-
HLA-C Expression Pattern Is Spatially Differentbetween Psoriasis
and Eczema Skin LesionsLina Carlen1, Kazuko Sakuraba1, Mona Stahle1
and Fabio Sanchez1
Interactions between genetic and environmental factors underlie
the immune dysregulation and keratinocyteabnormalities that
characterize psoriasis. Among known psoriasis susceptibility loci
(PSORS), PSORS1 onchromosome 6 has the strongest association to
disease. Altered expression of some PSORS1 candidate geneshas been
reported but little is known about HLA-C expression in psoriasis.
This study compared expression ofmajor histocompatibility complex
class Ia and HLA-C in psoriasis, allergic contact eczema, and
normal skin.Although HLA-C was abundant in protein extracts from
both eczema and psoriasis, a consistent and intriguingdifference in
the expression pattern was observed; strong immunoreactivity in the
basal cell layer, polarizedtowards the basement membrane in
psoriasis, whereas in eczema lesions HLA-C immunostaining was
presentmostly in suprabasal cells. Inflammatory cells in the dermis
were strongly stained in both diseases. Normal skinepithelium
showed less intense but similar HLA-C staining as eczema lesions.
HLA class Ia expression overallresembled that of HLA-C in all
samples. The distinct HLA-C expression patterns in psoriasis and
eczema suggesta functional role in the specific psoriasis immune
response and not only a general feature of inflammation.
Journal of Investigative Dermatology (2007) 127, 342348.
doi:10.1038/sj.jid.5700549; published online 28 September 2006
INTRODUCTIONAssociation between psoriasis and the major
histocompat-ibility complex (MHC) region is firmly established
(McMi-chael et al., 1978; Laurentaci et al., 1982; Henseler
andChristophers, 1985). A minimal genomic segment definingthe
psoriasis susceptibility locus 1 (PSORS1) on chromosome6 (Jenisch
et al., 1998; Balendran et al., 1999; Oka et al.,1999; Nair et al.,
2000) contains a number of proposedcandidate genes, for example
HLA-C, CDSN (corneodesmo-sin), CCHCR1 (coiled-coil alpha-helical
rod protein 1), SEEK,and PSORS1C3 (Bowcock and Krueger, 2005). All
of themhave been investigated for strength of association to
psoriasisin genetic studies but only some have been studied
forexpression in skin (Holm et al., 2003; Suomela et al.,
2003;Capon et al., 2004). There are no published studies
thatdescribe HLA-C expression in psoriasis despite the fact that
itsassociation to disease has been known for several decades.
It is known that under non-inflammatory conditions allnucleated
cells express class I molecules, and that one oftheir important
functions is to suppress self-reactivity byengaging inhibitory
receptors on natural killer (NK), NK-Tand CD8 cells (Martin and
Carrington, 2005; Rajagopalan
and Long, 2005). HLA-C but not HLA-A and B is expressed
bytrophoblast where it regulates maternal NK cell
reactivity(Parham, 2004). HLA-C has recently gained attention
intransplantation biology because it has been recognized
thatmismatches in the killer immunoglobulin-like receptor (KIR)-HLA
class I system can lead to transplant rejection (Petersdorfet al.,
1997). This interaction is strongly influenced by theamino acids
present at position 80 and 44 of the HLA-C andKIR molecules,
respectively (Boyington et al., 2001) and isalso influenced by the
glycosylation state of the ligands (Babaet al., 2000). HLA-Cw*0602
has a lysine at position 80 andinteracts mostly with KIRs that
possess a methionine atposition 44 (Winter and Long, 1997), such as
KIR2DL1(inhibitory) and KIR2DS1 (activating). Upon
interaction,ligand and receptor are internalized into the effector
cell(Carlin et al., 2001) which is followed by cellular
activationor inhibition (Moretta and Moretta, 2004). HLA-C
expressionin the skin has not been described yet, although it is
generallyassumed to be low based on expression in other
tissues(McCutcheon et al., 1995).
