Embed Size (px)
Citation preview

Prcg. New-o-Psychopharmncd dl3bL Psychfat 1993. Vol. 17. pp. 87-104 0278 - 5845/93 $15.00
Printed tn Great Brttatn. AU rights reserved 0 1992 Per@mon Press Ltd
5-HT RECEPTOR LIGANDS IN ANIMAL MOD& OF ANXIETY, IMPULSMTYAND DEPRESSION: MULTIPLE MECHANISMS
OF ACTION?
RUDY SCHREIBER and JEAN DE VRY
Institute for Neurobiology, Department of Psychopharmacology, Troponwerke, Cologne, Germany
(Final form, February 1992)
1. 2. 2.1 2.2 2.3 2.4 3.
4.
5.
6.
Abstract Introduction
Contents
5-HT,,, Receptor Ligands in Anxiety, Impulsivity and Depression S-HT,,, Receptors: Selective Ligands and Distribution 5-HT, Receptor Ligands and Anxiety S-HT, Receptor Ligands and Impulsivity 5-HT, Receptor Ligands and Depression Are 5-HT,, Receptors dysfunctional in Anxiety, Impulsivity and Depression? Are 5-HT, Receptors Involved in the Mechanism of Action of Classical Anxiolytics and Antidepressants? Are 5-HT,/5-HT, Receptor Interactions Involved in the Therapeutic Effects of 5-HT,, Receptor Ligands ? Conclusions References
87 88 89 89 89 91 92
94
96
97 99 99
Abstract
Schreiber, Rudy and De Vry, Jean: 5-HT,, Receptor Ligands in Animal Models of Anxiety, Impulsivity and Depression: Multiple Mechanisms of Action? Prog. Neuro-Psychopharmacol. & Biol. Psychiat. 1993 17( 1): 87-104.
1. Preclinical and clinical studies suggest that 5-HT, receptor agonists are a new class of mixed axiolytics/antidepressants with, possibly, impulsivity reducing properties.
2. The anxiolytic effects of 5-HT, receptor agonists result predominantly from an interaction with presynaptic 5-HT, receptors (resulting in a decrease of serotonergic transmission), whereas the antidepressive and, possibly, the impulse control enhancing effects, result predominantly from an interaction with postsynaptic 5-HT,, receptors.
87

88 R. Schreiber and J. De Vry
3. These proposed mechanism(s) of action fit well with the generally held view that anxiety is the result of a hypersensitive S-HT system; whereas impulsivity and depression is the result of a hyposensitive 5-HT system.
4. However, it appears very likely that activation of pre- and postsynaptic S-HT, receptors is additionally involved in the antidepressive and impulse control enhancing effects, on the one hand, and in the anxiolytic effects of these compounds, on the other hand.
5. These latter, seemingly paradoxical, findings can be explained by assuming that (1) the presynaptic mechanism reflects an anxiolytic component in the animal models of impulsivity and depression, (2) antagonism of postsynaptic 5-HT,, receptors by these compounds contributes to their anxiolytic effects, (3) postsynaptic 5-HT,,and 5-HT, receptors have functionally opposing effects or, alternatively, that (4) downregulation of postsynaptic 5-HT, receptors contributes to the therapeutic effects of these compounds.
Keywords: animal models, anxiety, depression, 5-HT, 5-HT, receptors, 5-HT, receptors, impulsivity.
Abbreviations: 5,7_dihydroxytryptamine (5,7-DHT), Shydroxy-2-(di-n-propylamino)tetralin (&OH-DPAT), ethanol (EtOH), para-chlorophenylalanine (pCPA), serotonin (5-HT).
1. Introduction
Both, from clinical and preclinical studies, evidence is accumulating that serotonergic (5-HT)
neurotransmission may be involved in the etiology, expression and treatment of anxiety, impulsivity
and depression (Brown and van Praag 1991). However, many contradictory findings concerning
the dysfunction of 5-HT in these disorders have been reported. The discovery of multiple 5-HT
receptor subtypes in recent years and the development of selective ligands for these receptors, may
offer an opportunity to clarify the ambiguous role of 5-HT in these mental disorders and to treat
them more effectively (Cowen 1991, Marsden 1989). Currently, 5-HT receptors are classified into
seven subtypes, called 5-HT,.5-HT,,5-HT,,S-HT,,, 5-HT,, 5-HT, and 5-HT, receptors, each of
them having a particular brain distribution (Glennon 1990). 5-HT, receptors have been of
particular interest because they are involved in the regulation of emotional and behavioral
processes and possibly in the mechanism of action of a number of currently used treatments of
anxiety, impulsivity and depression (Dourish et al 1987, De Vry et al 1992, Lesch 1991). Therefore,
5-HT, receptor ligands may consist of a new class of psychotherapeutic agents with improved
efficacy and/or side-effect profile (Traber and Glaser 1987, De Vry et al 1991a, 1992). Concerning
the other 5-HT receptor subtypes, some evidence has been reported which suggests an
involvement of 5-HT, receptors and, possibly, 5-HT, and 5-HT,, receptors in (some of) these

