Upload
angellisimal
View
227
Download
2
Embed Size (px)
Citation preview
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
1/24
Horse-Mounted Invaders From the Russo-KazakhSteppe or Agricultural Colonists From Western Central
Asia? A Craniometric Investigation of the Bronze AgeSettlement of Xinjiang
Brian E. Hemphill1*and J.P. Mallory2
1Department of Sociology and Anthropology, California State University at Bakersfield,Bakersfield, California 93311-10992School of Archaeology and Palaeoecology, Queens University, Belfast, Northern Ireland BT7 1NN, UK
KEY WORDS craniometry; phenetic distance; Tarim Basin; Bactria; China
ABSTRACT Numerous Bronze Age cemeteries in the
oases surrounding the Taklamakan Desert of the TarimBasin in the Xinjiang Uyghur Autonomous Region, west-ern China, have yielded both mummified and skeletalhuman remains. A dearth of local antecedents, coupledwith woolen textiles and the apparent Western physicalappearance of the population, raised questions as to wherethese people came from. Two hypotheses have been offeredby archaeologists to account for the origins of Bronze Agepopulations of the Tarim Basin. These are the steppehypothesis and the Bactrian oasis hypothesis. Eightcraniometric variables from 25 Aeneolithic and Bronze
Age samples, comprising 1,353 adults from the TarimBasin, the Russo-Kazakh steppe, southern China, Central
Asia, Iran, and the Indus Valley, are compared to testwhich, if either, of these hypotheses are supported by the
pattern of phenetic affinities possessed by Bronze Ageinhabitants of the Tarim Basin. Craniometric differencesbetween samples are compared with Mahalanobis gener-
alized distance (d2), and patterns of phenetic affinity are
assessed with two types of cluster analysis (the weightedpair average linkage method and the neighbor-joiningmethod), multidimensional scaling, and principal coordi-nates analysis. Results obtained by this analysis providelittle support for either the steppe hypothesis or the Bac-trian oasis hypothesis. Rather, the pattern of pheneticaffinities manifested by Bronze Age inhabitants of theTarim Basin suggests the presence of a population ofunknown origin within the Tarim Basin during the earlyBronze Age. After 1200 B.C., this population experiencedsignificant gene flow from highland populations of thePamirs and Ferghana Valley. These highland populationsmay include those who later became known as the Sakaand who may have served as middlemen facilitatingcontacts between East (Tarim Basin, China) and West
(Bactria, Uzbekistan) along what later became known asthe Great Silk Road. Am J Phys Anthropol 124:199222,2004. 2004 Wiley-Liss, Inc.
Schoolchildren in the United States are taughtthat the peoples of western Asia and Europe re-mained ignorant of the populations of East Asia formany centuries, but that this changed with the in-credible journey of Marco Polo. According to popularaccount, Marco Polo, a Venetian merchant, leftVenice along with his father and uncle in A.D. 1269and traveled east across Central Asia along the
route of what later came to be known as the GreatSilk Road, arriving at the court of Kublai Khan atBeijing in A.D. 1275. There they were received bythe great Khan and remained for the next 16 years,only to return to Venice with news of the wonders ofthe East in A.D. 1295 (Komroff, 1930, p. viixx). Yet,despite popular perception, the numerous inaccura-cies, apparent plagiarisms, and profound gaps ofobservation evident in The Travels of Marco Poloraise serious doubts as to whether Polo ever went toChina at all (Mallory and Mair, 2000, p. 71).
Those with more than a passing familiarity oftenidentify the date 132 B.C. as the beginning of con-
tacts between East and West along the Great SilkRoad (Barber, 1999, p. 19; Di Cosmo, 1996, p. 87;Mair, 1995, p. 302; Mallory and Mair, 2000, p. 56). Itwas at this time that the Chinese explorer ZhangQian embarked on a 13-year mission westward fromGansu, through Xinjiang, and across the FerghanaValley into Central Asia. Soon the Emperor Wudigathered an army of some 60,000 soldiers to secure
the Great Silk Road, so that between 114 and 108B.C. no less than 10 caravans a year were making
*Correspondence to: Brian E. Hemphill, Department of Sociologyand Anthropology, California State University at Bakersfield, Bakers-field, CA 93311-1099. E-mail: [email protected]
Received 25 October 2002; accepted 2 June 2003.
DOI 10.1002/ajpa.10354Published online 19 September 2003 in Wiley InterScience (www.
interscience.wiley.com).
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 124:199222 (2004)
2004 WILEY-LISS, INC.
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
2/24
the journey from China in the east to the FerghanaValley in the west.
Other scholars maintain that the beginning ofcontacts between East and West occurred even ear-lier, for silk appears in Europe by the sixth centuryB.C., throughout the northern Mediterranean basin
by the fifth century B.C, and perhaps as early as1000 B.C. in North Africa (Mair, 1995, p. 285). In thefifth century B.C., Herodotus mentioned transittrade occurring across great distances in CentralAsia along a route that stretched from the River Donin the Urals in the west to the Altai and the Mi-nusinsk Basin in the east (Chlenova, 1983).
Archaeological excavations at the site of Sapallitepe, located in the North Bactrian oasis of southernUzbekistan, during the 1960s and 1970s yielded theearliest evidence of silk outside of China and raisedthe possibility that contacts between East and Westalong the Great Silk Road may be far older thanpreviously thought (Askarov, 1973, p. 133134,
1974, 1977, 1981, 1988). No longer were contactsdated to 13th century A.D., nor to the second cen-tury B.C., nor even to the 10th century B.C. Rather,the presence of silk at Sapalli tepe raised the possi-bility that contacts along the Great Silk Road mayhave occurred near the end of the third and thebeginning of the second millennia B.C. (Hiebert,1994; Kohl, 1984). Yet, as provocative as this discov-ery was, few scholars outside the former SovietUnion knew of these discoveries (Kohl, 1981, 1992).
One of the major archaeological events of the pastdecade has been the proliferation of popular articles,books, and television documentaries devoted to thediscovery of prehistoric Bronze Age Caucasoid or
Europoid populations in the western Chinese Xin-jiang Uyghur Autonomous Region (Mair, 1995, p.281). Here along the oases of the Tarim Basin havebeen recovered some 300 mummies, many of whichhave been found along with their clothes in an ex-traordinary state of preservation, dating from ca.1800 B.C. until the Chinese conquest of the region inthe first centuries B.C. A far greater assemblage ofskeletal remains has been recovered from Bronzeand Iron Age cemeteries of the region (an assem-blage that should be numbered in the many hun-dreds, if not thousands), though only a few morethan 300 have been examined. Where it has beenpossible to identify a phenotypic pattern, the major-
ity of these individuals are identified as possessing astronger resemblance to a Western pattern (e.g.,fair hair, high-bridged noses, heavy beard) ratherthan the phenotypic pattern common to the Han ofChina (Barber, 1999, p. 19; Mallory and Mair, 2000,p. 16; Wang, 2001). Not surprisingly, such identifi-cations led many to ask (Mair, 1995, p. 289), Whowere the[se] corpses from the Tarim Basin? Wheredid they come from? And how did they get there?
Three lines of evidence have been offered to dem-onstrate that the Bronze Age inhabitants of Xinjiangwere neither long-term indigenous inhabitants ofthis region, nor ethnic Han Chinese immigrants
from the East. These lines of evidence include thetextiles worn by these individuals, the evidence ofIndo-European languages in Xinjiang, and previousbiological analyses of the human remains them-selves. The purpose of this paper is to comparecraniometric variation among Bronze Age inhabit-
ants of Xinjiang, western China, with Aeneolithicand Bronze Age samples from the Russo-Kazakhsteppe, south Central Asia, southern China, Iran,and the Indus Valley, in order to test which of thehypotheses best explain the origins and subsequentinteractions of the Bronze Age inhabitants of theTarim Basin.
Mair (1995, p. 295) considered the textiles wornand associated with the Xinjiang remains to behighly diagnostic, perhaps still more so than DNAanalysis, for identifying the origins and affiliationsof the Tarim Basin Bronze Age people. These tex-tiles encompass an array of items including stringskirts, fur-lined and fur-trimmed coats, long stock-
ings, and pants that Mair (1995, 1998), Mallory andMair (2000), and Barber (1999) claim are indicativeof a close relationship with Indo-European-speakingpastoralist nomads from the Russian steppe. Themost ancient Bronze Age remains found in Xinjiangderive from the site of Qawrighul (ca. 1800 B.C.),located along the Konchi River at the southeasternedge of the Tarim Basin (Fig. 1). These remains werefound clad in simple cloaks, mantles, and wraps inshades of natural brown or beige that lack any evi-dence of piping, sleeves, or trouser legs. Barber(1999, p. 71) maintained that these individuals notonly appear to have introduced weaving into theTarim Basin, but the mere presence of woolen tex-
tiles reveals that domestic sheep from the West hadbeen introduced into the Tarim Basin by the begin-ning of the second millennium B.C. (Mallory andMair, 2000, p. 219).
Some 500 years later, during the closing centuriesof the second millennium B.C., mummies recoveredfrom Zaghunluq (near Charchan), located along thesouthern margin of the Taklamakan Desert in thesouthern Tarim Basin, document the introduction ofan entire array of new techniques in clothing man-ufacture (Barber, 1999). Textiles from the site ofQizilchoqa (near Qumul/Hami), in the easternmostpart of the region, are marked by weaving of im-proved quality that includes twill as well as plain
weave. A detailed examination of a textile fragmentby Good (1995) yielded evidence of the same decora-tive technique as that found in Scottish tartanswhich, in turn, exhibited similarities to tartansfound at Hallstatt in Austria dated to the late sec-ond millennium B.C. (Mallory and Mair, 2000).
The second line of evidence is linguistic, and in-volves the discovery of written documents in theTarim Basin that attest to the presence of a series ofIndo-European languages collectively known as To-charian (Barber, 1999, p. 115; Jettmar, 1998, p. 216;Mallory, 1998, p. 189; Renfrew, 1988, p. 6366).These languages exhibit no close similarities to
200 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
3/24
Indo-Iranian (which is well-represented by a num-ber of languages in the Tarim Basin) or any othereastern (satem) Indo-European languages (otherthan loan words). Within the phylogeny of the Indo-European languages, Tocharian languages are ei-ther placed with the western (centum) languagesof Europe (Adams, 1984; Hamp, 1998), or are re-
garded as languages that split from the rest of theIndo-European languages at such an early date thatthey lack many of the isoglosses found betweenother Indo-European languages (Ringe et al., 1998).Mallory and Mair (2000, p. 240246) suggested thatthe separation of Tocharian speakers from otherIndo-European-speaking communities may have oc-curred as early as the fourth millennium B.C., andthey identified the Afanasievo culture (ca. 3500 2500 B.C.), a primarily pastoralist culture found inthe Altai and Minusinsk regions of the Eurasiansteppe, as a possible source for a Tocharian presencein the Tarim Basin (Mallory, 1989, p. 62, 263, 1995,
p. 380381, 1998, p. 189; see also Parpola, 1998).Wall paintings of Tocharian speakers depict theseindividuals as possessing red or blonde hair, longnoses, blue or green eyes, and wearing broadswordsinserted in scabbards hanging from their waists(Mair, 1995, p. 299).
