Embed Size (px)
Citation preview

A U2-snRNP–independent role of SF3b in promotingmRNA exportKe Wanga,b, Changping Yina,b, Xian Duc, Suli Chena,b, Jianshu Wanga,b, Li Zhanga,b, Lantian Wanga,b, Yong Yud,Binkai Chia,b, Min Shia,b, Changshou Wanga,b, Robin Reedd, Yu Zhouc, Jing Huange, and Hong Chenga,b,1
aState Key Laboratory of Molecular Biology, CAS Center for Excellence in Molecular Cell Science, Shanghai Institute of Biochemistry and Cell Biology,Chinese Academy of Sciences, University of Chinese Academy of Sciences, 200031 Shanghai, China; bShanghai Key Laboratory of Molecular Andrology, CASCenter for Excellence in Molecular Cell Science, Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences, University of ChineseAcademy of Sciences, 200031 Shanghai, China; cHubei Key Laboratory of Cell Homeostasis, College of Life Sciences, Wuhan University, 430072 Wuhan,China; dDepartment of Cell Biology, Harvard Medical School, Boston, MA 02115; and eShanghai Institute of Precision Medicine, Ninth People’s Hospital,Shanghai Jiao Tong University School of Medicine, 200025 Shanghai, China
Edited by Alberto R. Kornblihtt, University of Buenos Aires, Buenos Aires, Argentina, and approved February 28, 2019 (received for review November11, 2018)
To ensure efficient and accurate gene expression, pre-mRNA pro-cessing and mRNA export need to be balanced. However, how thisbalance is ensured remains largely unclear. Here, we found thatSF3b, a component of U2 snRNP that participates in splicing and 3′processing of pre-mRNAs, interacts with the key mRNA export adap-tor THO in vivo and in vitro. Depletion of SF3b reduces THO bindingwith the mRNA and causes nuclear mRNA retention. Consistently,introducing SF3b binding sites into the mRNA enhances THO recruit-ment and nuclear export in a dose-dependent manner. These datademonstrate a role of SF3b in promoting mRNA export. In support ofthis role, SF3b binds with mature mRNAs in the cells. Intriguingly,disruption of U2 snRNP by using a U2 antisense morpholino oligonu-cleotide does not inhibit, but promotes, the role of SF3b in mRNAexport as a result of enhanced SF3b–THO interaction and THO re-cruitment to the mRNA. Together, our study uncovers a U2-snRNP–independent role of SF3b in mRNA export and suggests that SF3bcontributes to balancing pre-mRNA processing and mRNA export.
SF3b | THO | mRNA export | pre-mRNA processing | U2 snRNP
In eukaryotes, the nascent pre-mRNA transcripts undergo multi-ple processing steps in the nucleus before mRNAs are exported
to the cytoplasm for translation. Accumulating evidence suggeststhat pre-mRNA processing and mRNA export need to be balancedto ensure efficient and accurate gene expression (1–7). Whensplicing factors are limited or when mRNA export factors arepresent in excess, even unspliced pre-mRNAs are leaked to thecytoplasm (1–3). On the contrary, down-regulation of nuclearexport factors results in nuclear retention of fully processedmRNAs that are ultimately subject to degradation (4–7). Thus,maintenance of the balance between pre-mRNA processing andmRNA export is of significant importance. However, how thisbalance is achieved remains largely unknown.U2 snRNP is a core component of the spliceosome. It is comprised
of U2 snRNA, multisubunit SF3a and SF3b complexes, U2-snRNP–specific proteins A′ and B″, as well as the seven Sm proteinscommon to the spliceosomal snRNPs (8–10). During splicing, SF3bproteins contact the pre-mRNA at and near the branch site (BS) ina sequence-independent manner, stabilizing the U2 snRNA/BS in-teraction (11–18), and thereby regulate BS recognition and se-lection. Except for splicing, U2 snRNP also functions in the 3′processing of polyadenylated and nonpolyadenylated mRNAs(19, 20). On polyadenylated pre-mRNAs, U2 snRNP componentsincluding SF3b interact with the cleavage and polyadenylationspecificity factor and enhance the rate of 3′ processing (19). Onnonpolyadenylated histone pre-mRNAs, SF3b155, the largestSF3b subunit, together with Prp43, directly makes contact withthe 7-nt motif, C/GAAGAAG, present in the coding region andfacilitates 3′ processing as a component of U2 snRNP (20). Todate, all known roles of SF3b are executed in the context of
U2 snRNP, and whether it has a U2-snRNP–independent roleremains unknown.The highly conserved TREX complex plays key roles in mRNA
export (21, 22). It mainly contains three parts: the THO sub-complex (THOC1/2/3/5/6/7) and proteins ALYREF and UAP56(21, 23–26). ALYREF and THO serve as mRNA export adaptorsthat link the mRNA to the export receptor, a heterodimer ofNXF1/NXT1 (27–29). In higher eukaryotes, splicing significantlypromotes TREX recruitment and mRNA export (23, 24, 30). Ithas been thought that physical interaction(s) between splicing andmRNA export machineries mediate splicing-dependent TREXrecruitment. However, these interactions remain to be identified.In this work, we found that SF3b interacts with the THO com-
plex in vivo and in vitro. Loss-of-function and gain-of-functiondata demonstrate a role of SF3b in promoting mRNA export byrecruiting THO/TREX. In support of this role, SF3b bindsintronless and spliced mRNAs in the cells. Intriguingly, disrup-tion of U2 snRNP formation does not inhibit, but promotes,SF3b-mediated mRNA export by enhancing the SF3b–THO in-teraction and boosting THO recruitment to the mature mRNA.Thus, our work identifies SF3b as a linker between splicing andmRNA export machineries and suggests that the balance betweenpre-mRNA processing and mRNA export can be regulated throughalteration of SF3b distribution in U2 snRNPs and mature mRNPs.
Significance
To ensure efficient and accurate gene expression, pre-mRNAprocessing and mRNA export need to be balanced. SF3b is acore component of U2 snRNP that participates in splicing and 3′processing of pre-mRNAs. Here, we uncovered a role of SF3b inpromoting mRNA export by directly binding with the maturemRNA and recruiting the key mRNA export adaptor THO. In-triguingly, this role is not dependent on U2 snRNP, but is ratherenhanced upon U2 snRNP disruption as a result of enhancedSF3b–THO interaction and recruitment to the mRNA. Together,our work uncovers an important role of SF3b in promotingmRNA export and suggests that competition between U2 snRNPand the mature mRNP for SF3b contributes to balancing pre-mRNA processing and mRNA export.
Author contributions: K.W., Y.Z., J.H., and H.C. designed research; K.W., C.Y., S.C., J.W.,L.Z., L.W., Y.Y., B.C., M.S., and C.W. performed research; R.R. contributed new reagents/analytic tools; K.W., X.D., Y.Z., and H.C. analyzed data; and H.C. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Published under the PNAS license.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1818835116/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1818835116 PNAS Latest Articles | 1 of 10
BIOCH
EMISTR
Y

