Embed Size (px)
Citation preview

Biomedicine & Pharmacotherapy 67 (2013) 58–65
Original article
6r, a novel oxadiazole analogue of ethacrynic acid, exhibits antitumor activityboth in vitro and in vivo by induction of cell apoptosis and S-phase arrest
Peng Zhang a, Jin-Hua Chen a, Xue Dong a, Ming-Tan Tang a, Li-Yan Gao a, Gui-Sen Zhao b,*,Lu-Gang Yu c, Xiu-Li Guo a,**a Department of Pharmacology, Key Laboratory of Chemical Biology (Ministry of Education), School of Pharmaceutical Sciences, Shandong University, No. 44, WenHuaXi Road,
250012 Jinan, PR Chinab Department of Medicinal Chemistry, School of Pharmaceutical Sciences, Shandong University, No. 44, WenHuaXi Road, 250012 Jinan, PR Chinac Department of Gastroenterology, Institute of Translational Medicine, University of Liverpool, L69 3BX Liverpool, United Kingdom
A R T I C L E I N F O
Article history:
Received 6 September 2012
Accepted 22 October 2012
Keywords:
5-[2,3-Dichloro-4-(2-methylene-1-
oxobutyl) phenoxymethyl]-
3-methyl-1,2,4- oxadiazole (6r)
Human cancer cell lines
Glutathione S-transferase P1-1
Apoptosis
Cell cycle
A B S T R A C T
This study investigated the in vitro and in vivo antitumor effects of 5-[2,3-Dichloro-4-(2-methylene-1-
oxobutyl) phenoxymethyl]-3-methyl-1,2,4- oxadiazole (6r), a novel ethacrynic acid (EA) derivative. The
in vitro effect of 6r on cell proliferation of human colon, leukemia, prostate, lung, breast, ovarian and
cervical tumor cell lines was assessed using MTT assay and the in vivo effect was determined with an
SW620 xenografts nude mice model. The effect of 6r on expressions of GST P1-1 and apoptosis-related
proteins were measured by western blotting and the effect on cell apoptosis was analysed by Hoechst
33258 nuclear staining as well as by cell surface staining of annexin V/propidium iodide. The effect on
cell cycle was assessed by flow cytometry. Results showed that 6r inhibit proliferation of a range of
human cancer cells in vitro and growth of SW620 tumor xenografts in vivo. The anti-proliferative effect
of 6r is associated with cell apoptosis as a result of increased ratio of cellular Bax/bcl-2 expression and
subsequent cytochrome-c and caspase-3 activation. Unlike EA, 6r did not show any influence on cellular
GST P1-1 expression and its anti-proliferative action was associated with cell cycle arrest in G1/S-phase.
In conclusion, 6r has the potential to be developed as a chemotherapeutic agent by induction of cell
apoptosis but not regulating GST P1-1.
� 2012 Elsevier Masson SAS. All rights reserved.
Available online at
www.sciencedirect.com
1. Introduction
Cancer is a major public health concern and life-threateningdisease that affects millions of people around the world each year.In spite of the discoveries of new biological molecules andimmune-mediated therapies, chemotherapy is still the mainclinical method to treating cancer [1]. However, most of thechemotherapeutic agents currently used in clinic have either lowspecificity or high side effect [2]. Thus, development of novelchemotherapeutic agents with high specificity and low side effectsis highly.
Glutathione S-transferases (GSTs) are a family of phase IIdetoxification enzymes that catalyze the S-conjugation of gluta-thione (GSH) with a wide variety of compounds. Glutathione S-transferases P1-1 (GST P1-1) is one of the GST isozymes and hasbeen reported to be overexpressed in a wide range of cancers, such
* Corresponding author. Fax: +86 531 8838 2490.
** Corresponding author. Fax: +86 531 8838 2009.
E-mail addresses: [email protected] (G.-S. Zhao),
[email protected] (X.-L. Guo).
0753-3322/$ – see front matter � 2012 Elsevier Masson SAS. All rights reserved.
http://dx.doi.org/10.1016/j.biopha.2012.10.011
as colon, prostate, breast, skin and lung [3]. GST P1-1 has shown toact as an endogenous inhibitor that suppresses activation of c-JunN-terminal kinase (JNK) and protects cells against stress-inducedapoptosis [4], thus is a potential therapeutic target for cancertreatment.