The third line of evidence comes from analyses of
the biological features of the human remains them-selves. This evidence derives from the obviouslyWestern appearance of many of the mummifiedremains, analyses of ancient DNA, and craniometry.Francalacci (1995) obtained tissue samples from 11mummies, but only two of these samples were per-mitted out of China, and the DNA from one was toodamaged for analysis. Hence, at present, the geneticevidence for the history of the Bronze Age inhabit-ants of Xinjiang rests on results obtained from asingle individual (Mallory and Mair, 2000, p. 246247). The mtDNA of this mummy was identified byFrancalacci (1995) as belonging to haplogroup H,
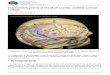
Fig. 1. Geographic location of craniometric samples. Sample abbreviations from Table 1. Xinjiang samples (QAW, ALW, and KRO)and Chinese sample from Hainan (HAI) are represented by asterisks; North Bactrian samples, by stars; Iranian samples, bypentagons; Turkmenian, Caucasus, and Tajik samples, by triangles; Indus Valley samples, by circles; and Russo-Kazakh samples, bysquares.
INHABITANTS OF XINJIANG 201
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
4/24
one of nine subtypes of mitochondrial lineageslargely associated with Europeans. Yet, while hap-logroup H is the most common marker of Europeanpopulations (40%), it is also found in 15% of individ-uals from the Near East. Hence, despite claims tothe contrary (see Cavalli-Sforza, 2000), the ancient
DNA evidence currently available offers little reso-lution concerning the precise origins of the BronzeAge inhabitants of Xinjiang.
Over the course of the last two decades, Han(1994a,b; see also Mair, 1998) conducted craniomet-ric analyses of some 302 adult Bronze Age inhabit-ants of Xinjiang. Han (1998, p. 568) concluded thatmetric variation among these individuals revealedthat the majority (89%) may be attributed to at leastthree branches of the Caucasoid type, while a mi-nority (11%) may be ascribed to two branches of theMongoloid type. Han (1994a, 1998, p. 566568)contended that the temporal and geographic pat-terns observed among these cranial types document
a three-stage settlement of Xinjiang during theBronze Age. According to Han (1998), the first waveof immigration involved colonization by a proto-Europoid type with Nordic characteristics that heattributed to Afanasievo and Andronovo populationswho migrated to Xinjiang and the Tarim Basin fromthe steppelands located to the north and northwest.Han (1998) maintained that a second wave of immi-gration, this time involving an Eastern Mediterra-nean type and possibly a Pamir-Fergana type,entered Xinjiang and the Tarim Basin from the westaround 500 B.C. The third wave of immigration in-volved a westward migration of an Eastern Mon-goloid type into the eastern regions of Xinjiang and
the Tarim Basin, most likely from the adjacent prov-ince of Gansu and points further east (see also An,1992b; Shui, 1993).
MODELS FOR XINJIANG POPULATION ORIGINS
Contemporary researchers are divided over themost likely explanation for the origins of the BronzeAge inhabitants of Xinjiang, and this division isreflected by the wide array of explicative accountsadvanced by archaeologists, biological anthropolo-gists, historical linguists, and others. Nevertheless,these explanations can be grouped into two generalmodels. These models may be designated the steppehypothesis and the Bactrian oasis hypothesis.
Proponents of the steppe hypothesis maintainthat the Tarim region experienced at least two pop-ulation influxes from the Russo-Kazakh steppe re-gion (Anthony, 1998; Han, 1998; Kuzmina, 1994,1998; Mallory, 1995; Mallory and Mair, 2000; Par-pola, 1998). They suggest that the initial wave ofimmigration into Xinjiang may have originatedamong members of the Afanasievo culture found inthe Altai and Minusinsk regions of the steppe northof the Tarim Basin. This attribution is based onnumerous similarities in material culture, includingbronze metallurgy, burial practices, and textiles,found between Afanasievo culture sites and the ear-
liest Bronze Age sites in Xinjiang, such as Qawri-ghul (Kuzmina, 1994, p. 241, 1998, p. 69; Mallory,1995; Mallory and Mair, 2000).
The Afanasievo culture itself is believed intrusiveto the eastern steppe and is held to share the closestsimilarities to the Yamnaya culture (35003000
B.C.; Mallory and Mair, 2000) found in the Pontic-Caspian region of the Russian steppe (Alexeev,1961; Anthony, 1998, p. 104; Chernykh, 1992;Gryaznov and Vadezkaya, 1968; Kuzmina, 1998;Mallory, 1989, p. 62, 1995, 1998, p. 189; Mallory andMair, 2000; Parpola, 1998; Posrednikov, 1992; butsee Shishlina and Hiebert, 1998, p. 222223). Asnoted by Mallory (1995) and Mallory and Mair(2000, p. 381382), an eastward emigration and sub-sequent isolation of Yamnaya-derived Afanasievopopulations in the eastern steppe provides a possibleexplanation for the appearance of Tocharian in theTarim Basin of Xinjiang. Hence, the apparent simi-larities between Tocharian and such western Indo-
European languages as Celtic and Italian are due toa common retention of proto-Indo-European archai-cisms (Mallory, 1989, p. 61), while the differencesbetween Tocharian and neighboring Indo-Iranianmight be explained by the peripheral position ofproto-Tocharian populations with respect to theemergence of Indo-Iranian languages. This asser-tion appeared to be supported by Hans (1998) con-trast of cranial and facial indices which indicatedthat the earliest individuals from Qawrighul (type Itombs; see below) were most similar to individualsfrom Afanasievo cultural contexts (see Mallory andMair, 2000, p. 240243).
Many proponents of the steppe hypothesis con-
tend that the immigration of Afanasievo populationsto the Tarim Basin was followed by a later influx ofpopulations derived from the Late Bronze Age An-dronovo culture complex (ca. 2100900 B.C.; Mal-lory and Mair, 2000) found to the west, northwest,and north in the Pamirs, the Ferghana Valley, Ka-zakhstan, and the Minusinsk/Altai region (Chenand Hiebert, 1995; Kuzmina, 1998; Mei and Shell,1998; Parpola, 1998). As with earlier Afanasievopopulations, those accompanied by artifacts as-signed to the Andronovo culture are believed to haveoriginated in the western Russian steppe. In thislatter case, ultimate stylistic origins of the artifactsare traced to the Sintashta culture (ca. 23001900
B.C.; Mallory and Mair, 2000) of the southeast Urals(Anthony, 1998; Anthony and Vinogradov, 1995, p.36; Kuzmina, 1994; Mallory, 1998). The eastwardexpansion of these likely Indo-Iranian-speaking peo-ples was facilitated by the development of the warchariot (Anthony, 1998, p. 94; Di Cosmo, 1996, p.9091; Mallory, 1989; Parpola, 1998) and is re-flected by the rapid appearance of such regionalvariants of the Andronovo culture designated asAlakul, Federovo, Tazabagyab, Beshkent, and Va-khsh (Gupta, 1979; Hiebert, 1994; Kohl, 1984, p.183184; Kuzmina, 1994; Masson, 1992b, p. 350351; Sulimirski, 1970, p. 261, 263; Zdanovich, 1988).
202 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
5/24
The initial appearance of Andronovo-derived popu-lations in the Tarim Basin is held to be signaled bythe introduction of new clothing styles, ceramicwares, and burial customs as well as objects of tinbronze, and objects associated with horses around1200 B.C. (Barber, 1999; Kuzmina, 1998, p. 73; Mei,
2000, p. 7275; Mei and Shell, 1998).The second model for the origin of the Bronze Ageinhabitants of Xinjiang is the Bactrian oasis hypoth-esis. Proponents of this model assert that settlementof this region of western China came not from no-madic pastoralists of the steppe, but from sedentary,agriculturally based populations of the Oxus civili-zation (the Bactrian-Margiana archaeological com-plex (BMAC); Hiebert, 1994) found west of Xinjiangin Uzbekistan (north Bactria), Afghanistan (southBactria), and Turkmenistan (Margiana) (Barber,1999; Chen and Hiebert, 1995, p. 287). Proponents ofthis model emphasize the environmental similari-ties between the desert basins of eastern (Xinjiang)
and western Central Asia (Bactria, Margiana) andmaintain that the sophisticated irrigation tech-niques developed in the oases of Margiana and Bac-tria permitted the colonization of the river deltasand oases surrounding the north, east, and southernmargins of the Taklamakan Desert (Barber, 1999;Chen and Hiebert, 1995).
Barber (1999) suggested that the archaeologicalrecord provides better evidence that the initial col-onizers of the Tarim Basin were agriculturalistsfrom Bactria than nomadic pastoralists from theRussian steppe. This evidence not only includes ir-rigation systems, evidence of western cultigens suchas wheat, and bones of sheep and goats, but also
evidence of carefully bundled bags containingEphe-dra sp. found accompanying many Bronze Age Xin-jiang burials. Use of ephedra is well-known in Oxuscivilization urban centers, where Hiebert (1994) andSarianidi (1987, 1990, 1993a,b, 1994; see alsoKussov, 1993; Meyer-Melikyan, 1998; Meyer-Me-likyan and Avetov, 1998) found evidence of special-ized areas known as white rooms where it is be-lieved a ritual drink, known as haoma in Iranianand soma in Indic, was consumed (but see Nyberg,1995, p. 400; Parpola, 1995, p. 371). Ephedra doesnot grow on the Russo-Kazakh steppe, nor is it as-sociated with either Afanasievo or Andronovo cul-tures (Barber, 1999, p. 165; but see Parpola, 1998, p.
126127).Barber (1999) suggested that the Bronze Age set-
tlement of the Tarim Basin was a two-step processin which initial immigration came from the oases ofBactria and Margiana, and is represented by re-mains found at such southeastern sites as Qawri-ghul. This was followed by a second wave of immi-gration soon after 1200 B.C. This time, immigrantscame from Andronovo populations located to thenorthwest, and participants in this wave of immi-gration may be associated with the remains found atnorthern Tarim Basin sites such as Alwighul, Hami,Turfan, and Kucha. Barber (1999) claimed that ev-
idence of these two separate waves of immigration isprovided not only by dramatic differences in textilemanufacture, but also by textual evidence from thefirst centuries A.D. which documents that inhabit-ants of the southern Tarim oases spoke Iranian lan-guages (such as Saka and Sogdian), while those
inhabiting the northern oases spoke Tocharian.Measurements of the neurocranium and facialskeleton have been used for many years to providean assessment of the degree of biological relatednessamong samples of past and living populations. Al-though it is clear that these measurements actuallyprovide assessment of an unknown combination ofenvironmental and hereditary factors (Cavalli-Sforza and Bodmer, 1971), and may be affected bymasticatory mechanics (Carlson and Van Gerven,1977; Van Gerven, 1982) and environmental varia-tion (Beals, 1972; Guglielmino-Matessi et al., 1979),twin studies (Clark, 1956; Lundstrom, 1954; Nakataet al., 1974a; Orczykowska-Swiatkowska and Leb-
ioda, 1975; Saunders et al., 1980), familial studies(Devor, 1987; Howells, 1966; Nakata et al., 1974b;Susanne, 1975, 1977), and worldwide comparisonsof craniometric variation revealed a moderate de-gree of genetic control (Susanne, 1975, 1977), anddemonstrated the utility of such variables for recon-structing patterns of biological interactions amongpopulations (Howells, 1973, 1989). Since all of thesamples included in this study derive from eithersedentary, agricultural communities or pastoralistpopulations who received regular supplies of agri-cultural produce, and from sites that differ little inlatitude, a comparison of craniometric variationshould suffer no systemic biases due to differences in
masticatory stresses or natural selection for dramat-ically different environments (Hemphill, 1998,1999).
