Embed Size (px)
Citation preview

Matrix Biology 34 (2014) 1–12
Contents lists available at ScienceDirect
Matrix Biology
j ourna l homepage: www.e lsev ie r .com/ locate /matb io
A rapid increase in macrophage-derived versican and hyaluronan ininfectious lung disease
Mary Y. Chang a,⁎, Yoshinori Tanino c, Veronika Vidova a, Michael G. Kinsella d, Christina K. Chan d,Pamela Y. Johnson d, Thomas N. Wight d, Charles W. Frevert a,b
a Comparative Pathology Program, Department of Comparative Medicine, University of Washington School of Medicine, Seattle, WA, United Statesb Division of Pulmonary/Critical Care Medicine, Department of Medicine, University of Washington School of Medicine, Seattle, WA, United Statesc Fukushima Medical University School of Medicine, Department of Pulmonary Medicine, Fukushima, Japand Hope Heart Matrix Biology Program, Benaroya Research Institute at Virginia Mason, Seattle, WA, United States
Abbreviations: TLRs, toll-like receptors; LPS, lipopolysapolymerase chain reaction; IHC, immunohistochemistrbiotinylated hyaluronan binding protein; BMDMs, boneHas, hyaluronan synthase; Hyal, hyaluronidase; ELSA,SEM, standard error of the means.⁎ Corresponding author at: University of Washington
Program, Campus Box: 358052, 815 Mercer Street, SeaStates. Tel.: +1 206 616 6979.
E-mail address: [email protected] (M.Y. Ch
0945-053X/$ – see front matter © 2014 Elsevier B.V. All rhttp://dx.doi.org/10.1016/j.matbio.2014.01.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 19 November 2013Received in revised form 21 January 2014Accepted 21 January 2014Available online 26 January 2014
Keywords:LungMacrophageVersicanHyaluronanTLR-4
The goals of this study were to characterize the changes in chondroitin sulfate proteoglycans and hyaluronan inlungs in acute response to gram-negative bacterial infection and to identify cellular components responsible forthese changes. Mice were treated with intratracheal (IT) live Escherichia coli, E. coli lipopolysaccharide (LPS), orPBS. Both E. coli and LPS caused rapid selective increases in mRNA expression of versican and hyaluronan syn-thase (Has) isoforms 1 and 2 associated with increased immunohistochemical and histochemical staining forversican and hyaluronan in the lungs. Versicanwas associatedwith a subset of alveolarmacrophages. To examinewhether macrophages contribute to versican and hyaluronan accumulation, in vitro studies with primary cul-tures of bone marrow-derived and alveolar macrophages were performed. Unstimulated macrophagesexpressed very low levels of versican and hyaluronan synthase mRNA, with no detectible versican protein orhyaluronan product. Stimulation with LPS caused rapid increases in versicanmRNA and protein, a rapid increasein Has1mRNA, and concomitant inhibition of hyaluronidases 1 and 2, themajor hyaluronan degrading enzymes.Hyaluronan could be detected following chloroquine pre-treatment, indicating rapid turnover and degradationof hyaluronan by macrophages. In addition, the effects of LPS, the M1 macrophage classical activation agonist,were compared to those of IL-4/IL-13 or IL-10, the M2a and M2c alternative activation agonists, respectively.Versican and Has1 increased only in response to M1 activation. Finally, the up-regulation of versican and Has1in thewhole lungs ofwild-typemice following IT LPSwas completely abrogated in TLR-4−/−mice. Thesefindingssuggest that versican and hyaluronan synthesis may play an important role in the innate immune response togram-negative lung infection.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
In the acute response to bacterial lung infection, gram-negativebacteria are recognized by toll-like receptors (TLRs) on the surface ofalveolar macrophages triggering a cascade of events that leads to pul-monary inflammation and, ultimately, bacterial clearance and healing.TLR activation stimulates resident macrophages to secrete a variety ofchemokines, cytokines and other molecules that induce recruitment ofneutrophils and monocytes from the bloodstream. This leukocyte re-sponse is essential to the propagation and resolution of inflammation.
ccharide; IT, intratracheal; PCR,y; Ct, threshold cycle; b-HABP,marrow derived macrophages;enzyme-linked sorbent assay;
, N301/Comparative Pathologyttle, WA 98109-4714, United
ang).
ights reserved.
We are interested in the potential for proteoglycans and related mole-cules secreted by macrophages to have a role in promoting the acuteinflammatory response to bacterial infections in the lung.
Proteoglycans are important biological modifiers that influence bothhomeostasis and the response to injury (Esko and Lindahl, 2001; Parish,2006; Gill et al., 2010). In the lung, proteoglycans also have a role inorgan development and contribute to the innate immune response toinfection (Gill et al., 2010; Tanino et al., 2012). Following the treatmentof lungs with lipopolysaccharide (LPS), the glycosaminoglycan compo-sition changes from predominantly heparan sulfate in healthy lungs topredominantly chondroitin and dermatan sulfate in inflamed lungs(Karlinsky, 1982; Blackwood et al., 1983). Changes in the compositionof proteoglycans and their glycosaminoglycan chains, resulting inincreases in versican, decorin and biglycan, are also documented inboth animal models and human conditions of chronic lung disease(Karlinsky, 1982; Blackwood et al., 1983; Bensadoun et al., 1996,1997; Malmstrom et al., 2002; de Medeiros Matsushita et al., 2005).Hyaluronan is a glycosaminoglycan that is not attached to a core pro-tein, but binds to a number of chondroitin sulfate proteoglycans,
2 M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
including versican, to form large molecular weight complexes (Day andde la Motte, 2005). Similarly to versican, hyaluronan also is increased inchronic lung diseases (Hallgren et al., 1989; Nettelbladt and Hallgren,1989; Nettelbladt et al., 1989; Jiang et al., 2005) and has been shownto have an important role in airway mucosal defense (Forteza et al.,2001).
Temporal and spatial changes in the expression of specific chondroi-tin sulfate proteoglycans and hyaluronan in the lungs during the innateimmune response have not been systematically studied. Thus, the goalsof this study were to characterize the changes in chondroitin sulfateproteoglycans and hyaluronan in lungs of mice with gram-negativebacteria and to identify specific cellular components that contribute tothese changes. While multiple cell types are involved in the inflamma-tory response of the lungs (Strieter et al., 2002), this study focuses onthe contribution of macrophages and epithelial cells as these are thecells that first encounter airborne pathogens and have been shown tohave key roles in the initiation and regulation of inflammation in thelungs (Berg et al., 1993; Koay et al., 2002; Skerrett et al., 2004).
2. Results
2.1. Versican mRNA is Selectively Increased in Lungs of E. coli- andLPS-treated mice
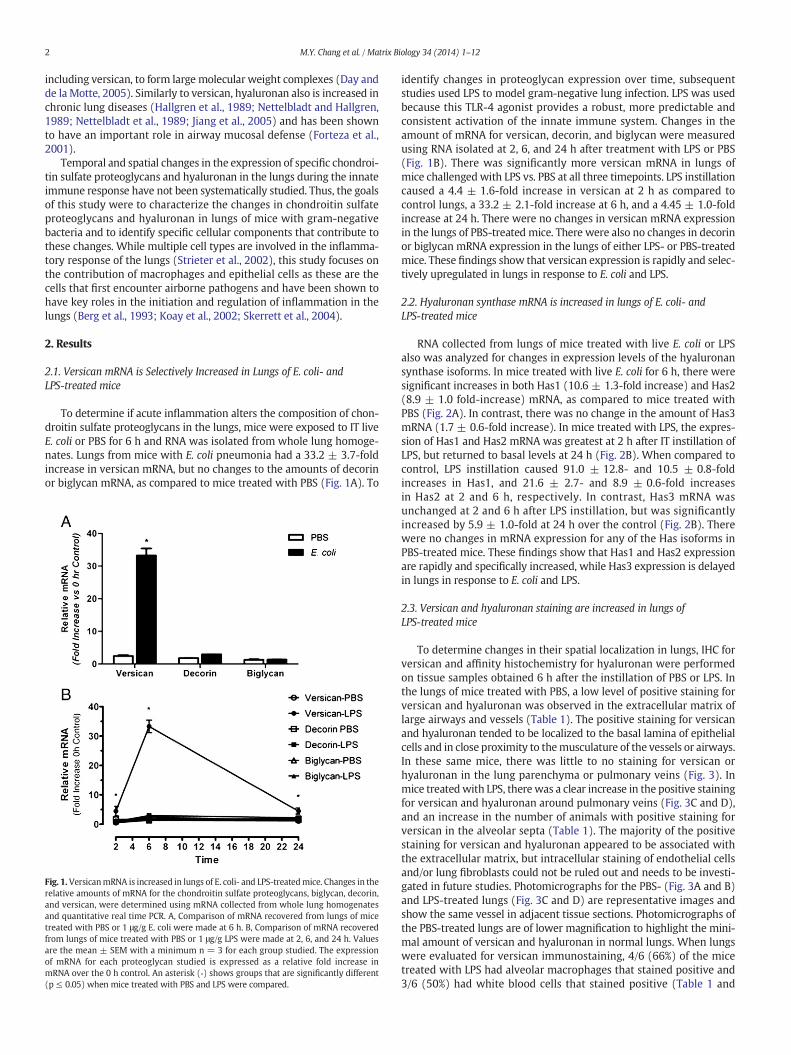
To determine if acute inflammation alters the composition of chon-droitin sulfate proteoglycans in the lungs, mice were exposed to IT liveE. coli or PBS for 6 h and RNA was isolated from whole lung homoge-nates. Lungs from mice with E. coli pneumonia had a 33.2 ± 3.7-foldincrease in versican mRNA, but no changes to the amounts of decorinor biglycan mRNA, as compared to mice treated with PBS (Fig. 1A). To
Fig. 1.VersicanmRNA is increased in lungs of E. coli- and LPS-treatedmice. Changes in therelative amounts of mRNA for the chondroitin sulfate proteoglycans, biglycan, decorin,and versican, were determined using mRNA collected from whole lung homogenatesand quantitative real time PCR. A, Comparison of mRNA recovered from lungs of micetreated with PBS or 1 μg/g E. coli were made at 6 h. B, Comparison of mRNA recoveredfrom lungs of mice treated with PBS or 1 μg/g LPS were made at 2, 6, and 24 h. Valuesare the mean ± SEM with a minimum n = 3 for each group studied. The expressionof mRNA for each proteoglycan studied is expressed as a relative fold increase inmRNA over the 0 h control. An asterisk (*) shows groups that are significantly different(p ≤ 0.05) when mice treated with PBS and LPS were compared.
identify changes in proteoglycan expression over time, subsequentstudies used LPS to model gram-negative lung infection. LPS was usedbecause this TLR-4 agonist provides a robust, more predictable andconsistent activation of the innate immune system. Changes in theamount of mRNA for versican, decorin, and biglycan were measuredusing RNA isolated at 2, 6, and 24 h after treatment with LPS or PBS(Fig. 1B). There was significantly more versican mRNA in lungs ofmice challengedwith LPS vs. PBS at all three timepoints. LPS instillationcaused a 4.4 ± 1.6-fold increase in versican at 2 h as compared tocontrol lungs, a 33.2 ± 2.1-fold increase at 6 h, and a 4.45 ± 1.0-foldincrease at 24 h. There were no changes in versican mRNA expressionin the lungs of PBS-treated mice. There were also no changes in decorinor biglycan mRNA expression in the lungs of either LPS- or PBS-treatedmice. These findings show that versican expression is rapidly and selec-tively upregulated in lungs in response to E. coli and LPS.
