Embed Size (px)
Citation preview

A CYTOCHEMICAL STUDY OF .ACID PHOSPH.\TASE AND NUCLEASE ACTIVITY IN T H E PLXSTID OF CLOSTERIUi1.I ACEROSUiII'
Philip I.V. Cook Department of Botany, University of L'ermont. Burlington, Vermont 05401
SUMMARY
Acid phosphatase activity in Closterium acerosum hcis been studied using t h e C o m o r i rind the azo dye procedures. A modif icat ion of t h e G o m o r i m e t h o d was used for detecting t h e dis tr ibut ion of acid nu- cleases. T h e plastid is t h e major site of acid phos- phntase activity which may be primari ly w i t h i n py- renoids, betrcven pyrenoids, or throughout t h e plas- t id . T h e Gomori Procedlire showed actiuity w i t h i n / l i e pyrenoids or in the central core of t h e plastid, whereas the NZO dye method showed activity through- orrt w i t h a n orcasionrrl tendcncy to be localized near t h e ends. N o other cytoplasmic activity was ob- served but evidence for occasiond activity i n t h e nriclezis is presented. Alkaline phosphatase could n o t be detected. Acid nrrclense activity, which results in the d e p d a t i o n or D N A , R N A , rind RXA-Core , h s beerr f o u n d i r 7 both the niiclezis and the plnstid. Plastid activity is heat k b i l e , iuliereas n riclear ac- tivity is only slightly diminished af ter 3 min at 100 C . T h e results (ire interpreted NS indicat ing at k i s t 2 rrcid phosphiitases rind 2 niiclenses in C . acerosum. T h e f indings are disciissed w i t h respect to t h e distri- Di/tio,i of siinilar enzymes itz olher orpnisms.
INTRODL'CTlON
Although many enzymes have been isolated from algae and characterized biochemically ( I I ) , there has been relatively little attention devoted to their intra- cellular distribution apart from data derived from cell fractionation studies. U'hile fractionation is use- ful for those algae which can be grown in quantity, not all forms lend themselves to mass culture. Pro- cedures now exist for the cytochemical detection of a variety of enzymes (5,15) and many of the larger, slower growing algae are admirably suited for cyto- chemical studies. Such an approach can be expected to yield information on changes in the activity or distribution of selected enzymes during normal mor- phogenesis or during various abnormal physiological conditions. T h e present report is concerned with acid phosphatase and nuclease activity in healthy vegetative cells of Closteriztm aceroszim (Schrank) Ehrbg. This study is designed to provide a basis for
1 This investigation was supported by National Science Foun- dation Institutional Grant GU-1505. I would like to thank Dr. M. Nasitir for donating certain of the nucleic acid preparations, Dr. A. Gel-shov for reading and criticizing the manuscript, and several colleagues for their discussions and constructive criti- cism. Rrrrivrtf Jnniin? 35, 196;: revised Xlorch 10, 1967.
future investigation of enzyme distribution in cells parasitized by various phycomycetes.
MATERIALS AND METHODS
T h e alga (clone So. 994, of personal collection) used in this study is characterized bv slightly curved elongate cells rang- ing from 300-330 p in length and 36-42 p in diameter (Fig. 1). T h e alga was grown in soil-water cultures lacking calcium carbonate and was maintained on a 12-hr light : I?-hr dark cycle at 25 C. The light intensity varied somewhat but cul- tures were generally kept at 50-100 ft-c. Under these conditions starch accumulation was reduced and the plastid sti-ucture was easilv observed. Cclls from cultures 2-8 weeks old were used for the cytochemical tests. There was some variability in enzyme activity between different cultures but this could not tic correlated with culture age. T h e general mot-phological appenrnnce of cells in a l l cultures was normal.
Cells werc fiver1 i n cold formol-calcium (1% CaCl, in a 10% formalin solution) or in cold acetone. Various fixation times were rricd, a short fixation being as satisfactory as the usual overnight fixation. T h e results described in this paper are Ixised upon cells fixed for 15-20 min in formol-calcium or for 3 ( ! 4 5 min in acctone, both fixations carried out in an ice bath. The best cytoplasmic fixation was obtained using formol- calcium. Acetone fixation resulted in many lysed cells and much intracellular distortion. However, acetone-fixed cells provided a useful comparison and, because tlie chlorophyll was extracted, were better suited for the azo dye acid phos- phatase method. Unfixed cells failed to reveal any enzymatic activity, presumably due to the impermeability of living cells to certain of the reagents. Following fixation, the cells were washed twice by centrifugation in distilled water. Then 20-50 cells were transferred by capillary pipette to depression slides containing the appropriate incubation mixture.