ResultsSF3b Associates with THO in Vivo, and This Association Is Strengthenedin the Absence of RNAs. With the interest to understand the mech-anism for splicing-dependent mRNA export, we examined the in-teractions of TREX components with splicing factors. In an attemptof THOC2 immunoprecipitation (IP) from RNase A-treated HeLanuclear extract (NE), SF3b components, including SF3b155 andSF3b130, were specifically present in the THOC2 immunoprecip-itate but not the control (Cntl; i.e., IgG) immunoprecipitate (Fig.1A and Dataset S1). RRP6, a component of the exosome that is not
known to interact with THO or SF3b, was not detected in eitherimmunoprecipitate (Fig. 1A). Considering that THO and SF3b aretightly bound complexes (15, 21, 23, 26, 31), this result suggests thatTHO might interact with SF3b. To further investigate this in-teraction, we carried out reverse IPs by using an SF3b155 antibodyfrom HeLa NE treated or not treated with RNase A, followed byMS. An antibody against all subunits of SF3a, another U2 snRNPcomponent, was also included. As expected, SF3a and SF3b sub-units, as well as the two other U2 snRNP components, A′ and B″,were present in both immunoprecipitates, and RNase A treatment
Fig. 1. SF3b interacts with THO in vivo and in vitro. (A) The THOC2 antibody coprecipitated SF3b. Western blot analysis to detect components of SF3b (SF3b155,SF3b130) and THO (THOC2, THOC5) complexes immunoprecipitated by the Cntl (i.e., IgG) and THOC2 antibodies. RRP6 serves as a negative control. (B) MS to detectproteins associated with SF3a and SF3b155. Total peptide counts of components of U2 snRNP and THO in the immunoprecipitates of SF3a and SF3b155 in the presenceor absence of RNase A are shown. (C) The SF3b155 antibody coprecipitated THO. Western blot analysis to detect components of SF3b (SF3b155, SF3b130), SF3a(SF3a120, SF3a66, SF3a60), and THO (THOC2, THOC1, THOC5) complexes immunoprecipitated by the Cntl (i.e., IgG) and SF3b155 antibodies with or without RNaseA treatment. RRP6 serves as a negative control. The numbers shows the ratio of each protein coimmunoprecipitated from NE in the presence (+) or absence (−) ofRNase A. (D) Strep-tagged THO pulled down the SF3b complex. Inputs and proteins pulled down were detected by Coomassie staining and Western blotting. Strep-tagged MBP was used as a negative control. The white line delineates the boundary where the irrelevant lane has been removed from the same gel and blots.(E) Strep-tagged THO pulled down the SF3b core. Same as D, except that, instead of the whole SF3b complex, the SF3b core was used.
2 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1818835116 Wang et al.
weakened their coprecipitations, as indicated by reduced peptideenrichment (Fig. 1B and Dataset S2). In support of the SF3b–THOinteraction, several THO proteins were present in the immuno-precipitate of SF3b155 but not that of SF3a. Intriguingly, theamounts of all these proteins apparently increased upon RNase Atreatment (Fig. 1B).To validate the MS data, SF3b155 and Cntl IPs were repeated in
the absence or presence of RNase A, followed by Western analysisusing antibodies to subunits of SF3b (SF3b155 and SF3b130), SF3a(SF3a120, SF3a66, and SF3a60), and THO (THOC1, THOC2, andTHOC5). The RRP6 antibody was used as a negative control. Asshown in Fig. 1C, all of these proteins except RRP6 were appar-ently precipitated by the SF3b155 antibody, but not the Cntl.Consistent with the MS data, RNase A treatment resulted in re-duced association of SF3b155 with SF3a subunits and concomi-tantly enhanced association with THO subunits (Fig. 1C). Thesedata indicate that SF3b associates with THO and that this associ-ation is strengthened in the absence of RNAs.
SF3b Directly Interacts with THO via the Core Proteins. We nextasked whether SF3b interacts with THO in vitro. To this end, weexpressed and purified THO and SF3b complexes in insect cells.In the THO complex, THOC1 was strep-tagged and used forpull-downs. Strep-tagged MBP was used as a negative control.Significantly, Western blotting showed that SF3b155 and SF3b130were apparently pulled down by the THO complex but not byMBP, indicating that THO and SF3b interact in vitro (Fig. 1D).To further validate this interaction and narrow down the THO-interacting SF3b components, we next used the SF3b core (32),which is composed of SF3b155 (residues 454–1304), SF3b130,SF3b14b, and SF3b10, for pull-downs. Because the SF3b155 an-tibody recognizes two N-terminal fragments (residues 330–344,450–463) that are completely or partially deleted in the SF3b core,we used only the SF3b130 antibody for Western blot analysis. Asshown in Fig. 1E, SF3b130 was more apparently detected in thepull-down of strep-tagged THO even though significantly morestrep-tagged MBP was pulled down. Together, these data indicatethat SF3b directly interacts with THO via the core proteins.
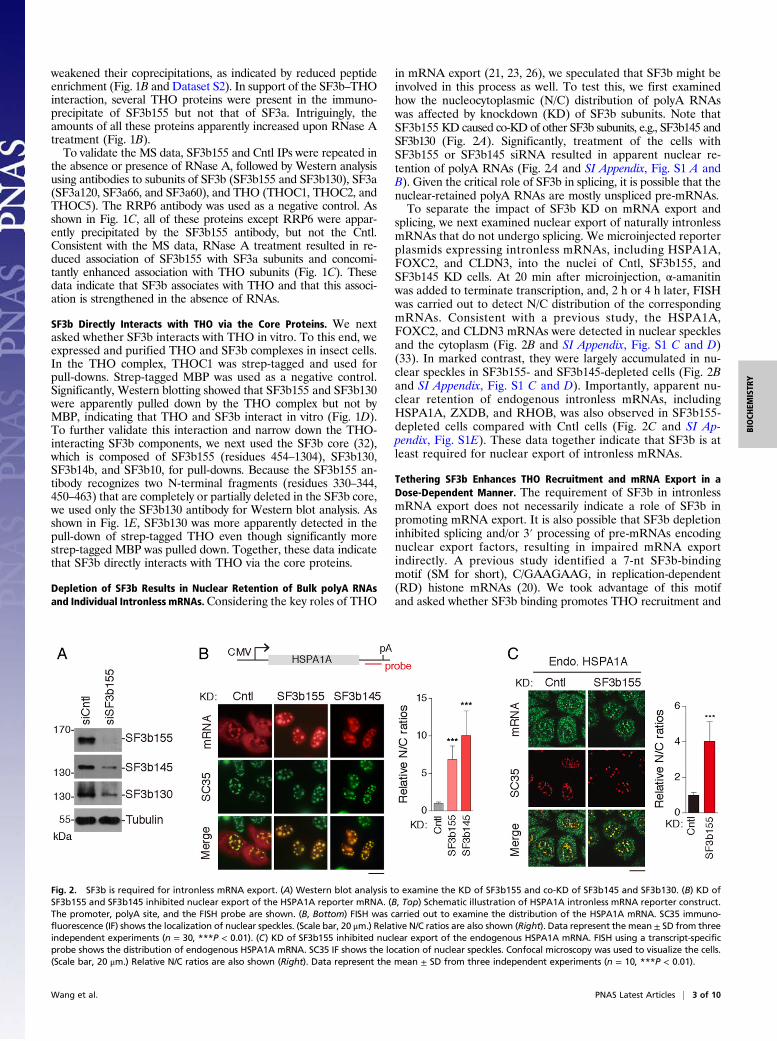
Depletion of SF3b Results in Nuclear Retention of Bulk polyA RNAsand Individual Intronless mRNAs.Considering the key roles of THO
in mRNA export (21, 23, 26), we speculated that SF3b might beinvolved in this process as well. To test this, we first examinedhow the nucleocytoplasmic (N/C) distribution of polyA RNAswas affected by knockdown (KD) of SF3b subunits. Note thatSF3b155 KD caused co-KD of other SF3b subunits, e.g., SF3b145 andSF3b130 (Fig. 2A). Significantly, treatment of the cells withSF3b155 or SF3b145 siRNA resulted in apparent nuclear re-tention of polyA RNAs (Fig. 2A and SI Appendix, Fig. S1 A andB). Given the critical role of SF3b in splicing, it is possible that thenuclear-retained polyA RNAs are mostly unspliced pre-mRNAs.To separate the impact of SF3b KD on mRNA export and
splicing, we next examined nuclear export of naturally intronlessmRNAs that do not undergo splicing. We microinjected reporterplasmids expressing intronless mRNAs, including HSPA1A,FOXC2, and CLDN3, into the nuclei of Cntl, SF3b155, andSF3b145 KD cells. At 20 min after microinjection, α-amanitinwas added to terminate transcription, and, 2 h or 4 h later, FISHwas carried out to detect N/C distribution of the correspondingmRNAs. Consistent with a previous study, the HSPA1A,FOXC2, and CLDN3 mRNAs were detected in nuclear specklesand the cytoplasm (Fig. 2B and SI Appendix, Fig. S1 C and D)(33). In marked contrast, they were largely accumulated in nu-clear speckles in SF3b155- and SF3b145-depleted cells (Fig. 2Band SI Appendix, Fig. S1 C and D). Importantly, apparent nu-clear retention of endogenous intronless mRNAs, includingHSPA1A, ZXDB, and RHOB, was also observed in SF3b155-depleted cells compared with Cntl cells (Fig. 2C and SI Ap-pendix, Fig. S1E). These data together indicate that SF3b is atleast required for nuclear export of intronless mRNAs.
Tethering SF3b Enhances THO Recruitment and mRNA Export in aDose-Dependent Manner. The requirement of SF3b in intronlessmRNA export does not necessarily indicate a role of SF3b inpromoting mRNA export. It is also possible that SF3b depletioninhibited splicing and/or 3′ processing of pre-mRNAs encodingnuclear export factors, resulting in impaired mRNA exportindirectly. A previous study identified a 7-nt SF3b-bindingmotif (SM for short), C/GAAGAAG, in replication-dependent(RD) histone mRNAs (20). We took advantage of this motifand asked whether SF3b binding promotes THO recruitment and
Fig. 2. SF3b is required for intronless mRNA export. (A) Western blot analysis to examine the KD of SF3b155 and co-KD of SF3b145 and SF3b130. (B) KD ofSF3b155 and SF3b145 inhibited nuclear export of the HSPA1A reporter mRNA. (B, Top) Schematic illustration of HSPA1A intronless mRNA reporter construct.The promoter, polyA site, and the FISH probe are shown. (B, Bottom) FISH was carried out to examine the distribution of the HSPA1A mRNA. SC35 immuno-fluorescence (IF) shows the localization of nuclear speckles. (Scale bar, 20 μm.) Relative N/C ratios are also shown (Right). Data represent the mean ± SD from threeindependent experiments (n = 30, ***P < 0.01). (C) KD of SF3b155 inhibited nuclear export of the endogenous HSPA1A mRNA. FISH using a transcript-specificprobe shows the distribution of endogenous HSPA1A mRNA. SC35 IF shows the location of nuclear speckles. Confocal microscopy was used to visualize the cells.(Scale bar, 20 μm.) Relative N/C ratios are also shown (Right). Data represent the mean ± SD from three independent experiments (n = 10, ***P < 0.01).
Wang et al. PNAS Latest Articles | 3 of 10
BIOCH
EMISTR
Y

mRNA export. To this end, we inserted one copy or three copiesof SM into the AdML cDNA (cA) construct, which does notcontain any SM (SI Appendix, Fig. S2). To exclude the possibilitythat SF3b binding impacts THO recruitment through promoting 3′processing, we used in vitro transcribed mRNAs that do not un-dergo 3′ processing. The cA, cA-1SM, and cA-3SM mRNAs wereincubated separately in HeLa NE, followed by IPs using antibodiesto Cntl, SF3b155, and THOC2. As expected, the cA mRNA wasbarely bound by SF3b155, and insertion of SM progressively
promoted SF3b155 binding as the number increased (Fig. 3A).Consistent with previous studies, the cA mRNAwas not efficientlyassociated with THOC2 (24, 30). Significantly, 1SM slightlystrengthened THOC2 binding and 3SM showed much moreapparent effect (Fig. 3A). These results together indicate thattethering SF3b promotes THO recruitment in a dose-dependentmanner. We next asked whether SF3b is required for efficientTHO recruitment to SM-containing mRNAs by depleting SF3b155from the NE. Consistent with the notion that SF3b is a tightly
Fig. 3. Tethering SF3b enhances THO recruitment and mRNA export in a dose-dependent manner. (A) Insertion of SM to the mRNA enhanced THO re-cruitment. (A, Top) Schematic illustration of cA, cA-1SM, and cA-3SM mRNAs. (A, Bottom) RIPs of in vitro-transcribed cA, cA-1SM, and cA-3SM mRNAs fromHeLa NE with indicated antibodies. IgG was used as the control. One fourth of the input was loaded. Relative enrichments are also shown (Bottom). Datarepresent the mean ± SEM (n = 3, *P < 0.05 and ***P < 0.01). (B) SF3b was required for the effect of SM on promoting THO recruitment. In vitro-transcribedcA-3SM mRNA was used for RIPs from mock- or SF3b155-depleted HeLa NE with indicated antibodies. The CBP80 antibody and the IgG were used as positiveand negative controls, respectively. One fourth of the input was loaded. Relative enrichments are also shown (Bottom). Data represent the mean ± SEM (n = 3,*P < 0.05 and ***P < 0.01; n.s., not significant). (C) Same as B, except that, instead of the cA-3SM mRNA, the FOXC2 mRNA was used. (D) Insertion of SM to themRNA enhanced its nuclear export. (D, Top) Schematic illustration of the cA, cA-1SM, and cA-3SM expression constructs. The promoter, polyA site, and probe areshown. (D, Bottom) FISH to detect the cA, cA-1SM, and cA-3SM mRNAs transcribed from microinjected constructs. DAPI staining was used to mark the nucleus.(Scale bar, 20 μm.) Relative N/C ratios are also shown (Right). Data represent the mean ± SD from three independent experiments (n = 30, ***P < 0.01). (E) Theeffect of SM on promoting THO recruitment depended on SF3b and TREX. (E, Top) Schematic illustration of the cA-3SM expression construct. The promoter,polyA site, and probe are shown. (E, Bottom) FISH to detect the cA-3SMmRNA in Cntl, SF3b155, and UAP56 KD cells. DAPI staining was used to mark the nucleus.(Scale bar, 20 μm.) Relative N/C ratios are also shown (Right). Data represent the mean ± SD from three independent experiments (n = 30, ***P < 0.01).
4 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1818835116 Wang et al.

