Embed Size (px)
Citation preview

J. exp. Biol. 149, 361-378 (1990) 3 6 1Printed in Great Britain © The Company of Biologists Limited 1990
A VIBRATION-SENSITIVE DESCENDING STATOCYSTINTERNEURONE IN THE CRAYEISH PROCAMBARUS CLARKII
BY H. NAKAGAWA AND M. HIS AD A
Zoological Institute, Faculty of Science, Hokkaido University, Sapporo 060,Japan
Accepted 24 October 1989
Summary
1. An interneurone specifically sensitive to substratum vibration was identifiedin the crayfish circumoesophageal connective. The interneurone, called B± in thispaper, received excitatory input from the statocysts on both sides. Electricalstimulation of the statocyst nerve elicited several spikes in the interneurone withlatencies that depended on which side was stimulated.
2. !$! responded phasically to artificial bending of the statocyst sensory hairs.The response was similar to that of the phasic-type receptor in the statocyst.
3. The morphology of B1 was studied by an intracellular staining techniqueusing nickel chloride and subsequent silver intensification. The interneuroneprojects its neurite arborization to the dorsal part of the deutocerebrum andparolfactory lobe on both sides, where the statocyst primary afferents also project.The overlapping of central projections, together with the properties of theresponse of Bl 5 suggests that the interneurone receives excitatory input from thephasic-type receptors and transmits information about phasic body movement,but not static positional information, to the posterior ganglia.
4. Branches of Bi also project to the antennal and tegumentary lobes ipsilateralto the axon. B1 may receive additional mechanosensory information from thecuticular sensory hairs on the antennae and the cephalic body surface.
Introduction
The statocyst sensory nerve of crustaceans includes various functional types ofsensory afferent (Cohen, 1955, 1960; Ozeki et al. 1978; Takahata and Hisada,1979). In the crayfish Procambarus clarkii Girard, the afferents may be classifiedas being of either the tonic type or the phasic type according to the time course oftheir excitatory responses. Although most of them respond only to statocyst hairdeflection towards the centre of the crescent, some of the phasic-type receptorsrespond transiently regardless of the direction of hair deflection (Takahata andHisada, 1979).
In a preceding paper (Nakagawa and Hisada, 1989), we identified seven
words: crayfish, statocyst interneurone, vibration reception.

362 H. NAKAGAWA AND M. H I S AD A
statocyst interneurones which responded tonically to hair deflection towards thecentre of the crescent. We suggested that three of them would correspond to thestatocyst interneurones identified physiologically by tilting experiments (Takahataand Hisada, 1982^). The tonic-type receptors are most likely to be functionallyconnected to these statocyst interneurones and convey positional informationabout body tilting. However, it is still unclear how the information from thephasic-type receptors is processed and represented in the central nervous system.In the crab Scylla serrata a semicircular canal interneurone, fibre 5, has beenshown to carry not only directional low-frequency information but also high-frequency vibrational information (Fraser, 1975).
In this study, we report a descending interneurone which responds transiently tostatocyst hair deflection regardless of the direction of deflection.
Materials and methods
Animals
Adult crayfish, Procambarus clarkii Girard, of either sex, measuring 8-11 cm inlength, were obtained commercially and kept in laboratory tanks before use.
Preparations
Both chelipeds and all walking legs were cut away and stomach contents wereremoved by suction with a pipette. For experiments involving mechanicalstimulation, the animal was placed dorsal-side-up in a chamber filled with crayfishsaline (Van Harreveld, 1936). A small portion of the dorsal carapace was removedto expose the circumoesophageal connectives. To stimulate the statocyst sensoryhairs mechanically, the rostrum and the eyestalk on the side of the body to bestimulated were removed and the hairs covering the opening of the statocyst werethen cut away. The statolith was washed out with a fine saline jet directed into thelumen. Finally, the chitinous dorsal covering over the statocyst was carefullyremoved to expose the statocyst floor and sensory hairs. The antennules wererestrained by a rubber band to prevent them from moving during the experiment.
For experiments using electrical stimulation, the animals were placed ventral-side-up in a chamber filled with crayfish saline. The mouthparts and epistome wereremoved to expose the brain and circumoesophageal connectives. Then theantenna and the proximal joint of the antennule were cut away to expose thestatocyst nerve.
Stimulation
Mechanical vibration of the substratum was achieved by tapping the side of thechamber using an acrylic rod attached to a solenoid driven by a waveformgenerator through a current driver. Statocyst hairs were stimulated using aloudspeaker, as previously described (Nakagawa and Hisada, 1989). Both types ofmechanical stimulation were monitored by recording the current passing througthe solenoid or the loudspeaker. For stimulation of the hairs, the monitor show

Vibration-sensitive statocyst interneurone 363
the time course and polarity of the stimulation, as in the previous paper(Nakagawa and Hisada, 1989). For tapping of the chamber, however, the monitoronly showed that the acrylic rod had struck the side of chamber around the top ofthe 'hill' appearing on the monitoring trace. In later experiments, a stimulusintensity that was critical to elicit Bj spikes only was adopted, unless otherwisenoted (Figs 3A,B, 4D, 5D). To determine the effective stimulus at this intensity,the substratum vibration was calibrated using a piezoelectric transducer (Fig. 1).The stimulus waveform was found to be a quickly damped oscillation of severalsuccessive wave peaks at about 125 FIz. The maximal amplitude of the stimulusvibration was found to be about 1.4 /jm by extrapolation. Thus, except whenspecifically noted, the preparation was stimulated by this vibration regardless ofthe shape of the hill.
To stimulate the statocyst nerve electrically, the nerve was placed on a silverhook electrode enclosed in an oil-filled capillary. A square pulse of 0.1 ms durationwas used for stimulation (Fig. 2A).