In all cases tlie cells were incubated at 25 C rather than at tlie standard 35 C used for mammalian tissues. The lower tem- perature was found to be more convenient and to produce equally good results. Optimum incubation times were 1-2 hr for the Gornori acid phosphatase method, 30 min for the azo dye procedure, and 4 8 hr for the nuclease method. For those tests which required the visualization of lead phosphate deposits. the cells were transferred by capillary pipette through 2 changes of distilled water, a 2% acetic acid rinse, 2 addi- tional rinses in distilled water, and finally into 0.5% ammonium sulfide. Rinses were made in depression slides under a binocu- lar dissecting microscope.
.Acid phosphatase activity was detected using the Gomori metal- salt procedure or the naphthol-AS phosphate azo dye method (5 ) . The Gomori incubation mixture consisted of 0.01 hi
sodium glvcerophosphate (60% @-form; Nutritional Biochem. Co.) and 0.003 >I Pb (NO3), in 0.05 r.5 acetate buffer at pH 5.0. The precipitate formed upon mixing was allowed to settle and the clear supernatant was used for the tests. T h e mixture was kept refrigerated and prepared fresh every month. T h e azo dye mixture was prepared using naphthol AS-TR phos- phate (Ps'utritional Biochem. CO.) as a substrate and Fast Red Violet LB salt (Dajac) as a coupling agent. T h e solution was used immediately after mixing. T h e naphthol AS-TR phosphate

ES%Y;\IE LOC,\Ll%.-\TION I N .\ C L O S T E K l f 'i\f
T.\BLE 1. Sotiices of niicleic ctcids used ns m h s t m t e s . -
Type of nucleic Biological Commercial
nc1d source source Result
DNA Calf thymus Worthington Biochem. Corp. - (highly
merizetl)
highly
m eri ze (I)
POlY-
DNA Sperm (not Nutritional Biochem. Co. + POIY -
R N A Yeast U'orthington Biochem. Corp. + R N;Z Yeast Nutritional Biochem. Co. + RNA-Core Yeast Sigma Chemical Co. +
gave much better results than either naphthol AS-BI or AS-AN phosphate. Controls consisted of cells which had been heated, incubated in the presence of 0.001 11 NaF or, in the case of the Gomori procedure, incubated in the absence of the sub- strate.
Both the metal-salt and the azo dye procedures for alka- line phosphatase ( 5 ) were tried but no activity was detected.
i\ modification of the mctliocl of Aronson et nl. (190) was used for the detection of acid nuclease activity. This method is based upon the assumption th:it mononucleotitles are re- leased as a result of nuclease activity. In the presence of adtlctl acid phosphatase or intracellular nucleotidase, the nucleotides arc hydrolyzcd and the liberated phosphate immediately pre- cipitates in the form of lead phosphate. T h e site of lead phosphate is then visualizcd as for the Gomori acid phos- plintase method 2nd the black dcposits are assumetl to rcp- rcscnt tlic site of nuclease. Several sources of DNA and RNA wcrc used :IS sulxtratcs, and arc listed in Table 1. The final conccntration~ i n thc incubation misture wcrc: substrate, 20 nig/lOO ml; acid phosphatase (wheat germ. Nutritional Biochcm. Co.) 10 mg/100 ml; lead nitrate, 0.002 *I: and acetate buffer pH 5.0. 0.05 > I . A stringy precipitate formed if the sub- strate and acid phosphatase were initially dissolved in the same solution. T h e following procedure eliminated this diffi- culty. Twenty mg of substrate were dissolved in 50 ml of 0.1 M acetate buffer, pH 5.0. Ten mg of acid phosphatase were dissolved in 50 ml distilled water and then added to the buffer solution containing the substrate. This was followed by the addition of 2 ml of a 0.1 XI lead nitrate solution. A flocculent precipitate developed after mixing but did not appear to interfere with the reaction. T h e incubation misture was always used immediately after preparation. Controls consisted of cells incubated without the substrate.