bound complex, SF3b155 depletion led to codepletion of otherSF3b subunits, e.g., SF3b49 (SI Appendix, Fig. S1F). Signifi-cantly, SF3b depletion significantly reduced the binding of THO,but not that of CBP80, with the cA-3SM mRNA (Fig. 3B).Similarly, THO recruitment to the FOXC2 mRNA, which con-tains three copies of SM (SI Appendix, Fig. S2), was also ap-parently weakened by SF3b depletion (Fig. 3C). Thus, SF3bbinding with the mRNA promotes THO recruitment.To examine whether SF3b binding promotes mRNA export,
we microinjected the same set of cA reporter constructs intoHeLa cell nuclei. As expected, the cA mRNA was exclusivelynuclear at 2 h postinjection (Fig. 3D). In contrast, the cA-1SMmRNA could be detected in the cytoplasm, and more significantcytoplasmic accumulation was observed with the cA-3SM mRNA(Fig. 3D), indicating that SF3b binding indeed promotes mRNAexport. Previously, it was shown that an SM-containing 22-ntRNA element promotes mRNA export by recruiting NXF1through SR proteins (34, 35). Thus, it was also possible that theSM promoted mRNA export via SR-mediated NXF1 recruit-ment. This possibility was not supported by the observation thatdepletion of SF3b155 or the TREX component UAP56 blockedthe cA-3SM mRNA in the nucleus (Fig. 3E). In addition toSF3b155, the SM also interacts with Prp43 (20). However, Prp43is unlikely a key factor in SM-mediated mRNA export, as Prp43KD showed only moderate impact on cA-3SM mRNA export (SIAppendix, Fig. S1G and H). Together, these data indicate that SF3bpromotes mRNA export by recruiting the THO/TREX complex.
SF3b Binds Intronless and Spliced mRNAs in the Cells. Does SF3bactually bind intronless mRNAs in the cells? We analyzed thepresence of SM in exons of intronless and intron-containingprotein-coding genes from an Ensembl gene set (human hg19).This analysis revealed that one third of intronless genes containat least one copy of SM, supporting the possibility that SF3bbinds intronless mRNAs in the cells (Fig. 4A). Interestingly, twothirds of intron-containing genes harbor SM in their exon se-quences (Fig. 4A), suggesting that SF3b might bind splicedmRNAs as well. To examine these possibilities, we analyzed SF3b155enhanced cross-linking and IP (eCLIP) data (36) (SI Appendix, Fig.S3A). As expected, the vast majority (94%) of eCLIP peaks weremapped to protein-coding genes (SI Appendix, Fig. S3B), andmost of these peaks (74.6%) were distributed in introns (Fig.4B). Importantly, in agreement with the notion that SF3b bindson intronless and spliced mRNAs, a significant fraction (22.5%)of SF3b binding sites were mapped to intronless genes as well asexons of intron-containing genes (Fig. 4B). More importantly,eCLIP reads were apparently enriched at exon–exon junctions inboth replicates (3.38% and 3.97%) compared with the input(1.31%; SI Appendix, Table S1). Motif analysis identified threeapparently enriched motifs in SF3b155 binding peaks on exonsequences (SI Appendix, Fig. S3C). Although the top one GAGGAresembles the SM, the other two (GUUGGU and CGGGGGC)do not, suggesting that SF3b-binding sequences on exons arehighly heterogenous. This provides an explanation for why noapparent enrichment of SF3b binding was detected on SM-containing intronless genes compared with SM-lacking ones (SIAppendix, Fig. S3D).To confirm SF3b binding on mature mRNAs, we next carried
out RNA IPs (RIPs) with the SF3b155 antibody, followed byquantitative RT-PCRs (RT-qPCR; Fig. 4 C–E). As expected,SF3b155 was apparently enriched on the U2 snRNA but not the18S rRNA (Fig. 4D). Consistent with previous studies (11–13,20), pre-mRNAs, including UAP56 and URH49, and RD his-tone mRNAs, including H1-H2AG, H1-H3H, and H3-H2A, wetested were specifically enriched by the SF3b155 antibody (Fig.4E). Significantly, SF3b association on nonhistone intronless mRNAs,including RHOB, HSPA1A, and HSPA6, and spliced mRNAs,including UAP56 and URH49, was also reproducibly detected,
although to less of an extent compared with that on pre-mRNAs(Fig. 4E). Together, these data indicate that SF3b binds not onlyon pre-mRNAs but also on mature mRNAs.
Inhibition of U2 snRNP Function Does Not Inhibit, but Promotes,mRNA Export. To date, all known roles of SF3b are executed inthe context of U2 snRNP (8–10, 19, 20). To investigate whether
Fig. 4. SF3b prevalently binds intronless and spliced mRNAs. (A) Pie chartsshow that more than one third of intronless mRNAs and two thirds of splicedmRNAs contain the SM motif in the exon sequences. (B) Pie chart shows thedistribution of SF3b155 eCLIP peaks detected on protein-coding genes. (C)Western analysis to detect SF3b155 immunoprecipitated by the Cntl (i.e.,IgG) and SF3b155 antibodies. (D) SF3b associated with U2 snRNA. RT-qPCRsto examine the associations of SF3b155 with U2 snRNA and 18S rRNA. RNAsenriched with Cntl were set as 1. Data represent the mean ± SEM (n = 3,***P < 0.01; n.s., not significant). (E) SF3b associated with mature mRNAs.RT-qPCRs to examine the associations of SF3b155 with pre-mRNAs, intronlessmRNAs, and spliced mRNAs. RNAs enriched with Cntl were set as 1. Datarepresent the mean ± SEM (n = 3, *P < 0.05 and ***P < 0.01).
Wang et al. PNAS Latest Articles | 5 of 10
BIOCH
EMISTR
Y

this is also true for mRNA export, we used an antisense mor-pholino oligonucleotide (AMO) against U2 snRNA to inhibitU2 snRNP functions (37–39). Treatment of the cells with the U2AMO, but not the Cntl AMO, significantly inhibited splicing ofβ-globin and Smad reporter pre-mRNAs (SI Appendix, Fig. S4A),indicative of inhibited U2 snRNP functions.We next coinjected the FOXC2 expression construct with dif-
ferent amounts of Cntl or U2 AMO. To allow sufficient U2 snRNPinhibition, we incubated the cells for 8 h, with α-amanitin omittedto avoid cell toxicity resulting from long-term treatment. In cellstreated with low or high Cntl AMO, the FOXC2 mRNA waslargely detected in the nucleus, probably because of its continuoustranscription (Fig. 5A). Unexpectedly, apparent cytoplasmic accu-mulation of the FOXC2 mRNA was detected with low U2 AMO,and the mRNA was mostly detected in the cytoplasm with high U2AMO treatment, indicating that U2 snRNP inhibition did not im-pair, but enhanced, FOXC2 mRNA export (Fig. 5A). Similarly, U2AMO treatment also promoted cytoplasmic accumulation of theCLDN3 reporter mRNA that contains one copy of SM (SI Ap-pendix, Figs. S2 and S4B). Importantly, the effect of U2 AMO onenhancing mRNA export was also confirmed with endogenousintronless mRNAs, HSPA1A and RHOB. Although Cntl AMO didnot apparently impact their N/C distribution, apparently enhancedcytoplasmic accumulation was observed in U2 AMO-treated cellscompared with the surrounding nontreated cells (Fig. 5B and SIAppendix, Fig. S4C).Notably, KD of SF3b subunits, including SF3b155, SF3b145,
and SF3b49, completely inhibited the effect of U2 AMO onmRNA export (Fig. 5C and SI Appendix, Fig. S4 D and E), in-dicating that U2 AMO treatment induced mRNA export pro-motion was dependent on the SF3b complex. It was possible thatthis promotion was caused by increased availability of nuclear ex-port factors for intronless mRNAs as a result of reduced abundanceof spliced mRNAs caused by splicing inhibition. However, althoughU1 AMO treatment also blocked splicing, it did not enhance cy-toplasmic accumulation of the FOXC2 mRNA (SI Appendix, Fig.S5). Together, these data indicate that inhibition of U2 snRNPfunctions promotes intronless mRNA export.As SF3b also associates with spliced mRNAs, we next sought
to examine how nuclear export of spliced mRNAs was affectedupon U2 snRNP inhibition. Considering the difficulty in dis-tinguishing pre-mRNAs and spliced mRNAs based on FISHanalysis, we carried out cell fractionation and RT-qPCRs. Con-sistent with the previously described FISH data, U2 AMOtreatment resulted in apparently increased cytoplasm/total-cell(C/T) ratios of intronless mRNAs, including HSPA1A andRHOB. Significantly, the C/T ratios of spliced mRNAs we ex-amined, including UAP56, URH49, FOS, FOXF2, and SF3b130,were all enhanced in U2 AMO-treated cells compared with Cntlcells (Fig. 5 D and E). Together, these data indicate that the roleof SF3b in promoting mRNA export is not dependent on U2snRNP, but is rather enhanced upon U2 snRNP inhibition. Infurther support of a U2 snRNP-independent role of SF3b inmRNA export, SF3a and B″ did not apparently associate withthe intronless or spliced mRNAs we examined, although theywere easily detected on the U2 snRNA and pre-mRNAs (Fig. 5Fand SI Appendix, Fig. S6).
Disruption of U2 snRNP Formation Enhances the SF3b–THO Interactionand Their Recruitment to Mature mRNAs. How does U2 AMOtreatment enhance mRNA export? Our initial observation thatRNase A treatment led to enhanced SF3b–THO interactionreminded us of the possibility that U2 AMO treatment mightdisrupt U2 snRNP. Indeed, IPs with the B″ antibody showedweakened bands corresponding to all SF3a and some SF3bsubunits in the U2 AMO-treated NE compared with the Cntl NEon the silver-stained gel (SI Appendix, Fig. S7A). These weak-ened associations of SF3a and SF3b with B″ were validated by
Western blotting (Fig. 6A). Further, the associations of SF3a,SF3b, and B″ with U2 snRNA were significantly reduced in thepresence of U2 AMO (SI Appendix, Fig. S7 B and C). These datasuggest that U2 AMO treatment disrupted the formation of asignificant fraction of U2 snRNP. The disrupted U2 snRNP wasfurther validated by a more direct method, the sedimentation ofthe U2 snRNPs on glycerol gradients. U2 AMO treatment causedan apparent shift of U2 snRNA and B″, but not U1 snRNA or theU1 snRNP protein U1A, to low-molecular weight (MW) fractions(SI Appendix, Fig. S8A). Similarly, U1 AMO treatment alsocaused a specific shift of U1 snRNA and U1-70K, a specific U1snRNP component, to low-MW fractions (SI Appendix, Fig. S8B),suggesting that U1 snRNP was also disrupted by U1 AMO.In agreement with the view that U2 snRNP disruption leads to
enhanced SF3b–THO interaction, reproducibly strengthenedassociation of SF3b155 with THO proteins, including THOC2,THOC1, and THOC5, and reduced interaction with SF3a weredetected in the U2 AMO NE compared with the Cntl NE (Fig.6B). Further, SF3b155, THOC2, and ALYREF RIPs using invitro-transcribed FOXC2 mRNA and HeLa NE treated withCntl or U2 AMO revealed that U2 snRNP disruption promotesrecruitments of SF3b and THO/TREX to the mRNA (Fig. 6C).Notably, when the cA and cA-1SM mRNAs were used for IPs, U2AMO treatment enhanced the bindings of SF3b155 and THOC2with the cA-1SMmRNA but not the cA mRNA (SI Appendix, Fig.S9), indicating that U2 snRNP disruption specifically promotesTHO/TREX recruitment to mRNAs with SF3b-binding sites.Together, these data indicate that SF3b interacts with THO andpromotes mRNA export independent of U2 snRNP, and suggestthat the pool of SF3b in mature mRNPs competes with thatpresent in U2 snRNP.
DiscussionTo ensure efficient and accurate gene expression, nuclear RNAexport needs to be tightly controlled. Accumulating evidenceindicates that nuclear export machinery is limited to ensure onlyRNAs that are fully processed and properly assembled intomRNPs could be exported to the cytoplasm (3, 40). As the price,a fraction of mature mRNAs are retained in the nucleus andsubject to degradation even in normal cells (7, 41). Producing alarge number of mature mRNAs that ultimately undergo deg-radation is obviously not economic, and one way to avoid that isto ensure the balance between pre-mRNA processing andmRNA export.Our study raises the possibility that the usage of common
factors in pre-mRNA processing and mRNA export could pro-vide a mechanism for balancing these processes. SF3b preva-lently binds on pre-mRNAs, where it participates in splicing and3′ processing in the context of U2 snRNP. Here we found that italso binds on mature mRNAs to promote nuclear export byrecruiting THO/TREX through direct interactions. Intriguingly,the role of SF3b in mRNA export does not rely on, but competeswith, that of U2 snRNP. These findings suggest that SF3b islimited and that alteration in SF3b distribution in U2 snRNPsand mature mRNPs impacts the balance between mRNA pro-cessing and export. In line with this, compared with the totalabundance of U2 snRNA and mRNA (∼106 and 2–4 × 105, re-spectively; refs. 42–44), the concentration of SF3b is relatively low(∼3 × 105; SI Appendix, Fig. S10). This SF3b-mediated balancecould be important to ensure the majority of fully processedmRNAs are transported to the cytoplasm in normal condition, asonly when released from mature mRNAs could SF3b enter thenext cycle for pre-mRNA processing as a component of U2 snRNP.Also, it might be vital for guaranteeing mRNA export and proteinexpression when cells are under stressed conditions, such as heatshock, which causes U snRNP disassembly (45). Many other splicingfactors, such as SR proteins, are also bound on mature mRNAs andpromote mRNA nuclear export (22, 33–35, 46–51). It would be
6 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1818835116 Wang et al.