Although several inflammatory diseases associate
withpolymorphisms at MHC class I genes, for example, ankylos-ing
spondylitis with the HLA-B27 allele and subacutethyroiditis with
the HLA-B35 allele (Nyulassy et al., 1977;Rubin et al., 1994),
psoriasis is the only inflammatory diseasethat associates with
HLA-C (HLA-Cw*0602). The role ofMHC class Ia in the
pathophysiological mechanisms ofpsoriasis is not well understood;
however, it is clear thatinflammation is a marker of disease
activity (Krueger andBowcock, 2005) and that environmental factors
such asinfections can trigger this disease (Wolf and Ruocco,
1999;Langley et al., 2005), which provides an immunological
ORIGINAL ARTICLE
342 Journal of Investigative Dermatology (2007), Volume 127
& 2006 The Society for Investigative Dermatology
Received 22 February 2006; revised 17 May 2006; accepted 22 May
2006;published online 28 September 2006
1Dermatology and Venereology Unit, Department of Medicine,
KarolinskaInstitutet, Stockholm, Sweden
Correspondence: Dr Fabio Sanchez, Molecular Dermatology, CMM
L8:02,Karolinska University Hospital, SE-171 76 Stockholm,
Sweden.E-mail: [email protected]
Abbreviations: KIR, killer cell immunoglobulin-like receptor;
MHC, majorhistocompatibility complex; NK, natural killer; PSORS,
psoriasissusceptibility locus
-
playground for HLA-C. In particular, streptococcal
throatinfections are often concomitant with onset of
guttatepsoriasis, which is the psoriasis subphenotype with
thestrongest association to HLA-Cw*0602 (Mallon et al., 2000;Holm
et al., 2005). Whether these associations are circum-stantial or do
indeed contribute to pathophysiology remainsto be established. In
the late 1970s, Gross et al. (1977, 1983)reported that, in
psoriasis, T-cell functions in response to invitro exposure to
streptococcal factors are influenced byHLA-C genotypes. No
additional reports addressing thisphenomenon are available since
then. It is likely thattechnical difficulties arising from the high
degree ofhomology between MHC class I genes and from the highdegree
of polymorphism within each gene group explain thescarcity of
studies on this subject. Nonetheless, the role ofHLA-C in immune
surveillance (Boyington and Sun, 2002)and its importance in
relation to infection grants a closer lookat its expression pattern
in psoriasis.
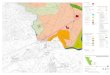
RESULTSHLA-C is expressed in skin, tonsil, and adult colon
epitheliumbut not in fetal colon epithelium
The level and pattern of HLA-C and HLA-ABC expressionwere
investigated in normal specimens of skin, tonsil, and
colon using antibodies recognizing HLA-C (L31) andHLA-A,B and C
(W6/32). Immunohistochemical analysisshowed that HLA-C staining in
tonsil was strong aroundgerminal follicles and in epithelial cells
(Figure 1a). Therewas no HLA-C staining in fetal colon epithelium
(Figure 1b),although strong staining in neighboring lymphoid tissue
onthe same specimen was evident, in contrast, there was verystrong
staining in adult colon epithelium (Figure 1c). HLA-Cexpression in
normal skin epithelium was predominantlysuprabasal with a
membrane-like staining pattern (Figure 1d).Strong immunoreactivity
for HLA-ABC was observed in allcells, including epithelial cells,
in all tissues and in leukocyte-like cells, especially large
dendritic-like cells (data notshown), except in fetal colon
epithelium where the stainingwith this antibody was faint.
Psoriasis and allergic contact eczema skin lesions show
spatiallydisparate HLA-C expression patterns
Next, we compared the HLA-C expression between plaquepsoriasis,
allergic contact eczema, and normal skin. Epider-mal HLA-C
expression in normal and eczematous tissue wasstronger than in
plaque psoriasis. Immunoreactivity in plaquepsoriasis was
consistently stronger in the basal layer,especially at the tips of
rete ridges where the signal often
a
d
g h i j
e f
b c
Figure 1. HLA-C expression detected by immunohistochemistry with
mAb L31. (a) Tonsil, (b) adult colon epithelium, (c) fetal colon
epithelium, (d) normal
skin, (e) plaque psoriasis, (f) allergic contact eczema
(original magnification 20, Bar 100mM, insets: 100, Bar20 mm) (g)
LCL721.221 cells expressingHLA-Cw3 before lipopolysaccharide
stimulation, and after (h) 16, (i) 48, and (j) 96 hours
post-stimulation (original magnification 40, Bar 50 mm).
www.jidonline.org 343
L Carlen et al.HLA-C Expression Pattern in Psoriasis
-
polarized towards the basement membrane (Figure 1e).
Clearpericellular delineation, compatible with membrane
staining,was observed in suprabasal keratinocytes of normal
andallergic contact eczema (Figure 1f). Not all cell types in
skinstained with the L31 anti-HLA-C antibody; fibroblast-likecells
were mostly negative. Leukocyte-like cells as well aslarge
dendritic-like cells stained strongly with anti-HLA-C inall sample
groups.
MHC class Ia expression was intense and had a
similardistribution pattern to that of HLA-C, showing less
intensestaining in the epithelium of plaque psoriasis compared
tonormal and eczema skin but with relatively
strongerimmunoreactivity in the basal cell layer. An inverse
relation-ship between the number of infiltrating cells and the
averagestaining density at the basal layer was observed.
HLA-C expression on transfected LCL721.221 cells is
induciblewith E. coli-derived lipopolysaccharide
In order to determine if HLA-C immunoreactivity correlatedwith
changes in cellular functional state, LCL721.221 cells,which lack
HLA class I expression, were used. LCL721.221cells transfected with
HLA-Cw3 and Cw4, as well as, non-transfected cells were stimulated
with lipopolysaccharide forup to 96 hours and then tested for
reactivity with the L31antibody. There was a time-dependent
increase in thefrequency of L31-positive cells in HLA-Cw3 and
Cw4transfectants, which reached a peak at 48 hours (Figure1gj).
