Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY,0022-538X/00/$04.0010
Mar. 2000, p. 2227–2238 Vol. 74, No. 5
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
A Structured RNA Motif Is Involved in Correct Placement ofthe tRNA3
Lys Primer onto the Human ImmunodeficiencyVirus Genome
NANCY BEERENS, BEP KLAVER, AND BEN BERKHOUT*
Department of Human Retrovirology, Academic Medical Center, University of Amsterdam,Amsterdam, The Netherlands
Received 3 September 1999/Accepted 22 November 1999
Human immunodeficiency virus type 1 (HIV-1) reverse transcription is primed by the cellular tRNA3Lys
molecule that binds with its 3*-terminal 18 nucleotides to the fully complementary primer-binding site (PBS)on the viral RNA genome. Besides this complementarity, annealing of the primer may be stimulated byadditional base-pairing interactions between other parts of the tRNA molecule and viral sequences flanking thePBS. According to the RNA secondary structure model of the HIV-1 leader region, part of the PBS sequenceis involved in base pairing to form a small stem-loop structure, termed the U5-PBS hairpin. This hairpin maybe involved in the process of reverse transcription. To study the role of the U5-PBS hairpin in the viralreplication cycle, we introduced mutations in the U5 region that affect the stability of this structured RNAmotif. Stabilization and destabilization of the hairpin significantly inhibited virus replication. Upon prolongedculturing of the virus mutant with the stabilized hairpin, revertant viruses were obtained with additionalmutations that restore the thermodynamic stability of the U5-PBS hairpin. The thermodynamic stability of theU5-PBS hairpin apparently has to stay within narrow limits for efficient HIV-1 replication. Transient trans-fection experiments demonstrated that transcription of the proviral genomes, translation of the viral mRNAs,and assembly of the virions with a normal RNA content is not affected by the mutations within the U5-PBShairpin. We show that stabilization of the hairpin reduced the amount of tRNA primer that is annealed to thePBS. Destabilization of the hairpin did not affect tRNA annealing, but the viral RNA-tRNA complex was lessstable. These results suggest that the U5-PBS hairpin is involved in correct placement of the tRNA primer onthe viral genome. The analysis of virus mutants and revertants and the RNA structure probing experimentspresented in this study are consistent with the existence of the U5-PBS hairpin as predicted in the RNAsecondary structure model.
The replication cycle of human immunodeficiency virus type1 (HIV-1) and other retroviruses is characterized by reversetranscription of the viral RNA genome into a double-strandedDNA, which subsequently becomes integrated into the hostcell genome (42). This process is mediated by the virion-asso-ciated enzyme reverse transcriptase (RT), and the cellulartRNA3
Lys molecule is used as a primer by HIV-1 (35). ThetRNA primer binds with its 39-terminal 18 nucleotides (nt) toa complementary sequence in the viral genome, the primer-binding site (PBS), which is located in the untranslated leaderregion of the viral genome (Fig. 1A). Besides the complemen-tarity between the PBS and the 39 end of tRNA3
Lys, annealingof the primer has been proposed to be stimulated by additionalbase-pairing interactions between other parts of the tRNAmolecule and viral sequences flanking the PBS (31).
Extensive secondary structure in the 59 untranslated leaderregion of the HIV-1 genome has been suggested by electronmicroscopy, replication studies with mutant viruses, and bio-chemical RNase probing studies (3, 11, 17, 21, 22, 37). Theseresults, combined with phylogenetic analyses and computer-assisted structure prediction, led to a model of the secondaryRNA structure of the complete leader region of the HIV-1genome (4). According to this model, the PBS is flanked by an
upstream small stem-loop structure, the U5-PBS hairpin (Fig.1A). This HIV-1 hairpin structure was modeled primarilybased on the fact that phylogenetic analysis of different HIVand simian immunodeficiency viruses (SIV) demonstrated aconservation of the hairpin structure, despite considerable di-vergence in sequence (5, 7). A striking feature of the U5-PBShairpin of different HIV and SIV isolates is that part of thePBS sequence is involved in base pairing (Fig. 1B). SeveralRNA secondary structures in the leader RNA have been re-ported to regulate important viral replication steps of HIV-1;examples are transcriptional transactivation by Tat (8, 20, 30),mRNA polyadenylation (16, 28), and dimerization of the viralRNA genome (9, 12, 38). A stem-loop structure at a similarposition as the U5-PBS hairpin of HIV-1 was predicted forRous sarcoma virus. This structure is required for efficientinitiation of reverse transcription in Rous sarcoma virus (2,13). In addition, an interaction between U5 RNA and se-quences of the primer tRNA has been proposed (1) and wasconfirmed recently by RNA structure probing studies (37a). Adetailed structure has also been proposed for the HIV-1 RNA-tRNA3
Lys complex based on biochemical experiments (24, 25a).Several sequences in the U5 region upstream of the PBS weresuggested to interact with different parts of the tRNA3
Lys
primer. According to this model, base pairing occurs betweenthe U-rich anticodon loop of tRNA3
Lys and the A-rich loop ofthe U5-PBS hairpin. These combined observations suggest aspecific role for the U5-PBS hairpin structure in the process ofreverse transcription.
To study the role of the U5-PBS hairpin in the viral repli-
* Corresponding author. Mailing address: Department of HumanRetrovirology, Academical Medical Center, University of Amsterdam,P.O. Box 22700, 1100 DE Amsterdam, The Netherlands. Phone: 31-20-5664822. Fax: 31-20-6916531. E-mail: [email protected].
2227
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

cation cycle, we introduced mutations in this structured RNAmotif of the HIV-1 genome. Stabilization or destabilization ofthe U5-PBS hairpin significantly reduced virus replication.Analysis of revertant viruses, obtained through prolonged cul-turing of the mutant viruses, revealed that the thermodynamicstability of the hairpin has to stay within narrow limits forefficient HIV-1 replication. Biochemical assays demonstratedthe involvement of the U5-PBS hairpin in the correct place-ment of the tRNA3
Lys primer onto the viral genome.
MATERIALS AND METHODS
DNA constructs. A derivative of the full-length proviral HIV-1 clone pLAI wasused to produce wild-type and U5-mutated viruses. This construct, pLAI-R37,was described previously (17). The 39 long terminal repeat (39LTR) was trun-cated at the SacI site within the R region, and the chloramphenicol acetyltrans-ferase (cat) gene and simian virus 40 polyadenylation site were inserted at thisposition. Nucleotide numbers refer to positions on the genomic RNA transcript,with 11 being the capped G residue. For mutation of the U5-PBS hairpin, weused the construct pBlue-59LTR (29), which contains an XbaI-ClaI fragment ofHIV-1 encompassing the 59LTR, PBS, and 59 end of the gag gene (positions2454 to 1376) cloned into pBluescript (Stratagene). The U5-PBS hairpin se-quence was mutated by oligonucleotide-directed in vitro mutagenesis with aMuta-Gene phagemid in vitro mutagenesis kit (Bio-Rad). Oligonucleotides usedare Ts (59-AGACCCTTTTAGTCACTGCTGGAAAATCTCTAGC-39) and Td(59-CCTCAGACCCTTTTACAAAGTGTGGAAAATCTC-39 (mutagenic posi-
tions underlined). The mutations introduced were verified by sequence analysis.Sequencing was performed with the primer AD-SD (positions 1269 to 1290),using a Thermo Sequenase dye terminator cycle sequencing kit (Amersham) andan Applied Biosystems 373 DNA sequencer. Subsequently, the mutated XbaI-ClaI fragments were introduced into the proviral clone pLAI-R37, which againwas verified by sequence analysis. For transcription studies, the pBlue-39LTR-luciferase reporter construct was generated by the exchange of theHindIII-BamHI fragment of pBlue-39LTR-CAT (29), encompassing the catgene, by the HindIII-BamHI fragment of pGL3 (Promega), encoding the lucif-erase gene. For construction of the pBlue-59LTR-luciferase reporter construct,the 59LTR-leader region of HIV-1 was PCR amplified to introduce an NcoIrestriction site overlapping the gag translation start codon. The primers used areAD-R1 (positions 16 to 130) and SP6-ATG (59-ATTTAGGTGACACTATAGCCATGGCTCTCCTTCTAGCC-39 (start codon underlined, mutagenic posi-tions in bold). The PCR fragment was digested with HindIII/NcoI and insertedinto HindIII/NcoI-digested pBlue-39LTR-luciferase. The control luciferase con-struct was generated by deletion of the XhoI/NcoI fragment, encompassing allHIV-1 sequences, filling of the recessed termini by the Klenow fragment of DNApolymerase I, and self-ligation of the vector. The expression vector pcDNA3-Tatwas described previously (44).