interesting to investigate whether these factors also contribute tobalancing pre-mRNA processing and mRNA export. What otheradvantages could there be for utilizing SF3b, rather than U2 snRNP,in promoting mRNA export? It might be important to avoid pre-mRNA leakage by preventing THO recruitment to pre-mRNAs bythe U2 snRNP. Also, U2 snRNP bound on mature mRNAs mightbe recognized by the surveillance machinery at the nuclear pore (52,53) that likely precludes efficient mRNA export.
Our findings could also provide mechanistic insights into se-quence- and splicing-dependent mRNA export. We found thatSM exists in one third of naturally intronless mRNAs, and SF3bbindings are detected on them. Importantly, we provide evidencethat this motif indeed promotes intronless mRNA export by en-hancing THO/TREX recruitment. In addition to SM, we identifiedother motifs based on SF3b eCLIP data. In the future, it would beinteresting to investigate whether these motifs are functional for
Fig. 5. Inhibition of U2 snRNP promotes mRNA export. (A) U2 AMO treatment promoted FOXC2 mRNA export. FISH to examine the distribution of theFOXC2 mRNA derived from the construct coinjected with a low amount (1×) or high amount (3×) of Cntl or U2 AMO. The injection marker was used to markinjected cells. (Scale bar, 20 μm.) Relative N/C ratios are also shown (Right). Data represent the mean ± SD from three independent experiments (n = 30, ***P <0.01). (B) U2 AMO treatment promoted nuclear export of endogenous HSPA1A mRNA. Distribution of the endogenous HSPA1A mRNA was detected with itsspecific probe in cells injected with Cntl or U2 AMO. GFP was used to indicate injected cells. DAPI staining was used to mark the nuclei. (Scale bar, 20 μm.) RelativeN/C ratios are also shown (Right). Data represent the mean ± SD from three independent experiments (n = 30, ***P < 0.01). (C) The effect of U2 AMO treatment onpromoting mRNA export required SF3b155. FISH to examine the distribution of the FOXC2 mRNA derived from the construct coinjected with Cntl or U2 AMO incells treatedwith Cntl or SF3b155 siRNA. DAPI staining was used to mark the nuclei. (Scale bar, 20 μm.) Relative N/C ratios are also shown (Right). Data represent themean ± SD from three independent experiments (n = 15, ***P < 0.01; n.s., not significant). (D) U2 AMO treatment enhanced nuclear export of spliced mRNAs. RT-qPCRs examining the cytoplasmic distribution of intronless and spliced mRNAs with Cntl or U2 AMO. mRNA levels in the cytoplasm (marked as “C”) and total cells(“T”) were examined, and the relative C/T ratios are shown. The C/T ratios in the Cntl AMO sample were set as 1. Data represent the mean ± SEM (n = 3, *P <0.05 and ***P < 0.01). (E) Western analysis to assess the purities of the nuclear and cytoplasmic fractions. UAP56 and tubulin are markers for nucleus and cytoplasm,respectively. (F) SF3a and B″ did not associate with mature mRNAs. RT-qPCRs to examine the associations of SF3b155, SF3a, and B″ with pre-mRNAs, intronlessmRNAs, and spliced mRNAs. RNAs enriched with IgG were set as 1. Data represent the mean ± SEM (n = 3, *P < 0.05 and ***P < 0.01; n.s., not significant).
Wang et al. PNAS Latest Articles | 7 of 10
BIOCH
EMISTR
Y

SF3b binding and mRNA export. Although, in the present work, wecould not directly examine whether SF3b promotes spliced mRNAexport as a result of its critical role in splicing, we speculate that theSF3b–THO interaction possibly contributes to connecting mRNAexport to splicing. SF3b is recruited to the BS region of pre-mRNAsas a component of U2 snRNP. During splicing, when SF3b is re-leased from U2 snRNP (17, 18, 54–58), it might be handed over tospliced mRNAs via specific sequences present on exons and recruitTHO (see model in Fig. 6D). Further studies are needed to in-vestigate this hypothesis in future.In recent years, recurrent mutations have been identified in
SF3b155 in many types of cancers (32, 59, 60). Since this dis-covery, many efforts have been made to detect altered splicingevents that are causative for tumorigenesis (2, 60–62). However,to date, no convincing evidence has been obtained supporting
that SF3b155 mutations lead to cancer development as a resultof splicing defects. The role of SF3b in mRNA export describedhere could open a new direction for investigation of the un-derlying mechanism for SF3b155 mutations causing cancers.
Materials and MethodsCell Culture and Transfection. HeLa cells were cultured in DMEM supple-mented with 10% FBS and penicillin/streptomycin. For siRNA transfection,HeLa cells seeded in 12-well plates or glass-bottomed dishes were transfectedwith 10 pmol of siRNA per well using Lipofectamine RNAi Max (Invitrogen)following the manufacturer’s protocol. DNA transfection was performed byusing Lipofectamine 2000 (Invitrogen) following the manufacturer’s pro-tocol. AMOs (1 μM) were introduced to HeLa cells by using electroporation(NEPA21) according to the manufacturer’s recommendations. The siRNA andAMO target sequences are listed in SI Appendix, Table S2.
Fig. 6. Disruption of U2 snRNP enhances the SF3b–THO interaction and their recruitment to the mRNA. (A) U2 AMO treatment reduced the association ofSF3b and SF3a with B”. Western blot analysis to detect SF3b and SF3a subunits immunoprecipitated by the Cntl (i.e., IgG) and the B″ antibody from HeLa NEtreated with Cntl or U2 AMO. (B) Western blot analysis to detect components of SF3b, SF3a, and THO complexes immunoprecipitated by the Cntl (i.e., IgG)and SF3b155 antibodies from HeLa NE treated with Cntl or U2 AMO. (C) U2 AMO treatment enhanced TREX recruitment to the FOXC2 mRNA. In vitro-transcribed FOXC2 mRNA was incubated under splicing condition with HeLa NE and treated with Cntl or U2 AMO, followed by IPs with the indicated an-tibodies, IgG was used as the Cntl. One fourth of the input was loaded. Relative enrichments are also shown (Bottom). Data represent the mean ± SEM (n = 3,*P < 0.05 and ***P < 0.01). (D) A model for two distinct pools of SF3b in U2 snRNP and the mature mRNP. In the normal condition, two distinct SF3b pools existin the cells: SF3b bound on the pre-mRNA participates in its processing as a component of U2 snRNP, and SF3b bound on mature mRNAs promotes nuclearexport by recruiting the THO. Upon U2 snRNP disassembly or disruption, SF3b is released and increasingly available for the assembly of export-competentmRNPs, resulting in reduced pre-mRNA processing and enhanced mRNA export.
8 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1818835116 Wang et al.