Very faint expression was observed before lipopoly-saccharide
stimulation for both constructs. No increase instaining intensity
or in the frequency of positive cells wasobserved in
non-transfected cells at the set time points.
HLA-C protein levels are increased in lesional allergic
contacteczema and plaque psoriasis skin
Using the L31 antibody, a band of approximately 45
kDa,compatible with the denatured form of HLA-C, was detectedby
Western blot in protein extracts from lesional allergiccontact
eczema, plaque and guttate psoriasis, non-lesionalsamples for these
three sample types, and normal skinsamples. The range of HLA-C
intensity values among thedifferent samples was between 0.10 and
1.08 expressionunits. Analysis of variance revealed inter-group
differencesaccording to sample category (P 0.0039). Allergic
contacteczema had the most intense HLA-C signal (averageexpression
index 0.665, standard deviation (0.343),being significantly higher
than in the other samples exceptfor plaque psoriasis (average
expression index0.425,standard deviation0.189) (Figure 2a). Overall
HLA class Iexpression assessed by b2-microglobulin staining was
alsoincreased in allergic contact eczema compared to the otherskin
samples (average expression index0.256, standarddeviation0.151,
P0.0002) (Figure 2b). A representativeimage of an L31-stained
membrane is presented in Figure 3.
DISCUSSIONThe contribution of the PSORS1 locus to psoriasis
suscept-ibility has been a subject of intense research over the
past twodecades (Bowcock and Krueger, 2005). Although several
1.0
0.8
0.6
0.4
0.2
LE LG LP NPhenotype
Exp
ress
iom
inde
x
0.5
0.4
0.3
0.2
0.1
0.0
Exp
ress
iom
inde
x
NLE NLG NLP
LE LG LP NPhenotype
NLE NLG NLP
a
b
Figure 2. HLA-C protein expression levels in total protein
extracts from
psoriasis, eczema and normal skin biopsies. (a) Box plot
representation of
HLA-C expression according to sample group, measured by
densitometry on
Western blot analysis. (b) Box plot representation of HLA class
I expression,
measured by densitometry on Western blot analysis and detected
by b2-microglobulin staining. LE lesional eczema, LG lesional
guttate psoriasis,LP lesional plaque psoriasis, Nnormal, NLE
non-lesional eczema,NLGnon-lesional guttate psoriasis,
NLPnon-lesional plaque psoriasis.
45 kDa
C NLG LE N LG N LP NLE LP
a
b
Figure 3. Representative Western blot membrane showing total
protein
smears and specific HLA-C bands. (a) Ponceau staining of the
skin protein
extracts after transfer to a membrane. (b) Specific L31 signal.
CControl,LE lesional eczema, LG lesional guttate psoriasis, LP
lesional plaquepsoriasis, N normal, NLE non-lesional eczema, NLG
non-lesionalguttate psoriasis.
344 Journal of Investigative Dermatology (2007), Volume 127
L Carlen et al.HLA-C Expression Pattern in Psoriasis
-
genes in this locus are expressed in skin, some withdifferential
expression between psoriasis and normal skin(HCR, CDSN), there is
no conclusive explanation about theirfunctional role in psoriasis
pathogenesis (Bowcock, 2005).HLA-C has been considered a
susceptibility gene in itself or agenetic marker in linkage
disequilibrium with a psoriasisgene that has not yet been
identified (Helms et al., 2005). Ifintrinsic defects in this gene
are at the base of psoriasispathogenesis then there should be
evidence of altered HLA-Cexpression or function, otherwise this
molecule mightfacilitate the effect of third party factors such as
autoantigens.
Expression analysis has been critical to understanding therole
of HLA-C in other contexts than psoriasis but withthe common
denominator of immune regulatory functions forthis molecule. For
example, soluble MHC class I moleculesseem to regulate lymphocyte
survival through apoptosis,tumors can evade immune surveillance by
modifyingexpression of MHC class I molecules, and HLA-C
specificallycontributes to regulate maternal immune reactivity
againstfetal tissues (Le Bouteiller, 2001; Puppo et al., 2002;
Garcia-Lora et al., 2003). Global expression studies of RNA
andprotein do not indicate differential expression of HLA-C
inpsoriasis compared with normal or non-lesional skin (Zhouet al.,
2003; Bowcock, 2005; Carlen et al., 2005), incon-gruent with HLA-C
being a psoriasis susceptibility gene.However, qualitative as well
as quantitative differences areimportant in defining the role of
HLA-C in psoriasispathogenesis. Different patterns of expression
can emergeeven in the presence of similar levels of total RNA or
protein.Our findings indicate that HLA-C expression in
psoriasisdepends predominantly on infiltrating inflammatory
cells,whereas non-lesional and normal samples express higherlevels
of the protein in the epidermis.
It is expected that all nucleated cells express MHC class
Iamolecules because they contribute to preserving peripheralself
tolerance under normal circumstances, that is, noinflammation, no
infection (Zimmer et al., 2005). However,it is also expected that
the constitutive expression of HLA-Cshould be low (Neisig et al.,
1998). The present studydemonstrated intense staining on
infiltrating inflammatorycells and variable expression levels in
the epidermis.