RESULTS
1. Nonspeci f ic lead precipitation. Since the Go- mori method is based upon lead phosphate precipita- tion and its subsequent visualization, i t was neces- sary to examine cells incubated in the absence of a substrate to determine the extent of lead precipita- tion due to nonenzymatic factors. In some control cells, small granules (ca. 0.5 I-L diam.) are scattered throughout the plastid and often show a tendency to be concentrated at the ends of the plastid near the nucleus and the terminal vacuole (Fig. 2 , 3). T h e number of granules varies from cell to cell but they are never abundant enough to obscure results when a substrate is present in the incubation mixture. It is not known whether the granules represent poly- phosphates or are derived from some other source.
FIG. 1. Diagram o f C. I K C ~ O S I L ) I I showing the prominent cel- lular fcatures. X 275.
2 . Acid phosphntase activity. The intensity of acid phosphatase activity varies between different cultures and often between cells from the same cul- ture. In some tests scarcely any reaction product was observed. Despite this variation, a general pattern of distribution can be established for a given type of fixation and for each method of detection.
With the Gomori method, enzyme activity is indi- cated by a nonparticulate blackish-brown stain which contrasts with the discrete granules of the control. In formol-calcium-fixed cells the stain is confined to the plastid (Fig. 4). The site of the heaviest stain varies from within the pyrenoid (Fig. 5) to the cen- tral core of the plastid between adjacent pyrenoicls (Fig. 6). Often, both types of distribution may be found within the same cell. As the incubation time is increased, the entire cell becomes blackened, in- cluding the wall. The gradual spread of the lead deposit with time is most likely due to diffusion of acid phosphatase and/or the released phosphate. With one exception, there was no evidence of enzyme activity outside of the plastid in formol-calcium- fixed cells. A pronounced nuclear stain was observed in a single test, but this observation has not been re- peated in numerous trials.
Intra- and interpyrenoid staining is also observed in acetone-fixed cells. However, a major difference after acetone fixation is the appearance of some cells with a nuclear stain in nearly all tests (Fig. 7). In many nuclei the stain is strongest in the nucleolar region. T h e proportion of cells showing nuclear activity has ranged from less than 5% to over 70%

26 PHILIP it'. COOK
ancl, in a few cells, activity was confined to the nu- cleus (Fig. 8). I t was initially thought that perhaps a formalin-sensitive enzyme was present within the nucleus. However, when acetone-fixed cells were placed in cold formol-calcium for an additional 15 min before incubation, the sporadic nuclear staining persisted. T h e variability oE nuclear activity is puz- zling but independent evidence for the occasional presence of acid phosphatase in the nucleus is pre- sented later.
T h e azo dye acid phosphatase method results in a nonparticulate pink stain which is usually evenly distributed in the plastitl with little o r no stain in the remaining cytoplasm except after prolonged in- cubation. Occasionally, some staining of the nucleus occurs (Fig. 9). .I few of the cells show regions of intense staining near the ends of the plastitl (Fig. 10, 11). These sites are not associated with any structure but they do correspond with the areas of greatest 1e:itl precipitation in control cells. Strong intr;ipyrenoitl staining was uncommon although the smie poptilation of cells often showed pyrenoitl ac- t i v i t y when the Gomori method w x used.
Heating the cells for 10 min in ;I boiling water b;ith completely destroys ;ill ;icitl phosphntxe activ- i t y ;inti the presence of 0.001 XaF strongly in- hibit.\ [he activity tletectetl by either oE the two me thotls.
3. Acid ririclr,cc.sc ctctivilg. Xlthough the following observations (lo not indicate specific DNases or RNases, i t should be kept i n mind that i f present they could contribute to the results. IVith the ex- ception of c ~ l f thymus DN.4, the pattern ot' nuclease activity is similar with either DN.4 or RN.\ as a sub- strate. Some minor variations in intensity antl tlis- tribution were observed but they have not been con- sistent. Considerable difficulty was encountered in tlisolving the highly polymerized thymus DN.4 and lack of activity with this substrate is assumed to be due to poor penetration into the ClostrTi irn cell. Vorbrodt (20) notes that a much longer incubation time is required for highly polymerized DNA and suggests that diffusion of the substrate is slow.