MATERIALS AND METHODS
Materials
Analyzed Bronze Age skeletal samples from Xin-jiang are few and are underrepresented in the liter-ature. Although many individuals included in thecurrent study were the subject of craniometric com-parisons by Han (1990, 1994b, 1998, 2001) and Mal-lory and Mair (2000, p. 236244), these comparisonsare limited to contrasts of cranial and facial indices
that provide no assessment of covariation amongcranial indicators or of the significance of pheneticseparation between samples. Further, results ob-tained from these contrasts are interpreted within atypologically grounded ethnogenetic paradigm thatidentifies human variation, even in individual cem-eteries, as attributable to the presence of multiplephysical types and subsequent interbreedingamong them (Han, 1994b, 1998; Mair, 1995, p. 291292; Mallory and Mair, 2000, p. 235). Hence, reifi-cation of such fixed physical types encourages astatic perspective of human populations that fails toaccommodate the known evolutionary forces of nat-
INHABITANTS OF XINJIANG 203
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
6/24
ural selection, mutation, gene flow, and genetic drift.It is these processes that result in the inherent dy-namism in the genetic foundation of all populations,as emphasized in modern population genetics (Fal-coner, 1981; Hartl and Clark, 1997; Hedrick, 2000).
Through the efforts of Mallory (1995), measure-
ments made in accordance with standards estab-lished by Martin (1928) by Han (1998) on craniarecovered from three sites from the Tarim Basin ofXinjiang were made available to B.E.H. for furtherstatistical analyses and interpretation. These sitesinclude Qawrighul, Alwighul, and Kroran (Fig. 1).
Discovered in 1979 and excavated by Wang (1987,1996), Qawrighul represents the most ancientBronze Age cemetery in Xinjiang (Debaine-Francfort, 1988, p. 1516; Han, 1994a; Jettmar,1992; Kuchera, 1988; Wang, 1987, 1996). A series offive radiocarbon dates places this cemetery between23001430 B.C. (An, 1998; Chen and Hiebert, 1995).The cemetery is located on the bank of the Konchi
River, 70 km west of ancient Lake Lopnur along theeastern edge of the Taklamakan Desert (Kuzmina,1998; Mallory and Mair, 2000, p. 137). Excavation ofthe cemetery resulted in the recovery of 42 burials,each containing a single individual, from two differ-ent types of tombs (Wang, 1982, 1983).
The first, designated as Qawrighul type I tombs,account for 36 of the burials. All are shaft pit graves,and a minority of these graves feature wooden polesplaced at east and west ends. The actual burialchamber was lined with small wooden boards, andthe top of the chamber was sealed with animalskins, carpets, or a basket-shaped cover. The bodieswere placed in a supine, extended position, with
their heads to the east and their feet to the west.Qawrighul type II tombs account for six burials, andthey appear stratigraphically later than type Itombs. Type II tombs are identical to type I tombswith respect to the use of small wooden boards toline the burial pit and placement of the body, butdiffer by featuring an elaborate arrangement ofwooden poles embedded in the ground surface. Themost elaborate of these type II tombs featured sevenconcentric rings of wooden stakes that radiate out-ward in what some (Wang, 1983, 1984) interpretedas a solar pattern from the center of the burialchamber, to encompass an area 5060 m in diame-ter.
Individuals interred in the Qawrighul cemeterywore no clothing apart from leather shoes and cloaksmade of plain woven textiles fastened at the frontwith bone pins. Felt hats were placed on the head,and in several instances, small bags containingtwigs of Ephedra sp. were found on their chests(Barber, 1999; Chen and Hiebert, 1995, p. 253). Noceramics accompany any of the burials, but one boneand five wooden anthropomorphic figurines, somestill wearing fragments of textiles, were recovered. Afew jade beads and fragments of either copper orbronze represent the extent of additional burial ac-coutrements (Wang, 1982, 1983, 1984). All 18 adult
crania recovered from this site (11 male, 7 female)were identified by Han (1986a, 1994a, 1998) asproto-European and possessing closest affinities tocrania recovered from southern Siberia, Kazakh-stan, and the Volga River region of southern Russia(Han, 1998, p. 559560; Mair, 1995, p. 290; Mallory
and Mair, 2000, p. 241). Han (2001, p. 234) sug-gested that the earliest (type I) burials were mostclosely related to the Afanasievo type, while laterburials bore greater similarity to Andronovo popu-lations.
The cemetery of Alwighul is located near the Or-dos grasslands along the southern slopes of the TianShan mountains (Han, 1998; Ma and Wang, 1994, p.213). Featuring a series of radiocarbon dates thatcluster between 800200 B.C., the Alwighul ceme-tery dates to the Late Bronze Age, a period that ismarked by an increase in the frequency of bronzeobjects and, in some regions of Xinjiang, the initialappearance of large bronze objects (An, 1998, p. 58;
Ma and Wang, 1994, p. 213).Three major inhumation types were observed atAlwighul. The first two feature distinctive burialchambers in which the perimeter walls are linedwith pebbles. The early pebble graves (type I) fea-ture multiple interments of at least 1020 individ-uals piled atop one another in a supine position,with the head to the west and the feet to the east.Late pebble graves (type II) are similar in construc-tion to those of type I, but feature an additionalwooden bench supported by four pillars and containthe remains of only 12 individuals. Type III gravesappear as heaps of stones at ground level beneathwhich there is a large vertical pit that leads to a
wooden coffin-chamber.In total, 58 adult crania (33 males, 25 females)
were recovered from Alwighul. All of these remainswere recovered from a single type I mass intermentgrave and were associated with a wide array ofburial goods, including hand-made gray-red ceramicvessels decorated with triangular, net, whorl andpine-needle motifs painted in light black; a consid-erable number of bronze plates and knives; manystrings of beads made from bone, shell, agate, andjade; and earrings of both bronze and gold (Ma andWang, 1994, p. 213215). The craniometric analysisby Han (1994b, 1998, p. 560561, 2001, p. 231232)of these remains revealed the presence of at least
three different racial types, including two differentforms of Caucasoids (Pamir-Ferghana type andEastern Mediterranean type) and a single form ofMongoloid. Han (1990, 1998) suggested that theapparent lack of conformity of some of the crania tothese racial types indicated some degree of mix-ture between the two Caucasoid types as well asbetween Caucasoids and Mongoloids.
The cemetery of Kroran is located in one of thesoutheastern group of oases on the southern bank ofthe Konchi River near the outskirts of Loulan, thecapital of the Shan-Shan state during the first cen-tury B.C. (Ma and Wang, 1994, p. 211). There is
204 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
7/24
considerable controversy over the correct dates forthe human remains from this cemetery. According tosome researchers, the six adult crania recoveredfrom this cemetery derive from the Western Hanperiod (202 B.C.A.D. 220; Han, 1998; Mair, 1995),but recent Chinese excavations suggest that theseremains are associated with artifacts that span theperiod between the seventh to first centuries B.C.(Ma and Wang, 1994, p. 211). Nevertheless, a pair ofradiocarbon dates obtained from the cemetery sug-gest that the Western Han period is most likelycorrect (Mallory and Mair, 2000, p. 335).
Individuals recovered from the cemetery at Kro-ran were interred in graves featuring a chamber of
wooden planks within a shallow pit. The body wasplaced in an extended position, and wrapped in awoolen cloth. One extremely well-preserved individ-ual was buried wearing hide boots and a peakedbrown felt hat with bird feathers. In this case, thewoolen cloth was gathered into a pouch on the upperchest and filled with fragments ofEphedra sp. (Maand Wang, 1994, p. 211212). The graves also con-tained wooden and stone figurines with long, roundfaces, but most funerary objects represent articles ofeveryday use and decorative ornaments. In earliertombs, there is no pottery, and utensils are made ofwoven grass, wood, bone, or horn. Ornaments in-
clude beads made of bone, amber, agate, or jade, andwere usually found encircling the neck or ankles.Groups of bone tubes about 10 cm long were some-times linked together and worn around the waist. Inaddition to these locally produced items, many arti-facts such as brocades, rough silk, silk floss, bronzemirrors, lacquerware, andwuzhucoins typical of theHan Dynasty of the middle-lower Yellow River werealso recovered.
Of the six adult crania recovered, only one, a fe-male, was identified by Han (1986b, 1994a, 1998, p.562563) as Mongoloid. The remaining five indi-viduals were identified by Han as possessing East-ern Mediterranean characteristics most similar tothose found among sixth century B.C. Saka of thesouthern Pamirs (Ma and Wang, 1994, p. 212; Mair,1995, p. 292).
Cranial series used to provide a comparative foun-dation for the Xinjiang remains encompass 22 sam-ples, numbering 1271 individuals (665 males, 606females) from the Russo-Kazakh steppe, southernChina, Central Asia, Iran, and Indus Valley. To-gether, all 25 skeletal samples span a timeframefrom 3500 B.C. to the present. Abbreviations, sam-ple sizes, sources, and sample locations for all cra-nial samples are provided in Table 1 and Figure 1.