Recording and staining
The activity of the circumoesophageal connective was recorded extracellularlyas previously described (Nakagawa and Flisada, 1989). To record the spikedischarge from the interneurone Bi, the circumoesophageal connective was splitinto a smaller bundle containing the axon so that spikes of this neurone could beclearly distinguishable from others.
SM
20 ms
Fig. 1. Actual vibratory movement of the substratum caused by the current method ofstimulation. The upper trace is the recording from the medio-ventral small bundle ofthe circumoesophageal connective (CC). Only Bt spikes (dots) are elicited by tappingat 10 Hz. The middle trace shows the waveform of the substratum vibration recordedby a piezoelectric transducer (SV). The lower trace is the current applied to thesolenoid. Throughout the following figures, this current profile was used as thestimulus monitor (SM).

364 H. NAKAGAWA AND M. HISADA
In the ventral preparation, intracellular recordings were made from the smallbundle of the circumoesophageal connective, using a glass microelectrode placedanterior to the extracellular recording site. The connective was stabilized on aplatform. The microelectrode was filled with 0.25moll"1 NiC^ (d.c. resistance,30-80 MQ in saline) (Fig. 2A). The best recordings were obtained using elec-trodes having a d.c. resistance of 50-55 MQ. Confirmation that the impaledneurone was the descending intemeurone whose response was recorded extracel-lularly was given by the 1:1 correspondence between the extracellular andintracellular spikes (Fig. 2B).
After the response of the intemeurone had been examined, nickel ions wereinjected into the intemeurone by applying 10 nA depolarizing current pulses of0.5 s in duration at 1 Hz for 1 h. After incubation at room temperature, the nickelwas precipitated by rubeanic acid. Silver intensification was performed routinelyon whole-mount specimens (Bacon and Altman, 1977). The intensified brain wasdehydrated in alcohol and cleared in methyl salicylate. Drawings of the stainedinterneurones were made with a camera lucida. The description of the morphologywas based on four successful stains of the intemeurone.
Transverse paraffin sections, 10fjm thick, were made of the brain and theconnective containing the stained neurone. Drawings were made with a cameralucida to reconstruct the entire structure of the intemeurone and to examine itscentral projection. The nomenclature of the cell body clusters within the brain wastaken from the scheme devised by Tautz and Tautz (1983).
I 1Circumoesophageal
connective
Fig. 2. (A) Experimental arrangement for extracellular and intracellular recording ofB! spikes. The electrode for stimulation was placed on the statocyst nerve. Intracellu-lar recordings were made from the circumoesophageal connective anterior to theextracellular recording site. (B) The response of interneurone Bi to electricalstimulation of the statocyst nerve on the side contralateral to the axon. In thispreparation, B] generated two spikes (arrows) in response to the stimulation. Note the1:1 correspondence of the intracellular spikes with the extracellular ones.

Vibration-sensitive statocyst interneurone 365
A
<X~-—-»
SM-
B
CC
SM--
20 ms
CCU—SM--
Fig. 3. The responses of several descending interneurones to a soft tapping of thechamber at 10 Hz with decreasing amplitude (A-C) . The upper record in each panel isthe extracellular recording from the medio-ventral small bundle of the circumoeso-phageal connective (CC). The lower trace monitors the applied current (SM). In A andB, several descending interneurones respond to the stimulation, whereas in C only onedescending interneurone, identified as interneurone Bj , generates five spikes inresponse to the stimulation. The dots mark the Bi spikes.
Results
A vibration-sensitive descending statocyst interneurone
Descending interneurones which responded to tapping of the experimentalchamber were identified in the drcumoesophageal connectives. An interneuronewhich was specifically sensitive to the mode of vibratory stimulation was found toreceive excitatory input from statocysts on both sides (see below). We designatedit as B1 ; based on its bilateral input.
Several units including Bi responded to hard tapping. As the intensity of thetapping was decreased gradually, the number of responding units decreased untilonly interneurone Bi responded to the stimulus (Fig. 3A-C). We adopted this
^ intensity for the following experiments. Removal of the statolith on the

366 H. NAKAGAWA AND M. HISADA
side ipsilateral to the interneurone axon had little effect on its response to tapping.However, when the contralateral one was subsequently washed away, theinterneurone response disappeared completely (Fig. 4A-C). No response of theinterneurone was observed after this bilateral statolith removal, even with adrastic increase in stimulus intensity (Fig. 4D). Similar results were obtained whenstatolith removal was performed in the reverse order, i.e. removal of the first(contralateral) statolith had little effect, whereas removal of the second (ipsilat-eral) statolith abolished the response (Fig. 5A-D). This observation indicatedthat the interneurone received vibratory input exclusively, but bilaterally, fromthe statocysts.
Difference in response between interneurones Cj and Bj
Of the crayfish statocyst interneurones reported previously, interneurone Ci hasbeen examined most thoroughly because of its large axon diameter (Takahata and
A Intact C Bilateral statolith removal
20 msB Ipsilateral statolith removal D Stimulus intensity increase
1 M̂̂—\f— - ^ / ^
SM-
Fig. 4. The effect of statolith removal on the response of interneurone B1 to a softtapping at 10 Hz. In each panel the upper four traces are extracellular recordings fromthe medio-ventral small bundle of the circumoesophageal connective (CC). Theresponses to four successive cycles of stimulation are shown. The lowest trace monitorsthe applied current (SM). In the top trace, Bi spikes are indicated by dots. (A) Bothstatocysts intact. (B) The statolith on the side ipsilateral to the axon of theinterneurone is removed. Note the slightly reduced response of interneurone Bi.(C) The contralateral statolith is also removed subsequently. The Bi responsedisappears completely. (D) The amplitude of stimulation is increased after removal ofboth statoliths. Bj never responds to the stimulation. All recordings are from the sameanimal.

