Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Oct. 2010, p. 10844–10851 Vol. 84, No. 200022-538X/10/$12.00 doi:10.1128/JVI.01045-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
A gC1qR Prevents White Spot Syndrome Virus Replication in theFreshwater Crayfish Pacifastacus leniusculus�
Apiruck Watthanasurorot,1 Pikul Jiravanichpaisal,1,2 Irene Soderhall,1 and Kenneth Soderhall1*Department of Comparative Physiology, Uppsala University, Uppsala, Sweden, and National Center for Genetic Engineering and
Biotechnology, National Science and Technology Development Agency, Pathumthani, Thailand
Received 14 May 2010/Accepted 15 July 2010
The gC1qR/p32 protein is a multiple receptor for several proteins and pathogens. We cloned a gC1qR homologuein a crustacean, Pacifastacus leniusculus, and analyzed the expression of P. leniusculus C1qR (PlgC1qR) in varioustissues. The gC1qR/p32 transcript was significantly enhanced by white spot syndrome virus (WSSV) infection 6 hafter viral infection both in vitro in a hematopoietic tissue cell culture (Hpt) and in vivo compared to appropriatecontrols. Moreover, PlgC1qR silencing in both the Hpt cell culture and live crayfish enhanced the WSSV replication.In addition, by making a recombinant PlgC1qR protein we could show that if this recombinant protein was injectedin a crayfish, Pacifastacus leniusculus, followed by injection of WSSV, this significantly reduced viral replication invivo. Furthermore, if the recombinant PlgC1qR was incubated with Hpt cells and then WSSV was added, this alsoreduced viral replication. These experiments clearly demonstrate that recombinant PlgC1qR reduce WSSV repli-cation both in vivo and in vitro. The results from a far-Western overlay and glutathione S-transferase pull-downassays showed that PlgC1qR could bind to VP15, VP26, and VP28. Altogether, these results demonstrate a role forPlgC1qR in antiviral activity against WSSV.
White spot syndrome virus (WSSV), an enveloped double-stranded DNA virus, is a serious pathogen to shrimp and manyother crustaceans, including freshwater crayfish. Several genesand proteins that are affected by a WSSV infection have beenreported (18). Some of these appear to play a critical role inanti-WSSV activity since they are associated with an ability ofinfected crustaceans to survive viral infection (5, 17, 20, 30).Moreover, some proteins of shrimp, such as a chitin-bindingprotein and �-integrin, have been proposed as cellular recep-tors for WSSV (4, 14, 30). However, the mechanism by whichthey function as receptors is not known.
The first gC1qR, also known as p32, C1qBP, and HABP1,was identified as the globular head of the C1q binding protein(3). This protein is detected in various tissues and many com-partments of a cell, including the cell surface (9, 11). ThegC1qR on the cell surface can serve as a receptor for numerousextra- and intracellular proteins and microbial and viral pro-teins, and it was originally called a multifunctional chaperone(16, 23, 44). In general, the binding of complement proteinC1q on the receptor gC1qR induces early defense responsesagainst viral infections (11, 45). After binding of a ligand,gC1qR can induce the generation of proinflammatory by-prod-ucts from the complement and kinin/kallikrein pathways, but itis also able to serve as a vehicle for many pathogens to enterinto the host cells. Recently, for example, it has been shownthat the core proteins of hepatitis B virus, hepatitis C virus, andadenovirus can bind to gC1qR, and there it plays an importantrole in promoting viral infection and maintaining virus persis-tence (22, 45).
In the present study, a full-length cDNA of gC1qR wascloned, and its tissue distribution in crayfish was studied. ThisgC1qR of Pacifastacus leniusculus (PlgC1qR) was also found tobe upregulated upon WSSV infection. Both in vivo and in thehematopoietic tissue (Hpt) cell culture in vitro, semiquantita-tive PCR showed that knockdown of the PlgC1qR by RNAinterference (RNAi) resulted in WSSV replication levels thatwere higher than those treated with control double-strandedRNA (dsRNA). In addition, we showed that a recombinantPlgC1qR protein decreased WSSV VP28 expression both invitro and in vivo. Moreover, by using a far-Western overlay andglutathione S-transferase (GST) pull-down assays, we showthat the PlgC1qR could bind to VP15, VP26, and VP28. It istherefore likely that PlgC1qR plays an essential role in theimmune defense against WSSV replication in crayfish.
MATERIALS AND METHODS
Crayfish. Healthy intermolt crayfish, P. leniusculus, from Nils Fors, Torsang atLake Vattern, Sweden, were maintained in aerated tap water at 10°C.
Tissue distribution of PlgC1qR mRNA. RNA from various tissues, includinghepatopancreas, stomach, intestine, heart, Hpt, muscle, brain, hemocytes, andnerve, was extracted according to the instructions of the GenElute mammaliantotal RNA miniprep kit (Sigma), followed by treatment with RNase-free DNaseI (Ambion, Austin, TX). cDNA was synthesized by using ThermoScript (In-vitrogen). Gene-specific forward and reverse primers for PlgC1qR and 40Sribosomal genes (PlgC1qR-F [5�-AATCACACGGTAGACACTGAAATGCC-3�] and PlgC1qR-R [5�-CATCATCCCATCTAAAATGTCCCCTG-3�]; 40S-F[5�-CCAGGACCCCCAAACTTCTTAG-3�] and 40S-R [5�-GAAAACTGCCACAGCCGTTG-3�]) were designed from P. leniusculus Lambda Zap Expresslibrary Hpt cDNA (GenBank accession numbers GR930855 and CF542417,respectively) and used in reverse transcription-PCR (RT-PCR). The 40S ribo-somal gene was used as an internal control in all PCR experiments. PCR con-ditions were as follows: 94°C for 2 min, followed by 30 cycles of 94°C for 20 s,58°C for 20 s, and 72°C for 30 s for the PlgC1qR gene and 25 cycles for the 40Sgene. The PCR products were analyzed on 1.2% agarose gel stained withethidium bromide.
Cloning of full-length PlgC1qR cDNA. Total RNA (at least 1 �g) was extractedfrom heart and converted into 5� and 3� RACE-Ready first-stand cDNA accord-ing to the SMARTer RACE cDNA amplification kit user manual (Clontech).