2.2. Hyaluronan synthase mRNA is increased in lungs of E. coli- andLPS-treated mice
RNA collected from lungs of mice treated with live E. coli or LPSalso was analyzed for changes in expression levels of the hyaluronansynthase isoforms. In mice treated with live E. coli for 6 h, there weresignificant increases in both Has1 (10.6 ± 1.3-fold increase) and Has2(8.9 ± 1.0 fold-increase) mRNA, as compared to mice treated withPBS (Fig. 2A). In contrast, there was no change in the amount of Has3mRNA (1.7 ± 0.6-fold increase). In mice treated with LPS, the expres-sion of Has1 and Has2 mRNA was greatest at 2 h after IT instillation ofLPS, but returned to basal levels at 24 h (Fig. 2B). When compared tocontrol, LPS instillation caused 91.0 ± 12.8- and 10.5 ± 0.8-foldincreases in Has1, and 21.6 ± 2.7- and 8.9 ± 0.6-fold increasesin Has2 at 2 and 6 h, respectively. In contrast, Has3 mRNA wasunchanged at 2 and 6 h after LPS instillation, but was significantlyincreased by 5.9 ± 1.0-fold at 24 h over the control (Fig. 2B). Therewere no changes in mRNA expression for any of the Has isoforms inPBS-treated mice. These findings show that Has1 and Has2 expressionare rapidly and specifically increased, while Has3 expression is delayedin lungs in response to E. coli and LPS.
2.3. Versican and hyaluronan staining are increased in lungs ofLPS-treated mice
To determine changes in their spatial localization in lungs, IHC forversican and affinity histochemistry for hyaluronan were performedon tissue samples obtained 6 h after the instillation of PBS or LPS. Inthe lungs of mice treated with PBS, a low level of positive staining forversican and hyaluronan was observed in the extracellular matrix oflarge airways and vessels (Table 1). The positive staining for versicanand hyaluronan tended to be localized to the basal lamina of epithelialcells and in close proximity to themusculature of the vessels or airways.In these same mice, there was little to no staining for versican orhyaluronan in the lung parenchyma or pulmonary veins (Fig. 3). Inmice treatedwith LPS, there was a clear increase in the positive stainingfor versican and hyaluronan around pulmonary veins (Fig. 3C and D),and an increase in the number of animals with positive staining forversican in the alveolar septa (Table 1). The majority of the positivestaining for versican and hyaluronan appeared to be associated withthe extracellular matrix, but intracellular staining of endothelial cellsand/or lung fibroblasts could not be ruled out and needs to be investi-gated in future studies. Photomicrographs for the PBS- (Fig. 3A and B)and LPS-treated lungs (Fig. 3C and D) are representative images andshow the same vessel in adjacent tissue sections. Photomicrographs ofthe PBS-treated lungs are of lower magnification to highlight the mini-mal amount of versican and hyaluronan in normal lungs. When lungswere evaluated for versican immunostaining, 4/6 (66%) of the micetreated with LPS had alveolar macrophages that stained positive and3/6 (50%) had white blood cells that stained positive (Table 1 and

Fig. 2. Hyaluronan synthase mRNA is increased in lungs of E. coli- and LPS-treated mice.Changes in the relative amounts of mRNA for the 3 isoforms of hyaluronan synthase,Has1, Has2 and Has3, were determined using mRNA collected from whole lung homoge-nates and quantitative real time PCR. A, Comparison of mRNA recovered from lungs ofmice treatedwith PBS or 1 μg/g E. coliweremade at 6 h. B, comparison ofmRNA recoveredfrom lungs of mice treated with PBS or 1 μg/g LPS were made at 2, 6, and 24 h. Values arethemean± SEMwith a minimum n=3 for each group studied. The expression ofmRNAfor each isoform is expressed as a relative fold increase in mRNA over the 0 h control. Anasterisk (*) shows groups that are significantly different (p ≤ 0.05) when mice treatedwith PBS and LPS were compared.
3M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
Fig. 3D and E). In contrast, there was no immunostaining of versican inwhite blood cells or alveolar macrophages in mice treated with PBS(Table 1 and Fig. 3B). Colloidal carbon was co-instilled with LPS to
identify the gross and microscopic locations where LPS was depositedin lungs. The uptake of colloidal carbon by alveolar macrophages wasused to identify the cells that were exposed to LPS. Positive stainingfor hyaluronan was not observed in macrophages or in white bloodcells in adjacent tissue sections from the same animals (Fig. 3C andTable 1). Thus, in contrast to control mice, significant positive stainingfor versican or hyaluronan was detected in the lungs by 6 hoursafter intratracheal administration of LPS, indicating that versican andhyaluronan are early responses to infection.
2.4. Versican mRNA and protein are increased in BMDMs treated with LPS
The finding of versican in a subset of alveolar macrophages exposedto LPS was examined in in vitro studies initially using BMDMs, assufficient numbers of these macrophages are easily obtained formRNA and protein analyses. Versican mRNA was detected at low levelsin untreatedmacrophages (Ct= 33.6± 0.3 for 15 ng cDNA, n=6), andexpression levels did not change in control cells over the time course ofthese experiments. However, versican mRNA was markedly increasedin response to LPS treatment (Fig. 4A). VersicanmRNAwas significantlyincreased from 6 to 16 h after treatment with 10 ng/ml of LPS, with amaximal increase of 180.9 ± 9.3-fold after 8 h, as compared to thePBS control (n = 3, p b 0.0001), and returned to basal levels after 24h. Treatment with a higher dose of LPS (100 ng/ml) had similar impactson the fold-increase and time course of versicanmRNA expression (datanot shown).
Proteoglycanswere isolated fromboth themedia and cell-associatedcompartments of BMDMs and evaluated by Western immunoblottingfor versican protein (Fig. 4B). Using an antibody specific for the versicanglycosaminoglycan-β domain, no versican was detected in the mediafrom cells treated with PBS or from cells treated for 4 h with LPS.However, versican protein was strongly induced after 24 h of treatmentwith either 10 or 100 ng/ml LPS. Using the same glycosaminoglycan-βantibody, no versican protein was detected in the cell-associated com-partment at either 4 or 24 h (data not shown). Also, no versican proteinwas detected in either compartment using an antibody specific for theversican glycosaminoglycan-α domain at 4 or 24 h (data not shown).These data indicate that the N250 kDa protein detected using theglycosaminoglycan-β antibody is the V1 isoform of versican which con-tains the glycosaminoglycan-β domain, but not the glycosaminoglycan-α domain of versican (Wight, 2002). Thus, LPS induces synthesis ofversican mRNA and secretion of the V1 isoform of versican by BMDMs.
2.5. Has1mRNA and hyaluronan synthesis are increased in BMDMs treatedwith LPS
The effects of LPS treatment on expression of the hyaluronansynthases (Has 1, 2 and 3), their product (hyaluronan), and the majorenzymes that degrade hyaluronan (hyaluronidases 1 and 2), were alsoexamined in vitro using BMDMs. Has1 mRNA was present at verylow levels in untreated macrophages (Ct = 38.1 ± 0.3 for 15 ng cDNA,n = 6) and expression levels did not change in control cells over thetime course of these experiments. However, Has1 mRNA was markedlyincreased in response to LPS treatment (Fig. 4C). Has1mRNAwas signif-icantly increased from 4 to 8 h after treatment with 10 ng/ml of LPS,with a maximal increase of 44.8 ± 11.6-fold after 6 h, as compared tothe PBS control (n = 3, p b 0.0001), and returned to basal levels after16 h. Treatment with a higher dose of LPS (100 ng/ml) had similar im-pacts on the fold-increase and time course of versicanmRNA expression(data not shown). Neither Has2 nor Has3 mRNA were detected in con-trol or LPS-treated BMDMs over the time course of these experiments(data not shown).
Hyaluronidase 1 and 2mRNAwere present at high levels in untreat-ed macrophages (Hyal1 Ct = 28.2 ± 0.1 and Hyal2 Ct = 23.3 ± 0.3 for15 ng cDNA, n = 3). Hyaluronidase 1 mRNA levels decreased in bothcontrol and LPS-treated cells over the time course of these experiments

Table 1Spatial location of positive staining for HA and versican in lungs of mice (6 h).
Bronchioles Bronchiole vessels Alveolar septa Pulmonary veins Alveolar macrophages White blood cells
PBS LPS PBS LPS PBS LPS PBS LPS PBS LPS PBS LPS
HA 4/4 6/6 4/4 6/6 1/4 4/6 1/4 5/6 0/4 0/6 0/4 0/6Versican 2/4 5/6 3/4 6/6 1/4 5/6 0/4 5/6 1/4 4/6 0/4 3/6
4 M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
(p b 0.001 at all times vs. control), with no significant differencesbetween the effects of PBS and LPS (Fig. 4D). In contrast, hyaluronidase2 mRNA levels did not change in control cells over the time course ofthese experiments, while LPS caused a transient decrease in hyaluroni-dase 2 that wasmaximally inhibited to 14.5% of control levels at 8 h andwas significantly lower from 2 to 12 h (p b 0.0001 at 4, 6, and 8 h vs.control), after which hyaluronidase 2 levels returned to basal levels(Fig. 4E). It should be noted that while hyaluronidase 1 and 2 levelswere significantly lower relative to control as early as 2 h after exposureto LPS, abundant message was still detected for both hyaluronidases atall times.
