Embed Size (px)
Citation preview

8.9Metallothioneins
P. GONZALEZ-DUARTE
Universitat Autonoma de Barcelona, Spain
8.9.1 INTRODUCTION 2138.9.1.1 Sources, Classification, and Function 2148.9.1.2 Spectroscopic Characterization and Stoichiometry of the Metal–Sulfur
Aggregates in Metallothioneins 2158.9.1.3 Three-dimensional Structures 2158.9.1.4 Metal Thiolates as Models of the Metal–Sulfur Aggregates in Metallothioneins 2208.9.1.5 Dynamic Aspects and Reactivity 223
8.9.2 REFERENCES 225
8.9.1 INTRODUCTION
Metallothioneins (MTs) constitute a very wide family of ubiquitous proteins with features that areexceptional from the perspectives both of biology and coordination chemistry. If compared withall other metalloproteins, MTs are wholly unconventional because of their small size (2–7 kDa),their extremely high thiolate sulfur and metal content, both in the order of 10% (w/w), and thelack of tertiary structure in their apo form (apo-MT or thionein). The structural properties thatdistinguish all MTs are based on the metal-thiolate aggregates that give rise either to a singlemetal-binding domain, as in yeast, or to two such domains, as in mammals. These can beregarded as polynuclear homo- or heterometallic complexes of a multidentate ligand, involvingterminal and/or bridging cysteinyl thiolate groups, and thus their study could be related to thefield of metal thiolate complexes. However, from the perspective of coordination chemistry, MTsare unique as well as intriguing, given the different number and nature of the metal centers inMTs, the possible coexistence of several coordination geometries even for the same metal, and theinfluence of the peptide chain on the ligation modes of the thiolate sulfur.
Even though MTs exist naturally with zinc and/or copper bound to them, the discovery of thefirst MT in 1957 from horse kidney was the result of a search for a cadmium protein.1 Since then,MTs have continuously challenged the interest of chemists and life scientists. A search in theSciFinder database with metallothionein as the entry yields about 15,000 publications and revealsmore than 700 articles per year over the 1991–2001 decade. It also shows that developments inMT research have been covered by about 300 reviews. The widespread occurrence of MTsin nature suggests that they serve an important biological function not yet completely established.It would appear that MTs have no enzymatic activity, nor do they perform any catalytic role inknown metabolic processes. Precise identification of the function of MTs accounts for the out-standing number of works available (as indicated by the search results) and prompts most of theresearch currently being undertaken.
This section is intended as a broad overview of the present knowledge of MTs from theperspective of coordination chemistry; the main developments with respect to their metal content,structure, spectroscopic features and chemical properties will therefore be summarized. Moreover,the structural patterns of the metal–sulfur aggregates in MTs will be compared with those found
213

in synthetic metal thiolates. A recent review of the structure and chemistry of MTs includes themost relevant literature prior to 2001.2 Progress in the study of MTs by scientists from differentfields has been covered in the proceedings of the four international conferences devoted exclu-sively to MTs.3–6 Standard operating procedures for the isolation, quantification, and the chem-ical, physical, and spectroscopic characterization of MTs have been compiled in a methodologicalcontribution.7 General reviews covering all aspects of MT research are also available.8–12
8.9.1.1 Sources, Classification, and Function
MTs have been isolated from a wide range of living systems, from eukaryotic—and someprokaryotic—microorganisms through plants and invertebrates to mammals. A remarkable bio-logical feature of MTs is that their genes are induced by a variety of agents. Their biosynthesis istherefore greatly enhanced both in vivo and in cultured cells by metal ions, certain hormones, andmany cytotoxic agents. In animals, MTs are most abundant in parenchymatous tissues, i.e., liver,kidney, pancreas, and intestines.8 Procedures for the isolation and characterization of MTs frombiological sources, and special precautions that avoid oxidation of the cysteinyl residues formingdisulfide bridges, as well as the randomization of metals, have all been reported.7
Less frequently, conventional chemical peptide synthesis has been used to produce the entireapo-MT protein (when the amino acid sequence is relatively short) or in the production of theseparate MT domains. Thus, the metal-binding features of synthetic MTs from fungi13,14 or theindividual domains of crustacean15 and mammalian13,16–19 MTs have been characterized. Add-itionally, as peptide synthesis provides the methodology for a facile sequence modification, studieson designed MT-related peptides have also been carried out.20,21
The expression of MTs in recombinant systems has recently been proven as an effectivealternative to their isolation from natural sources, thereby giving further stimulus to the studyof MTs. After some early attempts,22–24 current heterologous expression systems (essentially inE. coli) provide significant quantities of the desired MT, avoiding whatever constraint may beimposed by the availability of the corresponding native organisms.19,25–31 In addition, byadapting the metal content in the culture media, MTs containing different metal ions canbe recovered.32,33 Furthermore, the expression and purification of separate MT domains affordan adequate alternative to proteolytic digestion procedures in the entire protein, which maycompromise the integrity of the isolated peptides.21,34–36 Finally, the DNA recombinanttechniques allow the expression and purification both of wild-type and mutant forms, thusfacilitating the analysis of the relative contribution that individual residues make to the in vivoand in vitro functions of MTs.19,37–41
Historically, three different classes were proposed in order to classify the extremely heterogeneousamino acid sequences of MTs: class I, comprising all MTs with clear sequence similarity tomammalian MT; class II, including polypeptides that are only distantly related to mammalianMT; and class III, for the enzymatically synthesized peptides of general formula (�-Glu-Cys)nGly(the values of n ranging from 2 to 8), variously known as phytochelatins and cadystins and denotedas (�-Glu-Cys)nGly(�-EC) peptides.42 More recently, a new classification consisting of a singularset for each individual taxon has been proposed (www.unizh.ch/�mtpage/classif.html). Bothschemes are based on primary structure data, but do not include any functional or evolutionaryinformation.26 With respect to the primary structure of all MTs, the most remarkable feature is therecurrence of Cys–X–Cys tripeptide sequences, where X stands for an amino acid residue other thancysteine. To date, all vertebrates examined contain two or more distinct MT isoforms, denotedMT-1 through MT-4, which differ in amino acid residues other than the conserved cysteines.In mammals, MT-1 and MT-2 are found in all organs, MT-3 is expressed mainly in the brain,and MT-4 is most abundant in certain stratified tissues.8
Due to both the origin of their discovery and to their high metal-inducibility, it has long beensuggested that one of the primary roles of MTs is to serve as a heavy-metal detoxification system.43
However, it seems likely that this is not the primary function of MTs, but rather is simply aconsequence of their high cysteine content.44 A more physiological view generally considers thatMTs could be involved in the homeostasis of copper and (especially) zinc ions.45,46 There has beenmounting evidence in recent years to suggest that MTs could be significant antioxidant andantiapoptotic proteins,47,48 being especially important within the central nervous system.49 Further-more, other studies suggest that these proteins are also involved in the control of the redox status ofcells and in energy metabolism.50–53
214 Metallothioneins