Pull-Downs. Untagged SF3b or SF3b core complex (50 μg) was mixed with50 μg of the strep-tagged THO complex or the strep-tagged MBP proteinand 40 μL of Strep-Tactin Superflow beads (IBA) in a buffer of 50 mM Tris·HCl, pH 7.5, and 150 mM (for SF3b complex) or 500 mM (for SF3b corecomplex) NaCl and incubated for 4 h at 4 °C. Then, the beads were exten-sively washed four times, and the bound proteins were eluted with 5 mMD-desthiobiotin and separated on a 12% SDS/PAGE gel.
RNA Extraction and RT-qPCRs. Total RNA was extracted by using TRI reagent(Sigma-Aldrich) and treated with RNase-free RQ1 DNase I (Promega) toremove genomic DNA. Random primer was used for reverse transcription byM-MLV reverse transcriptase (Promega). Quantitative PCR was carried out byusing GoTaq Master Mix (Promega) according to the manufacturer’s in-structions. The primers used for quantitative PCR are listed in SI Appendix,Table S3.
DNAMicroinjections.HeLa cells used for microinjection were plated on 20-mmcoverslips at the bottom of 35-mm dishes. Plasmid DNA (100 ng/μL) wascoinjected with FITC-conjugated 70-kDa dextran (injection marker). For eachexperiment, ∼300 cells were microinjected, followed by incubation at 37 °C.After 20 min of incubation, transcription was terminated with α-amanitin(4 μg/mL; Sigma-Aldrich), and incubation was continued for the desiredperiod of time before fixation. For U1- or U2-snRNP disruption, control, U1,or U2 AMO was coinjected with plasmid DNA and α-amanitin was omitted.
Protein IPs. To identify SF3a- and SF3b155-interacting proteins, the antibodyto the whole SF3a complex or to SF3b155 was covalently cross-linked tonProtein A Sepharose (GE Healthcare) by dimethyl pimelimidate. A total of350 μL of HeLa NE was treated or not treated with RNase A under splicingcondition (6 mM Tris, pH 7.6, 30 mM KCl, 3.2 mM MgCl2, 20 mM creatinephosphate, 0.05 mM ATP) at 30 °C for 20 min and then incubated with an-tibody cross-linked beads in IP buffer (1× PBS solution, 0.1% Triton, 0.2 mMPMSF, and protease inhibitor) overnight. The beads were washed withwashing buffer (1× PBS solution, 0.1% Triton, 0.2 mM PMSF) three times,and the proteins were eluted with SDS loading buffer and analyzed by ion-trap MS. For protein IPs followed by Western blot, 75 μL of HeLa NE was usedfor IPs. For IPs following disruption of U2 snRNP formation, HeLa NE wascoincubated with control or U2 AMO (4 μM) at 30 °C for 20 min. For THOC2IP-Western experiment, the HeLa NE was diluted with IP buffer and treatedwith RNase A at 30 °C for 20 min and then incubated with antibody cross-linked beads overnight.
Immunodepletion and RIPs from NEs. For immunodepletions, antibodies werecovalently cross-linked to protein A Sepharose beads (GE Healthcare) at a
4:1 ratio (crude serum volume:packed beads volume) by using dimethylpimelimidate (Sigma). A total of 100 μL of high-salt HeLa NE (350 mM) wasmixed with 25 μL of beads and rotated for 2 h at 4 °C. The procedure wasrepeated three times. RIPs were carried out as previously described (30).Phosphorus-32–labeled in vitro-transcribed mRNAs were incubated in splic-ing reaction mixtures containing 30% HeLa NE for 2 h (63). To disruptU2 snRNP formation, HeLa NE was coincubated with Cntl or U2 AMO at 30 °Cfor 2 h. For IPs, 5 μL of splicing reaction and 100 μL of binding buffer (20 mMHepes, pH 7.9, 150 mM KCl, 0.1% Triton, 2.5 mM EDTA, and 5 mM DTT) wereincubated with antibody-coupled protein A Sepharose beads at 4 °C for 2 h.After extensive washing with the binding buffer, immunoprecipitates weretreated with proteinase K (Roche) at 37 °C for 10 min, and RNAs were re-covered by phenol/chloroform extraction and ethanol precipitation. RNAwas analyzed on denaturing polyacrylamide gels and visualized by Phos-phorImager. One fourth of the input was loaded.
RIPs from Whole-Cell Lysates. HeLa cell pellets were resuspended in 1 mL ofNET-2 buffer [50 mM Tris·HCl, pH 7.4, 150 mM NaCl, 0.1% Tergitol (NP-40),0.2 mM PMSF], followed by sonication and centrifuge. The lysates were in-cubated with the indicated antibodies for 2 h at 4 °C, followed by rotationwith nProtein A Sepharose (GE Healthcare) for another 2 h at 4 °C. Theimmunoprecipitates were washed three times with the NET-2 buffer. Onefifth of the immunoprecipitate was analyzed by Western blotting. The restof the immunoprecipitate was treated with proteinase K, and RNAs wererecovered by phenol/chloroform extraction and ethanol precipitation. ForRIPs following disruption of U2 snRNP formation, HeLa lysates were coin-cubated with Cntl or U2 AMO at 30 °C for 20 min before incubation with theindicated antibodies.
Sedimentation Experiments. A total of 400 μL of NE preincubated with Cntl,U1, or U2 AMO was layered on a 10–35% linear glycerol gradient containing150 mM KCl, 20 mM Hepes/KOH, pH 8.0, and 1.5 mM MgCl2. After centrifu-gation at 32,000 rpm for 17 h in a Beckman SW41 rotor, fractions were col-lected. The RNA of each gradient fraction was extracted by TRI reagent andvisualized by SYBR Gold (Invitrogen) staining. The protein of each fractionwas acetone-precipitated, followed by SDS/PAGE and Western blotting.
ACKNOWLEDGMENTS. We thank Ming Lei for providing THO expressionconstructs and Dangsheng Li and Jing Fan for useful discussion and criticalcomments on the manuscript. This work was funded by National Key R&DProgram of China Grant 2017YFA0504400 and National Natural ScienceFoundation of China Grants 31570822, 31770880, 31800686, and 91640115.This work was also supported by “Strategic Priority Research Program” ofthe Chinese Academy of Sciences Grant XDB19000000.
1. Kaida D, et al. (2007) Spliceostatin A targets SF3b and inhibits both splicing and nu-clear retention of pre-mRNA. Nat Chem Biol 3:576–583.
2. Yoshimoto R, et al. (2017) Global analysis of pre-mRNA subcellular localization fol-lowing splicing inhibition by spliceostatin A. RNA 23:47–57.
3. Zhou Z, et al. (2000) The protein Aly links pre-messenger-RNA splicing to nuclearexport in metazoans. Nature 407:401–405.
4. Rougemaille M, et al. (2007) Dissecting mechanisms of nuclear mRNA surveillance inTHO/sub2 complex mutants. EMBO J 26:2317–2326.
5. Saguez C, et al. (2008) Nuclear mRNA surveillance in THO/sub2 mutants is triggered byinefficient polyadenylation. Mol Cell 31:91–103.
6. Herold A, Teixeira L, Izaurralde E (2003) Genome-wide analysis of nuclear mRNAexport pathways in Drosophila. EMBO J 22:2472–2483.
7. Fan J, et al. (2017) Exosome cofactor hMTR4 competes with export adaptor ALYREF toensure balanced nuclear RNA pools for degradation and export. EMBO J 36:2870–2886.
8. Will CL, Lührmann R (2011) Spliceosome structure and function. Cold Spring HarbPerspect Biol 3:a003707.
9. Shi Y (2017) Mechanistic insights into precursor messenger RNA splicing by the spli-ceosome. Nat Rev Mol Cell Biol 18:655–670.
10. Fica SM, Nagai K (2017) Cryo-electron microscopy snapshots of the spliceosome:Structural insights into a dynamic ribonucleoprotein machine. Nat Struct Mol Biol 24:791–799.
11. Gozani O, Feld R, Reed R (1996) Evidence that sequence-independent binding ofhighly conserved U2 snRNP proteins upstream of the branch site is required for as-sembly of spliceosomal complex A. Genes Dev 10:233–243.
12. Gozani O, Potashkin J, Reed R (1998) A potential role for U2AF-SAP 155 interactions inrecruiting U2 snRNP to the branch site. Mol Cell Biol 18:4752–4760.
13. Wang C, et al. (1998) Phosphorylation of spliceosomal protein SAP 155 coupled withsplicing catalysis. Genes Dev 12:1409–1414.
14. Das BK, et al. (1999) Characterization of a protein complex containing spliceosomalproteins SAPs 49, 130, 145, and 155. Mol Cell Biol 19:6796–6802.
15. Will CL, et al. (2001) A novel U2 and U11/U12 snRNP protein that associates with thepre-mRNA branch site. EMBO J 20:4536–4546.
16. Bertram K, et al. (2017) Cryo-EM structure of a pre-catalytic human spliceosomeprimed for activation. Cell 170:701–713.e11.
17. Haselbach D, et al. (2018) Structure and conformational dynamics of the humanspliceosomal Bact complex. Cell 172:454–464.e11.
18. Zhang X, et al. (2018) Structure of the human activated spliceosome in three con-formational states. Cell Res 28:307–322.
19. Kyburz A, Friedlein A, Langen H, Keller W (2006) Direct interactions between subunitsof CPSF and the U2 snRNP contribute to the coupling of pre-mRNA 3′ end processingand splicing. Mol Cell 23:195–205.
20. Friend K, Lovejoy AF, Steitz JA (2007) U2 snRNP binds intronless histone pre-mRNAs tofacilitate U7-snRNP-dependent 3′ end formation. Mol Cell 28:240–252.
21. Strässer K, et al. (2002) TREX is a conserved complex coupling transcription withmessenger RNA export. Nature 417:304–308.
22. Reed R, Cheng H (2005) TREX, SR proteins and export of mRNA. Curr Opin Cell Biol 17:269–273.
23. Chi B, et al. (2013) Aly and THO are required for assembly of the human TREX complexand association of TREX components with the spliced mRNA. Nucleic Acids Res 41:1294–1306.
24. Masuda S, et al. (2005) Recruitment of the human TREX complex to mRNA duringsplicing. Genes Dev 19:1512–1517.
25. Jimeno S, Rondón AG, Luna R, Aguilera A (2002) The yeast THO complex and mRNAexport factors link RNA metabolism with transcription and genome instability. EMBOJ 21:3526–3535.
26. Rehwinkel J, et al. (2004) Genome-wide analysis of mRNAs regulated by the THOcomplex in Drosophila melanogaster. Nat Struct Mol Biol 11:558–566.
27. Viphakone N, et al. (2012) TREX exposes the RNA-binding domain of Nxf1 to enablemRNA export. Nat Commun 3:1006.
28. Segref A, et al. (1997) Mex67p, a novel factor for nuclear mRNA export, binds to bothpoly(A)+ RNA and nuclear pores. EMBO J 16:3256–3271.
29. Strässer K, Hurt E (2000) Yra1p, a conserved nuclear RNA-binding protein, interactsdirectly with Mex67p and is required for mRNA export. EMBO J 19:410–420.
30. Cheng H, et al. (2006) Human mRNA export machinery recruited to the 5′ end ofmRNA. Cell 127:1389–1400.
Wang et al. PNAS Latest Articles | 9 of 10
BIOCH
EMISTR
Y