Hyaluronan synthesis by BMDMs was examined by quantifyinghyaluronidase-sensitive [3H]-glucosamine incorporation. Two consider-ations were incorporated into this assay. First, because of the timingof the mRNA responses of Has1 and Hyal2, which were maximallyhighest and lowest, respectively, within hours after exposure to LPS,we chose to evaluate [3H]-hyaluronan accumulation at the 2 and 4 htimepoints. Second, because no hyaluronan product could be detectedby hyaluronan ELSA (data not shown) we considered that hyaluronanmight be rapidly internalized and degraded bymacrophages. Therefore,chloroquine treatment was used to inhibit hyaluronan degradationin lysosomal and endosomal cellular compartments (Mapleson andBuchwald, 1981). A low level of [3H]-hyaluronan was measured in themedia of control cells (744 ± 290 dpm/200 μl media) and no effect ofLPS treatment could be detected by evaluating [3H]-hyaluronan in themedia at 2 or 4 h (Fig. 4F). However, when cells were pre-treatedwith chloroquine prior to LPS treatment, a 4.67 ± 1.75-fold increasein [3H]-hyaluronan production was measured at 4 h, as compared towithout chloroquine pre-treatment. No [3H]-hyaluronan was detectedin the cellular fraction with or without chloroquine treatment (datanot shown).
Thus, LPS specifically and temporally stimulates Has1 while concur-rently diminishing both hyaluronidases 1 and 2. Further inhibition ofpH-sensitive hyaluronan degradation with chloroquine allowed us todemonstrate increased hyaluronan synthesis by BMDMs in responseto LPS treatment. However, hyaluronan was not found to accumulatein either intracellular or extracellular compartments, suggesting thathyaluronan product is rapidly degraded and utilized for metabolicprocesses by the macrophages.
2.6. Versican and Has1 mRNA are increased in alveolar macrophagestreated with LPS
In a limited number of experiments, murine alveolar macrophageswere isolated and treated with LPS to verify that these lung macro-phages have responses similar to BMDMs macrophages. In responseto 4 h-treatment with LPS, versican mRNA increased 30.2 ± 3.2-fold(n = 4, p b 0.001) and Has1 mRNA increased 116.8 ± 24.0-fold vs.PBS control treatment (n = 4, p b 0.001) in alveolar macrophages(Fig. 5). Thus, alveolar macrophages and BMDMs respond similarly toLPS with respect to versican and Has1 mRNA regulation.
2.7. Versican and HAS1 mRNA are not increased in epithelial cells treatedwith LPS
In addition to macrophages, airway epithelial cells are important inthe acute response to lung infection. Thus, we also examined the effects
of LPS on murine tracheal epithelial cell expression of versican andhyaluronan synthase. Unlike the response by macrophages, LPS (10or 100 ng/ml) had no significant effect on either versican (Fig. 6A) orHas1 (Fig. 6B)mRNA at 4 or 24 h vs. control cells. Interestingly, where-as LPS had no effects on Has2 or Has3 mRNA in either bone marrow-derived or alveolar macrophages, this TLR4 agonist caused a signifi-cant increase in both Has2 and Has3 mRNA levels in epithelial cells(Fig. 6B). Has2 increased by 4.0 ± 0.7-fold (p b 0.0001) and 7.9 ±0.9-fold (p b 0.0001) in response to 10 ng/ml LPS vs. control at 4 and24 h, respectively. Has3 increased by 2.3 ± 0.4-fold and 6.0 ± 0.6-fold(p b 0.0001) vs. control at 4 and 24 h, respectively. Similar responseswere obtained with a higher concentration of LPS (100 ng/ml). Thesestudies indicate that macrophages and epithelial cells have distinct re-sponses to LPS in terms of versican and hyaluronan synthase regulation.
2.8. Versican and Has1 mRNA are Increased by M1 stimulation of BMDMs
As shown in Fig. 3 and Table 1, positive immunostaining for versicanwas found in themajority of, but not in all, alveolarmacrophages inmicetreated with LPS. In vitro studies have shown that alveolar macrophagesare functionally heterogeneous (Brannen and Chandler, 1988) andexhibit bothM1 andM2 polarizationmarkers (Duan et al., 2012). There-fore, we chose to examine whether versican expression might correlatewith M1 or M2macrophage polarization. BMDMswere treated for 4, 24or 48 h with (i) LPS, the classical M1 macrophage agonist; (ii) a combi-nation of IL-4 and IL-13, alternative M2a macrophage agonists; or(iii) IL-10, the deactivated M2c macrophage agonist (Mantovani et al.,2004) (Fig. 7). Versican mRNA was rapidly and strongly induced(132.0 ± 28.6-fold, p b 0.0001 vs. control cells) only in response to theM1 agonist, LPS (Fig. 7A). Has1 mRNA also was rapidly and strongly in-duced (156.6 ± 52.7-fold, p b 0.0001 vs. control cells) only in responseto the M1 agonist (Fig. 7B). Neither the combination of IL-4 and IL-13,nor IL-10 had any effect on versican or Has1 mRNA expression even atthe later time point of 48 h. None of these agonists stimulated Has2 orHas3 mRNA expression in BMDMs over the time course of these exper-iments (data not shown). iNOS served as the positive control for M1activation (Fig. 7C); Arg1 as the positive control for M2a activation(Fig. 7D); and CD163 as the positive control for M2c activation(Nielsen et al., 2006) (Fig. 7E). Thus, upregulation of both versican andHas1 genes reflect classical M1 activation.
2.9. Versican and Has1 are regulated through TLR-4
Two approaches were taken to evaluate the role of toll-like receptor4, the canonical LPS receptor, in mediating the versican and Has1 re-sponse to LPS. First, we considered that standard LPS preparationsprimarily consist of the polysaccharide Lipid A, or endotoxin, but maybe contaminated with bacterial lipopeptides. While endotoxins modu-late the immune response via TLR-4, lipopeptides interact with TLR-2.Therefore, mice were exposed to a preparation of ultra-pure LPSwhich only activates the TLR-4 pathway (Hajjar et al., 2012). FollowingIT administration of ultra-pure LPS, RNA was isolated from wholelung homogenates. Treatment with ultra-pure LPS (1 μg/g) caused a25.6 ± 0.9-fold increase in versican (p b 0.001) and a 21.9 ± 3.1-foldincrease in Has1 (p b 0.001) as compared to untreated lungs (Fig. 8A),comparable to results with IT administration of standard LPS (Figs. 1B& 2B). No further increases were observed with a higher dose of ultra-

Fig. 3.Versican and hyaluronan staining are increased in lungs of LPS-treatedmice. IHC for versican and affinity histochemistry for hyaluronan localization in lung tissue from PBS- or LPS-treatedmice. A & B, Lungs frommice treated for 6 h with PBS, 1 μg/g LPS (C & D), or 1 μg/g LPS (E) were stained with an antibody specific for the glycosaminoglycan-β domain of versican(B & D) or with b-HABP protein (A & C). Representative micrographs are shown. Gray arrows indicate alveolar macrophages, identified by colloidal carbon uptake, that do not showpositive staining for hyaluronan (C) or versican (D & E). The black arrows indicate alveolar macrophages that also show positive staining for versican (D & E). The black arrow headindicates a circulating white blood cell that shows positive staining for versican (D).
5M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
pure LPS (2 μg/g). There were no changes to the amounts of decorin orbiglycanmRNA in the lungs of mice treated with IT ultra-pure LPS (datanot shown), similar to findings with standard LPS (Fig. 1B). Second, weevaluated the effects of IT ultra-pure LPS on versican and Has1 geneexpression in the lungs of WT vs TLR-4−/− mice. The ability of IT LPSto stimulate both versican (Fig. 8B) and Has1 (Fig. 8C) was completelyabrogated in TLR-4−/− mice. These findings show that TLR-4 is criticalfor up-regulation of both versican and Has1 by LPS.
3. Discussion
The goals of this work were to characterize the changes to chondroi-tin sulfate proteoglycans and hyaluronan in the lungs of mice withgram-negative pneumonia and to identify specific cellular components
that contribute to these changes. These extracellular matrix compo-nents have been shown to be key mediators in events associated withthe inflammatory response (Day and de la Motte, 2005; Wight, 2008).Of particular interest are the roles of macrophages and epithelial cells,as these are the cells that first encounter airborne pathogens and havekey roles in the innate immune response of the lungs (Berg et al.,1993; Koay et al., 2002; Skerrett et al., 2004). Live E. coli instilled intothe airways of mice induced rapid and selective increases in wholelung mRNA expression of versican and two hyaluronan synthases,Has1 and Has2. To measure changes in the expression of these genesover time and to reduce animal-to-animal variability resulting fromlive bacteria, we chose to study mice instilled with LPS, a componentof the cell wall of gram-negative bacteria that signals through TLR-4(Poltorak et al., 1998). As with live E. coli, LPS induced rapid and
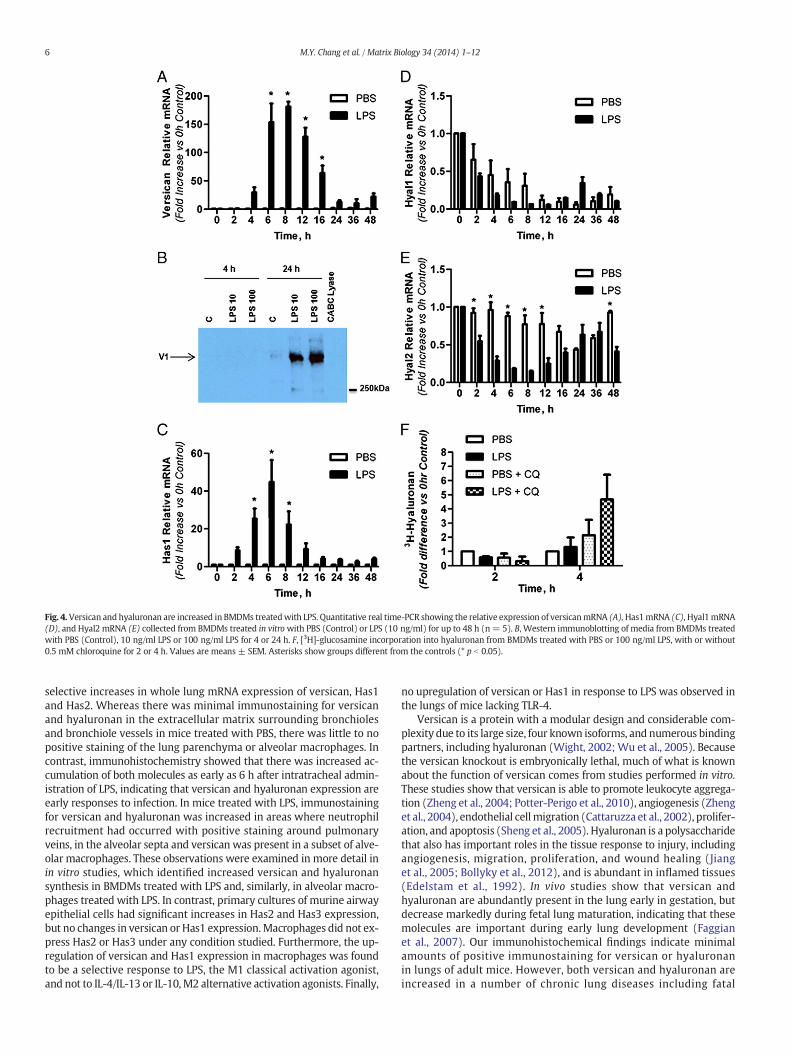
Fig. 4. Versican and hyaluronan are increased in BMDMs treatedwith LPS. Quantitative real time-PCR showing the relative expression of versicanmRNA (A),Has1mRNA (C), Hyal1mRNA(D), and Hyal2 mRNA (E) collected from BMDMs treated in vitrowith PBS (Control) or LPS (10 ng/ml) for up to 48 h (n= 5). B, Western immunoblotting of media from BMDMs treatedwith PBS (Control), 10 ng/ml LPS or 100 ng/ml LPS for 4 or 24 h. F, [3H]-glucosamine incorporation into hyaluronan from BMDMs treated with PBS or 100 ng/ml LPS, with or without0.5 mM chloroquine for 2 or 4 h. Values are means ± SEM. Asterisks show groups different from the controls (* p b 0.05).
