Embed Size (px)
Citation preview

LETTERS
Abscisic acid controls calcium-dependent egress anddevelopment in Toxoplasma gondiiKisaburo Nagamune1{, Leslie M. Hicks2, Blima Fux1, Fabien Brossier1{, Eduardo N. Chini3 & L. David Sibley1
Calcium controls a number of critical events, including motility,secretion, cell invasion and egress by apicomplexan parasites1.Compared to animal2 and plant cells3, the molecular mechanismsthat govern calcium signalling in parasites are poorly understood.Here we show that the production of the phytohormone abscisicacid (ABA) controls calcium signalling within the apicomplexanparasite Toxoplasma gondii, an opportunistic human pathogen. Inplants, ABA controls a number of important events, includingenvironmental stress responses, embryo development and seeddormancy4,5. ABA induces production of the second-messengercyclic ADP ribose (cADPR), which controls release of intracellularcalcium stores in plants6. cADPR also controls intracellular cal-cium release in the protozoan parasite T. gondii7,8; however, pre-vious studies have not revealed the molecular basis of thispathway9. We found that addition of exogenous ABA inducedformation of cADPR in T. gondii, stimulated calcium-dependentprotein secretion, and induced parasite egress from the infectedhost cell in a density-dependent manner. Production of endogen-ous ABA within the parasite was confirmed by purification (usinghigh-performance liquid chromatography) and analysis (by gaschromatography-mass spectrometry). Selective disruption ofABA synthesis by the inhibitor fluridone delayed egress andinduced development of the slow-growing, dormant cyst stage ofthe parasite. Thus, ABA-mediated calcium signalling controls thedecision between lytic and chronic stage growth, a developmentalswitch that is central in pathogenesis and transmission. The path-way for ABA production was probably acquired with an algalendosymbiont that was retained as a non-photosynthetic plastidknown as the apicoplast. The plant-like nature of this pathway maybe exploited therapeutically, as shown by the ability of a specificinhibitor of ABA synthesis to prevent toxoplasmosis in the mousemodel.
Calcium-mediated protein secretion in T. gondii controls bothmotility and cell invasion, and previous studies have demonstratedthat these processes utilize the second messenger cADPR, yet thesignals triggering this pathway remain unresolved7,8. In plants6,hydra10 and sponges11, ABA stimulates release of intracellular calciumthrough elevation of the cyclic nucleotide cADPR. Addition of exo-genous ABA proved to be a potent agonist of secretion in T. gondii asshown by the release of the protein MIC2 (microneme protein 2), aparasite adhesin that is discharged into the supernatant in response toincreases in intracellular calcium (Fig. 1a). Induction of MIC2 secre-tion by ABA was highly specific to (6)-ABA and was not inducedby (–)-ABA, the precursor b-carotene, or retinoic acid (Fig. 1b). Inthis regard, the response of T. gondii to ABA shows similar specificityto higher plants5. Treatment with ABA lead to a dose-dependentincrease in the second messenger cADPR in T. gondii, suggesting that
ABA may be a natural agonist for calcium signalling in parasites(Fig. 1c). Finally, chelation of intracellular calcium in the parasiteblocked secretion induced by ABA, confirming that it acts throughrelease of an intracellular calcium pool (Fig. 1d). Collectively, theseresults indicate that ABA in T. gondii controls release of calcium fromintracellular stores through production of cADPR, thus parallelingthe pathway seen in plants6, and early metazoans10,11.
Consistent with its physiological effects, ABA was detectedin lysates of T. gondii by direct biochemical analyses. Extracts of
1Department of Molecular Microbiology, Washington University School of Medicine, 660 S. Euclid Avenue, St Louis, Missouri 63110, USA. 2Donald Danforth Plant Science Center, 975N. Warson Road, St Louis, Missouri 63132, USA. 3Department of Anesthesiology, Mayo Clinic and Foundation, Rochester, Minnesota 55905, USA. {Present addresses: Department ofImmunoregulation, Research Institute for Microbial Diseases, Osaka University, Osaka 565-0871, Japan (K.N.); Linstitut National de la Researche Agronomique, Centre de Recherchede Tours, 37380 Nouzilly, France (F.B.).