Vibration-sensitive statocyst interneurone
A Intact C Bilateral statolith removal
CC v ^ « H V V + ^ CC v-
•V"
SM-
367
SM-
. * » * • •
B Contralateral statolith removal20 ms
D Stimulus intensity increase
C C > " • - - « • - ^ . • ^ • o . . ^
SM-
Fig. 5. The effect of statolith removal in the reverse order to that in Fig. 4 on theresponse of interneurone BL. The experimental conditions and data presentation arethe same as those in Fig. 4. (A) Both statocysts intact. (B) The contralateral statolith isremoved. Note the slight reduction in the Bi response. (C) The ipsilateral statolith isalso removed subsequently. The Bj response almost disappears. (D) The amplitude ofstimulation is increased after removal of both statoliths. Bt hardly responds to thestimulation.
Hisada, 1982#; Nakagawa and Hisada, 1989), and it can be considered asrepresentative of the position-sensitive interneurones. The responses of inter-neurone Bi were compared with those of Q . Their responses to artificial bendingof the statocyst hairs at various frequencies were examined first. For interneuroneCi, the stimulation was applied only to the contralateral statocyst, since C\ isknown to receive dominant input from the contralateral statocyst (Takahata andHisada, 1982a). For interneurone By, which receives bilateral input from thestatocysts, stimulation was applied to each statocyst alternately.
Interneurone Q showed a directional tonic response to stimulation at 0.1 Hz.However, this directionality became ambiguous at 1 Hz, and there was noresponse to stimulation at 10 Hz (Fig. 6A). Further experiments showed thatdirectional sensitivity was preserved well at 0.5 Hz, but disappeared at around2Hz(Fig. 6B,C).
Interneurone Bi did not show any directional tonic response to stimulation ateither 0.1 or 1 Hz. When a response was observed it was transient and occurred atthe onset or the end of mechanical stimulation. Furthermore, unlike interneuroneCi, interneurone Bt responded reliably to stimulation at 10Hz (Fig. 7). It showeda reliable response with a fixed latency to stimulation at 5 Hz, and a labile response
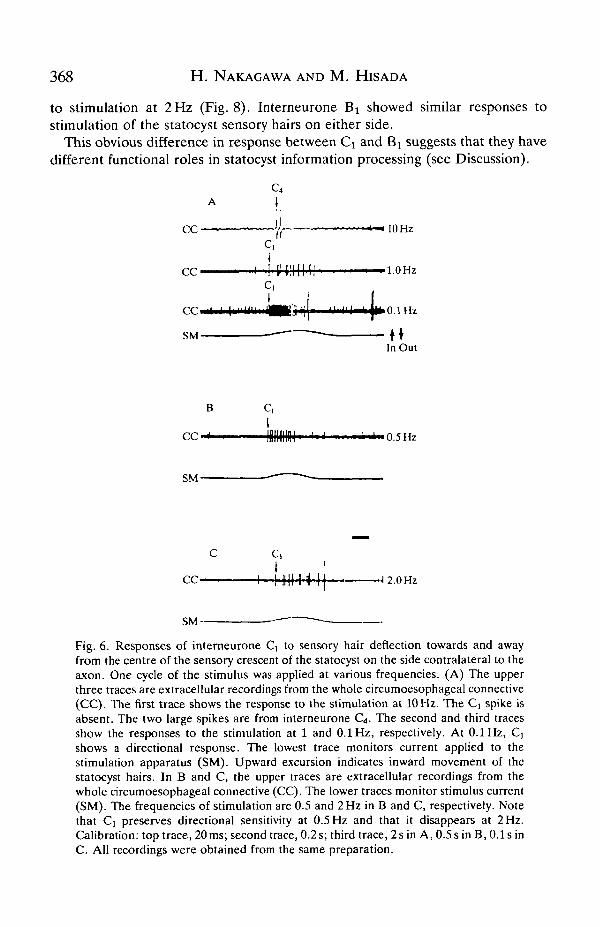
368 H. NAKAGAWA AND M. HISADA
to stimulation at 2 Hz (Fig. 8). Interneurone Bi showed similar responses tostimulation of the statocyst sensory hairs on either side.
This obvious difference in response between Ci and Bj suggests that they havedifferent functional roles in statocyst information processing (see Discussion).
c4
cc-
CO
C O
SM-
C,I
c,I
• l.OHz
.0 .1 Hz
-HIn Out
»•«*•• 0.5 Hz
CC- -•12.0 Hz
SM--
Fig. 6. Responses of interneurone Cj to sensory hair deflection towards and awayfrom the centre of the sensory crescent of the statocyst on the side contralateral to theaxon. One cycle of the stimulus was applied at various frequencies. (A) The upperthree traces are extracellular recordings from the whole circumoesophageal connective(CC). The first trace shows the response to the stimulation at 10 Hz. The Q spike isabsent. The two large spikes are from interneurone C4. The second and third tracesshow the responses to the stimulation at 1 and 0.1 Hz, respectively. At 0.1 Hz, Qshows a directional response. The lowest trace monitors current applied to thestimulation apparatus (SM). Upward excursion indicates inward movement of thestatocyst hairs. In B and C, the upper traces are extracellular recordings from thewhole circumoesophageal connective (CC). The lower traces monitor stimulus current(SM). The frequencies of stimulation are 0.5 and 2 Hz in B and C, respectively. Notethat Ci preserves directional sensitivity at 0.5 Hz and that it disappears at 2 Hz.Calibration: top trace, 20 ms; second trace, 0.2 s; third trace, 2 sin A, 0.5 s in B, 0.1 sinC. All recordings were obtained from the same preparation.