8.9.1.2 Spectroscopic Characterization and Stoichiometry of the Metal–SulfurAggregates in Metallothioneins
Unlike other metalloproteins, there is no specific metal-to-MT stoichiometry that defines MTs.Consequently, determination of the metal content is a primary step in the characterization of anew member of this large family of proteins, consisting of 197 sequences from 103 organisms(Swiss-Prot: www.expasy.ch/sprot/sprot-top.html). Within the MT family, differences in thelength of the primary sequence and in the number of cysteines they encompass account for thehighly variable metal-to-MT stoichiometries reported to date. Furthermore, not only is there nounique metal preference shown by mammalian and nonmammalian MTs15,26,54 but—addition-ally—they can bind a mixture of different metal ions.10 This complexity is added to by the factthat proteins with different metal contents are obtained depending both on the source and on theinduction, isolation, and purification procedures.7
The analysis of the metal content in MTs includes determination of the number and nature ofthe bound metal ions55 and elucidation of the coordination geometry around the differentmetal centers enfolded by the polypeptide chain. To this end, optical spectroscopies (UV-Vis, c.d.MCD and luminescence) have played a significant role,10,56–58 despite providing information onthe predominant species present in solution. Conversely, electrospray mass spectrometry (ES MS)allows determination of the molecular distribution of the various complex species coexistingwithin the sample.59–61 However, in ES MS, the formation of artifacts due to operational featuresshould not be disregarded.62–64 A summary of the techniques more commonly used in the study ofthe metal-binding features of MTs can be found elsewhere.65 Additionally, relevant informationon the application and possible limitations of the X-ray absorption spectroscopy within the studyof MTs has been reported.10,66–69
As regards the metal preferences of MTs, vertebrates and higher invertebrates naturally giverise either to Zn- or mixed Zn,Cu-loaded species, while yeast and fungi give rise to Cu-containingproteins. Concomitantly, it is widely accepted that in higher organisms the binding of theessential metals, Zn and Cu, occurs in two separate domains as opposed to lower organisms, whereCu is accommodated in only one domain.2,70 Both features, the nature of the bound metal (Zn or Cu)and the one- or two-domain cluster structure, are probably essential for the MTs’ different potentialroles.26 Within this context and in the absence of 3D structures of mixed Zn,Cu–MT species,recent studies suggest that ZnII is essential for the in vivo and in vitro folding of mouse copperMT-1 in two domains.33
Whereas extensive information on the metal-binding abilities of mammalian MTs has beencompiled,10 current knowledge of those of non-mammalian species is more limited and ratherdispersed. In order to fill this gap, existing data have been summarized in Table 1. A featurecommonly found in several MTs is that they contain heterometallic mixtures of two (Zn,Cd; Cu,Cd;Zn,Cu) or three (Zn,Cd,Cu) metal ions. Still more striking is the great diversity of stoichiometriesreported for certain homometallic metal-MT species, as shown in S. cerevisiae, where the copper-to-MT mole ratios range from 4 to 10, and in N. crassa, with a cadmium-to-MT mole ratio of 2 or 3.With this information on the metal-binding properties of MTs, it seems reasonable to conclude thatthe systematization of these data is hard indeed to achieve.
8.9.1.3 Three-dimensional Structures
Despite the discovery of MTs in 1957,1 and also many of the subsequent studies being directedtowards its metal content,113 the first detailed knowledge of the 3D structure for the metal-thiolate clusters was not reported until almost 30 years later. Currently, this knowledge is onlyavailable for a small number of MTs, if compared with the already reported primary structuresequences (Swiss-Prot), and includes mammalian, invertebrate, and bacterial forms exclusivelycontaining Zn and/or Cd ions (Table 2). To date, only limited structural data are available oncopper-containing MTs,85 attributable to the failure to obtain single crystals of the protein and tothe unsuitability of the 63Cu and 65Cu isotopes for NMR studies.18
The key to understanding the 3D structure of MTs was the detailed analysis of the 113Cd NMRspectra of rabbit liver Cd,Zn-MT-2,114which was reconfirmed by studies of rabbit liver Cd7 -MT-2.115
Subsequent multidimensional/multinuclear NMR studies on 113Cd-reconstituted Cd7 -MT-2 fromrabbit,116 rat,117 and human,118 and X-ray crystallographic studies of native rat Cd5,Zn2-MT-2119
(Figure 1) allowed determination of the first 3D MT structures. These showed identical metal-thiolate cluster architectures and closely comparable polypeptide folds in crystals and in solution.
Metallothioneins 215

Table
1Metal-to-M
Tstoichiometries
reported
forallknownMTsexceptformammalianform
s.
Taxon/organism
M-M
Tstoichiometry
Taxon/organism
M-M
Tstoichiometry
Bacteria:
Nem
atoda:
SynechococcusPCC
7942
Zn4-;Cd4,Zn-SmtA
71
Caenorhabditis
elegans
MII6-C
eMT-II(M¼Cd,Zn)29,62
Fungi:
Gastropoda:
Neurospora
crassa
Cu6-M
T;13,72,73
Cd2-M
T13M
II3-M
T(M¼Zn,Cd,Hg,Co,Ni)72,74
Helix
pomatia
(RomansnailorEdible
snail)
Cd5,Zn-M
T75
Cu12-M
T;62
Cu6-M
T54
Agaricusbisporus
Cu6-M
T76;Cu4.4-M
T14
Bivalvia:
Mytilusedulis
Cd7-M
T-10-IV62
Yeast:
Mytilusgalloprovincialis
Cd5-M
T77
Saccharomycescerevisiae
Cu10-C
UP178,79
M8-C
UP1(M¼Cu,Ag)80–83
M7-C
UP1(M¼Cu,Ag)83–86
Cu4-C
UP187M
II4-C
UP1
(M¼Zn,Cd,Fe)
81,88
Cu11–12-C
RS5;Cd6-C
RS589
Crassostreavirginica
Cd5,Zn1.7,Cu0.1-C
dBP190
Cd4.7,Zn1.6,Cu0.1-C
dBP290
Candidaglabrata
Cu11–12-M
TI;
Cu10-M
TII
91
Insecta:
Orchesella
cincta
Cd7-8-M
T92
Plants:
Pisum
sativum
(Pea)
Cd5.6–6.1-PsM
TA
93
Cd4-;Zn6–11.5-;Cu3.5-PsM
TA
94
Drosophilamelanogaster
MII3-þM
II4-M
TN
(M¼Zn,Cd)25
Cu8-M
TN
25
Lycopersiconesculentum
(Tomato)
Cd5.8-M
T95
Echinodermata:
Strongylocentrotuspurpuratus
(Sea
urchin)
Cd7-M
TA
[(Cd4)�(C
d3)�–MTA]62,97

Fucusvesiculosus(A
lgae)
Cd7-M
T;Cu13–MT96
Tritricum
aestium
(Wheat)
Zn5-M
T98
Crustacea:a
Vicia
fava
(Broadbean)
Zea
mays(M
aize)
Cd1.7,Cu,Zn0.1-M
T1a99
Cd2,Cu0.4,Zn0.02-M
T1b99
Cd2,Cu0.4,Zn0.1-M
T299
Cd8-M
T100
Homarusamericanus
(Americanlobster)
Cd3-��MTH;Cd3-��MTH
101,102
MII6- þ
MII7-M
TH
(M¼Zn,Cd)26
MII3- þ
MII4-��;
MII3- þ
MII4-��26
Cu9,Cd6-M
T1;103
Cd9,Cu2�x,Znx-M
T21
03
Cd5,Cu–;Cd6-M
T21
01
Arabidopsisthaliana
Cu8.4-M
T1,Cu7.3-M
T2,
Cu5.5-M
T3104
Cancerpagurus
MII6-þM
II7-M
T(M¼Cd,Co)105
Protozoa:
Tetrahymenapigmentosa
Cd12-M
T1;Cd12-M
T2106
Cu7-M
T107
Callinectessapidus(Bluecrab)
Scillaserrata
(Mudcrab)
Cd6-M
TI6
2
Cd6-M
TI;
Cd6-M
TII
108
Cd3.1–3.7,Zn1.6–1.8-M
T109
Tetrahymenapyriform
isCd12-M
T1;Cd12-M
T2106
Fishes:
Annelida:
Notothenia
coriiceps
(Antarcticfish)
MII7-M
TA
(M¼Zn,Cd)110
Lumbricusrubellus
Cd6-w
MT-1;Cd6-w
MT-2
31
Aves:
Eisenia
foetida
Cd4-M
T62,111
Gallusgallus(C
hicken)
Cd9�x,Znx-M
T;Cu15-M
T112
aIn
theseorganismstheM
II3(SCys)9cluster
intheN-terminal�domain
isvariouslydenotedas�Nor��.Analogously,theM
II3(SCys)9cluster
intheC-terminal�domain
isdenotedas�Cor��.