The most striking feature of the HLA-C staining pattern inthis
study was the polarization of the signal towards thebasement
membrane and the reduced expression on supra-basal keratinocytes in
lesional psoriasis samples. It is knownthat overexpression of MHC
class I molecules can down-regulate effector immune cell functions
(Parham, 2004);therefore, reduced expression in psoriasis
epithelium mightcontribute to maintaining inflammation owing to
inefficientdownregulation of activated lymphocytes.
Why would HLA-C be restricted to the basal layer inpsoriasis
epidermis? MHC class I clustering into lipid rafts hasbeen reported
as a requirement to binding T-cell receptors(Lebedeva et al., 2004)
and lymphocytes are known to hometowards the skin during psoriasis
plaque formation (Davisonet al., 2001), but how these cells
interact with the epitheliumis less well documented. HLA-C
polarization in psoriasisepidermis might lead to a directional flow
of inflammatory
cells towards the tip of papillae, above which microabscessesare
typically formed (Kaneko et al., 1991). A relevantobservation
regarding this possibility is that the inflammatoryinfiltrate in
psoriasis does not seem to affect the structure ofthe basal cell
layer, except at the uppermost section ofpapillae where HLA-C
expression seems lower. In contrast,the basal cell layer in eczema
is characterized by infiltratinginflammatory cells throughout the
epidermis. We attemptedto measure the HLA-C signal intensity at the
basal layer ofpsoriasis papillae and counted the number of
infiltrating cellsthrough this layer; we observed an inverse
relationshipbetween these variables (data not shown). Since some
classI molecules, including HLA-C, are strong inhibitors of T andNK
cells, the reduced epithelial HLA-C expression might leadto
increased vulnerability to inflammation (Carosella et al.,2001; Le
Discorde et al., 2002; Trowsdale and Betz, 2006),therefore the
polarized HLA-C expression might be acompensatory mechanism to
reduce lymphocyte infiltration.Support for this view comes from
reproduction researchindicating that HLA-C and KIR mismatches
appear to fail atinhibiting NK/NK-T cells in pre-eclampsia, (Hiby
et al., 2004;Parham, 2004). Also in idiopathic bronchiectasis,
combina-tions of HLA-C and KIR associate with the
progressiveinflammatory damage to the lungs (Boyton et al., 2006).
Inaddition, viral regulation of class I gene expression plays arole
in viral persistence; for example, the UL18 protein
fromcytomegalovirus can induce HLA-C expression and inhibitHLA-A
and B, which can inhibit antiviral NK/NK-T cellresponses (Gewurz et
al., 2001).
There is mounting evidence indicating that immunesurveillance of
HLA-C by KIR-positive cells might be acontributing factor to
psoriasis pathogenesis (Martin et al.,2002; Luszczek et al., 2004;
Holm et al., 2005), butfunctional studies on this subject are still
lacking. However,it is important to remember that an antigen-driven
theory ofpsoriasis has been extensively discussed, where HLA-Cw6
isregarded as a cofactor in the initiation and/or maintenanceof
inflammation (Nickoloff et al., 2000; Lin et al., 2001;Johnston et
a.l, 2004).
What would be particular about HLA-Cw*0602 comparedto other
HLA-C alleles in this scenario? It is known that HLA-C expression
varies in relation to the identity of the codingalleles, but it is
not yet known if HLA-Cw*0602 has uniquefeatures that would affect
its protein expression. Although thisstudy does not specifically
target the psoriasis-associatedallele HLA-Cw*0602, it shows that
HLA-C expression innormal tissues varies depending on developmental
stage andhistological location and reveals that skin
epitheliumexpresses HLA-C.
The distinct HLA-C expression patterns in psoriasis andeczema
suggest a functional role in the specific psoriasisimmune response
and not only a general feature ofinflammation. It will be important
to obtain a detaileddescription of coding and non-coding sequences
of all allelesin order to identify differences at regulatory sites
and designfunctional studies that compare HLA-Cw*0602 to
otheralleles. This will increase our understanding of the role
ofHLA-C in psoriasis pathogenesis.
www.jidonline.org 345
L Carlen et al.HLA-C Expression Pattern in Psoriasis
-
MATERIALS AND METHODSTissue samples
Details regarding patients and controls were reported
previously
(Carlen et al., 2005). All study subjects provided written
consent and
all research procedures were performed in accordance with
the
Declaration of Helsinki Principles and with permission from
the
Ethics Committee of the Karolinska Institutet. Biopsies were
taken
from lesional and non-lesional skin from guttate and plaque
psoriasis
patients, lesional and non-lesional skin from nickel-induced
contact
eczema patients, and normal skin of healthy individuals. For
immunohistochemistry, five normal, 12 plaque psoriasis,
three
non-lesional plaque psoriasis, five lesional eczema, and five
non-
lesional eczema samples were analyzed. In addition, one
tonsil
sample from a guttate psoriasis patient affected by
recurrent
streptococcal throat infections, two normal adult colon and
one
fetal colon samples were used for immunohistochemistry. For
Western blot analysis, 10 normal, 12 plaque psoriasis, three
non-
lesional plaque psoriasis, seven guttate psoriasis, four
non-lesional
guttate, five lesional eczema, and four non-lesional eczema
samples
were considered.