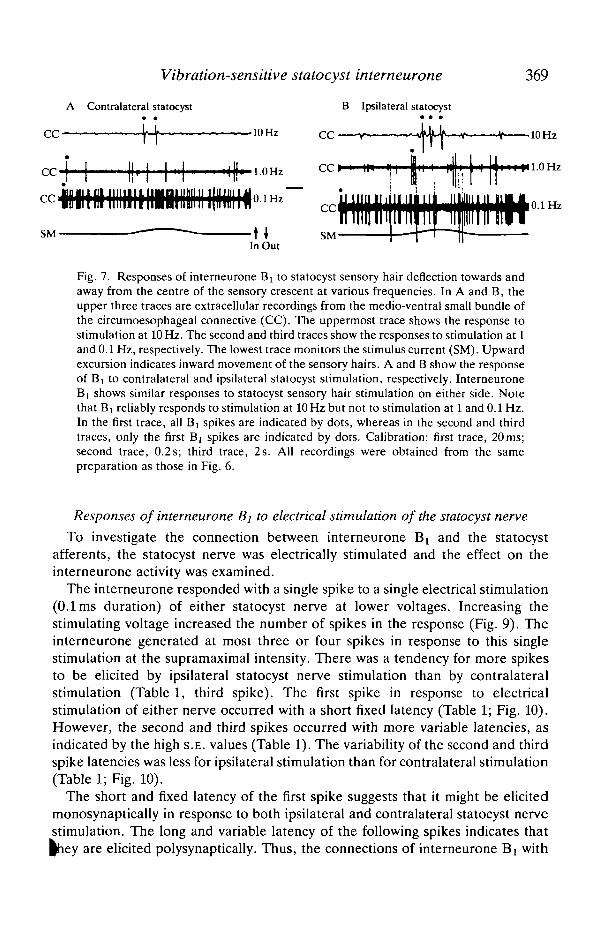
Vibration-sensitive statocyst interneurone 369
A Contralateral statocyst B Ipsilateral statocyst
CC ^ lOHz C C
4. •1.0 HzCC ,
' ijiiniiiiiiHiiiiauHinii'SM • 1 I
In Out
Fig. 7. Responses of interneurone Bi to statocyst sensory hair deflection towards andaway from the centre of the sensory crescent at various frequencies. In A and B, theupper three traces are extracellular recordings from the medio-ventral small bundle ofthe circumoesophageal connective (CC). The uppermost trace shows the response tostimulation at 10 Hz. The second and third traces show the responses to stimulation at 1and 0.1 Hz, respectively. The lowest trace monitors the stimulus current (SM). Upwardexcursion indicates inward movement of the sensory hairs. A and B show the responseof B] to contralateral and ipsilateral statocyst stimulation, respectively. InterneuroneB] shows similar responses to statocyst sensory hair stimulation on either side. Notethat B! reliably responds to stimulation at 10 Hz but not to stimulation at 1 and 0.1 Hz.In the first trace, all Bi spikes are indicated by dots, whereas in the second and thirdtraces, only the first Bi spikes are indicated by dots. Calibration: first trace, 20 ms;second trace, 0.2 s; third trace, 2 s. All recordings were obtained from the samepreparation as those in Fig. 6.
Responses of interneurone Bi to electrical stimulation of the statocyst nerve
To investigate the connection between interneurone Bt and the statocystafferents, the statocyst nerve was electrically stimulated and the effect on theinterneurone activity was examined.
The interneurone responded with a single spike to a single electrical stimulation(0.1ms duration) of either statocyst nerve at lower voltages. Increasing thestimulating voltage increased the number of spikes in the response (Fig. 9). Theinterneurone generated at most three or four spikes in response to this singlestimulation at the supramaximal intensity. There was a tendency for more spikesto be elicited by ipsilateral statocyst nerve stimulation than by contralateralstimulation (Table 1, third spike). The first spike in response to electricalstimulation of either nerve occurred with a short fixed latency (Table 1; Fig. 10).However, the second and third spikes occurred with more variable latencies, asindicated by the high S.E. values (Table 1). The variability of the second and thirdspike latencies was less for ipsilateral stimulation than for contralateral stimulation(Table 1; Fig. 10).
The short and fixed latency of the first spike suggests that it might be elicitedmonosynaptically in response to both ipsilateral and contralateral statocyst nervestimulation. The long and variable latency of the following spikes indicates thatJhey are elicited polysynaptically. Thus, the connections of interneurone Bi with
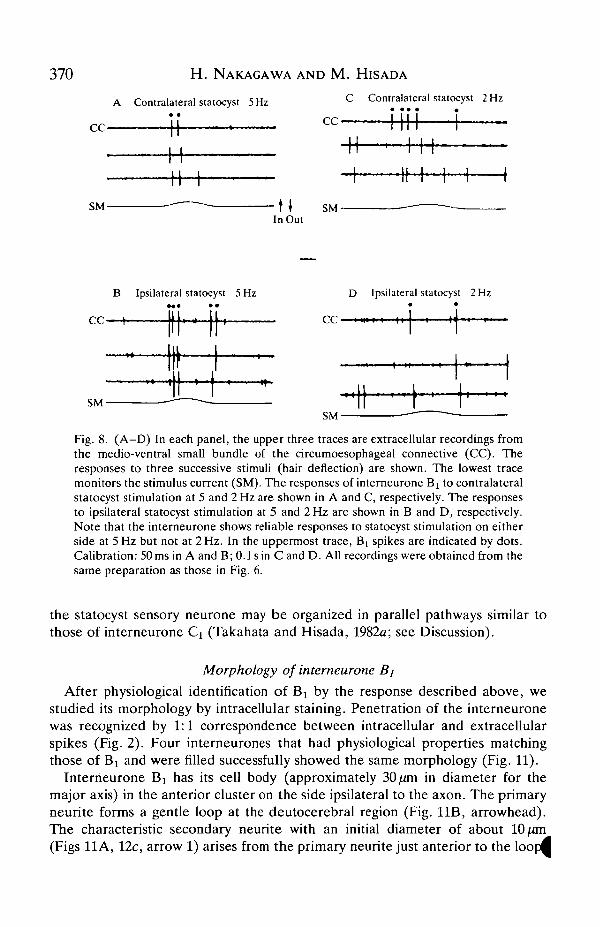
370 H. NAKAGAWA AND M. HISADA
A Contralateral statocyst 5 Hz C Contralateral statocyst 2 Hz
CC M > * •• •cc IJ
SM ~ H SM-InOut
B Ipsilateral statocyst 5 Hz D Ipsilateral statocyst 2 Hz••• • •
CC-H M..« U, CC
SM-
Fig. 8. (A-D) In each panel, the upper three traces are extracellular recordings fromthe medio-ventral small bundle of the circumoesophageal connective (CC). Theresponses to three successive stimuli (hair deflection) are shown. The lowest tracemonitors the stimulus current (SM). The responses of interneurone Bx to contralateralstatocyst stimulation at 5 and 2 Hz are shown in A and C, respectively. The responsesto ipsilateral statocyst stimulation at 5 and 2 Hz are shown in B and D, respectively.Note that the interneurone shows reliable responses to statocyst stimulation on eitherside at 5Hz but not at 2Hz. In the uppermost trace, B[ spikes are indicated by dots.Calibration: 50 ms in A and B; 0.1 sin Cand D. All recordings were obtained from thesame preparation as those in Fig. 6.
