Embed Size (px)
Citation preview

A
APPLICA
D
Pr
ATION O
SURFAC
DissertatiE
resident: PrSupe
Me
OF AQUA
CE-GROU
Mar
ion to obnvironm
rof. José Maervisor: Prof
Co‐supeembers: Pro
Dr
J
ATIC BIO
UNDWAT
ryam Shap
btain themental En
Jury
anuel de Saf. Luis Filipeervisor: Dr. Aof.Luis Cancr. Tibor Stig
June 2012
DIVERSI
TER ECO
pouri
e Master ngineerin
ldanha Gone Tavares RiAna Silva ela de Fonster
2
TY TO M
OTONES
Degree ng
nçalves Matbeiro
seca
MONITOR
S
in
tos
1
R

2
Acknowledgment:
When I started to investigate for this work I knew very little in this subject. After two years working with
a group of specialist in this filed I end up learning so many things. I am grateful for all the support and
help that I received from my professors, friends and family.
First I would like to thank my previous professor and my dear friend Paula Tavares who initiated this
work and taught me many things in this field. She kindly passed me her valuable experiment and
knowledge and I am so happy that I started my first steps with her.
I do like to thank Professor Luis Ribeiro for accepting the supervision of this thesis. He never
hesitated to help me when I needed and gave me the possibility to work on a topic that I really liked.
I also like to thank Dr. Ana Silva for her valuable advices during the work. She was always available
and helpful specially for running the Primer software and fundamental revision of the thesis. This work
has been done under her supervision in all the steps.
I truly want to thank Professor Luis Fonseca, for teaching me great experiment in the field, laboratory
and specially identification of fauna. I will always be grateful for his generous welcome in Algarve
University, the full integration in the Lab and all the support that he provided me.
I would like to thank Dr. Paula Chainho for hosting me in the Oceanographic Centre in Lisbon
University. I always enjoyed from discussion with her and I am so grateful for her helpful advice for
writing this thesis.
I appreciate my dear friends help in Benthic Ecology Laboratory, in Lisbon University. They assisted
me identifying the benthic fauna and they were always keen to pass their knowledge.
I would like to thank Dear Margarida Machado for her sincere help in the identification of benthic
fauna. I enjoyed spending time with her and working in her laboratory in Algarve University. Without
her help it was not possible to finish the identification.

3
Dr. Sanda Lepure, and their group in Imdea Centre in Universidad de Alcalá in Spain. Sanda had
given me great hand in identification of groundwater fauna. I am more than grateful for her help in the
laboratory and valuable scientific discussion that made me motivated to continue working on this field.
I am thankful to Dr.Tibor Stigter, for his company in the field and passing great information on the
geology and hydrogeology aspects of the study area.
Geosystem center (CVRM) is appreciated for having hosted this study. In particular, the financial
support by CLIMWAT project facilitated the samplings and field trips.
I do like to thank my dear friends from CVRM and master degree study to make these two years such
a memorable and joyful experiment. I want to thank my friends out of university for their sweet and
warm encouragements and being with me every moment of my life.
My last but not least thanks are for my family, my dear parents, my brothers and my sister to give me
the courage to travel abroad and expend my knowledge in a global experiment. I want to thank Vahid
my dearest friend, company and husband that has been always push me through the difficult moment
of my scientific life. The accomplishment of this work owes to you for giving me your brilliant idea,
warmest company and kindest help.
Maryam Shapouri, June 2012

4
Abstract
This study follows two experiments with two different objectives. The first experiment aimed to
characterise the estuarine benthic community along a salinity gradient reflecting the conditions of
groundwater dependent ecosystems. The second experiment aimed to examine the sensitivity of
stygofauna to variation in groundwater salinity/conductivity accessed through wells. In the first
experiment, the spatial and temporal differences in the structure and composition of benthic
invertebrates were studied at a branching channel of the main estuarine channel of Arade river in
south of Portugal, which receives significant groundwater input and is also influence by semi diurnal
tides from the Arade estuary. Benthic macroinvertebrates were sampled at the end of the wet and dry
periods in 2009, along the salinity gradient created by the groundwater entrance in the channel.
Physico-chemical variables were measured to determine their association in the benthic fauna
distribution. Results indicated that the abundance of species Isopode Cyathura carinata and the
Polychaeta Heteromastus filiformis varied significantly with the salinity gradient created by the
groundwater discharge into the estuarine habitat and with sampling time. The Polychaeta were more
abundant at the end of dry season than wet season, and also are more abundant at the points of high
salinity, suggesting their tendency for being more associated to saline water. However, the
Polychaeta Hediste diversicolor and Alkmaria romijni were more abundant in areas of lower salinity
and at the end of the wet season. Some taxa such as Oligochaeta did not display any orientated
distribution pattern as a response to both period and location factors, being abundant at the end of
wet season and simultaneously at the saline water points. The Polychaeta Alkmaria romijni was the
dominant species and ubiquitous throughout sampling stations .Among environmental variables,
salinity was the most explaining abiotic variable of the community distribution pattern.
In the second experiment, six wells were selected for sampling in which four of them were more
associated to salinization risk as they are located close to Arade estuary, whereas the remaining wells
are relatively far from the sea. Wells fauna was sampled with the use of a phreatobiological net and
well sediment sampling device developed for the present study (WSSD). Wells were grouped into two
categories based on their salinity. A total of 612 number of individual were collected and 19 species.
were identified. For all the 6 wells a total of 240 species were identified as true groundwater fauna.
Identification to species level is still on going. The Order Cyclopodia dominated in all 6 wells with
relatively high diversity. There was an apparent relationship between salinity level of well water and
stygofauna presence. The species Eucyclops speratus, Eucyclops hadjebensis, Megacyclops viridis
and Acanthocyclops sensitivus were particularly associated with low salinity conditions, hence being
potential indicators for saline intrusion if their abundances decrease greatly. Conversely, the taxa
Halicyclops sp. was identified as a possible indicator of high salinity conditions. The greatest diversity
and highest abundance were found in the lowest salinity condition. The comparison between the two
sampling methods indicated that the Phreatobiological net is more effective in gathering
representative samples, particularly from Cyclopodia which are generally good swimmers, while the
WSSD net is more suitable to have sample groups adapted to benthic life such as Harpacticoids and
Ostracods.

5
Keywords: Benthic fauna, salinity gradient, groundwater habitat interface, climatic and human
pressures
Resumo
Este trabalho é composto por duas grandes linhas de investigação. A 1ª teve como objectivo
caracterizar a comunidade estuarina bentónica ao longo de um gradiente salino que recria as
condições de um ecossistema dependente de águas subterrâneas. A 2ª grande temática teve com
objectivo examinar a sensibilidade da stygofauna a variações na salinidade/condutividade eléctrica
através de amostragens em poços artesanais. Na experiência estuarina, as diferenças na estrutura e
composição de invertebrados bentónicos foram estudadas no canal adjacente ao canal principal do
estuário do Arade no Sul de Portugal. Este recebe uma contribuição importante e directa de água
subterrânea e simultaneamente é influenciado por marés semi-diurnas do estuário. Os
macroinvertebrados bentónicos foram amostrados no final dos períodos seco e húmido em 2009, ao
longo do gradiente salino criado pela entrada de água subterrânea no canal. Variáveis físico-
químicas foram medidas para determinar a sua associação à distribuição da fauna bentónica. Os
resultados indicam que a abundância do isópode Cyathura carinata e do polychaeta Heteromastus
filiformis variou significativamente com o gradiente de salinidade criado pela descarga de água
subterrânea para o habitat estuarino e com o período de amostragem. Os poliquetas foram mais
abundantes no final do período seco do que no húmido, e nos locais de amostragem de maior
salinidade, sugerindo uma associação com águas mais salinas. No entanto, os poliquetas Hediste
diversicolor e Alkmaria romijni foram mais abundantes em áreas de menor salinidade e no final da
época húmida. Alguns taxa como sejam os Oligoquetas não revelaram nenhuma tendência no seu
padrão de distribuição espacial ou temporal. O poliqueta A. romijni foi a espécie dominante e
distribuída por todas as estações de amostragem. De entre as vaiáveis ambientais medidas, a
salinidade foi a que mais contribuiu para explicar o padrão de distribuição das comunidades.
Na segunda linha de investigação, 6 poços artesanais foram seleccionados na mesma área de
estudo, em que 4 deles estão associados a risco de salinização por se encontrarem perto do estuário
do ri Arade, enquanto que os outros estão relativamente longe do mar. A fauna existente no fundo
dos poços foi amostrada com uma rede freatobiológica e com um aparelho de amostragem em poços
desenvolvido para o presente estudo (WSSD. Os poços foram agrupados em 2 categorias com base
na salinidade da sua água. No total, foram amostrados nos poços 612 organismos que representam
19 espécies. A ordem Cyclopodia foi a dominante em todos os poços com uma relativamente
elevada diversidade. Foi verificada uma aparente relação entre o nível de salinidade da água do poço
e a presença de stygofauna. As espécies Eucyclops speratus, Eucyclops hadjebensis, Megacyclops
viridis e Acanthocyclops sensitivus foram encontradas associadas a condições de reduzida
salinidade, sendo potenciais indicadoras de intrusão salina costeira se a sua abundância decrescer
acentuadamente. Por oposição, o taxa Halicyclops sp. foi identificado como um potencial indicador
de condições de mais elevada salinidade. A maior abundância e diversidade foram encontradas na
salinidade mais baixa. A comparação entre os dois métodos de amostragem indicou que a rede

6
freatobiológica é mais eficaz na recolha de amostras representativas, particularmente de Cyclopoids
que são geralmente bons nadadores, enquanto que o WSSD é mais adequado para amostragem de
grupos adaptados a vida bentónica como sejam os harpaticóides e os ostracódes.
Palavras de chaves: Fauna bentica, gradiente de salinidade, interface de habitat de águas
subterrâneas , Pressões humanas e climáticas

7
ContentsChapter 1 1.1 Introduction ................................................................................................................... 11 1.2 State of art ..................................................................................................................... 14 1.2.1 Groundwater/surface-water ecotones (transitional zone) .......................................... 14 1.2.1.1 Concept of groundwater, water table and flow system ......................................... 16 1.2.1.2 Recharge -discharge behaviour of coastal aquifers ............................................... 17 1.2.1.3 Groundwater dependent Ecosystems ..................................................................... 18 1.2.1.4 GW-surface water interaction and salinity variation in the wetland ..................... 19 1.2.2 Climate change and groundwater surface water variability ...................................... 20 1.2.2.1 Climate change impact and aquifer vulnerability in the Mediterranean area (study area): ……………………………………………………………………………………21 1.2.2.2 Biological response to climate change on aquatic ecosystems .............................. 22 1.2.3 Monitoring of water resources and dependent ecosystems using bioindicators ....... 22 1.2.3.1 Ecological assessment and environmental policies “The Water Framework Directive”…………………………………………………………………………………… 24 1.2.3.2 Benthic Macroinvertebrates as an indicators of ecological status ......................... 24 1.2.3.2.1 Benthic community response to salinity gradients in estuaries ............................. 25 1.2.3.3 Seasonal and spatial patterns of benthic invertebrates .......................................... 27 Chapter 2 2.1 Abstract ......................................................................................................................... 29 2.2 Introduction ................................................................................................................... 30 2.3 Methods......................................................................................................................... 32 2.3.1 Study area .................................................................................................................. 32 2.3.1.1 Hydrology and hydrogeology: ............................................................................... 33 2.3.1.2 Land use ................................................................................................................. 33 2.3.2 Sampling design ........................................................................................................ 33 2.3.2.1 Environmental variables measurement:................................................................. 35 2.3.3 Data analysis ............................................................................................................. 37 2.3.3.1 Spatio-Temporal variation ..................................................................................... 37 2.3.3.2 Relationships between environmental and biological variables ............................ 37 2.4 Results ........................................................................................................................... 38 2.4.1 Benthic macrofauna general characterization ........................................................... 38 2.4.2 Species distribution ................................................................................................... 38 2.4.3 Species contribution for spatial and temporal differences ........................................ 42 2.4.4 Environmental variables ............................................................................................ 43 2.4.5 Abiotic variables contribution for community distribution ....................................... 44 2.5 Discussion ..................................................................................................................... 46 2.5.1 Groundwater availability and community predictions .............................................. 46 2.5.2 Variation in groundwater discharge and food-web implications .............................. 47 Chapter 3 3.1 Abstract ......................................................................................................................... 50 3.2 Introduction ................................................................................................................... 51 3.3 Methods......................................................................................................................... 54 3.3.1 Study area: ................................................................................................................. 54 3.3.2 Field abiotic measurements ....................................................................................... 54 3.3.3 Sampling design: ....................................................................................................... 55 3.3.4 Wells fauna assessment ............................................................................................. 57 3.4 Preliminary Results and interpretation .......................................................................... 57

8
3.4.1 Fauna general characterization .................................................................................. 57 3.4.2 Species distribution ................................................................................................... 60 3.4.3 Sampling device comparison .................................................................................... 61 Chapter 4 4.1 Main Conclusions and future works ............................................................................. 64
Bibliography ............................................................................................................................ 66
IndexofFigures
Figure 1.1- GW-SW interact throughout all landscapes K; Karst; M: Mountain, R: riverine (small); C: Costal; G: Glacial; V: riverine (large)…………………………………………………15
Figure 1.2- Location of the saturated and unsaturated zones in relation to the water table and processes involved in the water movements (USCS)……………………………………...17
Figure 1.3- Remane curve (after Remane, 1934), showing quantitative relations between freshwater, brackish and marine invertebrate species…………………………………………26
Figure 2.1-The enlarged map the Algarve province and the main water courses (blue) and corresponding catchment area (orange line). The main area of Querença-Silves aquifer is shown by light blue area……………………………………………………………………………32
Figure 2.2-Sediment sampling points. First location on the right side of the bottom image corresponds to the GW discharge point. The salinity gradient is also marked by colours from the lowest point (blue) to the highest point (red)…………………………………………………34
Figure 2.3- Specific sampling locations. Point C refers to the location of GW discharge and Point E is located in the connection of the branching channel to the main estuary canal…..35
Figure 2.4- Estômbar channel , a branching channel of the Arade estuary at low tide(A), and the location of sampling points A,B, and C in low tide(B); the Estômbar channel in high tide(C) and location of groundwater input at high tide(D)……………………………………… 36
Figure 2.5-MDS ordination of fauna sediment samples collected at the end of the dry and wet times, in the distance gradient originating at the location of groundwater input (A -E)…39
Figure 2.6- Temporal variations on the densities of taxa that contributed the most on the dissimilarities between periods…………………………………………………………………….40
Figure 2.7- dbRDA ordination of the sample variation during wet (Wet) and Dry (Dry) season and in five locations (A, B, C, D, E) for the abundance of species ……..……………………. 42
Figure 2.8- Variation of water level, electrical conductivity (EC) and temperature at the location of groundwater discharge into the Estômbar channel of the Arade estuary; also indicated are the tidal fluctuations measured in Lagos, located 20 km towards the west…..43

9
Figure 2.9- PCO analysis of biota distribution accordingly to sampling points with overlaid vectors of environmental variables contribution to biota distribution…………………………..45
Figure 3.1- Location of the inventoried wells. Wells number 2,3, 22,23,25 and 26 were sampled………………………………………………………………………………………………54
Figure 3.2- Phreatobiological net and its deployment………………………………………… 55
Figure3.3- Well Sediment Sampling Device and its deployment………………………………56
Figure3.4-some representive Stygofauna, sequently from left up to right down: gamarus pulex/Eucyclops serrulatus serrulatus / Cypridopsis vidua/ Harpacticoida/ Macrocyclops albidus/ Eucyclops / Megacyclops Viridis/ Gammarus pulex…………………………………..60
Figure 3.5-MDS ordination of wells fauna samples collected from 6 different wells (Q1-Q6), with three replicate from each well (R1-R3)grouped into two groups of salinity categories (H: high and L:low salinities)…………………………………………………………………………. 60
IndexofTablesTable 2.1- Permanova analysis of the sediment fauna for factors Period (dry and wet) and Location (A-E). The number of permutations used was 9999 (Ti=period, Lo=Location)……38
Table 2.2- SIMPER analysis identifying the species that contributed the most for differences between seasons for all locations…………………………………………………………………39
Table 2.3. SIMPER analysis identifying the taxa that contributed the most to differences from point D to the freshwater point (C )…………………………………………………………41
Table 2.4- SIMPER analysis identifying the species that contributed the most for dissimilarities between point D and point B……………………………………………………...41
Table2.5. Biota and environmental matching according to the BEST modelling…… …..…..44
Table3.1-. Wells abiotic and morphologic characteristics. Wells numbers 1, 2,3,4,5 and 6 refers to wells illustrated in “figure 1” by codes 2, 3, 22, 23, 2 and 26 respectively. n.a.-not available……………………………………………………………………………………………...54
Table 3.2- Identified species of Cyclopodia, Ostracod, Amphipod and Copepod sampled rom wells. Shaded cells represent absence and unshaded cells represent a taxa absence…….57
Table 3.3- Taxa abundance sampled by Phreatobiological net and WSSD…………………60
Table 3.4- SIMPER analysis identifying the species that contributed the most for differences between wells with high and low salinities………………………………………………………..61
Table3.5- Taxa abundance sampled by Phreatobiological net and WSSD……………….…61

10
Chapter 1: General introduction and state of art

11
1.1 Introduction
Habitats located at groundwater–surface water interfaces, i.e. transition zones, are considered to be
spatially and temporally dynamic (e.g. Malard et al., 2003; Bork et al., 2009). These transition zones
often display complex temporal and spatial variation on benthic macroinvertebrates community
(Rundle et al., 1998; Bork et al., 2009). Ecotones are characterized as dynamic components of a
landscape, providing habitat for many transient organisms that explore highly shifting environments
(Senft, 2009). Transitional zones in the estuary may have lower diversity in terms of benthic
communities when compared to freshwater and higher salinity areas, due to natural stress (e.g.
salinity variation)(Medeiros et al., 2011). The conservation of these habitats offers some challenges
because ecotones may appear to have lower diversity if they undergo through frequent or intense
disturbance events (Chapman, 1960 and Senft, 2009). Groundwater and surface water are connected
in a hydrological continuum and often originate transient zones, which provide refuge conditions, sites
of high biodiversity and habitat for the macrofauna, microbial production, and energy transfer
(Tomassoni, 2000). Climate is an important factor affecting water quality and availability in the
interaction zone. The change in the precipitation and temperature future regimes, induced by climate
change, will lead to change in the runoff, aquifer recharge, flood and drought frequency and
magnitude, as well as in the quality of the water resources (Santos et al., 2002; Ajami et al., 2008).
Groundwater resources are currently under severe threat due to Human usage and pollution
(Danielopol et al., 2003). As depletion of superficial water occurs and because climate change effects
are predicted to greatly enhance the demand on usable freshwater, groundwater is thought to be the
primary provider meeting the Human multiple demands on this limited resource (Danielopol et al.,
2003; Santos et al., 2002). Substantial aquifer exploitation threatens the wetlands that are also
groundwater dependent ecosystems (GDEs) (Boulton, 2009; Humphreys, 2009). In the south regions
of Portugal, where the present study was made, groundwater represents 60% of drinking water and
fulfils 80% of agricultural demand (Stigter et al., 2009). This demand is likely to be enhanced in this
geographical area as a result of global warming (Santos et al., 2002).
The benthic invertebrate community has been considered a very useful tool to monitor and assess the
Human induced impact on the aquatic environment due to their measurable response to natural shifts
and anthropogenic impacts (Chainho, 2008). Benthonic macroinvertebrates are identified by the
Water Framework Directive (WFD) as useful bioindicators of the ecological status of water bodies
(2000/60/CE) (Sandin and Hering, 2004; Borja et al., 2009, Silva et al., 2012). Many species of the
benthic community such as Polychaeta but not amphipods, show reduced mobility and in case of
contamination, they will be exposed to the stress for longer time periods, hence therefore they can
reflect also the effects of long term environmental disturbances (Chainho, 2008). In the context of
habitat management, biological monitoring methods present several advantages: biological response
reflects the conjugated action of environmental conditions and make the impact easily detected,
evaluate factors not directly measurable such as biological complexity and ecological value and, no
expensive laboratorial chemical analysis are required (Ambrogi and Forni, 2004, Silva et al., 2012).
Some biological characteristics of benthic species have to be taken into account when interpreting