They also evidenced an astonishing property of MTs, namely, the flexibility of the polypeptidechain and the dynamic fluctuation of the metal ions within the clusters.120
The 3D NMR solution structure of Cd7 -MT-1 from recombinant mouse has recently beenreported.121 It shows the same metal-to-Cys connectivities and the same structural organization ofclusters as those found in Cd7-MT-2. Therefore, all currently structurally characterized mammalianMT-1 and MT-2 containing either ZnII or CdII ions122 share common structural features. Theyare monomeric, dumb-bell-like shaped proteins with uniformly sized and almost sphericalC-terminal �- and N-terminal �-domains. The �-domain is composed of amino acid residues 31–61and enfolds the M4(SCys)11 aggregate, while the �-domain includes residues 1–30 and harborsthe M3(SCys)9 cluster, both being connected by the conserved Lys30–Lys31 segment. The absenceof inter-domain constraints in all the reported NMR structures of mammalian MTs has beenattributed to an increased flexibility in the linker region between the two domains of MT insolution, as compared with the crystal structure of rat liver MT-2.44 In each domain, all the metalcenters are tetrahedrally coordinated to both terminal and �2-bridging thiolate sulfur ligands(Figure 1), the central core of the metal–sulfur aggregates being formed by two fused M3(�-S)3rings (�-domain) or by one M3(�-S)3 ring (�-domain), whose conformation is commented onbelow.
Relevant information is also available on the structure of the mammalian isoform MT-3,123
having unique functional features when compared to MT-1 and 2.8 Solution NMR ofthe �-domain of Cd7-MT-3, which consists of the amino acid residues Lys32–Gln68, has revealeda tertiary fold that is very similar to MT-1 and 2, except for the Lys52–Glu60 loop thataccommodates an acidic insertion relative to these isoforms. The Cd-to-Cys connectivities pro-posed for this domain compare well to those in the Cd4(SCys)11 aggregates of the other twoisoforms. With respect to the �-domain of MT-3, NMR-detectable dynamic processes have
Table 2 Structurally characterized MTs and corresponding amino acid sequences.a,b
Mouse Cd7-MT-1121 (1DFT, 1DFS) MDPNCSCSTG GSCTCTSSCACKNCKCTSCK KSCCSCCPVGCSKCAQGCVC KGAADKCTCC A
Human Cd7-MT-2118 (1MHU, 2MHU) MDPNCSCAAG DSCTCAGSCKCKECKCTSCK KSCCSCCPVGCAKCAQGCIC KGASDKCSCC A
Rabbit Cd7-MT-2116 (1MRB, 2MRB) MDPNCSCAAA GDSCTCANSCTCKACKCTSC KKSCCSCCPPGCAKCAQGCICKGASDKCSCCA
Rat Cd5Zn2-MT-2 andCd7-MT-2117,119
(4MT2 and 1MRT,2MRT)
MDPNCSCATD GSCSCAGSCKCKQCKCTSCK KSCCSCCPVGCAKCSQGCIC KEASDKCSCC A
Mouse Cd7-MT-3c,123 (1JI9) MDPETCPCPT GGSCTCSDKCKCKGCKCTNC KKSCCSCCPAGCEKCAKDCV CKGEEGAKAEAEKCSCCQ
Blue crab Cd6-MT-1125 (1DMC, 1DME) MPGPCCNDKC VCQEGGCKAGCQCTSCRCSP CQKCTSGCKC
ATKEECSKTC TKPCSCCPKAmerican lobster Cd6–MT–1102,126 (1HZQ, 1HZR) PGPCCKDKCE CAEGGCKTGC
KCTSCRCAPC EKCTSGCKCPSKDECAKTCS KPCSCCXX
Sea urchin Cd7-MTA124 (1QJK, 1QJL) MPDVKCVCCK EGKECACFGQDCCKTGECCK DGTCCGICTNAACKCANGCK CGSGCSCTEGNCAC
Synechococcus Zn4-SmtA71 (1JJD) MTSTTLVKCA CEPCLCNVDPSKAIDRNGLY YCSEACADGHTGGSKGCGHT GCNCHG
S. cerevisiae Ag7-MT andCu7-MT84,85
(1AOO and 1AQR,d
1FMYd)QNEGHECQCQ CGSCKNNEQC
QKSCSCPTGC NSDDKCPCGNKSEETKKSCC SGK
a See text for details. b Protein Data Bank ID codes are given in parenthesis (www.rcsb.org). c Only the 3D structure of the �domain. d Only the polypeptide fold.
218 Metallothioneins

hindered its structure determination. However, models calculated by homology modeling areconsistent with this domain having the same backbone fold as MT-1 and 2.
Complete 3D solution structures have been determined by NMR experiments for three inverte-brate CdII-MT forms, one from echinoidean and two from crustacean species. The structure ofthe sea urchin (S. purpuratus) 113Cd7-MTA consists of two globular domains, an N-terminalCd4(SCys)11 and a C-terminal Cd3(SCys)9, whose metal cluster topology compares well with thecorresponding analogues in mammalian MT. Combined with this similarity, there is an invertedarrangement of the three and four metal clusters, and thus of the � and � domains, as well asa significantly different connectivity pattern for the Cys–metal coordination bonds in the twoproteins.124
Crustacean MTs differ from the above-mentioned structures in that they only have 18 Cysresidues within their sequences (Table 2). The blue crab (C. sapidus) 113Cd6-MT-1, whose solution
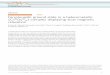
(a)
(b)
(c) (d)
Figure 1 (see color plate 14) The three-dimensional structure of various MTs: (a) Synechococcus Zn4-SmtA;(b) rat liver Cd5,Zn2-MT-2; (c) and (d), the isolated Cd3 �N and Cd3 �C domains of American lobster MTH,respectively. Zinc and cadmium ions are shown as blue and orange spheres, respectively. (Adapted from
entries made to the Protein Data Bank, ID codes given in Table 2.)
Metallothioneins 219

structure determination is the first reported for MTs from a non mammalian source, has twoseparate domains, each containing a Cd3(SCys)9 cluster with three bridging and six terminalthiolate ligands. The metal-to-Cys stoichiometry and the MII
3(�-S)3 ring framework in bothclusters compare well with those in the mammalian N-terminal � domain. In particular, thetertiary structure of crab MT-1 N-terminal � domain bears a close resemblance to the samedomain in mammalian MT-2.125 Previous studies on 111Cd6-MT-1 from lobster (H. americanus),which allowed only the metal-cluster topology to be characterized, had already confirmed that theprotein consists of two Cd3(SCys)9 clusters and established the corresponding Cd–SCys connect-ivities.126 This information has recently been extended to the full structure determination of thetwo isolated chemically synthesized 113Cd3 � domains from the same protein102 (Figure 1).
The recent NMR structure of the bacterial MT SmtA, synthesized by the cyanobacteriumSynechococcus PCC 7942, has provided evidence for the first metal core in MT where the Cysresidues are not uniquely responsible for metal coordination.71 In addition, the inorganic core ofthe Zn4(SCys)9(NHis)2 cluster in SmtA (Figure 1) strongly resembles the Zn4(SCys)11 cluster ofmammalian MT, despite the lack of sequence similarity and the presence of His ligands. Anotherinteresting finding in SmtA is the topology surrounding one of the Zn sites, whose unprecedentedelements of secondary structure in MTs resemble the zinc finger portions of GATA and LIMproteins.
The only reported structure for a copper metallothionein, also unique for an Ag-substitutedderivative, is for yeast (S. cerevisae), which includes 12 Cys and 1 His out of the 53 amino acidresidues. The NMR solution structure of both forms allowed determination of the backbone foldfor the first 40 residues of the protein. In addition, based on the fact that only seven major AgI
resonances were observed for the reconstituted protein, the Ag-to-Cys connectivities were derivedand assumed identical for Cu-MT. Overall, it was concluded that yeast binds seven CuI or AgI
ions in a single cluster, using 10 out of the 12 available Cys residues.84,86 A recent NMRreinvestigation of Cu7-MT from the same species has shown that a number of different arrange-ments of the seven copper ions is consistent with the highly refined structure of the polypeptidefold. This, together with the different coordination preferences of CuI and AgI with thiolate sulfurligands (see below), casts doubt on the isostructurality of both metalloforms in yeast MT.85
Existing data on phytochelatins and related polypeptides show that they differ from MT notonly in biosynthesis but also in structure. Whereas no 3D structure has yet been reported,electronic spectroscopy (c.d. and UV-Vis) and EXAFS data on Cd-�-EC peptides led to differentproposals as to the nature of the cadmium binding in such peptides, polynuclear cadmiumthiolate aggregates versus isolated Cd(SCys)4 units, respectively.9 However, contrary to theconclusion of the previous work, a recent X-ray absorption spectroscopy study on Cd-�-ECpeptides and model systems provides evidence for the formation of a polynuclear cadmiumcluster.67
8.9.1.4 Metal Thiolates as Models of the Metal–Sulfur Aggregates in Metallothioneins
Present structural and spectroscopic data on the metal coordination in MTs indicate that metalbinding is almost exclusively due to the cysteinyl thiolate sulfur atoms of the peptide chain: thecorresponding metal aggregates should therefore find a parallel in metal-thiolate chemistry. Theonly exception to this is the bacterial SmtA, in which it might be considered that two histidineresidues play the role of two terminal cysteine ligands.71 In fact, interest in the extensive family ofmetal thiolates has mainly been fuelled in recent decades by structural or bioinorganic objec-tives,127,128 the high cysteine content in MTs serving as an important impetus.
Although binary transition metal thiolates [M(SR)n], most frequently of polymeric nature,have been long known, it is since the 1970s that significant advances have been made in thesynthesis and structural chemistry of discrete thiolate complexes of general formula [Mx(SR)y]
z.127
Different synthetic approaches, such as the use of thiol ligands (RSH) including a low-coordin-ating solubilizing group (i.e., amine function) or sterically demanding substituents—or alternatively,of chelating dithiol ligands—have been most successful for the structural characterization of metalthiolate complexes of intermediate nuclearity (2� x� 14). The investigation of mononuclear andpolymeric complexes is more difficult because of the high solubility of the former and the well-known insolubility of the latter in common solvents. However, while the structural patterns foundin [Mx(SR)y]
z complexes have been systematized,127 it is still difficult to predict the stoichiometryand the stereochemistry around the metal centers in the complexes obtained at a given metal-to-thiol
220 Metallothioneins