Antibodies
The following antibodies were used: (1) Murine mAb L31 which
binds linear a1 domain epitope at position 67 of human
HLA-Calleles Cw1 through Cw8 and HLA-B8 and B51 (Setini et al.,
1996).
Alleles from Cw9 to Cw17, and B8 and B51 do not associate
with
psoriasis, therefore, under-or overestimation of expression
levels is
expected to cancel out between patients and controls. (2)
Murine
mAb W6/32 (Serotec, Oxford, UK) (Brodsky et al., 1979) which
binds MHC class I heavy chain associated with
b-2-microglobulin.(3) Murine mAb MCA2243 (NKVFS1) (Serotec)
(Spaggiari et al.,
2002) which recognizes a common epitope on CD158a and
CD158b (KIR2DL1, KIR2DL2), and (4) horse radish peroxidase-
conjugated rabbit polyclonal P0174 (DakoCytomation,
Glostrup,
Denmark) which binds human b2m. As secondary antibody,
horseradish peroxidase-conjugated goat anti-mouse IgG (Santa
Cruz,
sc-2005) was used.
Sample preparation
Skin biopsies were prepared as described previously (Carlen et
al.,
2005). In brief, the biopsies were kept frozen in liquid
nitrogen and
mechanically homogenized using a mortar and pestle. The
samples
were then solubilized in a buffer containing 7 M urea, 2 M
thiourea,
4% SDS, reducing agents, and protease inhibitors. Protein
concen-
tration was determined using the Bradford method (BioRad
Protein
Assay, BioRad, Stockholm, Sweden) (Bradford, 1976).
Western blot
Total protein extracts were separated, adding 7mg of each sample
to12% SDS-PAGE gels, and then electroblotted onto
nitrocellulose
membranes (Whatman Schleicher & Schuell, Kent, UK). The
membranes were then stained with 1% Ponceau S (Sigma, St
Louis,
MO) and scanned to check for loading and gel run quality.
After
blocking with 2% goat serum (Vector Laboratories, Burlingame,
CA)
and 1% BSA (Sigma, St Louis, MO) in Tris-buffered saline for 1
hour,
the membranes were incubated separately overnight with L31,
W6/
32, MCA2243, or b2m antibody diluted 1:500 in TBS with 1%Tween
20. The membranes were then incubated with horse radish
peroxidase-conjugated goat anti-mouse IgG (Santa Cruz
Biotechno-
logy, Santa Cruz, CA) (except for b2m) diluted 1:2,000 in
blockingbuffer followed by development using enhanced
chemilumines-
cence (Amersham Pharmacia Biotech, Sunnyvale, CA) and the
image acquired with a CCD camera (Fujifilm, Sendai, Japan).
The
immunoreactivity of the HLA-C and b2m antibodies was
testedagainst recombinant HLA-C and b2-microglobulin.
Unspecificbinding of the secondary antibody was prevented by
pre-incubation
in normal goat serum.
Immunohistochemistry
All biopsies were snap-frozen and stored at 701C until
furtherprocessed. Skin samples from lesional and non-lesional
eczema and
plaque and guttate psoriasis, and healthy skin were included in
the
same experiments. In short, 67-mm thick cryostat sections
wereincubated with L31, W6/32, and MCA2243 antibody at 1:200,
1:20,000, and 1:200 dilutions, respectively. All sections
were
stained using the avidin: biotinylated enzyme complex
technique
(Vector Laboratories, Burlingame, CA) following the
manufacturers
instructions, and counterstained with Gills III hematoxylin
solution.
As controls, tissue sections were processed in parallel
without
adding primary antibody. No reactivity for secondary antibodies
or
signal developing reagents was observed on control
histological
sections.
Cell culture
LCL721.221 cells were maintained in DMEM (Gibco-BRL Life
Technologies, Paisley, UK) supplemented with 10% FCS (fetal
calf
serum; Hy-Clone, Boule Nordic AB Huddinge, Sweden) and
antibiotics (PEST _ penicillin 50 U/l and streptomycin 50
mg/ml;
Gibco-BRL) at 5% CO2 at 371C. HLA-Cw3 and Cw4 transfectantswere
kept under G418 (GibcoBRL) selection. During stimulation
experiments, about 105 cells/well were grown in 12-well
plates
without selection with G418 for 2 days and then exposed to
1mg/mlE. coli-derived lipopolysaccharide. HLA-C expression was
detected
with the L31 antibody at time 0 and 16, 48 and 96 hours
after
stimulation. The cells were then transferred to slides, fixed
with
ethanol, and further processed as described in the
immunohisto-
chemistry section above.