* Corresponding author. Mailing address: Department of Compar-ative Physiology, Uppsala University, Norbyvagen 18A, SE-75236Uppsala, Sweden. Phone: 46 18 471 2818. Fax: 46 18 471 6425. E-mail:[email protected].
� Published ahead of print on 4 August 2010.
10844
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Then, 5� RACE (5� rapid amplification of cDNA ends) PCR was performed byusing the gene specific primer of PlgC1qR-R (above experiment) and theSMART universal primer A mix. The 3� RACE PCR was performed using 3�RACE-Ready first-stand cDNA template, with the PlgC1qR-F and SMARTuniversal primer A mix. Both RACE PCR thermal cycling protocols were asfollows: 25 cycles of 94°C for 30 s, 68°C for 30 s, and 72°C for 3 min. The 5� and3� RACE PCR products were cloned into TOP10 vector (Invitrogen) and se-quenced.
PlgC1qR sequence analysis, domain search, and phylogenetic analysis. Thenucleotide sequence of PlgC1qR was compared to other gC1qRs in GenBankby using BlastX. Multiple sequence alignment was performed by usingCLUSTALW (http://www.ebi.ac.uk/tools/clustalw/index.html). The deducedamino acid domain and mitochondrial targeting sequences were predictedwith the SMART (http://smart.embl-heidelberg.de/) and MITOPROT pro-grams (http://ihg2.helmholtz-muenchen.de/ihg/mitoprot.html), respectively.A phylogenetic cladogram representing the relationship between PlgC1qRand other gC1qR proteins was constructed by PHYLIP version 3.69 withbootstrap resampling, the neighbor-joining algorithm (7), and illustrated byusing TreeView (http://taxonomy.zoology.gla.ac.uk/rod/treeview.html).
Crayfish Hpt cell culture and maintenance. The Hpt was dissected accordingto the method of Soderhall et al. (28). The Hpt was washed with CPBS (crayfishphosphate-buffered saline: 10 mM Na2HPO4, 10 mM KH2PO4, 150 mM NaCl,10 �M CaCl2, 10 �M MnCl2 [pH 6.8]) and incubated in 600 �l of 0.1% colla-genase (types I and IV; Sigma) in CPBS at room temperature for 45 min toseparate the Hpt cells. The separated cells were washed twice with CPBS byspinning down at 800 � g for 5 min at room temperature. The cell pellet wasresuspended in modified L-15 medium (29) and subsequently seeded at a densityof 2.5 � 106 cells/150 �l in 96-well plates. Hpt cells were supplemented withpartially purified plasma (29) after 1 h of attachment at room temperature, theculture plates were incubated at 16°C, and one-third of the medium was changedat 48-h intervals.
PlgC1qR transcription in response to WSSV infection in vitro and in vivo. AWSSV stock was prepared from plasma of viral infected crayfish and controlWSSV was UV treated and killed according to the method of Jiravanichpaisalet al. (13). In the in vitro experiments, Hpt cell cultures were prepared andincubated at 16°C for 12 h. The medium was then replaced with 150 �l of L-15medium containing 5 �l of the WSSV stock suspension (the control group wassupplemented with 5 �l of UV-killed WSSV [equivalent to 2.5 � 104 copies]) and5 �l of crude astakine, followed by incubation for 0, 6, 12, and 24 h at 20°C.Thereafter, the cells were harvested at each time point for extraction of totalRNA. In vivo, normal and UV-inactivated WSSV preparations were dilutedthree times in crayfish saline buffer (CFS; 0.2 M NaCl, 5.4 mM KCl, 10 �MCaCl2, 10 mM MgCl2, and 2 mM NaHCO3 [pH 6.8]). In the in vivo experiments,200 �l of WSSV or control (equivalent to 2 � 106 copies) was injected via thebase of the fourth walking leg. The hemolymph of three crayfish from each groupwere bled at 0, 6, 12, and 24 h postinjection, and the hemocytes were separatelypreserved for RNA extraction. The transcription level of PlgC1qR in vitro and invivo was detected with semiquantitative RT-PCR. The amplification programswere the same as in the tissue distribution experiment, and the PCR productswere detected on a 1.2% agarose gel. The intensity of the PlgC1qR band fromeach group and time was measured by using QuantityOne (Bio-Rad).
Generation of dsRNA. Gene-specific primers for PlgC1qR and green fluores-cent protein (GFP) were incorporated with T7 promoter (italic letters) at the 5�ends (gC1Q 100� [5�-TAATACGACTCACTATAGGGGTCCTCTCCTCCAACAACCG-3�] and gC1Q 621� [5�-TAATACGACTCACTATAGGGCCCATCTAAAATGTCCCCTGA-3�]; GFP 63� [TAATACGACTCACTATAGGGCGACGTAAACGGCCACAAGT] and GFP 719� [TAATACGACTCACTATAGGGTTCTTGTACAGCTCGTCCATG]) and used to amplify PCR products as a templatefor dsRNA synthesis. A GFP transcript was amplified with the pd2EGFP-1vector (Clontech) as a template and used as a control. The amplified productswere then purified by using a GenElute gel extraction kit (Sigma), followed by invitro transcription using a MegaScript kit (Ambion). The dsRNA was purifiedwith the TRIzol LS reagent (Invitrogen).
dsRNAi in vitro and WSSV infection. The Hpt cells were divided into threegroups with four replicates in each group. The Hpt cells received differenttreatments as follows: group 1, GFP dsRNA plus UV-killed WSSV; group 2,GFP dsRNA plus WSSV; and group 3, PlgC1qR dsRNA plus WSSV. ThedsRNA transfection and WSSV infection into Hpt cell cultures was performed asdescribed by Liu et al. (17). Briefly, 4 �l of dsRNA (250 ng/�l) was mixed with3 �l of histone H2A (1 mg/ml) and with 20 �l of modified L15 and then addedto one well of 1-day-old Hpt cell cultures. The cells were then incubated for 3days at 16°C. After incubation, one replicate of groups 2 and 3 were subjected toRNA extraction to determine the RNAi efficiencies. For the other cells, the
medium was replaced with 150 �l of L15 medium, together with 5 �l of WSSVstock suspension (13), and 5 �l of crude astakine preparation and incubated foranother 36 h at 20°C, followed by RNA preparation.
dsRNAi in vivo. Small intermolt crayfishes (15 � 2 g of fresh weight) weredivided into three groups, with three crayfish in each group (n � 3). The first andsecond groups were injected with 300 �g of GFP control dsRNA, and the thirdgroup was injected with 300 �g of dsPlgC1qR, via the base of the fourth walkingleg. After 24 h of the first dsRNA injection, four drops of crayfish hemolymphwere bled for total RNA isolation to test the efficiency of the RNAi. Then, 200-�ldoses of UV inactivated WSSV were injected into the first group, while the othertwo groups were injected with normal live WSSV. At 12 h after WSSV injection,dsRNA was injected a second time into all three groups as described above. After36 h from the first WSSV infection, the total hemocyte RNA was extracted todetermine PlgC1qR and WSSV VP28 transcripts by semiquantitative RT-PCR.