31. Will CL, et al. (2002) Characterization of novel SF3b and 17S U2 snRNP proteins, in-cluding a human Prp5p homologue and an SF3b DEAD-box protein. EMBO J 21:4978–4988.
32. Cretu C, et al. (2016) Molecular architecture of SF3b and structural consequences of itscancer-related mutations. Mol Cell 64:307–319.
33. Wang K, et al. (2018) Intronless mRNAs transit through nuclear speckles to gain ex-port competence. J Cell Biol 217:3912–3929.
34. Huang Y, Steitz JA (2001) Splicing factors SRp20 and 9G8 promote the nucleocyto-plasmic export of mRNA. Mol Cell 7:899–905.
35. Huang Y, Gattoni R, Stévenin J, Steitz JA (2003) SR splicing factors serve as adapterproteins for TAP-dependent mRNA export. Mol Cell 11:837–843.
36. Van Nostrand EL, et al. (2016) Robust transcriptome-wide discovery of RNA-bindingprotein binding sites with enhanced CLIP (eCLIP). Nat Methods 13:508–514.
37. Kaida D, et al. (2010) U1 snRNP protects pre-mRNAs from premature cleavage andpolyadenylation. Nature 468:664–668.
38. Matter N, König H (2005) Targeted ‘knockdown’ of spliceosome function in mam-malian cells. Nucleic Acids Res 33:e41.
39. König H, Matter N, Bader R, Thiele W, Müller F (2007) Splicing segregation: The minorspliceosome acts outside the nucleus and controls cell proliferation. Cell 131:718–729.
40. Kallehauge TB, Robert MC, Bertrand E, Jensen TH (2012) Nuclear retention preventspremature cytoplasmic appearance of mRNA. Mol Cell 48:145–152.
41. Silla T, Karadoulama E, Makosa D, Lubas M, Jensen TH (2018) The RNA exosomeadaptor ZFC3H1 functionally competes with nuclear export activity to retain targettranscripts. Cell Rep 23:2199–2210.
42. Craft J (1992) Antibodies to snRNPs in systemic lupus erythematosus. Rheum Dis ClinNorth Am 18:311–335.
43. Eberwine J, Sul JY, Bartfai T, Kim J (2014) The promise of single-cell sequencing. NatMethods 11:25–27.
44. Shapiro E, Biezuner T, Linnarsson S (2013) Single-cell sequencing-based technologieswill revolutionize whole-organism science. Nat Rev Genet 14:618–630.
45. Bracken AP, Bond U (1999) Reassembly and protection of small nuclear ribonucleo-protein particles by heat shock proteins in yeast cells. RNA 5:1586–1596.
46. Fu XD (1993) Specific commitment of different pre-mRNAs to splicing by single SRproteins. Nature 365:82–85.
47. Chandler SD, Mayeda A, Yeakley JM, Krainer AR, Fu XD (1997) RNA splicing specificitydetermined by the coordinated action of RNA recognition motifs in SR proteins. ProcNatl Acad Sci USA 94:3596–3601.
48. Liu HX, Zhang M, Krainer AR (1998) Identification of functional exonic splicing en-hancer motifs recognized by individual SR proteins. Genes Dev 12:1998–2012.
49. Liu HX, Chew SL, Cartegni L, Zhang MQ, Krainer AR (2000) Exonic splicing enhancermotif recognized by human SC35 under splicing conditions. Mol Cell Biol 20:1063–1071.
50. Müller-McNicoll M, et al. (2016) SR proteins are NXF1 adaptors that link alternativeRNA processing to mRNA export. Genes Dev 30:553–566.
51. Chanarat S, Seizl M, Strässer K (2011) The Prp19 complex is a novel transcriptionelongation factor required for TREX occupancy at transcribed genes. Genes Dev 25:1147–1158.
52. Galy V, et al. (2004) Nuclear retention of unspliced mRNAs in yeast is mediated byperinuclear Mlp1. Cell 116:63–73.
53. Coyle JH, Bor YC, Rekosh D, Hammarskjold ML (2011) The Tpr protein regulates ex-port of mRNAs with retained introns that traffic through the Nxf1 pathway. RNA 17:1344–1356.
54. Bessonov S, Anokhina M, Will CL, Urlaub H, Lührmann R (2008) Isolation of an activestep I spliceosome and composition of its RNP core. Nature 452:846–850.
55. Zhan X, Yan C, Zhang X, Lei J, Shi Y (2018) Structure of a human catalytic step Ispliceosome. Science 359:537–545.
56. Ohrt T, et al. (2012) Prp2-mediated protein rearrangements at the catalytic core ofthe spliceosome as revealed by dcFCCS. RNA 18:1244–1256.
57. Lardelli RM, Thompson JX, Yates JR, 3rd, Stevens SW (2010) Release of SF3 from theintron branchpoint activates the first step of pre-mRNA splicing. RNA 16:516–528.
58. Warkocki Z, et al. (2009) Reconstitution of both steps of Saccharomyces cerevisiaesplicing with purified spliceosomal components. Nat Struct Mol Biol 16:1237–1243.
59. Malcovati L, et al.; Chronic Myeloid Disorders Working Group of the InternationalCancer Genome Consortium and of the Associazione Italiana per la Ricerca sul CancroGruppo Italiano Malattie Mieloproliferative (2011) Clinical significance of SF3B1mutations in myelodysplastic syndromes and myelodysplastic/myeloproliferative neo-plasms. Blood 118:6239–6246.
60. Wang L, et al. (2011) SF3B1 and other novel cancer genes in chronic lymphocyticleukemia. N Engl J Med 365:2497–2506.
61. Fan L, Lagisetti C, Edwards CC, Webb TR, Potter PM (2011) Sudemycins, novel smallmolecule analogues of FR901464, induce alternative gene splicing. ACS Chem Biol 6:582–589.
62. Kesarwani AK, et al. (2017) Cancer-associated SF3B1 mutants recognize otherwiseinaccessible cryptic 3′ splice sites within RNA secondary structures. Oncogene 36:1123–1133.
63. Krainer AR, Maniatis T, Ruskin B, Green MR (1984) Normal and mutant human beta-globin pre-mRNAs are faithfully and efficiently spliced in vitro. Cell 36:993–1005.
10 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1818835116 Wang et al.

Supplemental Information
Plasmids and Antibodies
To construct FOXC2 reporter plasmid, the coding sequence of FOXC2 was
inserted to pcDNA3 (Invitrogen). HSPA1A, CLDN3, wG, and wS constructs
were described previously (1-3). The AdML cDNA (cA) sequence (4) was
inserted into pcDNA3 to construct the cA plasmid. One or three tandem
repeats of GAAGAAG were inserted into the cA construct to make the cA-1SM
and cA-3SM constructs. The coding sequences of SF3b or THO complex
components were inserted to pRSFduet (ThermoFisher) to construct the
expression plasmids. The coding sequences of MBP or GST were inserted
into pRSFduet respectively.
The detailed information of antibodies against SF3a (SF3a120, SF3a66,
SF3a60), SF3b155, SF3b130, and B’’ were described previously (5-8). The
ALYREF, CBP80, THOC1, THOC2 and THOC5 antibodies were the same as
the ones used in previous studies (9). The RRP6 (Sigma-Aldrich), SF3b145
www.pnas.org/cgi/doi/10.1073/pnas.1818835116

(ProteinTech), Tubulin (Sigma-Aldrich), SF3b49 (ProteinTech), Prp43
(ProteinTech), SC35 (Sigma-Aldrich), GFP (Sigma-Aldrich), Dig (Roche),
U1A (SANTA CRUZ) and U1-70K (ABclonal) antibodies, and IgG (SANTA
CRUZ) were purchased. Alexa Fluor 546, Alexa Fluor 488, or Alexa Fluor
647 conjugated secondary antibodies were purchased from Life Technologies.
Protein Preparation for Pull-down Assays
The human THO complex, including THOC1-Strep, His6-THOC2 (residues
1-1190), THOC3, THOC5, THOC6, and THOC7, was co-expressed with
Bac-to-Bac Baculovirus Expression System (ThermoFisher) and purified using
Ni-NTA (Qiagen) and Strep-Tactin (GE Healthcare) resins sequentially. After
a final step of gel-filtration chromatography on Superdex 200 10/300 GL (GE
Healthcare), the purified peak fractions were directly used for the in vitro
pull-down assay. The human SF3b complex, composed of
Twin-StrepII-3C-SF3b155, SF3b145, His6-3C-SF3b130, SF3b49, SF3b14a,
SF3b14b and SF3b10, and the SF3b core complex, composed of

Twin-StrepII-3C-SF3b155 (residues 454-1304), His6-3C-SF3b130, SF3b14b,
and SF3b10, was expressed and purified in a similar way as that of the THO
complex, except that the Twin-StrepII tag was removed through the Protease 3C
digestion. The MBP-Twin-StrepII proteins were expressed in E. coli and
purified through a Strep-Tactin affinity chromatography and a gel-filtration
chromatography on Superdex 200 10/300 GL (GE Healthcare).
Fluorescence in situ Hybridization and Immunofluorescence
FISH for detection of polyA RNAs was performed as previously described (10).
To detect the mRNAs derived from microinjected plasmids, an HPLC-purified
Alexa 548 conjugated probe (3’ vector probe) that hybridizes to pcDNA3 vector
sequence upstream of the polyA site was used. Images were captured with a
DP72-CCD camera (Olympus) on an inverted microscope using DP-BSW
software (Olympus). FISH quantitation was carried out using ImageJ software
(National Institutes of Health), and N/C ratios were calculated as described (2).
For immunofluorescence, the SC35 and GFP antibodies were diluted 1: 2,000

and 1: 1,000 in blocking buffer, respectively.
FISH for detection of endogenous mRNAs was performed as previously
described (1). The endogenous HSPA1A, ZXDB and RHOB mRNAs were
detected using the in vitro transcribed digoxin-labeled probe. HeLa cells
treated with siRNAs or AMOs were fixed with 3.6% formaldehyde plus 10%
acetic acid in 1x PBS solution for 20 min, followed with three washes with 1 x
PBS solution and permeabilized with 1 x PBS solution/0.1% Triton/2 mM VRC
for 15 min. The cells were incubated with dig-labeled probes in hybridization
buffer at 50 0C for 16 h. After extensive wash, cells were incubated with
digoxin antibody for 1 h. After three washes with 1x PBS solution, the cells
were incubated with the Alexa-488 labeled anti-sheep antibody for 1 h,
followed by DAPI staining. Confocal imaging was performed using an A1
plus laser-scanning confocal microscope and acquired with NIS Elements
(Nikon) software. The detailed information of the probes is shown in SI
Appendix, Table S4.

eCLIP Data Analysis
SF3b155 eCLIP-seq and the size matched input (SMInput) fastq files were
downloaded from ENCODE with accessions ENCSR133QEA and ENCSR496UBM,
respectively. The pair-end reads were processed by following the ENCODE
released pipeline (11). Briefly, adaptors in reads were trimmed with cutadapt
program and mapped against human RepBase with STAR to remove reads with
potential repeat elements. The remaining reads were mapping to the human genome
(hg19) including splicing junctions from GENCODE gene annotation (v19) with
STAR. Usable reads were obtained by removing potential PCR duplicates from
uniquely mapped reads. The SF3b155 binding peaks were detected by computing
eCLIP read 2 signal enrichment and fold change compared with SMInput with
CLIPPer (12). For motif analysis, only eCLIP peaks on exons were used.
Backgrounds were produced by bedtools shuffle with peaks. Then findMotifs.pl in
homer program was used to produce motif logos with background and peaks (13).