Ethacrynic acid (EA) is a GST P1-1 inhibitor with an a,b-unsaturated carbonyl functional group and has shown toenhance the therapeutic efficiencies of several chemotherapeuticdrugs [5]. But its low efficacy and high side effects have limited itspharmacological development as an effective chemotherapeuticagent [6]. Investigations of the EA structure-activity relationshiphave indicated that its a,b-unsaturated carbonyl group maylargely be the determinant of its ability to inhibit GST P1-1 activity,while its carboxyl group may primarily govern the main diureticside effect [7]. With this in mind, we designed and synthesized aseries of new EA oxadiazole derivatives using the principle ofbioisosterism. One of the compounds, 5-[2,3-Dichloro-4- (2-methylene-1-oxobutyl) phenoxymethyl]- 3-methyl-1,2,4-oxadia-zole (6r) (Fig. 1) showed to one of the most effective EA oxadiazole-containing analogues in inhibiting growth of cancer cells includingPC3, SW620, DU145 and HL-60 cells [8].

Fig. 1. Chemical structures of ethacrynic acid and 6r.
P. Zhang et al. / Biomedicine & Pharmacotherapy 67 (2013) 58–65 59
The present study investigated the effects of 6r on cancer cellgrowth in vitro and tumor xenografts in nude mice. It was foundthat 6r induces cell cycle rest of tumor cells in G1/S-phase arrestand induces cell apoptosis by activation of bcl-2/Bax-Cyt-c-caspase-3 mitochondrial signaling pathway, an effect thatindependent on GST P1-1.
2. Materials and methods
2.1. Cells and animals
Human colon cancer SW620 cells were kindly provided by Dr.Lugang Yu (Liverpool University, UK), human prostate cancer PC3cells, human colon cancer Lovo and LS174-T cells, human prostatecancer PC3 cells, human lung adenocarcinoma epithelial cell lineA549, human breast cancer MCF-7 and Bacp37 cells, humanepithelial ovarian cancer HO 8910 cells and human cervical cancerHela 229 cells were obtained from the Institute of Biochemistryand Cell Biology, Chinese Academy of Science (Shanghai, China),human myeloid leukemia K562 cells was obtained from theDepartment of Pharmacology, the Institute of Hematology ofChinese Academy of Medical Sciences (Tianjin, China). All thesecells were cultured in Dulbecco’s Modified Eagle’s Medium(DMEM) supplemented with 10% (v/v) heat-inactivated FBS at37 8C in a humid atmosphere (5% CO2 – 95% air). For allexperiments, cells were used at passages 3-8. Six-week-old femaleBalb/c-nu mice were purchased from the Animal Centre of ChinaAcademy of Medical Sciences (Beijing, China) and acclimated andcaged in groups of 6. Animals were housed under pathogen-freeconditions. The research protocol was in accordance with theInstitutional Guidelines of Animal Care and Use Committee atShandong University.
2.2. Reagents
The design and synthesis of 6r was reported previously [8]. 6rcompound was dissolved in cremophor and enthanol (v/v, 1:1) asstock solution and was diluted in saline for the in vitro andanimal experiment. Trypsin, 3-(4,5-dimethyl-thiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), dimethyl sulfoxide(DMSO), 5-Fluorouracil, propidium iodide (PI) and Hoechst 33258were purchased from Sigma (St. Louis, MO, USA). Annexin V/FITCand PI apoptosis detection kit was purchased from MultisciencesBiotech (Hanzhou, China). RNase was obtained from SolarbioScience and Technology (Beijing, China). Monoclonal antibodies
against Bax, bcl-2, caspase-3, cytochrome-c and GST P1-1 werepurchased from Cell Signaling Technology (Boston, MA, USA).Monoclonal antibody against b-actin was purchased from Abcam(Boston, MA, USA) Biotinylated goat antirabbit IgG, goat anti-mouseIgG and nitrocellulose membranes were purchased from Amersham(Buckinghamshire, UK). The LumiGLO Reserve ChemiluminescentSubstrate Kit was purchased from KPL (Gaithersburg, MD, USA).
2.3. Cell proliferation assay
Cells (5 � 103 per well) were seeded into 96-well plates andgrown for 24 h before treatment with various concentrations of 6r(1.0 � 40 mM) for a further 48 h. The culture medium was thenremoved and the wells were washed with PBS. The MTT assay wasperformed by adding 20 ml of MTT (5 mg/mL) to each well for 4 h.Then 200 ml of dimethyl sulfoxide (DMSO), the solubilization/stopsolution, was added to dissolve the formazan crystals before theabsorbance was measured using a THERMOmax microplate reader(Molecular Devices, Sunnyvale, CA, USA) at a wavelength of570 nm. The GI50 value was obtained by the SPSS/Win 13.0software (SPSS, Inc., Chicago, Illinois, USA) [9,10].
2.4. Analysis of cell apoptosis by change of nuclear morphology
SW620 cells seeded in 24-well plates (8.0 � 104 per well) weretreated with various concentrations of 6r for 24 h. The cells werefixed in 4% paraformaldehyde in PBS for 30 min, and then stainedwith 10 mg/ml Hoechst 33,258 in dark at room temperature for10 min. After three washes with PBS, the cells were visualized withfluorescence microscope (excitation, 340 nm; emission, 460 nm)(NIKON, Ti-U, Tokyo, Japan). Viable cells were identified by theirintact nuclei with blue fluorescence, apoptotic cells by theirchromosome condensation and fragmentation of nuclei, exhibitinghighlight staining [11].