a
123
80
123
0 1 10 100 1,000 EtOH
5 10 5 10 5 10 5 10 5 10 5 10 cMIC
2
Incubation (min)
Supernatant
Cell pelletAnti-β-Gal
** **
b
c
Supernatant
Cell pellet
175
83
175
83
(±)-ABABAPTA-AM
BAPTA – –––
– – ––
– – – –
– –++++
++
EtOH cMIC
2
150
100
150
0.1 1.0 0.1 1.0 0.1 0.11.0 1%
InducerConcentration (µM)
Supernatant
Cell pellet
3.53.02.52.01.51.00.5
00 10 100 1,000
d
(±)-ABA (nM)
Anti-MIC2 (kDa)
Anti-β-Gal
Anti-β-Gal
Anti-MIC2 (kDa)
Anti-MIC2
β-caroteneRetinoic
acid (–)-ABA (±)-A
BA
EtOH
(–)(–) cM
IC2
cAD
PR
pro
duc
tion
(pm
ol p
er m
g p
rote
in)
Treatment with (±)-ABA (nM)
(kDa)
Figure 1 | ABA induced cADPR production and calcium-dependent proteinsecretion by T. gondii. a, Protein secretion induced by ABA was compared tothe positive control ethanol (EtOH) as detected by western blotting withanti-MIC2 (supernatant) and anti-b-galactosidase antibodies (cell pellet)7,8.b, Induction of MIC2 secretion was specific for (6)-ABA and was not seenwith (–)-ABA, b-carotene, or retinoic acid. c, cADPR production in T. gondiiwas increased by treatment with ABA. Student’s t-test; *P , 0.02, **P , 0.01versus control, mean 6 s.e.m., N 5 3 experiments. d, ABA-induced secretionrequired intracellular calcium. Treatment with the membrane permeablecalcium chelator, BAPTA-AM, blocked ABA-induced MIC2 release, whereasnon-permeant BAPTA had no effect. Cellular MIC2 (cMIC2) migratesslower owing to absence of processing.
Vol 451 | 10 January 2008 | doi:10.1038/nature06478
207Nature ©2007 Publishing Group

T. gondii were purified by high-performance liquid chromatography(HPLC) and analysed by gas chromatography to detect the methylester of ABA (methyl-ABA), which migrated with the same retentiontime (8 min) as the authentic methyl-ABA standard (Fig. 2a). Massspectrometric analysis of T. gondii samples yielded the expectedproduct of mass-to-charge ratio (m/z) 190 as well as peaks atm/z 91 and 162, characteristic of cis-ABA, and m/z 91 and 134, char-acteristic of trans-ABA, as described previously12 (Fig. 2b and Supple-mentary Fig. 1). The slightly nosier spectrum obtained from T. gondiiis consistent with the lower level of ABA found in the sample, none-theless its signature unambiguously matches ABA. Similar levels ofABA were detected in parasite extracts using a highly specific ELISA(enzyme-linked immunosorbent assay) kit (Fig. 2c). To confirm thatABA was indeed synthesized by the parasite, we used the herbicidefluridone, an inhibitor that specifically blocks phytoene destauraseactivity, and hence prevents synthesis of ABA by the indirect pathwaythat predominates in plants5. Treatment with fluridone reducedthe level of ABA in parasites by more than fourfold (Fig. 2c).Interestingly, when the level of ABA was monitored during one cycleof intracellular growth, it remained relatively constant during thefirst 30–36 h and then spiked shortly before egress of parasites fromthe infected host cell, which begins around 40–44 h (Fig. 2d).
Toxoplasma is an obligate intracellular pathogen and followinginvasion, the parasite replicates by binary fission within a vacuole,ultimately leaving the depleted host cell by active egress. Intracellularcalcium is central for several of these steps, leading us to examinewhether ABA-dependent calcium signalling was essential for growthin vitro. Treatment of parasite cultures with increasing concentra-tions of fluridone led to inhibition of parasite growth (effector con-centration for half-maximum response, EC50, ,15 mM; Fig. 3a).Surprisingly, fluridone treatment did not block invasion or replica-tion, but instead inhibited exit of mature parasites from the infectedcell. Whereas parasites had naturally egressed by ,60 h, those treatedwith 15 mM fluridone were delayed, and those treated with 50 mMfluridone were completely blocked out to 84 h (Fig. 3b), after whichthe integrity of the monolayer declined (data not shown). Previousstudies have implicated calcium signalling in parasite egress frominfected cells13,14, and the rapid increase in ABA concentrations inthe parasite just before egress (Fig. 2d) suggested this molecule maybe the natural signal for activating exit. Consistent with this, whenexogenous ABA was added to late-stage vacuoles, it induced pre-mature egress (Fig. 3c). Finally, ABA was able to overcome the
inhibition of egress caused by treatment with fluridone (Fig. 3d),consistent with it being able to bypass the metabolic block in biosyn-thesis. Although the responses to exogenously added ABA are some-what modest, this may reflect the relatively low permeability of thecharged anion that exists at physiological pH (pKa 4.8), and the factthat it has to cross several membranes to reach the parasite. WhetherABA produced by the parasite accumulates within the parasite or isexported to the parasite-containing vacuole is unknown, and themechanism(s) whereby ABA is sensed remain to be elucidated.