Data analysis
Image analysis was carried out using Science Lab 99 L
Process
software, version 1.95 (Fujifilm, Sendai, Japan). Intensity
measure-
ments of specific signals were corrected by the total amount
of
protein in their corresponding lanes as derived from measurement
of
Ponceau staining intensities of the membranes. Variations in
luminosity between images were adjusted by using an HLA-C-
positive cell line as internal control on every membrane and
the density values were adjusted using the following
formulae:
Expression Index (corrected sample reading)/(corrected
Ponceaureading).
Corrected sample reading (sample density (Strongest
intergelcontrol/samples gel control))
Corrected Ponceau reading reading of control lane/sum ofreadings
from all lanes in gel
Statistical analysis was performed using the R language and
statistics package (Ihaka and Gentleman, 1996). Mean
expression
indexes from different sample groups were compared by fitting
a
346 Journal of Investigative Dermatology (2007), Volume 127
L Carlen et al.HLA-C Expression Pattern in Psoriasis
-
one-way analysis of variance model to the data, assuming a
significance level of 0.05. Then a paired t-test was performed
in
order to identify comparisons with significant differences.
CONFLICT OF INTERESTThe authors state no conflict of
interest.
ACKNOWLEDGMENTSWe thank Dr Eniko Sonkoly for critically
reviewing this manuscript, Anna-Lena Kastman for skilful technical
assistance with immunohistochemistry,Dr Gunther Weber for providing
HLA-C-positive cell lines and for assistancewith immunoblotting, Dr
Alberto Beretta for providing us with the L31HLA-C antibody, Dr
Jack Strominger, and Dr Hugh Reyburn for providingHLA-C constructs,
and Dr Kalle Malmberg and Dr Hans Gustav Ljunggren forHLA-C
transfected LCLs. This study was supported by funds from theSwedish
Research Council, the Swedish Heart and Lung Foundation, theSwedish
Psoriasis Association, Ake-Wibergs foundation, Serono,
Biogen,Schering-Plough, the National Psoriasis Foundation,
Karolinska UniversityHospital, and Karolinska Institutet.
REFERENCES
Baba E, Erskine R, Boyson JE, Cohen GB, Davis DM, Malik P et al.
(2000)N-linked carbohydrate on human leukocyte antigen-C and
recogni-tion by natural killer cell inhibitory receptors. Hum
Immunol 61:120218
Balendran N, Clough RL, Arguello JR, Barber R, Veal C, Jones AB
et al. (1999)Characterization of the major susceptibility region
for psoriasis atchromosome 6p21.3. J Invest Dermatol 113:3228
Bowcock AM (2005) The genetics of psoriasis and autoimmunity.
Annu RevGenomics Hum Genet 6:93122
Bowcock AM, Krueger JG (2005) Getting under the skin: the
immunogeneticsof psoriasis. Nat Rev Immunol 5:699711
Boyington JC, Brooks AG, Sun PD (2001) Structure of killer cell
immunoglo-bulin-like receptors and their recognition of the class I
MHC molecules.Immunol Rev 181:6678
Boyington JC, Sun PD (2002) A structural perspective on MHC
class Irecognition by killer cell immunoglobulin-like receptors.
Mol Immunol38:100721
Boyton RJ, Smith J, Ward R, Jones M, Ozerovitch L, Wilson R et
al. (2006)HLA-C and killer cell immunoglobulin-like receptor genes
in idiopathicbronchiectasis. Am J Respir Crit Care Med
173:32733
Bradford MM (1976) A rapid and sensitive method for the
quantitation ofmicrogram quantities of protein utilizing the
principle of protein-dyebinding. Anal Biochem 72:24854
Brodsky FM, Parham P, Barnstable CJ, Crumpton MJ, Bodmer WF
(1979)Monoclonal antibodies for analysis of the HLA system. Immunol
Rev47:361
Capon F, Allen MH, Ameen M, Burden AD, Tillman D, Barker JN et
al. (2004)A synonymous SNP of the corneodesmosin gene leads to
increasedmRNA stability and demonstrates association with psoriasis
acrossdiverse ethnic groups. Hum Mol Genet 13:23618
Carlen LM, Sanchez F, Bergman AC, Becker S, Hirschberg D,
Franzen B et al.(2005) Proteome analysis of skin distinguishes
acute guttate from chronicplaque psoriasis. J Invest Dermatol
124:639
Carlin LM, Eleme K, McCann FE, Davis DM (2001) Intercellular
transfer andsupramolecular organization of human leukocyte antigen
C at inhibitorynatural killer cell immune synapses. J Exp Med
194:150717
Carosella ED, Moreau P, Aractingi S, Rouas-Freiss N (2001)