6 M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
selective increases in whole lung mRNA expression of versican, Has1and Has2. Whereas there was minimal immunostaining for versicanand hyaluronan in the extracellular matrix surrounding bronchiolesand bronchiole vessels in mice treated with PBS, there was little to nopositive staining of the lung parenchyma or alveolar macrophages. Incontrast, immunohistochemistry showed that there was increased ac-cumulation of both molecules as early as 6 h after intratracheal admin-istration of LPS, indicating that versican and hyaluronan expression areearly responses to infection. In mice treated with LPS, immunostainingfor versican and hyaluronan was increased in areas where neutrophilrecruitment had occurred with positive staining around pulmonaryveins, in the alveolar septa and versican was present in a subset of alve-olar macrophages. These observations were examined in more detail inin vitro studies, which identified increased versican and hyaluronansynthesis in BMDMs treated with LPS and, similarly, in alveolar macro-phages treated with LPS. In contrast, primary cultures of murine airwayepithelial cells had significant increases in Has2 and Has3 expression,but no changes in versican or Has1 expression.Macrophages did not ex-press Has2 or Has3 under any condition studied. Furthermore, the up-regulation of versican and Has1 expression in macrophages was foundto be a selective response to LPS, the M1 classical activation agonist,and not to IL-4/IL-13 or IL-10,M2 alternative activation agonists. Finally,
no upregulation of versican or Has1 in response to LPS was observed inthe lungs of mice lacking TLR-4.
Versican is a protein with a modular design and considerable com-plexity due to its large size, four known isoforms, and numerous bindingpartners, including hyaluronan (Wight, 2002; Wu et al., 2005). Becausethe versican knockout is embryonically lethal, much of what is knownabout the function of versican comes from studies performed in vitro.These studies show that versican is able to promote leukocyte aggrega-tion (Zheng et al., 2004; Potter-Perigo et al., 2010), angiogenesis (Zhenget al., 2004), endothelial cellmigration (Cattaruzza et al., 2002), prolifer-ation, and apoptosis (Sheng et al., 2005). Hyaluronan is a polysaccharidethat also has important roles in the tissue response to injury, includingangiogenesis, migration, proliferation, and wound healing (Jianget al., 2005; Bollyky et al., 2012), and is abundant in inflamed tissues(Edelstam et al., 1992). In vivo studies show that versican andhyaluronan are abundantly present in the lung early in gestation, butdecrease markedly during fetal lung maturation, indicating that thesemolecules are important during early lung development (Faggianet al., 2007). Our immunohistochemical findings indicate minimalamounts of positive immunostaining for versican or hyaluronanin lungs of adult mice. However, both versican and hyaluronan areincreased in a number of chronic lung diseases including fatal
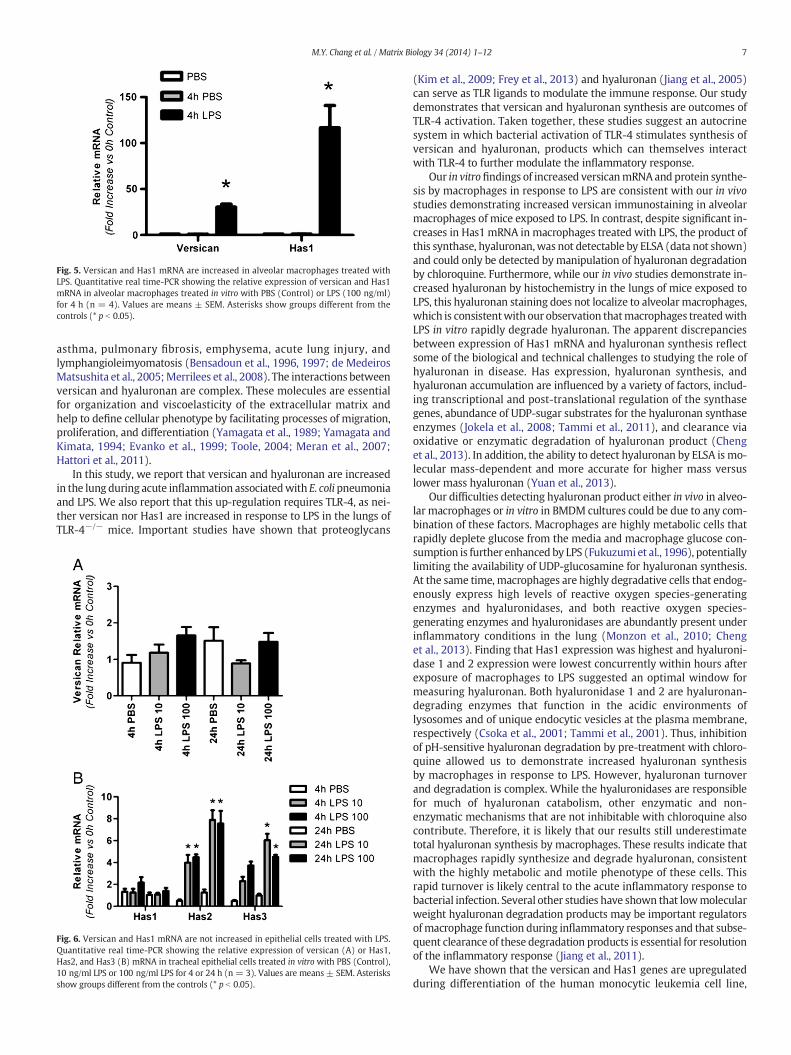
Fig. 5. Versican and Has1 mRNA are increased in alveolar macrophages treated withLPS. Quantitative real time-PCR showing the relative expression of versican and Has1mRNA in alveolar macrophages treated in vitro with PBS (Control) or LPS (100 ng/ml)for 4 h (n = 4). Values are means ± SEM. Asterisks show groups different from thecontrols (* p b 0.05).
7M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
asthma, pulmonary fibrosis, emphysema, acute lung injury, andlymphangioleimyomatosis (Bensadoun et al., 1996, 1997; de MedeirosMatsushita et al., 2005;Merrilees et al., 2008). The interactions betweenversican and hyaluronan are complex. These molecules are essentialfor organization and viscoelasticity of the extracellular matrix andhelp to define cellular phenotype by facilitating processes of migration,proliferation, and differentiation (Yamagata et al., 1989; Yamagata andKimata, 1994; Evanko et al., 1999; Toole, 2004; Meran et al., 2007;Hattori et al., 2011).
In this study, we report that versican and hyaluronan are increasedin the lung during acute inflammation associatedwith E. colipneumoniaand LPS. We also report that this up-regulation requires TLR-4, as nei-ther versican nor Has1 are increased in response to LPS in the lungs ofTLR-4−/− mice. Important studies have shown that proteoglycans
Fig. 6. Versican and Has1 mRNA are not increased in epithelial cells treated with LPS.Quantitative real time-PCR showing the relative expression of versican (A) or Has1,Has2, and Has3 (B) mRNA in tracheal epithelial cells treated in vitro with PBS (Control),10 ng/ml LPS or 100 ng/ml LPS for 4 or 24 h (n = 3). Values are means ± SEM. Asterisksshow groups different from the controls (* p b 0.05).
(Kim et al., 2009; Frey et al., 2013) and hyaluronan (Jiang et al., 2005)can serve as TLR ligands to modulate the immune response. Our studydemonstrates that versican and hyaluronan synthesis are outcomes ofTLR-4 activation. Taken together, these studies suggest an autocrinesystem in which bacterial activation of TLR-4 stimulates synthesis ofversican and hyaluronan, products which can themselves interactwith TLR-4 to further modulate the inflammatory response.
Our in vitrofindings of increased versicanmRNAand protein synthe-sis by macrophages in response to LPS are consistent with our in vivostudies demonstrating increased versican immunostaining in alveolarmacrophages of mice exposed to LPS. In contrast, despite significant in-creases in Has1 mRNA in macrophages treated with LPS, the product ofthis synthase, hyaluronan,was not detectable by ELSA (data not shown)and could only be detected bymanipulation of hyaluronan degradationby chloroquine. Furthermore, while our in vivo studies demonstrate in-creased hyaluronan by histochemistry in the lungs of mice exposed toLPS, this hyaluronan staining does not localize to alveolarmacrophages,which is consistentwith our observation thatmacrophages treatedwithLPS in vitro rapidly degrade hyaluronan. The apparent discrepanciesbetween expression of Has1 mRNA and hyaluronan synthesis reflectsome of the biological and technical challenges to studying the role ofhyaluronan in disease. Has expression, hyaluronan synthesis, andhyaluronan accumulation are influenced by a variety of factors, includ-ing transcriptional and post-translational regulation of the synthasegenes, abundance of UDP-sugar substrates for the hyaluronan synthaseenzymes (Jokela et al., 2008; Tammi et al., 2011), and clearance viaoxidative or enzymatic degradation of hyaluronan product (Chenget al., 2013). In addition, the ability to detect hyaluronan by ELSA is mo-lecular mass-dependent and more accurate for higher mass versuslower mass hyaluronan (Yuan et al., 2013).