molar ratio. The diverse coordination preferences of metal ions and their coexistence in the samecomplex species, the various ligation modes of the thiolate ligands (i.e., terminal, �2- or �3-bridging) and the possibility of secondary metal–sulfur interactions account for the main difficul-ties encountered in the synthesis of a particular complex. Overall, the search for synthetic modelsof metal–sulfur aggregates in MTs, which should facilitate knowledge of their chemical andspectroscopic features, is hampered by the inherent difficulties of metal-thiolate chemistry.
With respect to the metal–sulfur aggregates in MTs, it is remarkable that, despite the significantdifferences in amino acid composition and in the number and location of cysteine residues amongthe structurally characterized MTs (Table 2), only two basic types of ZnII and/or CdII metal–sulfur aggregates have been found: M3(SCys)9 and M4(SCys)11. As previously indicated, the lattercould also represent the M4(SCys)9(NHis)2 cluster found in SmtA. Surprisingly, among theextensive family of homoleptic thiolate complexes, those of formula [M3(SR)9]
3�
and [M4(SR)11]3� (M¼Zn or Cd) are unreported not only in the solid phase, according to the
Cambridge Structural Database (CSD),129 but also in solution.130 On the other hand, structuralinformation on CuI MTs is limited to the solution NMR studies of Cu7-MT from yeast. Aspreviously commented upon, these studies assumed that this metalloform was isostructural withthe 107Ag-substituted protein.84,131 Overall, both issues—the lack of synthetic models for themetal(II)–sulfur aggregates in MT and the recently questioned2,85 suitability of the 107Ag isotopeas a probe for the study of Cu-MTs—deserve further examination.
The steric arrangement of the ZnII and/or CdII ions and bridging thiolate sulfur ligands in theM3(SCys)9 and M4(SCys)11 clusters of MT can be described according to the six torsion anglesthat define the conformation of a six-membered ring (Figure 2).132 These values in structurallycharacterized MTs, obtained from Protein Data Bank (PDB) data (Table 2), show that most ofthe cyclohexane-like M3S3 rings in the M3(SCys)9 clusters have a distorted boat conformation,except for the �N and �C domains in lobster MTH, which show a sofa and a twist-boatconformation, respectively.102 Moreover, the bicyclo[3.3.1]nonane-like cluster in M4(SCys)11may be considered as being formed by two-fused six-membered M3S3 rings in twist-boat/sofaconformations (Figure 3), except for two cases where it is boat/twist-boat71 or boat/chair.124
This predominance of boat-type conformations in MTs contrasts with the chair-type usuallyfound in metal thiolate complexes with simple thiolate ligands and tetrahedrally coordinated Znor Cd.128 Thus, a search of the CSD129 on synthetic models of the M3(SCys)9 cluster reveals thatthe M3
II(�-S)3 ring is mainly found as a fragment of polynuclear metal thiolates with adaman-tanoid structure (19 out of 28 examples), and therefore in chair conformation. In a fewcomplexes, the chelating nature of the thiolate ligand or the high nuclearity of the complexspecies accounts for the sofa or boat conformation of the fragment mentioned; overall, however,no structurally related synthetic model of the M3(SCys)9 cluster has been reported. Analogously,for the M4(SCys)11 cluster, most of the metal thiolate complexes (17 out of 24) contain theM4
II(�–S)5 unit as a fragment of adamantanoid structures, where the two-fused M3S3 rings arein chair conformations.133,134 By extending the search to species containing the same unit butreplacing the thiolate sulfur that belongs to both of the two-fused M3S3 rings by either a sulfideor a halide ion, complexes with the core [M8(�4-X) (SR)16]
135–139 and [S4M10(SPh)16]4�,140
M¼Zn, Cd; X¼ S, halide, can be included. In all these examples, the two-fused rings showthe same conformation, i.e., twist-boat/twist-boat, as opposed to the twist-boat/sofa conform-ations in the M4(SCys)11 clusters of MTs (Figure 3). Overall, the different conformationalpreferences shown by the M3
IIS3 rings in the M3(SCys)9 and M4(SCys)11 clusters of MTs andin the inorganic metal thiolates emphasize the special role of the peptide chain enfolding themetal-thiolate cluster. Notwithstanding this, the presence of only two basic types of metal–sulfuraggregates in evolutionarily distant MTs emphasizes the functional relevance of the M3(SCys)9and M4(SCys)11 structural motifs.
An analysis of the data deposited on the CSD129 for the structurally characterized homolepticthiolate complexes of general formula Mx
I(SR)y, M¼Ag or Cu, allows the following conclusions
Boat (C2v) Sofa(Cs) Chair(D3d) Half-chair(C2) Twist boat(D2)
(52,0,-52,52,0,-52) (7,28,-62,62,-28,-7) (54,-54,54,-54,54,-54) (12,8,-47,67,-47,8) (30,-63,30,30,-63,30)
Figure 2 Interconversion between the conformational isomers of a six-membered ring with indication ofthe corresponding symmetry and set of torsion angles.
Metallothioneins 221

to be drawn: (i) the number of thiolates with known structure is significantly higher for Cu thanfor Ag, this difference being mainly due to the high number of Cu4
I(SR)6 species; (ii) thecoordination geometry around copper is exclusively trigonal in more than 50% of the complexes;in the rest it is either linear or there is coexistence of both trigonal and linear (however, there is noexample of tetrahedral coordination); (iii) the coordination geometry around silver is exclusivelylinear in 50% of the complexes, followed by the coexistence of different combinations of linear,trigonal, and tetrahedral geometries; (iv) unlike for copper, silver thiolates show a tendency toform polymeric species141 and secondary metal–sulfur interactions;142 and (v) copper and silvercomplexes with the same thiolate ligand may have different formulae143,144 and, even with thesame formula, may show a different structure142,145,146 as is shown in Figure 4. All these datastrongly support the uncertainty surrounding the notion of Cu/Ag isomorphous replacement inMTs.
(a)
(b)
Figure 3 Comparison of: (a) the Cd4(SCys)11 cluster of the � domain of rat liver Cd5,Zn2-MT (Figure 1) with(b) a fragment (darkest shading) of the cage skeleton of the [ClCd8{SCH(CH2CH2)2N(H)Me}16]
15þ cation.135
The two fused Cd3S3 rings (see text) show twist-boat (left)/sofa (right) conformations in the M4(SCys)11 clusterof MT and twist-boat/twist-boat conformations in the cation.
222 Metallothioneins