RNAi efficiency. In order to estimate RNAi efficiency, PCR was performedwith three oligonucleotide primers (PlgC1qR ORF-F [5�-CCTGCGTGTGTCTCAAGCCC-3�] and PlgC1qR ORF-R [5�-TGCCTCCGTTACTTCCGCTT-3�];WSSV VP28 [GenBank accession no. AF502435]-F [5�-TCACTCTTTCGGTCGTGTCG-3�] and WSSV VP28-R [5�-CCACACACAAAGGTGCCAAC-3�];and the previous primer for 40S ribosomal gene). After RNAi and virus infectionin vitro and in vivo, the PCR conditions were as follows: 94°C for 2 min, followedby 28 cycles of 94°C for 20 s, 58°C for 20 s, and 72°C for 30 s for PlgC1qR andVP28 and by 25 cycles for the 40S ribosomal gene.
Recombinant PlgC1qR protein. The coding sequence of PlgC1qR without 59amino acids at the N terminus was amplified using mature_gC1qR_expBamHI-Forward (5�-TTTTGGATCCATGCACACCAGAGGTGATCGTG-3�) and mature_gC1qR_exp XhoI-Reverse (5�-CTCGAGTTACTTCCGCTTCACAAAGTCCT-3�) primers. This insert was cloned into pGEX-4T-1(GE healthcare) at BamHI and XhoI and transformed into BL21 E. coli. Single colonieswere grown in LB medium containing 100 �g of ampicillin/ml to an opticaldensity at 600 nm of 0.6 and induced with 1 �M IPTG (isopropyl-�-D-thioga-lactopyranoside) for 5 h at 37°C. The protein was expressed as a fusion productwith GST partly at the N terminus of rPlgC1qR. After purifying this GST fusionprotein on a GST-trap FF column (GE Healthcare), the presence of recombi-nant protein was confirmed by Western blotting. The protein samples weresubjected to SDS–12% PAGE and then transferred electrophoretically to poly-vinylidene difluoride (PDVF) membrane. The membrane was blocked by im-mersion in 10% skimmed milk in TBST for 1 h and washed three times in 1�TBST (10 mM Tris-HCl [pH 7.5] containing 150 mM NaCl and 0.1% Tween 20).The membrane was then incubated with a 1: 2,000 dilution of a primary antibodyfor GST (Sigma) in TBST for 1 h. The previous washing procedure was thenrepeated before the membrane was incubated with anti-mouse IgG peroxidase-linked species-specific whole antibody from sheep (GE Healthcare) at 1:3,000 in1� TBST for 1 h and washed with TBST for 3 � 10 min. For detection, an ECLWestern blotting reagent kit (Amersham Biosciences) was used according to themanufacturer’s instructions.
The fusion protein with GST tag was used in a GST pull-down assay andfar-Western overlay assay as described below. For antiviral assay, the GST tagwas removed on a column by incubating with thrombin (10 U of thrombin foreach mg of fusion protein). The recombinant protein without GST tag wascleared of thrombin using HiTrap Benzamidine FF (high sub) (GE Healthcare).
WSSV purification. WSSV purification was modified from the method de-scribed by Xie et al. (40). Briefly, 5 g of gills from WSSV-infected crayfish wascollected and homogenized in 36 ml of TNE buffer (50 mM Tris-HCl, 400 mMNaCl, 5 mM EDTA [pH 8.5]) containing a complete protease inhibitor (usingcomplete, mini, EDTA-free; Roche). After centrifugation at 3,500 � g for 5min at 4°C, the preparation was filtered by a nylon net (400 mesh). Thesupernatant was centrifuged at 30,000 � g for 30 min at 4°C; the supernatantwas then carefully discarded, and the lower white pellet was suspended in 1ml of PBS buffer (140 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mMKH2PO4 [pH 7.3]).
GST pull-down assay. WSSV envelope proteins were solubilized by incubationwith Triton X-100 (0.4% Triton X-100 for 50 �l of purified WSSV) for 1 h atroom temperature with gentle shaking. The envelope fraction was collected bycentrifugation at 30,000 � g for 20 min at 4°C and suspended in PBS buffer. Theinteraction of WSSV envelope protein and PlgC1qR was examined by incubating5 �g of purified GST-PlgC1qR, 5 �g of WSSV envelope fraction, and glutathi-one-Sepharose 4B resin (50 �l of 50/50 bed slurry) for 2 h at 4°C. In the controlreaction, GST was displacing GST-PlgC1qR. After incubation, the samples werewashed 10 times with PBS, and then fusion proteins were eluted by adding PBScontaining 10 mM reduced glutathione. The fusion proteins were detected inSDS–12% PAGE and stained with Coomassie blue. The presence of GST andGST-PlgC1qR was confirmed by Western blot analysis. The protein bands that
VOL. 84, 2010 PlgC1qR PREVENTS WSSV REPLICATION 10845
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

were not found in the control were cut and used for mass spectrophotometeranalysis.
Far-Western overlay assay. To identify proteins in WSSV that could bind toPlgC1r, a far-Western overlay assay was used. These samples were subjected toSDS–12% PAGE, transferred to PDVF membranes, and blocked with 10% skim
milk in TBST for 1 h at room temperature. After three washes with TBST, themembranes were incubated with 25 nM GST-PlgC1r in 10 ml of TBST for 1 h atroom temperature. The blots were washed twice and incubated for 1 h with1:1,000 dilution of anti-GST-antibody and then washed twice, incubated for 1 hwith 1: 2,000 dilutions of anti-mouse antibody for 1 h, and washed three timesbefore detection with an ECL Western blotting reagent kit. The control reactionwas incubated with GST instead of GST-PlgC1r.