Synthesis of RNA templates. Plasmid pBlue-59LTR was used as a template forPCR amplification and subsequent in vitro transcription. The 59LTR region ofHIV-1 was PCR amplified with the sense primer T7-2 (positions 11 to 120)containing the T7 RNA polymerase promoter sequence and the antisense primerAUG (positions 1348 to 1368). The PCR fragments were phenol extracted,precipitated, and dissolved in water. The in vitro transcription reaction wasperformed in 10 ml of transcription buffer (40 mM Tris [pH 7.5], 2 mM spermi-dine, 10 mM dithiothreitol [DTT], 12 mM MgCl2) containing 0.5 mg of DNAtemplate, 0.06 mmol of ATP, GTP, CTP, and UTP, 10 U of T7 RNA polymerase(Boehringer), and 20 U of RNase inhibitor (Boehringer) and incubated for 4 hat 37°C. Upon DNase treatment and phenol extraction, the unincorporated freenucleotides were removed by passage through a Sephadex G-50 column. Subse-quently, the RNA was ethanol precipitated and dissolved in renaturation buffer(10 mM Tris-HCl [pH 7.5], 100 mM NaCl). The RNA was renatured by incu-bation at 85°C for 2 min, followed by slow cooling to room temperature.
RNA structure probing. The renatured RNA (25 ng) was treated with 0.5%diethyl pyrocarbonate (DEPC) or 0.1% dimethyl sulfate (DMS) in 25 ml of 10mM Tris (pH 7.5)–10 mM MgCl2–50 mM NaCl buffer. After incubation for 10min at 37°C, the RNA sample was recovered by ethanol precipitation anddissolved in 5 ml of renaturation buffer. The antisense primer BB-3 (positions1216 to 1245) was used to map the modified RNA positions in a primerextension reaction. This primer was end labeled with [g-32P]ATP and T4 polynu-cleotide kinase (Boehringer). The labeled oligonucleotide (2 ng) was mixed withthe RNA sample in a total volume of 10 ml of annealing buffer (83 mM Tris-HCl[pH 7.5], 125 mM KCl), incubated for 2 min at 85°C and for 10 min 65°C, andslowly cooled to 25°C. The primer was extended by addition of 5 ml of RT buffer(9 mM MgCl2, 30 mM DTT, 150 mg of actinomycin D per ml, 30 mM dATP,dGTP, dTTP, and dCTP) and 12.5 U of avian myeloblastosis virus (AMV) RT(Boehringer) in an incubation at 42°C for 15 min. The samples (2.5 ml) weremixed with formamide loading buffer (2.5 ml), denatured at 90°C, and analyzedon a 6% polyacrylamide–7 M urea gel.
Cells, viruses, and transfection. SupT1 T cells were grown in RPMI 1640medium supplemented with 10% fetal calf serum at 37°C and 5% CO2. SupT1cells (5 3 106) were transfected with 1 and 2 mg of the HIV-1 proviral constructsby electroporation (250 V, 960 mF). After transfection, 0.5 3 106 fresh SupT1cells were added to support viral replication. Cells were split 1 to 10 twice a week.For the selection of revertant viruses, the transfected cells were passaged up to124 days. At the peak of virus production, 100 to 0.1 ml of the culture supernatantwas used to infect fresh SupT1 cells. At each passage, cells and supernatantsamples were stored at 270°C. For transcription studies, 5 3 106 SupT1 cellswere transfected with 5 mg of the 59LTR-luciferase constructs by electroporation.We added 100, 500, and 1,000 ng of pcDNA-Tat, which is within the linear rangeof LTR transcriptional activation. To have an equal amount of 6 mg DNA in eachtransfection, we added the empty pcDNA3 vector.
C33A and HeLa cells were grown in Dulbecco’s modified Eagle’s mediumcontaining 10% fetal calf serum at 37°C and 5% CO2. For the transient produc-tion of virions, C33A cells were transfected by the calcium phosphate method.Cells were grown in 20 ml of culture medium in a 75-cm2 flask to 60% conflu-ency. Thirty micrograms of the proviral construct in 880 ml of water was mixedwith 1 ml of 50 mM HEPES (pH 7.1)–250 mM NaCl–1.5 mM Na2HPO4 and 120ml of 2 M CaCl2, incubated at room temperature for 20 min, and added to theculture medium. The culture medium was changed after 16 h. For transcriptionstudies, HeLa cells were transfected by the DEAE-dextran method. Cells weregrown in a 60- by 15-mm tissue culture dish to 60% confluency. Cells werewashed two times with Tris-buffered saline (TBS) and incubated for 30 min atroom temperature with the DEAE-dextran-DNA mixture, containing 1 mg of59LTR-luciferase construct with or without 1, 10, and 100 ng of the pcDNA3-Tatexpression vector, which is within the linear range of LTR transcriptional acti-vation, in 475 ml of TBS and 25 ml of DEAE-dextran (10 mg/ml in TBS). Finally,the cells were washed two times with TBS to remove the DEAE-dextran-DNAmixture, and culture medium was added.
FIG. 1. Annealing of the tRNA3Lys primer to the PBS of the HIV-1 RNA
genome. (A) The tRNA3Lys primer binds with its 39 terminus to the complemen-
tary sequence of the PBS to form an 18-bp duplex that is shown in detail (PBSsequence is marked in grey). The remainder of the tRNA cloverleaf structure isshown (AC, anticodon loop; D, D loop). Besides the base-pairing interactionwith the PBS, sequences in the U5 region may interact with different parts oftRNA3
Lys to stimulate primer annealing. Directly upstream of the PBS is a smallhairpin structure, the U5-PBS hairpin, which is the topic of this study. (B) Shownis the wild-type U5-PBS hairpin, which was mutated to change the thermody-namic stability. In mutant Ts, the hairpin was stabilized by the introduction of anadditional C nucleotide at position 165 and one nucleotide change at position162 (G to C). In mutant Td, the hairpin is destabilized by three nucleotidesubstitutions at positions 158 to 160. The introduced mutations are marked byopen boxes, and the PBS sequence is marked by a grey box. The thermodynamicstability of the hairpins, indicated at the bottom (DG in kilocalories per mole),was calculated using the Zuker algorithm (53).
2228 BEERENS ET AL. J. VIROL.
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Analysis of phenotypic revertants. SupT1 cells transfected with the Ts proviralconstruct were pelleted by centrifugation at 4,000 rpm for 4 min and washed withphosphate-buffered saline (PBS). The cells were resuspended in 10 mM Tris-HCl(pH 8.0)–1 mM EDTA–0.5% Tween 20 and incubated with 200 mg of proteinaseK per ml at 56°C for 1 h and at 95°C for 10 min to isolate total cellular DNA. The59LTR-leader region was PCR amplified from the total cellular DNA with the 59U3 region primer 59X (positions 2454 to 2434) and the 39 gag primer AD-gag(positions 1442 to 1463). To provide this fragment with a T7 tail for sequencingwith the universal T7 dye primer, a second PCR was performed with the 59 Rregion primer T7-1 (positions 254 to 234) and the 39 primer AUG (positions1123 to 1151, with six additional nucleotides at its 59 end). These PCR productswere sequenced directly with a DYEnamic Direct cycle sequencing kit (Amer-sham) and an Applied Biosystems 373 DNA sequencer. In addition, the 59X/AD-gag PCR product was cloned into pBlue-59LTR as a HindIII/NarI fragment.For analysis of individual clones, a PCR was performed with the primers T7-1and AD-SD (positions 1269 to 1290), and this PCR fragment was subsequentlysequenced. Finally, for insertion of the revertant sequences into the proviralplasmid pLAI-R37, XbaI/ClaI fragments of the specific clones were used toreplace the corresponding wild-type sequences. Introduction of the revertantsequences into the proviral plasmid pLAI-R37 was verified by sequence analysis.Therefore, a PCR was performed with the T7-1 and AD-SD primers and thePCR fragment was sequenced.
CA-p24 and RT assay. CA (capsid protein)-p24 levels in the culture mediumwere determined by enzyme-linked immunosorbent assay. RT assays were per-formed as described previously (49). The virus sample (10 ml) was added to 50 mlof RT buffer (60 mM Tris-HCl [pH 8.0], 1 mM EDTA, 75 mM KCl, 5 mMMgCl2, 0.1% Nonidet P-40, 4 mM DTT) supplemented with 0.25 mg of poly(A)and 8 ng of oligo(dT)18 primer and 2.5 mCi (3,000 Ci/mmol) of [a32P]dTTP.Samples (10 ml) were taken after 1, 2, and 3 h of incubation at 37°C and spottedonto DE-81 paper. The samples were dried for 5 min; the paper was subse-quently washed three times in 5% Na2HPO4, washed two times in ethanol, andair dried. RT activity was quantified on a Molecular Dynamics PhosphorImager.