Our difficulties detecting hyaluronan product either in vivo in alveo-lar macrophages or in vitro in BMDM cultures could be due to any com-bination of these factors. Macrophages are highly metabolic cells thatrapidly deplete glucose from the media and macrophage glucose con-sumption is further enhanced by LPS (Fukuzumi et al., 1996), potentiallylimiting the availability of UDP-glucosamine for hyaluronan synthesis.At the same time, macrophages are highly degradative cells that endog-enously express high levels of reactive oxygen species-generatingenzymes and hyaluronidases, and both reactive oxygen species-generating enzymes and hyaluronidases are abundantly present underinflammatory conditions in the lung (Monzon et al., 2010; Chenget al., 2013). Finding that Has1 expression was highest and hyaluroni-dase 1 and 2 expression were lowest concurrently within hours afterexposure of macrophages to LPS suggested an optimal window formeasuring hyaluronan. Both hyaluronidase 1 and 2 are hyaluronan-degrading enzymes that function in the acidic environments oflysosomes and of unique endocytic vesicles at the plasma membrane,respectively (Csoka et al., 2001; Tammi et al., 2001). Thus, inhibitionof pH-sensitive hyaluronan degradation by pre-treatment with chloro-quine allowed us to demonstrate increased hyaluronan synthesisby macrophages in response to LPS. However, hyaluronan turnoverand degradation is complex. While the hyaluronidases are responsiblefor much of hyaluronan catabolism, other enzymatic and non-enzymatic mechanisms that are not inhibitable with chloroquine alsocontribute. Therefore, it is likely that our results still underestimatetotal hyaluronan synthesis by macrophages. These results indicate thatmacrophages rapidly synthesize and degrade hyaluronan, consistentwith the highly metabolic and motile phenotype of these cells. Thisrapid turnover is likely central to the acute inflammatory response tobacterial infection. Several other studies have shown that lowmolecularweight hyaluronan degradation products may be important regulatorsofmacrophage function during inflammatory responses and that subse-quent clearance of these degradation products is essential for resolutionof the inflammatory response (Jiang et al., 2011).
We have shown that the versican and Has1 genes are upregulatedduring differentiation of the human monocytic leukemia cell line,

Fig. 7. Versican and Has1 mRNA are increased by M1, but not by M2, stimulation of BMDMs. Quantitative real time-PCR showing the relative expression of versican (A), Has1 (B), iNOS(C) andArg1 (D) inmacrophages treatedwith PBS (control), 10 ng/ml LPS, 10 ng/ml IL-4 plus 10 ng/ml IL-13, or 10 ng/ml IL4 for 4, 24 or 48 h (n=3). Values aremeans± SEM. Asterisksshow groups different from the controls (* p b 0.05).
8 M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
THP1, from monocytes into macrophages (Chang et al., 2012). Tran-scription profiling of human monocyte-to-macrophage polarizationindicates that versican (i.e., chondroitin sulfate proteoglycan 2) is oneof the genes that is induced by M1 activation using the combination ofLPS and interferon gamma in vitro (Martinez et al., 2006). Similarly,the present in vitro studies indicate that versican and Has1 are inducedby M1 macrophage activation with LPS. Hyaluronan interactionswith its cell surface receptors, CD44 and RHAMM, are important toinflammatory rolling, adhesion, and cell activation (Hall et al., 1994;DeGrendele et al., 1996) and hyaluronan-containing matrices havebeen shown to possess immunomodulating properties that dampenM1 inflammatory macrophage functions (Franz et al., 2013). Ourin vivo studies indicate positive versican staining in the majority of,but not all, alveolar macrophages in mice exposed to LPS, suggestingthat these alveolar macrophages are heterogeneous and differentiallyactivated. This is in keeping with the understanding that alveolar mac-rophages have diverse roles inmaintaininghomeostasis and respondingto inflammatory crises (Duan et al., 2012) and are heterogeneous ina number of ex vivo functional assays indicative of differential macro-phage activation, e.g., release of soluble mediators and migrationtowards a chemotactic peptide (Brannen and Chandler, 1988). Thus,versican and hyaluronan appear to be markers of pro-inflammatoryconditions.
The current data show that LPS exerts differential effects on macro-phages and epithelial cells with respect to versican and hyaluronansynthase expression. Thus, it is likely that the selective acute increasesin versican and Has1 seen in the whole lung after IT administration ofLPS reflect, at least in part, the macrophage response, whereas thewhole lung increase in Has2 reflects, at least in part, the epithelialresponse. The differential regulation of the hyaluronan synthases inmacrophages and epithelial cells treated with LPS suggests that theregulation of the genes for these three membrane-bound enzymesdiffers. The relative specific activities of these Has isoforms and thesize distributions of their hyaluronan products are different (Itanoet al., 1999; Jiang et al., 2007). We find that LPS induces Has1mRNA ex-pression by macrophages. Has1 synthesizes a relatively high molecular
weight product (Itano et al., 1999). The anti-inflammatory properties ofhigh molecular weight hyaluronan are evidenced by its ability todecrease LPS-induced inflammation in microglial cells (Austin et al.,2012), to promote immune tolerance by augmenting regulatory T cellfunction cells (Bollyky et al., 2009), and to protect against lung injuryin a variety of models (Lennon and Singleton, 2011). We also find thatLPS induces Has2 mRNA expression by airway epithelial cells. Lung ep-ithelial cell-specific overexpression of Has2 and its high molecularweight hyaluronan product has been shown to be protective againstacute lung injury, in part through TLR-dependent interactions (Jianget al., 2005).
While macrophages and epithelial cells are the first airway cells toencounter and respond to airborne pathogens, endothelial cells and fi-broblasts in the underlying tissue are also likely to respond to infection.The studies reported here, and work of others, indicate that theseunderlying cells also are involved in the expression and accumulationof versican and hyaluronan. Our immunohistochemical studies ofwhole lung demonstrate versican in the cytoplasm of endothelial cellswithin pulmonary veins, and in vitro studies have shown that activationof primary human pulmonary endothelial cells with tumor necrosisfactor-α or vascular endothelial growth factor increases the expressionof the versican isoforms, V1, V2, and V3 (Cattaruzza et al., 2002). Analy-ses of bronchoalveolar lavagefluid show that LPS stimulates theproduc-tion of both tumor necrosis factor-α and vascular endothelial growthfactor (data not shown), suggesting that these two inflammatorymedi-ators may contribute to the increase in versican expression by pulmo-nary endothelial cells. In addition, lung fibroblasts treated with theviral mimetic, poly I:C, secrete a matrix rich in versican and hyaluronanto which T cells bind avidly (Evanko et al., 2012). Activation of fibro-blasts by poly I:C requires the adaptor molecule Trif, a signaling mole-cule that becomes activated when LPS binds to TLR4 (Yamamoto et al.,2003). These findings suggest that activation of TLR4 signalingpathways in lung fibroblasts results in a pericellular matrix rich inversican and hyaluronan. Thus, multiple cell types within the lunghave the capacity for increased production of versican and hyaluronanin response to inflammatory conditions.

9M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
In summary, these findings suggest that versican and hyaluronansynthesis play an important role in the innate immune response togram-negative lung infection. Whether versican and hyaluronanare themselves pro-inflammatory molecules or simply a response toinflammation is not yet clear. Understanding the functional roles thatthese molecules have in the innate immune response to lung infectionis the focus of ongoing studies.
4. Materials and methods
4.1. Reagents
E. coli 91–100696 serotype K1 was a clinical isolate obtained from apatient with bacteremia due to biliary sepsis. LPS from E. coli serotype0111:B4 was purchased from List Biological Laboratories (Campbell,CA). Ultrapure LPS-EB from E. coli serotype 0111:B4 was purchasedfrom Invivogen (San Diego, CA). Colloidal carbon was prepared aspreviously described (Frevert et al., 2000). The polyclonal rabbitanti-versican antibody (β-GAG) was from Chemicon International(Temecula, CA). Gene-specific TaqMan primer-probe mixes used forquantitative real time PCR of versican, decorin, biglycan, Has1, Has2,Has3, hyaluronidase 1 and 2, and 18S mRNA were purchased from Ap-plied Biosystems (Foster City, CA). IL-4, IL-13, and IL-10were purchasedfrom Peprotech (Rocky Hill, NJ).
4.2. Preparation of bacteria
The use of E. coli serotype K-1 in mice was previously described(Matute-Bello et al., 2001). Methods used to pass and store the bacteriahave been detailed elsewhere (Frevert et al., 2000). On the daybefore anexperiment, a frozen aliquot of E. coliwas thawed, inoculated into 50mlLennox-B broth, and incubated overnight at 37 °C in a shaking incuba-tor. The bacteria were recovered by centrifugation, washed once inPBS, and resuspended in sterile water to 2 × 1010 CFU/ml. Bacterial con-centrationswere confirmed by quantitative culture using the pour platemethod.
Fig. 8. Versican and Has1are regulated through TLR-4, changes in the relative amounts ofmRNA for versican and Has1 were determined using mRNA collected from whole lunghomogenates and quantitative real time PCR. A, Comparison of mRNA recovered fromlungs of mice treated with PBS, 1 μg/g or 2 μg/g of ultra-pure LPS were made at 4 h.B, Comparison of versican mRNA recovered from lungs of WT or TLR-4−/− mice treatedwith PBS or 1 μg/g ultra-pure LPS were made at 4 h. C, Comparison of Has1 mRNA recov-ered from lungs of WT or TLR-4−/− mice treated with PBS or 1 μg/g ultra-pure LPS weremade at 4 h. Values are the mean ± SEM with a minimum n= 3 for each group studied.The expression of mRNA is expressed as a relative fold increase in mRNA over the 0 hcontrol. * Shows groups that are significantly different (p ≤ 0.05) when mice treatedwith PBS and LPS were compared. # shows groups that are significantly different(p ≤ 0.05) when WT vs TLR4−/− mice were compared.
4.3. Animal protocols
C57BL/6 and TLR-4−/−mice (7–8 weeks age) (Jackson Laboratories)were housed in an SPF animal facility at the University of Washingtonand the UW Institutional Animal Care and Use Committee approvedall studies performed. Using methods that were previously described,the intratracheal (IT) instillation of live E. coli, E. coli 0111:B4 LPS(1 μg/g), or PBS was performed in mice anesthetized with 3–4%isoflurane (Matute-Bello et al., 2001). To identify the location wherePBS, bacteria, or LPS or ultrapure LPS-EB were instilled in lungs at nec-ropsy and in tissues sections, 1% colloidal carbon was mixed into thesesolutions as previously described (Frevert et al., 2000). Following theIT instillation, mice were recovered and returned to a cage where theywere allowed access to food and water for the remainder of the study.At 2, 6, and 24 h after the IT instillation, mice were euthanized with120 mg/kg pentobarbital.Whole bloodwas obtained by direct punctureand the plasma was collected and stored at −70 °C until use. Thethoracic cavity was opened by amidline incision and the lungs were re-moved and placed into RNAlater (Ambion, Austin, TX) and stored at 4 °Covernight for the extraction of RNA. For animal studies where tissueswere obtained for immunohistochemistry (IHC), C57BL/6 mice wereinstilled with LPS, euthanized at specified times, then the lungs werecannulated with a 24-gage catheter and perfusion fixed using 10%neutral buffered formalin and a perfusion pressure of 21 cm H2O.Lungs were then processed for lightmicroscopy as previously described(Jiang et al., 2005).