Mass spectrophotometer analysis. Selected bands from different gels stainedwith Coomassie blue were excised and cleaved with trypsin by in-gel digestion.The peptides were analyzed by ESI-MS on a Q-TOF mass spectrometer (Waters,United Kingdom) using MassLynx software. Sequence homology search wasperformed with the BLAST program and the MS-BLAST program.
Antiviral activity of recombinant PlgC1qR. The experimental setup for both invitro and in vivo experiments were four groups with three replicates in eachgroup. These experimental groups received different treatments as follows: group1, GFP dsRNA with WSSV; group 2, GFP dsRNA with WSSV and rPlgC1qR;group 3, dsRNA and PlgC1qR with WSSV and rPlgC1qR; and group 4, PlgC1qRdsRNA with WSSV. The concentration of dsRNA and the dose and time ofaddition of WSSV in both in vitro and in vivo were as described above. Portions(2 and 10 �g) of rPlgC1qR were incubated with Hpt cell culture and injected intocrayfish. The recombinant protein was injected or incubated together withWSSV, followed by RNAi. The transcription of PlgC1qR, WSSV, and 40S ribo-somal was then detected by using semiquantitative RT-PCR.
Statistical analysis. The relative expression levels of different time groupswere examined by one-way analysis of variance, followed by Duncan’s newmultiple-range test and the Tukey test. Differences were considered statisticallysignificant at P 0.05. The results are expressed as the mean � the standarderror.
RESULTS
A full-length cDNA of PlgC1qR was obtained by RACEPCR. The open reading frame of PlgC1qR was 771 bp, encod-ing 256 amino acid residues (Fig. 1). The nucleotides fromRACE PCR also contained 244 and 440 bp of the 5� and 3�untranslated regions, respectively. A mitochondrial acidic ma-trix protein domain of 33 kDa (MAM33), which binds to theglobular “heads” of the C1q complementary protein, was pre-dicted from residues 72 to 256 of the deduced protein by theSMART program. With a prediction from the MITOPROTprogram, the deduced protein also had a mitochondrial target-ing sequences at the first 39 amino acids of its N-terminal end.In addition, the deduced PlgC1qR protein was found to havean Arg-Gly-Asp (also know as the RGD motif or cell adhesivemotif) at residues 62 to 64, and this motif plays an importantrole in binding to a family of cell surface receptors calledintegrins (2). The RGD motif is also found in the gC1qR of thesilk worm (Bombyx mori), whereas it is absent in gC1qRs ofother species.
The PlgC1qR transcript was detected in several tissues, andthe highest expression was found in the Hpt and the heart (Fig.2). The amino acid sequences of gC1qR from nine species,containing five vertebrates and four invertebrates, were ana-lyzed by using the neighbor-joining distance method. This anal-
FIG. 1. Nucleotide and amino acid sequence of P. leniusculusgC1qR. Boldface and single-underlined letters represent start and stopcodons, while double-underlined letters are the polyadenylation signal(AATAAA). The mitochondrial cleavage site and MAM domain ofthis protein are shown with boldface italics and gray shaded, respec-tively. In addition, a RGD motif is found in this protein, and its aminoacid residues are shown in a box.
FIG. 2. Tissue distribution of PlgC1qR in P. leniusculus. The experimental tissues examined included hepatopancreas (HP), stomach (ST),intestine (IN), gill (G), testis (TT), heart (HE), muscle (M), brain (B), hemocyte (HC), and abdominal nerve (N). A 40S ribosomal gene is usedas internal control.
10846 WATTHANASUROROT ET AL. J. VIROL.
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

ysis shows that this protein can be separated into one verte-brate and one invertebrate group, and the PlgC1qR was in theinvertebrate group (Fig. 3).
To investigate the response to WSSV infection in vitro andin vivo, PlgC1qR expression was detected by semiquantitativeRT-PCR. In the Hpt cell cultures the addition of WSSV orUV-inactivated WSSV resulted in a significant increase inPlgC1qR expression 12 h postinfection (Fig. 4A). If crayfishwere injected with WSSV or UV-inactivated WSSV as a con-trol, a significant difference (P 0.05) of PlgC1qR expressionin vivo between WSSV-challenged and control crayfish at 12 hpostinjection was observed (Fig. 4B).
The role of PlgC1qR during a WSSV infection was investi-gated using dsPlgC1qR RNAi to suppress the PlgC1qR expres-sion. The PlgC1qR gene was completely knocked down in theHpt cells in vitro (Fig. 5A), whereas the 40S ribosomal genewas unaffected. As shown in Fig. 5B and C, the WSSV infec-tion of Hpt cells resulted in an increased expression of thePlgC1qR. Furthermore, when the PlgC1qR was completelysilenced, the WSSV replication was dramatically increased, whichis shown by an increase in WSSV VP28 expression (Fig. 5B andC). dsRNA of gC1qR or dsRNA of GFP was also injected intocrayfish, and the efficiency of RNAi of the PlgC1qR was de-tected by RT-PCR before the crayfish received WSSV by in-jection (Fig. 6A). After crayfish were injected with WSSV, theVP28 of the WSSV in PlgC1qR-silenced animals was signifi-
cantly different and higher than the control group (Fig. 6B andC). These experiments therefore support the results from thein vitro experiments.