5-I-IT,, ligands - anxiolytics/antidepressants 89
disorders (Cowen 1991). The present paper will briefly review preclinical studies on the
mechanism of action of SHT, receptor ligands in anxiety, impulsivity and depression and discuss
the possibility that an interaction between SHT,, and 5-HT, receptors may contribute to the
therapeutic effects of SHT,,, receptor ligands.
2. 5_HT,.,Receptor Ligands in Animal Models of Anxiety. Impulsivity and Depression
2.1. 5_HT,, Recemors: Selective Lipands and Distribution
The most widely used high affinity ligands for the 5-HT,, receptor are the pyrimidinylpiperazine
partial agonists buspirone (which has additional affinity for dopamine D, receptors), ipsapirone
and gepirone, and the aminotetralin full agonist 8-OH-DPAT. 5-HT,receptors are located both
presynaptically (somatodendritic autoreceptors) on the 5-HT cell bodies in the dorsal and median
raphe nuclei of the brainstem which innervate the forebrain; and postsynaptically, predominantly
in limbic structures like the hippocampus and the septum. Activation of presynaptic 5-HT,
receptors results in an inhibition of cell firing activity and a consequent decrease in 5-HT
neurotransmission. Therefore the physiological function of these receptors is to provide the brain
with an autoinhibitory feedback system controlling 5-HT neurotransmission. Activation of
postsynaptic 5-HT,, receptors results in neuronal inhibition of particular parts of the limbic system.
Due to their particular distribution, 5-HT,, receptors offer the possibility to control 5-HT
neurotransmission in a general manner (mediated by the presynaptic receptors) and neuronal
activity in the limbic system in a selective manner (mediated by the postsynaptic receptor). For
a review and further references on the localization, biochemistry, electrophysiology, and selective
ligands of 5-HT,, receptors; see: Glaser and De Vry 1992.
2.2. 5-HT, Receotor Lieands and Anxiety
Anxiolytic properties of 5-HT,, receptor ligands have been shown in a number of animal models
of anxiety (including models based on unconditioned and conditioned anxiety, conflict, novelty,
exploration and social behavior), as well as in man (for reviews; see: De Vry et al 1991a, 1992,
Schweizer and Rickels 1991). Although it is not firmly established whether 5-HT,, receptor ligands
are equally effective in all anxiety disorders (e.g., general anxiety disorder, panic disorder,
obsessive-compulsive disorder); until now, the majority of clinical studies supports a role for these
90 R. Schreiber and J. De Vry
ligands in the treatment of general anxiety disorder and, possibly, obsessive-compulsive disorder.
Most preclinical studies on the mechanism of action of these compounds in animal models of
anxiety have focussed on the relative contribution of pre- and postsynaptic S-HT, receptors after
acute administration. E.g., in a rat model of conditioned anxiety, local application of ipsapirone
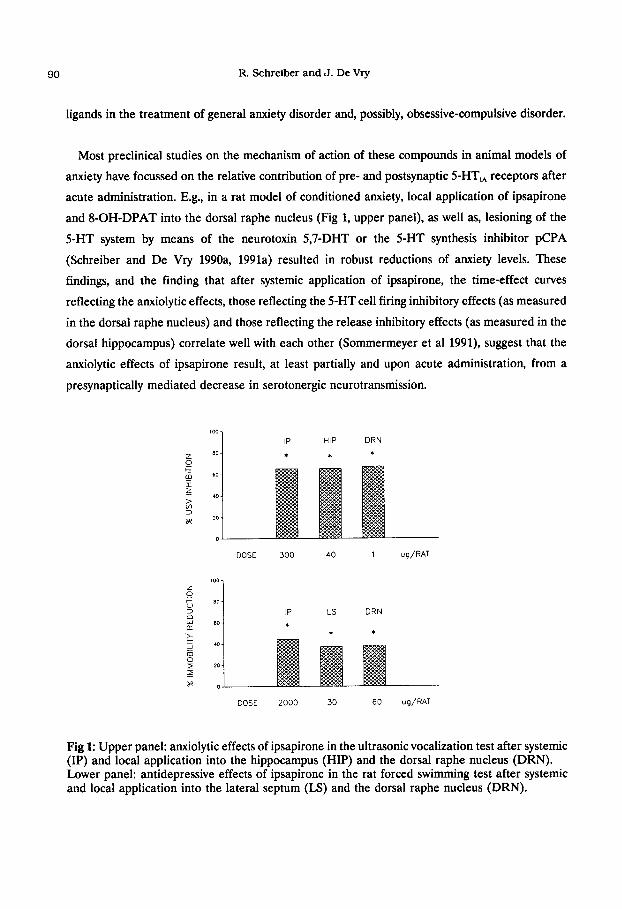
and 8-OH-DPAT into the dorsal raphe nucleus (Fig 1, upper panel), as well as, lesioning of the
5-HT system by means of the neurotoxin 5,7-DHT or the 5-HT synthesis inhibitor pCPA
(Schreiber and De Vry 1990a, 1991a) resulted in robust reductions of anxiety levels. These
findings, and the finding that after systemic application of ipsapirone, the time-effect curves
reflecting the anxiolytic effects, those reflecting the 5-HT cell firing inhibitory effects (as measured
in the dorsal raphe nucleus) and those reflecting the release inhibitory effects (as measured in the
dorsal hippocampus) correlate well with each other (Sommermeyer et al 1991), suggest that the
anxiolytic effects of ipsapirone result, at least partially and upon acute administration, from a
presynaptically mediated decrease in serotonergic neurotransmission.
1”“.
,P HIP DRN
DOSE 300 40 1 ug/RAT
DOSE 2000 30 60 ug/RAl
Fig 1: Upper panel: anxiolytic effects of ipsapirone in the ultrasonic vocalization test after systemic (IP) and local application into the hippocampus (HIP) and the dorsal raphe nucleus (DRN). Lower panel: antidepressive effects of ipsapirone in the rat forced swimming test after systemic and local application into the lateral septum (LS) and the dorsal raphe nucleus (DRN).

533~~~ ligands - anxiolytics/ant.tdepressants 91
However, an additional involvement of postsynaptic S-HT, receptors is very likely, because
microinjection of 8-OH-DPAT and ipsapirone into the hippocampus leads to a similar, although
less potent, anxiolytic response as compared to dorsal raphe application (Fig 1, upper panel;
Schreiber and De Vry 1990a, 199la).Therefore it appears that both presynaptic and (possible to
a lesser extent) postsynaptic S-HT,receptors are involved in the acute anxiolytic effects of 5-HT,
receptor ligands (De Vry et al 1992, Marsden 1989). This dual contribution of pre- and
postsynaptic receptor systems can, however, lead to an apparent paradox. Indeed, how can it be
understood that a (presynaptically mediated) reduction of S-HT neurotransmission has a similar
functional effect as a (postsynaptically mediated) increase in S-HT neurotransmission?
A possible explanation is based on the assumption that the intrinsic activity of these S-HT,
receptor ligands is different at pre- and postsynaptic receptors (i.e., that these compounds have
ago&tic properties at presynaptic receptors and antagonistic (or partial agonistic) properties at
postsynaptic receptors (Glaser and De Vry 1992)). According to this view, the anxiolytic activity
is the additive result from a reduction of S-HT neurotransmission (mediated by an agonistic
activity at presynaptic receptors) and an antagonistic activity at postsynaptic receptors.
Alternatively, anxiolytic activity is explained by assuming that activation of different postsynaptic
5-HT receptor subtypes has functionally opposite effects. According to this hypothesis, activation
of S-HT, receptors, which results in hyoeroolarisation of the postsynaptic neuron (neuronal
inhibition) is anxiolytic; whereas activation of non-5-HT,, receptors (5HT,, S-HT,, or 5-HT,?)
which results in deoolarisation of the postsynaptic neuron (neuronal excitation) is “anxiogenic” (see
further). A common feature of both hypotheses, however, is that overactivity of the 5-HT system
in particular brain areas is involved in pathological anxiety and that reduction of this overactivity
is anxiolytic. In line with this suggestion is the recent finding that rats in an aversive environment
(elevated X-maze) have increased 5-HT levels in the ventral hippocampus and that systemic
application of ipsapirone attenuates the elevated 5-HT activity (Wright et al 1991a).
2.3. 5-HT2.y
Preclinical studies have supported the concept that 5-m dysfunction may be involved in
impulsivity and impulse control disorders and have suggested that 5-HT, receptor ligands may
affect impulsivity (for reviews; see: Soubrie 1986, De Vry et al 1991a, 1992). E.g., anti-aggressive