Luciferase assay. Luciferase assays were performed according to Promega’sluciferase assay system protocol. Two days posttransfection, HeLa cells werewashed with PBS and lysed in 200 ml of reporter lysis buffer (Promega). SupT1cells were collected by centrifugation 3 days after transfection, washed with PBS,and lysed in 200 ml of reporter lysis buffer. Luciferase activity in the samples (50ml) was determined by addition of luciferase assay reagent (Promega) in aBerthold model LB 9501 luminometer.
Isolation of viral RNA. Three days after transfection of C33A cells, the culturemedium (20 ml) was centrifuged at 1,600 rpm for 15 min to remove cells.Subsequently the supernatant was filtered through a 0.45-mm-pore-size filter(Schleicher & Schuell), and the virions were pelleted by centrifugation at 25,000rpm for 30 min in a Beckman SW28 rotor. Virions were resuspended in 500 mlof 10 mM Tris-HCl (pH 8.0)–100 mM NaCl–1 mM EDTA. To isolate viral RNA,the viruses were incubated for 30 min at 37°C in the presence of 100 mg ofproteinase K per ml and 0.5% sodium dodecyl sulfate, followed by extractionwith phenol-chloroform-isoamyl alcohol (25:24:1) and precipitation in 0.3 Msodium acetate (pH 5.2) and ethanol at 220°C. The viral RNA was pelleted bycentrifugation (18,000 rpm, 20 min), washed with 70% ethanol, and dried. Thepellet was dissolved in 20 ml of 10 mM Tris-HCl (pH 8.0)–1 mM EDTA andstored at 270°C.
Oligonucleotide and tRNA primer extension assays. In the oligonucleotideand tRNA primer extension assays, viral RNA corresponding to 30 ng CA-p24was incubated with 20 ng of oligonucleotide primer in 12 ml of 83 mM Tris-HCl(pH 7.5)–125 mM KCl at 85°C for 2 min and 65°C for 10 min, followed by coolingto room temperature in 1 h to allow annealing of the primer. The primer wasextended by addition of 6 ml of RT buffer (9 mM MgCl2, 30 mM DTT, 150 mgof actinomycin D per ml, 30 mM dATP, dGTP, and dTTP, 1.5 mM dCTP), 0.5 mlof [a32P]dCTP, and either 0.5 U of HIV-1 RT (U.S. Biochemical) or 12.5 U ofAMV RT (Boehringer) and incubation at 42°C for 3 min; then 10 mM (each)deoxynucleoside triphosphate (dNTP) was added, and incubation was continuedfor 30 min. The cDNA product was precipitated in 25 mM EDTA–0.3 M sodiumacetate (pH 5.2)–70% ethanol at 220°C. For degradation of the tRNA part ofthe extended product in the tRNA primer extension assay, the samples wereincubated with 0.5 N NaOH for 20 min at 55°C, neutralized with 0.5 M HCl, andprecipitated as described above. The products were analyzed on a denaturing 6%polyacrylamide-urea sequencing gel. The antisense primers used are CN1 (po-sitions 1123 to 1151) and AUG (positions 1348 to 1368, with six additionalnucleotides at its 59 end).
RESULTS
Design of the U5-PBS hairpin mutants. To study the role inHIV-1 replication of the U5-PBS hairpin that is located di-rectly upstream of the PBS, we introduced mutations in thisstem-loop structure. The U5 region is encoded by the LTR thatis present at both the 59 and 39 ends of the HIV-1 proviralgenome. Mutations introduced into the U5 region of the59LTR will be inherited in both LTRs of the progeny. How-
ever, the presence of a wild-type 39LTR may result in reversionof the mutant virus to the wild-type sequence by recombinationwith the wild-type 39LTR sequences. For production of themutant viruses, we therefore used a derivative of the proviralclone pLAI in which part of the 39LTR, including the polyad-enylation signal and the complete U5 region, is deleted. AnSV40 polyadenylation site was placed downstream of theHIV-1 sequences to allow efficient polyadenylation of the viraltranscript. Transfection of the SupT1 T-cell line with this vec-tor results in the production of viruses with a mutant U5 regionin the untranslated leader RNA. Subsequent infection ofSupT1 cells by these viruses, followed by reverse transcriptionof the viral RNA genome, will produce proviral genomes witha complete 59LTR and 39LTR that both have the mutated U5sequence.
Mutations that affect the stability of the U5-PBS hairpinwere introduced in the U5 region (Fig. 1B). The mutants werecarefully designed not to affect important sequence motifs,such as the attachment site for integration (positions 170 to181) (18, 36) and the PBS sequence. All mutations were there-fore introduced on the left side of the hairpin. We stabilizedthe hairpin in mutant Ts by generating two extra C-G basepairs. This was done by replacement of the unpaired G162 byC and by insertion of an additional C at position 165 (Fig. 1B).This results in an increase in the thermodynamic stability of thehairpin (DG) from 25.4 kcal/mol for the wild type to 218.2kcal/mol for mutant Ts. In mutant Td, the hairpin was desta-bilized by substitution of three nucleotides at position 158 to160. As a result, base pairing in the lower part of the stem islost, and a relative short and instable hairpin structure is left(DG 5 21.6 kcal/mol).
Structure probing of the wild-type and mutant U5-PBS hair-pins. To demonstrate that the introduced mutations have noeffect on folding in other parts of the leader RNA, we per-formed structure probing experiments. In vitro-synthesizedHIV-1 leader RNA was treated with structure-specific probes,followed by primer extension analysis to localize the sites ofmodification. Nucleotides sensitive to DMS or DEPC are as-sumed not to be involved in base-pairing or base-stacking in-teractions. The sites of modification were identified by primerextension analysis using the DNA primer BB-3 (positions1216 to 1245). No striking differences in reactivity toward thechemicals was observed between the wild-type and mutantU5-PBS hairpins (Fig. 2). The A-rich loop of the wild-typeU5-PBS hairpin as well as that of the two mutants is modifiedby both DMS and DEPC, whereas the flanking sequences arenot. The A-rich loop is even visible in the Td mutant, in whichthe U5-PBS hairpin was destabilized but apparently not de-stroyed. No major differences were observed in the upstreamleader region (Fig. 2) and the region downstream of the PBS,which was also probed and analyzed with the DNA primersDIS (positions 1246 to 1269) and AUG (positions 1348 to1368) (results not shown). Furthermore, computer modelingof a larger region of the 59LTR leader RNA (positions 1111 to1244) suggests that the introduced mutations do not affect theRNA secondary structure in this part of the genome. Thesecombined results indicate that the mutations do not lead to anoverall structural rearrangement of the HIV-1 leader.
Replication capacity of viruses with a mutated U5-PBS hair-pin. To study the replication potential of the mutant viruses,we transfected wild-type and mutant proviral genomes intoSupT1 cells. These cells express the CD4-CXCR4 receptorsand are fully susceptible for replication of the LAI strain. Virusproduction was followed by measuring CA-p24 levels in theculture medium at several days after transfection. Transfectionwith 2 mg of the proviral constructs showed that the replication
VOL. 74, 2000 ROLE OF U5-PBS IN HIV-1 REPLICATION 2229
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

capacity of both mutants was reduced compared with the wild-type virus (Fig. 3A). This defect is even more pronounced intransfections with 1 mg of proviral construct (Fig. 3B). Noreplication of mutant Ts was observed in transfections with lessthan 1 mg of the proviral construct. Thus, stabilization of theU5-PBS hairpin affected the replication potential of the virusmore severely than destabilization of this RNA structure.These results demonstrate the importance of the U5-PBS hair-pin in viral replication.
Reversion of the stabilized U5-PBS hairpin mutant. Duringprolonged culturing of replication-defective viruses, pheno-typic revertants with an increased replication capacity canarise. The genomes of such revertant viruses should be alteredin order to replicate more efficiently, and analysis of suchrevertant genomes may allow the identification of importantRNA sequences and/or structures. This forced evolution ap-proach can be used for most retroviruses due to their highmutation rate and has proven to be valuable in the analysis ofregulatory RNA motifs (6, 15, 30). Mutant Td replicated tooefficiently to allow the selection of faster-replicating revertantswithin a reasonable time span. We therefore focused on theevolution of the severely defective mutant Ts.
SupT1 cells transfected with the Ts proviral construct weresplit into several independent cultures that were maintainedfor 7 weeks. The replication kinetics of the viruses present inseveral cultures increased after a variable time. To determinethe sequence of the U5-PBS hairpin of these phenotypic re-vertants, total cellular DNA was isolated from infected cells.The 59LTR-leader region was PCR amplified, and we per-
formed population-based sequencing of the DNA fragment.The predicted RNA structures for the revertant sequences areshown in Fig. 4, with the thermodynamic stability indicatedbelow the hairpins. Remarkably, the nucleotide changes intro-duced in mutant Ts were frequently found to be altered in therevertant genomes, although no true wild-type reversions wereobserved. All acquired mutations are located on the left side ofthe hairpin, which is consistent with the presence of importantsequence motifs on the right side. Analysis of the revertantsdemonstrated a variation in repair strategies, but all mutationsreduced the stability of the hairpin. This is most striking inrevertant w6, with two reversion-based mutations that result ina hairpin with a stability very similar to that of the wild-typeU5-PBS structure. It is likely that the other revertants, all withonly one nucleotide substitution within the hairpin, had not yetattained the optimal configuration and will evolve toward hair-pin structures with wild-type stability over time.