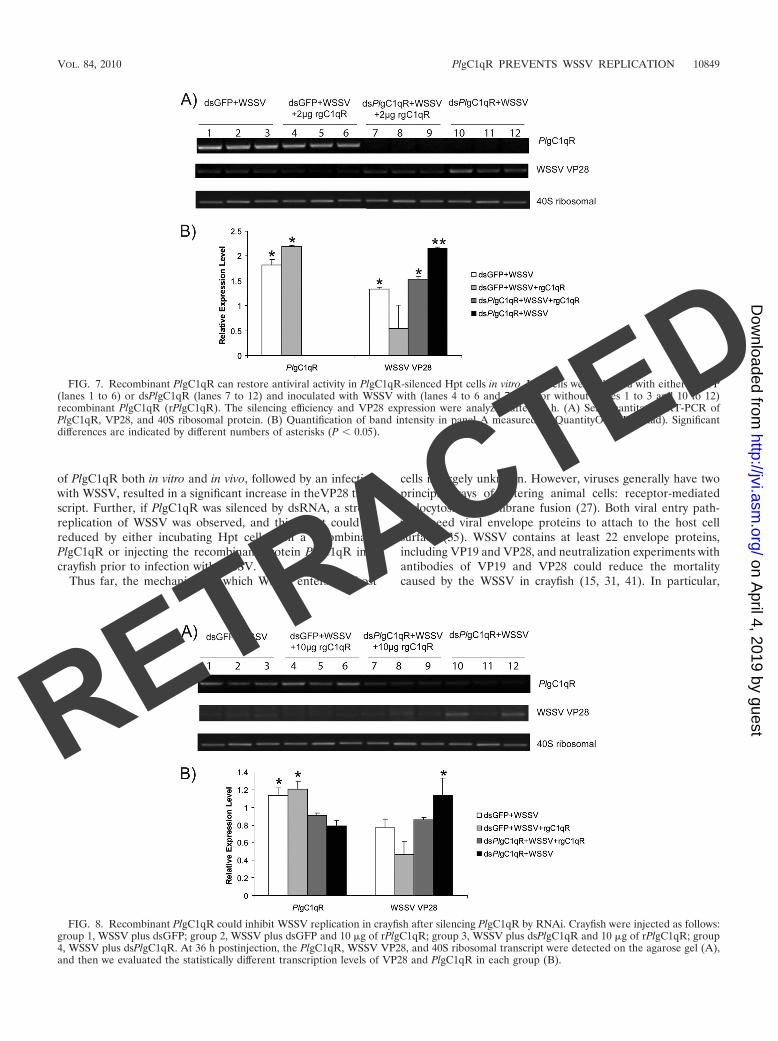
Our previous experiments showed that PlgC1qR is upregu-lated after WSSV infection and, when the PlgC1qR gene wassilenced by dsRNA, replication of WSSV was increased. Takentogether, these results indicate a role for the PlgC1qR in an-tiviral protection against WSSV. To investigate whether thePlgC1qR could interfere with the WSSV replication, we addeda recombinant protein (rPlgC1qR) to Hpt cells that had beenPlgC1qR silenced and then infected with WSSV. Figure 7A(lanes 7 to 9) shows that the addition of recombinant PlgC1qRdecreased the expression of WSSV VP28, whereas in the con-trol where PlgC1qR was knocked down but no recombinantprotein was added, WSSV replication was instead increased, aswas expected (lanes 10 to 12). In crayfish where PlgC1qR wassilenced and then they were injected with WSSV the replica-tion of this virus was increased, but if recombinant PlgC1qRwas injected this decreased WSSV replication (Fig. 8), al-though the silencing efficiency is lower compared to in vitro-cultured cells, and therefore the obtained results are morevariable.
In order to find out whether any WSSV protein could inter-act with PlgC1qR, we used a GST pull-down assay and afar-Western overlay assay. We found that the PlgC1qR couldbind to VP26 and VP28 of the WSSV, and this was demon-
FIG. 3. Evolutionary relationship of gC1qR in P. leniusculus andother species. The following protein sequences of gC1qR were used:Bombyx mori (DQ311376), Aedes aegypti (XM_001661360), Nasoniavitripennis (XM_001607453), Ixodes scapularis (XM_002400551), Bostaurus (NM_001034527), Mus musculus (NP_031599), Danio rerio(NP_001017858), Salmo salar (BT047985), and Homo sapiens(NP_001203).
FIG. 4. Expression of PlgC1qR in response to WSSV in vitro and invivo. (A) PlgC1qR expression in an Hpt cell culture and exposure toWSSV for 6 h. (B) In vivo, the expression level of PlgC1qR in hemo-cytes of WSSV-injected crayfish is higher than in hemocytes of controlcrayfish at 6 h after WSSV injection. The asterisk represents significantdifferences (P 0.05).
VOL. 84, 2010 PlgC1qR PREVENTS WSSV REPLICATION 10847
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

strated using both methods (Fig. 9). In addition, the result ofthe far-Western overlay blotting also indicated that bindingoccurred between PlgC1qR and VP15 (Fig. 9A).
DISCUSSION
The gC1qR was identified as the globular “head” bindingprotein of the C1q protein (24), and this protein contains adomain that can bind to C1q. This domain is also called themitochondrial acidic matrix protein of 33 kDa (MAM33) do-main and was named after the yeast homologue of the gC1qR,MAM33 protein, the function of which in Saccharomyces cer-evisiae remains to be defined (26). The MAM33p was identi-fied as a homologue of the human gC1qR, with 53 and 24%sequence similarity and identity, respectively (6, 21). BothgC1qR and MAM33p contain a mitochondrial targeting se-quence, which directs proteins to the mitochondrial matrix. Amitochondrial targeting sequence was predicted at the 1 to 39amino acid residues of the N-terminal of PlgC1qR using theMITO program. Interestingly, P. leniusculus, as well as B. mori,gC1qR contain an RGD motif that may serve as binding sitesor receptors for integrin (1, 25). RGD-containing proteins arefound in several virus, and they seem to play a role in promot-
ing virus entry and virus-cell interaction (37, 39). The attach-ment of virus to host cells can be inhibited by many factors, forexample, proteins or synthetic peptides containing an RGDsequence (10, 37). Recently, an RGD-containing peptide hasbeen identified as a protein inhibiting WSSV infection inshrimp (14). However, there are a number of proteins thatcontain this motif, and thus the use of synthetic RGD-peptidewill not show conclusively that this motif is involved in antiviralactivity. Upregulation of gC1qR can be induced by many fac-tors, such as inflammatory mediators, microbes, mitogenicagents, and virus (11, 12, 44). A significant PlgC1qR responseto WSSV infection in crayfish, P. leniusculus, both in vitro andin vivo was detected at 6 h after viral infection. In mammals,gC1qR is coexpressed with calreticulin (CRT; also known ascC1qR) inside and on the membrane of numerous cells, in-cluding different immune cells such as B- or T-cell lympho-cytes, macrophages, and dendritic cells (6, 33). Moreover,CRT, which has a high homology with the gC1qR (32), hasbeen found to be upregulated early as a response to WSSVinfection in shrimp (8, 19, 38). We have clearly shown here thatPlgC1qR is upregulated strongly by WSSV infection.