TABLE 1. Samples considered in study
Code
Maximum samplesize
Site/region Period Dates ReferenceMales Females
ADM 22 11 Andronovo/Minusinsk Late Bronze Age 2100900 B.C. Alexeev (1961)AFA 17 7 Afanasievo/Altai Early Bronze Age 35002500 B.C. Alexeev (1961)
AFM 18 11 Afanasievo/Minusinsk Early Bronze Age 35002500 B.C. Alexeev (1961)AND 25 31 Andronovo/Kazakhstan Late Bronze Age 2100900 B.C. Alekseev (1967)ALT 40 42 Altyn depe/Turkmenistan Namazga V 25002200 B.C. Kiiatkina (1967);
Hemphill (1999)ALW 33 25 Alwighul/Xinjiang, China Late Bronze Age 650200 B.C. Han (1998)CEMH 10 18 Harappa/Indus Valley Late Harappan 19001600 B.C. Gupta et al. (1962)DJR 17 33 Djarkutan/North Bactria Djarkutan phase 20001800 B.C. Hemphill (1998)GKS 38 32 Geoksyur/Turkmenistan Namazga III 35003000 B.C. Kiiatkina (1987);
Hemphill (1999)HAI 45 38 Hainan/South China Living Living Howells (1989)HAR 23 41 Harappa/Indus Valley Mature Harappan 25002000 B.C. Gupta et al. (1962);
Hemphill et al. (1991)KAM 133 118 Karasuk/Minusinsk Late Bronze Age Rykusina (1976)KAR 14 13 Kara depe/Turkmenistan Namazga III 35003000 B.C. Ginzberg and Trofimova
(1972)KOK 14 10 Kokcha III/Turkmenistan Late Bronze Age 18001500 B.C. Trofimova (1961)KRO 4 2 Kroran/Xinjiang, China Bronze/Iron Age 202 B.C.A.D. 150 Han (1998)KUZ 13 14 Djarkutan/North Bactria Kuzali Phase 18001650 B.C. Hemphill (1998)
KUZ 13 14 Djarkutan/North Bactria Kuzali phase 18001650 B.C. Hemphill (1998)MOL 18 28 Djarkutan/North Bactria Molali phase 16501500 B.C. Hemphill (1998)QAW 11 7 Qawrighul/Xinjiang, China Early Bronze Age 1800 B.C. Han (1998)SAMB 14 13 Samtavro/Caucasus Late Bronze Age 1400800 B.C. Abduselisvili (1954)SAP 13 28 Sapalli tepe/North Bactria Sapalli phase 22002000 B.C. Hemphill (1998)SHS 45 43 Shahr-i Sokhta/Eastern Iran SHS I, II, III 30002200 B.C. Pardini and Sarvari-
Negahban (1976)Pardini (1977, 19791980)
TH2 9 7 Tepe Hissar/Northern Iran Tepe Hissar II 33002500 B.C. Krogman (1940)TH3 102 36 Tepe Hissar/Northern Iran Tepe Hissar III 25001700 B.C. Krogman (1940)TMG 9 11 Timargarha/Indus Valley Late Bronze/Early
Iron Age1400800 B.C. Bernhard (1967)
TMM 26 21 Tigrovaja-Makoni Mor/ Tajikistan
Molali phase 16501500 B.C. Kiiatkina (1976)
INHABITANTS OF XINJIANG 205
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
8/24
Eight cranial variables (two for the neurocranium,and six for the facial skeleton) of those defined byMartin (1928) provide the metrical basis for thecurrent study (Table 2). While this small battery ofmeasurements is far from representing the ideal
array of variables for capturing the morphologicalcomplexity of the human cranium, increases in thenumber of variables do not automatically result ingreater insight into the patterning of phenetic dis-tances (Kowalski, 1972, p. 121; Oxnard, 1973, p. 39).In addition, when employing remains recoveredfrom archaeological contexts, the often fragmentarynature of these remains leads to a concomitant de-crease in sample size for every increase in variablesconsidered. Quite simply, these eight variables rep-resent the best combination of those measurementsfor which data were available for all of the samplescompared and those adequately represented, due todifferential preservation relative to other measure-
ments, within each of these samples.When utilizing data collected by other workers,
the degree of interobserver differences in assess-ment of these variables represents an importantsource of potential error that can compromise mean-ingful results. In this study, the degree of interob-server error between the authors and describers ofcomparative cranial series could be assessed forTepe Hissar (Krogman, 1940), Harappa (CemeteryR37), Cemetery H at Harappa (Gupta et al., 1962),Altyn depe, and Geoksyur (Ginzberg and Trofimova,1972; Trofimova, 1961; Kiiatkina, 1976, 1977, 1987).Repeated-measures analysis of variance (Hemphill,1998, 1999; Hemphill et al., 1991) indicated no sig-
nificant measurement differences between differentobservers. Although the degree of interobserver er-ror could not be directly assessed between the au-thors and samples obtained from Alekseev andGochman (1983), these researchers incorporatedmeasurements taken by Trofimova (1961) and Kiiat-kina (1976, 1977, 1987) with those of Alexseev(1961, 1967) and Abduselisvili (1954, 1960, 1966)and found no significant differences. Logically, then,there should be no significant differences betweenmeasurements taken by the author and those ob-tained by Alexseev (1961, 1967) and Abduselisvili(1954, 1960, 1966) as well. Interobserver error could
not be assessed for published measurements for in-dividuals recovered from Shahr-i Sokhta, Timar-garha, or Hainan (southern China).
Methods
The covariance matrix for each sample was obtained
for males and females pooled together with listwisedeletion. Although pairwise deletion permits greatereffective sample sizes within each sample, listwise de-letion was used to avoid systematic biases caused byoverrepresentation and underrepresentation of indi-vidual variables (Wilkinson, 1990). A pooled covari-ance matrix was obtained for all samples for whichindividual data were available and bias-adjusted toaccommodate differences in sample size. Hence, thosesamples represented by group-level data only (An-dronovo-Minusinsk (ADM), Afanasievo-Altai (AFA),Afanasievo-Minusinsk (AFM), Andonovo-Kazakhstan(AND), Karasuk-Minusinsk (KAM), Kara depe (KAR),Kokcha III (KOK), Samtavro (SAMB), and Tigrovaja
Balka-Makoni Mor (TMM)) were not used to constructthe pooled covariance matrix. Variable averages werecalculated for both males and females. Sex-standard-ized group values for each variable were obtained bytaking the average of male and female mean values foreach sample (Table 3). The bias-adjusted pooled co-variance matrix and sex-standardized group valueswere used to obtain Mahalanobis generalized dis-tances (d2) between each pair of samples. The diagonalmatrix of Mahalanobis d2 values is provided in Table4. The significance of pairwise d2 distance contrasts forthose samples in which individual data were availablewere assessed by means of F-tests, conducted accord-ing to the method of Konigsberg et al. (1993).
The diagonal matrix of Mahalanobis d2
values wasused as input for cluster analyses. Different associ-ating algorithms were used to provide two perspec-tives on the patterning of intersample phenetic af-finities. These associating algorithms include theweighted pair average linkage method (WPGMA)(Sneath and Sokol, 1973) and the neighbor-joiningmethod (Felsenstein, 1989; Saitou and Nei, 1987).The cophenetic correlation coefficient, rcs (Sneathand Sokol, 1973), was computed with the NTSYS-pcstatistical package to measure the degree of corre-spondence between the obtained phenogram fromWPGMA cluster analysis and the original resem-blance matrix.
The diagonal matrix of Mahalanobis d2
values wasused as input for nonmetric multidimensional scal-ing, to provide a third perspective on the patterningof intersample affinities. The coefficient of alienationof Guttman (1968) was used to calculate distancesbetween individual points. The goodness of fit ob-tained by multidimensional scaling was assessedthrough calculation of the degree of stress through100 iterations. Multidimensional scaling was accom-plished with the SYSTAT statistical package(Wilkinson, 1990). Results obtained were ordinatedin three-dimensional space, and a minimum span-ning tree (Hartigan, 1975) was imposed on the array
TABLE 2. Craniometric variables used to generate mahalanobisgeneralized distances between samples
Variable1
NeurocraniumMaximum cranial length (GOL) 1Maximum cranial breadth (BEB) 8
Facial skeleton
Upper facial height (NPH) 48Nasal height (NH) 55Nasal breadth (NB) 54Orbital height (OH) 52Orbital breadth (OB) 51Bizygomatric breadth (BZB) 45
1 Numbers of variables as defined by Martin (1928)
206 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
9/24
TABLE 3. Mean values of craniometric variables1
GOL BEB NPH NH NB OH OB BZB
Male sampleADM 186.0 145.0 67.8 50.2 25.8 31.7 44.4 140.7AFA 191.7 142.4 71.7 53.1 27.1 32.3 43.7 141.6AFM 192.1 144.1 71.8 52.1 26.1 32.9 44.9 138.4ALT 189.5 135.9 70.7 51.6 25.2 32.4 40.7 129.1
ALW 184.2 141.9 68.7 52.2 25.0 33.1 39.0 130.8AND 186.4 140.4 69.2 51.5 25.4 32.9 42.1 131.8CEMH 188.2 141.3 67.9 50.8 26.3 32.9 41.3 134.8DJR 186.9 134.7 69.9 50.7 24.8 30.9 37.5 131.3GKS 190.1 134.5 71.1 52.0 25.5 33.0 40.1 127.6HAI 176.4 138.4 69.7 52.4 27.3 33.6 38.7 134.0HAR 187.3 134.5 69.2 51.4 26.5 33.2 41.4 131.5KAM 183.0 147.4 73.4 51.6 25.8 33.7 44.1 139.7KAR 194.8 134.9 72.6 51.2 26.6 31.8 42.5 129.9KOK 186.1 138.1 68.4 51.5 23.5 30.9 43.2 133.4KRO 187.9 139.1 74.4 53.6 24.6 34.1 38.6 131.0KUZ 190.9 138.9 68.7 49.6 26.4 30.9 39.9 134.0MOL 185.6 138.1 69.4 51.5 25.1 31.8 38.3 126.6QAW 183.0 137.9 66.5 50.9 26.2 31.6 40.5 136.2SAMB 189.3 137.1 76.7 53.8 23.8 35.0 42.0 128.3SAP 183.5 134.9 70.2 51.3 24.2 32.7 37.7 129.1SHS 185.8 136.4 70.2 50.6 25.7 31.8 42.1 129.4TH2 188.8 132.0 70.3 50.4 25.1 31.6 41.0 125.3TH3 188.4 134.1 69.8 50.6 25.4 32.1 41.2 127.3