the statocyst sensory neurone may be organized in parallel pathways similar tothose of interneurone Q (Takahata and Hisada, 1982a; see Discussion).
Morphology of interneurone Bj
After physiological identification of B! by the response described above, westudied its morphology by intracellular staining. Penetration of the interneuronewas recognized by 1:1 correspondence between intracellular and extracellularspikes (Fig. 2). Four interneurones that had physiological properties matchingthose of Bi and were filled successfully showed the same morphology (Fig. 11).
Interneurone Bi has its cell body (approximately 30 /xm in diameter for themajor axis) in the anterior cluster on the side ipsilateral to the axon. The primaryneurite forms a gentle loop at the deutocerebral region (Fig. 11B, arrowhead).The characteristic secondary neurite with an initial diameter of about 10/an(Figs 11 A, 12c, arrow 1) arises from the primary neurite just anterior to the loopB

Vibration-sensitive statocyst interneurone 371
A 3V •
B 4V • •
10 ms
C 6V •
Fig. 9. Resjx>nses of intemeurone Bi to electrical stimulation of the ipsilateralstatocyst nerve. The stimulus intensity was increased gradually (A, 3 V; B, 4 V; C,6 V). Each panel shows the responses to three or four successive trials. Extracellularspikes of interneurone Bi (dots) were identified by a soft tapping beforehand.Increasing stimulus intensity both caused the short-latency response and subsequentlyrecruited the long-latency responses.
extends medio-ventrally across the midline, and terminates to branch in theparolfactory lobe on the side contralateral to the axon (Fig. 126,c).
The secondary neurite gives rise to 9-10 orthogonal tertiary neurites (approxi-mately 3^m in diameter) which occur at equal distances (50-80 ^un) along thesecondary neurite. These tertiary neurites, except the longest one (Figs 11B, 12a,arrow 2), project into the parolfactory lobe on both sides (Fig. 11A). The longestone proceeds further towards the ventral side. The branching pattern of Bj in thedeutocerebrum thus shows an orthogonal arrangement of secondary and tertiary
pheurites. This branching pattern resembles those of interneurones in the antennal
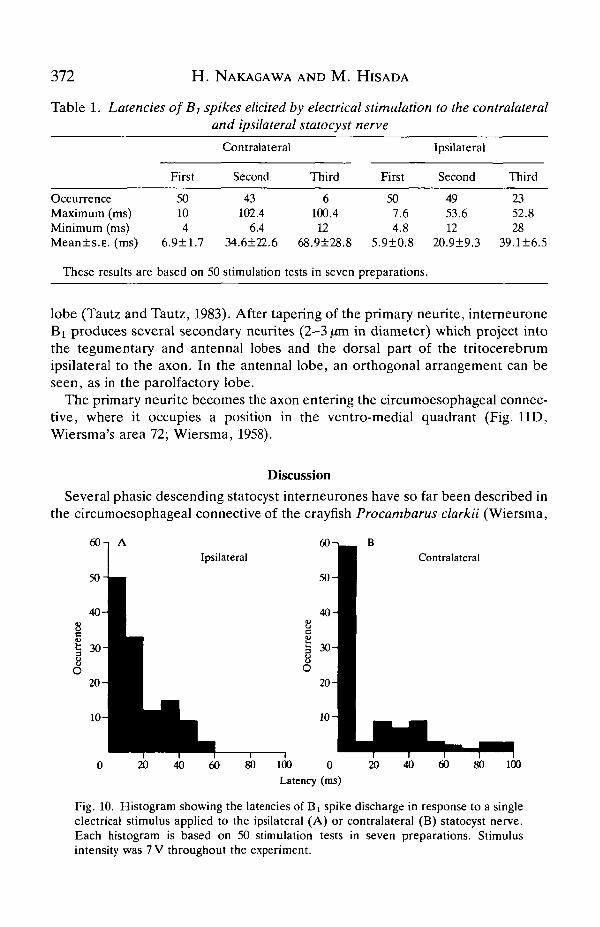
372 H. NAKAGAWA AND M. HIS AD A
Table 1. Latencies of Bj spikes elicited by electrical stimulation to the contralateraland ipsilateral statocyst nerve
OccurrenceMaximum (ms)Minimum (ms)Mean±s.E. (ms)
These results are
First
50104
6.9±1.7
based on
Contralateral
Second
43102.4
6.434.6±22.6 68
50 stimulation tests
Third
6100.4
12.9±28.8
in seven
First
507.64.8
5.9±0.8
preparations.