Recently, it was shown that gC1qR has a role in defenseagainst bacterial invasion by inhibiting hyaluronidase of S.pneumoniae (43). Moreover, gC1qR is a receptor which pro-motes entry of bacteria and virus to host cells (23, 42). RNAi
FIG. 5. PlgC1qR silencing influences WSSV replication in vitro.(A) The PlgC1qR dsRNA completely silenced endogenous PlgC1qRtranscripts in a Hpt cell culture before WSSV addition (M, molecularweight marker). (B) At 36 h after WSSV infection, WSSV VP28 andPlgC1qR were analyzed by semiquantitative RT-PCR, and the PCRproducts were visualized on 1.2% agarose gel stained with ethidiumbromide. (C) Diagram representing the band intensity of the results inpanel B measured by the QuantityOne program and statistical analysisbetween each group. Significant differences are indicated by differentnumbers of asterisks (P 0.05).
FIG. 6. PlgC1qR silencing results in enhanced viral replication invivo. (A) Silencing of PlgC1qR by dsRNA was detected by RT-PCRbefore WSSV injection. (B) WSSV VP28 expression in PlgC1qR (lastthree lanes) and control (middle three lanes) silenced animals. UV-killedWSSV was injected as a control (first three lanes) and was analyzed bysemiquantitative RT-PCR. (C) Statistical analysis comparing differentband intensities of the results shown in panel B. Significant differences areindicated by different numbers of asterisks (P 0.05).
10848 WATTHANASUROROT ET AL. J. VIROL.
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
of PlgC1qR both in vitro and in vivo, followed by an infectionwith WSSV, resulted in a significant increase in theVP28 tran-script. Further, if PlgC1qR was silenced by dsRNA, a strongreplication of WSSV was observed, and this effect could bereduced by either incubating Hpt cells with a recombinantPlgC1qR or injecting the recombinant protein PlgC1qR intocrayfish prior to infection with WSSV.
Thus far, the mechanism by which WSSV enters the host
cells is largely unknown. However, viruses generally have twoprinciple ways of entering animal cells: receptor-mediatedendocytosis or membrane fusion (27). Both viral entry path-ways need viral envelope proteins to attach to the host cellsurface (35). WSSV contains at least 22 envelope proteins,including VP19 and VP28, and neutralization experiments withantibodies of VP19 and VP28 could reduce the mortalitycaused by the WSSV in crayfish (15, 31, 41). In particular,
FIG. 7. Recombinant PlgC1qR can restore antiviral activity in PlgC1qR-silenced Hpt cells in vitro. Hpt cells were silenced with either dsGFP(lanes 1 to 6) or dsPlgC1qR (lanes 7 to 12) and inoculated with WSSV with (lanes 4 to 6 and 7 to 9) or without (lanes 1 to 3 and 10 to 12)recombinant PlgC1qR (rPlgC1qR). The silencing efficiency and VP28 expression were analyzed after 36 h. (A) Semiquantitative RT-PCR ofPlgC1qR, VP28, and 40S ribosomal protein. (B) Quantification of band intensity in panel A measured by QuantityOne (Bio-Rad). Significantdifferences are indicated by different numbers of asterisks (P 0.05).
FIG. 8. Recombinant PlgC1qR could inhibit WSSV replication in crayfish after silencing PlgC1qR by RNAi. Crayfish were injected as follows:group 1, WSSV plus dsGFP; group 2, WSSV plus dsGFP and 10 �g of rPlgC1qR; group 3, WSSV plus dsPlgC1qR and 10 �g of rPlgC1qR; group4, WSSV plus dsPlgC1qR. At 36 h postinjection, the PlgC1qR, WSSV VP28, and 40S ribosomal transcript were detected on the agarose gel (A),and then we evaluated the statistically different transcription levels of VP28 and PlgC1qR in each group (B).
VOL. 84, 2010 PlgC1qR PREVENTS WSSV REPLICATION 10849
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

VP28 plays a key role in the systemic infection of shrimp byWSSV (35). Both GST pull-down assay and far-Western over-lay blotting showed that PlgC1qR could bind to the majorenvelope protein VP28. With the ability of PlgC1qR to bindVP28 and since it is localized on the cell surface, the PlgC1qRmight interfere with the virus-cell binding process during aWSSV infection. In addition, PlgC1qR was also found to bindto two nucleocapsid proteins (VP15 and VP26) of WSSV.These proteins, VP15 and VP26, are identified as histone-likeproteins and act as tegument protein (34, 36). These proteinsprobably play important roles in WSSV replication and matu-ration and hence, again, the PlgC1qR might interfere withWSSV replication through binding both these nucleocapsidproteins.
ACKNOWLEDGMENT
This study was supported by grants from the Swedish Science Re-search Council.
REFERENCES
1. Ao, J., and X. Chen. 2006. Identification and characterization of a novel geneencoding an RGD-containing protein in large yellow croaker iridovirus.Virology 355:213–222.
2. Berinstein, A., M. Roivainen, T. Hovi, P. W. Mason, and B. Baxt. 1995.
Antibodies to the vitronectin receptor (integrin alpha V beta 3) inhibitbinding and infection of foot-and-mouth disease virus to cultured cells.J. Virol. 69:2664–2666.
3. Braun, L., B. Ghebrehiwet, and P. Cossart. 2000. gC1q-R/p32, a C1q-bindingprotein, is a receptor for the InlB invasion protein of Listeria monocytogenes.EMBO J. 19:1458–1466.
4. Chen, L. L., L. C. Lu, W. J. Wu, C. F. Lo, and W. P. Huang. 2007. White spotsyndrome virus envelope protein VP53A interacts with Penaeus monodonchitin-binding protein (PmCBP). Dis. Aquat. Organ. 74:171–178.
5. Chen, W. Y., K. C. Ho, J. H. Leu, K. F. Liu, H. C. Wang, G. H. Kou, and C. F.Lo. 2008. WSSV infection activates STAT in shrimp. Dev. Comp. Immunol.32:1142–1150.
6. Dedio, J., W. Jahnen-Dechent, M. Bachmann, and W. Muller-Esterl. 1998.The multiligand-binding protein gC1qR, putative C1q receptor, is a mito-chondrial protein. J. Immunol. 160:3534–3542.