c250
100
50
T.gondii+ fluridone
200
0
150
*P < 0.005
a Toxoplasma
48
121620
Methyl-ABA
6.0 7.0 8.0 9.0 10.0 11.0 12.0 6.0 7.0 8.0 9.0 10.0 11.0 12.0
10
20
30
40
Time (min) Time (min)
100 200 30010
30507090
110
m/z100 200 300
m/z
190
134
16255 91106
13579
1113
190
57162
91134119
bd
0 10 20 30
4
3
2
1
Incubation period (h)
Egress
Ab
und
ance
× 1
04A
bun
dan
ce ×
102
AB
A (µ
M)
AB
A (n
M)
T.gondii
40
Figure 2 | Biochemical evidence for the production of ABA in T. gondii.a, Gas chromatography traces of ABA standards (left) and T. gondii samples(right). A single peak with similar retention time (8.0 min) to authenticmethyl-ABA was detected in extracts of the parasite following chemicalextraction and methylation. b, Mass spectra of methyl-ABA standard and T.gondii sample from a, showing characteristic fragment at m/z 190, with
additional minor peaks in common. c, Detection of ABA by ELISA in controlparasites or treated with 50mM of fluridone. Student’s t-test; *P , 0.005versus control, mean 6 s.e.m., N 5 3 experiments. d, Time course of ABAproduction by T. gondii. Values are intracellular concentration in theparasite. Time 0 refers to newly invaded parasites at the start of infection.Mean 6 s.e.m., N 5 3 experiments.
(±)-ABA
Hours
0
22
*
*
*****
12
10
8
6
4
2
1
2
4
8
16
32
64
0 12 24 36 48 60 72 84Incubation period (h)
(±)-ABAFluridone (µM) 0 50
*****
*
25
50
75
100
a b
c d
15 µM0 µM fluridone
50 µM
(µM)
100
75
50
25
0
Fluridone concentration (µM)0.1 1.0 10 100
Egress
Intact
Par
asite
gro
wth
(%)
Egr
esse
d v
acuo
les
(%)
Egr
esse
d v
acuo
les
(%)
Num
ber
of p
aras
ites
per
vac
uole
25 28 30
101 0 101 0 101 0 10 1001 0 10 10010 101
Figure 3 | Fluridone blocked parasite growth by preventing egress.a, Fluridone prevented parasite growth as monitored by b-galactosidaseactivity in the parasite. b, Continuous culture in fluridone did not affectreplication but inhibited egress from the vacuole. c, Treatment withexogenous ABA induced egress of parasites from mature vacuoles. Hoursrefers to length of culture before addition of ABA for 1 h and scoring ofegress. Student’s t-test; *P , 0.02, **P , 0.05. d, ABA reversed theinhibition of parasite egress caused by fluridone. Parasite-infected cells weretreated with fluridone for 38 h, then stimulated with ABA for 1 h, andevaluated for egress. Student’s t-test; *P , 0.002, **P , 0.05, ***P , 0.02.a–d, Data shown are means 6 s.e.m., N 5 3 experiments.
LETTERS NATURE | Vol 451 | 10 January 2008
208Nature ©2007 Publishing Group
Nonetheless, these results indicate that ABA is a natural agonist forhost cell egress and that in the absence of its production, parasitesremain quiescent within the host cell.
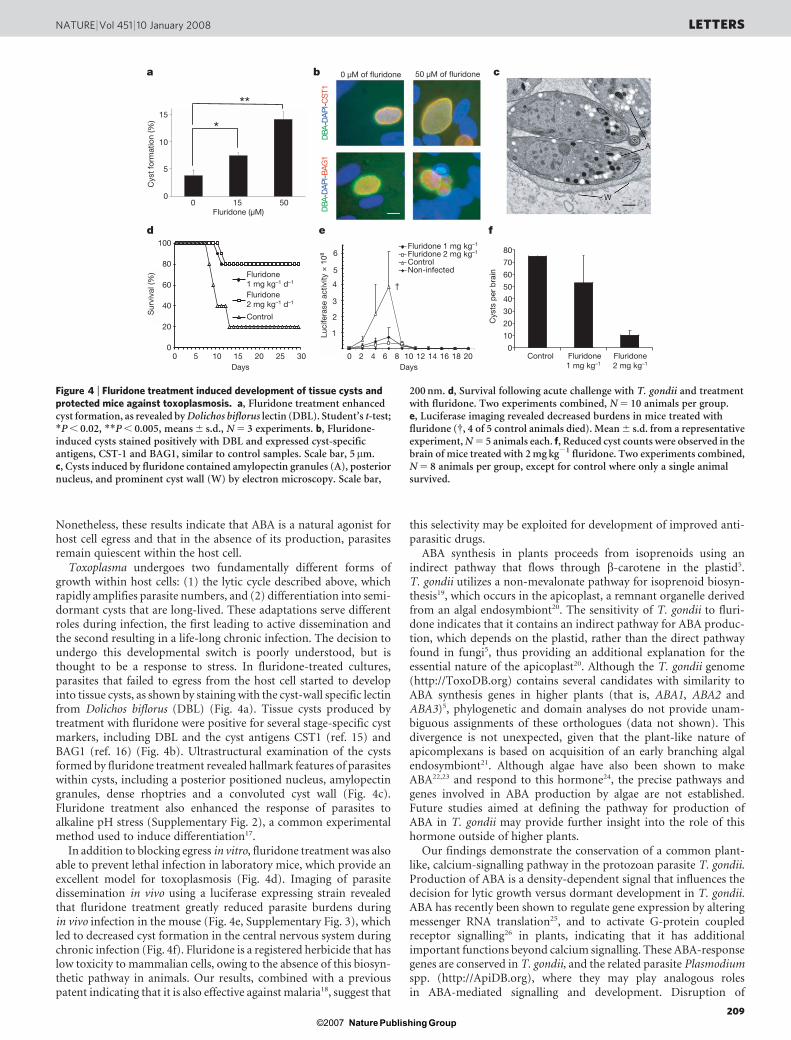