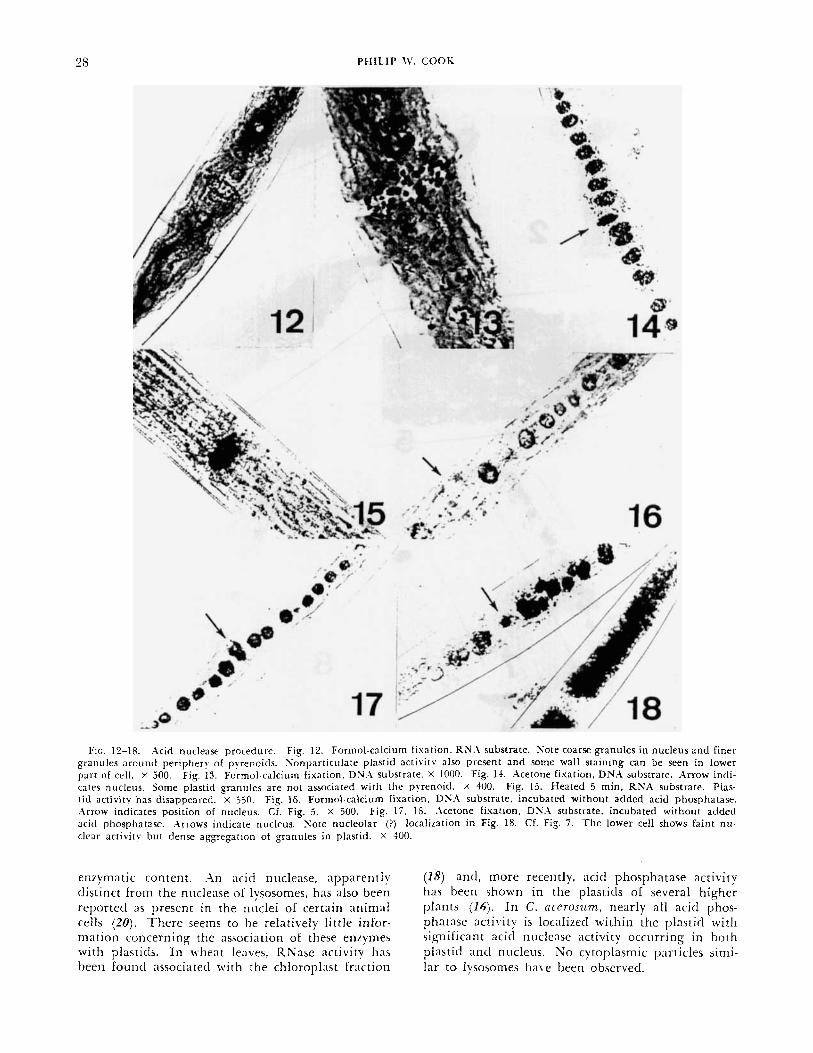
Formol-calcium-fixed cells consistently show nu- clease activity in the plastic1 and in the nucleus (Fig. 12). Nuclear activity is characterized by discrete and somewhat irregular granules, about 1-2 p diam., which tend to be spaced near the surface of the nu- cleus (Fig. 12, 13). I t is tempting to equate the granules with chromosomes. Preliminary counts are similar to the lower chromosome numbers reported for C. ncerosiiin (8). T h e distribution of nuclease activity during mitosis should provide some interest- ing information. Two types of plastitl activity were noted, granular and nonparticulate. Granules in the plastitl tend to be arranged around the periphery of the pyrenoid (Fig. 12, 13). Occ;isionally, granules are present in other portions of the plasticl (Fig.
14) antl are sometimes dense enough to entirely ob- scure the pyrenoids (Fig. IS). T h e nonparticulate activity resembles the stain obtained with the Go- mori acid phosphatase method. This could be the result of nucleoticles diffusing from the original site of nucleic acid digestion before they are hydrolyzed. There is also a tendency for the older segments of the wall to become blackened, especially when RNX is used as a substrate (Fig. 12). T h i s may represent adsorption of a diffusing enzyme rather than the in . r i tu presence of a nuclease.
A\ marked difference between plastitl and nuclear activity is shown by their heat sensitivity. UnEixetl cells which have been heated in a boiling water bath for 5 min retain most of their nuclear activity whereas the plasticl staining completely disappears (Fig. 15). Heated cells often exhibit a strong clarken- ing of the cell walls. Localization remains the same with either RNX or DNA as a substrate.
.\cetone-fixetl cells show the same localization of activity (Fig. 1-1) but the granules in the nucleus ;ire iisunlly coalesced and the nonparticulate stain in the plastid is diminished.