12
results of benthic community as a whole. Benthic fauna show high spatial heterogeneity related to
their tolerance to the influence of different environmental factors such as salinity, sediment type,
temperature, etc. Moreover, invertebrate communities also show important temporal variations related
to seasonal and interannual variation. Part of the seasonal fluctuation is associated with their
biological cycle due to recruitment processes that occur during spring and autumn for most species,
but also due the control of extreme climate event such as low temperatures, floods and droughts
(Alden et al., 1997; Attrill and Power, 2000; Salen-Picard and Arlhac, 2002; Chainho, 2008).
Moreover the organisms living within the shallow groundwater zone can serve as indicators of the
quality of the groundwater resource, particularly at the interaction and influence area of the surface
water systems (Claret et al. 1999). In fact, groundwater fauna reflect structural conditions of their
habitat such as hydraulic conductivity, heterogeneity of habitats in an aquifer and provide information
on surface/subsurface hydrological exchanges (Danielopol et al., 2007). Therefore, the relative
presence or absence of different communities or populations of organisms may reflect the impact of
changes in water quality, in similarity with the bio indicator function that many surface taxa display.
The CLIMWAT project aimed at evaluating the climate change impacts on the Querença-Silves
aquifer, which is a costal aquifer with dependent ecosystems , and one of eight transnational
research projects funded under the CIRCLE-MED network (Stigter, 2011). Querença-Silves was the
aquifer studied in the present thesis which was developed within the project. Different climate
scenarios were used to predict the future climate change impact on aquifer net recharge and
consequently the output to GDEs and water quality condition in different wells. Results from the
project indicated a significant decrease in recharge in the present study area for the future years. The
models calculated in the project, predict a significant increase in the mean temperature for future
years and, consequently it is expected an increase groundwater demand for crop cultures. Due to
increased extraction of aquifer water and less recharge flow into the aquifer, an overall decreasing
trend of groundwater levels is foreseen, which is likely to reduce the groundwater outflow into coastal
wetlands, threatening the ecosystem stability. Climate change has the potential to greatly influence
interface or border aquatic habitats such as estuaries and wetlands that are dependent on
groundwater, by altering environmental variables such as temperature, salinity, etc. (Bates et al
2008). The combined effect of prolonged and large extractions from the aquifer and reduced recharge
in coastal areas can lead to seawater intrusion, a serious problem worldwide, including the
Mediterranean countries.
This study follows two experiments with different objectives and study site. The first experiment was
made in an estuarine branch which receives groundwater from the coastal aquifer and brackish tidal
water originated from the main estuary channel. In fact, the branching channel was located
approximately half-way between the river and sea points of the estuary. It represents an interface
area between the main estuary channel and a point of considerable groundwater direct discharge
from the Querença-Silves aquifer into the estuary. This interface ecosystem is a perfect case study to
evaluate the estuarine benthic community response to changes in groundwater input from aquifers.
The main objective of the present study was to test the hypothesis of there being a relationship
between the distribution pattern of benthic estuarine taxa and the groundwater availability in the

13
estuary. The present study also aimed to identify benthic estuarine taxonomic groups and/or species
which can potentially be monitored to ecological impacts of changes in groundwater discharge. It was
expected that benthic communities responded in presence, abundance and potentially in population
structure, to the gradient in salinity originated by the freshwater input into the estuary, due to
discriminating salinity tolerances of organisms and species.
The second experiment aimed to examine the sensitivity of stygofauna to variation in groundwater
salinity/conductivity accessed through wells. Assessing the biodiversity and abundance response of
groundwater fauna sampled in aquifers of differing groundwater characteristics allows evaluating
impacts such as seawater intrusion on the vulnerable groundwater ecosystem. This experiment also
aimed to provide ground breaking biological data to integrate the use and conservation of
groundwater dependent fauna into aquifer management. Six wells were selected for sampling in
which four of them were more associated to salinization risk as they are located close to Arade
estuary, whereas the other two wells are relatively far from the sea. Wells fauna was sampled with the
use of a phreatobiological net developed for large diameter wells, as well as a well sediment sampling
device developed for the present study (WSSD).
This thesis was divided in four different chapters as it follows:
Chapter 1 is a general introduction that summarises the two experiments with their respective
objectives. It also contains the state of art in which some key themes of the study will be
explained by details.
Chapter 2 consists of an experiment that identifies Spacio-tempral variation of benthic
communities in GDEs influenced by climate change. Chapter 3 consists of second experiment in which stygofauna were used for assessing
groundwater with different conductivity condition.
Some final remarks are presented in Chapter 4, integrating the results obtained in each
chapter and presenting the major conclusions of the study.

14
1.2 Stateofart
Freshwater ecosystems fed by groundwater or/and surface water, have been important sources for
the development of environmental monitoring programmes (De Pauw et al., 1992). The management
of the freshwater ecosystem needs to integrate the groundwater exploitation, as well as surface water
bodies and ecosystems. Wetlands are characterized by large land-water interfaces that regulate the
ecological status of ecosystems established there (Winter, 2003; Euliss et al., 2004). Wetlands that
are linked to aquifers also include surface-groundwater interfaces and are referred to as groundwater
dependent systems (GDEs). A wide range of disturbance sources including pollution may affect the
groundwater quality and availability (Danielopol et al., 2003). Climate change has direct impacts on
the availability, timing and variability of the water supply and demand, and is also related to the
significant consequences of these impacts on many sectors of our society. Bioindicators sensitive to
climate variability impacts on coastal waters (and ecosystems) quantify the impacts of climate change
on water quantity and quality. For this study, ecosystems have been studied with particular attention
for the ecotones where groundwater and surface water bodies interact. Assessing the bioindicators
representing different ecotones and trophic levels (e.g. invertebrates, vegetation and respective
predators) enables to integrate pollution effects at different spatial and time scales. Indicators of
ecosystem health improve the efficiency of ecosystem management such as that of southern
wetlands in Portugal, where climate changes are predicted to enhance eutrophication and droughts
effects. In a groundwater-brackish water ecosystem with high instability in environmental factors such
as salinity and conductivity, invertebrates are expected to respond, and therefore could be used as
potential important tools in the assessment and management of surface and groundwater availability
under different climate change scenarios and uses. The following review section analysis the current
knowledge of the role that groundwater–surface water (GW–SW) interactions play in the ecology of
arid/semi-arid wetlands, particularly those which are dependent on groundwater inputs. The key
themes of the review are as follows: (i) Groundwater/surface-water ecotones, (ii) Impact of climate
Change on water resources particularly in interaction zones and on the ecosystems depending on
groundwater input and, (iii) Application of biological indicators of climate change and anthropogenic
impact in the mentioned ecosystems.
1.2.1 Groundwater/surface‐waterecotones(transitionalzone)
Ground and surface waters are usually evaluated as separated water masses, but hydraulically they
are connected and the groundwater contacts and feeds all types of surface water aquatic habitats
such as lake, stream and wetland in different terrains from mountain to ocean (Fig.1.1) (Winte, 2000).

Figure 1.1
In the pr
water ty
measure
mixture.
with a h
2009). M
such as
changes
et al., 2
1‐ GW‐SW inter
resent study
ypes, such as
e the water
The transitio
high heteroge
Moreover, ch
s daily cycle
s, and long-t
2000). Unde
ract throughou
, the ecotone
s groundwat
type intera
on zone betw
eneity in spa
hemical and
es (e.g., tem
term climatic
erstanding th
ut all landscape
riverine (la
e concept re
ter/surface w
action since
ween surface
ace and time
biological pr
mperature a
c changes an
he interactio
es K; Karst; M: M
arge) (Winter e
efers to the tr
water and fre
many natur
e and ground
e (Boulton e
rocesses in t
nd transpira
nd events (s
n between
Mountain, R: ri
et al., 2000)
ransition zon
eshwater/salt
ral and anth
dwater is cha
t al., 1998; M
this zone ope
ation), short-
uch as extre
groundwater
iverine (small);
ne between tw
twater. It is c
hropogenic a
aracterised b
Malard et al
erate at diffe
-term weath
eme weather
r and surfac
; C: Coastal; G:
two water bo
considered d
activities aff
by dynamic g
., 2002; Bro
erent tempor
her events,
r events) (To
ce water ca
15
Glacial; V:
odies and
difficult to
fect their
gradients
ke et al.,
ral scales
seasonal
omassoni
an be an

16
important tool for water resources management, aiming to preserve the ecosystem stability and
determine the migration pathways of contaminants (Winter et al., 2003). The movement of surface
and groundwater is controlled by a natural condition of an area such as climate, through the effects of
precipitation and evapotranspiration that affects the distribution of water to—and removal from—
landscapes. The precipitation distribution pattern is highly variable in space and, evaporation and
evapotranspiration also vary under different climatic conditions, often even showing differing patterns
in a relatively small spatial scale from forest to nearby urban area. The physiography of the area such
as land-surface form and geology are also parameters controlling the groundwater and surface water
movement (Winter et al., 2003). Therefore, it is necessary to understand the effects of physiography
and climate on surface water runoff and groundwater flow systems, taking into account the impact of
human activities on their functioning and thus, on ecotones management policies. The mixture of two
water bodies in an ecotone may have major impacts on the individual ecosystems, if major
environmental factors such as temperature, acidity or dissolved oxygen are altered (Winter et al.,
2003). Due to the water interchange between surface-groundwater bodies, any shifts or
contamination of one commonly affects the other one. Surface waters are more vulnerable to pollution
due to their easy accessibility for disposal of wastewaters. Both the natural processes, such as
precipitation inputs, erosion, weathering of crustal materials, as well as the anthropogenic influences
such as urban, industrial and agricultural activities and exploitation of water resources, determine the
quality of surface water in a region. In turn, groundwater recharge is currently and in the future
predicted to be altered as a result of climate change and anthropogenic impacts (Ajami, et al., 2008).
Transitional zones are also ecologically important because some surface and groundwater organisms
have a life stage dependent on this zone, hence being vulnerable to any habitat shift. Ecotones
established at surface/groundwater or fresh/saltwater interfaces are flexible to changes in water mass
fluxes and have intermediate biodiversity (Vervier et al., 1992), but less is known of the unique
species that permanently inhabit in the transition zone and many have not been described.
Ecologically, the ground-water/surface-water transition zone is an important ecosystem affecting
several trophic levels from microbes to fish (Bear et al., 1979).
1.2.1.1 Conceptofgroundwater,watertableandflowsystem
Groundwater is found underground in the unsaturated and saturated zones. In the unsaturated zone,
the voids which represent the space between the soil grains are filled by air and water. The water
present in this zone does not have the potential to be easily used because it is not feasible to pump
this water from the wells, mostly due to capillary force holding the water into the soil. The “soil water
zone” occurs at the top of the unsaturated zone. This layer is typically crossed by voids created by
roots (live and decaying), animal and worm burrows, all which will increase the water infiltration into
the soil. Soil water is used by plants and goes back to atmosphere through their transpiration or
evaporation directly from the soil. On the contrary, the saturated zone is completely filled with water
and the water in this zone is referred as groundwater. The upper section of the saturated zone is
referred as water table (Fig.1.2) (Winter, 2000).

17
Saturated ZoneBelow the water table
(Ground Water)
Capillary fringe
Soil zone
Un
satu
rate
d z
on
e
Recharge to water table Water table
Evaporanspiration
Precipitation
Figure 1.2‐ Location of the saturated and unsaturated zones in relation to the water table and processes involved in the water movements
The water table meets the surface water at or near shorelines if the surface water body is connected
to groundwater systems. The depth to the water table is variable in different landscapes but is smaller
near permanent surface water bodies such as lake and wetlands. The water table configuration varies
seasonally because the recharge into the saturated zone is affected by the precipitation pattern that
also depends on seasons. The groundwater flow system can operate at the local, intermediate and
regional scales. The most dynamic and shallowest flow is the local flow and therefore more in
interchange with the surface flow. The deeper flow systems such as those at the regional and
intermediate scales reside longer underground and, are more in contact with subsurface material, so
that when they discharge into the surface water, it has augmented effect on the chemical
characteristics of the interaction zone. Three types of flow regime can be described for the GW–SW
ecotone, which occurs mostly in wetland ecosystems: (i) recharge: surface water penetrates into the
underlying aquifer; (ii) discharge: surface water gains water from the underlying aquifer; or (iii) flow-
through: the surface water gains water from the groundwater in some locations and loses it in others
(Winter, 2000).
1.2.1.2 Recharge‐dischargebehaviourofcoastalaquifers

18
The most dynamic boundary of great part of the groundwater flow systems is the water table. The
location of the water table changes continually in response to recharge and discharge from the
groundwater system (Winter, 2000). Changing meteorological conditions (e.g. precipitation) strongly
affect aquifer recharge, especially near the shoreline. The water table commonly intersects the
terrestrial surface at the shoreline, resulting in no unsaturated zone at this point. Because the water
table is near the surface adjacent to shoreline, the precipitation passes rapidly through a thin
unsaturated zone and recharges into the aquifer, potentially resulting in increased groundwater inflow
to surface water bodies. Transpiration by near-shore plants has the opposite effect, because plant
roots can penetrate into the saturated zone, allowing the plants to transpire water directly from the
groundwater system. The transpiration effect is very high in some areas whereby there is a
groundwater movement into the surface water during the night, and surface water discharge into
shallow groundwater during the day. These periodic changes in the direction of flow also can take
place on longer time scales such as, seasonally or annually. Recharge to the aquifer from
precipitation predominates during wet periods, and removal by transpiration predominates during dry
periods. As a result, the two processes—together with the geologic controls on seepage distribution—
can cause flow conditions at the beds of surface water bodies to be extremely variable. These
processes probably affect ecosystems depending on groundwater discharge more than other
ecosystems (Winter, 2000).
Costal and riverine wetlands have a relatively complex hydrologic pattern because they are subjected
to water level changes mostly due to tidal flow. The costal wetland is also likely to be affected by the
periodic tidal cycle, as well as the changes in water level due to seasonal changes in precipitation.
These combined changes, along with precipitation, evapotranspiration and surface-ground water
interactions, determine the water quality and availability in the wetland ecosystems. Among the
wetland, the present work focuses on coastal wetlands that depend on freshwater input mostly
originating from groundwater. These bordering wetlands ecosystems can more directly reflect the
consequences of any shifts in the aquifer dynamics (Day et al. 2008), constituting the so called
groundwater dependent ecosystems (GDEs). Shifts in the chemical and ecological status of these
GDEs may occur as a consequence of changes in groundwater availability and/or quality related to
climatic change effects (Danielopol et al., 2003; Eamus and Froend, 2006), hence affecting all
wetland biotic elements including vegetation, invertebrates, fish and birds.
1.2.1.3 GroundwaterdependentEcosystems
Groundwater dependent ecosystems (GDEs) are ecosystems that require the input of groundwater to
maintain their ecological structure and function (Murray et al., 2003; Murray et al., 2006). One of the
GDEs forms is wetlands which can be dependent on groundwater for all or part of the year.
Groundwater is essential to many ecological communities as a connector, not just in the aquifer itself,
but within, across, and between surface waters and many terrestrial ecosystems (Boulton, 2009).
Increasing awareness of groundwater issues worldwide are creating more opportunities to understand
how groundwater and their GDEs respond to altered hydrological regimes. Indeed, groundwater

19
resources have been threatened due to increased demands on groundwater for consumption,
industry and agriculture. These demands alter groundwater regimes of GDEs that have evolved over
millennia, resulting in the degradation of ecosystem health. As a consequence, the goods and
services (ecosystem services) that GDEs provide for humans, which include food production and
water purification, are at serious risk of being lost (Murray et al., 2003). Groundwater resource
managers commonly ask how much water can be taken from the aquifer while still maintaining a low
level of risk to GDEs. This requires quantified information on the relationship between groundwater
flux quality and pressure on biota and ecosystem processes of GDES (Eamus and Froend, 2006).
Groundwater exchange influencessurface ecology due to the multiple services it provides including
means of delivering oxygen and food to microbial and invertebrate communities. According to
Malcolm et al. year 2005 dissolved oxygen varies over very fine spatial scales in groundwater
exchange zones influencing salmon ecology. Thus, the areas of upwelling cool groundwater could
provide refugia for brown trout during summer (Hancock and Boulton, 2009). Ecologists have come to
recognise that many stream ecosystems in surface, subsurface, and lateral compartments which are
linked by hydrological exchanges, convey nutrients, organic matter, dissolved oxygen, and some
invertebrates among these zones (Boulton et al., 1997).
Therefore, declining groundwater quantity or quality and variation in groundwater-surface water
interaction will influence the ecological communities. The ecological, hydrological, and physical–
chemical links between groundwater, surface waters and associated ecosystems are seldom fully
understood even though true characterization and wise management will require a multidisciplinary
approach. This means biologists need to understand the importance of magnitude and timing of
groundwater flows for the system, integrated with hydrology background knowledge. For collaborative
research to improve our understanding of links between groundwater and ecosystems, it needs to
focus on spatial and temporal scales that are relevant to both the hydrogeological and the ecological
process being studied. Effective management of GDEs and their ecosystem services requires
prioritisation of the most valuable ecosystems, given that increasing human demands and limited time
and money prevent complete protection of all GDEs.
1.2.1.4 GW‐surfacewaterinteractionandsalinityvariationinthewetland
Precipitation is one of the most dominant sources of water in nearly all wetland systems, yet the
influence of the groundwater flow component into the ecosystem can be sufficient from an ecological
perspective to yield an entire new type of wetland. Influxes of groundwater to lakes, rivers, and
wetlands can change whole-system physical–chemical properties such as temperature and salinity,
and also influencing microenvironments and their ecological processes (Boulton, 2009). Infiltration of
water from surface aquatic ecosystems and rainfall can have a significant effect on aquifer ecology;
therefore whether the water is flowing into or out of an aquifer, or is moving from one part to another,
it is the extent and intensity of connectivity that often determines its importance to ecosystems.
Coastal wetlands receiving tidal brackish water and groundwater, can range in salinity from sea water
(>30,0) to freshwater (0-0.5) (Redeke,1935).Salinity varies according to freshwater discharge, thus

20
there are many differences between estuaries with different climatic regimes. In arid/semi-arid
environments, such as in the south part of Portugal examined in the present study, rainfall is seasonal
and significantly less than the evaporation rate. Hence, groundwater discharge into surface wetlands
can be a major component of the water quality and salt balance, which are major determinants of
wetland ecology. In semi-arid regions, the salinity in wetlands environments varies naturally due to
high evaporative conditions, infrequent rainfall, groundwater inflows, and after rains or floods (Jolly et
al., 2008). As a result of the human activities, including changes in land use, surface water regulation
and water resource availability, wetlands in arid/semi-arid environments are now often experiencing
extended periods of high salinity (Jolly et al., 2008). For the present study, the location where the
groundwater flows into surface systems is located in a channel connected to an estuary. The surface
water body is brackish closer to the estuary while it is less saline closer to the groundwater discharge
area. The canal is influenced by tidal brackish water daily, hence even in the point where groundwater
discharges into the wetland, the influence of the brackish water is high. When the groundwater inflows
increase, due to raised groundwater levels originated by factors such as land use change and river
regulation, this may have a major influence on the ecology of a wetland and its surrounding areas. On
the contrary, if the groundwater input to the wetland decreases, a scenario predicted for the region
(CLIMWAT Results), it can be expected that the ecosystem may alter strongly in the point with more
freshwater input. There are many knowledge gaps, particularly related to time-series data on the
salinity tolerance/sensitivity of freshwater aquatic biota and riparian vegetation.
1.2.2 Climatechangeandgroundwatersurfacewatervariability
It is expected that predicted global changes in temperature and precipitation will alter regional
climates and hydrologic regimes in most areas of the world. The change in the precipitation and
temperature regimes, induced by climate change, will lead to changes in the runoff, aquifer recharge,
flood and drought frequency and magnitude, as well as in the quality of the water resources in
Portugal (Santos et al., 2001). In assessing the impacts of climate change on water resources, most
research has been directed at forecasting the potential impacts to surface water hydrology. For
groundwater hydrology, large regional and coarse-resolution models have been used to determine the
sensitivity of groundwater systems to changes in critical input parameters, such as precipitation and
runoff ( York et al., 2002). Although the effect of temporal variability in surface waters is more visible,
groundwater recharge is likely to be altered as a result of climate change and anthropogenic impacts
(Ajami, et al., 2008), especially in areas where it is more sensitive to climate variability such as
Mediterranean areas. Groundwater resources in the studied area, south of Portugal, are under
increasing pressure due to large extraction rates for various water-consuming activities, particularly
agriculture (irrigation), public water supply (consumption) and industry. Among the aquatic
ecosystems, wetland systems are more vulnerable to changes in quantity and quality of their water
supply, and it is expected that climate change will have a pronounced effect on wetlands through
alterations in hydrological regimes with great global variability (Erwin, 2009). Many wetlands depend
on groundwater flux throughout annual and seasonal weather changes, which direct to couple change

21
on hydrological system of both water bodies. Because wetlands water level variation has an impact
on the surface flow and groundwater recharge. Substantial aquifer exploitation threatens the wetlands
that constitute groundwater dependent ecosystems (GDEs), and in coastal areas it can lead to
seawater intrusion, a serious problem worldwide, including the Mediterranean countries (Stigter et al.,
2009). It should be noted that intensive and uncontrolled groundwater exploitation can have similar or
more severe impacts on aquifers and related ecosystems than climate change. A correct
implementation of future adaptation measures requires a more detailed insight into the way climate
change affects aquifer recharge and discharge patterns to mitigate the expected disturbance
1.2.2.1 ClimatechangeimpactandaquifervulnerabilityintheMediterraneanarea(studyarea):
Climate change is predicted to be very noticeable in the Mediterranean region, due to the magnitude
of expected changes in temperature and rainfall patterns (Giorgi, 2006). Aquifers located in these
regions (e.g. Querença-Silves) are expected to be affected by climate change, particularly in arid and
semi-arid regions where decreases in recharge can become very significant in the following decades
(e.g.,Santos et al. 2002, Giorgi 2006). The aquifers may be more vulnerable to climate change if they
are located in regions where many Human sectors depend on them for their water supply. Drought
and increased water demand for agricultural activity affects the availability of surface water and will
lead to increased groundwater usage. Portugal is characterized by its mild Mediterranean climate,
with well-known water vulnerability to climate fluctuation, namely to droughts and desertification in the
southern sector (Santos et al., 2002). Severe droughts have occurred in the study area namely during
the hydrological year of 1991/92 and also in 2005 (Barros et al., 1995; Stigter, 2011). Climate change
may particularly aggravate this problem in the Mediterranean region (e.g. Giorgi 2006), due to the
combined effect of rising sea levels and reduced recharge of aquifers associated with the expected
decrease in precipitation and average temperature increase. In the CLIMWAT project it was examined
the effect of climate change in the present case study. Climate scenarios were calculated using the
available scenarios from the ENSEMBLES project as a starting point .The data of temperature and
precipitation for years 1960-1990 or 1980-2010 were used as a reference period, and aquifer
recharge was calculated for 2020-2050 and 2069-2099. Results for the calibrated period and
predicted climate scenarios indicated that discharge from the Querença-Silves aquifer into the Arade
estuary are to decline and therefore, it is modelled an increase in probability of occurrence of
seawater intrusion and the drying out of groundwater dependent wetlands. Continued climate stress
would increase the frequency with which ecosystem thresholds would be exceeded, leading to
chronic water-quality changes (Murdoch et al., 2000; Loáiciga et al., 2000).