92 R. Schreiber and J. De Vry
effects of these compounds have been reported in animal models for territorial aggression
(Schuurman et al 1991), shock-induced aggression (De Vry et al 1991a), maternal aggression
(Olivier et al 1989) and isolation-induced aggression (White et al 1991). Reductions in the intake
of (and possible craving for) alcohol and nicotine have been obtained in preclinical and some
clinical studies (for reviews; see: De Vry et al 1992, Tollefson 1991).
The finding that local application of 8-OH-DPAT into the dorsal raphe nucleus has
anti-aggressive effects (Mos and Olivier 1991) suggests that a reduction in 5-HT neurotransmission
is involved in the anti-aggressive effects of 5-HT, receptor ligands. Evidence for a similar
mechanism of action for the ethanol (EtOH) preference reducing activity of 8-OH-DPAT and
ipsapirone has been found in selectively inbred Wistar rats with high preference for EtOH over
water in a two bottle free choice situation (Schreiber et al 1991). However, because anxiety may
play a role in these animal models of impulsivity, it remains unclear to what extent the suggested
presynaptic mechanism does not merely reflect such an anxiolytic component (see above).
Moreover, it has been suggested that a reduction in 5-HT neurotransmission may actually increase
impulsivity (Soubrie 1986, Roy et al 1987). Such a dysfunction of 5-HT could possibly explain the
observed supersensitivity of postsynaptic 5-HT,, receptors in isolated rats and alcohol preferring
rats (Wright et al 1991b, Wong and Murphy 1989). However, it remains to be shown that local
application of 5-HT,, receptor ligands in brain regions rich in postsynaptic 5-HT, receptors leads
to efficacy in animal models of impulsivity.
2.4. 5-HT,. Receotor Lieands and Depression
Antidepressive activity has been reported for several 5-HT,, receptor ligands, both at the
preclinical and clinical level (for reviews; see: De Vry et al 1991a, 1992). In general, antidepressive
effects have been obtained in a variety of animal models based on stress induced behavioral
deficits (i.e., forced swimming test, learned helplessness and restraint stress).
Studies on the mechanism of action for the antidepressive effects of 5-HT, receptor ligands
have, as in case of the anxiolytic effects, focussed on the relative contribution of pre- and
postsynaptic receptors. Similar to the effects observed in an animal model of anxiety,
antidepressive effects were obtained after microinjection of ipsapirone and 8-OH-DPAT into the
dorsal raphe nucleus in the forced swimming depression test (Fig 1, lower panel; Schreiber and

5-HT,, ligands - anxiolytks/antidepressants 93
De Vry 199Ob). However, as compared to the anxiolytic effects, considerably higher doses were
needed after local application of ipsapirone (and 8-OH-DPAT) in the dorsal raphe; suggesting that
the contribution of presynaptic 5-HT, receptors is relatively less important in the case of the
antidepressive effects. Additionally, lesioning of the brain 5-HT system by means of either
5,7-DHT or pCPA had only small antidepressive effects on its own (Schreiber and De Vry 1991b),
again suggesting that presynaptic 5-HT, receptors are only weakly involved in the antidepressive
effects of these compounds. Moreover, as in the case of the animal models of impulsivity, it cannot
be ruled out that the apparent contribution of presynaptic 5-HT, receptors reflects an anxiolytic
component of the test. Alternatively, it has been suggested that activation of presynaptic 5-HT,
receptors leads to disinhibition of dopaminergic activity, which, in turn, would be responsible for
the effects of 5-HT, receptor agonists in this animal model of depression (Cervo et al 1988).
Because the antidepressive effects of 5-HT,, receptor ligands appear to correlate with their
intrinsic activity at postsynaptic 5-HT,, receptors (De Vry et al 1991a, 1992) and because these
compounds appear to be more potent after local application into the lateral septum (Fig 1, lower
panel) than after local application into the dorsal raphe, it is suggested that postsynaptic 5-HT,
receptors are strongly involved in the antidepressive effects. This suggestion is supported by the
finding that in the learned helplessness model of depression, local application of buspirone and
8-OH-DPAT in the septum, but not in the raphe nuclei, shows antidepressive effects (Martin et
al 1991). As in the case of impulsivity, a major contribution of postsynaptic 5-HT, receptors
appears to be in accordance with the hypothesis which states that 5-HT neurotransmission is
reduced in depression and that an increase in 5-HT neurotransmission results in antidepressive
effects (Coppen 1967).
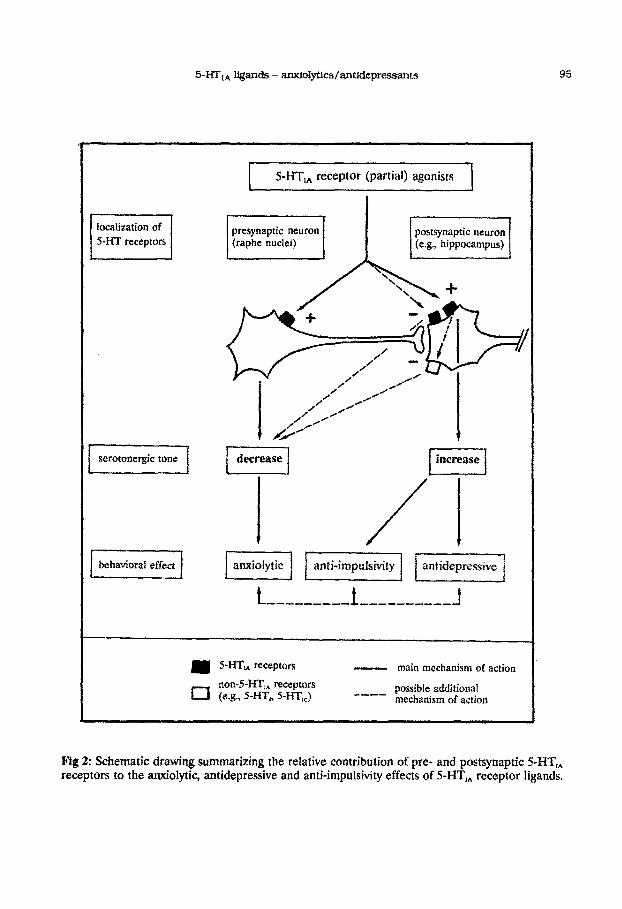
Taken together, it is concluded that 5-HT,, receptor ligands exert their anxiolytic effects
predominantly by activation of presynaptic 5-HT,, receptors (resulting in a general decrease of
serotonergic activity, Fig 2) and their antidepressive, and possibly impulse control enhancing
effects, predominantly by activation of postsynaptic 5-HT, receptors (resulting in-a selective
increase of serotonergic activity in particular brain structures, Fig 2). Because ample evidence
suggests that serotonergic activity is enhanced in anxiety and reduced in impulsivity and
depression, the proposed mechanism of action of 5-HT,, receptor ligands in anxiety, impulsivity
and depression is in line with currently accepted hypotheses concerning the role of 5-HT in these
disorders.