Statistics
All results were repeated at least three times in independent experiments. Data
were presented as mean ± SD or mean ± SEM as figure legend mentioned and
statistical significance was determined using unpaired t-test which was
two-sided. The variance is similar between the groups that are being
statistically compared. Data sets were analyzed for statistical significance
using GraphPad Prism software (version 6). P < 0.05 was considered as
statistically significant.
Supplemental Figure and Figure Legend

Figure S1. KD of SF3b155 and SF3b145 both result in nuclear retention
of polyA RNAs as well as individual intronless mRNAs, in support of Fig.
2.
(A) Western blot analysis to examine the KD efficiency of SF3b145.

(B) KD of SF3b155 and SF3b145 resulted in nuclear retention of bulk polyA
RNAs. FISH using an oligo dT probe to examine nucleocytoplasmic
distribution of polyA RNAs in Cntl, SF3b155 and SF3b145 KD cells. Scale
bar, 20 µm. Relative N/C ratios are shown on the right. Data represent the
mean ± SD from three independent experiments; n = 30. ***P < 0.01.
(C) KD of SF3b155 resulted in nuclear retention of reporter intronless mRNAs.
Top: schematic of naturally intronless mRNA reporter constructs. The
promoter, polyA site, and probe are shown. Bottom: FISH was carried out to
examine the distribution of the FOXC2 and CLDN3 mRNA. SC35 IF shows
the localization of nuclear speckles. Scale bar, 20 µm. Relative N/C ratios
are shown on the right. Data represent the mean ± SD from three independent
experiments; n = 30. ***P < 0.01.
(D) Same as (C), except that SF3b145 KD cells were used.
(E) SF3b155 KD inhibited endogenous intronless mRNA export. FISH using
a transcript-specific probe to examine distribution of the endogenous ZXDB
and RHOB mRNAs. SC35 IF shows the location of nuclear speckles.

Confocal microscopy was used to visualize the cells. Scale bar, 20 µm.
Relative N/C ratios are shown on the right. Data represent the mean ± SD
from three independent experiments; n = 10. ***P < 0.01.
(F) Western blot analysis to examine the levels of SF3b155, SF3b49, THOC2
and CBP80 in SF3b155-immunodepleted NE.
(G) Western blot analysis to examine the KD efficiency of Prp43.
(H) Prp43 KD moderately inhibited nuclear export of the cA-3SM mRNA.
FISH to detect the cA-3SM mRNA transcribed from microinjected construct.
DAPI staining was used to mark the nucleus. Scale bar, 20 µm. Relative
N/C ratios are shown on the right. Data represent the mean ± SD from three
independent experiments; n = 30. ***P < 0.01.

Figure S2. Numbers of SM present in reporter and endogenous mRNAs, in
support of Figs 2-6.

Figure S3. SF3b binds intronless and spliced mRNAs, in support of Fig. 4.
(A) Correlation of SF3b155 eCLIP-seq replicates.
(B) Pie chart shows that more than 94% of SF3b155 binding targets are
mRNAs.
(C) SF3b-binding motifs identified based on SF3b155 eCLIP data.
(D) SF3b155 eCLIP read counts mapped to SM-containing and SM-lacking
intronless genes.

Figure S4. Inhibition of U2 snRNP function promotes nuclear export of
intronless mRNAs, in support of Fig. 5.
(A) U2 AMO treatment inhibited splicing. RT-qPCRs to examine splicing of
β-globin (wG) and Smad (wS) pre-mRNAs in Cntl or U2 AMO treated cells
with primers targeting intron-exon and exon-exon junctions, respectively.
Data represent the mean ± SEM, n=3. ***P < 0.01.

(B) U2 AMO treatment enhanced CLDN3 reporter mRNA export. Top:
schematic of the CLDN3 reporter construct. The promoter, polyA site, and
probe are shown. Bottom: FISH was carried out to examine the distribution of
the CLDN3 mRNA in cells injected with Cntl or U2 AMO. Scale bar, 20 µm.
Relative N/C ratios are shown on the right. Data represent the mean ± SD
from three independent experiments; n = 30. ***P < 0.01.
(C) U2 AMO treatment enhanced nuclear export of the endogenous RHOB
mRNA. Distribution of endogenous RHOB intronless mRNAs was detected
with their specific probes in cells injected with Cntl or U2 AMO. GFP was
used to indicate injected cells. DAPI staining was used to mark the nuclei.
Scale bar, 20 µm. Relative N/C ratios are shown on the right. Data represent
the mean ± SD from three independent experiments; n = 30. ***P < 0.01.
(D) Western blot analysis to examine the KD efficiency of SF3b49. Tubulin was
used as a loading control. The * represents the SF3b49 signal from previous
blotting.

(E) Depletion of SF3b49 and SF3b145 eliminated the effect of U2 AMO on
promoting mRNA export. FISH to examine the distribution of the FOXC2
mRNA derived from the construct co-injected with Cntl or U2 AMO in Cntl
SF3b49, and SF3b145 KD cells. Scale bar, 20 µm. Relative N/C ratios are
shown on the right. Data represent the mean ± SD from three independent
experiments; n = 30. ***P < 0.01; n.s., not significant.
Figure S5. Inhibition of U1 snRNP functions does not promote nuclear
export of intronless mRNAs, in support of Fig. 5.
(A) U1 AMO treatment inhibited splicing. The wG or wS expression
constructs was co-injected with Cntl or U1 AMO, followed by RT-qPCRs to
specifically detect the pre-mRNAs and spliced mRNAs with primers targeting

intron-exon and exon-exon junctions, respectively. Data represent the mean ±
SEM, n=3. ***P < 0.01.
(B) U1 AMO treatment did not enhance FOXC2 mRNA export. Equal amount
of FOXC2 expression construct was co-injected with Cntl, U1 or U2 AMO,
followed by FISH with the 3’ vector probe. DAPI staining was used to mark
the nuclei. Scale bar, 20 µm. Relative N/C ratios are shown on the right.
Data represent the mean ± SD from three independent experiments; n = 30.
***P < 0.01.
Figure S6. SF3b, SF3a, and B” efficiently associate with U2 snRNA, in
support of Fig. 5.
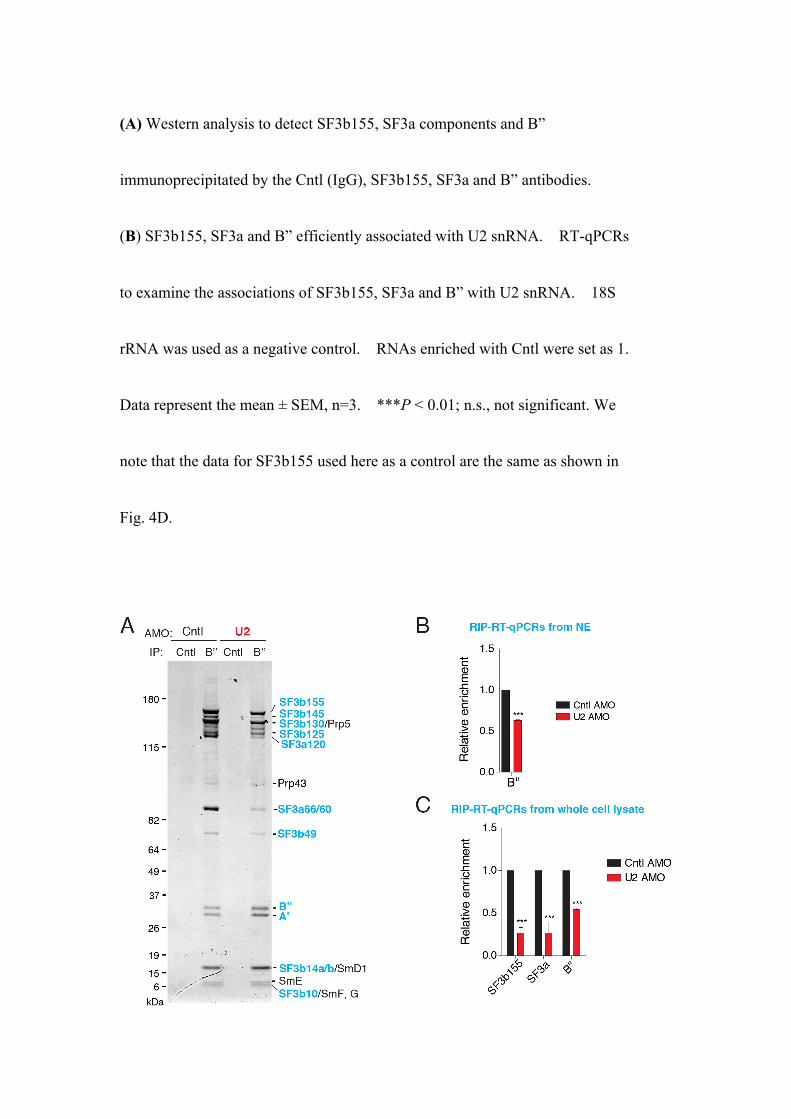
(A) Western analysis to detect SF3b155, SF3a components and B”
immunoprecipitated by the Cntl (IgG), SF3b155, SF3a and B” antibodies.
(B) SF3b155, SF3a and B” efficiently associated with U2 snRNA. RT-qPCRs
to examine the associations of SF3b155, SF3a and B” with U2 snRNA. 18S
rRNA was used as a negative control. RNAs enriched with Cntl were set as 1.
Data represent the mean ± SEM, n=3. ***P < 0.01; n.s., not significant. We
note that the data for SF3b155 used here as a control are the same as shown in
Fig. 4D.

Figure S7. U2 AMO inhibits the associations of SF3a and SF3b with B” and
U2 snRNA, in support of Fig. 6.
(A) Silver staining of immunoprecipitates of Cntl (IgG) and B” antibodies
from HeLa NEs treated with Cntl or U2 AMO.
(B) U2 AMO treatment inhibited the association of B” with U2 snRNA in the
NE. RIP-RT-qPCRs to detect the association of B” with U2 snRNA in HeLa
cell NE treated with Cntl or U2 AMO. Data represent the mean ± SEM, n=3.
***P < 0.01.
(C) U2 AMO treatment inhibited the associations of SF3a, SF3b, and B” with
U2 snRNA. RIP-RT-qPCRs to detect the association of SF3a, SF3b and B”
with U2 snRNA in cell lysates treated with Cntl or U2 AMO. Data represent
the mean ± SEM, n=3. ***P < 0.01.