Toxoplasma undergoes two fundamentally different forms ofgrowth within host cells: (1) the lytic cycle described above, whichrapidly amplifies parasite numbers, and (2) differentiation into semi-dormant cysts that are long-lived. These adaptations serve differentroles during infection, the first leading to active dissemination andthe second resulting in a life-long chronic infection. The decision toundergo this developmental switch is poorly understood, but isthought to be a response to stress. In fluridone-treated cultures,parasites that failed to egress from the host cell started to developinto tissue cysts, as shown by staining with the cyst-wall specific lectinfrom Dolichos biflorus (DBL) (Fig. 4a). Tissue cysts produced bytreatment with fluridone were positive for several stage-specific cystmarkers, including DBL and the cyst antigens CST1 (ref. 15) andBAG1 (ref. 16) (Fig. 4b). Ultrastructural examination of the cystsformed by fluridone treatment revealed hallmark features of parasiteswithin cysts, including a posterior positioned nucleus, amylopectingranules, dense rhoptries and a convoluted cyst wall (Fig. 4c).Fluridone treatment also enhanced the response of parasites toalkaline pH stress (Supplementary Fig. 2), a common experimentalmethod used to induce differentiation17.
In addition to blocking egress in vitro, fluridone treatment was alsoable to prevent lethal infection in laboratory mice, which provide anexcellent model for toxoplasmosis (Fig. 4d). Imaging of parasitedissemination in vivo using a luciferase expressing strain revealedthat fluridone treatment greatly reduced parasite burdens duringin vivo infection in the mouse (Fig. 4e, Supplementary Fig. 3), whichled to decreased cyst formation in the central nervous system duringchronic infection (Fig. 4f). Fluridone is a registered herbicide that haslow toxicity to mammalian cells, owing to the absence of this biosyn-thetic pathway in animals. Our results, combined with a previouspatent indicating that it is also effective against malaria18, suggest that
this selectivity may be exploited for development of improved anti-parasitic drugs.
ABA synthesis in plants proceeds from isoprenoids using anindirect pathway that flows through b-carotene in the plastid5.T. gondii utilizes a non-mevalonate pathway for isoprenoid biosyn-thesis19, which occurs in the apicoplast, a remnant organelle derivedfrom an algal endosymbiont20. The sensitivity of T. gondii to fluri-done indicates that it contains an indirect pathway for ABA produc-tion, which depends on the plastid, rather than the direct pathwayfound in fungi5, thus providing an additional explanation for theessential nature of the apicoplast20. Although the T. gondii genome(http://ToxoDB.org) contains several candidates with similarity toABA synthesis genes in higher plants (that is, ABA1, ABA2 andABA3)5, phylogenetic and domain analyses do not provide unam-biguous assignments of these orthologues (data not shown). Thisdivergence is not unexpected, given that the plant-like nature ofapicomplexans is based on acquisition of an early branching algalendosymbiont21. Although algae have also been shown to makeABA22,23 and respond to this hormone24, the precise pathways andgenes involved in ABA production by algae are not established.Future studies aimed at defining the pathway for production ofABA in T. gondii may provide further insight into the role of thishormone outside of higher plants.