94 R. Schreiber and J. De Vxy
However, some data which apparently do not fit into these hypotheses have also been obtained.
First, activation of presynaptic S-HT,, receptors leads to some efficacy in several animal models
of impulsivity and depression (Fig 1, lower panel). A possible explanation for these findings is that
an anxiety component is present in these animal models and that the presynaptic component
indentified in these models represents the anxiolytic activity of S-HT,, receptor ligands. Second,
activation of postsynaptic 5-HT, receptors leads to some efficacy in several animal models of
anxiety. In order to explain these findings, it has been suggested that antagonistic activity of these
compounds at postsynaptic 5-HT, receptors contributes to their anxiolytic effects. Alternatively,
activation of postsynaptic 5-HT,, and non-5HT,, receptors (5-HT,, 5-HT,, or 5-HT,?) may have
functionally opposite effects (i.e., activation of postsynaptic 5-HT, receptors and non-5-HT,
receptors leads to anxiolytic and anxiogenic effects, respectively; see Fig 2 and further).
3. Are 5-HT,. Receptors Dysfunctional in Anxiety. Imnulsivity and Depression?
The 5-HT, receptors which are involved in the hypothermic and ACTH/cortisol increase
induced by ipsapirone appear to be subsensitive in patients with panic disorder, but not in those
with obsessive-compulsive disorder (Lesch 1991). Although it may be premature to draw firm
conclusions about the pre- and postsynaptic location of the dysfunctional 5-HT,receptors in panic
disorder (because the synaptic localization of the 5-HT,, receptors involved in these responses is
not unequivocally established in human), subsensitivity of 5-HT,, receptors could tentatively be
interpretated as an adaptive response to increased 5-HT levels. It remains to be determined to
what extent dysfunctional 5-HT,, receptors are involved in other anxiety disorders (e.g., general
anxiety disorder) and to what extent challenge tests with 5-HT,,receptor ligands may become ”
diagnostic tools ” for these disorders.
Post-mortem studies revealed increased postsynaptic 5-HT, receptor densities in the frontal
cortex and hippocampus of suicides and smokers, respectively (Arango et al 1991, Benwell et al
1990). On the other hand, in patients with impulsive aggressive behaviour, buspirone-induced
prolactin release was attenuated compared to healthy controls (Coccaro et al 1990). In animal
models of impulsivity, increased densities of postsynaptic 5-HT,, receptors were found in the
cortex and hippocampus of alcohol preferring rats; whereas presynaptic 5-HT, receptor densities
in the brain stem were decreased (Wong and Murphy 1989).
95
S-HT, receptor (partial) agonists
m WIT, receptors
non-s-HT, receptors a (e.g., 5-ST, S-HT&
Fig 2: Schematic drawing summar~zin~ the relative ~ntribut~o~ of pre- and postsynaptic S-HI;, receptors to the anxiolytic, antidepressive and anti-impulsivity effects of 5-HT, receptor ligands.

96 R. Schrelber and J. De Yry
Similar MI’T.,receptor changes were found in isolation-reared rats (Wright et al 1989, 1991b).
These S-HT, receptor changes can tentatively be explained as adaptive responses to a
dysfunctional 5-HT system in impulsivity and impulse control disorders (for review; see: Coccaro
1989).
In depressed suicides, it has been suggested that postsynaptic 5-HT, receptor densities are
increased in the frontal cortex (Lowther et al 1990). In addition, postsynaptic 5-I-IT1, receptors
were found to be supersensitive in an animal model of depression (Kennett et al. 1985). As in the
case of the impulse control disorders, these preliminary data can be interpretated as supportive
evidence of a dysfunctional serotonergic system in depression (Coppen 1967). However, in patients
with unipolar depression, ipsapirone-induced hypothermia and ACTH/cortisol increase was also
blunted; suggesting that pre- and/or postsynaptic 5-HT,, receptors were subsensitive in this
disorder (Lesch 1991). Therefore it may be that upregulated and/or supersensitive 5-HT,
receptors are not a common feature of all types of depression. Clearly, as in the case of anxiety
disorders, further clinical and preclinical studies are needed to clarify this issue and the complex
relationship between adaptation of the 5-HT,, receptor system and apparent hyper- or
hyposensitivity of the 5-HT system.
4. Are 5-HT,, R ce r Inv Iv 1
Antidemessants?
Because 5-HT, receptor agonists and benzodiazepines reduce serotonergic activity, it has been
suggested that this effect may be involved in the anxiety reducing properties of these compounds
(Section 2, Stein et al 1975, Thiebot et al 1982; but see also: Thiebot 1986). Accordingly, the
anxiolytic activity of diazepam in the elevated X-maze coincides with a decrease of 5-HT release
in the ventral hippocampus, as measured by in vivo microdialysis (Wright et al 1991a) and, as
mentioned before, it was found in a test of conditioned anxiety that the time-effect curves
reflecting the anxiolytic effects of ipsapirone correlated well with those reflecting the release
inhibitory effects in the dorsal hippocampus (Sommermeyer et al 1991). However, the effects of
benzodiazepines in animal models of anxiety cannot be antagonized with 5-HT, receptor
antagonists; suggesting that 5-HT,, receptors are not involved in the anxiolytic effects of
benzodiazepines (Glaser et al 1992). Because benzodiazepines and 5-HT, receptor agonists induce
a desynchronization of hippocampal rhythmical slow activity, a phenomenon which possible results
from reduced 5-HT activity in the hippocampus and which is thought to reflect anxiolysis (Hirose