TMG 190.2 132.0 70.3 50.0 22.9 33.3 41.5 133.0TMM 188.4 136.9 71.8 51.6 24.7 31.2 42.4 131.8Female sampleADM 175.9 140.6 67.5 48.9 23.9 33.0 42.5 127.4AFA 182.6 138.2 64.8 47.8 25.4 31.1 46.5 129.8AFM 180.4 135.8 67.4 49.5 25.6 33.1 44.3 131.8ALT 181.4 135.1 67.3 49.6 24.2 32.7 38.9 121.4ALW 173.9 135.3 64.9 49.4 24.2 32.0 37.5 124.4AND 177.6 136.0 67.3 48.4 24.6 32.2 41.8 128.2CEMH 179.2 132.4 62.7 46.0 24.4 33.3 39.9 119.5DJR 184.7 134.0 69.5 50.2 25.6 33.0 38.5 123.9GKS 185.8 132.9 69.8 50.9 25.2 32.9 39.0 123.4HAI 170.6 135.0 65.4 49.3 26.0 32.8 37.6 125.6HAR 180.9 132.1 66.2 48.3 24.2 34.1 40.6 123.9KAM 173.2 143.4 68.0 48.4 24.6 32.9 42.1 131.8KAR 183.0 132.1 67.5 47.9 24.7 32.0 41.6 123.8KOK 177.6 136.4 66.2 49.4 23.8 31.8 41.2 128.5KRO 181.0 133.5 69.0 50.6 24.1 33.2 36.8 129.5KUZ 179.3 132.6 65.1 46.8 23.6 30.7 36.3 122.4
MOL 183.5 134.2 70.6 49.7 25.0 32.6 38.8 126.5QAW 178.1 128.8 62.6 47.4 24.5 32.3 38.9 125.0SAMB 180.1 136.5 69.5 48.4 23.4 32.1 39.3 122.5SAP 181.5 134.1 67.5 49.2 24.8 33.0 37.2 124.4SHS 179.1 133.3 67.6 50.0 24.5 31.9 40.7 122.7TH2 178.3 132.1 67.6 48.3 23.7 33.6 38.7 118.7TH3 179.4 131.8 66.1 48.3 23.9 31.7 39.6 120.2TMG 180.2 130.9 66.6 48.1 22.9 33.1 40.0 122.3TMM 179.8 133.2 69.2 49.2 23.7 31.9 40.9 124.5Sex-standardized sampleADM 181.0 142.8 67.7 49.6 24.9 32.4 43.5 134.1AFA 187.3 140.3 68.3 50.5 26.3 31.7 45.1 135.7AFM 186.3 140.0 69.6 50.8 25.9 33.0 44.6 135.1ALT 185.5 135.5 69.0 50.6 24.7 32.6 39.8 125.3AND 182.0 138.2 68.3 50.0 25.0 32.2 42.6 132.4ALW 179.0 138.6 66.8 50.8 24.6 32.6 38.3 127.6CEMH 183.7 136.8 65.3 48.4 25.3 33.1 40.6 127.1DJR 185.8 134.3 69.7 50.5 25.2 32.0 38.0 127.6GKS 190.1 134.5 71.1 52.0 25.5 33.0 40.1 127.6
HAI 173.5 136.7 67.5 50.8 26.7 33.2 38.1 129.8HAR 184.1 133.3 67.7 49.9 25.4 33.6 41.0 127.7KAM 178.1 145.4 70.7 50.0 25.2 33.3 43.1 135.8KAR 188.9 133.5 70.1 49.6 25.7 31.9 42.1 126.9KOK 181.9 137.3 67.3 50.5 23.7 31.4 42.2 131.0KRO 184.4 136.3 71.7 52.1 24.4 33.7 37.7 130.2KUZ 185.1 135.7 66.9 48.2 25.0 30.8 38.1 128.2MOL 184.5 136.1 70.0 50.6 25.1 32.2 38.6 126.5QAW 180.5 133.3 64.5 49.1 25.3 32.0 39.7 130.6SAMB 184.7 136.8 73.1 51.1 23.6 33.6 40.7 125.4SAP 182.5 134.5 68.8 50.2 24.5 32.9 37.5 126.7SHS 182.5 134.8 68.9 50.3 25.1 31.8 41.4 126.0TH2 183.5 132.1 69.0 49.4 24.4 32.6 39.9 122.0TH3 183.9 133.0 69.9 49.4 24.7 31.9 40.4 123.8TMG 185.2 131.5 68.4 49.1 22.9 33.2 40.8 127.7TMM 184.1 135.1 70.5 50.4 24.2 31.6 41.7 128.2
1 Abbreviations for craniometric variables are from Table 2. Abbreviations for samples are from Table 1.
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
10/24
of data points to ease interpretation of the pattern-ing of intersample associations.
Principal coordinates analysis was used to providea fourth perspective on intersample craniometricvariation (Hair et al., 1971). The symmetric matrixof Mahalanobis d2 values was double-centered prior
to entry into NTSYS-pc statistical software (Rohlf,2000). The first three principal coordinate axes wereretained, group scores were calculated along theseaxes, and ordinated into three-dimensional space.As with results from multidimensional scaling, aminimum spanning tree was imposed on the array ofprincipal coordinate scores to ease interpretation ofintersample associations. The cophenetic correlationcoefficient was computed to assess the goodness of fitof the obtained eigenvectors with the matrix of Ma-halanobis d2 values. This latter step is especiallyimportant, because the cophenetic correlation coef-ficient provides more information on the patterningof relative phenetic distances among samples than
the absolute distance (as indicated by the percentageof total variation explained by the first three eigen-vectors) (Rohlf, 1972, 2000), and it is the patterningof these relative distances that is most useful forunderstanding processes of past population interac-tions.
As a final step in assessment of the nature ofintersample craniometric variation, spatial distanceand temporal distance matrices were computedamong all sample pairs. Congruence between theMahalanobis d2 matrix and these latter two matri-ces was assessed by means of the Mantel test (Man-tel, 1967) and Mantel correlation coefficient (Smouseet al., 1986). These procedures provide a test to
determine if differences between samples may sim-ply be a product of geographical propinquity or dif-ferences in antiquity. The significance of these asso-ciations was obtained through 1,000 permutationsat random by rows and columns.
RESULTS
The bias-adjusted matrix of Mahalanobis d2 val-ues was calculated according to the procedures out-lined above (Table 4). F-tests for those 16 samples inwhich individual data are available (Table 5) revealthat the majority of d2 values between samples aresignificant (98/120; 81.7%). Of the 98 pairwise con-trasts exhibiting a significant difference, 7 (7.1%)are significant at the 0.05 level, while 91 (92.9%) aresignificant at the 0.01 level.
WPGMA cluster analysis
The dendrogram obtained by means of theWPGMA associating algorithm (Fig. 2) indicatesthat the Hainan sample (HAI) from south Chinarepresents the most divergent of all samples consid-ered. The major division among remaining samplesoccurs between steppe samples (except SAMB andTMM) and all other samples. Bactrian samples aresegregated from samples obtained from Iran, Turk-
TABLE4.
Matrix
ofmahalanobisd2
generalizeddistances1
ADM
AFA
AFM
ALT
ALW
AND
CEMH
DJP
GKS
HAI
HAR
KAM
KAR
KOK
KRO
KUZ
MOL
QAW
SAMB
SAP
SHS
TH2
TH3
TMGTMM
ADM
0.0
AFA
2.146
0.0
AFM
1.423
0.651
0.0
ALT
7.482
8.216
6.208
0.0
ALW
7.237
11.187
8.787
2.879
0.0
AND
1.094
1.900
0.953
3.870
4.961
0.0
CEMH
5.033
5.942
4.134
2.006
3.396
2.864
0.0
DJR
11.681
12.879
10.467
1.671
3.169
6.790
4.367
0.0
GKS
10.844
9.969
7.823
0.969
4.836
6.037
3.465
1.429
0.0
HAI
10.554
14.863
11.760
7.800
3.043
7.931
6.800
6.824
9.582
0.0
HAR
8.735
8.921
6.318
3.114
5.479
4.355
2.029
3.727
3.435
6.989
0.0
KAM
1.752
6.396
4.352
10.608
9.040
3.695
8.526
14.107
14.601
9.924
12.042
0.0
KAR
8.138
5.592
4.677
2.526
9.317
4.242
4.002
5.058
2.576
14.058
4.187
12.367
0.0
KOK
2.481
3.160
2.804
4.381
5.416
1.076
4.722
7.647
6.834
10.962
6.375
6.724
6.035
0.0
KRO
13.039
16.850
12.914
3.878
2.903
8.714
6.607
1.593
3.804
11.857
5.530
13.567
9.922
9.700
0.0
KUZ
9.531
10.922
9.408
2.519
3.537
5.703
3.748
0.952
3.154
7.896
4.176
12.471
5.182
6.472
3.445
0.0
MOL
9.209
10.987
8.567
0.759
2.327
5.304
3.399
0.428
1.296
6.218
3.979
11.105
4.189
6.280
1.954
1.3
42
0.0
QAW
6.914
7.599
6.263
4.270
3.055
3.435
2.581
4.154
5.180
4.693
2.069
10.941
7.091
4.269
5.783
3.5
58
4.544
0.0
SAMB
8.160
10.378
7.277
2.009
6.324
5.297
5.477
4.431
3.313
5.297
6.149
9.208
3.535
2.158
9.440
5.8
38
2.676
9.440
0.0
SAP
11.583
14.700
11.396
2.298
1.963
7.188
4.240
0.669
2.731
4.981
3.544
13.146
7.571
8.194
0.597
1.8
12
0.962
3.814
4.895
0.0
SHS
4.751
4.366
3.326
1.296
4.932
1.865
2.815
4.432
2.866
8.819
3.893
8.204
1.521
2.439
7.851
4.7
00
2.952
4.579
2.854
5.733
0.0
TH2
9.455
9.497
7.309
0.726
5.768
5.137
3.163
2.963
1.617
10.480
3.077
12.896
1.573
5.886
6.162
4.0
35
2.019
6.049
1.873
4.016
1.427
0.0
TH3
8.245
6.399
7.828
0.608
6.390
4.106
2.410
3.045
1.656
12.592
2.941
14.296
1.152
2.811
10.382
3.4
26
2.122
4.669
5.995
4.387
0.586
0.38
4
0.0
TMG
8.486
9.195
6.706
3.044
6.898
4.411
4.033
4.462
3.696
12.528
2.211
12.344
4.028
4.318
5.786
4.6
66
4.459
4.688
3.790
4.593
3.940
2.82
8
5.276
0.0
TMM
4.901
4.800
3.713
2.106
6.273
2.000
4.564
4.608
3.535
11.403
4.880
8.076
1.675
6.790
7.567
4.5
65
3.367
5.782
2.403
6.126
0.754
2.12
1
0.689
2.863
0.0
1
Abbreviationsforsamplesarefrom
Table1.
208 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
11/24
menistan, and the Indus Valley. The two later sam-ples from Xinjiang (ALW and KRO) are associated
with the Bactrian samples. Kroran (KRO) features avery close affinity with the earliest of the Bactriansamples (SAP), while Alwighul (ALW) joins the laterBactrian samples (DJR, KUZ, and MOL) at a moredistant remove. Indus Valley samples are identifiedas sharing slightly closer affinity to samples fromIran and Turkmenistan than to Bactrian samples.Affinities among Indus Valley samples are ratherdiffuse. In fact, the early sample from westernChina, Qawrighul (QAW), is identified as possessingcloser affinities to the two samples from Harappa(HAR and CEMH) than exhibited by the third IndusValley sample, Timargarha (TMG). The remaining
samples form a loose cluster composed of sedentary
agricultural groups from Iran (TH2, TH3, and SHS)and Turkmenistan (GKS, ALT, and KAR), as well assteppe samples from the Caucasus (SAMB) and Ta-jikistan (TMM). Affinities are closest between thetwo northern Iranian samples from Tepe Hissar(TH2 and TH3), followed by the latest sample fromTurkmenistan (ALT). The eastern Iranian sample(SHS) and the steppe Bronze Age sample from Ta-jikistan (TMM) exhibit close affinities to one an-other, and moderate affinities to the two samplesfrom Tepe Hissar (TH2 and TH3) and Altyn depe(ALT). The two earlier samples from Turkmenistan(KAR and GKS) join these samples at a more distantremove. The steppe Bronze Age sample from the
Caucasus (SAMB) exhibits a peripheral associationwith these Iranian, Turkmenian, and Tajik samples.