8.9.1.5 Dynamic Aspects and Reactivity
As research continues to seek out the physiological functions of MTs, their unusual dynamic
properties acquire greater significance.2,44,123 These were evidenced from 2D NMR117 and crystal-lographic119 studies on mammalian MII
7-MT-2, where the calculated root-mean-square deviation(rmsd) values and the B-factors, respectively, showed a significant degree of dynamic structuraldisorder. Present data on MTs from different sources indicate that they are flexible moleculesundergoing structural fluctuations within the metal clusters, as well as involving the polypeptideloops between the cysteine residues of the protein chain. Detailed analysis of the structural
(a)
(b)
Figure 4 Comparison of copper and silver thiolate complexes of the same thiolate ligand and with the sameformula unit. Perspective view of: (a) [Cu(SC6H4-o-SiMe3)]12 and (b) [{Ag(SC6H4-o-SiMe3)}4]2, highlighting
the Cu12S12 core and the fusing of two Ag4S4 units via Ag � � � S secondary interactions (3.05 A).142
Metallothioneins 223

flexibility and reactivity of MTs2 and the study of the dynamics of interdomain and inter-molecular interactions in mammalian MT have recently been reported.44
Structural non rigidity is often manifested in the MxSy cores of [Mx(SR)y]z aggregates.127 The
degree of mechanical flexibility in these [Mx(SR)y]z species is inversely related to the connectiv-
ity, i.e., the number of framework M—S bonds, and increases with the size of the Mn(SR)n ringsthat constitute the aggregate. As a result, inherently floppy [Mx(SR)y]
z complexes give rise toconfigurational isomers (those which can be distinguished in terms of the relative orientation ofbonds)147 as has been crystallographically observed for the [MI
5(SR)6]z complexes with M¼Cu
(z¼�1),148 and Ag (z¼ 2)149 (Figure 5). Furthermore, in the case of metal thiolate complexeswith an adamantane-like cluster structure, a mechanism for the intramolecular interconversion ofterminal and bridging thiolate ligands has been proposed.150 In addition to these dynamicfeatures, the group 12 metal thiolates in solution show kinetic lability and (consequently)continuous breaking and re-forming of the metal–sulfur bonds.65 By extending these featuresof metal thiolate complexes to MTs, it could be concluded that both the high structuralflexibility and the kinetic lability of the MxSy cores in the [Mx(SCys)y] aggregates shouldsubstantially contribute to the dynamic properties of the entire protein. In other words, itseems likely that the metal-thiolate nature of the metal clusters in MTs is not only responsiblefor the unusual dynamic features of their metal cores when compared to other metalloproteins,but also for the mobility of the peptide chain, whose folding is essentially determined by thebinding of the cysteine residues to the metal centers.
In all the mammalian MT structures characterized to date, the amino-terminal �-domainshave been found to be more flexible than the carboxy-terminal �-domains.123 Similarly, thestructure of the backbone in the �-domain of the sea urchin MTA is less rigid than its �counterpart.124 This enhanced flexibility of the �-domain in mammalian MT and in MTA isconsistent with the predictions operating in [Mx(SR)y]
z species,127 where a greater degree ofmechanical flexibility should be expected for an M3S3 fragment consisting of six core atoms,six bonds, and one M3(SR)3 cycle than that for M4S5 with nine core atoms, ten bonds, and two-fused M3(SR)3 cycles. However, the strikingly different flexibility encountered in the �-domain ofmammalian MTs (MT-3�MT-2>MT- 1),123 the different kinetic stability of the two �-domainsin lobster MTH15 and the enhanced flexibility of cold-adapted MT as compared to mouse MT151
reveal—among other examples—the special role of the peptide chain in tuning the chemical andstructural features of the metal–sulfur aggregates.
MTs are unusually reactive metalloproteins, in which the high thermodynamic stability of themetal–sulfur bonds is coupled to a high kinetic lability. The latter accounts for the easyexchange of metal ions within the metal clusters, with metal ions in solution and also with
(a) (b) (c)
Figure 5 Perspective view of the metal–sulfur core in [M5(SR)6]z complexes: (a) [Ag5{�-S(CH2)3-
NHMe2}3{�-S(CH2)3NMe2}3](ClO4)2,149 (b) (Et4N)[Cu5(�-SBu
t)6] and (c) (Et3NH)[Cu5(�-SBut)6],
148 andcorresponding views along the Mtrig–Mtrig axis, showing the nonrigidity of the M5S6 cages. Carbon atoms
are shown as gray spheres.
224 Metallothioneins

metal ions in clusters of other MT molecules.2,44,65 The thermodynamic stability dictates theaffinity of the metal ions for the binding sites as well as the order of their replacement in MTs,which is consistent with that found for inorganic thiolates: HgII>AgI�CuI>CdII>ZnII.2 Inaddition to the metal binding and exchange processes, the removal of the metal ions in MTs bycompeting ligands has also been analyzed with the aim of elucidating the interactions of MTswith cellular components and the role of Zn-MT as an intracellular metal donor.152,153 In thiscontext, i.e., the putative cellular chemistry of MTs, the metal transfer between MTs andenzymes,154,155 and the nucleophilic reactivity of the coordinated cysteines toward electrophiles,including alkylating and oxidizing agents153 and radical species,156–162 are the object of intensestudy. In summary, current experimental data show that both the inherent kinetic lability of themetal thiolates and the nucleophilicity of the thiolate sulfur account for the chemical reactivity ofthe MTs. The influence of the peptide chain on the control and modulation of such propertiesis probably essential for the still undisclosed functional roles of these unique metalloproteins.
8.9.2 REFERENCES
1. Margoshes, M.; Vallee, B. L. J. Am. Chem. Soc. 1957, 79, 4813–4814.2. Romero-Isart, N.; Vasak, M. J. Inorg. Biochem. 2002, 88, 388–396.3. Kagi, J. H. R.; Nordberg, M., Eds. Metallothionein I; Experientia Suppl.; Birkhauser Verlag: Basel, 1979; Vol. 34.4. Kagi, J. H. R.; Kojima, Y., Eds. Metallothionein II; Experientia Suppl.; Birkhauser Verlag: Basel, 1987; Vol. 52.5. Suzuki, K. T.; Imura, N.; Kimura, M., Eds. Metallothionein III. Biological roles and medical implications; Birkhauser
Verlag: Basel, 1993.6. Klaasen, C. D, Ed. Metallothionein IV; Birkhauser Verlag: Basel, 1999.7. Riordan, J. F.; Vallee, B. L., Eds. Methods in Enzymology; Academic Press: San Diego, 1991; Vol. 205.8. Vasak, M.; Hasler, D. W. Curr. Opin. Chem. Biol. 2000, 4, 177–183.9. Vasak, M.; Kagi, J. H. R. Metallothioneins. In Encyclopedia of Inorganic Chemistry; Bruce King, R., Ed., Wiley:
Chichester, UK, 1994; Vol. 4, pp 2229–2241.10. Stillman, M. J. Coord. Chem. Rev. 1995, 144, 461–511.11. Stillman, M. J.; Shaw III, C. F.; Suzuki, K. T., Eds. Metallothioneins; VCH: New York, 1992.12. Coyle, P.; Philcox, J. C.; Carey, L. C.; Rofe, A. M. Cell. Mol. Life Sci. 2002, 59, 627–647.13. Kull, F. J.; Reed, M. F.; Elgren, T. E.; Ciardelli, T. L.; Wilcox, D. E. J. Am. Chem. Soc. 1990, 112, 2291–2298.14. Nishiyama, Y.; Nakayama, S.; Okada, Y.; Min, K.-S.; Onosaka, S.; Tanaka, K. Chem. Pharm. Bull. 1990, 38,
2112–2117.15. Munoz, A.; Petering, D. H.; Shaw III, C. F. Inorg. Chem. 2000, 39, 6114–6123.16. Okada, Y.; Ohta, N.; Iguchi, S.; Tsuda, Y.; Sasaki, H.; Kitagawa, T.; Yagyu, M.; Min, K.-S.; Onosaka, S.; Tanaka, K.
Chem. Pharm. Bull. 1986, 34, 986–998.17. Hartmann, H.-J.; Li, Y.-J.; Weser, U. Biometals 1992, 5, 187–191.18. Li, Y.-J.; Weser, U. Inorg. Chem. 1992, 31, 5526–5533.19. Sewell, A. K.; Jensen, L. T.; Erickson, J. C.; Palmiter, R. D.; Winge, D. R. Biochemistry 1995, 34, 4740–4747.20. Okada, Y.; Tanaka, K.; Sawada, J.; Kikuchi, Y. Synthetic Metallothioneins. In Metallothioneins; Stillman, M. J.;
Shaw III, C. F.; Suzuki, K. T., Eds.; VCH: New York, 1992, Chapter 9, pp 195–225.21. Pan, P. K.; Zheng, Z. F.; Lyu, P. C.; Huang, P. C. Eur. J. Biochem. 1999, 266, 33–39.22. Morooka, Y.; Nagaoka, T. Appl. Environ. Microb. 1987, 53, 204–207.23. Hou, T. M.; Kim, R.; Kim, S. H. Biochim. Biophys. Acta 1988, 951, 230–234.24. Berka, T.; Shatzman, A.; Zimmerman, J.; Strickler, J.; Rosenberg, M. J. Bacteriol. 1988, 170, 21–26.25. Valls, M.; Bofill, R.; Romero-Isart, N.; Gonzalez-Duarte, R.; Abian, J.; Carrascal, M.; Gonzalez-Duarte, P.;
Capdevila, M.; Atrian, S. FEBS Lett. 2000, 467, 189–194.26. Valls, M.; Bofill, R.; Gonzalez-Duarte, R.; Gonzalez-Duarte, P.; Capdevila, M.; Atrian, S. J. Biol. Chem. 2001, 276,
32835–32843.27. Shi, J.; Lindsay, W. P.; Huckle, J. W.; Morby, A. P.; Robinson, N. J. FEBS Lett. 1992, 303, 159–163.28. Wang, Y.; Mackay, E. A.; Kurasaki, M.; Kagi, J. H. R. Eur. J. Biochem. 1994, 225, 449–457.29. Erickson, J. C.; Sewell, A. K.; Jensen, L. T.; Winge, D. R.; Palmiter, R. D. Brain Res. 1994, 649, 297–304.30. You, C.; Mackay, E. A.; Gehrig, P. M.; Hunziker, P. E.; Kagi, J. H. R. Arch. Biochem. Biophys. 1999, 372, 44–52.31. Sturzenbaum, S. R.; Winters, C.; Galay, M.; Morgan, A. J.; Kille, P. J. Biol. Chem. 2001, 276, 34013–34018.32. Cols, N.; Romero-Isart, N.; Bofill, R.; Capdevila, M.; Gonzalez-Duarte, P.; Gonzalez-Duarte, R.; Atrian, S. Prot.
Eng. 1999, 12, 265–269.33. Bofill, R.; Capdevila, M.; Cols, N.; Atrian, S.; Gonzalez-Duarte, P. J. Biol. Inorg. Chem. 2001, 6, 408–417.34. Capdevila, M.; Cols, N.; Romero-Isart, N.; Gonzalez-Duarte, R.; Atrian, S.; Gonzalez-Duarte, P. Cell. Mol. Life Sci.
1997, 53, 681–688.35. Kurasaki, M.; Emoto, T.; Linde Arias, A. R.; Okabe, M.; Yamasaki, F.; Oikawa, S.; Kojima, Y. Prot. Eng. 1996, 9,
1173–1180.36. Kurasaki, M.; Yamaguchi, R.; Linde Arias, A. R.; Okabe, M.; Kojima, Y. Prot. Eng. 1997, 10, 413–416.37. Thrower, A. R.; Byrd, J.; Tarbet, E. B.; Mehra, R. K.; Hamer, D. H.; Winge, D. R. J. Biol. Chem. 1988, 263,
7037–7042.38. Chernaik, M. L.; Huang, P. C. Proc. Natl. Acad. Sci. USA 1991, 88, 3024–3028.39. Emoto, T.; Kurasaki, M.; Oikawa, S.; Suzuki-Kurasaki, M.; Okabe, M.; Yamasaki, F.; Kojima, Y. Biochem. Genet.
1996, 34, 239–251.40. Pan, P. K.; Hou, F.; Cody, C. W.; Huang, P. C. Biochem. Biophys. Res. Commun. 1994, 202, 621–628.
Metallothioneins 225