5-HTIA ligands - anxiolytics/antidepressants 97
et al 1990, MC Naughton and Coop 1991); it is concluded that the mechanism of action for the
anxiolytic effects of benzodiazepines and S-HT,,, receptor ligands may, at least partially, overlap
and share a common final pathway.
With respect to antidepressants, it has been reported that repeated administration of tricyclics,
MAO-inhibitors, 5-I-IT reuptake inhibitors and electroconvulsive shock affect pre- and/or
postsynaptic S-HT,, receptor function, suggesting that S-HT, receptors may be involved in the
mechanism of action of currently used antidepressants (for review; see: De Vry et al 1992).
However, although the evidence for such an involvement appears stronger in the case of the
antidepressants than in the case of the anxiolytics, the issue remains controversial and needs to
be clarified further.
5. Are 5-HT,.!5-HT, Receptor Interactions Involved in the Therapeutic Effects of
5_HT,, Receptor Ligands?
Chronic treatment with most antidepressants results in a downregulation of 5-HT, receptors
(Kendall and Nahorski 1985, Peroutka and Snyder 1980). In addition, it is well established that
many currently used antidepressants are effective in the treatment of panic disorder and obsessive-
compulsive disorder. Therefore, it has been proposed that 5-HT, receptor mechanisms may be
involved in anxiety disorders (Deakin 1989). Moreover, because (1) both repeated and acute
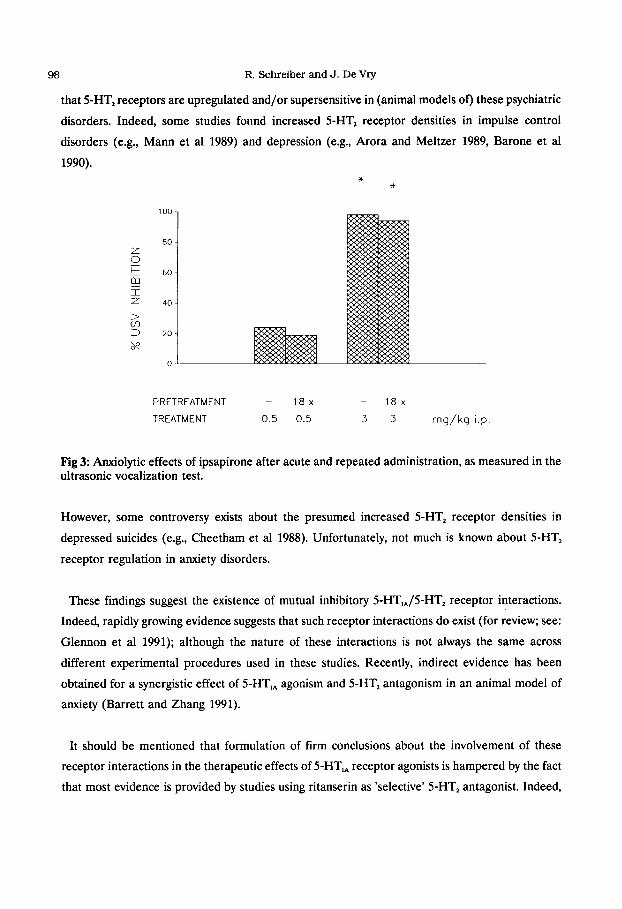
administration of 5-HT, receptor agonists (De Vry et al 1991b, Fig 3) and chronic application of
the 5-HT,,, antagonist ritanserin (Wright et al 1991~) reduces anxiety and (2) chronic application
of these compounds results in downregulation of 5-HT, receptors (Benjamin et al 1990, Leysen
et a1.1986, Schechter et al 1990, Frazer et al 1991), it is suggested that downregulation of 5-HT,
receptors may be a common mechanism of action for the anxiolytic effects of 5-HT, receptor
agonists and 5-HT, receptor antagonists. Therefore, 5-HT, receptor downregulation induced by
repeated administration of 5-HT,,, receptor ligands may add to, or substitute for, the anxiolytic
effects induced by 5-HT, receptor activation. It is tempting to suggest that such a mechanism is
related to the clinical observation that the anxiolytic effects of 5-HT, receptor ligands require at
least 2 weeks to fully develop.
If downregulation of 5-I-IT, receptors by 5-HT, receptor agonists contributes to the mechanism
of action of these compounds in anxiety and, possibly, impulsivity and depression, it is expected
98 R. Schreiber and J. De Vry
that S-I-IT, receptors are upregulated and/or supersensitive in (animal models of) these psychiatric
disorders. Indeed, some studies found increased .5-HT, receptor densities in impulse control
disorders (e.g., Mann et al 1989) and depression (e.g., Arora and Meltzer 1989, Barone et al
1990).
*
60
PRETREATMENT - 18 x _ 18 x
TREATMENT 0.5 0.5 3 3 mdb i.p
Fig 3: Anxiolytic effects of ipsapirone after acute and repeated administration, as measured in the ultrasonic vocalization test.
However, some controversy exists about the presumed increased S-HT, receptor densities in
depressed suicides (e.g., Cheetham et al 1988). Unfortunately, not much is known about S-I-IT,
receptor regulation in anxiety disorders.
These findings suggest the existence of mutual inhibitory S-HT,/S-HT, receptor interactions.
Indeed, rapidly growing evidence suggests that such receptor interactions do exist (for review; see:
Glennon et al 1991); although the nature of these interactions is not always the same across
different experimental procedures used in these studies. Recently, indirect evidence has been
obtained for a synergistic effect of S-HT,, agonism and S-HT, antagonism in an animal model of
anxiety (Barrett and Zhang 1991).
It should be mentioned that formulation of firm conclusions about the involvement of these
receptor interactions in the therapeutic effects of SHT, receptor agonists is hampered by the fact
that most evidence is provided by studies using ritanserin as ‘selective’ 5-HT, antagonist. Indeed,

