–L1 preadipocytes. J. Cell Sci. 122 : 4160 – 4167 .
21 . Kim , W. K. , H. Jung , E. Y. Kim , D. H. Kim , Y. S. Cho , B. C. Park , S. G. Park , Y. Ko , K-H. Bae , and S. C. Lee . 2011 . RPTP � tyrosine phosphatase promotes adipogenic differentiation via modulation of p120 catenin phosphorylation. Mol. Biol. Cell . 22 : 4883 – 4891 .
22 . Kang , T. H. , K-H. Bae , M. j. Yu , W. K. Kim , H-R. Hwang , H. Jung , P. Y. Lee , S. Kang , T-S. Yoon , S. G. Park , et al . 2007 . Phosphoproteomic analysis of neuronal cell death by glutamate-induced oxidative stress. Proteomics . 7 : 2624 – 2635 .
23 . Kim , S-Y. , P. Y. Lee , H-J. Shin , D. H. Kim , S. Kang , H-B. Moon , S. W. Kang , J. M. Kim , S. G. Park , B. C. Park , et al . 2009 . Proteomic analysis of liver tissue HBx -transgenic mice at early stages of hepa-tocarcinogenesis. Proteomics . 9 : 5056 – 5066 .
24 . Jang , M. , B. C. Park , S. Kang , S-W. Chi , S. Cho , S. C. Lee , K-H. Bae , and S. G. Park . 2009 . Far upstream element-binding protein-1, a novel caspase substrate, acts as a cross-talker between apoptosis and the c-myc oncogene. Oncogene . 28 : 1529 – 1536 .
25 . Kim , S-Y. , M. J. Kim , H. Jung , W. K. Kim , S. O. Kwon , M. J. Son , I. S. Jang , J. S. Choi , S. G. Park , B. C. Park , et al . 2012 . Comparative pro-teomic analysis of human somatic cells, induced pluripotent stem cells, and embryonic stem cells. Stem Cells Dev. 10.1089/scd.2011.0243.
26 . Jeon , Y-J. , D-H. Kim , H. Jung , S. J. Chung , S-W. Chi , S. Cho , S. C. Lee , B. C. Park , S. G. Park , and K-H. Bae . 2010 . Annexin A4 interacts with the NF- � B p50 subunit and modulates NF- � B transcriptional activity in a Ca 2+ -dependent manner. Cell. Mol. Life Sci. 67 : 2271 – 2281 .
27 . Norvell , A. , and S. B. McMahon . 2010 . Cell biology. Rise of the rival. Science . 327 : 964 – 965 .
28 . Tong , L. 2005 . Acetyl-coenzyme A carboxylase: crucial metabolic enzyme and attractive target for drug discovery. Cell. Mol. Life Sci. 62 : 1784 – 1803 .
29 . Yoo , E. Y. , J. J. Chung , S. S. Choe , K. H. Kim , and J. B. Kim . 2005 . Down-regulation of histone deactylases stimulates adipocyte differ-entiation. J. Biol. Chem. 281 : 6608 – 6615 .
30 . Schwer , B. , J. Bunkenborg , R. Q. Verdin , J. S. Andersen , and E. Verdin . 2006 . Reversible lysine acetylation controls the activity of the mitochondrial enzyme acetyl-CoA synthetase 2. Proc. Natl. Acad. Sci. USA . 103 : 10224 – 10229 .
31 . Lombard , D. B. , F. W. Alt , H. L. Cheng , J. Bunkenborg , R. S. Streeper , R. Mostoslavsky , J. Kim , G. Yancopoulos , D. Valenzuela , A. Murphy , et al . 2007 . Mammalian Sir2 homolog SIRT3 regulates global mitochondrial lysine acetylation. Mol. Cell. Biol. 27 : 8807 – 8814 .
32 . Natambi , J. M. , and Y. Kim . 2000 . Adipocyte differentiation and gene expression. J. Nutr. 130 : 3122 – 3126 .
33 . Rahman , M. M. , A. Kukita , T. Kukita , T. Shobuike , T. Nakamura , and O. Kohashi . 2003 . Two histone deacetylase inhibitors, tricho-stain A and sodium butyrate, suppress differentiation into osteo-clasts but not into macrophages. Blood . 101 : 3451 – 3459 .