Our findings demonstrate the conservation of a common plant-like, calcium-signalling pathway in the protozoan parasite T. gondii.Production of ABA is a density-dependent signal that influences thedecision for lytic growth versus dormant development in T. gondii.ABA has recently been shown to regulate gene expression by alteringmessenger RNA translation25, and to activate G-protein coupledreceptor signalling26 in plants, indicating that it has additionalimportant functions beyond calcium signalling. These ABA-responsegenes are conserved in T. gondii, and the related parasite Plasmodiumspp. (http://ApiDB.org), where they may play analogous rolesin ABA-mediated signalling and development. Disruption of
b 0 µM of fluridone 50 µM of fluridone
AB
D-
IP
AD
-1
GA
BA
BD
-I
PA
D-
1TS
C
c
W
A
0
10
20
30
40
50
60
70
80
Control
fe
Days
Fluridone 1 mg kg–1
Fluridone1 mg kg–1
Fluridone 2 mg kg–1
Fluridone2 mg kg–1
ControlNon-infected
6
5
4
3
2
1
†
0
5
10
15
0 15 50
*
**
Fluridone (µM)
Cys
t fo
rmat
ion
(%)
Cys
ts p
er b
rain
Sur
viva
l (%
)
Luci
fera
se a
ctiv
ity ×
108
a
0
20
40
60
80
100
0 105 15 20 25 30 0 42 6 8 10 12 14 16 18 20Days
Fluridone1 mg kg–1 d–1
Fluridone2 mg kg–1 d–1
Control
d
Figure 4 | Fluridone treatment induced development of tissue cysts andprotected mice against toxoplasmosis. a, Fluridone treatment enhancedcyst formation, as revealed by Dolichos biflorus lectin (DBL). Student’s t-test;*P , 0.02, **P , 0.005, means 6 s.d., N 5 3 experiments. b, Fluridone-induced cysts stained positively with DBL and expressed cyst-specificantigens, CST-1 and BAG1, similar to control samples. Scale bar, 5mm.c, Cysts induced by fluridone contained amylopectin granules (A), posteriornucleus, and prominent cyst wall (W) by electron microscopy. Scale bar,
200 nm. d, Survival following acute challenge with T. gondii and treatmentwith fluridone. Two experiments combined, N 5 10 animals per group.e, Luciferase imaging revealed decreased burdens in mice treated withfluridone ({, 4 of 5 control animals died). Mean 6 s.d. from a representativeexperiment, N 5 5 animals each. f, Reduced cyst counts were observed in thebrain of mice treated with 2 mg kg21 fluridone. Two experiments combined,N 5 8 animals per group, except for control where only a single animalsurvived.
NATURE | Vol 451 | 10 January 2008 LETTERS
209Nature ©2007 Publishing Group

ABA-mediated signalling in parasites offers a promising new targetfor development of improved interventions aimed at combatinginfection with apicomplexan parasites.
METHODS SUMMARYGrowth and egress assays. Intracellular parasite growth was monitored using
b-galactosidase activity7, or by microscopic examination determination of the
number of parasites per vacuole13,17. Egress was monitored by the percentage of
intact vacuoles after culturing for defined intervals with and without fluridone,
followed by treatment with ABA.
Secretion assay. Calcium-dependent secretion was monitored by release of
MIC2 (refs 7, 27), with b-galactosidase as a cell-loading and non-specific lysis
control.
ABA purification and analysis. ABA was extracted from parasites, purified by
HPLC, converted to methyl-ABA28, and analysed by tandem mass spectrometry
using an Agilent Technologies GC-MS instrument to detect the diagnosticfragments12.
ABA detection by ELISA. ABA concentrations in parasite extracts were deter-
mined by ELISA using the Phytodetek ABA Test Kit (Agdia) after subtraction ofthe background level of ABA in cultured host cells processed in parallel, and
expressed as intracellular concentrations29.
Cyclic ADPR. Freshly harvested parasites were snap frozen at –80 uC, extracted,
and cADPR levels determined as described previously8.
In vitro differentiation. Cyst formation was monitored following treatmentwith fluridone under normal culture conditions as detected using FITC-labelled
DBL (Sigma) or antibodies to the cyst antigens CST1 (ref. 15) or BAG1 (ref. 16).
Transmission electron microscopy was performed as described previously17.
Mouse infections. BALB/c mice were infected with type II strain PTG strain by
intraperitoneal (i.p.) injection and treated with fluridone by daily i.p. injection
for 12 d. To monitor tissue burdens, mice were challenged with the luciferase
expressing Pru-Luc strain and imaged as described previously30.
Full Methods and any associated references are available in the online version ofthe paper at www.nature.com/nature.
Received 2 October; accepted 13 November 2007.
1. Moreno, S. N. J. & Docampo, R. Calcium regulation in protozoan parasites. Curr.Opin. Microbiol. 6, 359–364 (2003).
2. Berridge, M. J., Bootman, M. D. & Roderick, H. L. Calcium signaling: Dynamics,homeostasis and remodelling. Nature Rev. Mol. Cell Biol. 4, 517–529 (2003).