7. DeSalle, R., C. Wray, and R. Absher. 1994. Computational problems inmolecular systematics. EXS 69:353–370.
8. Fagutao, F. F., M. Yasuike, C. M. Caipang, H. Kondo, I. Hirono, Y. Taka-hashi, and T. Aoki. 2008. Gene expression profile of hemocytes of kurumashrimp, Marsupenaeus japonicus following peptidoglycan stimulation. Mar.Biotechnol. 10:731–740.
9. Fogal, V., L. Zhang, S. Krajewski, and E. Ruoslahti. 2008. Mitochondrial/cell-surface protein p32/gC1qR as a molecular target in tumor cells andtumor stroma. Cancer Res. 68:7210–7218.
10. Fox, G., N. R. Parry, P. V. Barnett, B. McGinn, D. J. Rowlands, and F.Brown. 1989. The cell attachment site on foot-and-mouth disease virus in-cludes the amino acid sequence RGD (arginine-glycine-aspartic acid).J. Gen. Virol. 70(Pt. 3):625–637.
11. Ghebrehiwet, B., B. L. Lim, R. Kumar, X. Feng, and E. I. Peerschke. 2001.gC1q-R/p33, a member of a new class of multifunctional and multicompart-mental cellular proteins, is involved in inflammation and infection. Immunol.Rev. 180:65–77.
12. Guo, W. X., B. Ghebrehiwet, B. Weksler, K. Schweitzer, and E. I. Peerschke.1999. Up-regulation of endothelial cell binding proteins/receptors for com-plement component C1q by inflammatory cytokines. J. Lab. Clin. Med.133:541–550.
13. Jiravanichpaisal, P., K. Soderhall, and I. Soderhall. 2006. Characterizationof white spot syndrome virus replication in in vitro-cultured haematopoieticstem cells of freshwater crayfish, Pacifastacus leniusculus. J. Gen. Virol.87:847–854.
14. Li, D. F., M. C. Zhang, H. J. Yang, Y. B. Zhu, and X. Xu. 2007. Beta-integrinmediates WSSV infection. Virology 368:122–132.
15. Li, H. X., X. L. Meng, J. P. Xu, W. Lu, and J. Wang. 2005. Protection ofcrayfish, Cambarus clarkii, from white spot syndrome virus by polyclonalantibodies against a viral envelope fusion protein. J. Fish Dis. 28:285–291.
16. Lim, B. L., K. B. Reid, B. Ghebrehiwet, E. I. Peerschke, L. A. Leigh, and K. T.Preissner. 1996. The binding protein for globular heads of complement C1q,gC1qR: functional expression and characterization as a novel vitronectinbinding factor. J. Biol. Chem. 271:26739–26744.
17. Liu, H., P. Jiravanichpaisal, I. Soderhall, L. Cerenius, and K. Soderhall.2006. Antilipopolysaccharide factor interferes with white spot syndromevirus replication in vitro and in vivo in the crayfish Pacifastacus leniusculus.J. Virol. 80:10365–10371.
18. Liu, H., K. Soderhall, and P. Jiravanichpaisal. 2009. Antiviral immunity incrustaceans. Fish Shellfish Immunol. 27:79–88.
19. Luana, W., F. Li, B. Wang, X. Zhang, Y. Liu, and J. Xiang. 2007. Molecularcharacteristics and expression analysis of calreticulin in Chinese shrimpFenneropenaeus chinensis. Comp. Biochem. Physiol. B Biochem. Mol. Biol.147:482–491.
20. Luo, T., X. Zhang, Z. Shao, and X. Xu. 2003. PmAV, a novel gene involvedin virus resistance of shrimp Penaeus monodon. FEBS Lett. 551:53–57.
21. Mallick, J., and K. Datta. 2005. HABP1/p32/gC1qR induces aberrant growthand morphology in Schizosaccharomyces pombe through its N-terminal alphahelix. Exp. Cell Res. 309:250–263.
22. Matthews, D. A., and W. C. Russell. 1998. Adenovirus core protein Vinteracts with p32: a protein which is associated with both the mitochondriaand the nucleus. J. Gen. Virol. 79(Pt. 7):1677–1685.
23. Nguyen, T., B. Ghebrehiwet, and E. I. Peerschke. 2000. Staphylococcus aureusprotein A recognizes platelet gC1qR/p33: a novel mechanism for staphylo-coccal interactions with platelets. Infect. Immun. 68:2061–2068.
24. Peerschke, E. I., K. B. Reid, and B. Ghebrehiwet. 1994. Identification of anovel 33-kDa C1q-binding site on human blood platelets. J. Immunol. 152:5896–5901.
25. Ruoslahti, E. 1996. RGD and other recognition sequences for integrins.Annu. Rev. Cell Dev. Biol. 12:697–715.
26. Seytter, T., F. Lottspeich, W. Neupert, and E. Schwarz. 1998. Mam33p, anoligomeric, acidic protein in the mitochondrial matrix of Saccharomycescerevisiae is related to the human complement receptor gC1q-R. Yeast 14:303–310.
27. Sieczkarski, S. B., and G. R. Whittaker. 2002. Dissecting virus entry viaendocytosis. J. Gen. Virol. 83:1535–1545.
28. Soderhall, I., E. Bangyeekhun, S. Mayo, and K. Soderhall. 2003. Hemocyte
FIG. 9. Recombinant PlgC1qR could bind to VP15, VP26, andVP28 of WSSV. (A) Using far-Western overlay blotting, we found thatthe rPlgC1qR could bind to WSSV VP15 (arrowhead 1), 26 (arrow-head 2), and 28 (arrowhead 3). (B) The binding between rPlgC1qRwith VP26 (arrowhead 1) and VP28 (arrowhead 2) of WSSV wasconfirmed by GST pull-down assay. GST was used as a control in bothassays (lane 3 in panel A; lanes 1 and 3 in panel B).
10850 WATTHANASUROROT ET AL. J. VIROL.
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

production and maturation in an invertebrate animal: proliferation and geneexpression in hematopoietic stem cells of Pacifastacus leniusculus. Dev.Comp. Immunol. 27:661–672.
29. Soderhall, I., Y. A. Kim, P. Jiravanichpaisal, S. Y. Lee, and K. Soderhall.2005. An ancient role for a prokineticin domain in invertebrate hematopoi-esis. J. Immunol. 174:6153–6160.
30. Sritunyalucksana, K., W. Wannapapho, C. F. Lo, and T. W. Flegel. 2006.PmRab7 is a VP28-binding protein involved in white spot syndrome virusinfection in shrimp. J. Virol. 80:10734–10742.