To study the evolution of the stabilized Ts hairpin in moredetail, we performed an independent SupT1 transfection andmonitored this culture for up to 124 days. Total cellular DNAwas isolated from infected cells at several days posttransfec-tion. Figure 5A shows the results of direct sequence analysis ofthe PCR-amplified U5-PBS region of the revertants. Substitu-tions were initially observed at the two mutated residues inmutant Ts, but two additional mutations were acquired overtime. This so-called population sequencing provides informa-tion on the acquired mutations and their relative frequency in
FIG. 2. RNA structure probing of the U5-PBS region under native condi-tions. In vitro-transcribed HIV-1 leader RNA of the wild-type and U5-PBSmutants was treated with a limiting amount of the single-strand-specific reagentsDEPC (A specific) and DMS (A/C specific) as indicated. Modification sites weredetected using primer extension analysis with the BB-3 primer. The productswere analyzed on a 6% polyacrylamide–7M urea gel. For reference, the BB-3primer was used in a DNA sequencing reaction (lanes 4 to 7 and 11 to 14).Positions of the hairpin structures in this part of the HIV-1 leader RNA and thePBS sequence are shown schematically on the left. FIG. 3. Replication of wild-type (wt) and U5-PBS hairpin mutants Ts and
Td. SupT1 cells were transfected with 2 (A) or 1 (B) mg of the proviral constructs.Several days after transfection, CA-p24 production was measured in the culturemedium.
2230 BEERENS ET AL. J. VIROL.
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

the virus quasispecies population. However, this method doesnot determine genetic linkages if mixed sequences are presentat multiple positions (e.g., the day 27 sample). We thereforealso performed clonal sequencing. To do so, the PCR fragmentwas cloned into pBlue-59LTR, and multiple individual cloneswere sequenced for the samples obtained at day 27, 86, and 124(Fig. 5B). In the initial phase of the evolution experiment, two
revertants appeared to be present simultaneously. Both evolu-tion routes target one of the nucleotides introduced in mutantTs. The evolutionary pathways of Ts reversion are depicted inFig. 6, which shows the predicted RNA structure and thermo-dynamic stability of the observed revertants. Alteration ofC165 to A produces an A/G mismatch and destabilizes thehairpin from 218.2 to 211.1 kcal/mol. The other revertant
FIG. 4. Phenotypic revertants of the stabilized U5-PBS hairpin mutant Ts. Through prolonged culturing of the mutant virus Ts, several revertant viruses withnucleotide changes in the hairpin were obtained. The predicted RNA structures of these revertants and DG values (in kilocalories per mole) are shown. The mutationspresent in mutant Ts are marked by an open box, the acquired additional mutations are marked by black boxes, and the PBS is marked by a grey box. All reversion-basedmutations reduce the stability of the mutant U5-PBS hairpin.
FIG. 5. Sequence analysis of Ts revertant genomes. (A) The sequence of the U5-PBS hairpin was determined by population sequencing several days aftertransfection of SupT1 cells with the Ts proviral construct. Positions of the hairpin motif and the PBS are indicated at the top. Dashes indicate nucleotides that areidentical to that of the input Ts mutant. Acquired mutations are indicated in capitals if the majority of genomes carried the mutation; minor changes are in smallcharacters. (B) The genetic linkage of the various acquired mutations was determined by sequencing of multiple individual clones for the samples taken at days 27, 86,and 124 posttransfection. The frequency of each sequence is given in parentheses. The clones tested in the replication studies are marked by asterisks.
VOL. 74, 2000 ROLE OF U5-PBS IN HIV-1 REPLICATION 2231
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

changes C162 to U, thereby creating a weak U-G base pair thathas a moderate effect on stability (DG 5 15.6 kcal/mol). Al-though both genotypes are present at an approximatelyequimolar concentration at day 27 (Fig. 5A), the former seemsto outcompete the latter, as is evident from the populationsequence at days 36 and 41. However, the latter genotypereappears at day 65 due to the acquisition of another destabi-lizing mutation (G168 to A) that triggers a rearrangement ofthe upper part of the stem region and increases the loop size.This hairpin (DG 5 211.1 kcal/mol) acquires one more sub-stitution at day 124 that further reduces the hairpin stability(DG 5 27.3 kcal/mol) to a value that is similar to that of thewild-type structure (DG 5 25.4 kcal/mol).
The role of the acquired U5 mutations in the phenotypicreversion of mutant Ts was demonstrated by introduction ofrevertant sequences observed at days 27, 86, and 124 in thewild-type proviral genome for replication studies. Consecutiveintermediates in the evolutionary pathway showed graduallyimproved replication (Fig. 7). This finding demonstrates thatreplication of the mutant Ts is repaired by restoration of thehairpin stability. The combined results of the evolution studiesindicate that a U5-PBS hairpin of approximately wild-typestability is optimal for virus replication.
The U5-PBS hairpin is not involved in gene expression andvirus production. Binding sites for AP-1 and NF-AT/AP3 tran-scription factors were recently reported to be positioned on theproviral DNA genome directly upstream of the PBS (43).These transcription factor binding sites have been suggested tobe involved in HIV-1 transcription and replication. The up-stream AP-1 site is changed in mutant Td, and both binding
sites are affected in mutant Ts (Fig. 8A), raising the possibilitythat viral transcription is affected in these mutants. Further-more, the revertants obtained by prolonged culturing of mu-tant Ts possibly restore the binding of these transcription fac-tors and thereby virus replication. To test whether the U5mutations affect viral transcription, we transfected C33A cells
FIG. 6. Evolutionary pathway of the stabilized U5-PBS hairpin mutant Ts. The predicted structure for the sequences observed at several days posttransfection (Fig.5) is shown. The period in which the intermediates were observed is indicated at the top, and the calculated thermodynamic stability (in kilocalories per mole) isindicated below the hairpins. The introduced mutations in mutant Ts are marked by an open box, the acquired mutations are marked by black boxes, and the PBS ismarked by a grey box. The hairpin of mutant Ts acquires several mutations that reduce the thermodynamic stability and finally attains a stability similar to that of thewild-type U5-PBS hairpin.
FIG. 7. Replication of different intermediates in the reversion of mutant Ts.Revertant genomes observed at days 27, 86, and 124 (marked by an asterisk inFig. 5B) were introduced in the wild-type (wt) proviral genome. SupT1 cells weretransfected with 2 mg of the proviral constructs. Several days after transfection,CA-p24 production was measured in the culture medium.
2232 BEERENS ET AL. J. VIROL.
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

(human cervix carcinoma cells not expressing CD4) with thewild-type and mutant Ts and Td proviral vectors and analyzedthe level of viral gene expression and virion production. Virusproduction was monitored by measuring the amount of CA-p24 and virion-associated RT activity in the culture medium.Table 1 summarizes the results of two independent transfec-tions. No significant differences were measured, as the ob-served differences reflect experimental variation in the electro-poration protocol. The expression levels of viral proteins werealso found to be similar for all constructs by Western blotanalysis of total cell extracts (results not shown).
To study the transcriptional activity of the mutants Td andTs as well as two Ts revertants in more detail, we constructedLTR-reporter vectors with the complete LTR-leader region ofHIV-1 fused to the luciferase open reading frame. TransientLTR-luciferase transfection assays and cotransfections with aTat expression vector were performed in HeLa and SupT1cells. Transfection of episomal plasmids may not accuratelyreflect proviral transcription from an integrated position, butthe results shown in Fig. 8B to D indicate that the wild-type,mutant, and revertant LTR constructs do not differ signifi-cantly in basal or Tat-activated transcriptional activity. Thesecombined results suggest that the mutations in the U5 regiondo not affect viral gene expression (e.g., transcription andtranslation) and virion assembly.
The U5-PBS hairpin is not involved in packaging of the viralRNA. The untranslated leader RNA contains important signalsfor packaging of the viral RNA into virion particles. To deter-mine the RNA content of the wild-type and mutant viruses, weisolated RNA from purified virions that were produced inC33A cells. The viral RNA was measured by primer extensionanalysis with the CN1 oligonucleotide primer, which is com-plementary to the 1123 to 1151 region. The CA-p24 valueswere used to control for the amount of virions used per sample.The mutant virions contained a normal level of RT enzyme(Table 1). As summarized in Table 2, no significant differenceswere observed between the mutant and wild-type viruses inregard to the amount of viral RNA per virion. These resultsshow that the U5-PBS hairpin does not contribute to the pro-cess of packaging of genomic HIV-1 RNA.