3. Bothwell, J. H. F. & Ng, C. K. Y. The evolution of Ca21 signaling in photosyntheticeukaryotes. New Phytol. 166, 21–38 (2005).
4. Xiong, L. & Zhu, J. K. Regulation of abscisic acid biosynthesis. Plant Physiol. 133,29–36 (2003).
5. Schwartz, S. H., Qin, X. & Zeevaart, J. A. D. Elucidation of the indirect pathway ofabscisic acid biosynthesis by mutants, genes, and enzymes. Plant Physiol. 131,1591–1601 (2003).
6. Wu, Y. et al. Abscisic acid signaling through cyclic ADP ribose in plants. Science278, 2126–2130 (1997).
7. Lovett, J. L., Marchesini, N., Moreno, S. N. & Sibley, L. D. Toxoplasma gondiimicroneme secretion involves intracellular Ca21 release from IP3 / ryanodinesensitive stores. J. Biol. Chem. 277, 25870–25876 (2002).
8. Chini, E. N., Nagamune, K., Wetzel, D. M. & Sibley, L. D. Evidence that the cADPRsignaling pathway controls calcium-mediated secretion in Toxoplasma gondii.Biochem. J. 389, 269–277 (2005).
9. Nagamune, K. & Sibley, L. D. Comparative genomic and phylogenetic analyses ofcalcium ATPases and calcium-regulated proteins in the Apicomplexa. Mol. Biol.Evol. 23, 1613–1627 (2006).
10. Puce, S. et al. Abscisic acid signaling through cyclic ADP-ribose in hydroidregeneration. J. Biochem. 279, 39783–39788 (2004).
11. Zocchi, E. et al. The temperature-signaling cascade in sponges involves a heat-gated cation channel, abscisic acid, and cyclic ADP ribose. Proc. Natl Acad. Sci.USA 98, 14859–14864 (2001).
12. de Almeida, J. A. S., Kascheres, C. & Pereira, M. D. A. Ethylene and abscisic acid inthe control of development of the rhizome of Koleria eriantha (Benth.) Hanst.(Gesneriaceae). Braz. J. Plant Physiol. 17, 391–399 (2005).
13. Moudy, R., Manning, T. J. & Beckers, C. J. The loss of cytoplasmic potassium uponhost cell breakdown triggers egress of Toxoplasma gondii. J. Biol. Chem. 276,41492–41501 (2001).
14. Endo, T., Sethi, K. K. & Piekarski, G. Toxoplasma gondii: Calcium ionophoreA23187-mediated exit of trophozoites from infected murine macrophages. Exp.Parasitol. 53, 179–188 (1982).
15. Zhang, Y. W., Halonen, S. K., Ma, Y. F., Wittner, M. & Weiss, L. M. Initialcharacterization of CST1, a Toxoplasma gondii cyst wall glycoprotein. Infect.Immun. 69, 501–507 (2001).
16. Zhang, Y. W. et al. Disruption of the Toxoplasma gondii bradyzoite-specific geneBAG1 decreases in vivo cyst formation. Mol. Microbiol. 31, 691–701 (1999).
17. Fux, B. et al. Toxoplasma gondii strains defective in oral transmission are alsodefective in developmental stage differentiation. Infect. Immun. 75, 2580–2590(2007).
18. Leef, J. L. & Carlson, P. S. Carotenoid synthesis inhibiting herbicides and fatty acidsynthesis oxime herbicides as anti-apicomplexa protozoan parasite agents.Patent 847932, 1–8 (Potomax Ltd. Prtn) (1999).
19. Jomaa, H. et al. Inhibitors of the non-mevalonate pathway of isoprenoidbiosynthesis as antimalarial drugs. Science 285, 1573–1576 (1999).
20. Ralph, S. A. et al. Metabolic maps and functions of the Plasmodium falciparumapicoplast. Nature Rev. Microbiol. 2, 203–216 (2004).
21. Waller, R. F. & McFadden, G. I. The apicoplast: A review of the derived plastid ofapicomplexan parasites. Curr. Iss. Mol. Biol. 7, 57–79 (2005).
22. Hirsch, R., Hartung, W. & Gimmler, H. Abscisic acid content of algae under stress.Bot. Acta 102, 326–334 (1989).
23. Cowan, A. K. & Rose, P. D. Abscisic acid metabolism in salt stressed cells ofDunaliella salina. Plant Physiol. 97, 798–803 (1991).
24. Kobayashi, M., Hirai, N., Kurimura, Y., Ohigashi, H. & Tsuji, Y. Abscisic acid-dependent algal morphogenesis in the unicellular green alga Haematococcuspluvialis. Plant Growth Reg. 22, 79–85 (1997).