22
1.2.2.2 Biologicalresponsetoclimatechangeonaquaticecosystems
Variability within abiotic processes influences ecosystem properties. Continued climate changes can
threaten a large number of unique biological systems (Smith et al., 2001). While ecosystems have
coevolved with these abiotic disturbances and biotic disturbances such as insects, disease, and fire,
changes in these disturbance patterns at rates faster than ecosystems can evolve could potentially
affect ecosystem regeneration and resilience (Coulson and Joyce, 2006). Some climate change
effects projected on aquatic resources are expressed in a small and slow environmental response
such as change in surface runoff, while other changes like extreme drought events are likely to
exceed the ecosystem threshold and cause a drastic switch in ecosystem biota (Prowse et al., 2006).
Climate change will potentially alter physical and chemical parameters at the landscape scale, and
are very likely to affect aquatic community and ecosystem attributes (Wrona et al., 2006). Climate
change is expected to have effects on benthic macroinvertebrate communities’ parameters such as
species richness, biodiversity, range, and distribution, and as a result, alter corresponding food web
structures and primary and secondary consumers levels, such as aquatic birds and mammals (Wrona
et al., 2006). The magnitude and extent of the ecological consequences of climate change in
Mediterranean freshwater ecosystems will depend largely on the rate and magnitude of change in
primary environmental drivers such as temperature, precipitation and alterations in water quality
properties such as salinity and nutrient levels (Poff et al., 2002; Wrona et al., 2006).
1.2.3 Monitoringofwaterresourcesanddependentecosystemsusingbioindicators
In an ideal situation, quality control in aquatic systems should be assessed by the use of physical,
chemical and biological parameters in order to provide a complete spectrum of information for
appropriate water management, however, in many cases the focus is on chemical parameters. Yet,
direct chemical analyses of water and sediment, which are usually very sensitive and accurate, do not
necessarily reflect the actual ecological state (Simboura and Zenetos, 2002). The history of bio-
indicator as tools for environmental monitoring started more than a century ago by Kolenati (1848)
and Cohn (1853) for surface water quality assessment, when they observed that organisms occurring
in polluted water are different from those in clean water(Iliopoulou-Georgudaki et al., 2003). At the
European level, the development of bioindicators, as a tool for the knowledge of the environment and
hence the protection of biological diversity of coastal and marine ecosystems has been progressed
through the implementation of the HABITATS directive, the biological quality elements of the Water
framework Directive (WFD), the Integrated Coastal Zone Management (ICZM) and others (Simboura
and Zenetos, 2002). Moreover, New European rules (see Directive Proposal 1999/C 343/01, Journal
of the European Communities 30/11/1999) emphasize the importance of using biological indicators to
establish the ecological quality of European coasts and estuaries (Borja et al., 2000). Ecological
assessment based upon the status of the biological elements, consideres frequently phytoplankton,
macroalgae, angiosperms, benthic macroinvertebrates and fish. The Ecological status (ES) of a water

23
body is determined by comparing data obtained from monitoring networks (Current status of
macroinvertebrate) with reference (undisturbed) conditions (Borja et al., 2009; Ferreira et al., 2007).
Parameters of the biological quality elements that must be included in the Ecological status
assessment of a water body are described in the European Water Framework Directive, e.g. for the
marine macroinvertebrate community the elements include composition and abundance of
invertebrate taxa and the proportion of disturbance-sensitive and tolerant taxa.
Moreover, chronic long-term contamination at low concentrations may be not detected through direct
measurements of chemicals in water mass, but may have effect on the biota. Biodiversity and
community composition measures for singular species (functionality) may provide information not only
on the current state, but also at a “time integrated” state of the system (Ambrogi and Forni, 2004). A
representative indicator needs to be: (1) applicable in many areas and scales of measurements, (2)
repeatable and reproducible by others besides its authors, (3) sensitive to pressures acting on the
system, responding in a predictable manner, but be relatively insensitive to expected (natural)
sources of interference, (4) operationally easy to collect data , (5) representative of the changes that
can be mitigated through a correct management, (6) integrative and cover key ecological gradients,
(7) scientifically reliable, and (8) economically feasible and the benefits of the information provided by
the indicator outweigh the costs of usage (Chainho, 2008; Dale and Beyeler, 2001; Niemeijer and
Groot, 2008). The use of bioindicators in the assessment of environmental quality in GW-SW
ecotones represents also a very useful tool that was applied in the present study. The flow between
groundwater and surface water create a dynamic ecotone habitat for aquatic fauna in interface area.
Saturated, interstitial subsurface (like hyporheic zone in river beds) zone below many gravel-bed
streams harbours a diverse fauna of invertebrates including benthic surface water fauna as well as
obligate hyporheic fauna (Williams 1984). The distribution patterns of invertebrates in GW-SW
interaction area apparently correlate strongly with water chemistry and hydrological exchange with the
surface stream and groundwater (Boulton, 1997). Assessing the existing species in surface water and
groundwater may indicate the state of respective water bodies regarding water quality, and can
function as indicators of groundwater originated changes in wetland status. The integration of
bioindicators representing different ecotones and trophic levels enables to integrate pollution effects
at different spatial and time scales. They may indicate the resilience to over abstraction of water,
efficiency of restoration practices, and impacts of artificial water-recharge, thus improving the
management of southern wetlands where climate changes shall enhance effects of eutrophication
and periodic droughts. In ecosystems with tidal and brackish estuarine influence ( temperature and
salinity), particularly those which have the influence of groundwater such as the case study of the
present work, the necessity of developing biological conservation strategies in ecosystem is high.
Due to the increasing anthropogenic impact partly revealed as climatic variability in such an
ecosystem, the development of rapid tools for estuarine environmental monitoring is currently highly
desirable for Portuguese estuaries (Santos et al., 2002).

24
1.2.3.1 Ecologicalassessmentandenvironmentalpolicies“TheWaterFrameworkDirective”
The European Water Framework Directive (WFD, 2000/60/EC) establishes a working platform to
protect aquatic ecosystems. The Biological Quality Elements (BQEs) designed by the WFD for
assessing the ecological status in coastal waters, include phytoplankton, macroalgae, angiosperms
and macroinvertebrates. Among the biological quality elements for the definition of ecological status in
coastal waters in WFD are the composition and abundance of benthic invertebrate fauna (Simboura
and Zenetos, 2002). The application of the WFD has encouraged scientists to work on the design of
different methodologies for assessing ecological status (Díeza et al., 2012). The main objective of
WFD is to achieve a ‘good ecological status’ for all waters by 2015. The successful implementation of
this directive depends on an integrated approach to water problems, supported by some fundamental
requirements including: (1) a single approach of water protection for all water categories, including
surface and groundwater, (2) achieving Good status for all waters by a set deadline, (3) apply water
management based on river basins, (4) a combined approach of emission limit values and quality
standards, (5) using water pricing as an incentive for better use, (6) getting citizens involved more
closely and, (7) streamlining legislation.
The assessment of ecological status requires the development of adequate tools, based on the
identification of surface and groundwater types, the definition of type-specific reference conditions,
and the classification of all water bodies within five ecological quality classes. A common
implementation strategy for the WFD was agreed between European Member States and several
working groups developed guidance documents on different aspects of the WFD, including the
assessment of ecological status in transitional waters (i.e. estuaries). The integration of biological
criteria in the assessment and definition of water quality standards was one of the major changes
introduced by the WFD to European legislation on water issues. As pointed out by Dauer (1993), the
use of biological elements is very important because (1) they are direct measures of the condition of
the biota, (2) they may uncover problems undetected or underestimated by other methods, and (3)
such criteria provide measurements of progress of restoration efforts. However, biological criteria
should not replace toxicity and chemical assessment methods, but complement the information
produced by those, serving as independent evaluations of the quality of marine and estuarine
ecosystems (Dauer, 1993). Although different methods can be used by different countries to classify
the ecological status, the classifications have to be comparable (Chainho, 2008).
1.2.3.2 BenthicMacroinvertebratesasanindicatorsofecologicalstatus
Benthic fauna, through the long history of Mediterranean research, have been often used as
indicators for assessment of the habitat quality or biological integrity which can be reliably used for the
classification of coastal and transitional water bodies. This is due to the stability and consistency of
community structure and composition under given natural conditions and the relatively rapidly
respond to anthropic and natural stress (Simboura and Zenetos, 2002). There are several

25
characteristics of these communities that make them respond predictably to many kinds of natural
and human induced pressures: (1) Most benthic invertebrates are fixed in their habitat and have low
mobility, therefore being unable to avoid the potential local harmful impact and can thus reflect directly
the local habitat status; (2) Life cycles, long- life and the high recruitment potential of most benthic
macroinvertebrate species allows the community structure to integrate and reflect disturbances in a
long period of time; 3) Benthic species can be sensitive to different stress types, hence their
monitoring can reflect diverse type of stress; (4) those benthic organisms that their habitat is in
sediment have the potential to better reflect long term accumulation of contaminants in the
sediments; (5) Benthic organisms are a very important component of estuarine ecosystems, closely
coupled with the pelagic food web, constituting a link for the transport of contaminants to higher
trophic levels (Chainho, 2008; Borja et al., 2000, ; Veríssimoa et al., 2012).
Response of macroinvertebrate communities to different disturbance types is often evaluated using
‘‘metrics’’, which describe biological conditions from structural and/or functional assemblage
measures. Whereas single metrics reflect only one aspect of the assemblage such as number of
individual taxa and diversity and may not indicate effects of multiple stressors, a multimetric analysis
incorporates several single assemblage/habitat metrics that encompass multiple aspects of
assemblages and thus may provide a more powerful means of assessment (Maloney and Feminella,
2006). Besides these recently developed indices, which were especially developed to meet the
requirements of the WFD, several community-descriptive parameters and indices exist that have been
used in conjunction with the demands of the WFD (Wetzela., et al., 2012). Specifically for
macroinvertebrates, different European countries are adopting multimetric approaches, which try to
include different aspects of macroinvertebrate community structure, compliant with the WFD, such as
species richness, diversity and taxa composition. It is proposed that for benthic quality assessment in
transitional waters, it would be necessary to assess not only the structural attributes of the
community, but also its functional attributes (Elliott and Quintino, 2007). Functional features refer to
the holistic performance of ecosystems and are directly related with ecosystem processes (properties,
goods and services) and to the individual components involved (Gamito et al., 2012). Benthic
macroinvertebrate communities, considered in the present study to be organisms retained in a 0.5
mm screen, have been widely used as indicators for assessing and monitoring anthropogenic impact
on aquatic ecosystems. Impacts on aquatic ecosystems may be measured at different levels of
biological organization, which can include several components of the ecosystem (e.g. estuarine food
web), certain communities (e.g. benthic infaunal macroinvertebrates), a few indicator species (e.g.
pollution indicator species) or even populations (Chainho, 2008).
1.2.3.2.1 Benthiccommunityresponsetosalinitygradientsinestuaries
Although the distribution of faunal estuarine species is primarily determined by their responses to the
highly variable physical (e.g. sediment type) and chemical (e.g. oxygen concentration) environments,
their distribution can also reflect their response to different tolerances of freshwater and marine
species to salinity variations (Remane, 1934). Estuaries are the most productive marine coastal
environments because nutrient-rich freshwater mixes with highly oxygenated waters from the seas

26
(Wetzela et al., 2012). Estuaries are naturally stressed due to strong spatio-temporal variability in
water properties (e.g. salinity). Salinity is a major factor that influences environmental conditions along
the estuarine and its fluctuation can be an important disturbance factor for benthic communities
(Medeiros et al., 2011). In addition, estuaries are characterized mainly by strong gradients (salinity,
temperature), and by changes and fluctuations of these gradients due to the tidal regime making them
unique habitats for a variety of brackish-water species. The spatial extent of organism distribution
within estuaries is determined by the degree of freshwater entering from major tributaries coupled with
the physiological tolerance to salinity conditions made variable by the marine influence (Attrill and
Power, 2000). Remane (1934) proposed a first species distribution model, known as the “paradox of
brackish water” which depicts the quantitative relations between freshwater, brackish and marine
invertebrate species (Fig. 1.3). The paradox indicates that the abundance of freshwater species
decreases drastically with a slight increase in salinity, while a higher number of marine species are
more tolerant to salinity decrease. The two peaks of higher species abundance in the figure
correspond to freshwater and marine salinities.
Figure 1.3‐ Remane curve (after Remane, 1934), showing quantitative relations between freshwater, brackish and
marine invertebrate species. The relative number of species is indicated by the vertical extension of the respective area.

27
1.2.3.3 Seasonalandspatialpatternsofbenthicinvertebrates
The estuarine environments, particularly in Portugal, are characterized by both spatial and temporal
fluctuation not only between estuaries across different locations, but also within each estuary
(Chainho, 2008). Benthic communities show high spatial heterogeneity in estuaries, related to the
influence of natural gradients of different environmental factors. Many benthic species occur along a
wide spectrum of an estuarine environment, while others are confined to a narrower distribution,
according to their tolerance to environmental variables such as salinity, sediment type, depth, etc. In
addition to spatial patterns, there is a temporal variation in invertebrate community presence related
to seasonal and interannual changes. The abundance and composition of benthic community can
also vary seasonally, due to recruitment pulses that occur during spring and autumn for most species,
but also to the occurrence of extreme environmental conditions such as low temperatures, floods and
droughts (Attrill and Power, 2000; Chainho, 2008). Seasonal cycles of precipitation and river flows
contribute to spatial and temporal variability in the structure of estuarine invertebrate assemblages
(Attrill and Power, 2000; Rundle, 1998). Freshwater flow variability is one of the main factors
influencing the high temporal and spatial changes in physical, chemical and biological conditions in
estuaries, particularly in rivers that show strong seasonal changes (Kimmerer, 2002). These
hydrodynamic fluctuations have an important effect on the erosion and depositional cycles,
influencing the sediment composition and therefore the colonization by particular benthic
communities. According to Rundle et al. (1998), the effect of low flows on tidal freshwater
macroinvertebrates at the head of an estuary with small increases in salinity can cause dramatic
changes in community composition.

28
Chapter 2: Temporal and Spacial variation of benthic communities in GDEs influenced by climate change

29
2.1 Abstract
Communities located in the interface between marine/brackish and freshwater habitats are likely to be
early responders to climatic changes as they are exposed to both saline and freshwater conditions,
and thus are expected to be sensitive to any change in their environmental conditions. Climatic effects
are predicted to reduce the availability of groundwater, altering the hydrological balance on estuarine
aquifer interfaces. Here, we aimed to characterise the estuarine faunal community along a gradient
dependent on groundwater input, under a predicted climatic scenario of reduction in groundwater
discharge into the estuary. Sediment macrofauna was sampled along a salinity gradient following both
the wet and dry seasons in 2009. Results indicated that species abundance varied significantly with
the salinity gradient created by the groundwater discharge into the estuarine habitat and with
sampling time. The isopode Cyathura carinata and the polychaetes Heteromastus filiformis and
Hediste diversicolor were associated with the more saline locations, while oligochaeta and Spionidae
were more abundant in areas of lower salinity. The polychaete Alkmaria romijni was the dominant
species and ubiquitous throughout sampling stations. This study provides evidence for estuarine
fauna to be considered as a potentially valuable indicator of variation in the input of groundwater into
marine-freshwater interface habitats, expected from climatic pressures on aquifer levels, condition
and recharge rates. For instance, the abundance of the Spionidae, Alkmaria romijni and Hediste
diversicolor will diminish greatly under severe reduction of groundwater discharge into estuarine
ecosystems. These specimens can potentially be early warnings of a reduction in the input of
groundwater into estuaries. Estuarine benthic species are often the main prey for commercially
important fish predators such as in our case study, making it important to monitor the aquatic habitat
interfaces taking into consideration the estuarine macrobenthos and groundwater availability in the
system.

30
2.2 Introduction
Groundwater is currently considered a key resource under severe threat due to human consumption,
pollution and climatic pressures (Danielopol et al., 2003). Groundwater is also regarded as the
primary provider for human freshwater demands such as agriculture and industry (Santos et al., 2002;
Danielopol et al., 2003). In southern Portugal, where the present study was conducted, groundwater
represents 60% of freshwater human usage and nearly 80% of agricultural demand. These demands
are likely to increase in the future as a result of global warming (Santos et al., 2002). Estuaries often
interface with coastal aquifers and receive groundwater discharge, implying that communities therein
are subject to changes in the salinity due to both the marine and freshwater influences. The
groundwater discharge into estuarine habitats may be reduced by the climatic-driven pressures on its
availability, and thereby induce changes the ecosystem stability. The interface or border habitats such
as estuaries and wetlands are considered to be highly vulnerable to alterations in variables such as
salinity, sediment and nutrient availability (Bates et al., 2008). This is the case of the estuary
associated with the Arade River at the south coast of Portugal, a system of ecological and economic
importance. The estuary permanently receives groundwater from the Querença-Silves aquifer, the
largest and most productive aquifer in the south of Portugal. If the average temperature rises in the
near future as predicted in climate change scenarios for this region (Santos et al., 2002), more
drought periods will occur. In these scenarios, the groundwater withdraws will rise and its output to
adjacent habitats such as estuaries will be reduced and ecosystem stability be altered (Ranjan et al.,
2006). Hence, the fauna of associated habitats such as the Arade estuary will likely be exposed to
increased salinities, becoming important to understand how benthic estuarine species respond in the
present day to the groundwater-estuarine interaction. Invertebrate communities of transitional waters
(freshwater-saltwater ecotone) are highly influenced by freshwater discharge, showing marked
seasonal variation mainly related to salinity fluctuations (Chainho et al., 2006). Alterations in the
benthic community will likely have cascade trophic consequences for their predators, which in the
studied estuary correspond mainly to commercially important fish species (Cabral, 2000; Gonçalves
et al., 2004). The salinity variation related to the groundwater discharge into the estuary and the
benthic species tolerances to salinity levels have an important role in determining prey availability for
their predators. The aim of the study was to identify benthic estuarine taxonomic groups and/or
species which can potentially be monitored to ecological impacts of changes in groundwater
discharge. It was expected that benthic communities responded in presence, abundance and
potentially in population structure, to the gradient in salinity originated by the freshwater input into the
estuary, due to discriminating salinity tolerances of organisms and species. Identification of
bioindicators demands establishing a causal relationship and the tested factor ideally requires in situ
manipulative tests, whereby levels of groundwater would be applied to faunal communities and their
response (e.g. presence-absence, abundance, etc.) measured (Silva et al., 2012). Groundwater is
inaccessible for control in the estuarine locations of the present study mainly because it runs and
surfaces at the sub-estuary level and such manipulations would be logistically challenging and are out
of scope of the present work. The coastal aquifers of the Algarve region have been categorised as a

31
highly vulnerable to the threat of saline intrusion, and the fauna of associated habitats such as the
Arade estuary will likely be exposed to increased salinities, becoming of paramount importance to
understand how benthic estuarine species respond in the present day to the groundwater-estuarine
interaction. Therefore, the present work contributes to the development of a biologically based
method to evaluate cascading Climate Change impacts such as seawater intrusion on groundwater
associated ecosystems. The present work aimed to:
(i) Identify transition zone ecotones such as surface-groundwater and salt-fresh water
interfaces and, assess benthic community structure, so as use of the tool to assessment
of water quality in wetlands partly dependent on groundwater.
(ii) Assess the response of benthic macroinvertebrates community to shifts in salt-fresh
water availability (possible bioindicators), so as to provide the information needed to
broadly assess the condition of wetlands as groundwater dependent ecosystems
throughout different conservation states, especially with respect to changes in
groundwater- fresh water availability (wet and dry periods).