2.5. Analysis of cell apoptosis by annexin V cell surface binding
SW620 cells seeded in 6-well plates (2.0 � 105 per well) weretreated with various concentrations of 6r for 24 h. The cell wereharvested, washed with cold PBS and the cell surface phospha-tidylserine in apoptotic cells was quantitatively determined usingAnnexin V/FITC and PI apoptosis detection kit according tomanufacturer’s instructions with a FACSVantage flow cytometer(Becton Dickinson Immunocytometry System, San Jose, CA, USA)using emission filters of 525 and 575 nm, respectively [12,13].
2.6. Cell cycle analysis
SW620 cells were seeded into 6-well plates (2.0 � 105 per well)and treated with various concentrations of 6r for 24 h and 48 h. Thecells were harvested, washed twice with cold PBS and fixed withprecooled 70% ethanol at 4 8C for 24 h. One hundred microgram permillilitre RNase was then added for 30 min at 37 8C before additionof propidium iodide at a final concentration of 50 mg/mL for DNAstaining in the dark at 4 8C for 45 min. For each sample, more than1 � 104 cells were analyzed using a FACScan flow cytometry. Cellcycle distribution was analyzed using the Modifit’s program(Becton Dickinson) [14,15].
2.7. Assessment of 6r antitumor activity in vivo
SW620 xenografts were established by inoculating 2.0 � 106
SW620 cells s.c in nude mice. The experiments began when thexenografts volume reached approximately 1.0 cm3 in nude mice.The tumor were extracted and cut into 1 mm3 fragments (about20 mg/fragment) under a sterile condition. One fragment was

Fig. 2. 6r inhibits proliferation of a panel of human tumor cell lines in vitro. Cells in
96-well plates were treated with various concentration of 6r for 48 h before the cell
viability was determined by 3-(4,5-dimethyl-thiazol-2-yl)-2,5-diphenyl
tetrazolium bromide assay. The data represent mean � SD of three separated
experiments.
P. Zhang et al. / Biomedicine & Pharmacotherapy 67 (2013) 58–6560
transplanted s.c. into the right flank by trocar in each nude mouse[16]. When the tumor reached a volume of 100 mm3 to 200 mm3,the mice were randomed to six groups, one group as controlto receive vehicle, other five groups were treated with 6r or5-Fluorouracil (5-Fu) at indicated doses by intravenous. via tail vein.Administration was performed twice weekly for three consecutiveweeks. The size of tumor was measured each week with calipers.Tumor volume (V) was calculated as V = (length � width2 � p)/6[17]. At the end of the experiment, the mice were sacrificed and thetumors were removed and weighed. Effect of the compound ontumor growth was expressed as a percentage to that in the controlgroup [18,19].
2.8. Analysis of GST P1-1 and apoptosis-related protein expression in
tumor xenografts by immunoblotting
SW620 cells were seeded into 6-well plates (2.0 � 105 per well)and treated with various concentrations of 6r for 24 h. The cells ortumor tissues of SW620 xenografts were dispersed mechanically inlysis buffer (50 mM Tris-Hcl pH 7.5; 1 mM EDTA; 150 NaCl; 50 mMNaF; 1 mM Na3VO4; 1 mM PMSF, 1% NP-40). The suspension wasthen centrifuged at 13,000 rpm for 15 min before collection of thesupernatants and determination of protein concentration using theBradford method [20]. Tumor or cell lysates (50 mg protein/lane ofthe tumor lysates or 30 mg/lane of the cell lysates) were subjectedto SDS-polyacrylamide gel electrophoresis (PAGE) followed byprotein transfer to a nitrocellulose membranes and probed withmonoclonal antibodies against GST P1-1, caspase-3, Bax, bcl-2,Cytochrome-c and b-actin. Immunoblots were developed usinghorseradish peroxidase-conjugated secondary antibodies, andvisualized by the enhanced chemiluminescent (ECL) system(Millipore, Billerica, MA, USA) and quantified by densitometryusing a ChemiDoc XRS (Bio-Rad, Berkeley, California, USA). Theband density was normalized to b-actin. The percentages ofinhibition were estimated by comparison to the untreated vehicle(100%) [21].
2.9. Statistical analysis
The data are expressed as mean � SD. Statistical analysis wasperformed by one-way analysis of variance (Anova) followed byDunnett’s multiple range tests using the SPSS/Win 13.0 software.P < 0.05 was considered as statistically significant.