41. Romero-Isart, N.; Cols, N.; Termansen, M. K.; Gelpi, J. L.; Gonzalez-Duarte, R.; Atrian, S.; Capdevila M.; Gonzalez-Duarte, P. Eur. J. Biochem. 1999, 259, 519–527.
42. Fowler, B. A.; Hildebrand, C. E.; Kojima, Y.; Webb, M. Nomenclature of Metallothionein. In Metallothionein II;Kagi, J. H. R.; Kojima, Y., Eds.; Experientia Suppl.; Birkhauser Verlag: Basel, 1987; Vol. 52, pp 19–22.
43. Klaassen, C. D.; Liu, J.; Choudhuri, S. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 267–294.44. Zangger, K.; Armitage, I. M. J. Inorg. Biochem. 2002, 88, 135–143.45. Cousins, R. J. Physiol. Rev. 1985, 65, 238–309.46. Bremner, I. Nutritional and Physiological Significance of Metallothionein. In Metallothionein II; Kagi, J. H. R.;
Kojima, Y, Eds.; Experientia Suppl., Birkhauser Verlag: Basel, 1987; Vol. 52, pp 81–107.47. Sato, M.; Bremner, I. Free Radic. Biol. Med. 1993, 14, 325–337.48. Kang, Y. J. Proc. Soc. Exp. Biol. Med. 1999, 222, 263–273.49. Hidalgo, J.; Aschner, M.; Zatta, P.; Vasak, M. Brain Res. Bull. 2001, 55, 133–145.50. Maret, W.; Vallee, B. L. Proc. Natl. Acad. Sci. USA 1998, 95, 3478–3482.51. Beattie, J. H.; Wood, A. M.; Newman, A. M.; Bremner, I.; Choo, K. H.; Michalska, A. E.; Duncan, J. S.; Trayhurn, P.
Proc. Natl. Acad. Sci. USA 1998, 358–363.52. Maret, W.; Heffron, G.; Hill, H. A.; Djuricic, D.; Jiang, L. J.; Vallee, B. L. Biochemistry 2002, 41, 1689–1694.53. Zangger, K.; Oz, G.; Armitage, I. M. J. Biol. Chem. 2000, 275, 7534–7538.54. Dallinger, R.; Berger, B.; Hunziker, P.; Kagi, J. H. R. Nature 1997, 338, 237–238.55. Bongers, J.; Walton, C. D.; Richardson, D. E.; Bell, J. U. Anal. Chem. 1988, 60, 2683–2686.56. Vasak, M.; Kagi, J. H. R.; Hill, H. A. O. Biochemistry 1981, 20, 2852–2856.57. Willner, H.; Vasak, M.; Kagi, J. H. R. J. Chem. Soc., Chem. Commun. 1987, 26, 6287–6292.58. Schaffer, A. Absorption, Circular Dichroism, and Magnetic Circular Dichroism Spectroscopy of Metallothionein.
In Methods in Enzymology, Riordan, J. F.; Vallee, B. L., Eds.; Academic Press: San Diego, 1991; Vol. 205,pp 529–540.
59. Jensen, L. T.; Peltier, J. M.; Winge, D. R. J. Biol. Inorg. Chem. 1998, 3, 627–631.60. Polec, K.; Palacios, O.; Capdevila, M.; Gonzalez-Duarte, P.; Lobinski, R. Talanta 2002, 57, 1011–1017.61. Hathout, Y.; Reynolds, K. J.; Szilagyi, Z.; Fenselau, C. J. Inorg. Biochem. 2002, 88, 119–122.62. Gehrig, P. M.; You, C.; Dallinger, R.; Gruber, C.; Brouwer, M.; Kagi, J. H. R.; Hunziker, P. E. Protein Science 2000,
9, 395–402.63. Yu, X.; Wojciechowski, M.; Fenselau, C. Anal. Chem. 1993, 65, 1355–1359.64. Zaia, J.; Fabris, D.; Wei, D.; Karpel, R. L.; Fenselau, C. Protein Science 1998, 7, 2398–2404.65. Kagi, J. H. R. Overview of Metallothionein. In Methods in Enzymology; Riordan, J. F.; Vallee, B. L., Eds.; Academic
Press: San Diego, 1991, Vol. 205, pp 613–626.66. Clark-Baldwin, K.; Tierney, D. L.; Govindaswamy, N.; Gruff, E. S.; Kim, C.; Berg, J.; Koch, S. A.; Penner-Hahn, J. E.
J. Am. Chem. Soc. 1998, 120, 8401–8409.67. Pickering, I. J.; Prince, R. C.; George, G. N.; Rauser, W. E.; Wickramasinghe, W. A.; Watson, A. A.; Dameron, C. T.;
Dance, I. G.; Fairlie, D. P.; Salt, D. E. Biochim. Biophys. Acta 1999, 1429, 351–364.68. Winge, D. R. Copper Coordination in Metallothionein. In Methods in Enzymology; Riordan, J. F.; Vallee, B. L., Eds.;
Academic Press: San Diego, 1991; Vol. 205, pp 458–469.69. Bogumil, R.; Faller, P.; Binz, P.-A.; Vasak, M.; Charnock, J. M.; Garner, D. Eur. J. Biochem. 1998, 255, 172–177.70. Nielson, K. B.; Atkin, C. L.; Winge, D. R. J. Biol. Chem. 1985, 260, 5342–5350.71. Blindauer, C. A.; Harrison, M. D.; Parkinson, J. A.; Robinson, A. K.; Cavet, J. S.; Robinson, N. J.; Sadler, P. J. Proc.
Natl. Acad. Sci. USA 2001, 98, 9593–9598.72. Beltramini, M.; Lerch, K. Environ. Health Perspect. 1986, 65, 21–27.73. Lerch, K. Nature 1980, 284, 368–370.74. Beltramini, M.; Lerch, K.; Vasak, M. Biochemistry 1984, 23, 3422–3427.75. Dallinger, R.; Wang, Y.; Berger, B.; Mackay, E. A.; Kagi, J. H. R. Eur. J. Biochem. 2001, 268, 4126–4133.76. Munger, K.; Lerch, K. Biochemistry 1985, 24, 6751–6756.77. Raspor, B.; Pavicic, J. Fresenius J. Anal. Chem. 1996, 354, 529–534.78. Prinz, R.; Weser, U. Hoppe-Seyler’s Z. Physiol. Chem. 1975, 356, 767–776.79. Sayers, Z.; Brouillon, P.; Svergun, D. I.; Zielenkiewicz, P.; Koch, M. H. J. Eur. J. Biochem. 1999, 262, 858–865.80. Weser, U.; Hartman, H.-J.; Fretzdorff, A.; Strobel, G.-J. Biochim. Biophys. Acta 1977, 493, 465–477.81. Winge, D. R.; Nielson, K. B.; Gray, W. R.; Hamer, D. H. J. Biol. Chem. 1985, 260, 14464–14469.82. Sayers, Z.; Brouillon, P.; Vorgias, C. E.; Nolting, H. F.; Hermes, C.; Koch, M. H. J. Eur. J. Biochem. 1993, 212,
521–528.83. Byrd, J.; Berger, R. M.; McMillin, D. R.; Wright, C. F.; Hammer, D.; Winge, D. R. J. Biol. Chem. 1988, 263,
6688–6694.84. Peterson, C. W.; Narula, S. S.; Armitage, I. M. FEBS Lett. 1996, 379, 85–93.85. Bertini, I.; Hartman, H.-J.; Klein, T.; Liu, G.; Luchinat, C.; Weser, U. Eur. J. Biochem. 2000, 267, 1008–1018.86. Narula, S. S.; Winge, D. R.; Armitage, I. M. Biochemistry 1993, 32, 6773–6787.87. Bordas, J.; Koch, M. H. J.; Hartmann, H.-J.; Weser, U. FEBS Lett. 1982, 140, 19–21.88. Ding, X.-Q.; Bill, E.; Trautwein, A. X.; Hartmann, H. J.; Weser, U. Eur. J. Biochem. 1994, 223, 841–845.89. Jensen, L. T.; Howard, W. R.; Strain, J. J.; Winge, D. R.; Culotta, V.C. J. Biol. Chem. 1996, 271, 18514–18519.90. Roesijadi, G.; Kielland, S.; Klersks, P. Arch. Biochem. Biophys. 1989, 273, 403–413.91. Mehra, R. K.; Garey, J. R.; Butt, T. R.; Gray, W. R.; Winge, D. R. J. Biol. Chem. 1989, 264, 19747–19753.92. Hensbergen, P. J.; van Velzen, M. J. M.; Nugroho, R. A.; Donker, M. H.; van Straalen, N. M. Comp. Biochem.
Physiol. C 2000, 125, 17–24.93. Kille, P.; Winge, D. R.; Harwood, J. L.; Kay, J. FEBS Lett 1991, 295, 171–175.94. Tommey, A. M.; Shi, J.; Lindsay, W. P.; Urwin, P. E.; Robinson, N. J. FEBS Lett. 1991, 292, 48–52.95. Rauser, W. E. The Cd-Binding Protein from Tomato Compared to Those of Other Vascular Plants. In Metallothionein
II; Kagi, J. H. R., Kojima, Y., Eds.; Experientia Suppl.; Birkhauser Verlag: Basel, 1987; Vol. 52, pp 301–308.96. Morris, C. A.; Nicolaus, B.; Sampson, V.; Harwood, J. L.; Kille, P. Biochem. J. 1999, 338, 553–560.97. Wang, Y.; Hess, D.; Hunzinker, P. E.; Kagi, J. H. R. Eur. J. Biochem. 1996, 241, 835–839.
226 Metallothioneins