2.3
2.3.1
The stud
shown i
summer
2010 pe
the main
This es
channels
is the Q
approxim
area bet
average
total out
two year
discharg
discharg
0.5 ppt (
Estuary)
branchin
compari
discharg
Figure 2.1
Methods
Studyare
died area is
n fig. 2.1. Th
rs and cool
eriods were 1
n estuarine c
stuary comp
s, and freshw
Querença-Si
mately half-w
tween the m
e groundwate
tflow, though
rs (2008-201
ge has been
ge. Salinity m
(part per tho
) at low and
ng channel
son along a
ge can have
1‐The enlarged(orange line)
s
ea
located on
his region is
and wet win
17.5 °C and
channel of th
prises exten
water contrib
lves, the la
way between
main estuary
er discharge
h this percen
10), continuo
n performed
measuremen
ousand) (Loc
high tide re
which then
a salinity gra
on biologica
map the Algar. The main are
the costal fr
characterize
nters. The m
739 mm res
he Arade riv
sive intertid
butions from
rgest in the
n the river a
channel and
from the aq
ntage varies
ous salinity re
, providing
nts along the
cation close t
espectively. G
n flows into
adient, the p
l communitie
rve province ana of Querença‐
ringe of cent
ed by a war
mean annual
spectively. Sa
ver near Silve
dal soft-sedi
m submerged
e Algarve re
and sea poin
d a point of
quifer into the
between se
ecording in th
reliable mea
e estuary sh
to Groundwa
Groundwate
o the main
potential risk
es.
nd the main wa‐Silves aquifer
tral Algarve
rm Mediterra
temperature
ampling was
es (Estômba
iment banks
and surface
egion. The
nts of the es
groundwater
e branching
asons and y
his lateral ch
asurements
ow a relative
ater output) t
r is directly
estuarine c
k that an ev
ater courses (blis shown by lig
in the south
anean climate
e and precip
s made at a b
ar: 37° 9'42.8
s partially s
e aquifer disc
branching c
stuary. It rep
r discharge i
channel is a
years (Stigter
annel at the
of salinity v
ely large sco
to >30 ppt (l
discharged a
channel, allo
ventual shift
lue) and corresht blue area ( S
h part of Por
e with dry a
pitation for th
branching ch
87"N, 8°29'1
separated b
charges. Th
channel was
presents an
into the estu
approximately
r, 2011). In
point of grou
variation with
ope of variat
ocations clo
at the surfac
owing asses
t in the grou
sponding catchStigter, 2011).
32
rtugal, as
nd warm
he 1980-
hannel of
0.61"W).
by water
e aquifer
s located
interface
uary. The
y 25% of
the past
undwater
h aquifer
tion from
se to the
ce of the
ssing by
undwater
ment area

33
2.3.1.1 Hydrologyandhydrogeology:
The most productive and important groundwater reservoir in South Portugal is the Querença-Silves
aquifer due to its large area and significant recharge. It covers an irregularly E-W elongated area of
324 km2 .The aquifer is built up of carbonate sedimentary rock, has a total extent of 324 km2 and a
mean annual recharge of 100 ×106 m3 (Monteiro et al., 2007; Stigter et al., 2009). The aquifer is
mainly explored for agronomic irrigation purposes (31 × 106 m3/year) and public water supply (around
10 × 106 m3/year). In extremely dry years like in 2005, the total abstractions of the aquifer reached 60
× 106 m3/year, representing more than half of the aquifer annual recharge rate. Surface stream flow in
this region (Central western Algarve) is restricted to a small number of temporarily rivers, which only
flow during and shortly after heavy rainfall events. Stream flow in the region can be divided into three
major units: (i) the low permeability Paleozoic rocks in the North, where large surface water reservoirs
exist for urban water supply and irrigation: (ii) the area of the aquifers, where the infiltration from
surface water is high and surface stream flow depend on hydraulic connection with groundwater in
several influent and effluent stream reaches and, (iii) the areas of less permeable Upper Jurassic and
Cretaceous rocks, where Miocene carbonate and detritic aquifers are covered by low permeability
Pliocene to Quaternary sediments; here stream flow towards the sea occurs over areas with variable
infiltration rates (Stigter, 2011).The main aquifer discharge points into the estuary are springs located
at the aquifer boundaries, of which the Estômbar springs at the western boundary represent the main
aquifer discharge location.
2.3.1.2 Landuse
According to the CLIMWAT project investigation, land use in Querença-Silves aquifer is significantly
different between the northern and the southern area, driven essentially by the soil type and
topography of these distinct sub-regions. In the northern part, the land is mostly covered by oak
savannah and Mediterranean shrub land, while in the southern area; the land is intensively cultivated
and densely populated by trees and citrus orchards. These require large amounts of irrigation, for
which groundwater is used, though outside the aquifer area, surface water from the reservoirs is the
main source. The crucial role of this aquifer system for the support of Algarve’s water supply in urban
areas was clearly revealed during the severe drought that affected Portugal in 2004 and 2005,
causing the depletion of the surface water reservoirs (Stigter et al., 2009).
2.3.2 Samplingdesign
A total of five sampling stations were selected in the Estômbar site and their position reflected a
distance-based gradient, starting at the point of groundwater surface discharge into the estuary
(Fig.2.2. and Fig.2.3).

Figure
discharg
Location
A was ~
the sam
the grou
were sa
0.5 mm
formalin
examine
e 2.2‐ Sediment
ge point. The sa
n C was adja
~200 m away
mpled branch
undwater dis
mpled at eac
square me
stained wi
ed at the en
Arade E
t sampling poin
alinity gradient
acent to the p
y, location D
ing channel
scharge poin
ch location u
sh, for mac
ith Rose Be
nd of wet (
Est
Estuary
nts. First locatio
t is also marked
point of groun
was ~400 m
meets the m
nt. Sampling
sing hand co
crofaunal stu
engal until
(April 2010)
tômbar
on on the right
d by colours fro
(Stigter, 2011)
ndwater disc
m away, and l
main estuarin
occurred a
orer devices
udies. The r
further anal
and dry (O
t side of the bo
om the lowest p
).
charge, locat
location E wa
ne channel,
t low-tide an
. The sedime
retained ma
lysis. Macro
October- No
Ara
de
ottom image co
point (blue) to
tion B was ~1
as situated a
approximate
nd five replic
ents were sie
terial was fi
ofaunal time
vember 200
estu
ary
orresponds to t
the highest po
100 m away
at the endpoi
ely 600 m aw
cate cores (
eved in situ t
fixed in 4%
variation w
09) periods.
34
he GW
oint (red).
, location
int where
way from
(0.01 m2)
hrough a
buffered
was also
Prior to

identifica
groups,
2.3.2.1
The stud
hydrolog
oxygen)
Data So
Oxygen,
concent
factor th
accordin
sedimen
discharg
measure
correlati
limitation
sample w
(DW) wa
dried un
organic
method
The perc
– DW af
Figure 2.
ation, sampl
identified to
Environ
dy area was
gical regime
using histor
onde Survey
, water tem
ration was m
hat discrimin
ng to in situ o
nts, while po
ge were mud
ed because
on between
n factor (>1
was taken w
as determine
ntil constant w
matter in th
(LOI) at 450
centage of o
fter ignition) /
.3‐ Specific samth
les were wa
the lowest p
nmentalva
s characterise
e (e.g. preci
rical and cur
or 4 includin
mperature (C
mentioned in
ate the surfa
observation,
oint A had co
ddy-sandy ty
available lit
organic mat
mg/l). To
with Corer
ed for bioma
weight at 90
he surface s
0ºC for 4 ho
organic matte
/ DW before
mpling locationshe connection o
ashed and t
ractical taxo
ariablesme
ed through h
ipitation), an
rrent data. S
ng Bottom d
Cº), PH, sali
n the GW In
ace from gro
the main se
oarser grain
ype with som
erature (Hak
tter and faun
measure th
and frozen
ass estimatio
0ºC. Percenta
sediment we
urs (Kristens
er was given
ignitionx100
s. Point C referof the branchin
the organism
nomic level (
easurement
hydrogeolog
nd geochem
pecific abiot
issolved oxy
inity and El
ndex-Fauna
oundwater fa
ediment type
and muddy
me gravel (Fi
kenkamp an
na in GW-SW
e organic m
for posterior
on in macroi
age of organ
re determine
sen and And
by the equa
0
rs to the locationg channel to th
ms were ha
(mostly spec
t:
ical paramet
mical propert
ic parameter
ygen (DO (m
ectrical Con
developed b
auna. The g
e at Points D
y- type. The
g. 2.3). The
nd Morin, 20
W exchange
matter conten
r organic ma
nvertebrate
nic matter in
ed in sub-sa
derson, 1987
ation: Organ
on of GW dischhe main estuar
nd sorted in
cies level) an
ters (e.g. Ele
ties (e.g. sa
rs were mea
mg l−1)), perc
nductivity (A
by Hahn (20
grain size wa
and E were
points close
Organic ma
00) indicate
if oxygen c
nt at each s
atter content
samples wh
invertebrate
amples throu
7), after dry w
ic matter % =
arge and Pointry canal.
nto major ta
nd counted.
ectrical Cond
alinity and d
asured in situ
rcentage of d
Appendix 1).
006) as an i
as not meas
e pure muds
e to the grou
atter in sedim
ed that there
concentration
station, an a
analysis. Dr
hereby samp
es and perce
ugh ‘loss on
weight quan
= (DW befor
t E is located in
35
axonomic
ductivity),
dissolved
u using a
dissolved
Oxygen
mportant
sured but
with fine
undwater
ment was
e is good
n is not a
additional
ry weight
ples were
entage of
n ignition’
tification.
e ignition

Salinity
high tide
were pre
Figure 2points A,
and Electric
e and low tid
esented.
2.4‐ Estômbar ,B, and C in low
cal Conductiv
de in 2009.In
A
C
channel , a braw tide(B); the Es
vity measure
n Fig.2.4 som
anching channestômbar chann
ements were
me picture fr
el of the Arade nel in high tide(
e collected t
rom the sam
estuary at low(C) and location
two times in
mpling pints o
B
D
tide(A), and thn of groundwat
n each statio
on Estômbar
he location of ster input at hig
36
on during
r channel
sampling gh tide(D).

37
2.3.3 Dataanalysis
2.3.3.1 Spatio‐Temporalvariation
The present study considered a two factor design: Location (orthogonal, fixed, 5 levels (Salinity A to
salinity E) and Time (orthogonal, fixed, 2 levels: end of dry and wet seasons). This design does not
allow withdrawing conclusions on seasonal patterns as no replicated times were measured within
seasons. Five replicates were taken at each location and time combination. Multifactorial analysis was
used to determine Spatio-temporal variation of benthic community. The analyses was made in the
PRIMER+ package (Clarke and Warwick, 2007; Anderson et al., 2008). The similarity percentage
breakdown procedure (SIMPER) was used to determine the contribution of individual taxa to the
dissimilarity between and within the factors, and also to identify the species most contributing for
significant factorial differences. MDS and dbRDA ordination techniques were calculated to graphically
examine the samples distribution pattern. For all tests, the Bray-Curtis similarity measure, the fourth-
root transformation and 9999 permutations were used for all biotic data, while the abiotic data was
normalized and the Euclidean distance measure used in the PRIMER+ calculations. The fourth root
transformation was applied to give more weight to the rare species and reduced the weight of the
dominant species. The statistical software package IBMSPSS 19 was used to show graphically the
temporal variation on the densities of the eight dominant species in the benthic community.
2.3.3.2 Relationshipsbetweenenvironmentalandbiologicalvariables
Several environmental variables were measured in the study site during the experiment and the
PRIMER+ technique BEST was used to calculate the most parsimonious model explaining the
species distribution, in accordance with the measured abiotic variables. The model with the highest
correlation coefficient corresponds to the environmental variables that better explained the biotic
distribution pattern. Biplots of sampling points and environmental variables were displayed in
Ordination by principal coordinate’s analysis (PCO; Gower 1966), in which it is possible to graphically
examine the relation between environmental variables and community distributions. The PCO
analysis was made only for the location factor as only the wet period was considered in the
calculations because the abiotic variables were only measured at the end of the wet season.

38
2.4 Results
2.4.1 Benthicmacrofaunageneralcharacterization
A total of 6786 invertebrates were collected and 38 taxa identified (Appendix 2). The highest and
lowest densities were registered after the wet period at location B, with respectively 331 individuals/
0.01 m2 and 14 individuals/ 0.01m2. Annelida (Polychaeta and Oligochaeta) and Crustaceans
(Isopoda and Amphipods) were the dominant sampled groups of macroinvertebrates. Oligochaeta,
Spionidae and the species Alkmaria romijni Horst, 1919 and Cyathura carinata were the most
abundant throughout the sampled locations. Alkmaria romijni accounting for 29% of the total number
of individuals, while the Oligochaeta represented 26%, Spionidae 14% and the isopod Cyathura
carinata 11% of the total number of individuals. Dominant species found in wet and dry period were
typical brackish water species but some freshwater invertebrates (Castaneda and Drake, 2008) such
as Lekanesphaera hookeri leach 1814, Hediste diversicolor O.F. Müller, 1776 and Alkmaria romijni,
were collected.
2.4.2 Speciesdistribution
The species distribution was significantly explained by the interaction of the distance to the point of
groundwater discharge into the estuary, i.e. location, and sampling time (Table 2.1).
Table 2.1‐ Permanova analysis of the sediment fauna for factors Period (dry and wet) and Location (A‐E). The number of permutations used was 9999 (Ti=period, Lo=Location).
Source df SS MS Pseudo ‐F
P (perm)
U. perms
Ti 1 3347,7 3347,7 3,5409 0,0021 9946
Lo 4 12959 3239,7 3,4267 0,0001 9913
Ti x Lo 4 9498 2374,5 2,5115 0,0008 9899
Res 40 37818 945,44
Total 49 63622
The MDS ordination graphically represents the community variation in relation to factor time and
location. The plot showed that the community structure varied strongly with sampling time, and
relatively less with location, although location D is clearly separated from the remaining locations,
which are more homogenous in between (Fig. 2.5). Pairwise analysis was used to show where the
differences are (Time: wet-dry/Location: A-C). Pairwise comparisons for pairs of factor time showed
that the benthic community only varied significantly between the dry and wet times at locations near
the main estuary channel (Pair-wise tests, p (perm) < 0.05 for location E; p (perm) < 0.05 for location
D. As expected, pair-wise tests for term location only showed significant differences in the abundance
in pairs with locations D or E (including A, D; A, E; B, D; B,E; C,D; C,E).

Figure
The SIM
between
between
Fabriciu
than on
to be m
approxim
dominan
SIMPER
Table 2
SpeOligo
Spio
Alkm
Hedi
Capi
Scrob
2.5‐MDS ordin
MPER analy
n periods for
n seasons w
s,1780 and t
the wet time
more abunda
mately 50%
nt specimen
R analysis.
2.2‐ SIMPER an
ecies ochaeta
nidae
maria romijni
iste diversico
itella capitat
bicularia pla
ation of fauna gradient o
ysis identifie
all locations
were Oligoch
the bivalve S
es (Table 2.2
ant in the w
for the diffe
s for the dr
alysis identifyin
G
D
A
i
olor
a
ana
sediment samoriginating at th
ed which sp
s (Appendix 3
haeta (11%)
Scrobicularia
). The A. rom
wet sampling
erences betw
ry and wet
ng the species
Group
Dry
G
W
Av.Abund A
1,90
1,50
2,01
1,24
0,97
1,11
ples collected ahe location of g
pecies contr
3). The taxa
which, toge
a plana da Co
mijni, H. dive
g time. Tog
ween periods
periods we
that contributelocations
Group
Wet
Av.Abund
1,19
1,63
2,26
1,35
0,70
0,78
at the end of thgroundwater in
ributed the
which contr
ether with th
osta, 1778, h
ersicolor, 177
gether, these
s. In Fig 2.6
re illustrated
ed the most fo
Av.Diss D
5,29
4,89
4,35
3,61
3,59
3,15
he dry and wetnput (A ‐E).
most for si
ibuted the m
he Polychae
had higher ab
76 and the Sp
e taxonomic
6 the variatio
d and emph
r differences be
iss/SD Co
1,09 1
1,23 1
0,98 9
1,02 7
1,18 7
1,01 6
t times, in the d
ignificant dif
most for dissi
eta Capitella
bundances i
pionidae wer
c groups co
on in abund
hasize the r
etween season
ntrib% Cum
11,34 11,
10,48 21,
9,33 31,
7,75 38,
7,70 46,
6,74 53,
39
distance
fferences
milarities
capitata
n the dry
re shown
ontributed
ances of
esults of
ns for all
m.%
34
81
14
89
59
33

Figure 2.6
6‐ Temporal variations on the densities of ta(D‐dry
axa that contribperiod, W‐wet
buted the mostt period)
t on the dissimiilarities betwe
40
en periods

41
Taking into consideration the factor location there were small dissimilarities in community composition
between points A and B (around 37%), points A and C (around 35%) and C and B (around 44%). The
average dissimilarity was larger between locations D and C (around 60%), and also between points D
and B (around 60%) (Appendix3). Table 2.3 shows the taxa that contributed the most for
dissimilarities between points D and C. Location C (corresponding to where the freshwater output is)
differed from location D (Point close to estuary) mainly due to lower abundances of Oligochaeta, A.
romijni and H. diversicolor in location D. This location had, however, larger abundances of the bivalve
Hidrobia ulva Pennant, 1777.
Table 2.3. SIMPER analysis identifying the taxa that contributed the most to differences from point D to the freshwater point (C )
Group C Group D
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Oligochaeta 2,28 0,56 8,06 1,97 13,50 13,50
Alkmaria romijni 1,83 1,52 7,37 1,69 12,35 25,85
Spionidae 1,85 1,03 6,07 1,44 10,17 36,02
Hediste diversicolor 1,43 0,70 5,80 1,29 9,71 45,73
Hydrobia ulva 0,00 1,12 5,30 0,92 8,88 54,62
The SIMPER routine showed that the location closer to the freshwater source (location B) had higher
abundances of Oligochaeta, A. romijni, Spionidae and C. capitata than location D. The location closer
to the main estuary channel and therefore in more brackish conditions (Point D), differed from less
saline conditions locations (point B) mainly due to larger abundances of Hidrobia ulva.
The Polychaeta A. romijni was ubiquitous throughout all sampling locations although generally more
abundant in locations closer to freshwater conditions.
Table 2.4‐ SIMPER analysis identifying the species that contributed the most for dissimilarities between point D and point B
Group B Group D
Species Av. Abund Av. Abund Av. Diss Diss/SD Contri% Cum%
Oligochaeta 2,10 0,56 8,14 1,98 13,71 13,71
Alkmaria romijni 2,23 1,52 6,80 1,29 11,46 25,18
Spionidae 1,84 1,03 6,78 1,46 11,42 36,60
Hidrobia ulva 0,00 1,12 5,13 0,85 8,64 45,24
Capitella capitata 1,27 0,40 5,03 1,85 8,48 53,72
Melita palmata 1,11 0,27 4,40 1,09 7,42 61,14
Hediste diversicolor 1,20 0,70 4,11 1,05 6,93 68,07

2.4.3
The dbR
variation
species,
Hidrobia
capitata
location
F1,49=1,7
F4,49=1,7
Figure 2.C, D
Speciesc
RDA ordinat
n as both axe
, with correla
a ulva isolate
, Oligochaeta
E (fig.2.7).
78, P(perm)=
71, P(perm)=
.7‐ dbRDA ordiD, E) for the abu
ontributi
tion was co
es included
ations R>0.4
ed almost all
a and Scrob
No pattern
=0,17; Locat
=0,31).
nation of the sundance of spe
ionforspa
nsidered to
approximate
were consid
samples fro
icularia plan
was detecte
tion: Pseudo
ample variatioecies, where the
atialand
be a consi
ely 50% of th
dered. This re
om location D
na separated
ed for separa
o- F 4,49=1,87
on during wet (We overlying vec
temporal
iderably goo
he total varia
epresentatio
D, while Alkm
location A, B
ation of dry
7, P(perm)=0
Wet) and Dry (ctors are the sp
ldifferenc
od represent
tion (Fig 2.7
n clearly sho
maria romijni,
B and C from
and wet tim
0,08; Period
Dry) season anpecies which ha
ces
tation of the
7). Only varia
owed that the
, Spionidae,
m most of sa
mes (Period:
x Location:
nd in five locatiad correlation >
42
e sample
ables, i.e.
e species
Capitella
amples of
Pseudo-
Pseudo-
ons (A, B, >0.4.