3. Results
3.1. 6r inhibits cancer cell proliferation in vitro
The effect of 6r on cell proliferation was evaluated using theMTT assay against a panel of 10 human tumor cell lines includingthose from colon (SW620, Lovo, LS174-T), leukemia (K562),prostate (PC3), lung (A549), breast (MCF-7, Bacp37), ovarian(HO8910) and cervical cancer (Hela229). The presence of 6r
Table 1The inhibitory effect of 6r on SW620 xenografts in Balb/c-nu mice (n = 7).
Groups Dose (mg/kg) Body weight (initial/17 days, g)
Vehicle 0 21.7 � 1.7/18.0 � 2.0
6r 1 21.5 � 2.0/18.3 � 1.5
6r 2 21.0 � 1.0/17.7 � 2.0
6r 4 21.5 � 1.6/18.8 � 1.7
6r 8 21.0 � 1.6/17.8 � 2.3
5-Fu 15 21.1 � 1.5/15.7 � 1.6*
Inhibition rates were defined as percentage of vehicle treated-tumor weight.* P < 0.05.** P < 0.01, significant difference compared with the vehicle group.
showed to cause significant inhibition of proliferation of PC3,SW620, K562 and Hela 229 cells with GI50 value of 3.79, 4.89, 5.08and 4.45 mmol/L, respectively. Lovo, LS174-T, A549 and HO89100cell lines were moderate sensitive to 6r with GI50 value of 15.85,14.13, 12.11 and 13.65 mmol/L (Fig. 2). MCF-7 and Bacp 37 celllines were not sensitive to 6r. Results showed that all three coloncancer cell lines (SW620, Lovo, LS174-T) were sensitive to 6r withdifferent degree. Thus, we chose SW620 cells, one of the cell linesto which 6r showed to have the most effect, for the followingexperiments in this study.
3.2. 6r inhibits tumor growth in mice implanted with human SW620
colon cancer
Treatment of nude mice with 6r resulted in significantinhibition of growth of SW620 tumor xenografts in nude mice(Table 1, Fig. 3). Administration of 6r at 8 mg/kg by intravenous.resulted in 44.17% reduction of tumor growth and volume (Fig. 3b)after 17 days in comparison to the control group. A similar effectwas produced by administration of 5-Fu at 15 mg/kg (49.17%). Theanimals demonstrated general tolerance with 6r treatment and nosignificant loss of the body weight was observed (P > 0.05 vs.vehicle group). On the other hand, the animals administrated with5-Fu showed significant reduction of body weight (Table 1).
3.3. No effect of 6r on GST P1-1 expression in SW620 cells and in tumor
xenografts
Previous study has reported that 6r showed over 20 time higherinhibition than EA on the growth of HL-60 cells, but produced lessinhibition on GST P1-1 activity than EA [8,22]. We therefore
Tumor weight (mean � S.D, g) Inhibition rate (%)
1.20 � 0.20 0
1.09 � 0.12 9.17
0.85 � 0.15** 29.17
0.71 � 0.13** 35.00
0.67 � 0.10** 44.17
0.64 � 0.15** 49.17

Fig. 3. 6r inhibits the growth of SW620 xenografts in nude mice. SW620 cells were injected s.c into the right anterior flank of nude mice as described in ‘‘Materials and
methods’’. Mice were treated every two days via tail vein with 6r, 5-Fu or Mitoxantone for three consecutive weeks. Images of the subcutaneous tumors derived from the
SW620 xenografts are shown in a and tumor volume for SW620 xenografts are shown in b (means � SEM).
P. Zhang et al. / Biomedicine & Pharmacotherapy 67 (2013) 58–65 61
assessed the effect of 6r on GST P1-1 expression in SW620 cells andtumor xenografts. It was found that the presence of 6r had nosignificant effect on of GST P1-1 expression in either SW620 cells orxenografts (Fig. 4).
3.4. Effect of 6r on apoptosis in SW620 cells
In order to investigate the effect of 6r on SW620 apoptosis, wefirst examined the cell morphology in cell response to 6rtreatment. Treatment of SW620 cells with 1, 2 or 4 mmol/L 6rfor 24 h followed by nucleus staining with Hoechst 33258 showedincreased number of cells with reduced nuclear size, chromatincondensation, and nuclear fragmentation, characteristics ofapoptosis in comparison to the control cells (Fig. 5a–e).
Annexin V cell surface staining followed by flow cytometryanalysis also showed significant increase of annexin V positivecells following treatment of the cells with 2 mmol/L or 4 mmol/L 6r
Fig. 4. 6r has no effect on GST P1-1 expression in SW620 cells and in SW620 xenografts. T
after immunoblotting with monoclonal antibody against GST P1-1 (b). (c) Quantification
actin (mean � SD, n = 3). * P < 0.05 and ** P < 0.01 vs. vehicle group.
(Fig. 6) These results indicate that 6r is capable of inducing SW620cell apoptosis in a concentration-dependent manner.