T h e localization of nuclease is tlepentlent upon hytlrolysis of the liberated nucleotitle t i v i t y in the absence of atltled a shoiiltl indicate the simultaneous presence of nu- clease and endogcnoits acid phosphatase or niicleo- titlase. When acid phosphntase is omitted from the incubation mixture, nuclease activity parallels the distribution of acid phosphatase described earlier. I f formol-calcium-fixed cells are incubated with the substrate, but without acltletl acid phosphatase, there are granules around the pyrenoids but no nuclear ac- t iv i ty (Fig. 16). However, acetone-fixed cells often show nuclear activity in addition (Fig. 17, 18) antl occasional cells have nuclear activity only. There was often a definite reaction associated with the nircleolus (Fig. 18). Although the participation of ;in intracelliilar nucleotidase is a possibility, there is n o evidence of nucleotitlase activity in the nuclei of formol-calcium-fixed cells under the incubation conditions used. Furthermore, 5'-nucleotidase is ac- tivated by Mn++ ancl Mg-+ (5 ) antl these cations were not ndtled to the mixture. I t therefore seems likely that the observed activity is the combined result of endogenous acid phosphatase antl nuclease.
DISCUSSION
T h e results obtained in this study provide an in- teresting comparison with the normal distribution of acid hyclrolases in other organisms. Acid phos- phatase and the acid nucleases have been considered characteristic components of lysosomes in aninial cells (6,14). Cytochemical studies (2,7,12), supple- mented with biochemical investigations (9,13), have intlicaterl the presence of particles within p l a n t cells which are equivalent to lysosomes in their size and

ENZYME LOCALIZATION IS A CLO.YTERlC',\.I
FIG. 2. Control, formol-calcium fixation. Note nonspecific lead deposit in granules, especially near nucleus. T h e background i n this and other formol-calcium-fixed cells is due to the presence of chlorophyll. X 675. FIG. 3. Control, acetone fixation. Concen- tration of granules near the end of plastid. X 675. FIG. 6 6 . Gomori acid phosphatase procedure, formol-calcium fixation. Note absence of nuclear and cytoplasmic activity outside the plastid. Prominent intrapyrenoid activity in Fig. 5, interpyrenoid activity in Fig. 6. Fig. 4, X 325; Fig. 5 , 6, X 900. FIG. 7 , 8. Gomori acid phosphatase procedure, acetone fixation. Note definite intrapvrenoitl activity and nuclear (nucleolar?) activity (arrow) in Fig. 7 . Fig. 8 shows cell with nuclear activity only. X 400. FIC. 9-1 I . Azo dye acid phosphatase procedure, acetone fixation. Note diffuse activity in 110th plastids and nucleus in Fig. 9; regions of intense plastid activity in Fig. 10 and 11. Arrow indicates position of nucleus in Fig. 1 1 . Fig. 9, X 400; Fig. 10, 11, X 350.
PHILIP 11’. COOK
FIG. 12-18. .acid nuclease procedure. Fig. 12. Formol-calcium fixation. R N h substrate. Sore coarse granules in nucleus a n d finer granules around periphery of pyrenoicls. Sonpar t ic~r la te plastid activitv also present and some wall staining can be seen in lower part of cell. X 500. Fig. 13. Formol-calcium fixation, DN.1 substrate. X 1000. Fig. 14. Acetone fixation. DNA substrate. Arrow indi- cates nucleus. Some plastid g a n u l e s are not associated with the pyrenoid. X 400. Fig. 15. Heated 5 min, RNA substrate. Plas- Lid activity has disappeared. x 350. Fig. 16. Formol-calcium fixation, DS.1 substrate, incubated without added acid phosphatase. Arrow indicates position of nucleus. Cf. Fig. 5 . X 500. Fig. 17, IS. Acetone fixation, DXA substrate. incubated without added acid phosphatase. .\mows indicate nucleus. S o t e nucleolar (?) localization in Fig. IS. Cf. Fig. 7. T h e lower cell shows faint n u - clear activity but dense aggregation of granules in plastid. X 400.
enzymatic content. .in acid nuclease, apparently ( I S ) and, more recently, acid phosphatase activity distinct froin the niiclease of lysosomes, has also been has been shown in the plasticls of several higher reported a s present in the nuclei of certain animal plants (16). In c. ncerosum, nearly all acid phos- cells (20). T h e r e seeins to be relatively little infor- phntase activity is localized within the plastic1 with mation concerning the association of these e n q m e s significant acid nuclease activity occurring in both with plasticls. In wheat leaves, RNase iictivitv has plastic1 a n d nucleus. N o cytoplasmic particles simi- been found associated with the chloroplast traction lar to lysosomes ha\-e been observed.