Neighbor-joining cluster analysis
Neighbor-joining cluster analysis (Fig. 3) providesa different representation of the distance matrixthan that provided by WPGMA cluster analysis, be-cause it is an unrooted tree whose branches havedifferent lengths. Long branch lengths may be inter-preted as an indicator of a large degree of morpho-logical separation, while short branch lengths areindicative of a small degree of morphological sepa-ration between samples.
The neighbor-joining tree provides an array of
intersample associations that are largely in agree-ment with those depicted by WPGMA (Fig. 2). Onceagain, the south China sample from Hainan (HAI) isidentified as the most divergent of all samples con-sidered. All three western Chinese samples exhibitclosest affinities to samples from Bactria. The twolater western Chinese samples, Kroran (KRO) andAlwighul (ALW), feature closest affinities with theearliest of the Bactrian samples, Sapalli (SAP),while the earliest western Chinese sample, Qawri-ghul (QAW), is identified as possessing closer affin-ities to later Bactrian samples (DJR, KUZ, andMOL).
TABLE 5. F-tests and probability values of pairwise mahalanobis d2 generalized distances1
ALT ALW CEMH DJR GKS HAI HAR KRO KUZ MOL QAW SAP SHS TH2 TH3 TMG
ALT 0.000 0.003 0.000 0.001 0.000 0.000 0.067 0.000 0.081 0.000 0.000 0.000 0.425 0.004 0.000ALW 9.423 0.000 0.000 0.000 0.000 0.000 0.197 0.000 0.000 0.000 0.001 0.000 0.000 0.000 0.000CEMH 3.254 5.378 0.000 0.000 0.000 0.007 0.086 0.004 0.001 0.061 0.001 0.000 0.026 0.000 0.008DJR 4.512 8.280 6.102 0.000 0.000 0.000 0.660 0.228 0.554 0.000 0.358 0.000 0.002 0.000 0.000GKS 3.443 16.507 5.769 4.002 0.000 0.000 0.066 0.000 0.003 0.000 0.000 0.000 0.027 0.000 0.000HAI 31.612 11.799 12.222 21.312 40.812 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
HAR 9.300 15.797 3.031 9.003 10.669 24.431 0.018 0.000 0.000 0.014 0.000 0.000 0.014 0.000 0.009KRO 1.975 1.457 2.229 0.735 1.969 6.421 2.672 0.359 0.564 0.187 0.980 0.000 0.162 0.000 0.187KUZ 4.251 5.825 3.701 1.380 5.468 14.805 6.481 1.198 0.102 0.012 0.061 0.000 0.006 0.000 0.003MOL 1.839 5.467 4.406 0.861 3.247 17.226 8.291 0.854 1.802 0.000 0.183 0.000 0.038 0.000 0.000QAW 6.043 4.228 2.208 5.113 7.511 7.301 2.712 1.739 3.139 5.209 0.000 0.000 0.002 0.000 0.008SAP 4.585 3.812 4.768 1.130 5.613 11.172 6.447 0.233 2.107 1.497 3.816 0.000 0.002 0.000 0.001SHS 4.919 17.950 4.874 13.115 11.404 40.557 12.834 4.160 8.480 7.782 6.883 12.318 0.041 0.001 0.000TH2 1.027 7.983 2.706 3.647 2.345 16.304 4.033 1.853 3.560 2.314 4.638 4.019 2.145 0.733 0.070TH3 2.948 29.446 4.768 10.984 8.508 77.572 12.042 5.928 7.091 6.702 7.929 11.006 3.294 0.652 0.000TMG 4.308 9.547 3.450 5.492 5.359 19.491 2.898 1.740 4.117 5.111 3.594 4.596 5.922 2.168 8.960
1 F-values are below diagonal. Probability values (P-values) areabove diagonal.Abbreviations for samples are from Table 1. Only thosesamples with individual data are included. This resulted in elimination of Andronovo-Minusinsk (ADM), Afanasievo-Altai (AFA),Afanasievo-Minusinsk (AFM), Andronovo-Kazakhstan (AND), Karasuk-Minusinsk (KAM), Kara depe (KAR), Kokcha III (KOK),Samtavro (SAMB), and Tigrovija Balka-Makoni Mor (TMM).
Fig. 2. WPGMA cluster analysis of Mahalanobis d2 values.Branch points are Euclidean distances. Sample abbreviationsfrom Table 1.
INHABITANTS OF XINJIANG 209
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
12/24
Turkmenian samples from Geoksyur (GKS) andAltyn depe (ALT) serve as a phenetic link betweenIndus Valley samples (HAR, TMG, and CEMH) thatfeature the closest affinities to one another. In adeparture from the results obtained by WPGMAanalysis, the sample from Kara depe (KAR) occupiesa unique position among Turkmenian samples byexhibiting much closer affinities to Iranian samples(especially TH3 and SHS) than to samples fromBactria.
Andronovo and Afanasievo steppe samples occupythe left side of the array. Steppe samples fromTigrovaja Balka/Makoni Mor (TMM), Samtavro
(SAMB), and Kokcha III (KOK) occupy an interme-diate phenetic position; these samples, especiallyTMM, manifest some affinities to Iranian samples.Afanasievo samples (AFA and AFM) are identifiedas possessing the closest affinities to one another,and exhibit affinities to the Andronovo samples(AND and ADM) as well. The Karasuk sample fromthe Minusinsk region (KAM) stands apart as themost divergent of the steppe samples considered.
Cophenetic correlation coefficients
The cophenetic correlation coefficient for the de-gree of correspondence between the phenogram ob-tained by WPGMA cluster analysis and the bias-
adjusted matrix of Mahalanobis d2
values is low(rcs
0.496). This suggests that a fair amount ofdistortion is encountered when attempting to ar-range intersample differences in craniometric vari-ation in a hierarchical fashion through cluster anal-ysis (Rohlf, 2000). Sneath and Sokol (1973)recommended that alternative methods of data re-duction be used in cases where cophenetic correla-tions indicate that a fair amount of distortion of theoriginal data matrix is incurred by hierarchical clus-ter analyses. Specifically, Sneath and Sokol (1973)recommended the use of multidimensional scalingand principal coordinates analysis.
Multidimensional scaling
Multidimensional scaling of the bias-adjusted di-agonal matrix of d2 values into three dimensionswith the coefficient of alienation of Guttman (1968)is accomplished with a stress value of 0.097 after100 iterations. This value falls within acceptablelimits, and indicates that multidimensional scalingof these data into three dimensions provides an ar-ray of intersample associations only mildly affectedby distortion. A plot of multidimensionally scaledvalues, with a minimum spanning tree imposedbetween individual data points, is provided in Fig-ure 4.
An examination of this array confirms the pat-terns of interregional affinities identified by neigh-bor-joining cluster analysis (Fig. 3). Hainan (HAI)reflects the most divergent sample. The two laterwestern Chinese samples, Kroran (KRO) and Alwi-ghul (ALW), feature the closest affinities to Sapalli(SAP), the earliest of the Bactrian samples. Two ofthe samples from Turkmenistan (Altyn depe (ALT)and Geoksyur (GKS)) span the phenetic space be-tween Iranian samples and Bactrian samples, withGeoksyur exhibiting closer phenetic affinities toBactrians (especially the latest sample, Molali(MOL)), while Altyn depe shares closer phenetic af-
finities to Iranians. The steppe Bronze Age samplefrom the Caucasus (SAMB) represents a pheneticoutlier to all other samples, exhibiting only a verydistant affinity to the sample from Altyn depe. IndusValley samples share rather close affinities to oneanother but are strongly segregated from all othersamples, except the early western Chinese samplefrom Qawrighul (QAW).
All steppe Bronze Age samples, except Samtavro(SAMB), are found on the left side of the array.Intersample affinities among the Karasuk (KAM),Afanasievo (AFA and AFM), and Andronovo (ANDand ADM) samples are relatively close. However,
Fig. 3. Neighbor-joining tree based on Mahalanobis d2 values. Sample abbreviations from Table 1.
210 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
13/24
steppe Bronze Age samples from Turkmenistan(KOK) and Tajikistan (TMM), while exhibiting dis-tant affinities to other steppe Bronze Age samples,appear more closely aligned to sedentary agricul-tural samples from Turkmenistan (Kara depe(KAR)) and, to a lesser degree, eastern Iran (Shahr-iSokhta (SHS)).
Principal coordinates analysis
A principal coordinates analysis of the double-centered Mahalanobis d2 matrix yields three coordi-nate axes that combine to explain 89.9% of the totalvariance. Comparison of the eigenvector matrix withthe d2 matrix yields a cophenetic correlation coeffi-cient whose value (rcs 0.948) indicates that thefirst three eigenvectors provide an excellent fit of thedata (Rohlf, 2000). An ordination of group scores forthe first three coordinate axes is provided in Figure5, and a minimum spanning tree was imposed onthis array to clarify associations between samples.
The pattern of intersample variation provided bythis analysis confirms many of the major featurespreviously identified by neighbor-joining clusteranalysis (Fig. 3) and multidimensional scaling (Fig.4). Once again, the two later western Chinese sam-ples, Kroran (KRO) and Alwighul (ALW), exhibit theclosest affinities to the earliest Bactrian sample,
Sapalli (SAP). Bactrian samples (SAP, DJR, KUZ,and MOL) exhibit the closest affinities to one an-other. The two Turkmenian samples from Geoksyurand Altyn depe occupy an intermediate pheneticposition between Bactrians and northern Iranians,in which the former (GKS) shares the closest affin-ities with the latest Bactrian sample (MOL), whilethe latter (ALT) shares the closest affinities with theearlier northern Iranian sample (TH2). Indus Valleysamples (HAR, CEMH, and TMG) are located in thelower left of this array and, once again, the earliestwestern Chinese sample, Qawrighul (QAW), is iden-tified as possessing closer affinities to Indus Valley
Fig. 4. Minimally spanned plot of sample values for first three multidimensionally scaled dimensions. Sample abbreviations fromTable 1. Xinjiang samples (QAW, ALW, and KRO) and Chinese sample from Hainan (HAI) are represented by asterisks; NorthBactrian samples, by stars; Iranian samples, by pentagons; Turkmenian, Caucasus, and Tajik samples, by triangles; Indus Valleysamples, by circles; and Russo-Kazakh samples, by squares.