2.4.4
Variation
observe
location
highest
Figurdischargeseason (s
Environm
n in levels o
d in Fig.2.8.
C. Location
level of in
re 2.8‐ Variatioe into the Estômsummer, upper
tidal flu
mentalvar
of electrical
There was
n C is partic
fluence of
n of water levembar channel or graph) and in ctuations meas
riables
conductivity
a high level
cularly intere
groundwate
el, electrical coof the Arade esthe middle of tsured in Lagos,
y (EC, an in
of variation
esting to ob
r discharge
nductivity (EC)stuary. Two repthe rainy seaso, located 20 km
ndicator of s
of electrica
bserve as it
and to ty
and temperatpresentative timon (nearly wintm towards the w
alinity) and
l conductivity
simultaneou
pical estuar
ure at the locatmes are indicater, lower graphwest (Silva et a
temperature
y and tempe
usly respond
rine brackis
tion of groundted, at the end h); also indicatl., 2012).
43
e can be
erature in
ds to the
h water.
water of the dry ed are the
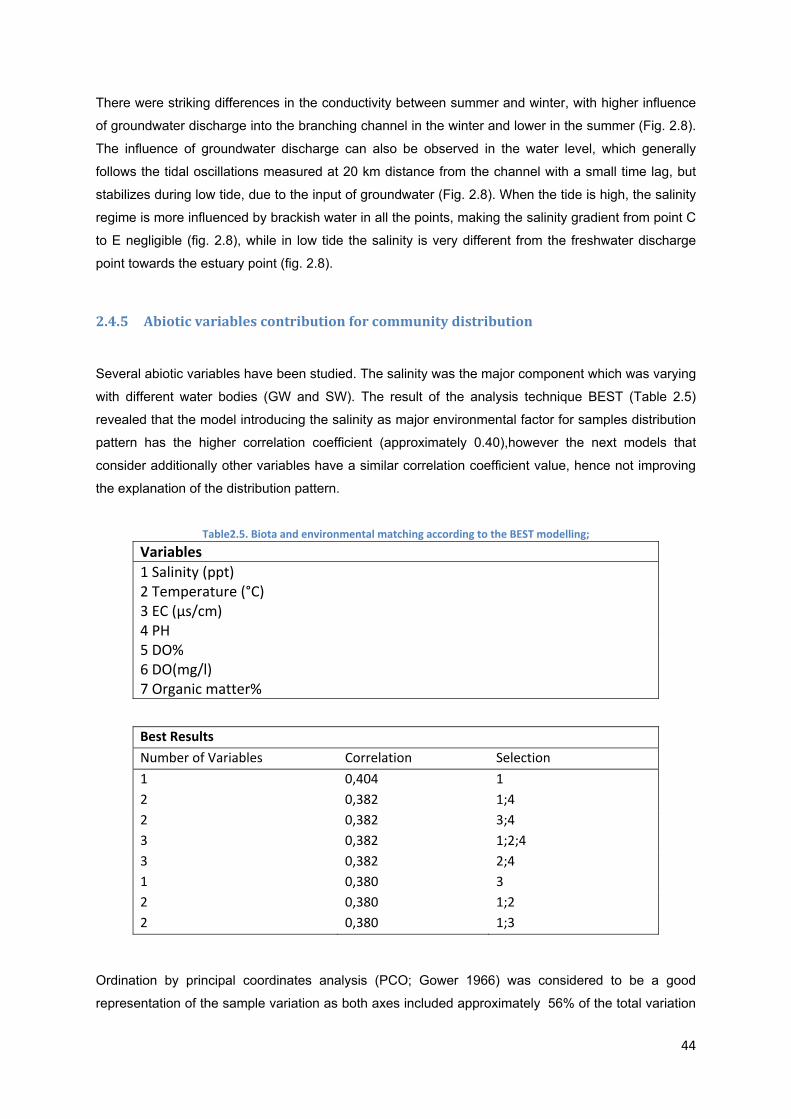
44
There were striking differences in the conductivity between summer and winter, with higher influence
of groundwater discharge into the branching channel in the winter and lower in the summer (Fig. 2.8).
The influence of groundwater discharge can also be observed in the water level, which generally
follows the tidal oscillations measured at 20 km distance from the channel with a small time lag, but
stabilizes during low tide, due to the input of groundwater (Fig. 2.8). When the tide is high, the salinity
regime is more influenced by brackish water in all the points, making the salinity gradient from point C
to E negligible (fig. 2.8), while in low tide the salinity is very different from the freshwater discharge
point towards the estuary point (fig. 2.8).
2.4.5 Abioticvariablescontributionforcommunitydistribution
Several abiotic variables have been studied. The salinity was the major component which was varying
with different water bodies (GW and SW). The result of the analysis technique BEST (Table 2.5)
revealed that the model introducing the salinity as major environmental factor for samples distribution
pattern has the higher correlation coefficient (approximately 0.40),however the next models that
consider additionally other variables have a similar correlation coefficient value, hence not improving
the explanation of the distribution pattern.
Table2.5. Biota and environmental matching according to the BEST modelling;
Variables
1 Salinity (ppt) 2 Temperature (°C) 3 EC (µs/cm) 4 PH 5 DO% 6 DO(mg/l) 7 Organic matter%
Best Results
Number of Variables Correlation Selection
1 0,404 1
2 0,382 1;4
2 0,382 3;4
3 0,382 1;2;4
3 0,382 2;4
1 0,380 3
2 0,380 1;2
2 0,380 1;3
Ordination by principal coordinates analysis (PCO; Gower 1966) was considered to be a good
representation of the sample variation as both axes included approximately 56% of the total variation
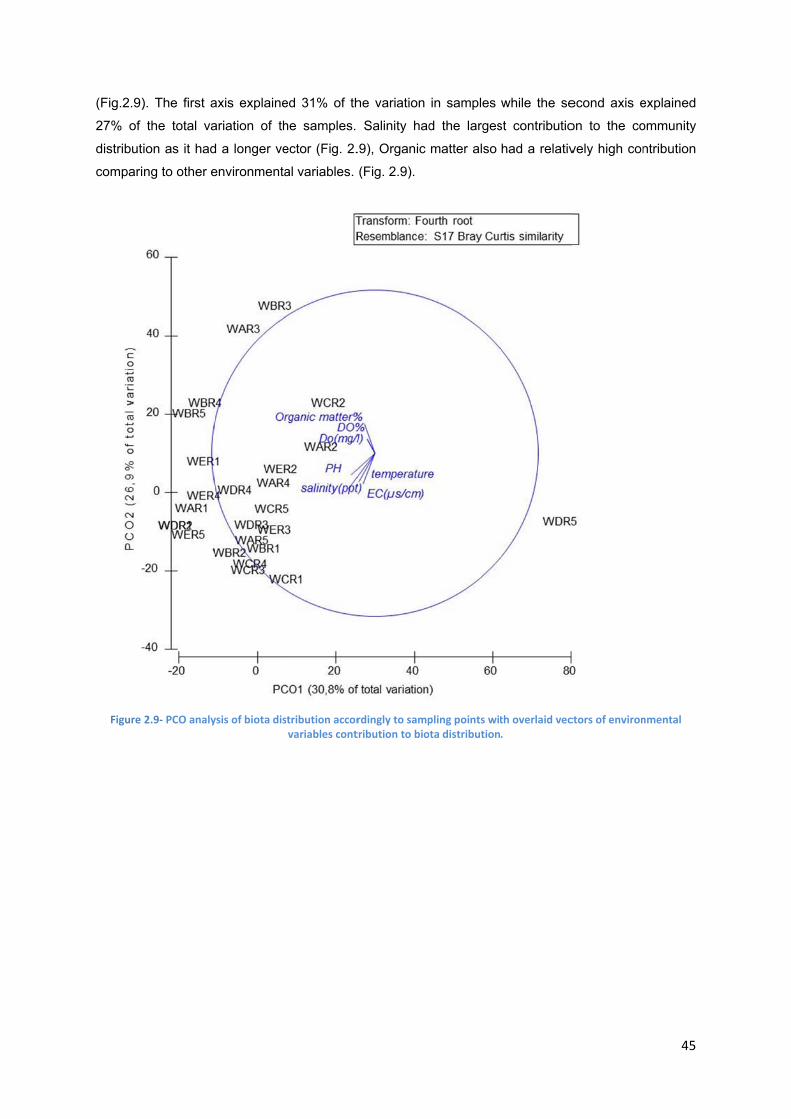
(Fig.2.9)
27% of
distribut
compari
Figure
). The first a
the total va
ion as it had
ng to other e
2.9‐ PCO analy
axis explaine
ariation of th
d a longer ve
environmenta
ysis of biota dis
ed 31% of th
he samples.
ector (Fig. 2.
al variables.
stribution accorvariables cont
he variation
Salinity had
.9), Organic
(Fig. 2.9).
rdingly to samptribution to bio
in samples
d the larges
matter also
pling points witota distribution
while the se
st contributio
had a relativ
th overlaid vec.
econd axis e
on to the co
vely high con
ctors of environ
45
explained
ommunity
ntribution
nmental

46
2.5 Discussion
2.5.1 Groundwateravailabilityandcommunitypredictions
The present work represents the first study in Portugal examining the influence of groundwater
availability on estuarine biodiversity. There was a correspondence of the community distribution
pattern to a salinity gradient established by groundwater discharge into the estuary. There was also
indication that temporal differences in macrobenthic presence may occur between in dry and wet
season, although further evidence is required. Taken together these results indicate that the benthic
estuarine community can be potentially used as a surrogate for evaluating changes in the ecosystem
salinity. Climatic predictions indicate that groundwater will be used in higher human demands in South
Portugal due to increased drought periods (Santos et al., 2002), and that global groundwater
availability and quality will diminish (Danielopol et al., 2003). There is a predicted scenario for the
studied area of reduced aquifer recharge and increased sea level rise followed by a potential saline
intrusion, which can change the salinity condition of the Querença- Silves aquifer (Santos et al., 2002;
Monteiro et al., 2007). This condition can be aggravated by the risk of overexploitation for irrigation
and public water supply and thus, the biological community is likely to suffer specific shifts related to
salinity, which can be detected under a biologically orientated monitoring programme.
It is known from literature that there is also a close relationship between sediment grain size and the
trophic structure of benthic communities in estuaries and, in general, suspension feeders are more
common in coarser sandy sediments while deposit feeders seem to have a preference for muddy
sediments (Chainho, 2008). In the present study, sediment type is primarily a function of flow, since
the transport and deposition of sediment particles from estuary into the channel are regulated by the
diurnal tidal movement. At the upper part of the channel, in the location of the GW input (Point C),
there was more coarse sandy sediment, while the other points toward the estuary (point D and E) had
more fine muddy sediment type. Seasonal or even daily changes of the sediment structure due to
fluctuations in tidal movements can be regarded as physical disturbances and affect the colonization
by benthic invertebrate species. For the present study, the grain size was not measured due to
logistic constrains, therefore it is possible that part of the 50% of the unexplained variation within the
total variation found in our data can be due to sediment type.
The organic matter content in the sediment and salinity are known to be the two most important
physic-chemical features influencing benthic distribution in other Portuguese estuarine systems
(Nunes et al., 2008; Teixeira et al., 2008). Certain aquatic species such as Capitella capitata has
been documented be a reliable indicator of organic enrichment pollutant (Macleod et al., 2004). In the
present study Capitella capitata found to be more abundant in location B with higher Organic
matter%.
The community showed clear evidence of responding to the influence of groundwater discharge and
times. For instance, several species of the Polychaeta class were more abundant at the freshwater
discharge location and at the end of the wet season. Similar groundwater dependence has been
found for rotifers in Brazilian estuaries but in their case for the dry season (Medeiros et al., 2010). The
present study allows the prediction that a decrease in the abundance of the some Polychaeta species

47
and increase in Isopoda, are potentially early warnings of a reduction in groundwater input into this
habitat interaction. The abundance of the Spionidae, Alkmaria romijni and Hediste diversicolor will
potentially diminish greatly under severe reduction of groundwater discharge into estuarine
ecosystems. This is potentially more visible at the end of the dry season when groundwater
availability is likely to be more limited .Conversely, the Isopod Cyathura carinata and Hediste
diversicolor, will potentially remain in the current abundance level unless other environmental
parameters such as organic matter or environmental variables (e.g. pH, temperature) shift. Some taxa
such as Oligochaeta, the second most abundant specimens of our samples, and Capitella Capitata
did not show any pattern with groundwater discharge as they were abundant both at the end of dry
season and freshwater discharge points. This macrofauna Polychaeta is described as a bacterial
pollution indicator and it corresponds to a brackish-saltwater species found in Mediterranean, Atlantic,
Arctic and pacific Oceans (Heyward and Ryland, 1995). Subclass Oligochaeta are known to tolerate a
wide range of salinity since some of their species are fresh water and some are brackish water
species (Chapman and Brinkhurst, 1980; Oikos, 1969). The present study sampled area is a
protected site where aquatic biodiversity is mainly influenced by natural shifts in the tidal cycle and
level of groundwater discharge directly from the aquifer. As discharges cause salinities as low as 0.5
ppt, the organisms adjacent and downstream to the point of groundwater input into the estuary are
directly exposed to sharp salinity shifts. This is analogous to the intertidal rocky shore habitat, as the
fauna therein is also subject to cyclic changes in the presence/absence of water coverage and salinity
levels. If the groundwater discharge is hindered and increased salinities occur, then the composition
and abundance of the faunal community characterised in the present study is likely to vary.
2.5.2 Variationingroundwaterdischargeandfood‐webimplications
The Polychaeta Hediste diversicolor and Alkmaria romijni were found to be sensitive to groundwater
deficit as they are assossiated to freshwater resources. Results indicated that the abundance of
species Isopode Cyathura carinata and the Polychaeta Heteromastus filiformis varied significantly
with the salinity gradient created by the groundwater discharge into the estuarine habitat and they can
potentially be more sensitive to variations in groundwater input into the estuary. These taxa are
important food items for many commercially important estuarine fish, for example the European
flounder Platichthys flesus (Linnaeus, 1758) and Solea solea (Linnaeus, 1758) (Cabral, 2000;
Pasquaud et al., 2010). There are 36 commercially important fish species in the Arade estuary,
indicating the importance of this ecosystem as a nursery for many commercial species (Gonçalves et
al., 2004). Hence, under a predicted climatic scenario of reduction in the groundwater discharge rate
into the estuary, the macrobenthic species found here to be more associated with the groundwater
discharge point such as the Oligochaeta; will likely have their abundance reduced. This may have
bottom-up cascade effects into their predatory fish mentioned above, which will see their prey
availability reduced. Such cascading trophic links and effects should be evaluated and taken into
consideration in a monitoring programme, as shifts in the abundance of those fish species can be also
early warnings signals of changes in prey availability, albeit sampling of invertebrates is logistically

48
easier. Global climate change effects are considered a major threat to estuarine fishes and their
fishery (Roessig et al., 2004), to the structure and dynamics of estuarine macrobenthic communities
(Grilo et al., 2011), and also to coastal fresh groundwater resources (Ranjan et al., 2006). Hence,
groundwater input into estuarine systems can be a key bottom-up factor regulating both food webs
and biodiversity levels. Benthic communities were shown here to provide an indirect method to
evaluate changes in salinity in the interface between aquatic habitats encompassing groundwater
systems. It is important that future monitoring programmes include an assessment of the spatial and
temporal variability of the species identified here as potential responders to groundwater availability.
.

49
Chapter 3: Accessing stygofauna through wells: a window for evaluating GW condition?

50
3.1 Abstract
In coastal areas, groundwater is vulnerable to salinization by intruding natural seawater which makes
it unsuitable for drinking and agricultural use. In arid /semiarid areas such as in the present study
case, climate is expected to aggravate this problem, due to the combined effect of rising sea levels
and reduced recharge of aquifers associated with the expected decrease in precipitation and average
temperature increase. The purpose of this study was to examine the sensitivity of wells fauna to
variation in groundwater salinity/conductivity accessed through wells. Also, it aimed to provide ground
breaking biological data to integrate the use and conservation of groundwater dependent fauna into
aquifer management. Six wells were selected for sampling in which four of them were more
associated to salinization risk as they are located close to Arade estuary, whereas the other two wells
are relatively far from the sea. Wells fauna was sampled with the use of a phreatobiological net
developed for large diameter wells, as well as a well sediment sampling device developed for the
present study (WSSD). Except for the three groups including Oligochaeta, Gastropoda and
Coleoptera, all sampled species belonged to the Crustacea taxa (Daphnia, Cladocera, Cyclopoida,
Harpacticoida, Ostracoda, Amphipoda). A total of 612 specimens comprising so far 19 species were
identified for all 6 wells. Fauna identification was only completed for some families in Crustaceans
including Amphipods, Copepods, and Ostrocods, but further studies are on-going. The Order
Cyclopodia dominated in all 6 wells with relatively high diversity. There was an apparent relationship
between salinity level of well water and stygofauna presence. The results of multivariate analysis
indicate that wells with high salinities were separated from wells with low salinity records. The species
Eucyclops speratus (Lilljeborg, 1901), Eucyclops hadjebensis (Kiefer, 1926), Megacyclops viridis
(Jurine 1820) and Acanthocyclops sensitivus (Graeter & Chappuis, 1914) were particularly associated
with low salinity conditions (Electrical conductivity<1500 µs/s), hence being potential indicators for
saline intrusion if their abundances decrease greatly. Conversely, the taxa Halicyclops sp. is a
possible indicator of high salinity conditions (Electrical conductivity>6000 µs/s). The greatest diversity
and highest abundance were found in the lowest salinity condition (well Q4). The comparison
between the two sampling methods indicated that the Phreatobiological net is more effective in
gathering representative samples, particularly from Cyclopodia which are generally good swimmers,
while the WSSD net is more suitable to have sample groups adapted to benthic life such as
Harpacticoids and Ostracods. Further studies on high and low salinity wells are necessary to
disentangle the relation between species presence and salinity conditions.

51
3.2 Introduction
Groundwater is one of the most essential basic resources for human life. In Europe, 65% of the
drinking water originates directly or indirectly from the subsurface (Steube et al., 2010). Climate
change will have a profound impact on all water resources including groundwater, however compared
to surface water resources, the underground water has not received as much as attention in climate
change risk issues (Bates et al., 2008). This may be because aquifers are considered more resilient
than surface water and, because the quality of surface water is seldom considered a serious threat to
groundwater quality, because it is considered to be filtrated and diluted as it enters the aquifer.
Conversely, surface waters are considered more vulnerable to quality degradation due to their easy
accessibility. The aquifer salinization is the most widespread phenomenon of groundwater
contamination (Kim et al., 2003) and may occur when groundwater levels are abnormally low. Climate
change aggravates this problem in arid and semi-arid regions due to predicted future scenario in
which the temperature and groundwater demand increase and precipitation decreases. Groundwater
resources are considered to be under increasing pressure due to large abstraction rates for various
water-consuming activities (e.g. agriculture activity such as irrigation), public water supply
(consumption) and industry (Stigter et al., 2009). Reductions in recharge can become very significant
due to acceleration in groundwater pumping in the following decades (Santos et al. 2002, Giorgi 2006:
Jackson et al., 2011). The combined effect of prolonged and large extractions from the aquifer and
reduced recharge in coastal areas can lead to seawater intrusion, a serious problem worldwide,
including the Mediterranean countries.
Groundwater ecology is a developing science in the last few decades, dealing with structural and
functional aspects of the organisms inhabiting the water subsurface and with the relationships
between these organisms and their surrounding aquatic environment (Danielopol et al., 2007). For
many years, groundwater was considered unlikely to be a habitat for diverse invertebrate
assemblages due to ‘extreme environment’ associated with absence of sunlight. However, recently
this hypothesis has been challenged and it is acknowledged that biological diversity of groundwater is
much greater than formerly appreciated and much more widespread (Humphreys, 2009; Danielopol et
al. 2001; Danielopol and Pospisil 2001). In 1986, Botosaneanu listed about 7000 groundwater species
worldwide (Ferreira et al., 2007). However, the number of subterranean taxa is likely to be severely
underestimated because of the lack of investigations in this area and the lack of taxonomic expertise
for several subterranean groups. In the groundwater ecosystem, different animal types occur and
there are several classification frameworks of groundwater invertebrates in the literature. A most
accepted classification considers their preferred habitat and resultant behavioural, physiological and
morphological features (Gibert, 2001). Three categories are distinguished in this classification
including (i) Stygoxenes- organisms that have no affinities with groundwater systems and but
occurring there accidentally, (ii) Stygophiles- encompass a number of species that actively exploit the
resources of the groundwater environment for part of their life cycle and, (iii) Stygobites- who are
specialised subterranean forms that complete their whole life cycle exclusively in subsurface water