ENZYXIE LOC.\LIZ.\TION I N A CLO.STERII',\[ ?!I
The frequent association of acid phosphatase and nuclease with the pyrenoicl suggests that this spe- cialized region may have functions in addition to starch synthesis. At the present time, the exact role of these enzymes in Closteritrm is unclear. It is gen- erally assumed that the main function of the ly- sosomal enzymes is hydrolytic and that they partici- pate in intracellular digestion and autolysis (6). This function would seem highly unlikely for the plastid enzymes. Acid phosphatases from various sources are reported to be capable of phosphate transfer as well as hydrolysis (cited in 19) and certain nucleases are reported to catalyze transfer of nucleoticles (17). It is possible that these enzymes coulcl play a part in the anabolic processes within the highly structured membrane system of an unfixed plastid.
With respect to acid phosphatase activity, the re- sults suggest the presence of a t least 2 enzymes which differ in substrate specificity antl localization. One, acting upon naphthol AS phosphates, shows a rather generalized distribution throughout the plastid with occasional ill-defined regions of intense activity. The second, detected by the Gomori procedure, is con- fined to the central core of the plastid antl often shows distinctive intrapyrenoitl localization. The question of a nuclear enzyme needs further study. Some investigators believe that sporadic nuclear ac- t iv i ty is ;in artifact (1O,Z2) but the following ob- servations provide more positive evidence. Since the nuclear staining is substrate dependent, i t cannot be clue to nonspecilic lead adsorption. If nuclear activity were clue to clilfusion of the enzyme from the plastid, one wouIcl not expect to find cells in which activity was confined to the nucleus. Further- more, by omitting the exogenous acid phosphatase, the nuclease procedure provides independent evi- dence for the occasional presence of acid phosphatase in the nucleus. Double fixation indicates that the nuclear enzyme is not inactivated by formalin but rather that acetone treatment results in its activation. Brachet (4 ) points out that, in contrast to m;iny other enzymes, acid phosphatase synthesis ceases when nuclei are removed from Amoeba and Acetabtr larin and he suggests that acid phosphatase might be synthesized primarily in the nucleus. If this were the case in Closterizim, acetone treatment could acti- vate the nascent enzyme during periods of nuclear synthesis. As a possible explanation for the vexing variability of activity, it is reported that acid phos- phatase synthesis is incluced by low phosphate con- centration (3) . Further studies of Closterizim grown in defined media should help explain some of the variation in both intensity and localization of acid phosphatase activity.
T h e success of the nuclease procedure was siir- prising since it seemed unlikely that the substrate and the acid phosphatase in the incubation mixture would penetrate the intact Closteriiim cell. Al-
though a relatively long incubation time was re- quired, the substrate apparently did reach all por- tions of the cell. X comparison of cells incubated with and without exogenous acid phosphatase showed that pyrenoid activity was more intense when the exogenous enzyme was present, indicating that the latter was diffusing into the center of the plastid as well as into the nucleus. T h e results show the pres- ence of a t least 2 enzymes, differing in their heat sensitivity antl intracellular distribution, which are capable of hydrolyzing nucleic acids. T h e observed activity could be the result of phosphodiesterases, nonspecific nucleases, or of enzymes which are spe- cific for a particular nucleic acid. RNA-Core, ,which is the nondialyzable residue after digestion with puri- fied RNase, was hydrolyzed in both the nucleus antl plastitl. This suggests that the Closterilrm enzymes have a relatively broad specificity. T w o nucleases with a broad specificity iire also intlicatetl by the parallel localization of activity with either DNA or RNA as a substrate, repardless of the type of fixa- tion. Whether these nucleases also possess a more general phosphotliesterase activity awaits further study. Preliminary attempts to use the phospho- diesterase substrate, calcium his-1)-nitrophenyl phos- phate, ;IS a cytochemical reagent have been unsuccess-
It should be emphasized that the results of this study have been based largely upon methods devised for animal tissues. Although the principles of cyto- chemical localization remain the same, enzymes in plant cells can be expected to differ in their substrate specificity, pH optimum, activators and inhibitors. Further investigation of these factors is necessary for a more complete characterization of the enzymes reported here.
rui.
1.
2.
3.
4.
5.
6.
-
8 .
9.