98. Robinson, N. J.; Tommey, A. M.; Kuske, C.; Jackson, P. J. Biochem. J. 1993, 295, 1–10.99. Foley, R. C.; Liang, Z. M.; Singh, K. B. Plant Mol. Biol. 1997, 33, 583–591.100. Bernhard, W. R.; Kagi, J. H. R. Purification and Characterization of Atypical Cadmium-Binding Polypeptides from
ZeaMays. In Metallothionein II; Kagi, J. H. R., Kojima, Y., Eds.; Experientia Suppl.; Birkhauser Verlag: Basel, 1987;Vol. 52, pp 309–315.
101. Zhu, Z.; Goodrich, M.; Isab, A. A.; Shaw III, C. F. Inorg. Chem. 1992, 31, 1662–1667.102. Munoz, A.; Forsterling, F. H.; Shaw III, C. F.; Petering, D. H. J. Biol. Inorg. Chem. 2002, 7, 713–724.103. Chou, C. L.; Guy, R. D.; Uthe, J. F. Sci. Total Environ. 1991, 105, 41–59.104. Murphy, A.; Zhou, J.; Golsbrough, P. B.; Taiz, L. Plant Physiol. 1997, 113, 1293–1301.105. Overnell, J.; Good, M.; Vasak, M. Eur. J. Biochem. 1988, 172, 171–177.106. Piccinni, E.; Staudenmann, W.; Albergoni, V.; de Gabrieli, R.; James, P. Eur. J. Biochem. 1994, 226, 853–859.107. Santovito, G.; Irato, P.; Palermo, S.; Boldrin, F.; Sack, R.; Hunziker, P.; Piccinni, E. Protist 2001, 152, 219–229.108. Otvos, J. D.; Olafson, R. W.; Armitage, I. M. J. Biol. Chem. 1982, 257, 2427–2431.109. Law, A. C. Y.; Cherian, M. G.; Stillman, M. J. Biochim. Biophys. Acta 1984, 784, 53–61.110. D’Auria, S.; Carginale, V.; Scudiero, R.; Crescenzi, O.; di Maro, D.; Temussi, P. A.; Parisi, E.; Capasso, C. Biochem.
J. 2001, 354, 291–299.111. Gruber, C.; Sturzenbaum, S.; Gehrig, P.; Sack, R.; Hunziker, P.; Berger, B.; Dallinger, R. Eur. J. Biochem. 2000, 267,
573–582.112. Rupp, H.; Voelter, W.; Weser, U. Hoppe-Seyler’s Z. Physiol. Chem. 1975, 356, 755–765.113. Vallee, B. L.; Maret, W. The Functional Potential and Potential Functions of Metallothioneins: A Personal
Perspective. In Metallothionein III. Biological roles and medical implications; Suzuki, K. T.; Imura, N.; Kimura, M.,Eds.; Birkhauser Verlag: Basel, 1993, pp 1–9.
114. Otvos, J. D.; Armitage, I. M. Proc. Natl. Acad. Sci. USA 1980, 77, 7094–7098.115. Otvos, J. D.; Engeseth, H. R.; Wehrli, S. Biochemistry 1985, 24, 6735–6739.116. Arseniev, A.; Schultze, P.; Worgotter, E.; Braun, W.; Wagner, G.; Vasak, M.; Kagi, J. H. R.; Wuthrich, K. J. Mol.
Biol. 1988, 201, 637–657.117. Schultze, P.; Worgotter, E.; Braun, W.; Wagner, G.; Vasak, M.; Kagi, J. H. R.; Wuthrich, K. J. Mol. Biol. 1988, 203,
251–268.118. Messerle, B. A.; Schaffer, A.; Vasak, M.; Kagi, J. H. R.; Wuthrich, K. J. Mol. Biol. 1990, 214, 765–779.119. Robbins, A. H.; McRee, D. E.; Williamson, M.; Collett, S. A.; Xuong, N. H.; Furey, W. F.; Wang, B. C.; Stout, C. D.
J. Mol. Biol. 1991, 221, 1269–1293.120. Braun, W.; Vasak, M.; Robbins, A. H.; Stout, C. D.; Wagner, G.; Kagi, J. H. R.; Wuthrich, K. Proc. Natl. Acad. Sci.
USA 1992, 89, 10124–10128.121. Zangger, K.; Oz, G.; Otvos, J. D.; Armitage, I. M. Protein Sci. 1999, 8, 2630–2638.122. Messerle, B. A.; Schaffer, A.; Vasak, M.; Kagi, J. H. R.; Wuthrich, K. J. Mol. Biol. 1992, 225, 433–443.123. Oz, G.; Zangger, K; Armitage, I. M. Biochemistry 2001, 40, 11433–11441.124. Riek, R.; Precheur, B.; Wang, Y.; Mackay, E. A.; Wider, G.; Guntert, P.; Liu, A.; Kagi, J. H. R.; Wuthrich, K.
J. Mol. Biol. 1999, 291, 417–428.125. Narula, S. S.; Brouwer, M.; Hua, Y.; Armitage, I. M. Biochemistry 1995, 34, 620–631.126. Zhu, Z.; DeRose, E. F.; Mullen, G. P.; Petering, D. H.; Shaw III, C. F. Biochemistry 1994, 33, 8858–8865.127. Dance, I. G. Polyhedron 1986, 5, 1037–1104.128. Dance, I. G.; Fisher, K.; Lee, G. Metal–Thiolate Compounds. In Metallothioneins; Stillman, M. J.; Shaw III, C. F.;
Suzuki, K. T. Eds.; VCH: New York, 1992, Chapter 13, pp 284–345.129. Allen, F. H.; Kennard, O. Chem. Des. Autom. News 1993, 8, 31–37.130. Gonzalez-Duarte, P.; Vives, J. Inorg. Chem. 1989, 28, 25–30.131. Narula, S. S.; Mehra, R. K.; Winge, D. R.; Armitage, I. M. J. Am. Chem. Soc. 1991, 113, 9354–9358.132. Dale, J. Stereochemistry and Conformational Analysis; Verlag Chemie: New York, 1978, Chapter 5, pp 147–153.133. Dean, P. A. W.; Vittal, J. J. Adamantane-like Cages. In Metallothioneins; Stillman, M. J.; Shaw III, C. F.; Suzuki, K. T.,
Eds.; VCH: New York, 1992, Chapter 14, , pp 346–386.134. Vittal, J. J. Polyhedron 1996, 15, 1585–1642.135. Gonzalez-Duarte, P.; Clegg, W.; Casals, I.; Sola, J.; Rius, J. J. Am. Chem. Soc. 1998, 120, 1260–1266.136. Dance, I. G. Aust. J. Chem. 1985, 38, 1391–1411.137. Guo, S.; Ding, E.; Chen, H.; Yin, Y.; Li, X. Polyhedron 1999, 18, 735–740.138. Burth, R.; Gelinsky, M.; Vahrenkamp, H. Inorg. Chem. 1998, 37, 2833–2836.139. Ka-Luo, T.; Xiang-Lin, J.; Shao-Juan, J.; You-Qi, T. Chin. J. Struct. Chem. 1995, 14, 399–404.140. Lee, G. S. H.; Fisher, K. J.; Craig, D. C.; Scudder, M. L.; Dance, I. G. J. Am. Chem. Soc. 1990, 112, 6435–6437.141. Casals, I.; Gonzalez-Duarte, P.; Sola, J.; Vives, J.; Font-Bardia, M.; Solans, X. Polyhedron 1990, 9, 769–771.142. Block, E.; Gernon, M.; Kang, H.; Ofori-Okai, G.; Zubieta, J. Inorg. Chem. 1989, 28, 1263–1271.143. Tang, K.; Aslam, M.; Block, E.; Nicholson, T.; Zubieta, J. Inorg. Chem. 1987, 26, 1488–1497.144. Ka-Luo, T.; Qingchuan, Y.; Jianping, Y.; You-Qi, T. Acta Sci. Nat. Univ. Pek. 1988, 24, 398–403.145. Tang, K.; Yang, J.; Yang, Q.; Tang, Y. J. Chem. Soc., Dalton Trans. 1989, 2297–2302.146. Schroter-Schmid, I.; Strahle, J. Z. Naturforsch., Teil B 1990, 45, 1537–1542.147. Harrowfield, J. M.; Wild, S. B. Isomerism in Coordination Chemistry. In Comprehensive Coordination Chemistry,
1st ed.; Wilkinson, G., Gillard, R. D., McCleverty, J. A., Eds.; Pergamon Press: New York, 1987; Vol. 1, pp 179–208.148. Bowmaker, G. A.; Clark, G. R.; Seadon, J. K.; Dance, I. G. Polyhedron 1984, 3, 535–544.149. Gonzalez-Duarte, P.; Sola, J.; Vives, J.; Solans, X. J. Chem. Soc., Chem. Commun. 1987, 1641–1642.150. Hagen, K. S.; Stephan, D. W.; Holm, R. H. Inorg. Chem. 1982, 21, 3928–3936.151. Capasso, C.; Abugo, O.; Tanfani, F.; Scire, A.; Carginale, V.; Scudiero, R.; Parisi, E.; D’Auria, S. Proteins 2002, 46,
259–267.152. Petering, D. H.; Krezoski, S.; Chen, P.; Pattanaik, A.; Shaw III, C. F. Kinetic Reactivity of Metallothionein. In
Metallothioneins; Stillman, M. J.; Shaw III, C. F.; Suzuki, K. T., Eds.; VCH: New York, 1992, Chapter 7, pp 164–185.153. Shaw III, C. F.; Savas, M. M.; Petering, D. H. Ligand Substitution and Sulfhydryl Reactivity of Metallothionein. In
Methods in Enzymology; Riordan, J. F., Vallee, B. L., Eds.; Academic Press: San Diego, 1991; Vol. 205, pp 401–419.
Metallothioneins 227