3.5. 6r-induced cell cycle arrest in S-phase
Treatment of SW620 cells with 4 mmol/L 6r for 24 or 48 hr wasfound to result in significant increase (46.42% and 66.85%,respectively) of the cells in S-phase in Fig. 7a and b. This suggeststhat the antiproliferative effect of 6r is related to the cell rest inS-phase in a concentration- and time-dependent manner (Fig. 7cand d).
3.6. 6r induces caspase-3 activation and cytochrome-c release in
tumor xenografts
It was found that administration of 6r at 8 mg/kg byintravenous for 17 days caused marked increase of the cleavage
he representative bands of GST P1-1 expression in SW620 cells (a) or tumor tissues
of the GST P1-1 bands in SW620 cells or (d) tumor tissues after normalization to b-

Fig. 5. 6r induces apoptosis in SW620 cells using Hoechst 33258 staining. SW620 cells were stained with DNA-binding fluorochrome Hoechst 33258 and treated with or
without 6r for 24 h. Representative images of SW620 cells after treatment with (a) vehicle, (b) 6r 1 mM, (c) 6r 2 mM, (d) 6r 4 mM groups. Arrows indicate characteristic
apoptotic features. Scale bar = 100 mm. The percentage of apoptotic cells in 100 cells was scored in four random observations and the results are shown in (e) (mean � S.D.). **
P < 0.01 vs. vehicle group.
Fig. 6. 6r induces apoptosis. Cells were treated with (a) vehicle, (b) 6r 1 mM, (c) 6r 2 mM, (d) 6r 4 mM before cell apoptosis was analysed by flow cytometry following Annexin
V/FITC and PI double-staining. The FL1 axis shows the Annexin V intensity, and the FL2 axis shows the PI staining. The percentages of apoptosis and necrosis cells are scored in
three separate experiments and the results are shown in (e) (mean � S.D.). * P < 0.05 and ** P < 0.01 vs. vehicle group.
P. Zhang et al. / Biomedicine & Pharmacotherapy 67 (2013) 58–6562
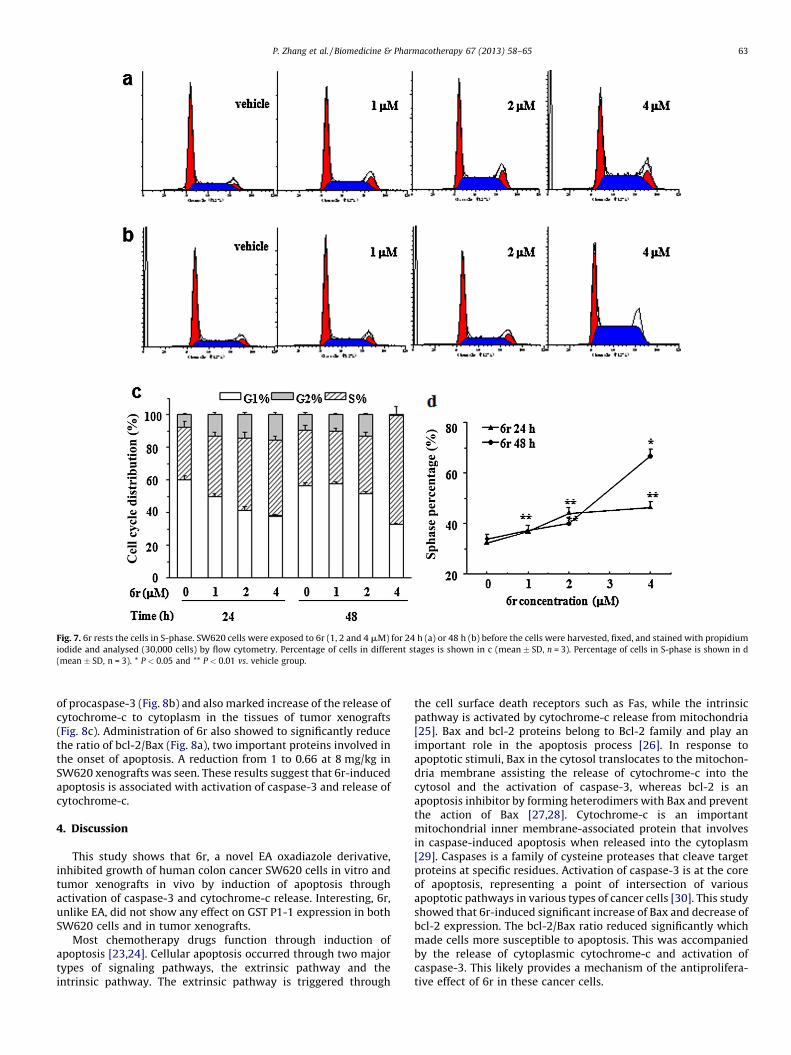
Fig. 7. 6r rests the cells in S-phase. SW620 cells were exposed to 6r (1, 2 and 4 mM) for 24 h (a) or 48 h (b) before the cells were harvested, fixed, and stained with propidium
iodide and analysed (30,000 cells) by flow cytometry. Percentage of cells in different stages is shown in c (mean � SD, n = 3). Percentage of cells in S-phase is shown in d
(mean � SD, n = 3). * P < 0.05 and ** P < 0.01 vs. vehicle group.