25. Razem, F. A., El-Kereamy, A., Abrams, S. R. & Hill, R. D. The RNA-binding proteinFCA is an abscisic acid receptor. Nature 439, 290–294 (2006).
26. Liu, X. et al. A G protein-coupled receptor is a plasma membrane receptor for theplant hormone abscisic acid. Science 315, 1712–1716 (2007).
27. Carruthers, V. B., Sherman, G. D. & Sibley, L. D. The Toxoplasma adhesive proteinMIC2 is proteolytically processed at multiple sites by two parasite-derivedproteases. J. Biol. Chem. 275, 14346–14353 (2000).
28. Tian, L., DellaPenna, D. & Zeevaart, J. A. D. Effect of hydroxylated carotenoiddeficiency on ABA accumulation in Arabidopsis. Physiol. Plant. 122, 314–320(2004).
29. Rodrigues, C. O., Ruiz, F. A., Rohloff, P., Scott, D. A. & Moreno, S. N. J.Characterization of isolated acidocalcisomes from Toxoplasma gondii tachyzoitesreveals a novel pool of hydrolyzable polyphosphate. J. Biol. Chem. 277,48650–48656 (2002).
30. Saeij, J. P., Boyle, J. P., Grigg, M. E., Arrizabalaga, G. & Boothroyd, J. C.Bioluminescence imaging of Toxoplasma gondii infection in living mice revealsdramatic differences between strains. Infect. Immun. 73, 695–702 (2005).
Supplementary Information is linked to the online version of the paper atwww.nature.com/nature.
Acknowledgements We thank J. Zeevaart for conducting the initial purificationand analysis of ABA and for supplying standards; J. Boothroyd and L. Weiss forproviding reagents; W. Beatty, S. Moreno, B. Striepen, A. Waters and L. Xiong forcomments; and J. Nawas and D. Gill for technical assistance. This work wassupported by the Uehara Medical Foundation (K.N.), the Mayo Clinic andAmerican Heart Association (E.N.C.) and the NIH (L.D.S.).
Author Contributions K.N. performed the experiments on the effects of ABA andfluridone on the parasite, L.M.H. performed the MS studies, B.F. performed theanimal studies, F.B. contributed to the analysis of ABA genes, E.N.C. performed themeasurements of cADPR, L.D.S. supervised the project and wrote the manuscriptwith input from all the authors.
Author Information Reprints and permissions information is available atwww.nature.com/reprints. Correspondence and requests for materials should beaddressed to L.D.S. ([email protected]).
LETTERS NATURE | Vol 451 | 10 January 2008
210Nature ©2007 Publishing Group

METHODSChemicals. Chemicals were purchased from Sigma-Aldrich, including: (6)-
abscisic acid, referred to as (6)-ABA or ABA, (2)-ABA, retinoic acid, and
b-carotene. BAPTA and BAPTA-AM were obtained from Molecular Probes.
Fluridone was obtained from Chem Service Inc. Chemicals were dissolved in
DMSO and stored at –80 uC.
Protein secretion. To examine the induction of microneme protein secretion,
purified 2F strain parasites, which express b-galactosidase, were treated with
compounds for 5–10 min at 18 uC, and separated into supernatant and cell
pellet by centrifugation. MIC2 secretion was detected by western blottingwith anti-MIC2 rabbit antibody (supernatant). Cell pellets were probed with
anti-b-galactosidase monoclonal antibody 40a-1 as a control for loading and
inadvertent lysis. Secreted MIC2 in the supernatant migrates slightly faster than
cellular MIC2 (cMIC2) owing to proteolysis during shedding27. To test the roles
of extracellular and intracellular calcium, respectively, purified parasites were
treated with 1 mM of BAPTA or 50mM of BAPTA-AM for 20 min at 18 uC before
treatment with 100 nM of ABA for 5 min, as described previously7.
Growth and egress assays. Parasite replication and egress were monitored by
microscopic examination of parasites grown in human foreskin fibroblast (HFF)
cells cultured on glass coverslips. Following culture for specific time intervals, the
number of intracellular parasites per vacuole, and the percentage of intact versus
disrupted vacuoles (egress), was determined by examining specific antibody-
stained cells (that is, SAG1 surface staining, DAPI nuclear staining) under
epifluorescence microscopy, as described previously13,17. Egress was monitored
following culture in normal medium for defined intervals followed by stimu-
lation with ABA for 1 h, and microscopic examination to determine the percent-
age of intact vacuoles as above. To test the ability of ABA to overcome inhibition
of egress by fluridone, cells were cultured in control medium or medium con-taining fluridone for 38 h, then treated with ABA for 1 h, and examined micro-
scopically to determine the percentage of intact vacuoles as above.