INHABITANTS OF XINJIANG 211
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
14/24
samples than to samples from any other region.Standing somewhat in contrast to results obtainedby other analyses, principal coordinates analysisidentifies an especially close affinity between theLate Bronze-Early Iron Age sample from the SwatValley of Pakistan (TMG) and the early northernIranian sample (TH2). As with other analyses, thisarray also indicates that the Turkmenian sample
from Kara depe (KAR) is strongly separated fromother sedentary Turkmenistan samples, but unlikeother analyses, principal coordinates analysis indi-cates that this sample possesses no close affinitieswith any of the other samples considered.
All steppe Bronze Age samples, regardless of geo-graphic location, occupy the right side of this array.In agreement with other analyses, the Afanasievosamples (AFA and AFM) exhibit the closest affini-ties to one another. However, unlike other analyses,the patterning of affinities yielded by principal coor-dinates analysis suggests a moderate degree of dis-tinctiveness between Afanasievo samples and An-
dronovo samples (AND and ADM). The sample fromTajikistan (TMM) is identified as the steppe samplewith closest affinities to nonsteppe samples in gen-eral, and with the eastern Iranian sample fromShahr-i Sokhta (SHS) in particular.
Mantel tests
The normalized Mantel statistic, which is equiva-
lent to a correlation coefficient (r), obtained betweenthe Mahalanobis d2 matrix and the matrix of chro-nological differences between samples, is 0.294. Thepermutational probability to observe a higher orequal correlation based on 1,000 permutations is
P 0.827. This value suggests that differences inantiquity, ranging from 3500 B.C. to the present, donot contribute significantly to the patterning ofcraniometric differentiation among these samples.Given ample opportunity for responses to changes inselection pressures due to exposure to agriculturaldiets and more sophisticated food preparation tech-niques during passage of the more than five millen-
Fig. 5. Minimally spanned ordination of sample scores for first three principal coordinate axes. Sample abbreviations from Table1. Xinjiang samples (QAW, ALW, and KRO) and Chinese sample from Hainan (HAI) are represented by asterisks; North Bactriansamples, by stars; Iranian samples, by pentagons; Turkmenian, Caucasus, and Tajik samples, by triangles; Indus Valley samples, bycircles; and Russo-Kazakh samples, by squares.
212 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
15/24
nia encompassed by these comparative samples, theabsence of any chronological effect on the patterningof phenetic distances suggests that either these se-lective pressures led to an alteration of cranio-gnathic dimensions prior to 3500 B.C. or that thisbattery of measurements is not affected in any ap-
preciable way by changes in masticatory pressures.A comparison between the Mahalanobis d2 matrixand the matrix of geographical distances betweensamples yields a correlation coefficient of r 0.560.The permutational probability to observe a higher orequal correlation is also not significant, with a valueof P 0.330. Contrary to standard expectations ofisolation by distance (Barbujani, 1987; Cavalli-Sforza et al., 1994; Fix, 1999; Kimura and Weiss,1964; Malecot, 1967; Morton et al., 1982; Piazza andMenozzi, 1983; Sokol et al., 1986; Sokal and Warten-burg, 1983; Wright, 1943, 1946, 1951), these resultsindicate that the amount of geographic distance be-tween individual samples does not provide an im-
portant contributing factor behind the patterning ofcraniometric differentiation among these samples.
DISCUSSION
Numerous specific hypotheses have been ad-vanced to account for the initial appearance ofBronze Age populations found at a series of oasesskirting the margins of the Taklamakan Desertwithin the Tarim Basin of Xinjiang, western China,during the final two millennia B.C. These individualhypotheses can be grouped into two general models,and the model currently favored by a small majorityof archaeologists working in Central Asia and west-
ern China is the steppe hypothesis (Han, 1998;Kuzmina, 1998; Mair, 1995; Mallory and Mair, 2000;Parpola, 1998). As a general model, this hypothesisholds that for reasons as yet unknown, Afanasievo-related steppe populations from the north andnorthwest began to emigrate southward, either di-rectly into the Tarim Basin (Kuzmina, 1998), orsubsequent to contact with more settled agriculturalpopulations in Central Asia (Mallory and Mair,2000). These immigrants are thought to be repre-sented by the human remains recovered from suchearly Bronze Age sites as Qawrighul (Kuzmina,1998; Mallory, 1995; Mallory and Mair, 2000). Later,beginning around 1200 B.C., the archaeological
record of Xinjiang reveals a series of changes intextile manufacture and clothing design. Althoughthere are always problems in equating changes inmaterial culture with population movements, propo-nents of the steppe hypothesis suggest that thesechanges signal the appearance of a second wave ofimmigration to the Tarim Basin from the Russo-Kazakh steppe. In this latter case, these immigrantsare held to be members of the widespread An-dronovo culture complex that appears throughoutthe south Russian steppe, Kazakhstan, and westernCentral Asia during the middle of the second mil-lennium B.C.
Given both the archaeological (Kuzmina, 1998)and craniometric (Han, 1998) arguments, remainsrecovered from the earliest sample, Qawrighul,should exhibit broad phenetic similarities to Afa-nasievo samples from the Altai and Minusinsk,while later Tarim Basin samples from Xinjiang (Al-
wighul and Kroran) should exhibit closer pheneticaffinities to the later Andronovo samples from Ka-zakhstan and Minusinsk. Since most proponents ofthe steppe hypothesis envision the immigration ofAndronovo populations as limited to several centu-ries spanning the end of the second and the begin-ning of the first millennia B.C., late Bronze Agepopulations of the Tarim Basin are expected to besequentially more divergent from their Andronovosource populations over time due to genetic drift.Hence, the Alwighul sample (ca. 800200 B.C.), if ittruly predates the sample from Kroran (ca. 202B.C.A.D. 220), should exhibit closer affinities toAndronovo samples, while the Kroran sample
should be more divergent. Bactrian, Iranian, andIndus Valley populations are thought to have playedlittle to no role in the origins of Bronze Age inhab-itants of the Tarim Basin of Xinjiang; therefore,samples from these latter regions should be mark-edly divergent phenetically.
Most advocates of the steppe hypothesis recognizean East Asian contribution to the Xinjiang gene poolsubsequent to that provided by Afanasievo-relatedsteppe populations, but contemporaneous with thatprovided by the later influx of Andronovo steppepopulations (Han, 1998; Mallory and Mair, 2000).Han (1998, 2001, p. 237239) maintained that thisinfluence is largely restricted to such eastern Tarim
Basin samples as Yanbulaq (Han, 1990), but identi-fied seven of the crania from the earlier graves atAlwighul (Han, 1998) and a single female from Kro-ran (Han, 1986b) as Mongoloid. If Han (1998) iscorrect that East Asian populations contributed toeastern Tarim Basin populations in general and ac-count for a minority of individuals encompassed byAlwighul (12%) and Kroran (17%) samples, theselater Tarim Basin samples should be marked by areduction in phenetic distance from the Han Chi-nese sample (HAI) relative to that found for theearlier sample from Qawrighul.
The results of all analyses provide abundant evi-dence in support of a migration of pastoralist popu-
lations across the Russo-Kazakh steppe. This is re-flected by the degree of phenetic cohesion foundamong steppe samples, regardless of the geographicdistances that separate them. Further, once steppesamples are removed from consideration, a Manteltest of the correlation between the Mahalanobis d2
matrix and the matrix of geographical distances be-tween samples yields a highly significant (P 0.001)correlation coefficient (r 0.871). Thus, the appar-ent departure of the patterning of phenetic distancefrom expectations of an isolation-by-distance modelappears to be due to a spread of steppe pastoralistpopulations across an enormous distance from the
INHABITANTS OF XINJIANG 213
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
16/24
trans-Ural region in the west to the Minusinsk Ba-sin in the east. In this case, the close similarities inarchaeological assemblages attributed to Andronovoand Afanasievo archaeological horizons do appear todocument an eastward and southward populationexpansion.
Nevertheless, there is no support for the hypoth-esis that steppe populations contributed signifi-cantly to Bronze Age populations of the Tarim Ba-sin. Despite numerous similarities betweenAfanasievo and Andronovo artifacts and Bronze Ageartifacts from Xinjiang (Bunker, 1998; Chen andHiebert, 1995; Kuzmina, 1998; Mei and Shell, 1998;Peng, 1998), all analyses of phenetic relationshipsconsistently reveal a profound phenetic separationbetween steppe samples and the samples from theTarim Basin (Qawrighul, Alwighul, and Kroran).Further, neither of the later Tarim Basin samplesfrom Alwighul or Kroran appears phenetically closerto the Han Chinese sample from Hainan, thereby
indicating an absence of East Asian influence inthese samples.The second model offered to account for the origins
of the Bronze Age inhabitants of the Tarim Basin isthe Bactrian oasis hypothesis (Askarov, 1973, 1981,1988; Barber, 1999). Proponents of this model em-phasize the similarity in environmental conditionsbetween the oases skirting the Taklamakan Desertand those found in Bactria and Margiana to thewest. Proponents of the Bactrian oasis model arguethat the very skills developed by the founders andoccupants of the urban centers of the Oxus civiliza-tion (irrigation agriculture, development of exten-sive trade networks between locally resource-impov-
erished oases, and domestication of sheep and goats)are exactly those that accompany the initial appear-ance of Bronze Age populations in the Tarim Basin(Barber, 1999; Chen and Hiebert, 1995). To explainthe changes in textile manufacture, clothing styles,and metallurgical technology found in the TarimBasin beginning around 1200 B.C., some proponentsof the oasis model concur with the steppe hypothesisand envisage a second influx of colonists from thesteppelands (Barber, 1999)
If this model is true, the earliest Tarim Basinpopulations, such as Qawrighul, should possessclose similarities to samples from Bactria. Giventhat the nature of this interaction is thought to have
been unidirectional, from Bactria to the Tarim Ba-sin, and limited in duration, phenetic affinities be-tween populations of the two regions should initiallybe close and then progressively decrease over timeas the two gene pools became increasingly distinctdue to genetic drift. Tarim Basin inhabitants thatpostdate 1200 B.C. should represent the impact ofAndronovo immigrants from the Russo-Kazakhsteppe. Later Bronze Age inhabitants of the TarimBasin should be marked by a reduction in pheneticdistance to steppe populations in general, and toAndronovo samples in particular. Phenetic affinitiespossessed by the samples from Alwighul and Kroran
should be markedly closer to those of steppe samplesthan those possessed by the earlier sample fromQawrighul.
The results obtained offer little support for theBactrian oasis hypothesis. While Tarim Basin sam-ples do exhibit closer affinities to samples from the
urban centers of the Oxus civilization than to steppesamples, three aspects of the patterning of interre-gional phenetic affinities run counter to the expec-tations of this model. First, rather than identifyingthat closest affinities occur between early TarimBasin (Qawrighul) and earlier or contemporaneousOxus civilization samples that antedate or are con-temporaneous (Sapalli and Djarkutan), the closestaffinities actually occur between the earliest of theOxus civilization samples, Sapalli, and the latest ofthe Tarim Basin samples, Kroran, followed by thelate sample from Alwighul. Second, none of the Ta-rim Basin samples, not even those that postdate1200 B.C., exhibit any phenetic affinities to any of
the steppe samples included in this analysis. Third,while neighbor-joining cluster analysis (Fig. 3) sug-gests a distant affinity between Qarwighul, the ear-liest Tarim Basin sample, and the Oxus civilizationsamples, this is not confirmed by any other analysis.While a case could be made for greater involvementof Bactrian oasis peoples in the population history ofthe Tarim Basin during the Bronze Age than bysteppe populations, the nature of this involvement isnot predicted by the Bactrian oasis hypothesis.