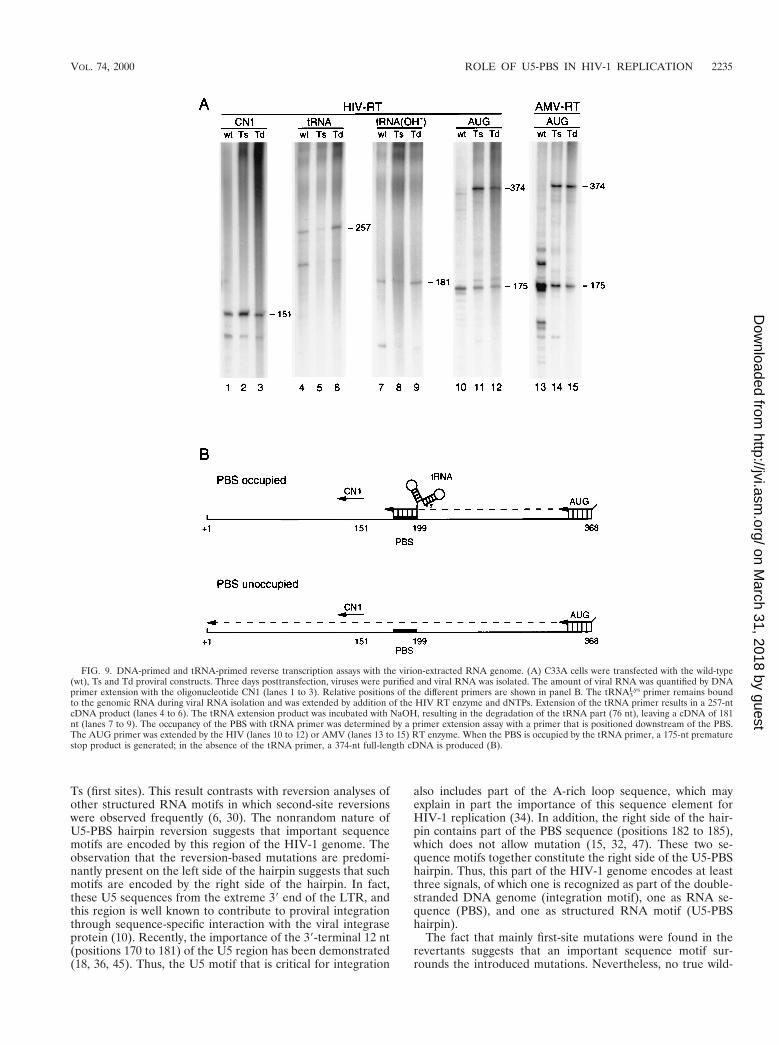
The U5-PBS hairpin is involved in reverse transcription.The U5-PBS structure of HIV-1 may be involved in regulationof reverse transcription, in particular because the hairpin in-cludes part of the PBS sequence. In addition, a similar hairpinstructure in avian sarcoma-leukosis virus has been found to beinvolved in the process of reverse transcription, and an inter-action between the anticodon of the tRNA primer and theA-rich hairpin loop was proposed for HIV-1 to play a role inthe process of reverse transcription. We therefore analyzed theamount of tRNA3
Lys primer annealed to the PBS of the wild-type and mutant virion particles. During the isolation of viralRNA, the tRNA primer remains bound to the PBS and can be
FIG. 8. Transcriptional activity of wild-type and mutant LTR promoters. (A)The Ts and Td mutations affect transcription factor binding sites in the U5 DNA.Marked are the AP-1 and NF-AT/AP-3 transcription factor binding sites locatedin the U5 region immediately upstream of the PBS (43). The nucleotide changesin mutant Td, mutant Ts, and two Ts revertants (rev), w3 and w3.1 (clonesobtained from revertant w3), are shown. (B) Relative basal transcriptional ac-tivity of wild-type (wt) and mutant LTRs in HeLa and SupT1 cells after trans-fection with 1 and 5 mg, respectively, of the different LTR-luciferase constructs.
The basal transcriptional activity of the wild-type LTR was set at 1. (C and D)Relative Tat-activated transcriptional activities of wild-type and mutant LTRs inHeLa (C) and SupT1 (D) cells. HeLa cells were transfected with 1 mg of theLTR-luciferase constructs with or without 1, 10, and 100 ng of Tat expressionvector. SupT1 cells were transfected with 5 mg of the LTR-luciferase constructswith or without 100, 500, and 1,000 ng of Tat expression vector. The amounts ofadded Tat expression vector are within the linear range of LTR transcriptionalactivation. The basal transcriptional activity of each individual LTR promoterwas set at 1. Transfection with the control luciferase construct, containing theluciferase gene but not the LTR, and cotransfection of this construct withpcDNA3-Tat as well as transfection with the empty Tat vector (pcDNA3) re-vealed no luciferase activity. Cotransfection of the LTR-luciferase constructswith pcDNA3 resulted in a low basal level of LTR transcription.
VOL. 74, 2000 ROLE OF U5-PBS IN HIV-1 REPLICATION 2233
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

visualized by extension upon addition of HIV-1 RT enzymeand dNTPs. Extension of the tRNA primer produces a 257-nt-long tRNA-cDNA product (Fig. 1A shows a schematic; Fig.9A, lanes 4 to 6). The identity of this product was confirmed byNaOH-mediated degradation of the tRNA part, leaving a181-nt cDNA product (Fig. 9A, lanes 7 to 9). The extendedtRNA-cDNA products were quantified and corrected for theamount of input viral RNA template as determined by CN1primer extension (Fig. 9A, lanes 1 to 3). As summarized inTable 3, the tRNA extension efficiency of mutant Ts was re-duced to 27% of the value measured for the wild-type tem-plate. The mutant Td was not affected in tRNA extension; weconsistently measured a small improvement compared with thewild-type (125%).
The reduced tRNA extension efficiency of mutant Ts may bethe result of less tRNA primer that is annealed to the PBS.Alternatively, normal levels of tRNA are bound, but theseprimers cannot be extended efficiently on the template with thestabilized U5-PBS hairpin. To discriminate between these twopossibilities, the tRNA occupancy of the PBS was determinedby a different assay. Viral RNA-tRNA complexes were used asa template for the extension of a primer that is positioneddownstream of the PBS. The oligonucleotide primer AUG,complementary to the 1348 to 1368 region with six additionalnucleotides at its 59 end, was used in this experiment. ViralRNA without a tRNA primer will produce a full-length cDNAproduct of 374 nt (Fig. 9B). When a tRNA primer is present onthe PBS, this primer will also be extended by the RT enzyme,and RNase H will subsequently degrade the RNA templatestrand. The AUG primer can be extended to the 39 end of thePBS, where it encounters the annealed tRNA, which will resultin a 175-nt cDNA product. Alternatively, when the tRNA isdisplaced by the RT enzyme, the AUG primer can be extendedto the 59 end of the PBS, producing a 193-nt cDNA product. Asshown in Fig. 9A (lanes 10 to 12), extension of the downstreamAUG primer produced predominantly the 175-nt stop product,indicating that the tRNA primer is not displaced by the elon-gating RT enzyme. A different result was reported for theavian leukosis virus, where the tRNA primer was efficientlydisplaced (48). Extension of the AUG primer on the wild-typetemplate (Fig. 9, lane 10) produced predominantly the 175-ntstop product and almost no 374-nt full-length product. Quan-titation of the stop and full-length cDNA products indicated
that approximately 90% of the wild-type template has bound atRNA primer (Table 3), suggesting that nearly all PBS sites ofthe wild-type HIV-1 RNA are occupied. In contrast, extensionof the AUG primer on the mutant Ts template producedrelatively less premature stop product (23%) and more full-length product (77%). This result demonstrates that the PBSof mutant Ts is only partially occupied by a tRNA. In fact, the23% PBS occupancy measured with this assay correlates wellwith the 27% tRNA extension efficiency (Table 3).
Surprisingly, the PBS occupancy of mutant Td was stronglyreduced (38%), even though this mutant showed no defect inthe assay (125%) (Fig. 9A; Table 3). Apparently, the tRNAprimer is present on the PBS and can be extended efficiently,but the tRNA is lost during the PBS occupancy assay with thedownstream AUG primer. The tRNA primer is probably re-leased during the heat denaturation step that is used to annealthe AUG primer. The results suggest that the PBS-associatedtRNA primer does not optimally interact with the Td template.In fact, this means that the PBS occupancy assay is not areliable method when RNA templates that differentially bindthe tRNA primer are compared. Similar results were obtainedin PBS occupancy assays performed with the RT enzyme ofAMV (Fig. 9A, lanes 13 to 15). However, upon extension ofthe tRNA primer, we observed several additional stop prod-ucts, specific for the wild-type template, at positions both up-stream and downstream of the PBS. This implies a conforma-tional difference between the viral RNA-tRNA duplex formedwith the wild-type and mutant templates. Annealing of thetRNA primer onto the wild-type genome is evidently morecomplex and more stable than interaction with the genome ofmutant Td. These results indicate that the U5-PBS hairpin isinvolved in the correct placement of the tRNA primer onto theviral RNA.