P. Zhang et al. / Biomedicine & Pharmacotherapy 67 (2013) 58–65 63
of procaspase-3 (Fig. 8b) and also marked increase of the release ofcytochrome-c to cytoplasm in the tissues of tumor xenografts(Fig. 8c). Administration of 6r also showed to significantly reducethe ratio of bcl-2/Bax (Fig. 8a), two important proteins involved inthe onset of apoptosis. A reduction from 1 to 0.66 at 8 mg/kg inSW620 xenografts was seen. These results suggest that 6r-inducedapoptosis is associated with activation of caspase-3 and release ofcytochrome-c.
4. Discussion
This study shows that 6r, a novel EA oxadiazole derivative,inhibited growth of human colon cancer SW620 cells in vitro andtumor xenografts in vivo by induction of apoptosis throughactivation of caspase-3 and cytochrome-c release. Interesting, 6r,unlike EA, did not show any effect on GST P1-1 expression in bothSW620 cells and in tumor xenografts.
Most chemotherapy drugs function through induction ofapoptosis [23,24]. Cellular apoptosis occurred through two majortypes of signaling pathways, the extrinsic pathway and theintrinsic pathway. The extrinsic pathway is triggered through
the cell surface death receptors such as Fas, while the intrinsicpathway is activated by cytochrome-c release from mitochondria[25]. Bax and bcl-2 proteins belong to Bcl-2 family and play animportant role in the apoptosis process [26]. In response toapoptotic stimuli, Bax in the cytosol translocates to the mitochon-dria membrane assisting the release of cytochrome-c into thecytosol and the activation of caspase-3, whereas bcl-2 is anapoptosis inhibitor by forming heterodimers with Bax and preventthe action of Bax [27,28]. Cytochrome-c is an importantmitochondrial inner membrane-associated protein that involvesin caspase-induced apoptosis when released into the cytoplasm[29]. Caspases is a family of cysteine proteases that cleave targetproteins at specific residues. Activation of caspase-3 is at the coreof apoptosis, representing a point of intersection of variousapoptotic pathways in various types of cancer cells [30]. This studyshowed that 6r-induced significant increase of Bax and decrease ofbcl-2 expression. The bcl-2/Bax ratio reduced significantly whichmade cells more susceptible to apoptosis. This was accompaniedby the release of cytoplasmic cytochrome-c and activation ofcaspase-3. This likely provides a mechanism of the antiprolifera-tive effect of 6r in these cancer cells.

Fig. 8. Effect of 6r on cellular expressions of procaspase-3, cleaved caspase-3, bcl-2, Bax and Cyt-c in SW620 xenografts. The SW620 tumor lysates were separated by SDS-
PAGE and probed with antibodies against bcl-2 and Bax (a), procaspase-3, cleaved caspase-3 (b), Cyt-C (c). (d) The densities of cleaved caspase-3, bcl-2/Bax bands were
quantified by densitometry after normalization to b-actin and are shown as the percentage to the controls (mean � SD, n = 3). * P < 0.05 and ** P < 0.01 vs. vehicle group.
P. Zhang et al. / Biomedicine & Pharmacotherapy 67 (2013) 58–6564
Another main regulatory mechanism of cell growth is cell cycleregulation [31]. Many chemopreventive and therapeutic agentshave shown to induce apoptosis after cell cycle arrest [32]. Forexample, SW620 cells have been shown to rest at S-phase inresponse to multiple such agents, 5-FU, Oxaliplatin and olive oilpolyphenols, etc. [33–35]. In this study, we found that 6r alsoinduced S-phase arrest of SW620 cells in a concentration-dependent and time-dependent manner. Remarkable cell cycleperturbations were observed after treatment of 6r with 4 mM for48 h, indicating that the drug primarily exerted its activity byinfluencing the cell cycle of SW620 cells. Although the mechanismof the influence of 6r on cell S-phase rest is unclear, it is possiblethat cyclins or CDKs may be involved in the process. Previousstudies have reported that cyclins Clb5 and Clb6, complexes ofCdk1 and Cln cyclins were involved in the initiation of S-phase incell cycle [36–38].
In conclusion, as a novel EA oxadiazole analogue, 6r inhibitsproliferation of human colon SW620 cancer in vitro and in tumorxenografts in vivo. This effect of 6r is associated with its regulationof cellular bcl-2/Bax expression and subsequent cytochrome-crelease and caspase-3 activation with cell rest in S-phase. UnlikeEA, 6r does not regulate GST P1-1 expression. 6r is therefore has thepotential to be developed as a chemotherapeutic agent.