31. Tsai, J. M., H. C. Wang, J. H. Leu, A. H. Wang, Y. Zhuang, P. J. Walker,G. H. Kou, and C. F. Lo. 2006. Identification of the nucleocapsid, tegument,and envelope proteins of the shrimp white spot syndrome virus virion. J. Vi-rol. 80:3021–3029.
32. van den Berg, R. H., M. C. Faber-Krol, R. B. Sim, and M. R. Daha. 1998. Thefirst subcomponent of complement, C1q, triggers the production of IL-8,IL-6, and monocyte chemoattractant peptide-1 by human umbilical veinendothelial cells. J. Immunol. 161:6924–6930.
33. van den Berg, R. H., F. Prins, M. C. Faber-Krol, N. J. Lynch, W. Schwaeble,L. A. van Es, and M. R. Daha. 1997. Intracellular localization of the humanreceptor for the globular domains of C1q. J. Immunol. 158:3909–3916.
34. van Hulten, M. C., M. Reijns, A. M. Vermeesch, F. Zandbergen, and J. M.Vlak. 2002. Identification of VP19 and VP15 of white spot syndrome virus(WSSV) and glycosylation status of the WSSV major structural proteins.J. Gen. Virol. 83:257–265.
35. van Hulten, M. C., J. Witteveldt, M. Snippe, and J. M. Vlak. 2001. Whitespot syndrome virus envelope protein VP28 is involved in the systemicinfection of shrimp. Virology 285:228–233.
36. Wan, Q., L. Xu, and F. Yang. 2008. VP26 of white spot syndrome virusfunctions as a linker protein between the envelope and nucleocapsid ofvirions by binding with VP51. J. Virol. 82:12598–12601.
37. Wang, F. Z., S. M. Akula, N. Sharma-Walia, L. Zeng, and B. Chandran.2003. Human herpesvirus 8 envelope glycoprotein B mediates cell adhesionvia its RGD sequence. J. Virol. 77:3131–3147.
38. Wang, H. C., H. C. Wang, J. H. Leu, G. H. Kou, A. H. Wang, and C. F. Lo.2007. Protein expression profiling of the shrimp cellular response to whitespot syndrome virus infection. Dev. Comp. Immunol. 31:672–686.
39. Williams, C. H., T. Kajander, T. Hyypia, T. Jackson, D. Sheppard, and G.Stanway. 2004. Integrin alpha v beta 6 is an RGD-dependent receptor forcoxsackievirus A9. J. Virol. 78:6967–6973.
40. Xie, X., H. Li, L. Xu, and F. Yang. 2005. A simple and efficient method forpurification of intact white spot syndrome virus (WSSV) viral particles. VirusRes. 108:63–67.
41. Xie, X., L. Xu, and F. Yang. 2006. Proteomic analysis of the major envelopeand nucleocapsid proteins of white spot syndrome virus. J. Virol. 80:10615–10623.
42. Xu, Z., A. Hirasawa, H. Shinoura, and G. Tsujimoto. 1999. Interaction of thealpha(1B)-adrenergic receptor with gC1q-R, a multifunctional protein.J. Biol. Chem. 274:21149–21154.
43. Yadav, G., R. L. Prasad, B. K. Jha, V. Rai, V. Bhakuni, and K. Datta.2009. Evidence for inhibitory interaction of hyaluronan-binding protein 1(HABP1/p32/gC1qR) with Streptococcus pneumoniae hyaluronidase.J. Biol. Chem. 284:3897–3905.
44. Yao, Z. Q., A. Eisen-Vandervelde, S. N. Waggoner, E. M. Cale, and Y. S.Hahn. 2004. Direct binding of hepatitis C virus core to gC1qR on CD4� andCD8� T cells leads to impaired activation of Lck and Akt. J. Virol. 78:6409–6419.
45. Yao, Z. Q., S. Ray, A. Eisen-Vandervelde, S. Waggoner, and Y. S. Hahn.2001. Hepatitis C virus: immunosuppression by complement regulatory path-way. Viral Immunol. 14:277–295.
VOL. 84, 2010 PlgC1qR PREVENTS WSSV REPLICATION 10851
RETRACTED
on April 4, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Retraction for Watthanasurorot et al., A gC1qR Prevents White SpotSyndrome Virus Replication in the Freshwater Crayfish Pacifastacusleniusculus
Apiruck Watthanasurorot,1 Pikul Jiravanichpaisal,1,2 Irene Söderhäll,1 and Kenneth Söderhäll1
Department of Comparative Physiology, Uppsala University, Uppsala, Sweden,1 and National Center for Genetic Engineering and Biotechnology, National Science andTechnology Development Agency, Pathumthani, Thailand2
Volume 84, no. 20, p. 10844 –10851, 2010. The authors and the journal hereby retract this article. After publication, this article wasfound to have multiple images that were unacceptably manipulated by Apiruck Watthanasurorot, a clear violation of ASM’s ethicalstandards. In Fig. 6B, the 40S panels are duplicated from Fig. 5B, and PlgC1qR bands in lanes 1 to 3 seem to be duplicated in lanes 7 to9 in reverse orientation. In Fig. 7A and Fig. 8A, the 40S panels seem to be duplicated from Fig. 2A (lanes 6 to 11), in both forward (lanes7 to 12) and reverse (lanes 1 to 6) orientations. WSSV VP28 lanes 9 and 10 seem to be duplicated in lanes 11 and 12 for Fig. 8A. Sincethe integrity of the data as presented was compromised and A. Watthanasurorot cannot provide reliable original files for these figures,this publication is retracted in its entirety. We apologize to the readers of Journal of Virology and regret any inconvenience this causes.A. Watthanasurorot could not be reached when asked to agree to the retraction.
Citation Watthanasurorot A, Jiravanichpaisal P, Söderhäll I, Söderhäll K. 2016.Retraction for Watthanasurorot et al., A gC1qR prevents white spot syndromevirus replication in the freshwater crayfish Pacifastacus leniusculus. J Virol90:1154. doi:http://dx.doi.org/10.1128/JVI.02628-15.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
RETRACTION
crossmark
1154 jvi.asm.org January 2016 Volume 90 Number 2Journal of Virology




