5-HT,, ligands - anxiolytics/antidepressants 99
because of the additional 5HT,,receptor blocking properties of this compound, it cannot be ruled
out that S-I-IT,&HT,,interactions were studied. The possible relevance of such a receptor
interaction is emphasized by the observation that activation of 5-I-IT,, receptors by the 5-
I-IT,&HT,, receptor agonist mCPP appears to he anxiogenic in the rat (for review; see: Curzon
et al 1991). Mutual inhibitory interactions between S-HT,, and 5-HT,, receptors have already been
proposed for some components of the S-OH-DPAT-induced 5-HT behavioural syndrome
(Berendsen 1991). Nevertheless, the receptor downregulation obtained after repeated treatment
with ipsapirone is observed for the 5-HT,, but not the 5-HT,,. receptor (Gozlan and Hamon,
personal communication).
6. Conclusions
It is suggested that - in line with current hypotheses about the role of 5-HT in anxiety on the
one hand (serotonergic hyperactivity) and impulsivity and depression on the other hand
(serotonergic hypoactivity) - the anxiolytic activity of 5-HT, receptor agonists is induced by a
reduction of serotonergic activity (predominantly mediated by activation of presynaptic 5-HT,
receptors, and probably also by inactivation of 5-HT, receptors, achieved either by mutual
inhibitory postsynaptic 5-HT,/5-HT, receptor interactions, or by downregulation of 5-HT,
receptors upon repeated administration); whereas the antidepressive activity and, possibly, the
impulse control enhancing activity, is predominantly induced by activation of postsynaptic 5-HT,,
receptors (either directly or by attenuation of the inhibitory effect of 5-HT, receptors). According
to this hypothesis a careful analysis of the interaction between 5-HT receptor subtypes may add
to a better understanding of the therapeutical effects of 5-HT,, receptor ligands and possibly lead
to the development of more efficient and safer drugs.
References
ARORA, RC., and MELTZER, H.Y. (1989) Increased serotonin, (5-HT,) receptor binding as measured by [“HI-lysergic acid diethylamide ([‘HI-LSD) in the blood platelets of depressed patients. Life Sci. 44: 725-734.
ARANGO, V., MILLER, W.E., MILLER, M.L., UNDERWOOD, M.D., SMITH, R.W., and MANN., J.J. (1991) Quantitative autoradiography of 5-HT, binding in suicide. Sot. Neurosci. Abstr. 12: 86.3.
BARONE, P., ATGER, F., MARTIN, F., PUECH, P., and FILLION, G. (1990) Increased number of 5-I-IT, receptors in the “learned helplessness” rat. Eur J Pharmacol m: 1900-1901

100 R. Scbreiber and J. De Vry
BARRETT, J.E. and ZHANG, L. (1991) Anticonfhct and discriminative stimulus effects of the 5-HT, compounds WY-47846 and WY-48,723 and the mixed 5-HT,, agonist/S-H’& antagonist WY- 50,324 in pigeons. Drug Dev. Res. 24: 179-188,
BENJAMIN, D,, SAIFF, EL, LAL, H., and COUPET, J, (1990) Downregulation of brain S-HT., receptors underhes anxiolytic effect produced by sustained treatment with gepirone. Sot. Neurosci. Abstr. M: 547.8.
BENWELL, M.E.M., BALFOUR, D.J.K., and ANDERSON, J.M. (1990) Smoking associated changes in the serotonergic systems of discrete regions of human brain. Psychopharmacalogy &X& 68-72.
BERE~SEN, H.M.G. (1991) Behavioural consequences of selective activation of 5-HT receptor subtypes; possible imp~i~t~ons for the mode of action of antidepressants. Ph. D. Thesis, Univers, of Groningen, The Netherlands.
BROWN, S.L., and VAN PRAAG, H.M. (1991) The Role of Serotonin in Psychiatric Disorders, Clin. and Exp. Psych. Monograph. Brunner/Mazel Publishers, New York, No.4.
CERVO, L,, GRIGNASCHI, G., and SAMANIN, R. (1988) 8-Hydro~-2-(di-u-propylamino)tetralin, a selective serotonin,~ receptor agonist, reduces the immobility of rats in the forced swimming test by acting on the nucleus raphe dorsalis. Eur. J. Pharmacol. m: 53-59.
CHEE~M, SC., CROMPTGN, M.R., KATONA, C.L.E., and HORTON, R.W. (1988) Brain 5-HT, receptor binding sites in depressed suicide victims. Brain Res. !l,&?$ 272-280.
COCCARO, E.F. f 1989) Central serotonin and impulsive aggression. Br. J. Fsy~hiat~ ~@uppl
8): 52-62.
COCCARO, E.F., GABRIEL, S., and SIEVER, L.J. (1990) Buspirone challenge: preliminary evidence for a role for central S-HT,, receptor function in impulsive aggressive behavior in humans. Psychopharmacol. Bull. a: 393-405.
COPPEN, A. (1967) The biochemistry of affective disorder, Br. J. Psychiat. X$: X237-1264.
COWEN, P.J. (1991) Serotonin receptor subtypes: impfications for psychopharmacoio~. Br. J. Psychiat. m (suppl. 12): 7-14,
CURZON, G., GIBSON, EL, ~NNEDY, A.J., KENNETH, G.A., SARNA, G.S., and WHITTON, P. (1991). Anxiogenic and other effects of mCPP* a S-H& agonist. In: New Concepts in Anxiety, M. Briley and SE. File (Eds.), pp 154-167, Mae Miflan Fress, London.
DE VRY, J., GLASER, T., SCHUURMAN, T., SCHREIBER, R., and TRABER, J. (1991a) 5-HT, receptors in anxiety. In: New Concepts in Anxiety, M. Briley and S.E. File (Eds.), pp 94-129, Mat Millan Press, London.
DE VRY, J, SCHREIBER, R., and GLASER, T. (1991b) Behavioral pha~a~o~o~ of the 5-HTti receptor ligands ipsapirone and &OH-DPAT in rats after repeated a~i~stration. Sot. Neurosci. Abstr. fz: 116.4.