4.4. Isolation of RNA from lung tissue
Total RNA was isolated from lung tissue using the Absolute RNAMiniprep Kit (Stratagene, La Jolla, CA). Genomic DNA was digested byincubation with DNAse I (Ambion) and RNA was reverse transcribedusing the High Capacity cDNA Archive Kit (Applied Biosystems) at25 °C for 10 min, at 37 °C for 2 h, and at 90 °C for 5 min. The resultingcDNAwasused for standard polymerase chain reaction (PCR) and quan-titative real-time PCR.

10 M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
4.5. Measurement of mRNA using quantitative real time PCR
Quantitative real-time PCR was performed using an ABI PRISM 7000Sequence Detector and TaqMan gene expression assays for versican,biglycan, decorin, the hyaluronan synthases, Has1, Has2, and Has3,and 18S (Life Technologies, Grand Island, NY). Real-time PCR wascarried out in a total volume of 25 μl with a master mixture includingall reagents required for PCR and gene-specific TaqMan primer-probemixes. Cycle parameterswere 50 °C for 2min, 95 °C for 10min, followedby 40 cycles of 95 °C for 15 s and 60 °C for 1 min. Delta Ct (ΔCt) was cal-culated as the difference in threshold cycles (Ct) for the target genescompared to 18S. Relative mRNA levels were calculated by the 2−ΔΔCt
method (Livak and Schmittgen, 2001) to express the fold increaseover values obtained from RNA collected from untreatedmice (i.e., nor-mal lungs) or from untreated cells. No template controls and no reversetranscriptase controls were performed simultaneously.
4.6. Preparation of tissue for immunohistochemistry
Lung tissuewas fixed by IT instillation of 4% paraformaldehyde at 21cm H2O pressure. After inflation, the lungs were immersed in 4% para-formaldehyde, then embedded in paraffin and sectioned using standardmethods and used for the immunohistochemical analysis.
4.7. Immunohistochemistry
IHC for versican and affinity histochemistry for hyaluronan wereperformed with paraffin-embedded tissue. After deparaffinization, en-dogenous peroxidases were blocked using H2O2 in methanol and tissuesectionswere rehydrated in a series of graded ethanol. For versican IHC,sections were digested with 0.2 U/ml of chondroitinase ABC to removechondroitin sulfate side chains and expose versican epitopes. Tissueswere then incubated with 5% Carnation non-fat milk in PBS for tissueblocking, followed by overnight incubation with either rabbit anti-versican polyclonal antibody or isotype-matched control IgG. This wasfollowed by biotinylated goat anti-rabbit IgG antibody for 2 h at roomtemperature. The tissue sections were rinsed twice with PBS andthen incubated with the Vector “Elite” ABC-HP kit (Burlingame, CA) ina moist chamber for 30 min at room temperature. Detection wasperformed using the Vector NovaRed substrate (Burlingame, CA) for10 min at room temperature. Slides were counterstained with Gills #3hematoxylin. The same protocol was used for hyaluronan affinity histo-chemistry, except that the tissue was not treated with chondroitinaseABC and biotinylated hyaluronan binding protein (b-HABP, the N-terminal hyaluronan binding region of aggrecan which has beenbiotinylated)was used instead of the primary and secondary antibodies.
4.8. Assessment of positive staining for versican and hyaluronan by IHC
The anatomical location of the positive staining for versican andhyaluronan in lungs was identified using tissues collected from animalstreated with PBS or LPS for 6 h. A comparative pathologist (CWF) eval-uated tissues collected from all animals and the positive staining forversican and hyaluronan was identified in six anatomical locations,bronchioles, bronchial vessels, pulmonary veins, alveolar septa, alveolarmacrophages and white blood cells. Colloidal carbon was co-instilledwith the PBS and LPS to identify the gross and microscope location ofLPS deposition in the lungs. Alveolar macrophages exposed to LPSwere identified by cellular morphology and the presence of colloidalcarbon in their cytoplasm. Each animal (n = 4 for PBS and n = 6 forLPS treated mice) were individually scored as having positive stainingfor versican or HA in the six anatomical locations. This analysis wasperformed in a blinded manner.
4.9. In vitro cell culture studies
Bone marrow-derived macrophages (BMDMs) were isolated fromfemurs and tibia of mice and cultured in “macrophage medium”
(RPMI 1640, 10% FBS, 30% L929 cell supernatant, 2 mM L-glutamine,100 IU/mL penicillin, and 100 μg/mL streptomycin) (Tanino et al.,2012). After 24 h, adherent cells were placed in macrophage medium,and cultured for 6 days. Macrophages were then re-plated in macro-phage medium in a 6-well tissue culture dish at a density of 2 × 106
cells/well for 24 h and then stimulated in the presence or absence ofLPS (10 or 100 ng/mL), ultrapure LPS-EB (10 or 100 ng/ml), a combina-tion of IL-4 (10 ng/ml) and IL-13 (10 ng/ml), or IL-10 (10 ng/ml) inRPMI with 10% FBS for 4, 24, or 48 h. Murine alveolar macrophageswere isolated from BAL fluid, cultured for 24 h in macrophage medium,and then stimulatedwith 100 ng/ml LPS for 4 h. Primarymouse trachealepithelial cells were cultured at an air–liquid interface, as previouslydescribed, and stimulated with LPS (10 or 100 ng/mL) for 4 and 24 h(Kassim et al., 2007).
4.10. Isolation of RNA from cell cultures and quantitative real time PCR
Total RNA was obtained from cell culture monolayers using theRNeasy Mini Kit from Qiagen (Valencia, CA). Complementary DNAwas prepared by reverse transcription using the High Capacity cDNAReverse Transcription Kit from Applied Biosciences (Carlsbad, CA).Quantitative real-time PCR was performed using an ABI PRISM 7000Sequence Detector (Applied Biosystems) to quantify the mRNA expres-sion of versican, the hyaluronan synthases (Has1, Has2, and Has3), thehyaluronidases (Hyal1 and Hyal2), iNOS, Arg1, and 18S as describedabove.
4.11. Isolation of proteoglycans
Cell medium was collected and combined with protease inhibitors(5 mM benzamidine, 100 mM 6-aminohexanoic acid, and 1 mMphenylmethylsulfonyl fluoride). The cell layer was rinsed withphosphate-buffered saline and solubilized in 8 M urea buffer (8 Murea, 2 mM EDTA, 0 or 0.25 M NaCl, 50 mM Tris–HCl, and 0.5% TritonX-100 detergent, pH 7.4) containing protease inhibitors (Schonherret al., 1991, 1993). Themedia contained secreted proteoglycans, where-as cell layers contained cell membrane-associated, intracellular andextracellular matrix proteoglycans. Media and cell layer extracts wereconcentrated and purified by ion-exchange chromatography ondiethylaminoethyl-Sephacel in 8 M urea buffer with 0.25 M NaCl andeluted with 8 M urea buffer containing 3 M NaCl (Schonherr et al.,1991, 1993).
4.12. Versican Western
ForWestern immunoblotting, proteoglycans isolated by ion-exchangechromatography were digested with 0.05 units chondroitin ABC lyase(North Star BioProducts, East Falmouth, MA) in 0.3 M Tris–HCl, pH8.0, 0.6 mg/ml bovine serum albumin and 18 mM sodium acetatewith protease inhibitors for 3 h at 37 °C) and run on SDS-PAGE (4–12% with 3.5% stacking gel) under reducing conditions (Laemmli,1970). Separated proteins were electrophoretically transferred to 0.2μm nitrocellulose membranes (GE Healthcare, Piscataway, NJ) using aBioRad Transblot SD Semi-Dry Transfer Cell (BioRad, Hercules, CA)(Olin et al., 1999). The transferred proteins were then detected withthe primary antibody to versican (Millipore), and enhanced chemilumi-nescence (Western-Light Chemiluminescent Detection System) withproprietary luminescent substrate (Applied Biosystems, Foster City,CA) (Lemire et al., 2007).

11M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
4.13. Quantification of [3H]-hyaluronan synthesis
Macrophage cultures were metabolically labeled with [3H]-glucos-amine (50 □Ci/ml) in the presence or absence of 100 ng/ml LPS and/or 0.5mMchloroquine for 2, 4, 6 and 24 h.Media and cell layer fractionswere harvested separately and digested with pronase (100 μg/ml) in0.5 M Tris pH 6.5 overnight at 37 °C. Following digestion, the pronasewas inactivated by heating to 100 °C for 20 min. Radiolabeled macro-molecules were then recovered and separated from unincorporatedprecursor by precipitation on nitrocellulose membranes using slotblot analysis (Agren et al., 1994). Briefly, 200 μl of sample was addedto an equal volume of 2% cetylpyridinium chloride, 50 mM NaCl (CPCwash solution) and the solution blotted onto 0.45 μm nitrocellulosemembrane (Schleicher and Schuell, Keene, NH). The membranewas washed six times in CPC wash solution and once in deionizedwater before air-drying at room temperature overnight. Incorporationof [3H]-glucosamine into hyaluronan was measured by digestingan equivalent radiolabeled aliquot with Streptomyces hyaluronidase(2 U/ml) for 24 h at 37 °C before slot blotting. Hyaluronan was mea-sured as the amount of hyaluronidase-sensitive material precipitatedto the nitrocellulose membrane. All scintillation counting was done onBeckman LS 6500 (Beckman Instruments, Fullerton, CA).
4.14. Hyaluronan Enzyme-linked Sorbent Assay (ELSA)
Media was digested with 300 μg/ml pronase for 18 h at 37 °C.Following digestion, the pronase was inactivated by heating to 100 °Cfor 20 min. The hyaluronan ELSA is a modification (Wilkinson et al.,2004) of a previously described (Underhill et al., 1993) competitiveELSA inwhich the samples to be assayed are first mixedwith biotinylat-ed hyaluronan binding protein and then added to hyaluronan-coatedmicrotiter plates; therefore the final signal is inversely proportional tothe amount of hyaluronan in the sample. Specifically, Nunc Maxisorp96-well plates were coated with an excess of hyaluronan (Sigma),which was covalently cross-linked to bovine serum album to enhanceits retention by the plastic, and blocked with PBS containing serum.Hyaluronan standards and media samples were pre-incubated with b-HABP. After incubation, the mixtures were added to the wells andexcess b-HABP bound to the hyaluronan in the wells, while b-HABPalready bound to hyaluronan was washed away. Thus, increasingamounts of hyaluronan resulted in decreasing amounts of b-HABP avail-able to be retained in the wells. Bound b-HABPwas detected colorimet-rically with peroxidase-labeled streptavidin, H2O2, and the peroxidasesubstrate 2,2 azinobis (3-ethylbenzthiazoline sulfonic acid) in sodiumcitrate buffer, pH 4.2. The reaction product was monitored spectropho-tometrically at OD405.