DISCUSSION
In this study, we demonstrate the importance of the U5-PBShairpin structure for efficient HIV-1 replication. Both stabili-zation and destabilization of this RNA structure decreased theviral replication capacity. Upon prolonged culturing of thestabilized mutant Ts, several revertant viruses were obtainedwith an increased replication potential. All of the phenotypicrevertants acquired additional mutations in the hairpin thatreduce its thermodynamic stability. Thus, the mutant virusesrevert by emulating the stability of the wild-type hairpin. Thisindicates that RNA structural effects rather than RNA orDNA sequence effects are responsible for the replication de-fect of these HIV-1 mutants. Apparently, the thermodynamicstability of the U5-PBS hairpin must stay within narrow limitsfor efficient HIV-1 replication.
Analysis of revertant viruses of the mutant Ts also revealedthat the reversion-based mutations are almost exclusivelypresent on the left side of the U5-PBS hairpin, and in partic-ular at the nucleotide positions that were altered in the mutant
TABLE 1. Virus production upon transfection of wild-type and mutant proviral constructs into C33A cells
Construct
Expt 1 Expt 2
CA-p24(ng/ml)
RT activity(counts) RT/CA-p24 CA-p24
(ng/ml)RT activity
(counts) RT/CA-p24
Wild type 180 39,201,163 1.0a 550 17,543,880 1.0a
Ts 310 66,584,699 1.0 300 11,657,850 1.2Td 410 98,476,183 0.9 380 11,970,842 1.0
a Set at 1.
TABLE 2. Genomic RNA content of wild-type and mutant virionsas determined by primer extension
Construct CA-p24(ng/ml)
Viral RNA(counts) RNA/CA-p24
Wild type 1,920 92,238 100a
Ts 2,000 92,498 100Td 4,200 198,688 98
a Set at 100%.
2234 BEERENS ET AL. J. VIROL.
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
Ts (first sites). This result contrasts with reversion analyses ofother structured RNA motifs in which second-site reversionswere observed frequently (6, 30). The nonrandom nature ofU5-PBS hairpin reversion suggests that important sequencemotifs are encoded by this region of the HIV-1 genome. Theobservation that the reversion-based mutations are predomi-nantly present on the left side of the hairpin suggests that suchmotifs are encoded by the right side of the hairpin. In fact,these U5 sequences from the extreme 39 end of the LTR, andthis region is well known to contribute to proviral integrationthrough sequence-specific interaction with the viral integraseprotein (10). Recently, the importance of the 39-terminal 12 nt(positions 170 to 181) of the U5 region has been demonstrated(18, 36, 45). Thus, the U5 motif that is critical for integration
also includes part of the A-rich loop sequence, which mayexplain in part the importance of this sequence element forHIV-1 replication (34). In addition, the right side of the hair-pin contains part of the PBS sequence (positions 182 to 185),which does not allow mutation (15, 32, 47). These two se-quence motifs together constitute the right side of the U5-PBShairpin. Thus, this part of the HIV-1 genome encodes at leastthree signals, of which one is recognized as part of the double-stranded DNA genome (integration motif), one as RNA se-quence (PBS), and one as structured RNA motif (U5-PBShairpin).
The fact that mainly first-site mutations were found in therevertants suggests that an important sequence motif sur-rounds the introduced mutations. Nevertheless, no true wild-
FIG. 9. DNA-primed and tRNA-primed reverse transcription assays with the virion-extracted RNA genome. (A) C33A cells were transfected with the wild-type(wt), Ts and Td proviral constructs. Three days posttransfection, viruses were purified and viral RNA was isolated. The amount of viral RNA was quantified by DNAprimer extension with the oligonucleotide CN1 (lanes 1 to 3). Relative positions of the different primers are shown in panel B. The tRNA3
Lys primer remains boundto the genomic RNA during viral RNA isolation and was extended by addition of the HIV RT enzyme and dNTPs. Extension of the tRNA primer results in a 257-ntcDNA product (lanes 4 to 6). The tRNA extension product was incubated with NaOH, resulting in the degradation of the tRNA part (76 nt), leaving a cDNA of 181nt (lanes 7 to 9). The occupancy of the PBS with tRNA primer was determined by a primer extension assay with a primer that is positioned downstream of the PBS.The AUG primer was extended by the HIV (lanes 10 to 12) or AMV (lanes 13 to 15) RT enzyme. When the PBS is occupied by the tRNA primer, a 175-nt prematurestop product is generated; in the absence of the tRNA primer, a 374-nt full-length cDNA is produced (B).
VOL. 74, 2000 ROLE OF U5-PBS IN HIV-1 REPLICATION 2235
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

type reversions were observed. AP-1 and NF-AT/AP3 tran-scription factor binding sites have been found in the U5 region(43). The mutations introduced in mutant Ts and Td affectthese sites, and we therefore analyzed the transcriptional ac-tivity of these mutants in transient LTR-luciferase transfectionassays. In addition, we tested whether two revertants of mutantTs increased the LTR activity of mutant Ts. Both basal andTat-activated transcriptional activities of the wild-type, mutant,and revertant LTRs were tested, but we measured no differ-ence among the different promoters. Furthermore, similar lev-els of virus production were measured in cells transiently trans-fected with the proviral constructs. Thus, these transcriptionfactor binding sites in the U5 region are either not affected bythe Ts and Td mutations or not important for viral replication.Nevertheless, the nonrandom pattern of reversion suggests thepresence of a sequence-specific motif in this part of the U5region.
Biochemical assays with virion-derived RNA-tRNA com-plexes showed that the reduced replication potential of mutantTs correlates with reduced tRNA priming efficiency, which isthe result of decreased tRNA occupancy of the PBS. We mea-sured nearly complete occupancy of the PBS for the wild-typetemplate, suggesting that both copies of the dimeric HIV-1RNA genome have an associated tRNA primer. This resultdiffers somewhat with studies on murine leukemia virus andavian leukosis virus, for which PBS occupancies of 50 and 70%have been reported (19, 48). We found the PBS occupancy ofthe mutant Ts template to be reduced to 23%. Thus, inclusionof part of the PBS in an excessively stable hairpin structureinhibits the annealing of the tRNA primer. This effect appar-ently restricts the U5-PBS hairpin from becoming excessivelystable in natural HIV and SIV isolates (5). In other words, theHIV-1 genome contains a structured RNA motif in the U5-PBS region that is at the threshold of becoming inhibitory tothe process of initiation of reverse transcription. In fact, thetRNA extension efficiency could be increased to 125% of thewild-type value by opening of the U5-PBS hairpin as in mutantTd. Although speculative, this hairpin may restrict prematuretRNA annealing to the viral RNA in the infected cell, but thisrestriction is apparently overcome in the context of the virionparticle, perhaps due to viral cofactors (see below). More ex-tended hairpin structures with greater thermodynamic stabilitywere predicted for the HIV-2 genome and several SIV vari-ants, but we previously stressed that these structures are char-acterized by having either a limited number of PBS nucleotidesthat are involved in base pairing or a large percentage ofrelatively weak G-U base pairs (7). There is recent evidence forribozymes that terminal G-U base pairs are involved in struc-tural rearrangements (50), which may explain the efficienttRNA annealing in HIV-2, despite the relatively stable U5-PBS structure. Obviously, there may be cofactors that facilitatetRNA annealing onto the PBS in the context of the viralparticle. One such a factor is the viral nucleocapsid (NC)
protein, which has been reported to facilitate the annealing ofthe tRNA primer to the PBS (23, 39). Apparently, the exces-sively stable Ts hairpin interferes with this process, and it willbe of interest to study this annealing reaction in more detail inin vitro assays in the absence and presence of NC protein. Suchstudies are currently being performed.