HLA-G: a shieldagainst inflammatory aggression. Trends Immunol
22:5535
Davison SC, Ballsdon A, Allen MH, Barker JN (2001) Early
migration ofcutaneous lymphocyte-associated antigen (CLA) positive
T cells intoevolving psoriatic plaques. Exp Dermatol 10:2805
Garcia-Lora A, Algarra I, Collado A, Garrido F (2003) Tumour
immuno-logy, vaccination and escape strategies. Eur J Immunogenet
30:17783
Gewurz BE, Gaudet R, Tortorella D, Wang EW, Ploegh HL (2001)
Virussubversion of immunity: a structural perspective. Curr Opin
Immunol13:44250
Gross WL, Packhauser U, Hahn G, Westphal E, Christophers E,
Schlaak M(1977) Lymphocyte activation by streptococcal antigens in
psoriasis. Br JDermatol 97:52936
Gross WL, Vorwerk I, Westphal E, Christophers E, Hahn G, Schlaak
M (1983)HLA-related lymphocyte responsiveness in psoriasis. Int
Arch AllergyAppl Immunol 70:1516
Helms C, Saccone NL, Cao L, Daw JA, Cao K, Hsu TM et al.(2005)
Localization of PSORS1 to a haplotype block harboringHLA-C and
distinct from corneodesmosin and HCR. Hum Genet 118:46676
Henseler T, Christophers E (1985) Psoriasis of early and late
onset:characterization of two types of psoriasis vulgaris. J Am
Acad Dermatol13:4506
Hiby SE, Walker JJ, OShaughnessy KM, Redman CW, Carrington
M,Trowsdale J et al. (2004) Combinations of maternal KIR and
fetalHLA-C genes influence the risk of preeclampsia and
reproductivesuccess. J Exp Med 200:95765
Holm SJ, Carlen LM, Mallbris L, Stahle-Backdahl M, OBrien KP
(2003)Polymorphisms in the SEEK1 and SPR1 genes on 6p21.3 associate
withpsoriasis in the Swedish population. Exp Dermatol 12:43544
Holm SJ, Sakuraba K, Mallbris L, Wolk K, Stahle M, Sanchez FO
(2005)Distinct HLA-C/KIR genotype profile associates with guttate
psoriasis.J Invest Dermatol 125:72130
Ihaka R, Gentleman R (1996) A language for data analysis and
graphics.J Comput Graph Statist 5:299314
Jenisch S, Henseler T, Nair RP, Guo SW, Westphal E, Stuart P et
al.(1998) Linkage analysis of human leukocyte antigen (HLA)
markersin familial psoriasis: strong disequilibrium effects provide
evidencefor a major determinant in the HLA-B/-C region. Am J Hum
Genet 63:1919
Johnston A, Gudjonsson JE, Sigmundsdottir H, Love TJ,
Valdimarsson H(2004) Peripheral blood T cell responses to keratin
peptides that sharesequences with streptococcal M proteins are
largely restricted to skin-homing CD8(+) T cells. Clin Exp Immunol
138:8393
Kaneko F, Itoh N, Yoshida H, Suzuki M, Ono I (1991) The
cell-componentsand cytokines in the subcorneal microabscess of
psoriasis. Fukushima JMed Sci 37:10312
Krueger JG, Bowcock A (2005) Psoriasis pathophysiology: current
concepts ofpathogenesis. Ann Rheum Dis 64(Suppl 2):ii306
Langley RG, Krueger GG, Griffiths CE (2005) Psoriasis:
epidemiology, clinicalfeatures, and quality of life. Ann Rheum Dis
64(Suppl 2):ii1823(discussion ii24ii25)
Laurentaci G, Lomuto M, Favoino B (1982) Immunogenetic analysis
ofassociation between HLA antigens and psoriasis vulgaris:
population andfamily studies. Dermatologica 165:591600
Lebedeva T, Anikeeva N, Kalams SA, Walker BD, Gaidarov I, Keen
JH et al.(2004) Major histocompatibility complex class
I-intercellular adhesionmolecule-1 association on the surface of
target cells: implications forantigen presentation to cytotoxic T
lymphocytes. Immunology113:46071
Le Bouteiller P (2001) [HLA molecules, immunity and gestation].
GynecolObstet Fertil 29:52331
Le Discorde M, Moreau P, Rouas-Freiss N, Carosella E (2002)
[HLA-G:immune tolerance in normal and pathological physiology].
Pathol Biol(Paris) 50:4551
Lin WJ, Norris DA, Achziger M, Kotzin BL, Tomkinson B (2001)
Oligoclonalexpansion of intraepidermal T cells in psoriasis skin
lesions. J InvestDermatol 117:154653
Luszczek W, Manczak M, Cislo M, Nockowski P, Wisniewski A, Jasek
M et al.(2004) Gene for the activating natural killer cell
receptor, KIR2DS1, isassociated with susceptibility to psoriasis
vulgaris. Hum Immunol65:75866
Mallon E, Bunce M, Savoie H, Rowe A, Newson R, Gotch F et al.
(2000)HLA-C and guttate psoriasis. Br J Dermatol 143:117782
www.jidonline.org 347
L Carlen et al.HLA-C Expression Pattern in Psoriasis
-
Martin MP, Carrington M (2005) Immunogenetics of viral
infections. CurrOpin Immunol 17:5106
Martin MP, Nelson G, Lee JH, Pellett F, Gao X, Wade J et al.