52
(Gibert, 2001). The risk of species extinction is expectedly high for Stygobites because they have a
narrow distribution range and are more in danger in face of the increase in multiple anthropogenic
pressures (Ferreira et al., 2007). The biogeographical distribution of groundwater fauna is different
around Europe and in southern Europe, where the climate stayed moderate during the various
glaciation periods; most of the old Tertiary fauna still persists. These regions are characterised by a
diverse, endemic, exclusively subterranean dwelling fauna (Pipan and Culver, 2007). In Europe, karst
groundwater systems contain a pervasive and significant fauna that compares with some of the
richest sites known anywhere in the world (Boulton et al., 2003; Danielopol et al., 2007). According to
Galassi (2009), crustaceans represent about 10% of the total number of invertebrate species known
from fresh water world-wide, of which Copepoda have 2814 described species, Ostracoda about 1936
species and Amphipods 1870 species, being by far the most species-rich crustacean groups (Dole-
Olivier et al., 2000; Galassi et al., 2009; Stoch and Galassi, 2010). The stygofauna in temperate
regions is dominated by crustaceans (65% of species) and molluscs (22%). Temporal trends in the
cumulative number of groundwater and surface water species suggested that groundwater comprised
more crustaceans than surface freshwater (Ferreira et al., 2007).
The qualitative and quantitative status of groundwater is assessed based on the chemical and
hydrogeological parameters (Steube et al., 2009). However, knowledge on the drivers for the often
patchy distribution of groundwater fauna is poor, which limits predictive conclusions. Besides that,
there is a lack of sufficient information on the taxonomy, autecology and physiology (e.g. sensitivity to
pollutants) with respect to groundwater organisms. Nevertheless, several authors have already
stressed the use of groundwater invertebrates for biomonitoring purposes and the assessment of the
ecological status (Steube et al., 2009; Bright et al., 1998; Claret et al. 1999). Although physical–
chemical monitoring and routine analyses cover most of the contaminants relevant for human health,
there are number of arguments which support the complementary use of bioindicators to assess
subterranean resources including i) Detection of effects by short-term (pulses of) contaminant impacts
because it is possible to collect organisms in short-term, mid-term and long-term impacts over time
can reflect an ‘integrative’ picture of water quality and ecosystem status, ii) Indirect detection of ‘new’
pollutants at low but ‘chronic’ concentrations which are not yet detectable by other methods (Steube
et al., 2009), iii) Evaluation of the hydrological connection between the surface and the aquifer. In
fact, groundwater fauna reflect structural conditions of their habitat such as hydraulic conductivity,
heterogeneity of habitats in an aquifer and provide information on surface/subsurface hydrological
exchanges (Danielopol et al., 2007). Recently, environmental policy has started to consider
groundwater not only as a resource for high quality water but as a living ecosystem. In particular, the
Swiss Water Protection Ordinance (GSchV, 1998) and the EU Groundwater Directive (EU-GWD,
2006) mention ecological objectives to keep the biocoenosis in a natural state and to foster research
in groundwater ecology, respectively (Steube et al., 2010). In 2002, the European Commission
launched the PASCALIS programme (Protocols for the Assessment and Conservation of Aquatic Life
in the Subsurface), aiming to improve the evaluation of GW biodiversity and to develop guidelines and
strategies for its conservation in six European regions (Gibert, 2001). Assessing the biodiversity and
abundance response of groundwater fauna sampled in aquifers of differing groundwater

53
characteristics allows evaluating impacts such as seawater intrusion on the vulnerable groundwater
ecosystem. Knowledge on groundwater biodiversity is currently biased towards penetrable karstic
habitats (caves), whereas other common habitats such as those found in porous aquifers have been
neglected! Planning an efficient sampling strategy is strongly constrained by difficulties in accessing
the subterranean realm, especially in deep phreatic habitats. As a consequence, sometimes it may
not be possible to distribute samples sites where it would be necessary, but only where access is
possible through a limited number of outputs (springs, resurgences) or few “windows” (caves,
wells)(Gibert, 2001). Within an existing fragmented frame of knowledge of subterranean biodiversity,
the present contribution aimed to (i) study the community structure of stygofauna sampled in wells
and, (ii) its relation to groundwater condition, i.e. salinity at the Central Algarve. Reliable assessments
of groundwater biodiversity are urgently needed to resolve current issues relating to the protection of
aquifers.

3.3
3.3.1
The stu
prelimina
the stud
groundw
(establis
selected
occurred
and one
Fig.3.1 a
3.3.2
Tempera
portable
measure
the cond
probes
measure
Methods
Studyare
udy area is lo
ary well inve
dy aims wer
water saliniti
shed with d
d: high-salini
d in Decemb
e spring was
and wells 2,
Figure 3.1‐
Fieldabio
ature (°C), d
e meters at
ed with fixed
ductivity mea
installed an
ements.
s
ea:
ocated in the
entory was m
re selected.
es so as to
ata loggers)
ty wells-EC
er 2010. At l
selected for
3, 22, 23, 25
Location of the
oticmeas
dissolved ox
each samp
probes in a
asurements
d their valu
e discharge a
made in study
The curren
o detect res
) of several
> 6000 μS/
east two we
r sediment sa
5 and 26 wer
e inventoried w
surements
xygen (mg.l-
pling occasi
long term su
value for ev
ue in the ta
area of the Q
y zone and w
t stygofauna
sponses to
l wells was
/cm, and low
lls per salinit
ampling. The
re sampled.
wells. Wells num
s
1 O2) and p
ion (Table
urvey for wel
very 6 minut
able is the a
Querença-Sil
wells with the
a status was
this parame
considered
w-salinity we
ty category w
e location of
mber 2,3, 22,23
pH were me
3.1). Elect
lls number 1
tes. Wells nu
average valu
ves aquifer.
e most suitab
s evaluated
eter. The a
and two w
ells -EC< 14
were sampled
the inventor
3,25 and 26 we
easured dire
trical condu
, 2 4 and 5. T
umber 3 and
ue of sever
In an initial
ble characte
in wells of
average yea
well categor
400 μS/cm. S
d, in a total o
ried wells is
ere sampled.
ectly in the f
ctivity (µm/c
The probes
d 6 did not
ral in situ re
54
phase, a
ristics for
differing
r salinity
ies were
Sampling
of 6 wells
shown in
field with
cm) was
recorded
have the
egistered

Table3.
W
S
3.3.3 Two sam
phreatob
(WSSD)
deep or
is descri
75
Mecha
.1‐. Wells abiot
ell code
Q5
Q1
Q3
Q6
Q4
Q2
Spring
Sampling
mpling device
biological ne
) developed
have shallow
ibed in Fig.3
5 cm
40µm
talic ains
tic and morpho
“figure 1” by co
PH
7.75
7.44
7.49
7.58
7.03
7.37
7.25
gdesign:
es were used
et for sampl
by the CVR
w water. The
.2 (Cvetkov
Figur
ologic character
odes 2, 3, 22, 2
Depth (m)
2
3
3.65
>30
>10
4
n.a.
d to assess g
ing in large
M team for d
e phreatobiol
1968, Gibert
Me
fr
20µm
Plasti
re 3.2‐ Phreato
ristics. Wells n
23, 25 and 26 re
EC (µm/cm)
12987
8162
6100
848
1390
9113
759
groundwater
diameter w
directly colle
logical net w
t, 2001).
etalic circular
rame 40cm
ic valve
obiological net
umbers 1, 2,3,4
espectively. n.a
Dis. Oxi.(mg/l)
1.5
4.4
0.88
6.15
3.61
3.72
6.38
r fauna asso
wells and a W
ecting sedim
which can onl
and its deploy
4,5 and 6 refers
a.‐not available
Temp(C)
15.3
19.63
15.71
15.43
17.6
16.2
20
ciated with th
Well Sedime
ent in the we
y be used in
ment.
s to wells illust
e.
p Hyd.h(m
1.7
3 1.3
1 3
3 n.a
5.6
0.9
n.a.
he well sedim
ent Sampling
ells that are
n large diame
55
trated in
ead )
.
5
_
ment: the
g Device
not very
eter wells

This net
(PASCA
circular f
to rubbe
exit. Th
moveme
encomp
column.
the well
PASCAL
sampling
develope
directly c
of benth
of the W
Decemb
The Phr
to collec
this dev
After ea
material
was add
were tak
t is an ackno
ALIS project)
frame of 40 c
er) which allo
he net creat
ents of the n
asses weigh
According t
l at least 10
LIS and sev
g stygofaun
ed another
collected. Th
hic animals. T
WSSD net is
ber 2010.
reatobiologic
ct sediment f
ice. Three re
ch replicate
stored into
ded to the co
ken for which
owledged sam
). As it is ill
cm diameter
ows the swim
tes an asce
net, subsequ
hts that distu
to the PASC
0 times thro
veral other w
a is verified
net- sample
his device wa
The use of tw
s illustrated i
al net was u
from the disc
eplicated net
collection, th
sealed conta
ontainer to p
h fauna ident
Figure3.3
mpling metho
ustrated in F
r. At the base
mming fauna
ending curre
uently captu
urb the sedi
CALIS projec
ough the ent
works (Giber
d for large d
r (WSSD) fo
as designed
wo different
n Fig 3.3. S
sed to collec
charge point
t deploymen
he net collect
ainers and fix
osteriorly sim
tification and
‐ Well Sedimen
od, with exis
Fig.3.2 the 4
e of the mes
a and the ben
ent in the w
uring the sus
iment and b
ct recommen
tire water co
rt, 2001) th
diameter we
or shallow w
so as to min
methods allo
Samplings co
ct samples in
of a spring a
ts and at lea
ting glass wa
xed with 97%
mplify the so
d quantificatio
nt Sampling De
ting samplin
40µm mesh
h cone there
nthic fauna t
well by suc
spended ani
bring the ani
dation (Gibe
olumn. With
he efficiency
ells. Howev
wells in whic
nimise the dis
owed an effic
ollections fro
n 6 wells, wh
and from 1 w
ast one WSS
as washed in
% ethanol al
orting of colo
on was made
evice and its de
g protocols a
cone is mo
e is a valve (a
o be collecte
cessive upw
mals. The lo
mals of sed
ert, 2001) the
in the frame
of the phre
ver, in the p
h the bottom
sturbance, an
ciency comp
m the wells
hile the WSS
well where its
SD replicate
nto the 50µm
cohol. The ro
ured fauna.
e.
ployment.
at the Europ
ounted on a
aluminium co
ed and preve
ward and d
ower end of
diment into t
e net must d
ework of the
eatobiologica
present wor
m sediment
nd potential
parison. The
were carrie
SD net was o
s depth allow
were taken
m mesh sieve
rose Bengal
In total, 20
56
ean level
a metallic
onnected
ents their
ownward
f the net
he water
draw into
e project
al net for
rk it was
could be
damage,
structure
ed out on
only used
wed using
per well.
e and the
chemical
samples

57
3.3.4 Wellsfaunaassessment
The Crustacean was selected as the target group because they are the dominant groups of the
stygofauna in temperate regions (65% of species). The presented study considered only the factor
salinity subdivided in high salinity and low salinity wells. This design does not allow withdrawing
conclusions on temporal patterns as no replicated were measured within along the time period. Three
replicates were taken at each location and time combination. Multifactorial analyses were made in the
PRIMER+ package (Clarke and Warwick, 2007; Anderson et al., 2008). The similarity percentage
breakdown procedure (SIMPER) (Clarke & Warwick, 1994) was used to determine the contribution of
individual taxa to the dissimilarity between and within the factors, and also to identify the species most
contributing for significant factorial differences. MDS and dbRDA ordination techniques were
calculated to graphically examine the samples distribution pattern. For all tests, the Bray-Curtis
similarity measure, the fourth-root transformation and 9999 permutations were used for all biotic data,
while for abiotic data was normalized and the Euclidean distance measure used in the PRIMER+
calculations. The fourth root transformation was applied to give more weight to the rare species and
reduced the weight of the dominant species. Obligate stygofauna species were separated from the
stygoxene and stygophile species. The wells with easy accessibility to bottom sediment due to its
shallow depth were selected to be sample of the two different devices (Phreato net and WSSD net)
allowing a comparison of fauna structure and composition.
3.4 PreliminaryResultsandinterpretation
3.4.1 Faunageneralcharacterization
A total of 612 crustacean organisms were sampled in total. A total number of 19 species was
identified and corresponds to brackish water organisms, Stygobites, Stygoxenes and Stygophiles of
the identified fauna corresponds to Amphipods, Copepods and Ostracod (Table 3.2.). The Order
Cyclopodia from the class Copepod was the most dominant taxa. With 53 families, the order
Cyclopodia revealed to be the most diverse taxa with 11 identified species in the present study. The
taxa richness for the wells was highest at the well number 4 with also had the lowest salinity recorded.
Faunal assemblages were dominated by Stygobites species and Stygophiles and/or Stygoxenes
never contained more than 32% of the total number of invertebrates.
The order Harpacticoida is well adapted to benthic life because they are typical crawlers, walkers, and
burrowers (Galassi et al., 2009). They can be found in freshwater and in saline water ecosystems
depending on the species. Here, Harpacticoida were found in the spring sampling location and in the
well Q1. The spring was sampled with the WSSD net, considered more likely to collect this type of
benthic animal. Well Q1 and 2 had always high salinity (> 7000 µs/cm) comparing to the other
sampling wells, potentially because they are closer to the Arade estuary.The Halicyclops sp. is a
cosmopolitan Cyclopodia genus of the family Cyclopidae, widely distributed in several kinds of surface
brackish water bodies (Chang, 2012).

58
Table 3.2‐ Identified species of Cyclopodia, Ostracod, Amphipod and Copepod sampled from wells. Shaded cells represent absence and unshaded cells represent a taxa absence.
Q1 Q2 Q3 Q4 Q5 Q6 Spring
Cyclopodia
33 0 34 48 209 42 6
Halicyclops sp.
Eucyclops graeteri (Chappuis 1927)
Eucyclops serrulatus (Fischer, 1851)
Eucyclops hadjebensis
Eucyclops speratus
Megacyclops brachypus (Kiefer, 1954)
Acanthocyclops sp.
Acanthocyclops sensitivus
Megacyclops viridis
Macrocyclops sp.
Macrocyclops albidus (Jurine, 1820)
Ostracoda
2 0 31 171 6 1 1
Cypria ophtalmica (Jurnine 1820)
Cypridopsis vidua (O. F. Müller, 1776)
Bradleycypris oblique (Brady, 1868)
Amphipoda 1 12 1 1 0 3
Gammaridae
Sphroma sp.
Gammarus sp.
Gammarus pulex (Linnaeus,1758)
Copepoda 2 2 3
Harpacticoida

Its occur
The gen
Eucyclop
number
of individ
influence
Eucyclop
found in
being a
moderat
Acantho
and Woj
setae an
6 wells
(stygoxe
Howeve
Cypridop
family G
pulex is
(oxygen
1990). In
rrence in we
nus Eucyclo
ps graeteri i
Q5 and Q6)
duals. Well n
e by fresh/gr
ps speratus
n the spring
a potential i
tely eutroph
ocyclops sen
jtasik, 2009)
nd antennal
. The most
ene, stygoph
er, they were
psis vidua) a
Gammaridae.
a freshwate
and ammon
n Fig. 3.4 so
ell Q1 highlig
ops was the
s a hypogea
). This was th
number Q4 h
roundwater.
were found
location. Th
indicator of
ic water, bu
nsitivus found
). Ostracods
swimming b
t dominant
hile). They w
e more num
are also epig
They have
r species tha
nia), inorgan
me pictures
hts the fact t
dominant s
an (Stygobite
he most abu
had the lowe
The two Sty
d in the men
is species w
the presen
ut it can als
d only in wel
are swimm
ristles for pro
Ostracod s
were found in
merous in we
gean (stygox
wide distribu
at is sensitive
nic ions (pH)
representing
that this hab
specimens o
es) found in
ndant specie
st salinity re
ygobite Eucy
ntioned well.
was also fou
nce of grou
so occur in
ll Q4, is a co
ers or crawl
opulsion. Th
species was
n wells with
ell Q4. The
xene, stygop
ution and can
e to several
) and organic
g stygofauna
itat is influen
of the subfa
wells with d
es which con
cord indicatin
yclops hadjeb
. The Eucyc
nd in the low
undwater. M
groundwate
ommon grou
lers. Swimm
ree species
s Cypria op
different ch
other two s
hile). All Am
n survive wid
water polluti
c compound
can be obse
nced by estu
mily Eucyclo
different salin
ntributed 37%
ng that this w
bensis (semi
clops speratu
w salinity we
Megacyclops
er as a styg
ndwater spe
ing species
of ostracod w
phtalmica, a
haracteristics
species (Cyp
phipod taxa
de range of s
on including
ds (dichloroa
erved.
arine brackis
opinae. The
nity categori
% of the tota
well is under
ibenthic spec
us species w
ells (Well Q4
viridis can
goxen (Reid
ecies (Miodu
use their an
were identifi
an epigean
s in terms of
pridopsis vid
corresponde
salinities. Ga
g metals (zinc
aniline) (Malt
59
sh water.
species
es (wells
l number
r a strong
cies) and
was also
4), hence
tolerate
d, 2001).
chowska
ntennular
ed for all
species
f salinity.
dua and
ed to the
ammarus
c), gases
by et al.,

Figure3serrulatus
3.4.2
The spe
Table3
So
SaLoRTo
The MD
categori
separate
the rem
species)
Figure 3
3.4‐some repres / Cypridopsis
Speciesd
ecies distribut
3.3‐ Permanova
ource
a o x Sa es otal
DS ordination
es (Fig 3.5)
ed in the ord
aining low s
).
3.5‐MDS ordinaeach well
esentive Stygofas vidua/ Harpac
distributio
tion was sign
a analysis of thenumber of
n showed tha
) Wells num
dination plot.
salinities poin
ation of wells f(R1‐R3)groupe
auna, sequentlcticoida/ Macr
on
nificantly exp
e wells fauna fopermutations u
df S
1 2
4 2
12 3
17 6
at the commu
mber 4 (Q4)
However o
nt. This is m
fauna samples ed into two gro
ly from left up rocyclops albidu
plained the s
or factors salinused was 9999
SS
2198 2
28243 7
30577 2
61016
unity structu
and 6 (Q6
ne replicate
mostly due to
collected fromups of salinity
to right down: us/ Eucyclops /
alinity gradie
nity (high and lo9 (Sa= salinity,L
MS Pse
2196 0,3
7060 2,7
2548
re varied be
), which are
(replicate 3)
o the low nu
6 different wecategories (H:
Gamarus pulex/ Megacyclops
ent (Table 3.
ow) and differeo‐ location).
eudo‐F P. (per
31102 1
771 0,00
tween wells
e wells with
) from well Q
umber of tax
ells (Q1‐Q6), wihigh and L:low
ex/Eucyclops seviridis/ Gamm
.3).
ent wells locati
rm) U. perms
15
001 9903
with differen
low saliniti
Q6 is separa
xa in this rep
ith three replicw salinities).
60
errulatus arus pulex
on. The
s
nt salinity
es, were
ated from
plicate (1
ate from

61
The SIMPER analysis reflected that there was a relatively high average dissimilarities (81,66)
between two group of wells based on their salinity properties. Within the context of this test, the
species that contributed the most for significant differences between two groups of wells are identified
in Table 3.4.
Table 3.4‐ SIMPER analysis identifying the species that contributed the most for differences between wells with high and low salinities
G.High G.Low
Av.Abund Av.Abud Av.Diss Diss/SD Contrib% Cum.%
Cyclopodia 3.26 2.81 20.02 1.35 24.52 24.52
Cypria
ophtalmica
0.70 3.12 18.11 0.73 22.18 46.70
Eucyclops
graeteri
2.31 0.17 11.07 0.79 13.56 60.26
Total Amphipod 0.64 0.57 7.04 0.66 8.62 68.88
Macrocyclops sp. 0.00 1.07 4.91 0.43 6.01 74.89
The taxa which contributed the most for dissimilarities between wells group were the total number of
Cyclopodia (25%) which, together with the species Cypria ophtalmica and Eucyclops graeteri
represented 50% of the dissimilarities. Cyclopodia were more numerous in the high salinities wells
which can be due to having more sampled wells with high salinities. However, the epigean Ostracod
Cypria ophtalmica was more abundant in low salinities wells. The obligate groundwater species
Eucyclops graeteri had higher abundance in the high salinities wells and, may have the same
justification as species Cypria ophtalmica. But here this species is very abundant in well Q5 which has
been characterised as high salinity well. The times series of electrical conductivity recorded periods
with very high salinities over 40000 µs/cm (months October and November) and relatively very low
salinity less than 2000 µs/cm (mostly in summer periods).The wide range of salinity variation in the
well may explain the presence of this true groundwater species in periods that have low salinity
measurements.
3.4.3 Samplingdevicecomparison
Well Q5 was sampled using the Phreato net and WSSD net methods. Three replicates were collected
with each device. Care was taken to use 1st the almost not disturbing WSSD so as to have
representative sub sequential nets samples. Also, the well was very wide, allowing sampling different
well areas with the two methods. The Total number of individuals, number of taxa, diversity and
richness captured with both devices were compared. The total number of individual was higher in
samples collected by the Phreato net; however the number of taxa was higher in samples collected by
WSSD device. Cyclopodia were more diverse in samples taken with the Phreato net while Ostracod
and Harpacticoida were more diverse in WSSD samples. This can be justified by the fact that
Harpacticoids and ostracods are well adapted to benthic life while Cyclopoids are generally good
swimmers (Dole-Oliver et al., 2000).