154. Jacob, C.; Maret, W.; Vallee, B. L. Proc. Natl. Acad. Sci. USA 1998, 95, 3489–3494.155. Maret, W.; Jacob, C.; Vallee, B. L.; Fisher, E. H. Proc. Natl. Acad. Sci. USA 1999, 96, 1936–1940.156. Thornalley, P. J.; Vasak, M. Biochim. Biophys. Acta 1985, 827, 36–44.157. Felix, K; Lengfelder, E.; Hartmann, H.-J.; Weser, U. Biochim. Biophys. Acta 1993, 1203, 104–108.158. Kroncke, K.-D.; Fehsel, K.; Schmidt, T.; Zenke, F. T.; Dasting, I.; Wesener, J. R.; Bettermann, H.; Breuing, K. D.;
Kolb-Bachofen, V. Biochem. Biophys. Res. Commun. 1994, 200, 1105–1110.159. Aravindakumar, C. T.; Ceulemans, J.; de Ley, M. Biochem. J. 1999, 344, 253–258.160. Hartmann, H.-J.; Weser, U. Biometals 2000, 13, 153–156.161. Montoliu, C.; Monfort, P.; Carrasco, J.; Palacios, O.; Capdevila, M.; Hidalgo, J.; Felipo, V. J. Neurochem. 2000, 75,
266–273.162. Warthen, C. R.; Hammes, B. S.; Carrano, C. J.; Crans, D. C. J. Biol. Inorg. Chem. 2001, 6, 82–90.
# 2003, Elsevier Ltd. All Rights ReservedNo part of this publication may be reproduced, stored in any retrieval system ortransmitted in any form or by any means electronic, electrostatic, magnetic tape,mechanical, photocopying, recording or otherwise, without permission in writingfrom the publishers
Comprehensive Coordination Chemistry IIISBN (set): 0-08-0437486
Volume 8, (ISBN 0-08-0443303); pp 213–228
228 Metallothioneins