5-HT,A ligands - anxiolytlcs/antidepressants 101
DE VRY, J., SCHREIBER, R., GLASER, T., and TRABER, J. (1992) Behavioural pharmacology of S-I-IT, agonists: animal models of anxiety and depression. In: Serotonin, Receptors in Depression and Anxiety, S.M. Stahl et al (Eds.), pp 55-81, Raven Press, New York.
DEAKIN, J.F.W. (1989) 5-HT receptor subtypes in depression. In: Behavioural Pharmacology of 5-I-H, P. Bevan, A.R. Cools and T. Archer (Eds.), pp 179-203, Lawrence Erlbaum associates, London.
DOLJRISH, C.T. (1987) Brain 5-HT, receptors and anxiety. In: Brain 5-HT, Receptors, Behavioural and Neurochemical Pharmacology, C.T. Dourish, S. Ahlenius and P.H. Hutson (Eds.), pp 261-277, Ellis Hot-wood Publishers.
FRAZER, A., HENSLER, J., and HAUPTMANN, M. (1991) Effects of typical and novel antidepressants on serotonergic responsiveness and receptors. Biol. Psychiat. B: 97s.
GLASER, T., DE VRY, J. (1992) Neurobiology of 5-HT,,-receptors. In: Serotonin, Receptors in Depression and Anxiety, S.M. Stahl et al (Eds.), pp 35-53, Raven Press, New York.
GIASER, T., GREUEL, J.M., and DE VRY, J. (1992) 5-HT,, Rezeptoren als Angriffspunkt fur neuartige Anxiolytika. In: G. Laux and P. Riederer (Eds.), Neuro-Psychopharmaka, Springer Verlag, Wien, in press, New York.
GLENNON, R. (1990) Serotonin receptors: clinical implications. Neurosci. 8c Behav. Rev. 14 : 35-47.
GLENNON, R., DARMANI, N.A., and MARTIN, B.R. (1991) Multiple populations of serotonin receptors may modulate the behavioral effects of serotonergic agents. Life Sci. 3: 2493-2498.
HIROSE, A., TSUJI, R, SHIMIZI, H., TATSUNO, T., TANAKA, H., KUMASAKA, Y., and NAKAMURA, M. (1990) Inhibition of 8-hydroxy-2-(di-n-propylamino)tetralin and SM-3997, a novel anxiolytic drug, of the hippocampal rhythmical slow activity mediated by 5-hydroxytryptamine, receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. &I: 8-13.
KENNETT, G.A., DICKINSON, S.L., and CURZON, G. (1985) Enhancement of some 5-HT-dependent behavioural responses following repeated immobilization in rats. Brain Res. &jQ: 253-263.
KENDALL, D.A. and NAHORSKI, S.R. (1985) 5-Hydroxytryptamine stimulated inositol phospholipid hydrolysis in rat cerebral cortex slices: Pharmacological characterization and effect of antidepressants. J. Pharmacol. Exp. Ther. m: 473
LESCH, K.P. (1991) 5-HT, receptor responsivity in anxiety disorders and depression. Prog. Neuro-Psychopharmacol. & Biol. Psychiat. 15: 723-733.
LEYSEN, J.E., VAN GOMPEI, P., GOMMEREN, G., WOESTENBORGHS, R., and JANSSEN, P.A.J.( 1986) Downregulation of serotonin-S* receptor sites in rat brain by chronic treatment with the serotonin-S, antagonists: ritanserin and setoperone. Psychopharmacology 88: 434-444.
LOWTHER, S., DE PAERMENTIER., CHEETHAM, S.C., CROMPTON, M.R., KATONA, C.L.E., and HORTON, R.W. (1990) 5-HT, receptor binding in post-mortem brain samples from

102 R. Schreiber and J. De Vry
antidepressant-free and antidepressant-treated suicides and controles. Proceedings of the 2nd IUPHAR Sattelite Meeting on Serotonin, Basel: P164.
MANN, J.J., ARANGO, V., MARZUK, P.M., THECCANAT, S., and REIS, D.J. (1989) Evidence for the 5-HT hypothesis of suicide: a review of post-mortem studies. Br. J. Psychiat. (suppl.) 155: 7-14.
MARSDEN, C.A. (1989) S-Hydroxytryptamine receptor subtypes and new anxiolytic drugs: an appraisal. In: Psychopharmacology of Anxiety, P. Tyrer (Ed.), pp 3-27, Oxford University Press.
MARTIN, P., TISSIER, M.H., ADRIEN, J., and PUECH, A.J. (1991) Antidepressant-like effects of buspirone mediated by the S-HT,, postsynaptic receptors in the learned helplessness paradigm. Life Sci. a: 25052511.
McNAUGHTON, N., and COOP, C.F. (1991) Neurochemically dissimilar anxiolytic drugs have common effects on hippocampal rhythmic slow activity. Neuropharmacol. 3: 855-863.
MOS, J., and OLIVIER, B. (1991) On the site of action of eltoprazine, TFMPP and 8-OH-DPAT in reducing aggression in rats. Biol. Psychiatry 3: 209s.
OLIVIER, B., MOS, J., TULP, M., SCHIPPER, J., and BEVAN, P. (1989) Modulatory action of serotonin in aggressive behaviour. In: Behavioural Pharmacology of 5-HT, P. Bevan, A.R. Cools and T. Archer (Eds.), pp 89-115, Lawrence Erlbaum Associates, London.
PEROUTKA, S.J., and SNYDER, S.H. (1980) Long term antidepressant treatment lowering spiperidol labelled serotonin receptor binding. Science 210: 88
ROY, A., VIRKKUNEN, M., and LINNOILA, M. (1987) Reduced central serotonin turnover in a subgroup of alcoholics. Prog. Neuro-Psychopharmacol. & Biol. Psychiat. 11: 173-177.
SCHECHTER, L.E., BOLANOS, F.J., GOZLAN, H., LANFUMEY, L., HAJ-DAHMANE, S., LAPORTE, A.M., FATTACCINI, C.M., and HAMON, M. (1990) Alterations of central serotonergic and dopaminergic neurotransmission in rats chronically treated with ipsapirone: biochemical and electrophysiological studies. J. Pharmacol. & Exp. Ther. z: 1335-1347.
SCHREIBER, R., and DE VRY, J. (1990a) Neuroanatomical correlate of the anxiolytic effects of the 5-HT, receptor ligands 8-OH-DPAT, ipsapirone and buspirone in the rat. Psychopharmacology Xl_l: 52.
SCHREIBER, R., and De Vry, J. (1990b) Neuroanatomical correlate of the antidepressant-like effects of ipsapirone and 8-OH-DPAT in the rat forced swimming test. Psychopharmacology _lQ& 52.
SCHREIBER, R., and De Vry, J. (1991a) Involvement of pre- and postsynaptic mechanisms in the anxiolytic effects of the 5-HT, receptor ligands 8-OH-DPAT, ipsapirone and buspirone in the rat. Proceedings of the 12th Low Countries Meeting, Cologne: 32.
SCHREIBER, R., and De Vry, J. (1991b) Involvement of pre- and postsynaptic mechanisms in the antidepressive effects of the 5-HT,, receptor ligands 8-OH-DPAT and ipsapirone in the rat