GC-MS analysis of ABA. Parasites from heavily infected cultures at 30–36 h post
inoculation were snap frozen in an ethanol-dry ice bath, lyophilized, and stored
at –80 uC. Tissues were extracted in organic solvents and resolved by HPLC using
a C18 reverse phase column as described previously28. Fractions containing ABA
were collected, dried under vacuum, methylated using diazomethane, and ana-
lysed by gas chromatography in comparison to standards for methyl-ABA (pro-
vided by J. Zeevaart), as described previously28. Tandem mass spectrometry
analyses were performed on an Agilent Technologies GC-MS (6890N network
GC system). Samples (dissolved in 10ml) and standards (appropriate dilutions)
were solubilized in ethyl acetate, and injected into the GC-MS instrument. The
samples were introduced through a DB-5 capillary column (30 m 3 0.250 mm
i.d., film thickness 0.25mm (J&W Scientific)) with helium as the carrier gas (flow
rate 1.3 ml min21). The GC oven was programmed from 60 to 210 uC at
25 uC min21 and from 210 to 280 uC at 12 uC min21. The ion-source temper-
ature was 150 uC. The mass analyser was scanned from m/z 50 to 400 at a data rate
of 20 Hz for the full scan analyses, and m/z 190 at a data rate of 20 Hz for singleion monitoring (SIM) analyses. Spectra from parasite extracts were compared to
the methyl-ABA standard and previously reported literature values of cis and
trans ABA12.
ABA detection by ELISA. ABA concentrations in parasite extracts were deter-
mined by ELISA using the Phytodetek ABA Test Kit (Agdia) after subtraction of
the background level of ABA in cultured host cells processed in parallel.
Concentrations of ABA in the parasite were expressed as intracellular concentra-
tions based on previous estimates of the parasite volume29.
Cyclic ADPR. T. gondii parasites were purified from culture and treated with
doses of ABA for 10 min. Parasites were snap frozen at –80 uC, extracted in 5%
v/v trichloroacetic acid at 4 uC, and cADPR levels determined using the cyclase
reaction, as described previously8.
In vitro differentiation. Type II strain ME49 parasites were induced to differ-
entiate by treatment with fluridone in normal culture medium (DMEM, 10%
FBS, 10 mM HEPES, gentamicin) during culture for 60 h at 37 uC, 5% CO2.
Alternatively, induction was performed by culture for 72–96 h in RPMI
(pH 8.1) in atmospheric air at 37 uC. Cysts were stained with FITC-labelled
Dolichos biflorus lectin (DBL) (Sigma), DAPI to label DNA, anti-CST1
mAb73.18 (ref. 15), and anti-BAG1 (ref. 16) (provided by L. Weiss) followed
by fluorescently conjugated secondary antibodies. For ultrastructural analysis,
cultures were induced as described above and processed for transmission elec-
tron microscopy, as described previously17. Cysts were classified on the basis of
the presence of a convoluted cyst wall, amylopectin granules, solid-staining
rhoptries, and the posterior position of the nucleus17.
In vivo infections in mice. BALB/c mice were purchased from Charles River
Laboratories, Inc. and infected with tachyzoites of the type II strain PTG, or the
type II PRU strain expressing firefly luciferase (LUC) under the control of the
T. gondii a-tubulin promoter (provided by J. Boothroyd). Parasites were pro-
pagated in vitro in monolayers of HFF cells and tachyzoites were injected by the
i.p. route. An inoculum capable of resulting in death in 90% of untreated animals
(LD90) was used for challenges that consisted of 2 3 103 parasites for PRU-LUC
and 2 3 105 for PTG strain parasites. Animals were treated for 12 d by daily i.p.
injection with 1 mg kg21 or 2 mg kg21 of fluridone (Chem Services) that
was dissolved in DMSO and diluted in PBS. Experiments were conducted with
3–5 animals per group and were repeated 3 or more times. Survival was recorded
for 30 d.
For bioluminescence experiments, animals were imaged at two day intervals,
following mild anaesthesia using isofluorene and i.p. injection of 0.15 mg per kg
body weight of firefly luciferin D (Biosynth AG). Animals were imaged using a
Xenogen IVIS 100 (Caliper LifeSciences). Data are expressed in relative light
units and are shown as mean 6 s.d. Experiments contained 5 animals per group
(except a single animal was used for the non-infected negative control). Animals
were followed for 30 d post-infection, after which chronic infection was con-
firmed by examining homogenates of the brain for tissue cysts, as described
previously17. Briefly, animals were humanely killed, the brains removed and
homogenized in 1 ml of sterile PBS. Homogenates were examined by bright
field or phase contrast microscopy at 403 and cysts were counted from 4 sepa-
rate 10-ml aliquots.
doi:10.1038/nature06478
Nature ©2007 Publishing Group