REFERENCES
.\RONSON, J., H E M P E L M A N N , L. H.. & OKADA, S. 1958. Pre- liminary studies on the histological demonstration of des- oxyribonuclease I1 by adaptation of the Gomori acid phosphatase method. J . Histochem. Cytocltena. 6:255-9. AVERS, C. L KING, E. E. 1960. Histochemical evidence of intracellular enzymatic heterogeneity of plant mitochondria. Ain. J. Bat. 47:220-5. BLUM, J. J. 1965. Observations on the acid phosphatases of Euglena gmcilis. J . Cell Biol. 24:2234. BRACHET, J. 1961. Nucleocytoplasrnic interactions in uni- cellular organisms. In Brachet. J. & hlirsky. A. [eds.], The Cell , Vol. 2. Academic Press, N. Y., 771-841. BURSTONE, M. S. 1962. Enzyme Histochettristr~~ and I t s Applicotion in the Study of Neo$lastiis, Academic Press, N. Y. DE DUVE, C. 1959. T h e function of intracellular h y - tlrolases. EsP. Cel l Res., Suppl. 7:169-82. G A H A N , P. B. 1965. Histochemical evidence for the pres- ence of lysosome-like particles in root meristem cells of l'icia fnbn. J . E x p . Bot. 16:350-5. GODWARD, M. B. E. L PUISEIJX-DAO, S. 1966. T h e Cliloro- pliyceae. Iiz Godward. 31. B. E. [ed.], The Chrottiosonm of the Algae, Edward Arnold Ltd., London, 1-T7. H A R R I N C T O N , J. F. & ALTSCHUL, A. M. 1963. Lysasome-

H I D E 0 l lV. \S . \Kl 30
like hehavior in germinating onion seeds. Federntion Proc. 223475. (.\hstr.)
10. HOI.T, S. J. 1959. Factors governing the validity of stain- ing methods for enzymes, and their hearing upon the Gomcri acid pliosphatase technique. E x P . CelL Res. , SUPPI.
11. J ~ C O B I , G. 1362. Enzyme systcms. 271 Lcwin. R. .\. [ed.]. Phyriologv a n d Biochemistrs of Algae, Academic Press, N. Y. , 12540.
12. JENSEN, t V . A. 1956. The c)toclicmical localization of acid phospliatase in root tip cells. Am. ]. Bot. 43:5&34.
13. MATILE, P.. BALZ. P. J., SEMADENI. E., k JOST, M. 1965. Isolation of splierosomes with lysosome characteristics from seedlings. 2. NntiirforSch. 2Ob:693-8.
14. SOVIKOFF. ‘4. 1961. Lysosomes and related particles. In Rrachet. J. k Xlirsky, A. [eds.]. The Cell , Vol. 2, .\cademic Press, N. Y., 423-88.
7:1-27.
13. PEARSE. A . G . E. 1960. Histoclieinistry: Theoret ical ~ n d .-IPfilied. Little, Brown 8c Co.. Boston.
16. RMETLI, H. W. J.. ~ V E I N T R \ U B . M.. & R I N K . U. M. 1366. Latent acid phosphatase in chloroplasts. Con. I. Bot. 44: 1723-5.
17. RAZZELL. l ,V. E. & KHOR.PNA, H. G. 19661. Studies on polynucleotides X. Enzymatic degradation. Some proper- ties and mode of action of spleen phosphodiesterase. ]. B i d . Chetn. 236:ll-M-9.
18. ROHRINCER. R., SANBORSKI, D. J.. & PERSON. C. 0. 1961. Rihonuclease activity in rusted wheat leaves. Con. ]. Rot. 39:775-04.
19. SCHMIDT. G. 1961. Nonspecific acid phosphomonoester- ases. I n Bover. P. D.. Lardy, H.. & Myrback. K . [etls.]. The Enzymes, 2nd ed., Vol. 5 . Academic Press, N. Y., 3 7 4 7 .
20. VORBRODT, A. 1961. Histochemical studies on the intra- cellular localization of acid deoxyrihonuclease. /. Histo- che in . Cyfochem. 9:64i -55 .