The results fail to demonstrate close phenetic af-finities between the early inhabitants of Qawrighuland any of the proposed sources for immigrants tothe Tarim Basin. The absence of close affinities to
outside populations renders it unlikely that the hu-man remains recovered from Qawrighul representthe unadmixed remains of colonists from the Afa-nasievo or Andronovo cultures of the steppelands, orinhabitants of the urban centers of the Oxus civili-zation of Bactria.
Three alternative possibilities remain once simplelarge-scale emigration from a known source popula-tion is ruled out. First, the human remains fromQawrighul may be those of a local, indigenous pop-ulation from the Tarim Basin itself or the surround-ing highlands. Second, the human remains fromQawrighul may be the product of emigration from asource area other than the Russo-Kazakh steppel-
ands or Oxus civilization urban centers. Third, thehuman remains from Qawrighul may derive fromone of the suggested source areas, but the separationof this emigrant population from the host populationinvolved fewer founding individuals or occurred ear-lier than currently thought by proponents of eitherthe steppe or Bactrian oasis hypotheses. Under suchconditions, a founder effect, coupled with subse-quent genetic drift, may have resulted in ameliora-tion of phenetic similarities detectable throughcraniometric analyses.
The first alternative is certainly possible, for ad-vocates of both the steppe and Bactrian oasis hy-
214 B.E. HEMPHILL AND J.P. MALLORY
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
17/24
potheses admit the presence, albeit scarce, of a se-ries of Neolithic sites in the basin that antedate theinitial Bronze Age (An, 1992a; Bergman, 1939;Chang, 1977; Chen and Hiebert, 1995; Chen, 1990;IAX, 1989; Mallory, 1995; Olsen et al., 1988; Teil-hard de Chardin and Young, 1932; Wang, 1985; Xu,
1995). The recovery of microliths on the surface inassociation with painted ceramics at Yierkabakeand with plain red wares at Xingeer led Wang(1985) to suggest that the transition from the Neo-lithic to the Bronze Age, in at least the easternportion of the Tarim Basin, was one of cultural con-tinuity rather than an unprecedented introductionof a foreign Bronze Age cultural complex.
If a resident population, regardless of ultimatederivation, was present in the Tarim Basin at theemergence of the Bronze Age, initiation of theBronze Age in this region may have been the productof a more subtle interaction between this local pop-ulation and groups from adjacent regions. From a
biological perspective, such interactions may haveinvolved limited levels of unidirectional or bidirec-tional gene flow. Under such conditions, the biolog-ical impact of emigrants from outside would likelybe muted relative to the impact of outright whole-sale colonization of an essentially unpopulated, orminimally populated, region (Ayala, 1982; Bodmerand Cavalli-Sforza, 1975; Cavalli-Sforza et al., 1994;Cummings, 1997; Fix, 1999; Weiss, 1988; Wijsmanand Cavalli-Sforza, 1984). Unless by some fortuitouscircumstance the remains recovered from Qawri-ghul are those of some foreign entrepot, these re-mains should reflect the impact of such gene flow bymoderate to low affinities to those adjacent, non-
Tarim Basin populations with whom they were incontact.
The results, however, fail to demonstrate even alow-level phenetic affinity between Qawrighul andeither steppe samples or samples from Oxus civili-zation urban centers. Not only is there no evidencefor substantial immigration into the Tarim Basin bypopulations of these two adjacent regions; it alsoappears unlikely that either steppe populations orOxus civilization populations served as a source ofany significant gene flow commensurate with theappearance of the Bronze Age occupation of Qawri-ghul. Given the paucity of contemporaneous skeletalremains from other parts of the Tarim Basin, the
presence of immigrants from either the steppelandsor the urban centers of the Oxus civilization cannotbe definitively ruled out.
The second alternative explanation to account forthe human remains from Qawrighul is that they arethe product of emigration from a source area otherthan the Russo-Kazakh steppelands or Oxus civili-zation urban centers. While the results obtainedindicate that there is no evidence that gene flowfrom either steppe or Oxus civilization populationsled to the establishment of the Qawrighul popula-tion, all analyses, except neighbor-joining clusteranalysis (Fig. 3), disclose a low-level affinity be-
tween the Qawrighul and Indus Valley samples.Such affinities could be indicative of some early in-teraction between the populations of these two re-gions. The implications of such early interaction arepotentially profound.
In a reversal of mainstream thought on a western
Asian homeland (Urheimat) and eastward dispersalof Indo-European languages into Central Asia andIndia (Burrow, 1973; Gamkrelidze and Ivanov, 1990;Mallory, 1989; Renfrew, 1988), there is a body ofscholars who have vigorously argued for an Indo-European homeland in the Indus Valley of India andPakistan (surveyed at length in Bryant, 2001), orthat Indo-European languages disseminated from alocus somewhere in the vicinity of ancient Bactria-Sogdiana (Nichols, 1997, p. 137; see also Sargent,1997). If true, the dispersal of these Indo-Europeanlanguages may have been accompanied by immigra-tion and some gene flow from the Indus Valleyhomeland to the various historical seats of the Indo-
European languages. In this way, Tocharian lan-guages found in the Tarim Basin would be attrib-uted to the influx of populations from Bactria whoseultimate derivation may be traced to the Indus Val-ley of India and Pakistan.
The results of this study offer little support forsuch a scenario. The problems are threefold. First,WPGMA cluster analysis (Fig. 2) identifies Qawri-ghul as possessing closer affinities to the two sam-ples from Harappa than are possessed by the LateBronze-Early Iron Age sample from Timargarha. Itis difficult to see any archaeological support for sucha connection, as there are no material artifacts ofmature Harappan or even late Harappan attribu-
tion found at Qawrighul (Chen and Hiebert, 1995;Wang, 1982, 1983). Likewise, there are no artifactsreflective of Tarim Basin derivation at either matureHarappan (HAR) or late Harappan (CEMH) levelsat Harappa (Allchin and Allchin, 1982; Kenoyer,1998). Second, if Indo-European-speaking popula-tions entered the Tarim Basin from Bactria, Bac-trian populations should also show evidence of geneflow from the Indus Valley. None of the analysespresented here or in previous assessments (Hemp-hill, 1998, 1999; Hemphill et al., 1998) provide anyevidence of significant interaction between Bactrianand Indus Valley populations prior to the latter halfof the first millennium B.C. Finally, greater insight
into the relationship between Indus Valley and Ta-rim Basin populations is provided by multidimen-sional scaling (Fig. 4) and principal coordinate anal-ysis (Fig. 5). Both merely identify Qawrighul asoccupying a peripheral and opposite phenetic posi-tion to whatever Indus Valley sample is least sepa-rated from other regional samples. Such positioningis best interpreted as evidence of outlier status to allsamples considered in this multidimensional array,rather than of any peripheral association to IndusValley samples per se.
With neither biological nor archaeological sup-port, there is no compelling evidence to uphold the
INHABITANTS OF XINJIANG 215
7/24/2019 A Craniometric Investigation of the Bronze Age Settlement of Xinjiang
18/24
idea that Indo-European languages were introducedinto the Tarim Basin from populations emigratingfrom the Indus Valley commensurate with the initi-ation of the Xinjiang Bronze Age. Given the infor-mation currently available, it is most likely that theearly Tarim Basin sample from Qawrighul occupies
an isolated phenetic position, because this samplerepresents a population of western China to whichnone of the potential regional contributors repre-sented in this analysis (Russo-Kazakh steppe, Bac-tria, Indus Valley, and south China) contributedsubstantially.
Yet while the cranial series from Qawrighul ex-hibits no distinct affinities to any of the other sam-ples included in this analysis, this is not the case forthe later Tarim Basin samples from Alwighul andKroran. Although results differ as to whether Kro-ran has closer affinities to the inhabitants of theearliest of the north Bactrian urban centers (Sapalli)than does Alwighul (Figs. 2, 3), or whether affinities
to the inhabitants of Sapalli are equally close (Figs.4, 5), all results indicate that these later inhabitantsof the Tarim Basin manifest a unique affinity toBactrians. None of the results revealed affinitiesbetween Tarim Basin samples and samples from theRusso-Kazakh steppelands, the Indus Valley, or theHan Chinese sample from Hainan.
It appears that neither Han Chinese nor steppepopulations played any detectable role in the initialestablishment or subsequent interregional biologi-cal interactions of Bronze Age Tarim Basin popula-tions. This conclusion, however, must be temperedwith a pair of caveats. First, East Asian populationsmay have played a late role in the eastern oases ofthe Tarim Basin (Han, 1994b, 1998). Until adequatesamples from such eastern Tarim Basin sites asYanbulaq (Han, 1990, 1998) and from temporallyand geographically more appropriate Han Chinesecontexts become available, this possibility must re-main open. Second, while Russo-Kazakh steppe pop-ulations appear to have played no role in the estab-lishment of southern (KRO and QAW) and northernTarim Basin oasis populations (ALW) for which dataare available, this does not rule out a role for steppepopulations in Xinjiang during the Bronze Age alltogether. It is possible that Afanasievo populations,Andronovo populations, or both may have played an
important role in the development of Bronze Agecultures of the Zhungeer Basin and the Yili Valley,located north of the Tian Shan Mountains in north-e

![Craniometric Analysis of the Hindbrain and Craniocervical ...epubs.surrey.ac.uk/813560/1/PLOS ONE Craniometric... · [25–27]. The Chihuahua is the smallest known dog breed for which](https://img.dokumen.tips/doc/110x75/5f6bfefecd418f439b108db3/craniometric-analysis-of-the-hindbrain-and-craniocervical-epubs-one-craniometric.jpg)




![FOREIGN BROADCAST If uirf/ INFORMATION …Textile Industry Adjusts Product Structure [XINJIANG RIBAO 16 Mar] 11 Xinjiang Official Describes Causes, Solutions to Inflation [XINJIANG](https://img.dokumen.tips/doc/110x75/5fb5b3e123d56a3a9d730569/foreign-broadcast-if-uirf-information-textile-industry-adjusts-product-structure.jpg)
![Prehistoric Interactions in Eurasia: A Re-evaluation of ...Xinjiang Wenwu Kaogu Yanjiusuo [Xinjiang Institute of Archaeology] (Urumqi: Xinjiang meishu shejing chubanshe 1997: 44-56)](https://img.dokumen.tips/doc/110x75/5f18bffb0b48650cc441aaeb/prehistoric-interactions-in-eurasia-a-re-evaluation-of-xinjiang-wenwu-kaogu.jpg)