A more complex defect was apparent for the Td mutant withthe destabilized U5-PBS hairpin. This mutant template was atleast as efficient in tRNA extension as the wild-type template,demonstrating that the PBS is occupied by tRNA. Despiteefficient tRNA extension, we measured a strongly reduced PBSoccupancy in tests in which reverse transcription is primed byan oligonucleotide from a position downstream of the PBS.Apparently, the tRNA primer was released during the primer-annealing step. This result indicates that the interaction be-tween the tRNA primer and the mutant Td genome is lessstable than the complex with the wild-type template, eventhough the two templates have identical PBSs. Moreover, sev-eral additional stop products upstream and downstream of thePBS were observed for the wild-type template during extensionof the downstream primer with the AMV RT enzyme. Thesestops are due to tRNA annealing because the signals are notobserved with the mutant Ts template. Most importantly, thesestops were not observed either for the mutant Td template.These combined results indicate that a different conformationof the viral RNA-tRNA complex is reached on the wild-typetemplate compared with mutant Td, suggesting that the U5-PBS hairpin is directly or indirectly involved in correct tRNAannealing onto the viral RNA genome. Several studies suggestthat the A-rich loop of the U5-PBS hairpin interacts directlywith the anticodon of tRNA3
Lys (24–27, 33, 34, 40, 41, 46, 51,52), but the interpretation of these experiments is complicatedbecause this U5 sequence encodes multiple, overlapping rep-lication signals. For instance, mutations in the U5-PBS regionmay affect the secondary structure of this part of the leaderRNA (5), and the presence of overlapping integration signals(18, 36) makes it difficult to analyze the proposed interactionsbetween the viral RNA and the tRNA primer. The phenotypeof the carefully designed RNA structure mutant Td in thisstudy does support the idea that structured RNA motifs in theU5 region contribute to functional tRNA annealing. The in-duction of multiple stop signals upon tRNA binding to thewild-type template suggests that a structural rearrangementoccurs in the region surrounding the PBS, but the molecularnature of the additional viral RNA-tRNA interactions remainsto be determined.
The mutant and revertant analysis presented in this study isconsistent with the existence of the U5-PBS hairpin as depictedin Fig. 1. We previously proposed this hairpin conformation aspart of a secondary RNA structure model of the completeleader region of the HIV-1 genome (4). In fact, this HIV-1hairpin structure is not very stable but was modeled basedprimarily on similar structures in the HIV-2 RNA (7) and thegenomes of several SIV viruses (5). Although a different con-formation was recently proposed for this part of the HIV-1genome based on RNA structure probing experiments (14),our structure probing results and in particular the functionaldata strongly support the existence of the U5-PBS hairpin.Furthermore, this stem-loop structure is supported by phylo-genetic evidence based on the sequence of different HIV-1subtypes (not shown). It may nevertheless be too simplistic tosuggest that this part of the RNA genome has one static con-formation. Several factors will bind to this region of the viralgenome during discrete steps of virus replication, e.g., thetRNA primer, the NC protein, and the RNA itself duringdimerization, and it is likely that the RNA conformation will
TABLE 3. tRNA-priming efficiency and tRNA occupancy of wild-type and mutant templates
Construct
RT activity (%) AUG primer
CN1 tRNA Full-lengthDNA
Stopproduct
Wild type 100a 100a 9.8 90.2Ts 170 27 77 23Td 90 125 62 38
a Set at 100%.
2236 BEERENS ET AL. J. VIROL.
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

change during consecutive steps of the viral replication cycle. Itcannot even be excluded that this region acts as a molecularswitch during replication by changing between alternativeRNA conformations. We recently obtained evidence for such aconformational polymorphism of the HIV-1 leader RNA (9a).
ACKNOWLEDGMENTS
We thank Atze Das for helpful discussions, Bianca Schuijt for per-forming the LTR-luciferase transcription assays, and Wim van Est forphotography work.
This work was supported in part by the Dutch AIDS Fund and by theNetherlands Foundation for Chemical Research with financial aidfrom the Netherlands Organization for Scientific Research.
REFERENCES
1. Aiyar, A., D. Cobrinik, Z. Ge, H. J. Kung, and J. Leis. 1992. Interactionbetween retroviral U5 RNA and the TYC loop of the tRNATrp primer isrequired for efficient initiation of reverse transcription. J. Virol. 66:2464–2472.
2. Aiyar, A., Z. Ge, and J. Leis. 1994. A specific orientation of RNA secondarystructures is required for initiation of reverse transcription. J. Virol. 68:611–618.
3. Baudin, F., R. Marquet, C. Isel, J. L. Darlix, B. Ehresmann, and C. Ehres-mann. 1993. Functional sites in the 59 region of human immunodeficiencyvirus type 1 RNA from defined structural domains. J. Mol. Biol. 229:382–397.
4. Berkhout, B. 1996. Structure and function of the human immunodeficiencyvirus leader RNA. Prog. Nucleic Acid Res. Mol. Biol. 54:1–34.
5. Berkhout, B. 1997. The primer-binding site on the RNA genome of humanand simian immunodeficiency viruses is flanked by an upstream hairpinstructure. Nucleic Acids Res. 25:4013–4017.
6. Berkhout, B., B. Klaver, and A. T. Das. 1997. Forced evolution of a regula-tory RNA helix in the HIV-1 genome. Nucleic Acids Res. 25:940–947.
7. Berkhout, B., and I. Schoneveld. 1993. Secondary structure of the HIV-2leader RNA comprising the tRNA-primer binding site. Nucleic Acids Res.21:1171–1178.
8. Berkhout, B., R. H. Silverman, and K. T. Jeang. 1989. Tat trans-activates thehuman immunodeficiency virus through a nascent RNA target. Cell 59:273–282.
9. Berkhout, B., and J. L. B. van Wamel. 1996. Role of the DIS hairpin inreplication of human immunodeficiency virus type 1. J. Virol. 70:6723–6732.
9a.Berkhout, B., and J. L. B. van Wamel. The leader of the RNA genome formsa compactly folded tertiary structure. RNA, in press.
10. Brown, P. O. 1997. Integration, p. 161–204. In J. M. Coffin, S. H. Hughes, andH. E. Varmus (ed.), Retroviruses. Cold Spring Harbor Laboratory Press,New York, N.Y.
11. Clever, J., C. Sassetti, and T. G. Parslow. 1995. RNA secondary structureand binding sites for gag gene products in the 59 packaging signal of humanimmunodeficiency virus type 1. J. Virol. 69:2101–2109.
12. Clever, J. L., and T. G. Parslow. 1997. Mutant human immunodeficiencyvirus type 1 genomes with defects in RNA dimerization or encapsidation.J. Virol. 71:3407–3414.
13. Cobrinik, D., A. Aiyar, Z. Ge, M. Katzman, H. Huang, and J. Leis. 1991.Overlapping retrovirus U5 sequence elements are required for efficient in-tegration and initiation of reverse transcription. J. Virol. 65:3864–3872.
14. Damgaard, C. K., H. Dyhr-Mikkelsen, and J. Kjems. 1998. Mapping theRNA binding sites for human immunodeficiency virus type-1 Gag and NCproteins within the complete HIV-1 and -2 untranslated leader regions.Nucleic Acids Res. 26:3667–3676.
15. Das, A. T., B. Klaver, and B. Berkhout. 1995. Reduced replication of humanimmunodeficiency virus type 1 mutants that use reverse transcription primersother than the natural tRNA3
Lys. J. Virol. 69:3090–3097.16. Das, A. T., B. Klaver, and B. Berkhout. 1999. A hairpin structure in the R
region of the human immunodeficiency virus type 1 RNA genome is instru-mental in polyadenylation site selection. J. Virol. 73:81–91.
17. Das, A. T., B. Klaver, B. I. F. Klasens, J. L. B. van Wamel, and B. Berkhout.1997. A conserved hairpin motif in the R-U5 region of the human immu-nodeficiency virus type 1 RNA genome is essential for replication. J. Virol.71:2346–2356.
18. Esposito, D., and R. Craigie. 1998. Sequence specificity of viral end DNAbinding by HIV-1 integrase reveals critical regions for protein-DNA inter-action. EMBO J. 17:5832–5843.
19. Fu, W., B. A. Ortiz-Conde, R. J. Gorelick, S. H. Hughes, and A. Rein. 1997.Placement of tRNA primer on the primer-binding site requires pol geneexpression in avian but not murine retroviruses. J. Virol. 71:6940–6946.
20. Harrich, D., G. Mavankal, A. Mette-Snider, and R. B. Gaynor. 1995. Humanimmunodeficiency virus type 1 TAR element revertant viruses define RNAstructures required for efficient viral gene expression and replication. J. Vi-rol. 69:4906–4913.
21. Harrison, G. P., and A. M. L. Lever. 1992. The human immunodeficiencyvirus type 1 packaging signal and major splice donor region have a conservedstable secondary structure. J. Virol. 66:4144–4153.
22. Hoglund, S., A. Ohagen, J. Goncalves, A. T. Panganiban, and D. Gabuzda.1997. Ultrastructure of HIV-1 genomic RNA. Virology 233:271–279.