Ipsilateral
Second
4953.612
20.9±9.3
Third
2352.828
39.1±6.5
lobe (Tautz and Tautz, 1983). After tapering of the primary neurite, interneuroneBi produces several secondary neurites (2-3 jan in diameter) which project intothe tegumentary and antennal lobes and the dorsal part of the tritocerebrumipsilateral to the axon. In the antennal lobe, an orthogonal arrangement can beseen, as in the parolfactory lobe.
The primary neurite becomes the axon entering the circumoesophageal connec-tive, where it occupies a position in the ventro-medial quadrant (Fig. 11D,Wiersma's area 72; Wiersma, 1958).
DiscussionSeveral phasic descending statocyst interneurones have so far been described in
the circumoesophageal connective of the crayfish Procambarus clarkii (Wiersma,
60 n AIpsilateral
0 20 40 60 80 100
20
10
0Latency (ms)
Contralateral
40 60 80 100
Fig. 10. Histogram showing the latencies of Bi spike discharge in response to a singleelectrical stimulus applied to the ipsilateral (A) or contralateral (B) statocyst nerve.Each histogram is based on 50 stimulation tests in seven preparations. Stimulusintensity was 7 V throughout the experiment.

Vibration-sensitive statocyst interneurone 373
1958; Wiersma and Mill, 1965). However, the stimulation technique employed wasnot sensitive enough to determine specifically which receptors in the antennulewere involved. In this study, we could identify the descending interneurone whichresponded phasically to statocyst hair stimulation. In the following sections we willdiscuss the functional and morphological differences between the phasic- andtonic-type descending statocyst interneurones by comparing interneurone Bi withthe previously identified statocyst interneurone Ci (Takahata and Hisada,
; Nakagawa and Hisada, 1989).
Range fractionation in the central representation of statocyst information
Interneurones Ci and Bi both respond to mechanical stimulation of the statocyst
Anterior
Ventral
Pa
Fig. 11. Morphology of interneurone Bi. (A) Dorsal view of the NiCl2-injectedinterneurone. Arrow 1 indicates the characteristic thick secondary neurite (see text).The dotted line shows the midline of the ganglion. (B) Lateral view of the sameinterneurone as in A. The arrowhead indicates a gentle loop, and arrow 2 indicates thelongest tertiary neurite (see text), a-d indicate the positions of the sections recon-structed in Fig. 12. (C) Dorsal view of the brain at a lower magnification showing theposition of the neuropile in the ganglion and the projecting area of Bj (stippled area).Op, optic lobe; Pa, parolfactory lobe; Ol, olfactory lobe; Ac, accessory lobe; An,antennal lobe. (D) Cross-section of the right circumoesophageal connective. Thedarkened rounded profile is the axon of the stained interneurone Bi (arrow), mg,medial giant fibre; Ig, lateral giant fibre. In A-C, anterior is at the top. In B, dorsal is tothe left. Calibration: 100/an in A and B, 50[an in D. B and D were obtained from thesame preparation.

374 H. NAKAGAWA AND M. H I S AD A
sensory hairs. However, their response frequency and input statocyst are quitedifferent, indicating that they perform different functions.
The statocyst sensory nerves of crayfish and lobster include various types ofsensory afferents. The first electrophysiological analysis of a single unit of thistype, in Homarus americanus, revealed a variety of unit responses (Cohen, 1955).The position receptor gives a nonadapting and tonic response with a frequencydepending on the absolute position of the sensory hair to which the unit isattached. The vibration receptor gives responses only to large acceleration orwhen the substratum is tapped. Recently, in the crayfish Orconectes limosus,vibration sensitivity of the statocyst has been investigated more quantitatively(Breithaupt and Tautz, 1988).
In the crayfish Procambarus clarkii, detailed study of the statocyst sensoryneurones has revealed that they can be classified into two types of tonic and phasiccells according to their discharge patterns in response to sensory hair deflection(Takahata and Hisada, 1979). The tonic-type receptor showed an initial transient
2 VPLC
DACDorsal
VPPC
DMC
Fig. 12. Drawings of transverse sections through interneurone Bi. Each sectionconsists of three superimposed serial sections at intervals of 10-20 fim. The sections areviewed from the anterior end of the brain. The labelling (a-d) of sections correspondsto that indicated in Fig. 11B. In a the longest tertiary neurite (arrow 2) does notterminate in the parolfactory lobe. In b the interneurone projects into the parolfactorylobe on both sides. In c the characteristic secondary neurite (arrow 1) extends acrossthe midline. In d the interneurone projects into the antennal lobe ipsilateral to theaxon. VPMC, ventral paired medial cluster; VUMC, ventral unpaired medial cluster;VPLC, ventral paired lateral cluster; DAC, dorsal anterior cluster; DMC, dorsalmedial cluster; VUPC, ventral unpaired posterior cluster; VPPC, ventral pairedposterior cluster; DPC, dorsal posterior cluster. Abbreviations for sensory neuropilesare the same as in Fig. 11C. Ventral is at the top.

Vibration-sensitive statocyst interneurone 375
response when a specific hair was deflected, and the response was followed by asustained spike discharge while the hair remained deflected. The phasic-typereceptor showed only a transient excitatory response when a specific hair wasdeflected. It was also shown that most statocyst receptors could follow stimulusfrequencies up to a maximum of about 5 Hz, but some could follow frequencies ashigh as 10 Hz.
Thus, the response properties of tonic- and phasic-type receptors are similar tothose of intemeurones Q and Bi, respectively. This suggests that the tonic-typereceptors connect with the tonic-type descending statocyst intemeurones such asCi, while the phasic-type receptors connect with the phasic-type intemeuronessuch as Bi.