Figure S8. U2- and U1-snRNPs are disrupted by U2- and U1-AMO,
respectively.
(A) U2 AMO treatment caused an apparent shift of U2 snRNA and B” on the
glycerol gradients. Sedimentation analysis of U2 snRNP in nuclear extract
treated with Cntl or U2 AMO by linear 10-35% glycerol gradient centrifugation.
The top panel shows SYBR Gold staining of the RNAs from the gradient
fractions. Proteins from each fraction were analyzed by western blotting using

antibodies to B” and U1A (lower panels). The peak positions of 18S and 28S
rRNAs, run in parallel, are indicated at the bottom.
(B) Same as (A), except that U1 AMO was used instead of U2 AMO and U1-70K
antibody was used for western blotting.
Figure S9. The effect of U2 disruption on THO recruitment depends on
SF3b binding, in support of Fig. 6.
In vitro transcribed cA and cA-1SM mRNA were incubated under splicing
condition with HeLa nuclear extract treated with Cntl or U2 AMO, followed by
IPs with the indicated antibodies. IgG was used as the control. One fourth of
the input was loaded. Quantification was shown on the right. Data represent
the mean ± SEM, n=3. *P<0.05, ***P<0.01; n.s., not significant.

Figure S10. Estimation of SF3b concentration in the cells, in support of Fig.
6.
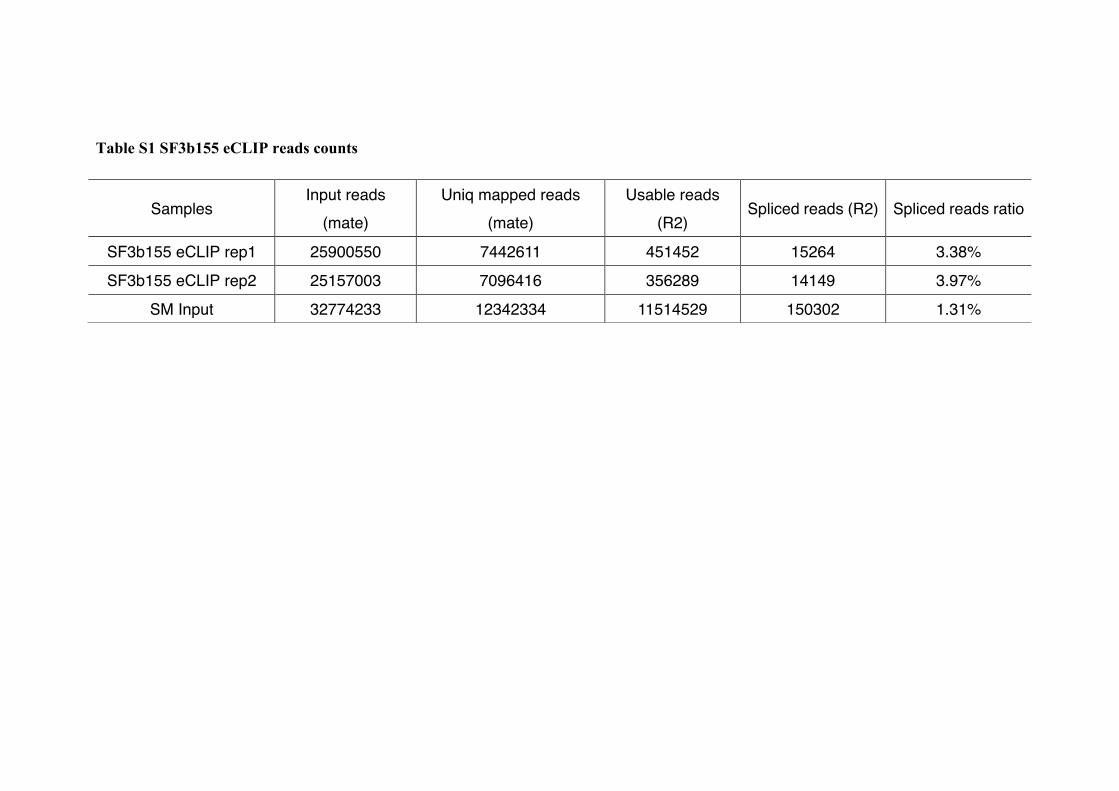
Table S1 SF3b155 eCLIP reads counts
Samples Input reads
(mate) Uniq mapped reads
(mate) Usable reads
(R2) Spliced reads (R2) Spliced reads ratio
SF3b155 eCLIP rep1 25900550 7442611 451452 15264 3.38%
SF3b155 eCLIP rep2 25157003 7096416 356289 14149 3.97%
SM Input 32774233 12342334 11514529 150302 1.31%

Table S2 siRNA and AMO targeting sequences
Name Target Sequence (5’ to 3’)
siCntl CGTACGCGGAATACTTCGA
siSF3b155 TTCTGACCAAGCAAACTCG
siSF3b145 GGACGTGTTTGGAACCAAT
siPrp43 GGTGACAGCTATTAGTAAA
siUAP56 AAGGGCTTGGCTATCACAT
siURH49 AAAGGCCTAGCCATCACTT
siSF3b49 GCATCAGCTCACAACAAAA
Cntl AMO TATAAATTGTAACTGAGGTAAGAGG
U1 AMO ATACTTACCTGGCAGGGGAGATACC
U2 AMO TCAAGTGTAGTATCTGTTCTTATCA

Table S3 Primer sequences
Name Sequence (5’ to 3’) β-globin pre F CAATAGAAACTGGGCATGTGGA β-globin pre R CCATAACAGCATCAGGAGTGGA
β-globin mature F AGAACTTCAGGCTCCTGGGC β-globin mature R TCTGGGACGTCGTATGGGTAG
Smad pre F GGTGGAGACACCAGGTGAGTAC Smad pre R CAAGAGGTGGCAGCTCTGTC
Smad mature F ACACCAGCAGTTTTACCACCTG Smad mature R ATGGAATGCGTGTAGTCATCAAG
UAP56 pre F GACAGCAGCTGGGGGAGATG UAP56 pre R CCAATGAGCACTACATGCC
UAP56 mature F GACAGCAGCTGGGGGAGATG UAP56 mature R CTCATGCTGGACTTCTGACG
URH49 pre F GTGTCTGACGAGAATGATGC URH49 pre R CTGCTCGACTGTAAGACATG
URH49 mature F GTGTCTGACGAGAATGATGC URH49 mature R CTCTGCTCGATGTATGTGG
U2 snRNA F TCTGATACGTCCTCTATCCGAG U2 snRNA R GTTCCTGGAGGTACTGCAATAC 18s rRNA F GGCCCTGTAATTGGAATGAGTC 18s rRNA R CCAAGATCCAACTACGAGCTT HSPA1A F TGCTCCGACCTGTTCCG HSPA1A R CTCGTCGGGGTTGATGC
RHOB F AAGGGTGGTGATGGGTG RHOB R CTGGTGGAGGGTTCGCA HSPA6 F ACCACCTACTCGGACAACC HSPA6 R CACGCTCAGGATGCCATTAG
HIST1H3H F CAGGACTTCAAGACCGAC HIST1H3H R ATGATAGTCACCCGCTTG
HIST1H2AG F AGTCACCATCGCACAGG HIST1H2AG R AGAGCCGTTGGTTTGGAC HIST3H2A F AAGGCTAAGTCGCGCTCG HIST3H2A R CCAGGATCTCGGCAGTCA
FOS mature F TCTCTTACTACCACTCACCCGC FOS mature R GCTCTGGTCTGCGATGGG
FOXF2 mature F CCTTACCTCAAGCAGCCG FOXF2 mature R TGGTGATAATACGACCCGCTA
SF3b130 mature F GCCCAGCCTGAGTCCTTGT SF3b130 mature R AGACAAATCCCCAGTGACAGG

Table S4 Probe sequences
Name Gene ID Sequences(5’ to 3’)
3’ Vector Probe - AAGGCACGGGGGAGGGGCAAACAACAGATGGCTGG CAACTAGAAGGCACAGTCGAGGCTGATCAGCGGGT
Oligo dT Probe - dT50
HSPA1A Probe NM_005345.5 Antisense sequence of human HSPA1A mRNA region (51-1206 nt)
RHOB Probe NM_004040.3 Antisense sequence of human RHOB mRNA region (408-1409 nt)
ZXDB Probe NM_007157.3 Antisense sequence of human ZXDB mRNA region (214-1206 nt)

Reference
1. Wang K, et al. (2018) Intronless mRNAs transit through nuclear speckles to
gain export competence. J Cell Biol 217(11):3912-3929.
2. Valencia P, Dias AP, & Reed R (2008) Splicing promotes rapid and efficient
mRNA export in mammalian cells. Proc Natl Acad Sci U S A
105(9):3386-3391.
3. Shi M, et al. (2015) Premature Termination Codons Are Recognized in the
Nucleus in A Reading-Frame Dependent Manner. Cell Discov 1.
4. Luo MJ & Reed R (1999) Splicing is required for rapid and efficient mRNA
export in metazoans. Proc Natl Acad Sci U S A 96(26):14937-14942.
5. Das BK, et al. (1999) Characterization of a protein complex containing
spliceosomal proteins SAPs 49, 130, 145, and 155. Mol Cell Biol
19(10):6796-6802.
6. Das R, Zhou Z, & Reed R (2000) Functional association of U2 snRNP with
the ATP-independent spliceosomal complex E. Mol Cell 5(5):779-787.
7. Wang C, et al. (1998) Phosphorylation of spliceosomal protein SAP 155
coupled with splicing catalysis. Genes Dev 12(10):1409-1414.
8. Habets WJ, Hoet MH, De Jong BA, Van der Kemp A, & Van Venrooij WJ
(1989) Mapping of B cell epitopes on small nuclear ribonucleoproteins that
react with human autoantibodies as well as with experimentally-induced
mouse monoclonal antibodies. J Immunol 143(8):2560-2566.
9. Chi B, et al. (2013) Aly and THO are required for assembly of the human
TREX complex and association of TREX components with the spliced mRNA.
Nucleic Acids Res 41(2):1294-1306.
10. Chi B, et al. (2014) A Sub-Element in PRE enhances nuclear export of
intronless mRNAs by recruiting the TREX complex via ZC3H18. Nucleic
Acids Res 42(11):7305-7318.
11. Van Nostrand EL, et al. (2016) Robust transcriptome-wide discovery of
RNA-binding protein binding sites with enhanced CLIP (eCLIP). Nat Methods
13(6):508-514.
12. Lovci MT, et al. (2013) Rbfox proteins regulate alternative mRNA splicing
through evolutionarily conserved RNA bridges. Nat Struct Mol Biol

20(12):1434-1442.
13. Heinz S, et al. (2010) Simple combinations of lineage-determining
transcription factors prime cis-regulatory elements required for macrophage
and B cell identities. Mol Cell 38(4):576-589.





