4.15. Statistical Analysis
A t-test and p b 0.05was used for statistical analysis of data collectedfrom studies where comparisons were made between PBS- and LPS-treated animals. Statistical analysis of multiple groups was performedusing log-transformed data and two-way ANOVA with Bonferroni'sMultiple Comparison Test and p b 0.05. Values are reported as means ±standard error of the means (SEM) unless otherwise specified.
Acknowledgements
This work was supported by National Institutes of Health grantsHL098067 (to C.W.F. and T.N.W.), RR030249-02 (to C.W.F.) and by theVA Medical Research Service (to C.W.F.).
The authors would like to thank Drs. Adeline Hajjar and Susan Perigofor helpful discussions. The authors also thank Dowon An, Gina Kiske,Vivian Lee, StevenMongovin, and TaraWigmosta for technical assistance.
References
Agren, U.M., Tammi, R., Tammi, M., 1994. A dot-blot assay of metabolically radiolabeledhyaluronan. Anal. Biochem. 217, 311–315.
Austin, J.W., Gilchrist, C., Fehlings, M.G., 2012. Highmolecular weight hyaluronan reduceslipopolysaccharide mediated microglial activation. J. Neurochem. 122, 344–355.
Bensadoun, E.S., Burke, A.K., Hogg, J.C., Roberts, C.R., 1996. Proteoglycan deposition inpulmonary fibrosis. Am. J. Respir. Crit. Care Med. 154, 1819–1828.
Bensadoun, E.S., Burke, A.K., Hogg, J.C., Roberts, C.R., 1997. Proteoglycans in granuloma-tous lung diseases. Eur. Respir. J. 10, 2731–2737.
Berg, J.T., Lee, S.T., Thepen, T., Lee, C.Y., Tsan, M.F., 1993. Depletion of alveolar macro-phages by liposome-encapsulated dichloromethylene diphosphonate. J. Appl. Physiol.74, 2812–2819.
Blackwood, R.A., Cantor, J.O., Moret, J., Mandl, I., Turino, G.M., 1983. Glycosaminoglycansynthesis in endotoxin-induced lung injury. Proc. Soc. Exp. Biol. Med. 174, 343–349.
Bollyky, P.L., Falk, B.A., Wu, R.P., Buckner, J.H., Wight, T.N., Nepom, G.T., 2009. Intact extra-cellular matrix and the maintenance of immune tolerance: high molecular weighthyaluronan promotes persistence of induced CD4 + CD25+ regulatory T cells.J. Leukoc. Biol. 86, 567–572.
Bollyky, P.L., Bogdani, M., Bollyky, J.B., Hull, R.L., Wight, T.N., 2012. The role of hyaluronanand the extracellular matrix in islet inflammation and immune regulation. Curr. Diab.Rep. 12, 471–480.
Brannen, A.L., Chandler, D.B., 1988. Alveolar macrophage subpopulations' responsivenessto chemotactic stimuli. Am. J. Pathol. 132, 161–166.
Cattaruzza, S., Schiappacassi,M., Ljungberg-Rose, A., Spessotto, P., Perissinotto, D.,Morgelin,M., Mucignat, M.T., Colombatti, A., Perris, R., 2002. Distribution of PG-M/versican vari-ants in human tissues and de novo expression of isoform V3 upon endothelial cellactivation, migration, and neoangiogenesis in vitro. J. Biol. Chem. 277, 47626–47635.
Chang, M.Y., Chan, C.K., Braun, K.R., Green, P.S., O'Brien, K.D., Chait, A., Day, A.J., Wight,T.N., 2012. Monocyte-to-macrophage differentiation: synthesis and secretion of acomplex extracellular matrix. J. Biol. Chem. 287, 14122–14135.
Cheng, G., Swaidani, S., Sharma, M., Lauer, M.E., Hascall, V.C., Aronica, M.A., 2013. Correla-tion of Hyaluronan Deposition with Infiltration of Eosinophils and Lymphocytes in aCockroach-Induced Murine Model of Asthma. Glycobiology 23, 43–58.
Csoka, A.B., Frost, G.I., Stern, R., 2001. The six hyaluronidase-like genes in the human andmouse genomes. Matrix Biol. 20, 499–508.
Day, A.J., de la Motte, C.A., 2005. Hyaluronan cross-linking: a protective mechanism ininflammation? Trends Immunol. 26, 637–643.
de Medeiros Matsushita, M., da Silva, L.F., dos Santos, M.A., Fernezlian, S., Schrumpf, J.A.,Roughley, P., Hiemstra, P.S., Saldiva, P.H., Mauad, T., Dolhnikoff, M., 2005. Airwayproteoglycans are differentially altered in fatal asthma. J. Pathol. 207, 102–110.
DeGrendele, H.C., Estess, P., Picker, L.J., Siegelman, M.H., 1996. CD44 and its ligandhyaluronate mediate rolling under physiologic flow: a novel lymphocyte-endothelialcell primary adhesion pathway. J. Exp. Med. 183, 1119–1130.
Duan, M., Li, W.C., Vlahos, R., Maxwell, M.J., Anderson, G.P., Hibbs, M.L., 2012. Distinctmacrophage subpopulations characterize acute infection and chronic inflammatorylung disease. J. Immunol. 189, 946–955.
Edelstam, G.A., Laurent, U.B., Lundkvist, O.E., Fraser, J.R., Laurent, T.C., 1992. Concentrationand turnover of intraperitoneal hyaluronan during inflammation. Inflammation 16,459–469.
Esko, J.D., Lindahl, U., 2001. Molecular diversity of heparan sulfate. J. Clin. Invest. 108,169–173.
Evanko, S.P., Angello, J.C., Wight, T.N., 1999. Formation of hyaluronan- and versican-richpericellular matrix is required for proliferation and migration of vascular smoothmuscle cells. Arterioscler. Thromb. Vasc. Biol. 19, 1004–1013.
Evanko, S.P., Potter-Perigo, S., Bollyky, P.L., Nepom, G.T., Wight, T.N., 2012. Hyaluronanand versican in the control of human T-lymphocyte adhesion and migration. MatrixBiol. 31, 90–100.
Faggian, J., Fosang, A.J., Zieba, M.,Wallace, M.J., Hooper, S.B., 2007. Changes in versican andchondroitin sulfate proteoglycans during structural development of the lung. Am.J. Physiol. Regul. Integr. Comp. Physiol. 293, R784–R792.
Forteza, R., Lieb, T., Aoki, T., Savani, R.C., Conner, G.E., Salathe, M., 2001. Hyaluronan servesa novel role in airway mucosal host defense. FASEB J. 15, 2179–2186.
Franz, S., Allenstein, F., Kajahn, J., Forstreuter, I., Hintze, V., Moller, S., Simon, J.C., 2013. Ar-tificial extracellular matrices composed of collagen 1 and high-sulfated hyaluronanpromote phenotypic and functional modulation of human pro-inflammatory M1macrophages. Acta Biomater. 9, 5621–5629.
Frevert, C.W., Matute-Bello, G., Skerrett, S.J., Goodman, R.B., Kajikawa, O., Sittipunt, C.,Martin, T.R., 2000. Effect of CD14 blockade in rabbits with Escherichia coli pneumoniaand sepsis. J. Immunol. 164, 5439–5445.
Frey, H., Schroeder, N., Manon-Jensen, T., Iozzo, R.V., Schaefer, L., 2013. Biological interplaybetween proteoglycans and their innate immune receptors in inflammation. FEBS J.280, 2165–2179.
Fukuzumi, M., Shinomiya, H., Shimizu, Y., Ohishi, K., Utsumi, S., 1996. Endotoxin-inducedenhancement of glucose influx into murine peritoneal macrophages via GLUT1.Infect. Immun. 64, 108–112.
Gill, S., Wight, T.N., Frevert, C.W., 2010. Proteoglycans: key regulators of pulmonaryinflammation and the innate immune response to lung infection. Anat. Rec. (Hoboken)293, 968–981.
Hajjar, A.M., Ernst, R.K., Fortuno III, E.S., Brasfield, A.S., Yam, C.S., Newlon, L.A., Kollmann,T.R., Miller, S.I., Wilson, C.B., 2012. Humanized TLR4/MD-2 mice reveal LPS recogni-tion differentially impacts susceptibility to Yersinia pestis and Salmonella enterica.PLoS Pathog. 8, e1002963.
Hall, C.L., Wang, C., Lange, L.A., Turley, E.A., 1994. Hyaluronan and the hyaluronan recep-tor RHAMM promote focal adhesion turnover and transient tyrosine kinase activity.J. Cell Biol. 126, 575–588.

12 M.Y. Chang et al. / Matrix Biology 34 (2014) 1–12
Hallgren, R., Samuelsson, T., Laurent, T.C., Modig, J., 1989. Accumulation of hyaluronan(hyaluronic acid) in the lung in adult respiratory distress syndrome. Am. Rev. Respir.Dis. 139, 682–687.
Hattori, N., Carrino, D.A., Lauer, M.E., Vasanji, A., Wylie, J.D., Nelson, C.M., Apte, S.S., 2011.Pericellular versican regulates the fibroblast–myofibroblast transition: a role forADAMTS5 protease-mediated proteolysis. J. Biol. Chem. 286, 34298–34310.
Itano, N., Sawai, T., Yoshida, M., Lenas, P., Yamada, Y., Imagawa, M., Shinomura, T.,Hamaguchi, M., Yoshida, Y., Ohnuki, Y., Miyauchi, S., Spicer, A.P., McDonald, J.A.,Kimata, K., 1999. Three isoforms of mammalian hyaluronan synthases have distinctenzymatic properties. J. Biol. Chem. 274, 25085–25092.
Jiang, D., Liang, J., Fan, J., Yu, S., Chen, S., Luo, Y., Prestwich, G.D., Mascarenhas, M.M., Garg,H.G., Quinn, D.A., Homer, R.J., Goldstein, D.R., Bucala, R., Lee, P.J., Medzhitov, R., Noble,P.W., 2005. Regulation of lung injury and repair by Toll-like receptors andhyaluronan. Nat. Med. 11, 1173–1179.
Jiang, D., Liang, J., Noble, P.W., 2007. Hyaluronan in tissue injury and repair. Annu. Rev.Cell Dev. Biol. 23, 435–461.
Jiang, D., Liang, J., Noble, P.W., 2011. Hyaluronan as an immune regulator in humandiseases. Physiol. Rev. 91, 221–264.
Jokela, T.A., Jauhiainen, M., Auriola, S., Kauhanen, M., Tiihonen, R., Tammi, M.I., Tammi,R.H., 2008. Mannose inhibits hyaluronan synthesis by down-regulation of the cellularpool of UDP-N-acetylhexosamines. J. Biol. Chem. 283, 7666–7673.
Karlinsky, J.B., 1982. Glycosaminoglycans in emphysematous and fibrotic hamster lungs.Am. Rev. Respir. Dis. 125, 85–88.