Fraser (1975) showed in the crab that a semicircular canal interneurone, fibre 5,exhibited two peaks of sensitivity to oscillations of the antennules. He concludedthat fibre 5 carried two sorts of information from the statocysts; low-frequencydirectional information and high-frequency vibrational information. In Procam-barus clarkii, by contrast, the positional information and vibrational informationare carried via two different channels of tonic-type and phasic-type statocystintemeurones, respectively.
Central connection between statocyst receptors and descending statocystintemeurones
In a previous study, it was shown that electrical stimulation of the contralateralstatocyst caused both short- and long-latency responses in interneurone Ci. As aresult, it has been suggested that connection of interneurone Ci with the inputstatocyst sensory neurone is organized in a parallel way, one connection beingmonosynaptic and the other polysynaptic (Takahata and Hisada, 1982a). In thepresent study we have shown that electrical stimulation of the statocyst nerve oneither side evoked several spikes with different latencies in interneurone B1
(Table 1). This result suggests that the input statocyst sensory neurone andinterneurone Bx are connected via parallel pathways.
Although stimulation of the statocyst sensory hairs on both sides could activateintemeurone Bi, more spikes were elicited in response to ipsilateral stimulationthan to contralateral stimulation, and the variability of the second and third spikelatencies was less for ipsilateral stimulation (Table 1). The functional significanceof this differential contribution from the two sides to activation of the inter-neurone is still to be investigated.
Relationship of morphology and function in two types of descending statocystintemeurones
Intemeurone Bi projects to the dorsal part of the deutocerebrum and theparolfactory lobe on both sides. The statocyst primary afferents also project tothese neuropile regions (Yoshino et al. 1983). The overlapping is consistent withthe current report that interneurone Bi receives short-latency, probably mono-
synaptic, input from bilateral statocysts (Figs 9, 10). Interneurone B t also extends

376 H. NAKAGAWA AND M. HISADA
its major branches to the antennal and tegumentary lobes on the ipsilateral side tothe axon. It is known that the primary afferents connected to the antennal sensoryhairs terminate in the antennal lobe (Taylor, 1975; Tautz and Tautz, 1983). It isalso known that the tegumentary lobe that is part of a continuum with the antennallobe is composed of axon terminals of mechanosensory afferents and inter-neurones whose receptive fields cover most of the animal's surface (Kinnamon,1979). This suggests that interneurone Bi also receives mechanosensory infor-mation from sensory hairs other than those of the statocysts. The threshold of non-statocyst receptors for activating B 1 ; however, was much higher than that ofstatocyst receptors (Figs 4, 5).
The projection pattern of interneurone Bi differs from that of the tonic-typestatocyst interneurone (Nakagawa and Hisada, 1989) in two ways.
First, all seven tonic-type interneurones extend their major branches to the opticlobe where the fibres from the eyestalk ganglia terminate (Bethe, 1897; Hanstrom,1925, 1948), while Bx lacks any branches in this neuropile. This difference in theirmorphology is consistent with physiological observations. It is well known that theequilibrium responses are controlled by complex interactions between thestatocyst and visual input (Alverdes, 1926; Hisada, 1975; Takahata and Hisada,1982a; Schone et al. 1983; Neil, 1985). Moreover, it has also been reported thatinterneurone Q receives visual input (Takahata and Hisada, 1982a). Tonic-typeinterneurones responsible for controlling equilibrium responses might well projectto the optic lobe. In contrast, the lack of B1 branches in the optic lobe is alsoconsistent with our conclusion that the phasic-type interneurone Bi is responsiblefor vibration reception rather than equilibrium responses.
Second, all seven tonic-type interneurones extend their major branches to thedeutocerebrum either ipsilateral or contralateral to their axons, while B^ branchesextend extensively and uniformly to the bilateral deutocerebrum. This appears tobe consistent with the physiological finding that tonic-type interneurones receivetheir dominant input from one of the statocysts, whereas Bi receives input fromboth statocysts. However, we should note here that (1) the statocyst sensoryafferents project to the deutocerebrum on both sides (Yoshino et al. 1983) and (2)the descending statocyst interneurones receive long-latency polysynaptic inputfrom the input statocyst (Takahata and Hisada, 1982a; Figs 9, 10). These findingsimply that the dendritic projection of statocyst interneurones in the deutoce-rebrum does not necessarily conform to the laterality of the input statocyst. In aprevious report, indeed, we showed that all branches of interneurone S3 wererestricted to the side ipsilateral to the axon, although this neurone received inputfrom the contralateral statocyst (Nakagawa and Hisada, 1989). Thus, it isimpossible to predict the input statocyst only from the projection patterns of aninterneurone in the deutocerebrum.
Function of interneurone Bj
Interneurone Cx and other tonic-type descending statocyst interneurones whichshow tonic and directional responses appear to be the principal channel fo^

Vibration-sensitive statocyst intemeurone 377
controlling equilibrium responses (Takahata and Hisada, 1982a; Nakagawa andHisada, 1989). Intemeurone Bi, however, does not seem to be responsible fordirectly controlling the equilibrium responses since the intemeurone shows onlyphasic responses and has no directional sensitivity. We conclude that intemeuroneBi transmits information about contact or boundary vibration (Markl, 1983) ratherthan positional information. This conclusion raises the question of the role ofvibrational information in controlling crayfish behaviour. Vibrational stimuligenerally evoke flight reactions or reflex-hke jumps directed away from the sourceof vibration. However, it is unlikely that the descending signals in Bi elicit theseescape responses because the threshold for firing of the intemeurone is very low.
One possible role of intemeurone Bx could be to provide a general excitatorybias to the motor control system for behaviour which is affected by vibrationalinformation - such as the escape response or target orientation (Tautz et al. 1981;Masters et al. 1982) - so that the following tonic signals could be readily integratedinto the motor control signals. Alternatively, it is well known that contact orboundary vibrations are an important means of communication of sexual oraggressive interactions throughout the animal kingdom (Markl, 1983). Therefore,intemeurone Bj might participate in vibrational communication, although weknow of no behavioural evidence for it in Procambarus clarkii. The role of Bx
should be subjected to further investigation, especially at the behavioural level.