34 . Catalioto , R. M. , C. A. Maggi , and S. Giuliani . 2009 . Chemically dis-tinct HDAC inhibitors prevent adipose conversion of subcutaneous human white preadipocytes at an early stage of the differentiation program. Exp. Cell Res. 315 : 3267 – 3280 .
35 . Hallows , W. C. , S. Lee , and J. M. Denu . 2006 . Sirtuins deacetylate and activate mammalian acetyl-CoA synthetases. Proc. Natl. Acad. Sci. USA . 103 : 10230 – 10235 .
36 . Schwer , B. , M. Eckersdorff , Y. Li , J. C. Silva , D. Fermin , M. V. Kurtev , C. Giallourakis , M. J. Comb , F. W. Alt , and D. B. Lombard . 2009 . Calorie restriction alters mitochondrial protein acetylation. Aging Cell . 8 : 604 – 606 .
37 . Ahn , B. H. , H. S. Kim , S. Song , I. H. Lee , J. Liu , A. Vassilopoulos , C. X. Deng , and T. Finkei . 2008 . A role for the mitochondrial deacetylase Sirt3 in regulating energy homeostasis. Proc. Natl. Acad. Sci. USA . 105 : 14447 – 14452 .
38 . Easlon , E. , F. Tsang , C. Skinner , C. Wang , and S. J. Lin . 2008 . The malate-aspartate NADH shuttle components are novel metabolic longevity regulators required for calorie restriction-mediated life span extension in yeast. Genes Dev. 22 : 931 – 944 .
39 . Wang , Y. H. , Y. G. Tsay , B. C. Tan , W. Y. Lo , and S. C. Lee . 2003 . Identifi cation and characterization of a novel p300-mediated p53 acetylation site, Lys 305. J. Biol. Chem. 278 : 25568 – 25576 .
40 . Feng , L. , T. Lin , H. Uranishi , W. Gu , and Y. Xu . 2005 . Functional analysis of the roles of posttranslational modifi cations at the p53 C-terminus in regulating p53 stability and activity. Mol. Cell. Biol. 25 : 5389 – 5395 .
41 . Marcotte , P. A. , P. L. Richardson , J. Guo , L. W. Barrett , N. Xu , A. Gunasekera , and K. B. Glaser . 2004 . Fluorescence assay of SIRT protein deacetylases using an acetylated peptide substrate and a secondary trypsin reaction. Anal. Biochem. 332 : 90 – 99. [Erratum. 2006. Anal. Biochem . 350: 316.]
42 . Jing , E. , S. Gesta , and C. R. Kahn . 2007 . SIRT2 regulates adipo-cyte differentiation through FoxO1 acetylation/deacetylation. Cell Metab. 6 : 105 – 114 .
43 . Flatt , J. P. 1970 . Conversion of carbohydrate to fat in adipose tissue: an energy-yielding and, therefore, self-limiting process. J. Lipid Res. 11 : 131 – 143 .
44 . Ingle , D. L. , D. E. Bauman , and U. S. Garrigus . 1972 . Lipogenesis on the ruminant: in vitro study of tissue sites, carbon source and reducing equivalent generation for fatty acid synthesis. J. Nutr. 102 : 609 – 616 .
45 . Pearson , D. J. , and S. S. Malde . 1985 . Cytoplasmic NADP-linked dehydrogenase activities in avian tissue. Comp. Biochem. Physiol. B . 81 : 727 – 731 .
46 . Suagee , J. K. , B. A. Corl , M. V. Crisman , J. G. Wearn , L. J. McCutcheon , and R. J. Geor . 2010 . De novo fatty acid synthesis and NADPH generation in equine adipose and liver tissue. Comp. Biochem. Physiol. B . 155 : 322 – 326 .
47 . Fell , D. A. , and J. R. Small . 1986 . Fat synthesis in adipose tissue. Biochem. J. 238 : 781 – 786 .
48 . Park , J. , H. K. Rho , K. H. Kim , S. S. Choe , Y. S. Lee , and J. B. Kim . 2005 . Overexpression of glucose-6-phosphate dehydrogenase is as-sociated with lipid dysregulation and insulin resistance in obesity. Mol. Cell. Biol. 25 : 5146 – 5157 .
by guest, on Decem
ber 4, 2018w
ww
.jlr.orgD
ownloaded from





