62
Table3.5‐ Taxa abundance sampled by Phreatobiological net and WSSD.
Replicates R1 R2 R3
Phreato net
Eucyclops graeteri 103 86 17
Megacyclops brachypus 2
Acanthocyclops sp. 1
Bradleycypris obliqua 6
WSSD net
Eucyclops graeteri 2
Acanthocyclops sp. 1
Harpacticoida 2
Cypria ophtalmica 2
Cypridopsis vidua 1

63
Chapter 4: Main conclusion and future works

64
4.1 MainConclusionsandfutureworks
This study initiates baseline data on stygofauna and benthic invertebrate, interfacing different saline
condition. Biological indicators can be a functional tool to predict climate change effects if we
understand the factors influencing the habitat communities.
Within the first experiment evidence for estuarine fauna to be considered as a potentially valuable
indicator of variation in the input of groundwater into marine-freshwater interface habitats were
provided. The shifts in precipitation and temperature regimes predicted by climate change will lead to
enhancement in the demand on groundwater, altering the hydrologic balance on the marine-
groundwater interface such as in estuaries. Substantial aquifer exploitation threatens the wetlands
that constitute groundwater dependent ecosystems. The habitat alteration may affect the communities
from primary producers such as macrophytes communities until the higher trophic level such as fish
communities .Hence, benthic community identification and protection before any anthropogenic
disturbance are highly recommended (Seilheimer et al., 2009). Regarding the presence of
macroinvertebrates in the sediment of the Arade estuary in the Central Algarve, diversity was
relatively high with a total of 38 taxa being identified. Results indicate that there was a qualitative and
quantitative response of macroinvertebrate community to the salinity gradient inherent to the sampling
locations in the channel: those which tolerate low salinity were the most abundant at the location of
groundwater input, locations A, B, C. There were clear differences in Electrical Conductivity
(considered a surrogate for salinity) between summer and winter, with higher influence of
groundwater discharge into the channel during winter. The macroinvertebrate community structure
responded to this seasonal hydrological variation. Groundwater input into estuarine systems can
therefore be a key bottom-up factor regulating both food webs and biodiversity levels. If the
groundwater outflow is reduced, as predicted by aquifer modelling under climate change, then the
composition and abundance of the faunal community is likely to vary. As these organisms adapt over
time to disturbance and display different biological sensitivities between species, they give an indirect
method to evaluate changes of environmental features and generalised impairments. It is crucial that
future monitoring programmes include an assessment of the spatial and temporal variability of the
species identified here as potential responders to groundwater influence, because it allows assessing
interferences from external factors such as pollution originated from the main estuarine channel or
shifts in groundwater availability into the estuary.
The results from the second experiment were indicated that stygofauna can be potential indicators to
predict changes in the biodiversity in situations where a reduction or deterioration in groundwater can
significantly impact biodiversity. Recent climate change and groundwater exploitation have a strong
impact on groundwater quality. Saline intrusion into the groundwater resources is one of the
consequences. This situation has a great impact on local social and economic systems and the loss
of regional biodiversity and aquatic refuges (Mischkea et al., 2012). The results of the present study
allowed for understanding the general stygofauna inhabiting in the wells receiving recharge water
from Querença-Silves aquifer. There has been a resistance in using biological indicators to assess

65
the groundwater conditions. This resistance reflects concerns about sampling costs, taxonomic
constraints, the lack of baseline data for comparison, and uncertainty about how to interpret
stygofauna data (Boulton, 2009).This work initiate the study on stygofauna in south of Portugal in the
hope of overcoming these restrictions and, providing baseline data for conservation planning for
future years. Within this experiment from well number 6, with low salinity condition some new species
were collected. They are small amphipods without eyes and have been identified to genus taxonomic
level and their identification into species level has not been completed since they are suspected to be
new to the science. In the climate scenario of decreased groundwater net recharge and intrusion of
brackish estuaries water into the aquifer, a shift in both identity and abundance of Stygofauna can
now be expected. This is in agreement with Boulton (2009) who indicate that there is a marked
difference among species in their response to water level changing (preliminary laboratory test).
Moreover, the ecological attributes of stygofauna makes them vulnerable to changes in habitat, which,
combined with their taxonomic affinities, makes them a significant issue to biodiversity conservation
(Humphreys, 2006).
In the same way, with this study we can start to understand some of the environmental factors such
as salinity and aquifer structure that correlate with groundwater fauna, particularly for the species
Eucyclops speratus, Eucyclops hadjebensis, Megacyclops viridis and Acanthocyclops sensitivus,
which were particularly associated with low salinity conditions, hence being potential indicators for
saline intrusion if their abundances decrease greatly.
The identification of this specialized fauna was made in collaboration with Dr. Sanda Iepure at the
Instituto Madrileño de Estudios Avanzados in Madrid, and its currently ongoing for the rest of the taxa,
taking longer than expected due to the detection of possible new species and due to the required
expertise to reach the ambitioned species level.

66
Bibliography Ajami, H., Hogan, J., Maddock, T., Meixner, T., Troch, P., 2008. Quantifying spatial and temporal variability of mountain system recharge in semi-arid catchments, in:AGU Fall Meeting Abstracts, p. 1011. Alden, R., Weisberg, S., Ranasinghe, J., Dauer, D., et al., 1997. Optimizing temporal sampling strategies for benthic environmental monitoring programs. Marine Pollution Bulletin 34, 913–922. Ambrogi, A.O., Forni, G., 2004. Biotic indices. Biologia Marina Mediterranea 11 (Suppl. 1), 545-572. Anderson, M.J., Gorley, R.N., Clarke, K.R., 2008. PERMANOVAþ for PRIMER: Guide to Software and Statistical Methods. PRIMER-E Ltd, Plymouth. Attrill, M., Power, M., 2000. Effects on invertebrate populations of drought-induced changes in estuarine water quality. Marine Ecology Progress Series 203, 133–143. Bates, B., Kundzewicz, Z., Wu, S., Palutikof, J., et al., 2008. Climate change and water. Intergovernmental Panel on Climate Change (IPCC). Barros, M., Mendo, M., Negrao, F., 1995. Surface water quality in portugal during a drought period. Science of the total environment 171, 69–76. Bear, J., 1979. Hydraulics of ground water. Borja, A., Franco, J., P´erez, V., 2000. A marine biotic index to establish the ecological quality of soft-bottom benthos within european estuarine and coastal environments. Marine Pollution Bulletin 40, 1100–1114. Borja, A., Miles, A., Occhipinti-Ambrogi, A., Berg, T., 2009. Current status of macroinvertebrate methods used for assessing the quality of european marine waters:implementing the water framework directive. Hydrobiologia 633, 181–196. Bork, J., Berkhoff, S., Bork, S., Hahn, H., 2009. Using subsurface metazoan fauna to indicate groundwater–surface water interactions in the nakdong river floodplain, south korea. Hydrogeology Journal 17, 61–75. Boulton, A., Scarsbrook, M., Quinn, J., Burrell, G., 1997. Land-use effects on the hyporheic ecology of five small streams near hamilton, new zealand. New Zealand journal of marine and freshwater research 31, 609–622. Boulton, A., Findlay, S., Marmonier, P., Stanley, E., Valett, H., 1998. The functional significance of the hyporheic zone in streams and rivers. Annual Review of Ecology and Systematics , 59–81. Boulton, A., Humphreys, W., Eberhard, S., 2003. Imperilled subsurface waters in australia: biodiversity, threatening processes and conservation. Aquatic Ecosystem Health & Management 6, 41–54. Boulton, A., 2009. Recent progress in the conservation of groundwaters and their dependent ecosystems. Aquatic Conservation: Marine and Freshwater Ecosystems 19, 731–735. Bright, J., Bidwell, V., Robb, C., Ward, J., 1998. Environmental performance indicators for groundwater. Report No 4306/1, May 1998.Chang, C., 2012. Discovery of halicyclops continentalis (cyclopidae, halicyclopinae) from estuaries and salt marshes on the west coast of south korea. Animal Systematics, Evolution and Diversity 28, 12–19. Cabral, H., 2000. Comparative feeding ecology of sympatric solea solea and s.senegalensis, within the nursery areas of the tagus estuary, portugal. Journal of Fish Biology 57, 1550–1562. Claret, C., Marmonier, P., Dole-Olivier, M., Castella, E., 1999. Effects of management works on the interstitial fauna of floodplain aquatic systems (river rhone, france). Biodiversity and Conservation 8, 1179–1204. Castaneda, E., Drake, P., et al., 2008. Spatiotemporal distribution of lekanesphaera species in relation to estuarine gradients within a temperate european estuary (swspain) with regulated freshwater inflow. ecological indicators 34, 125–141. Chainho, P., Costa, J.L., Chaves, M.L., Lane, M.F., Dauer, D.M., Costa, M.J., 2006. Seasonal and spatial patterns of distribution of subtidal benthic invertebrate communities in the Mondego River, Portugal - a poikilohaline estuary. Hydrobiologia, 555.59-74.

67
Chainho, M., 2008. Contribution to the development of biotic integrity assessment tools for Portuguese estuaries based on benthic communities. Ph.D. thesis. Universityof Lisbon, Portugal. Chang, C., 2012. Discovery of halicyclops continentalis (cyclopidae, halicyclopinae) from estuaries and salt marshes on the west coast of south korea. Animal Systematics, Evolution and Diversity 28, 12–19. Chapman, V., 1960. Salt marshes and salt deserts of the world. London: Leonard Hill (Books) Ltd. (Interscience Publ., Inc., New York). Chapman, P., Brinkhurst, R., 1980. Salinity tolerance in some selected aquatic oligochaetes. Internationale Revue der gesamten Hydrobiologie und Hydrographie 65, 499–505. Clarke, K.R., Warwick, R.M., 2007. In: Plymouth Marine Laboratory (Ed.), PRIMER-E, Version 6 (U.K.). Coulson, D., Joyce, L., 2006. Indexing variability: A case study with climate change impacts on ecosystems. Ecological Indicators 6, 749–769. Cvetkov, L., 1968. Un filet phr´eatobiologique. Bull. Inst. Zool. Mus. Acad. Bulgare. Sci., Sofia 27, 215–218. Dale, V., Beyeler, S., 2001. Challenges in the development and use of ecological indicators. Ecological indicators 1, 3–10. Danielopol, D., Pospisil, P., Rouch, R., 2000. Biodiversity in groundwater: a large scale view. Trends in ecology & evolution 15, 223. Danielopol, D., Pospisil, P., 2001. Hidden biodiversity in the groundwater of the danube flood plain national park (austria). Biodiversity and Conservation 10, 1711–1721. Danielopol, D., Griebler, C., Gunatilaka, A., Notenboom, A., et al., 2003. Present state and future prospects for groundwater ecosystems. Environmental Conservation 30, 104–130. Danielopol, D., Griebler, C., Gunatilaka, A., Hahn, H., Gibert, J., Mermillod- Blondin, G., Messana, G., Notenboom, J., Sket, B., 2007. Incorporation of groundwater ecology in environmental policy. Groundwater Science and Policy, London The Royal Society of Chemistry, RSC Publishing . Dauer, D.M. 1993. Biological criteria, environmental health and estuarine macrobenthic community structure. Marine Pollution Bulletin 26, 249–257. De Pauw, N., Ghetti, P., Manzini, P., Spaggiari, D., 1992. Biological assessment methods for running water. River water quality, Ecological assessment and control , 217–248. Díeza, I., Bustamante, M., Santolaria, A., Tajadura, J., Muguerza, N., Borja, A., Muxika, I., Saiz-Salinas, J., Gorostiaga, J., 2012. Development of a tool for assessing the ecological quality status of intertidal coastal rocky assemblages, within atlantic iberian coasts. Ecological Indicators . Dole-Olivier, M., Galassi, D., Marmonier, P., Creuz´e des Chˆatelliers, M., 2000. The biology and ecology of lotic microcrustaceans. Freshwater Biology 44, 63–91. Eamus, D., Froend, R., 2006. Groundwater-dependent ecosystems: the where, what and why of gdes. Australian Journal of Botany 54, 91–96. Elliott, M., Quintino, V., 2007. The estuarine quality paradox, environmental homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Marine Pollution Bulletin 54, 640–645. Erwin, K., 2009. Wetlands and global climate change: the role of wetland restoration in a changing world. Wetlands Ecology and Management 17, 71–84. EUCYCLOPINAE, K., EUCYCLOPS, C., Cyclopoida. available at http://www.luciopesce.net/copepods/eucyc2.htm. Euliss, N., LaBaugh, J., Fredrickson, L., Mushet, D., Laubhan, M., Swanson, G., Winter, T., Rosenberry, D., Nelson, R., 2004. The wetland continuum: a conceptual framework for interpreting biological studies. Wetlands 24, 448–458. Ferreira, D., Malard, F., Dole-Olivier, M., Gibert, J., 2007. Obligate groundwater fauna of france: diversity patterns and conservation implications. Biodiversity and Conservation 16, 567–596.

68
Ferreira, J., Vale, C., Soares, C., Salas, F., Stacey, P., Bricker, S., Silva, M., Marques, J., 2007. Monitoring of coastal and transitional waters under the eu water framework directive. Environmental monitoring and assessment 135, 195–216. Galassi, D., Huys, R., Reid, J., 2009. Diversity, ecology and evolution of groundwater copepods. Freshwater Biology 54, 691–708. Gamito, S., Patr´ıcio, J., Neto, J., Teixeira, H., Marques, J., 2012. Feeding diversity index as complementary information in the assessment of ecological quality status. Ecological Indicators . Gibert, J., 2001. Protocols for the assessment and conservation of aquatic life in the subsurface (pascalis): a european project. Mapping Subterranean Biodiversity , 19–21. Giorgi, F., 2006. Climate change hot-spots. Geophysical Research Letters 33, L08707. Goncalves, J., Veiga, P., Machado, D., Bentes, L., Monteiro, P., Coelho, R., Ribeiro, M.R.J., Erzini, K., 2004. Fish communities of the arade estuary. Poster presen- 3 tation in the Symposium IO-ECSA Towards an Integrated Knowledge for the Management of Estuarine Systems, FC/IO, Lisbon. Gower, J., 1966. Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 53, 325–338. Grilo, T., Cardoso, P., Dolbeth, M., Bordalo, M., Pardal, M., 2011. Effects of extreme climate events on the macrobenthic communities structure and functioning of a temperate estuary. Marine pollution bulletin 62, 303–311. Hahn, H., 2006. The gw-fauna-index: A first approach to a quantitative ecological assessment of groundwater habitats. Limnologica-Ecology and Management of Inland Waters 36, 119–137. Hakenkamp, C. C. and Morin, A., 2000. The importance of meiofauna to lotic ecosystem functioning. Freshwater Biology, 44, 165–175. Hancock, P., Hunt, R., Boulton, A., 2009. Preface: hydrogeoecology, the interdisciplinary study of groundwater dependent ecosystems. Hydrogeology Journal 17,1–3. Hayward, P., Ryland, J., 1995. Handbook of the marine fauna of North-West Europe. Oxford University Press, USA. Humphreys, W., 2006. Aquifers: the ultimate groundwater-dependent ecosystems. Australian Journal of Botany 54, 115–132. Humphreys, W., 2009. Hydrogeology and groundwater ecology: Does each inform the other Hydrogeology Journal 17, 5–21. Iliopoulou-Georgudaki, J., Kantzaris, V., Katharios, P., Kaspiris, P., Georgiadis, T., Montesantou, B., 2003. An application of different bioindicators for assessing water quality: a case study in the rivers alfeios and pineios (peloponnisos, greece). Ecological Indicators 2, 345–360. Jackson, C., Meister, R., Prudhomme, C., 2011. Modelling the effects of climate change and its uncertainty on uk chalk groundwater resources from an ensemble of global climate model projections. Journal of Hydrology 399, 12–28. Jolly, I., McEwan, K., Holland, K., 2008. A review of groundwater–surface water interactions in arid/semi-arid wetlands and the consequences of salinity for wetland ecology. Ecohydrology 1, 43–58. Jones, J., Mulholland, P., 2000. Streams and ground waters. Ecology 88, 727–731. Kim, J., Yum, B., Kim, R., Koh, D., Cheong, T., Lee, J., Chang, H., 2003. Application of cluster analysis for the hydrogeochemical factors of saline groundwater in kimje, korea. Geosciences Journal 7, 313–322. Kimmerer, W.J., 2002. Effects of freshwater flow on abundance of estuarine organisms: physical effects or trophic linkages?. Marine Ecology Progress Series 243, 39–55. Kristensen, E., Anderson, F.,1987. Determination of organic carbon in marine sediments: a comparison of two CHN-analyzer methods. Journal of Experimental Marine Biology and Ecology 109, 15–23.

69
Loaiciga, H., Maidment, D., Valdes, J., 2000. Climate-change impacts in a regional karst aquifer, texas, usa. Journal of Hydrology 227, 173–194. Malard, F., Tockner, K., DOLE-OLIVIER, M.,Ward, J., 2002. A landscape perspective of surface–subsurface hydrological exchanges in river corridors. Freshwater Biology 47, 621–640. Malard, F., Ferreira, D., Dol´edec, S., Ward, J., 2003. Influence of groundwater upwelling on the distribution of the hyporheos in a headwater river flood plain. Archiv f¨ur Hydrobiologie 157, 89–116. Maltby, L., Naylor, C., Calow, P., 1990. Effect of stress on a freshwater benthic detritivore: Scope for growth ingammarus pulex. Ecotoxicology and environmental safety 19, 285–291. Macleod, C., Crawford, C., Moltschaniwskyj, N., 2004. Assessment of long term change in sediment condition after organic enrichment: defining recovery. Marine Pollution Bulletin 49, 79–88. Malcolm, I., Soulsby, C., Youngson, A., Hannah, D., 2005. Catchment-scale controls on groundwater–surface water interactions in the hyporheic zone: implications for salmon embryo survival. River Research and Applications 21, 977–989. Maloney, K., Feminella, J., 2006. Evaluation of single-and multi-metric benthic macroinvertebrate indicators of catchment disturbance over time at the fort benning military installation, georgia, usa. Ecological Indicators 6, 469–484. Medeiros, A., Barbosa, J., Medeiros, P., Rocha, R., Silva, L., 2010. Salinity and freshwater discharge determine rotifer distribution at the mossor´o river estuary (semiarid region of brazil). Brazilian Journal of Biology 70, 551–557. Medeiros, J., Chaves, M., Silva, G., Azeda, C., Costa, J., Marques, J., Costa, M., Chainho, P., 2011. Benthic condition in low salinity areas of the mira estuary (portugal): Lessons learnt from freshwater and marine assessment tools. Ecological Indicators Mioduchowska, M., Wojtasik, B., 2009. Copepoda-cyclopoida of water bodies of the coast of Gdansk. Teka Kom. Ochr. Kszt. Srod. Przyr. OL PAN 6, 189–199. Mischkea, S., Ginat, H., Al-Saqarat, B., Almogi-Labin, A., 2012. Ostracods from water bodies in hyperarid israel and jordan as habitat and water chemistry indicators. Ecological Indicators 14, 87–99. Monteiro, J., Ribeiro, L., Reis, E., Martins, J., Matos Silva, J., 2007. Modelling stream-groundwater interactions in the Querença-Silves Aquifer System Groundwater and Ecosystems (Lisbon).,IAH Congress, Groundwater and Ecosystems, Lisbon, pp. 17–21. Murdoch, P., Baron, J., Miller, T., 2000. Potential effects of climate change on surface-water quality in north america1. JAWRA Journal of the American Water Resources Association 36, 347–366. Murray, B., Zeppel, M., Hose, G., Eamus, D., 2003. Groundwater-dependent ecosystems in australia: It’s more than just water for rivers. Ecological Management & Restoration 4, 110–113. Murray, B., Hose, G., Eamus, D., Licari, D., 2006. Valuation of groundwaterdependent ecosystems: a functional methodology incorporating ecosystem services. Australian journal of botany 54, 221–229. Niemeijer, D., de Groot, R., 2008. A conceptual framework for selecting environmental indicator sets. Ecological indicators 8, 14–25. Nunes, M., Coelho, J., Cardoso, P., Pereira, M., Duarte, A., Pardal, M., 2008. The macrobenthic community along a mercury contamination in a temperate estuarine system (ria de aveiro, portugal). Science of the Total Environment 405, 186–194. Occhipint, A., Forni, G., 2004. Biotic indices. Biologia marina mediterranea 11, 545–572. Pasquaud, S., Pillet, M., David, V., Sautour, B., Elie, P., 2010. Determination of fish trophic levels in an estuarine system. Estuarine, Coastal and Shelf Science 86, 237–246. Pipan, T., Culver, D., 2007. Regional species richness in an obligate subterranean dwelling fauna–epikarst copepods. Journal of biogeography 34, 854–861.