23. Huang, Y., A. Khorchid, J. Wang, M. A. Parniak, J.-L. Darlix, M. A. Wain-berg, and L. Kleiman. 1997. Effect of mutations in the nucleocapsid protein(NCp7) upon Pr160gag-pol and tRNALys incorporation into human immuno-deficiency virus type 1. J. Virol. 71:4378–4384.
24. Isel, C., C. Ehresmann, G. Keith, B. Ehresmann, and R. Marquet. 1995.Initiation of reverse transcription of HIV-1: secondary structure of theHIV-1 RNA/tRNA(3Lys) (template/primer). J. Mol. Biol. 247:236–250.
25. Isel, C., G. Keith, B. Ehresmann, C. Ehresmann, and R. Marquet. 1998.Mutational analysis of the tRNA3Lys/HIV-1 RNA (primer/template) com-plex. Nucleic Acids Res. 26:1198–1204.
25a.Isel, C., E. Westhof, S. F. Le Grice, B. Ehresmann, C. Ehresmann, and R.Marquet. 1999. Structural basis for the specificity of the initiation of HIV-1reverse transcription. EMBO J. 18:1038–1048.
26. Kang, S.-M., J. K. Wakefield, and C. D. Morrow. 1996. Mutations in both theU5 region and the primer-binding site influence the selection of the tRNAused for the initiation of HIV-1 reverse transcription. Virology 222:401–414.
27. Kang, S.-M., Z. Zhang, and C. D. Morrow. 1997. Identification of a sequencewithin U5 required for human immunodeficiency virus type 1 to stablymaintain a primer binding site complementary to tRNAMet. J. Virol. 71:207–217.
28. Klasens, B. I. F., M. Thiesen, A. Virtanen, and B. Berkhout. 1999. The abilityof the HIV-1 AAUAAA signal to bind polyadenylation factors is controlledby local RNA structure. Nucleic Acids Res. 27:446–454.
29. Klaver, B., and B. Berkhout. 1994. Comparison of 59 and 39 long terminalrepeat promoter function in human immunodeficiency virus. J. Virol. 68:3830–3840.
30. Klaver, B., and B. Berkhout. 1994. Evolution of a disrupted TAR RNAhairpin structure in the HIV-1 virus. EMBO J. 13:2650–2659.
31. Leis, J., A. Aiyar, and D. Cobrinik. 1993. Regulation of initiation of reversetranscription of retroviruses, p. 33–48. In A. M. Skalka and S. P. Goff (ed.),Reverse transcriptase. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, N.Y.
32. Li, X., J. Mak, E. J. Arts, Z. Gu, L. Kleiman, M. A. Wainberg, and M. A.Parniak. 1994. Effects of alterations of primer-binding site sequences onhuman immunodeficiency virus type 1 replication. J. Virol. 68:6198–6206.
33. Li, Y., Z. Zhang, J. K. Wakefield, S.-M. Kang, and C. D. Morrow. 1997.Nucleotide substitutions within U5 are critical for efficient reverse transcrip-tion of human immunodeficiency virus type 1 with a primer binding sitecomplementary to tRNAHis. J. Virol. 71:6315–6322.
34. Liang, C., X. Li, L. Rong, P. Inouye, Y. Quan, L. Kleiman, and M. A.Wainberg. 1997. The importance of the A-rich loop in human immunodefi-ciency virus type 1 reverse transcription and infectivity. J. Virol. 71:5750–5757.
35. Marquet, R., C. Isel, C. Ehresmann, and B. Ehresmann. 1995. tRNAs asprimer of reverse transcriptases. Biochimie 77:113–124.
36. Masuda, T., M. J. Kuroda, and S. Harada. 1998. Specific and independentrecognition of U3 and U5 att sites by human immunodeficiency virus type 1integrase in vivo. J. Virol. 72:8396–8402.
37. McBride, M. S., and A. T. Panganiban. 1996. The human immunodeficiencyvirus type 1 encapsidation site is a multipartite RNA element composed offunctional hairpin structures. J. Virol. 70:2963–2973.
37a.Morris, S., and J. Leis. 1998. Changes in Rous sarcoma virus RNA second-ary structure near the primer binding site upon tRNATrp primer annealing.J. Virol. 73:6307–6318.
38. Paillart, J.-C., L. Berthoux, M. Ottmann, J.-L. Darlix, R. Marquet, B. Ehres-mann, and C. Ehresmann. 1996. A dual role of the putative RNA dimer-ization initiation site of human immunodeficiency virus type 1 in genomicRNA packaging and proviral DNA synthesis. J. Virol. 70:8348–8354.
39. Prats, A. C., L. Sarih, C. Gabus, S. Litvak, G. Keith, and J. L. Darlix. 1988.Small finger protein of avian and murine retroviruses has nucleic acid an-nealing activity and positions the replication primer tRNA onto genomicRNA. EMBO J. 7:1777–1783.
40. Puglisi, E. V., and J. D. Puglisi. 1998. HIV-1 A-rich RNA loop mimics thetRNA anticodon structure. Nat. Med. 5:1033–1036.
41. Skripkin, E., C. Isel, B. Marquet, B. Ehresmann, and C. Ehresmann. 1996.Psoralen crosslinking between human immunodeficiency virus type 1 RNAand primer tRNAlys3. Nucleic Acids Res. 24:509–514.
42. Telesnitsky, A., and S. P. Goff. 1997. Reverse transcriptase and the genera-tion of retroviral DNA, p. 121–160. In J. M. Coffin, S. H. Hughes, and H. E.Varmus (ed.), Retroviruses. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
43. Van Lint, C., C. A. Amella, S. Emiliani, M. John, T. Jie, and E. Verdin. 1997.Transcription factor binding sites downstream of the human immunodefi-ciency virus type 1 transcription start site are important for virus infectivity.J. Virol. 71:6113–6127.
44. Verhoef, K., M. Koper, and B. Berkhout. 1997. Determination of the mini-mal amount of Tat activity required for human immunodeficiency virus type
VOL. 74, 2000 ROLE OF U5-PBS IN HIV-1 REPLICATION 2237
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

1 replication. Virology 237:228–236.45. Vicenzi, E., D. S. Dimitrov, A. Engelman, T.-S. Migone, D. F. J. Purcell, J.
Leonard, G. Englund, and M. A. Martin. 1994. An integration-defective U5deletion mutant of human immunodeficiency virus type 1 reverts by elimi-nating additional long terminal repeat sequences. J. Virol. 68:7879–7890.
46. Wakefield, J. K., S.-M. Kang, and C. D. Morrow. 1996. Construction of atype 1 human immunodeficiency virus that maintains a primer binding sitecomplementary to tRNAHis. J. Virol. 70:966–975.
47. Wakefield, J. K., H. Rhim, and C. D. Morrow. 1994. Minimal sequencerequirements of a functional human immunodeficiency virus type 1 primerbinding site. J. Virol. 68:1605–1614.
48. Whitcomb, J. M., B. A. Ortiz Conde, and S. H. Hughes. 1995. Replication ofavian leukosis viruses with mutations at the primer binding site: use ofalternative tRNAs as primer. J. Virol. 69:6228–6238.
49. Willey, R. L., D. H. Smith, L. A. Lasky, T. S. Theodore, P. L. Earl, B. Moss,
D. J. Capon, and M. A. Martin. 1988. In vitro mutagenesis identifies a regionwithin the envelope gene of the human immunodeficiency virus that iscritical for infectivity. J. Virol. 62:139–147.
50. Wu, M., and I. Tinoco. 1999. RNA folding causes secondary structure rear-rangement. Proc. Natl. Acad. Sci. USA 95:11555–11560.
51. Zhang, Z., S.-M. Kang, A. LeBlanc, S. L. Hajduk, and C. D. Morrow. 1996.Nucleotide sequences within the U5 region of the viral RNA genome are themajor determinants for an human immunodeficiency virus type 1 to maintaina primer binding site complementary to tRNAHis. Virology 226:306–317.
52. Zhang, Z., S.-M. Kang, Y. Li, and C. D. Morrow. 1998. Genetic analysis ofthe U5-PBS of a novel HIV-1 reveals multiple interactions between thetRNA and RNA genome required for initiation of reverse transcription.RNA 4:394–406.
53. Zuker, M. 1989. On finding all suboptimal foldings of an RNA molecule.Science 244:48–52.
2238 BEERENS ET AL. J. VIROL.
on March 31, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from











![MOTIF XF Editor VST Owner’s Manual7.Sélectionnez « MOTIF XF6 (MOTIF XF7 ou MOTIF XF8) » dans la colonne [FW Device] (Périphérique FW). 8.Sélectionnez « MOTIF XF6 (MOTIF XF7](https://img.dokumen.tips/doc/110x75/611158b13f31404d2d274378/motif-xf-editor-vst-owneras-manual-7slectionnez-motif-xf6-motif-xf7-ou.jpg)

