We thank Dr M. Takahata for his helpful comments on the manuscript. We arealso grateful to Dr R. M. Glantz for his invaluable advice of the ventral-side-upbrain preparation techniques. This work was supported by Grants-in-aid (nos61480316 and 61840023) from the Japanese Ministry of Education, Science andCulture to MH.
ReferencesALVERDES, F. (1926). Stato, Photo- und Tangoreactionen bei zwei Garneelenarten. Z. vergl.
Physiol. 4, 699-765.BACON, J. P. AND ALTMAN, J. S. (1977). A silver intensification method for cobalt-filled neurons
in wholemount preparations. Brain Res. 138, 359-363.BETHE, A. (1897). Das Nervensystem von Carcinus maenas, ein anatomischphysiologischer
Versuch. I. Theil. II. Mittheil. Arch, mikrosk. Anat. 50, 589-639.BREUHAUPT, TH. AND TAUTZ, J. (1988). Vibration sensitivity of the crayfish statocyst.
Naturwissenschaften 75, 310-312.COHEN, M. J. (1955). The function of receptors in the statocyst of the lobster Homarus
americanus. J. Physiol., Lond. 130, 9-34.COHEN, M. J. (1960). The response patterns of single receptors in the crustacean statocyst. Proc.
R. Soc. B 152, 30-48.FRASER, P. J. (1975). Free hook hair and thread hair input to fiber 5 in the mud crab, Scylla
serrata, during antennule rotation. J. comp. Physiol. 103, 291-313.HANSTROM, B. (1925). The olfactory centers in crustaceans. J. comp. Neurol. 38, 221-250.HANSTROM, B. (1948). The brain, the sense organs, and the incretory organs of the head in the
Crustacea Malacostraca. Bull. biol. France et Belg. Suppl. 33, 98-126.HISADA, M. (1975). Gravitational and visual control of eye movement in crayfish. Fortschr.
Zool. 23,162-173.

378 H. NAKAGAWA AND M. HIS AD A
KINNAMON, J. C. (1979). Tactile input to the crayfish tegumentary neuropile. Comp. Biochem.Physiol. 63A, 41-50.
MARKL, H. (1983). Vibration communication. In Neuroethology and Behavioral Physiology (ed.F. Huber and H. Markl), pp. 332-353. Berlin, Heidelberg: Springer-Verlag.
MASTERS, W. M., AJCHER, B., TAUTZ, J. AND MARKL, H. (1982). A new type of water vibrationreceptor on the crayfish antenna. II. Model of receptor function. J. comp. Physiol. 149,409-422.
NAKAGAWA, H. AND HISADA, M. (1989). Morphology of descending statocyst interneurons in thecrayfish Procambarus clarkii Girard. Cell Tissue Res. 255, 539-551.
NEIL, D. M. (1985). Multisensory interactions in the crustacean's equilibrium system. InFeedback and Motor Control in Invertebrates and Vertebrates (ed. W. J. P. Barnes and M.Gladden), pp. 277-298. London: Croom Helm.
OZEKI, M., TAKAHATA, M. AND HISADA, M. (1978). Afferent response patterns of the crayfishstatocyst with ferrite grain statolith to magnetic field stimulation. J. comp. Physiol. 123,1-10.
SCHONE, H., NEIL, D. M., SCAPINI, F. AND DREISSMANN, G. (1983). Interaction of substrate,gravity and visual cues in the control of compensatory eye responses in the spiny lobster,Palinurus vulgaris. J. comp. Physiol. 150, 23-30.
TAKAHATA, M. AND HJSADA, M. (1979). Functional polarization of statocyst receptors in thecrayfish Procambarus clarkii Girard. J. comp. Physiol. 130, 201-207.
TAKAHATA, M. AND HISADA, M. (1982a). Statocyst interneurons in the crayfish Procambarusclarkii Girard. I. Identification and response characteristics. J. comp. Physiol. 149, 287-300.
TAKAHATA, M. AND HISADA, M. (1982ft). Statocyst interneurons in the crayfish Procambarusclarkii Girard. II. Directional sensitivity and its mechanism. J. comp. Physiol. 149, 301-306.
TAUTZ, J., MASTERS, W. M., AICHER, B. AND MARKL, H. (1981). A new type of water vibrationreceptor on the crayfish antenna. I. Sensory physiology. J. comp. Physiol. 144, 533-541.
TAUTZ, J. AND TAUTZ, R. M. (1983). Antennal neuropile in the brain of the crayfish:Morphology of neurons. /. comp. Neurol. 218, 415—425.
TAYLOR, R. C. (1975). Integration in the crayfish antennal neuropile: Topographicrepresentation and multiple channel coding of mechanoreceptive submodalities. J. Neurobiol.6, 475-499.
VAN HARREVELD, A. (1936). A physiological solution for freshwater crustaceans. Proc. Soc. exp.Med. 34, 428-432.
WIERSMA, C. A. G. (1958). On the functional connections of single units in the central nervoussystem of the crayfish, Procambarus clarkii (Girard). /. comp. Neurol. 110, 421-471.
WIERSMA, C. A. G. AND MILL, P. J. (1965). "Descending" neuronal units in the commissure ofthe crayfish central nervous system; and their integration of visual, tactile and proprioceptivestimuli. /. comp. Neurol. 125, 67-94.
YOSHINO, M., KONDOH, Y. AND HISADA, M. (1983). Projection of statocyst sensory neuronsassociated with crescent hairs in the crayfish Procambarus clarkii Girard. Cell Tissue Res. 230,37-48.





