Disclosure of interest
The authors declare that they have no conflicts of interestconcerning this article.
Acknowledgements
This work was supported by The Major Projects of Science andTechnology for Drug Innovation (2009ZX09103-116) and TheNatural Science Foundation of China (30973548).
References
[1] Neidle S, Thurston DE. Chemical approaches to the discovery and developmentof cancer therapies. Nat Rev Cancer 2005;5:285–96.
[2] Newman DJ, Cragg GM. Natural products as sources of new drugs over the last25 years. J Nat Prod 2007;70:461–77.
[3] Ballerini S, Bellincampi L, Bernardini S, Lori R, Cortese C, Federici G. Analysis ofGSTP1-1 polymorphism using real-time polymerase chain reaction. Clin ChimActa 2005;329:127–32.
[4] Wang T, Arifoglu P, Ronai Z, Tew KD. Glutathione S-transferase P1-1 (GSTP1-1)inhibits c-Jun-N-terminal kinase (JNK1) signaling through interaction with theC terminus. J Biol Chem 2001;276:20999–1003.
[5] Aizawa S, Ookawa K, Kudo T, Asano J, Hayakari M, Tsuchida S. Characterizationof cell death induced by ethacrynic acid in a human colon cancer cell line DLD-1 and suppression by N-acetyl-L-cysteine. Cancer Sci 2003;94:886–93.
[6] Zhao G, Wang X. Advance in antitumor agents targeting glutathione-S-trans-ferase. Curr Med Chem 2006;13:1461–71.
[7] van Iersel ML, van Lipzig MM, Rietjens IM, Vervoort J, van Bladeren PJ. GSTP1-1stereospecifically catalyzes glutathione conjugation of ethacrynic acid. FEBSLett 1998;441:153–7.
[8] Yang XM, Liu GL, Li HC, Zhang Y, Song DD, Li CM, et al. Novel oxadiazoleanalogues derived from ethacrynic acid: design, synthesis, and structure-activity relationships in inhibiting the activity of glutathione S-TransferaseP1-1 and cancer cell proliferation. J Med Chem 2010;53:1015–22.
[9] Wesbuer S, Lanvers-Kaminsky C, Duran-Seuberth I, Bolling T, Schafer KL, BraunY, et al. Association of telomerase activity with radio- and chemosensitivity ofneuroblastomas. Radiat Oncol 2010;5:66.
[10] Kamal A, Sultana F, Ramaiah MJ, Srikanth YV, Viswanath A, Kishor C, et al. 3-substituted 2-phenylimidazo[2,1-b] benzothiazoles: synthesis, anticancer ac-tivity, and inhibition of tubulin polymerization. Chem Med Chem 2012;7:292–300.
[11] Chen JX, Zhao T, Huang DX. Protective effects of edaravone against cobalthlor-ide-induced apoptosis in PC12 cells. Neurosci Bull 2009;25:67–74.
[12] Bacso Z, Everson RB, Eliason JF. The DNA of annexin V-binding apoptotic cells ishighly fragmented. Cancer Res 2000;60:4623–8.
[13] Qi FH, Li AY, Lv H, Zhao L, Li JJ, Gao B, et al. Apoptosis-inducing effect ofcinobufacini. Bufo bufo gargarizans cantor skin extract on human hepatomacell line BEL-7402 Drug Discov Ther 2010;1:339–43.
[14] Lee SK, Kim HN, Kang YR, Lee CW, Kim HM, Han DC, et al. Obovatol inhibitscolorectal cancer growth by inhibiting tumor cell proliferation and inducingapoptosis. Bioorg Med Chem 2008;16:8397–402.
[15] Sikandar S, Dizon D, Shen XL, Li ZM, Besterman J, Lipkin SM. The class I hdacinhibitor Mgcd0103 induces cell cycle arrest and apoptosis in colon cancerinitiating cells by upregulating dickkopf-1 and non-canonical wnt signaling.Oncotarget 2010;1:596–605.
[16] Xiong X, Fu L, Wang L, Cai H, Li L, Jiang H, et al. Antitumor activity of a novelEGFR tyrosine kinase inhibitor against human lung carcinoma in vitro and invivo. Invest New Drugs 2009;27:1–11.
[17] Lennon FE, Mirzapoiazova T, Mambetsariev B, Salgia R, Moss J, Singleton PA.Overexpression of the m-opioid receptor in human non-small cell lung cancerpromotes Akt and mTOR activation, tumor growth, and metastasis. Anesthe-siology 2012;116:857–67.
[18] Chen WC, Kuo TH, Tzeng YS, Tsai YC. Baicalin induces apoptosis in SW620human colorectal carcinoma cells in vitro and suppresses tumor growth invivo. Molecules 2012;17:3844–57.