5-HT,, llgmds - anxiolytIcs/antidepressants 103
forced swimming test. Proceedings of the 12th Low Countries Meeting, Cologne: 79.
SCHREIBER, R., OPITZ, K., and De VRY, J. (1991) Neuroanatomical basis of the alcohol-preference-reducing effects of the 5-I-H,,, receptor ligands 8OH-DPAT and ipsapirone. Proceedings of 5-hydroxytryptamine-CNS Receptors and Brain Function, Birmingham, 132.
SCHUURMAN, T., SPENCER D.G., and TRABER, J. (1991) Effects of ipsapirone on social behavior and conflict-induced responses in rats: a comparison to diazepam (in press).
SCHWEIZER, E., and RICKELS, K. (1991) Serotonergic anxiolytics: a review of their clinical efficacy. In: 5-I-H,, Agonists, 5-HT, Antagonists and Benzodiazepines, Their Comparative Behavioural Pharmacology, R. J. Rodgers and S.J. Cooper (Eds.), pp 365-376, John Wiley & Sons Ltd..
SOMMERMEYER, H., SCHREIBER, R., GREUEL, J., DE VRY, J., and GLASER, T. (1991) Electrophysiological and biochemical correlates of the anxiolytic activity of ipsapirone in the rat. In: Monitoring Molecules in Neuroscience, Proceedings of the 5th International Conference on in vivo methods, H. Rollema, B. Westerink and W.J. Drijfhout (Eds.), pp 404-406, Krips Repro, Meppel.
SOUBRIE, P. (1986) Reconciling the role of central serotonin neurons in human and animal behavior. Behav. & Brain Sci. 9: 319-364.
STEIN, L., WISE C.D., and BELLUZI, J.D. (1975) Effects of benzodiazepines on central serotonergic mechanisms. In: Mechanism of Action of Benzodiazepines, E. Costa and P. Greengard (Eds.), pp 29-44, Raven Press, New York.
THIEBOT, M.H., HAMON, M., and SOUBRIE, P. (1982) Attenuation of induced-anxiety in rats by chlordiazepoxide: role of raphe dorsalis benzodiazepine binding sites and serotonergic neurons. Neurosci. 2: 2287-2294.
THIEBOT, M.H. (1986) Are serotonergic neurons involved in the control of anxiety and in the anxiolytic activity of benzodiazepines ? Pharmacol. Biochem. & Behav. 3: 1471-1477.
TOLLEFSON, G.D. (1991) Anxiety and alcoholism: a serotonin link. Br. J. Psychiat. 15e(suppl. 12) 34-39.
TRABER, J., and GLASER, T. (1987) 5-HT,, receptor-related anxiolytics, TIPS 8: 432-437.
WHITE, S.M., KUCHARIK, R.F., and MOYER, J.A. (1991) Effects of serotonergic agents on isolation-induced aggression. Pharmacol. Biochem. & Behav. 3: 729-736.
WONG, D.T., and MURPHY, J.M. (1989) Serotonergic mechanisms in alcohol intake. In: Molecular Mechanisms of Alcohol, G.Y. Sun (Eds.), pp 133-146, The Humana Press, Inc., Clifton.
WRIGHT, I.K., CRESPI, F., MARSDEN, C.A., JONES, G.H., and ROBBINS, T.W. (1989) Effect of isolation-rearing on 5-I-H release in the frontal cortex. J. Neurosci. Methods 29: 283.
WRIGHT, I.K., UPTON, N., and MARSDEN, C.A. (1991a) Effect of potential anxiolytics on

104 R. Schreiber and J. De Vry
extracellular 5-HT in the ventral hippocampus of rats observed on the elevated X-maze using in vivo microdialysis. In: Monitoring Molecules in Neuroscience, Proceedings of the 5th International Conference on in vivo methods, H. Rollema, B. Westerink and W.J. Drijfhout (Eds.), pp 208-211, Krips Repro, Meppel.
WRIGHT, I.K., ISMAIL, H., UPTON, N., and MARSDEN C.A. (1991b) Effect of isolation rearing on 5-HT agonist-induced responses in the rat. Psychopharmacology m: 259-263.
WRIGHT, I.K., HEATON, M., UPTON, N., and MARSDEN, C.A. (1991~) Comparison of acute and chronic treatment of diazepam and ritanserin in the elevated plus-maze model of anxiety. In: New Concepts in Anxiety. M. Briley and SE. File (Eds.), pp 277-287, Mac Millan Press, London.
Inquiries and reprint requests should be addressed to:
Dr. Jean De Vry Institute for Neurobiology Department of Psychopharmacology Troponwerke, Berliner Str. 156 5000 Cologne 80, F.R.G.





