NUTRITION.AL STCDIES OF T H E EDIBLE SEXIVEED PORPHI’RA T E N E R A . 11. N LIT RI T I 0 N 0 F C O N C H 0 C E LIS‘
Hideo I i ~ ~ m a k i Facti l tv of Fislici-ics ;inti ;\nimal Husl)antlry. Hirosliima Univcrsity. F u k u y m a . Hiroshima. Japan
S L: XI A I A R Y
T h e nutr i t ion of the free-living phase of Conch- ocelis of P. tenera i u m studied axenicall>l. Conch- ocelis preferred NO,, as nitrogen sotirce. Urea nnd h;H, i n low concentration, mp(iragine, nnd lysirze ruere very good A: solrrces. Seueral other amino acids were also utilized bitt groulth 7uas less abundant . Inorgaiiic crnd orgcrnic phosplzates zuere rttilized; t h q were required nl rel(itivr1y low concentrations. Gly- cerophosphnte ~ U I J C excellent growth in a cornpar(1- tiuely wide runge of concentrations (0.1-5 mg P 70). T h e opt imal Ca concentration was 10-100 m g ?c. Needs for boron, munganese, zinc, s t ront ium, rii- b id ium, l i th ium, and iodine were demonstrated. T h e iodine effect w a s remarkable (peak growth w i th 1 pg %); the effectiue concentration range was very narrow. Iron, cobalt, and bromine seemed to be adequately supplied as impuri t ies of the mncro- nutrients. A modif ied artificial m e d i u m (ASP, , I ) for the Conchocelis phnse is presented.
ISTRODUCTION
Although many ecological studies have been made on the Conchocebis phase of Porphyru tenera, knowl- edge of i ts nutrition is scanty. This ‘was clue mainly
‘This work was supported in par t hy the Science Research Fiind of the Ministry of Education. 2nd Contract Xonr 4062 of the Office of S a v n l Research, C.S..A.. to Haskins Laboratories. T h e author thanks Dr. L. Provasoli for help. R e c e i ~ ~ e d J w i e 1, 1966; revised Dereruber 10, 1966.
to the atrange inclination of Conchocelis to perfornte ant1 then grow in molluscan hells. Obviously, pre- cise nutritional stutlies can only he clone with nxenic free-living Coriclzocelis cultures ( I ) . This paper pre- sents data on Conchocelis nutrition.
SI.\TERIAL AND METHODS
Cenernl fiIocedrri-es. T w o or 3 newly tlevclopetl small colo- nies of bacteria-free Concliocelis were inoculated into flasks emploving Pasteilr pipettes connectetl with rubber tubing to a mout l i~~iece ; a glass hood provided with a germicidal lamp was used. Sterility tests in S T P ( I ) and ST, (2). both liquid and agarized. were included in each experiment. T h c colonies were grown in 100-ml Erlenmeyer flasks containing 40 ml of medium in a freezer-incubator illuminated with “natural” and
TABLE I . Basal inedium (mod i f i ed ASP,,iVTA).
Distilled water 100 ml Na, glycerophos-
NaCl 2.8 g Na,SiO,. 9 H 2 0 2 mg
MgCI,. 6H20 0.4 g P I1 metals” 1 ml KCI 70 m g S I1 metals” 1 m l
NaNO, 10 m g Nitrilotriacetic acid 10 mg KSPO, 1 m g PH 7.8-8.0
One ml of P I1 metals contains EDTA 1 mg, Fe (as C1-) 0.01 mg, B ( a s H,BO,) 0.3. mp, Mn ( a s Cl-) 0.04 nip, Zn (as C1-) 5 ,ug, Co ( 3 s CI-) 1 Fg.
One ml of S I1 metals contains Br ( a s Nil*) 1 ~ n p , Sr (as C1-) 0.9 mg. Rb (as CI-) 0.07 mg, Li ( a s Cl-), 0.02 mg. I (3s K * ) 1 f ig,
phate . jH,O 1 mg
MgS0, .SH20 0 . i g Vitamin B,? 0.1 ’g
Ca (as CI-) 40 m g Tris huffer 0.1 g
140 ( a s NahloO,) 0.05 mg.
![The conserved Fanconi anemia nuclease Fan1 and the SUMO E3 … · 2017. 2. 23. · FAN1 (Fanconi anemia-associated nuclease 1, or FANCD2/FANCI-associated nuclease 1) [13–18]. Human](https://img.dokumen.tips/doc/110x75/60c9d965c710eb0d72008d0e/the-conserved-fanconi-anemia-nuclease-fan1-and-the-sumo-e3-2017-2-23-fan1-fanconi.jpg)




