P. Zhang et al. / Biomedicine & Pharmacotherapy 67 (2013) 58–65 65
[19] Hou L, Xu B, Guo W, Ran FX, Liu JT, Yuan X, et al. (2012) Pseudolaric acid Binhibits inducible cyclooxygenase-2 expression via downregulation of the NF-kB pathway in HT-29 cells. J Cancer Res Clin Oncol Feb 8 [Epub ahead of print].
[20] Okutucu B, Dincer A, Habib O, Zihnioqlu F. Comparison of five methods fordetermination of total plasma protein concentration. J Biochem BiophysMethods 2007;70:709–11.
[21] Choi ES, Shim JH, Jung JY, Kim HJ, Choi KH, Shin JA, et al. Apoptotic effect oftolfenamic acid in androgen receptor-independent prostate cancer cell andxenograft tumor through specificity protein 1. Cancer Sci 2011;102:742–8.
[22] Zhao GS, Liu C, Wang R, Song DD, Wang XB, Lou HX, et al. The synthesis of a, b-unsaturated carbonyl derivatives with the ability to inhibit both glutathione S-transferase P1-1 activity and the proliferation of leukemia cells. Bioorgan MedChem 2007;15:2701–7.
[23] Diaial NN, Korsmeyer SJ. Cell death: critical control points. Cell 2004;116:205–19.
[24] Fesik SW. Promoting apoptosis as a strategy for cancer drug discovery. Nat RevCancer 2005;5:876–85.
[25] Ghobrial IM, Witzig TE, Adjei AA. Targeting apoptosis pathways in cancertherapy. CA Cancer J Clin 2005;55:178–94.
[26] Falschlehner C, Emmerich CH, Gerlach B, Walczak H. TRAIL signlling: decisionsbetween life and death. Int J Biochem Cell Biol 2007;39:1462–75.
[27] McNutt MC, Lagace TA, Horton JD. Catalytic activity is not required for secretedPCSK9 to reduce low density lipoprotein receptors in HepG2 cells. J Biol Chem2007;282:20799–803.
[28] Kauntz H, Bousserouel S, Gosse F, Marescaux J, Raul F. Silibinin, a naturalflavonoid, modulates the early expression of chemoprevention biomarkers in apreclinical model of colon carcinogenesis. Int J Oncol 2012;41:849–54.
[29] Tsuji T, Du W, Nishioka T, Chen L, Yamamoto D, Chen CY. Phellinus linteusextract sensitizes advanced prostate cancer cells to apoptosis in athymic nudemice. PLoS One 2010;5:e9885.
[30] Xing ZB, Yao L, Zhang GQ, Zhang XY, Zhang YX, Pang D. Fangchinoline inhibitsbreast adenocarcinoma proliferation by inducing apoptosis. Chem Pharm Bull2011;59:1476–80.
[31] Murray AW. Recycling the cell cycle: cyclins revisited. Cell 2004;116:221–34.[32] Schwartz GK, Shan MA. Targeting the cell cycle: a new approach to cancer
therapy. J Clin Oncol 2005;23:9408–21.[33] Matuo R, Sousa FG, Escargueil EE, Grivicich I, Garcia-Santos D, Chies JAB, et al.
5-Fluorouracil and its active metabolite FdUMP cause DNA damage in humanSW620 colon adenocarcinoma cell line. J Appl Toxicol 2009;29:308–16.
[34] Nannizzi S, Veal GJ, Giovannetti E, Mey V, Ricciardi S, Ottley CJ, et al. Cellularand molecular mechanisms for the synergistic cytotoxicity elicited by oxali-platin and pemetrexed in colon cancer cell lines. Cancer Chemother Pharmacol2010;66:547–58.
[35] Notarnicola M, Pisanti S, Tutino V, Bocale D, Rotelli MT, Gentile A, et al. Effectsof olive oil polyphenols on fatty acid synthase gene expression and activity inhuman colorectal cancer cells. Genes Nutr 2011;6:63–9.
[36] Palou G, Palou R, Moreno AG, Duch A, Travesa A, Quintana DG. Cyclin regula-tion by the S-phase checkpoint. J Biol Chem 2010;285:26431–40.
[37] Koivomagi M, Valk E, Venta R, Iofik A, Lepiku M, Balog ER, et al. Cascades ofmultisite phosphorylation control Sic1 destruction at the onset of S phase.Nature 2011;480:128–31.
[38] Tu YS, Kang XL, Zhou JG, Lv XF, Tang YB, Guan YY. Involvement of Chk1-Cdc23A-cyclin A/CDK2 pathway in simvastatin induced S-phase cell cycle arrest andapoptosis in multiple myeloma cells. Eur J Pharmacol 2011;670:356–64.





