(2002) Cuttingedge: susceptibility to psoriatic arthritis:
influence of activating killerIg-like receptor genes in the absence
of specific HLA-C alleles.J Immunol 169:281822
McCutcheon J, Gumperz J, Smith K, Lutz C, Parham P (1995) Low
HLA-Cexpression at cell surfaces correlates with increased turnover
of heavychain mRNA. J Exp Med 181:208595
McMichael AJ, Morhenn V, Payne R, Sasazuki T, Farber EM (1978)
HLA C andD antigens associated with psoriasis. Br J Dermatol
98:28792
Moretta L, Moretta A (2004) Killer immunoglobulin-like
receptors. Curr OpinImmunol 16:62633
Nair RP, Stuart P, Henseler T, Jenisch S, Chia NV, Westphal E et
al. (2000)Localization of psoriasis-susceptibility locus PSORS1 to
a 60-kb intervaltelomeric to HLA-C. Am J Hum Genet 66:183344
Neisig A, Melief CJM, Neefjes J (1998) Reduced cell surface
expression ofHLA-C molecules correlates with restricted peptide
binding and stableTAP interaction. J Immunol 160:1719
Nickoloff BJ, Schroder JM, von den Driesch P, Raychaudhuri SP,
Farber EM,Boehncke WH et al. (2000) Is psoriasis a T-cell disease?
Exp Dermatol9:35975
Nyulassy S, Hnilica P, Buc M, Guman M, Hirschova V, Stefanovic J
(1977)Subacute (de Quervains) thyroiditis: association with
HLA-Bw35antigen and abnormalities of the complement system,
immunoglobulinsand other serum proteins. J Clin Endocrinol Metab
45:2704
Oka A, Tamiya G, Tomizawa M, Ota M, Katsuyama Y, Makino S et al.
(1999)Association analysis using refined microsatellite markers
localizes asusceptibility locus for psoriasis vulgaris within a 111
kb segmenttelomeric to the HLA-C gene. Hum Mol Genet 8:216570
Parham P (2004) NK cells and trophoblasts: partners in
pregnancy. J Exp Med200:9515
Petersdorf EW, Longton GM, Anasetti C, Mickelson EM, McKinney
SK, SmithAG et al. (1997) Association of HLA-C disparity with graft
failure aftermarrow transplantation from unrelated donors. Blood
89:181823
Puppo F, Contini P, Ghio M, Indiveri F (2002) Soluble HLA class
I molecules/CD8 ligation trigger apoptosis of CD8+ cells by
Fas/Fas-ligandinteraction. Sci World J 2:4213
Rajagopalan S, Long EO (2005) Understanding how combinations of
HLA andKIR genes influence disease. J Exp Med 201:10259
Rubin LA, Amos CI, Wade JA, Martin JR, Bale SJ, Little AH et al.
(1994)Investigating the genetic basis for ankylosing spondylitis.
Linkage studieswith the major histocompatibility complex region.
Arthritis Rheum37:121220
Setini A, Beretta A, De Santis C, Meneveri R, Martayan A,
Mazzilli MCet al. (1996) Distinctive features of the alpha 1-domain
alpha helix ofHLA-C heavy chains free of beta 2-microglobulin. Hum
Immunol46:6981
Spaggiari GM, Contini P, Dondero A, Carosio R, Puppo F, Indiveri
F et al.(2002) Soluble HLA class I induces NK cell apoptosis upon
theengagement of killer-activating HLA class I receptors through
FasL-Fasinteraction. Blood 100:4098107
Suomela S, Elomaa O, Asumalahti K, Kariniemi AL, Karvonen SL,
Peltonen Jet al. (2003) HCR, a candidate gene for psoriasis, is
expressed differentlyin psoriasis and other hyperproliferative skin
disorders and is down-regulated by interferon-gamma in
keratinocytes. J Invest Dermatol121:13604
Trowsdale J, Betz AG (2006) Mothers little helpers: mechanisms
of maternal-fetal tolerance. Nat Immunol 7:2416
Winter C, Long E (1997) A single amino acid in the p58 killer
cell inhibitoryreceptor controls the ability of natural killer
cells to discriminatebetween the two groups of HLA-C allotypes. J
Immunol 158:40268
Wolf R, Ruocco V (1999) Triggered psoriasis. Adv Exp Med Biol
455:2215
Zhou X, Krueger JG, Kao MC, Lee E, Du F, Menter A et al. (2003)
Novelmechanisms of T-cell and dendritic cell activation revealed by
profilingof psoriasis on the 63,100-element oligonucleotide array.
PhysiolGenomics 13:6978
Zimmer J, Andres E, Donato L, Hanau D, Hentges F, de la Salle H
(2005)Clinical and immunological aspects of HLA class I deficiency.
QJM98:71927
348 Journal of Investigative Dermatology (2007), Volume 127
L Carlen et al.HLA-C Expression Pattern in Psoriasis