Kassim, S.Y., Gharib, S.A., Mecham, B.H., Birkland, T.P., Parks, W.C., McGuire, J.K., 2007.Individual matrix metalloproteinases control distinct transcriptional responses inairway epithelial cells infected with Pseudomonas aeruginosa. Infect. Immun. 75,5640–5650.
Kim, S., Takahashi, H., Lin,W.W., Descargues, P., Grivennikov, S., Kim, Y., Luo, J.L., Karin, M.,2009. Carcinoma-produced factors activate myeloid cells through TLR2 to stimulatemetastasis. Nature 457, 102–106.
Koay, M.A., Gao, X., Washington, M.K., Parman, K.S., Sadikot, R.T., Blackwell, T.S.,Christman, J.W., 2002. Macrophages are necessary for maximal nuclear factor-kappa B activation in response to endotoxin. Am. J. Respir. Cell Mol. Biol. 26, 572–578.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227, 680–685.
Lemire, J.M., Chan, C.K., Bressler, S., Miller, J., LeBaron, R.G., Wight, T.N., 2007. Interleukin-1beta selectively decreases the synthesis of versican by arterial smooth muscle cells.J. Cell. Biochem. 101, 753–766.
Lennon, F.E., Singleton, P.A., 2011. Role of hyaluronan and hyaluronan-binding proteins inlung pathobiology. Am. J. Physiol. Lung Cell. Mol. Physiol. 301, L137–L147.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods 25, 402–408.
Malmstrom, J., Larsen, K., Hansson, L., Lofdahl, C.G., Norregard-Jensen, O., Marko-Varga, G.,Westergren-Thorsson, G., 2002. Proteoglycan and proteome profiling of centralhuman pulmonary fibrotic tissue utilizing miniaturized sample preparation: a feasi-bility study. Proteomics 2, 394–404.
Mantovani, A., Sica, A., Sozzani, S., Allavena, P., Vecchi, A., Locati, M., 2004. The chemokinesystem in diverse forms of macrophage activation and polarization. Trends Immunol.25, 677–686.
Mapleson, J.L., Buchwald, M., 1981. Effect of cycloheximide and dexamethasone phos-phate on hyaluronic acid synthesis and secretion in cultured human skin fibroblasts.J. Cell. Physiol. 109, 215–222.
Martinez, F.O., Gordon, S., Locati, M., Mantovani, A., 2006. Transcriptional profiling of thehuman monocyte-to-macrophage differentiation and polarization: new moleculesand patterns of gene expression. J. Immunol. 177, 7303–7311.
Matute-Bello, G., Frevert, C.W., Liles, W.C., Nakamura, M., Ruzinski, J.T., Ballman, K., Wong,V.A., Vathanaprida, C., Martin, T.R., 2001. Fas/Fas ligand system mediates epithelialinjury, but not pulmonary host defenses, in response to inhaled bacteria. Infect.Immun. 69, 5768–5776.
Meran, S., Thomas, D., Stephens, P., Martin, J., Bowen, T., Phillips, A., Steadman, R., 2007.Involvement of hyaluronan in regulation of fibroblast phenotype. J. Biol. Chem. 282,25687–25697.
Merrilees, M.J., Ching, P.S., Beaumont, B., Hinek, A., Wight, T.N., Black, P.N., 2008. Changesin elastin, elastin binding protein and versican in alveoli in chronic obstructivepulmonary disease. Respir. Res. 9, 41.
Monzon, M.E., Fregien, N., Schmid, N., Falcon, N.S., Campos, M., Casalino-Matsuda, S.M.,Forteza, R.M., 2010. Reactive oxygen species and hyaluronidase 2 regulate airwayepithelial hyaluronan fragmentation. J. Biol. Chem. 285, 26126–26134.
Nettelbladt, O., Hallgren, R., 1989. Hyaluronan (hyaluronic acid) in bronchoalveolarlavage fluid during the development of bleomycin-induced alveolitis in the rat. Am.Rev. Respir. Dis. 140, 1028–1032.
Nettelbladt, O., Tengblad, A., Hallgren, R., 1989. Lung accumulation of hyaluronan parallelspulmonary edema in experimental alveolitis. Am. J. Physiol. 257, L379–L384.
Nielsen, M.J., Madsen, M., Moller, H.J., Moestrup, S.K., 2006. The macrophage scavengerreceptor CD163: endocytic properties of cytoplasmic tail variants. J. Leukoc. Biol. 79,837–845.
Olin, K.L., Potter-Perigo, S., Barrett, P.H., Wight, T.N., Chait, A., 1999. Lipoprotein lipaseenhances the binding of native and oxidized low density lipoproteins to versicanand biglycan synthesized by cultured arterial smooth muscle cells. J. Biol. Chem.274, 34629–34636.
Parish, C.R., 2006. The role of heparan sulphate in inflammation. Nat. Rev. Immunol. 6,633–643.
Poltorak, A., He, X., Smirnova, I., Liu, M.Y., Van Huffel, C., Du, X., Birdwell, D., Alejos, E.,Silva, M., Galanos, C., Freudenberg, M., Ricciardi-Castagnoli, P., Layton, B., Beutler, B.,1998. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4gene. Science 282, 2085–2088.
Potter-Perigo, S., Johnson, P.Y., Evanko, S.P., Chan, C.K., Braun, K.R.,Wilkinson, T.S., Altman,L.C., Wight, T.N., 2010. Polyinosine-polycytidylic acid stimulates versican accumula-tion in the extracellular matrix promoting monocyte adhesion. Am. J. Respir. CellMol. Biol. 43, 109–120.
Schonherr, E., Jarvelainen, H.T., Sandell, L.J., Wight, T.N., 1991. Effects of platelet-derivedgrowth factor and transforming growth factor-beta 1 on the synthesis of a largeversican-like chondroitin sulfate proteoglycan by arterial smooth muscle cells.J. Biol. Chem. 266, 17640–17647.
Schonherr, E., Jarvelainen, H.T., Kinsella, M.G., Sandell, L.J., Wight, T.N., 1993. Platelet-derived growth factor and transforming growth factor-beta 1 differentially affectthe synthesis of biglycan and decorin by monkey arterial smooth muscle cells.Arterioscler. Thromb. 13, 1026–1036.
Sheng, W., Wang, G., Wang, Y., Liang, J., Wen, J., Zheng, P.S., Wu, Y., Lee, V., Slingerland, J.,Dumont, D., Yang, B.B., 2005. The roles of versican V1 and V2 isoforms in cell prolif-eration and apoptosis. Mol. Biol. Cell 16, 1330–1340.
Skerrett, S.J., Liggitt, H.D., Hajjar, A.M., Ernst, R.K., Miller, S.I., Wilson, C.B., 2004. Respirato-ry epithelial cells regulate lung inflammation in response to inhaled endotoxin. Am.J. Physiol. Lung Cell. Mol. Physiol. 287, L143–L152.
Strieter, R.M., Belperio, J.A., Keane, M.P., 2002. Cytokines in innate host defense in thelung. J. Clin. Invest. 109, 699–705.
Tammi, R., Rilla, K., Pienimaki, J.P., MacCallum, D.K., Hogg, M., Luukkonen, M., Hascall, V.C.,Tammi, M., 2001. Hyaluronan enters keratinocytes by a novel endocytic route forcatabolism. J. Biol. Chem. 276, 35111–35122.
Tammi, R.H., Passi, A.G., Rilla, K., Karousou, E., Vigetti, D., Makkonen, K., Tammi, M.I., 2011.Transcriptional and post-translational regulation of hyaluronan synthesis. FEBS J. 278,1419–1428.
Tanino, Y., Chang, M.Y., Wang, X., Gill, S.E., Skerrett, S., McGuire, J.K., Sato, S., Nikaido, T.,Kojima, T., Munakata, M., Mongovin, S., Parks, W.C., Martin, T.R., Wight, T.N.,Frevert, C.W., 2012. Syndecan-4 regulates early neutrophil migration and pulmonaryinflammation in response to lipopolysaccharide. Am. J. Respir. Cell Mol. Biol. 47,196–202.
Toole, B.P., 2004. Hyaluronan: from extracellular glue to pericellular cue. Nat. Rev. Cancer4, 528–539.
Underhill, C.B., Nguyen, H.A., Shizari, M., Culty, M., 1993. CD44 positive macrophages takeup hyaluronan during lung development. Dev. Biol. 155, 324–336.
Wight, T.N., 2002. Versican: a versatile extracellular matrix proteoglycan in cell biology.Curr. Opin. Cell Biol. 14, 617–623.
Wight, T.N., 2008. Arterial remodeling in vascular disease: a key role for hyaluronan andversican. Front. Biosci. 13, 4933–4937.
Wilkinson, T.S., Potter-Perigo, S., Tsoi, C., Altman, L.C., Wight, T.N., 2004. Pro- and anti-inflammatory factors cooperate to control hyaluronan synthesis in lung fibroblasts.Am. J. Respir. Cell Mol. Biol. 31, 92–99.
Wu, Y.J., La Pierre, D.P., Wu, J., Yee, A.J., Yang, B.B., 2005. The interaction of versican withits binding partners. Cell Res. 15, 483–494.
Yamagata, M., Kimata, K., 1994. Repression of a malignant cell-substratum adhesionphenotype by inhibiting the production of the anti-adhesive proteoglycan, PG-M/versican. J. Cell Sci. 107 (Pt 9), 2581–2590.
Yamagata, M., Suzuki, S., Akiyama, S.K., Yamada, K.M., Kimata, K., 1989. Regulation of cell-substrate adhesion by proteoglycans immobilized on extracellular substrates. J. Biol.Chem. 264, 8012–8018.
Yamamoto, M., Sato, S., Hemmi, H., Hoshino, K., Kaisho, T., Sanjo, H., Takeuchi, O.,Sugiyama, M., Okabe, M., Takeda, K., Akira, S., 2003. Role of adaptor TRIF in theMyD88-independent toll-like receptor signaling pathway. Science 301, 640–643.
Yuan, H., Tank, M., Alsofyani, A., Shah, N., Talati, N., LoBello, J.C., Kim, J.R., Oonuki, Y., de laMotte, C.A., Cowman, M.K., 2013. Molecular mass dependence of hyaluronan detec-tion by sandwich ELISA-like assay and membrane blotting using biotinylatedhyaluronan binding protein. Glycobiology 23, 1270–1280.
Zheng, P.S., Vais, D., Lapierre, D., Liang, Y.Y., Lee, V., Yang, B.L., Yang, B.B., 2004. PG-M/versican binds to P-selectin glycoprotein ligand-1 and mediates leukocyte aggrega-tion. J. Cell Sci. 117, 5887–5895.





