70
Poff, N., Brinson, M., Day, J., 2002. Aquatic Ecosytems & Global Climate Change: Potential Impacts on Inland Freshwater and Coastal Wetland Ecosystems in the United States. Pew Center on Global Climate Change Arlington, Virginia. Prowse, T., Wrona, F., Reist, J., Gibson, J., Hobbie, J., L´evesque, L., Vincent, W., 2006. Climate change effects on hydroecology of arctic freshwater ecosystems. AMBIO: A Journal of the Human Environment 35, 347–358. Ranjan, P., Kazama, S., Sawamoto, M., 2006. Effects of climate change on coastal fresh groundwater resources. Global Environmental Change 16, 388–399. Remane, A., 1934. Die brackwasserfauna. Zoologischer Anzeiger (Supplement) 7, 34–74. Reid, J., 2001. A human challenge: discovering and understanding continental copepod habitats. Hydrobiologia 453, 201–226. Remane, A., Schlieper, C., 1971. Biology of brackish water. volume 25. Halsted Press. Redeke, H., 1935. Acartia (acanthacartia) tonsa dana, ein neuer copepode des niederl¨andischen brackwassers. Archives N´eerlandaises Zoollogie 1, 315–329. Roessig, J., Woodley, C., Cech, J., Hansen, L., 2004. Effects of global climate change on marine and estuarine fishes and fisheries. Reviews in Fish Biology and Fisheries 14, 251–275. Rundle, S., Attrill, M., Arshad, A., 1998. Seasonality in macroinvertebrate community composition across a neglected ecological boundary, the freshwater-estuarine transition zone. Aquatic Ecology 32, 211–216. Salen-Picard, C., Arlhac, D., 2002. Long-term changes in a mediterranean benthic community: Relationships between the polychaete assemblages and hydrological variations of the rhˆone river. Estuaries and Coasts 25, 1121–1130. Sandin, L., Hering, D., 2004. Comparing macroinvertebrate indices to detect organic pollution across europe: a contribution to the ec water framework directive intercalibration. Integrated Assessment of Running Waters in Europe , 55–68. Santos, F., Forbes, K., Moita, R., 2002. Climate change in Portugal: scenarios, impacts and adaptation measures: SIAM Project. Gradiva, Lisbon, Portugal. Seilheimer, T., Mahoney, T., Chow-Fraser, P., 2009. Comparative study of ecological indices for assessing human-induced disturbance in coastal wetlands of the laurentian great lakes. ecological indicators 9, 81–91. Senft, A., 2009. Species diversity patterns at ecotones. Ph.D. thesis. University of North Carolina. Singh, K., Malik, A., Mohan, D., Sinha, S., 2004. Multivariate statistical techniques for the evaluation of spatial and temporal variations in water quality of gomti river (india)–a case study. Water Research 38, 3980–3992. Silva, A.C.F., Tavares, P., Shapouri, M., Stigter, T.Y., Monteiro, J.P., Machado, M., Cancela da Fonseca, L., Ribeiro, L., 2012. Estuarine biodiversity as an indicator of groundwater discharge. Estuarine, Coastal and Shelf Science 97:38 43. Simboura, N., Zenetos, A., et al., 2002. Benthic indicators to use in ecological quality classification of mediterranean soft bottom marine ecosystems, including a new biotic index. Mediterranean Marine Science 3, 77–112. Smith, J., Schellnhuber, H., Mirza, M., Fankhauser, S., Leemans, R., Lin, E., Ogallo, L., Pittock, B., Richels, R., Rosenzweig, C., et al., 2001. Vulnerability to climate change and reasons for concern: a synthesis. Climate change , 913–967. Steube, C., Richter, S., Griebler, C., 2010. First attempts towards an integrative concept for the ecological assessment of groundwater ecosystems. Hydrogeology Journal 17, 23–35. Stigter, T., Monteiro, J., Nunes, L., Vieira, J., Cunha, M., Ribeiro, L., Nascimento, J., Lucas, H., 2009. Screening of sustainable groundwater sources for integration into a regional drought-prone water supply system. Hydrology and Earth System Sciences 13, 1185–1199. Stigter, T., 2011. Assessing and Managing the Impact of Climate Change on Coastal Groundwater Resources and Dependent Ecosystems: Final Report. Instituto Superior Tecnico, Lisbon. Stoch, F., Galassi, D., 2010. Stygobiotic crustacean species richness: a question of numbers, a matter of scale. Hydrobiologia 653, 217–234.

71
Teixeira, H., Salas, F., Borja, A., Neto, J., Marques, J., 2008. A benthic perspective in assessing the ecological status of estuaries: the case of the mondego estuary (portugal). Ecological Indicators 8, 404–416. Tomassoni, G., 2000. A federal statutory/regulatory/policy perspective on remedial decision-making with respect to ground-water/surface-water interaction, in: Surface-Water Interactions Workshop, p. 13. Tynen, M., 1969. Littoral distribution of lumbricillus reynoldsoni backlund and other enchytraeidae (oligochaeta) in relation to salinity and other factors. Oikos , 41–53. Ver´ıssimo, H., Neto, J., Teixeira, H., Franco, J., Fath, B., Marques, J., Patr´ıcio, J., 2012. Ability of benthic indicators to assess ecological quality in estuaries following management. Ecological Indicators . Vervier, P., Gibert, J., Marmonier, P., Dole-Olivier, M., 1992. A perspective on the permeability of the surface freshwater-groundwater ecotone. Journal of the North American Benthological Society , 93–102. Warren, L., 1977. The ecology of capitella capitata in british waters. Journal of the Marine Biological Association of the United Kingdom 57, 151–159. Wetzel, M., von der Ohe, P., Manz, W., Koop, J., Wahrendorf, D., 2012. The ecological quality status of the elbe estuary. A comparative approach on different benthic biotic indices applied to a highly modified estuary. Ecological Indicators , 118–129. Williams, D., 1984. The hyporheic zone as a habitat for aquatic insects and associated arthropods. The ecology of aquatic insects , 403–455 Winter, T., 2000. Interaction of ground water and surface water, in: Surface-Water Interactions Workshop, p. 15. Winter, T., 2003. Hydrological, chemical, and biological characteristics of a prairie pothole wetland complex under highly variable climate conditions: the Cottonwood Lake area, east-central North Dakota. 1675, USGPO. Wrona, F., Prowse, T., Reist, J., Hobbie, J., L´evesque, L., Vincent, W., 2006. Climate change effects on aquatic biota, ecosystem structure and function. AMBIO: A Journal of the Human Environment 35, 359–369. York, J., Person, M., Gutowski, W., Winter, T., 2002. Putting aquifers into atmospheric simulation models: An example from the mill creek watershed, northeastern kansas. Advances in Water Resources 25, 221–238.

72
Appendix
Appendix1: Environmental variables measurements in Estômbar channel in 2009-2010 (26/10/2009).
Locations temperature EC(µs/cm)
low tide
PH DO% DO(mg/l)
OM%
C 19.6 5363 7.38 0.069 0.63 2.9
B 19.6 5420 7.4 0.017 0.16 18.9
A 20 6300 7.5 0.299 2.7 8.8
D 21 6743 7.6 0.077 0.69 8.3
E 22 9300 7.95 0.173 1.58 5.5
Appendix 2: List of identified taxa and their corresponding abundance in Estômbar estuary.
DRY WET
A B C D E A B C D E
Phylum: Cnidaria
Class: Anthozoa
Subclass: Hexacorallia
Order: Actiniaria
1 12
12
Phylum: Platyhelminthes 2
Class: Turbellaria
Order: Tricladida
1
14
Kingdom: Animalia
Superphylm Lophotrochozoa
Phylum: Nemertea
2
Phylum: Mollusca
Class: Gastropoda
Superfamily: Haminoeoidea
Family: Haminoeidae 1 1 11
Species: H. ulvae 133 10 2

73
Phylum: Mollusca
Class: Bivalvia
Subclass: Heterodonta
Order: Veneroida
Superfamily: Tellinoidea
Family: Semelidae
Genus: Scrobicularia
Species: S. plana 21 11 25 33 6 9 5 27 8 2
Genus: Abra
Species: Abra tenuis 5 3 3 5 3
Superfamily: Cardioidea
Family: Cardiidae
Genus: Cerastoderma 1
Superfamily: Veneroidea
Family: Veneridae 1
Superfamily: Cardioidea 1 2 9
Family: Cardiidae
Subfamily: Lymnocardiinae
Genus: Cerastoderma
Species: C. glaucum 4
Phylum: Arthropoda
Subphylum: Crustacea
Class: Malacostraca
Subclass: Eumalacostraca
Order: Mysida
Family: Mysidae
Genus: Gastrosaccus
Species: G.sanctus 1 4
Genus: Mesopodopsis
Species: M. slabberi 1 1
Phylum: Arthropoda 2 1
subphylum: Crustacea
Class: Malacostraca
Subclass: Eumalacostraca
Order: Isopoda
Suborder: Cymothoida
Superfamily: Anthuroidea
Family: Anthuridae
Genus: Cyathura
Species: C. carinata 46 97 45 105 239 20 24 62 20 91
Family: sphaeromatidae
Genus: Lekanesphaera
Species: L. hookeri 1 5 2 1

74
Genus: Lekanesphaera
Species: L. rugicauda 1
Species: L.hoestlandti 1
Phylum: Arthropoda
Subphylum: Crustacea
Class: Malacostraca
Subclass: Eumalacostraca
Order: Amphipoda
suborder: Gammaridea
Family: Melitidae
Genus: Melita
Species: Melita palmata 3 47 9 1 2 19 13 14
Suborder: Corophiidea
Family: Corophiidae
Genus: Corophium 1 2 2
Family: Corophiidae 7
Phylum: Arthropoda
Subphylum: Crustacea
Class: Malacostraca
Order: Decapoda
Infraorder: Brachyura
Family: Portunidae
Genus: Carcinus
Species: C. maenas 1 1
Family: Palaemonidae
Genus: Palaemon
Species: P. adspersus 1
Family: Polybiidae
Genus: Liocarcinus
Species: L. arcuatus 1 3 6
Family: Palaemonidae
Genus: Palemon
Species: P. elegans 1

75
Phylum: Annelida
Class: Polychaeta
Family: Spionidae 97 161 97 2 20 53 160 151 108 53
Family: Capitellidae
Genus: Capitella
Species: C. capitata 19 82 18 1 11 6 17 12 3 4
Family: Nereididae
Genus: Hediste
Species: H. diversicolor 25 62 33 28 17 41 12 107 3 87
Family: Ampharetidae
Genus: Alkmaria
Species: Alkmaria romijni 285 164 184 26 176 167 192 51 359 324
Family: Capitellidae
Genus: Heteromastus
Species: H.filiformis 1 8 119 5 2
Order: Aciculata
Suborder: Phyllodocida
Family: Hesionidae 1
Family: Phyllodocidae
Genus: Eteone
Species: Eteone picta 4
Family: Spionidae
Genus: Polydorella sp. 1
Order: Terebellida
Family: Cirratulidae 1 3
Family: Spionidae
Genus: Streblospio
Species: S.shrubsoli 6 4
Phylum: Annelida
Class: Clitellata
Subclass: Oligochaeta 244 621 354 4 63 55 134 194 14 63
Family: Naididae 3

76
Appendix 3. Simper analysis results: Average Similarity (within groups) and Dissimillarity(between groups).
Examines Average similarity within Dry period; Average similarity: 62,79
Species Av.Abund Av.Sim Sim/SD Contrib% Cum.%
Cyathura carinata 1,92 11,51 1,87 18,33 18,33
Alkmaria romijni 2,01 10,69 1,76 17,02 35,35 Oligochaeta 1,90 9,26 1,28 14,74 50,09 Spionidae 1,50 7,79 1,62 12,41 62,50 Scrobicularia plana 1,11 5,26 1,08 8,38 70,89 Hediste diversicolor 1,24 5,14 1,07 8,18 79,06 Capitella capitata 0,97 3,63 0,85 5,78 84,84 Hydrobia ulva 0,38 2,55 0,36 4,05 88,89
Examines Average similarity within Wet period; Average similarity: 58,51
Species Av.Abund Av.Sim Sim/SD Contrib% Cum.%
Alkmaria romijni 2,26 16,50 1,81 28,20 28,20
Cyathura carinata 1,55 11,28 2,00 19,28 47,48 Hediste diversicolor 1,35 8,34 1,15 14,26 61,74 Spionidae 1,63 7,88 0,99 13,47 75,20 Oligochaeta 1,19 4,62 0,67 7,90 83,10 Scrobicularia plana 0,78 3,89 0,71 6,64 89,74 Capitella capitata 0,70 1,99 0,52 3,40 93,14 Examines Average dissimilarity between location A and B; Averagedissimilarity: 36,46
Group A Group B
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Oligochaeta 2,00 2,10 5,27 1,12 14,45 14,45 Spionidae 1,58 1,84 4,85 1,08 13,31 27,77 Melita palmata 0,34 1,11 4,05 1,05 11,11 38,88 Hediste diversicolor 1,53 1,20 3,71 0,98 10,19 49,07 Capitella capitata 0,80 1,27 3,22 1,14 8,84 57,91 Alkmaria romijni 2,41 2,23 3,13 1,38 8,58 66,49 Cyathura carinata 1,54 1,48 2,99 0,76 8,19 74,68 Scrobicularia plana 0,97 0,85 2,77 0,84 7,61 82,29 Abra tenuis 0,34 0,33 1,88 0,93 5,17 87,46 Haminoeidae 0,20 0,10 0,93 0,55 2,55 90,01

77
Examines Average dissimilarity between location A and C; Average dissimilarity: 34, 94
Group A Group C
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Oligochaeta 2,00 2,28 4,23 1,11 12,10 12,10 Spionidae 1,58 1,85 3,98 1,00 11,39 23,50 Alkmaria romijni 2,41 1,83 3,80 1,21 10,87 34,37 Hediste diversicolor 1,53 1,43 3,56 1,39 10,19 44,56 Capitella capitata 0,80 0,86 3,25 1,14 9,29 53,85 Scrobicularia plana 0,97 1,38 3,14 0,94 8,98 62,83
Examines Average dissimilarity between location B and C; Average dissimilarity: 43,67
Group B Group C
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Oligochaeta 2,10 2,28 5,62 1,15 12,87 12,87 Hediste diversicolor 1,20 1,43 4,98 1,16 11,39 24,27 Spionidae 1,84 1,85 4,50 1,18 10,30 34,57 Melita palmata 1,11 0,30 4,24 1,36 9,72 44,29 Capitella capitata 1,27 0,86 3,89 1,23 8,91 53,20 Scrobicularia plana 0,85 1,38 3,84 1,00 8,78 61,98 Alkmaria romijni 2,23 1,83 3,76 1,15 8,60 70,58 Cyathura carinata 1,48 1,63 3,49 0,88 7,99 78,58
Examines Average dissimilarity between location A and D; Average dissimilarity: 53, 78
Group A Group D
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Alkmaria romijni 2,41 1,52 7,32 1,48 13,61 13,61 Oligochaeta 2,00 0,56 7,21 1,80 13,41 27,02 Spionidae 1,58 1,03 6,86 1,46 12,75 39,77 Hydrobia ulva 0,00 1,12 5,18 0,92 9,63 49,40 Hediste diversicolor 1,53 0,70 5,05 1,37 9,40 58,79 Scrobicularia plana 0,97 0,89 3,66 1,13 6,80 65,60 Capitella capitata 0,80 0,40 3,65 1,35 6,79 72,39 Examines Average dissimilarity between location A and E; Average dissimilarity: 43, 69
Group A Group E
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cu%
Oligochaeta 2,00 0,81 5,47 1,70 12,52 12,52 Heteromastus filiformis 0,10 1,38 4,25 1,64 9,73 22,25 Spionidae 1,58 1,51 3,55 1,07 8,13 30,38 Cyathura carinata 1,54 2,39 3,04 3,39 6,96 37,34 Capitella capitata 0,80 0,85 2,85 1,08 6,53 43,86 Melita palmata 0,34 0,62 2,68 1,05 6,13 50,00 Alkmaria romijni 2,41 2,69 2,52 1,16 5,76 55,75 Scrobicularia plana 0,97 0,63 2,42 1,17 5,53 61,28 Cardioidea 0,00 0,51 1,90 0,76 4,34 65,63 Liocarcinus arcuatus 0,00 0,45 1,87 0,75 4,28 69,90

78
Examines Average dissimilarity between location B and E; Average dissimilarity: 47, 28
Group B Group E
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Oligochaeta 2,10 0,81 5,35 1,25 11,31 11,31 Hediste diversicolor 1,20 1,60 4,07 1,23 8,61 19,92 Cyathura carinata 1,48 2,39 3,80 0,95 8,05 27,97 Spionidae 1,84 1,51 3,79 1,18 8,01 35,98 Heteromastus filiformis 0,28 1,38 3,76 1,29 7,95 43,93 Capitella capitata 1,27 0,85 3,55 1,30 7,51 51,44 Melita palmata 1,11 0,62 3,41 1,40 7,21 58,65 Examines Average dissimilarity between location C and E; Average dissimilarity: 46, 03
Group C Group E
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Oligochaeta 2,28 0,81 5,95 1,60 12,94 12,94 Heteromastus filiformis 0,00 1,38 4,68 1,55 10,17 23,10 Alkmaria romijni 1,83 2,69 3,76 1,12 8,18 31,28 Scrobicularia plana 1,38 0,63 3,26 1,58 7,09 38,37 Spionidae 1,85 1,51 2,97 1,05 6,45 44,82 Cyathura carinata 1,63 2,39 2,91 0,94 6,32 51,14 Capitella capitata 0,86 0,85 2,87 1,17 6,24 57,38 Examines Average dissimilarity between location D and E; Average dissimilarity: 56, 03
Group D Group E
Species Av.Abund Av.Abund Av.Diss Diss/SD Contrib% Cum.%
Alkmaria romijni 1,52 2,69 5,46 1,15 9,65 9,65 Heteromastus filiformis 0,15 1,38 5,19 1,69 9,18 18,83 Hediste diversicolor 0,70 1,60 5,06 1,55 8,95 27,78 Hydrobia ulva 1,12 0,12 4,66 1,05 8,24 36,02 Spionidae 1,03 1,51 4,32 1,37 7,63 43,65 Oligochaeta 0,56 0,81 3,82 1,16 6,76 50,41 Melita palmata 0,27 0,62 3,21 1,19 5,68 56,09 Cyathura carinata 1,65 2,39 3,11 1,10 5,50 61,59 Capitella capitata 0,40 0,85 3,02 1,20 5,33 66,92
79
Appendix 4: List of identified taxa and their corresponding abundance in 6 wells and spring.
Ecology Q1 Q2 Q3 Q4 Q5 Q6 Spring
Phylum: Arthropoda
Subphylum: Crustacea
Class: Maxillopoda
Subclass: Copepoda
Order: Cyclopoida
Family: Cyclopidae
Genus: Halicyclops sp. Brackish water 1
Genus: Eucyclops
species Eucyclops graeteri
hypogean (stygobite)
17 206 1
Genus: Eucyclops
Species: Eucyclops serrulatus
epigean (stygoxene, stygophile)
2
Genus: Eucyclops
species Eucyclops hadjebensis
hypogean / epigean
11
Genus: Megacyclops
species Megacyclops brachypus
hypogean (stygobite)
2
Genus: Megacyclops epigean
(stygoxene, stygophile)
41
Genus: Megacyclops
species Megacyclops
viridis
epigean (stygoxene, stygophile)
2
Genus: Megacyclops
species Macrocyclops
albidus
epigean (stygoxene, stygophile)
10
Genus: Acanthocyclops
sp.
epigean (stygoxene, stygophile)
1
Genus: Acanthocyclops
species Acanthocyclops
sensitivus hypogean (stygobite)
1
Genus: Eucyclops
species Eucyclops speratus
epigean (stygoxene, stygophile)
24 3
Order: Harpacticoida 2 3

80
Phylum: Arthropoda
Subphylum: Crustacea
Class: Ostracoda
Subclass: Podocopa
Order: Podocopia
Suborder Cypridocopina
Superfamily Cypridoidae
Family Cypridae
Genus Cypria
Species Cypria
ophtalmica
epigean (stygoxene, stygophile)
2 30 171 1
Genus Cypria
Species Cypridopsis
vidua
epigean (stygoxene, stygophile)
1 1
Genus Bradleycypris
Species Bradleycypris
obliqua
epigean (stygoxene, stygophile)
6
Phylum: Arthropoda
Subphylum: Crustacea
Class: Malacostraca
Order: Isopoda
Family: Sphaeromatidae
Genus: Sphaeroma. Sp. 1
Family: Gammaridae
Genus: Gammarus
Species: G. pulex freshwater 1
Genus: Gammarus fresh/brackish
water 2 9 3





![Universiti Malaysia Sarawak · Brunei Darussalam ..... Martinah Bte Hj Tamil, Noral ]amalludin Bin Hj Mohd Yus~ 5. Current Issues on Biodiversi Lay Khim and N eth Barom 6. Sustainable](https://img.dokumen.tips/doc/110x75/5fa5bb1a40aeac6ada2b753c/universiti-malaysia-sarawak-brunei-darussalam-martinah-bte-hj-tamil-noral.jpg)























