Embed Size (px)
Citation preview

Rea, D.K., Basov, I.A., Scholl, D.W., and Allan, J.F. (Eds.), 1995Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 145
37. BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHY SUMMARY1
John A. Barron,2 Ivan A. Basov,3 Luc Beaufort,4 Gilles Dubuisson,5 Andrey Y. Gladenkov,3 Joseph J. Morley,6 Makoto Okada,7
Gunnar ólafsson,8 Dorothy K. Pak,9 Andrew P. Roberts,10 Valery V. Shilov,11 and Robin J. Weeks12
ABSTRACT
The magnetostratigraphic and biostratigraphic results of Ocean Drilling Program Sites 881-884 and 887 are summarizedand combined to form age vs. depth models for these sites. The recovery of continuous magnetostratigraphic records of theupper lower Miocene through Pleistocene section at Sites 884 and 887 allows the first direct calibration of diatom and radiolar-ian datum levels with magnetostratigraphy between magnetic Subchrons C5En and C3Bn (18.317-6.744 Ma) in the NorthPacific. Based on these records and temporally shorter magnetostratigraphic records at Sites 881-883, at least 25 diatom and 22radiolarian datum levels appear to be isochronous in the late early Miocene through Pleistocene of the North Pacific. The use ofcalcareous nannofossils for biostratigraphy of the post-early middle Miocene appears to be limited to the late Pliocene and Qua-ternary in sections sampled by Leg 145.
Biostratigraphy indicates that a composite section of Sites 883 and 884 represents a relatively complete record of the lowerEocene through the lower Miocene, although carbonate preservation is best and the section is less complicated by reworking atSite 883. Primary biostratigraphic control is provided by calcareous nannofossils. A large part of the upper middle Eocene andupper Eocene is missing at hiatuses at both Sites 883 and 884.
A condensed upper Paleocene section is present at Site 883, unconformably overlying uppermost Cretaceous sediments.Late Maestrichtian calcareous nannofossils are documented in Site 883 immediately above the basalt.
INTRODUCTION
Ocean Drilling Program (ODP) Leg 145 cored 25 holes at sevensites in a west to east transect of the subarctic North Pacific (Fig. 1).Three of these sites (882, 883, and 887) that were cored on top of sea-mounts, where carbonate-bearing sediments were relatively free ofterrigenous debris, offered the best possibility of high-resolution pa-leoceanographic studies. Site 884, on the east flank of the DetroitSeamount, was cored in order to obtain a comparative record of deep-sea sedimentation and to record the history of the Meiji sedimenttongue. Site 881 was selected to be a northern extension of the south-to-north paleoceanographic transect which began off Japan on DeepSea Drilling Project (DSDP) Leg 86, whereas Sites 885 and 886 werelocated in the low-biologic-productivity region of the central NorthPacific, where a good record of eolian deposition was expected (Rea,Basov, Janecek, Palmer-Julson, et al., 1993).
'Rea, D.K., Basov, LA., Scholl, D.W., and Allan, J.F. (Eds.), 1995. Proc. ODP, Sci.Results, 145: College Station, TX (Ocean Drilling Program).
2U.S. Geological Survey, MS 915, Menlo Park, CA 94025, U.S.A.3 Institute of the Lithosphere, Russian Academy of Sciences, Staromonetny per. 22,
Moscow 109180, Russia."Laboratoire de Géologie du Quatemaire, CNRS Luminy, case 907, 13288
Marseille cedex 09, France.'Laboratoire de Géologie, Ecole Normale Supérieure, 24 rue Lhomond, 75231 Paris
cedex 05, France.632 West End Ave., Westwood, NJ 07675, U.S.A.7 Ocean Research Institute, University of Tokyo 1-15-1 Minamidai, Nakano-ku,
Tokyo 164, Japan."Marine Research Institute, Hafrannsoknarstofnunin, Skulagata 4, P.O. Box 1390,
IS-121, Reykjavik, Iceland."Lamont-Doherty Earth Observatory, Palisades, NY 10964, U.S.A.'"Department of Geology, University of California, Davis, CA 95616, U.S.A."Institute of Geology and Mineral Resources of the Oceans, Maklina Ave. 1, St.
Petersburg 109121, Russia.12Department of Geological Sciences, University of Washington, Seattle, WA
98195, U.S.A.
The main purpose of this paper is to present age vs. depth modelsfor Sites 881-884 and 887 based on papers in this volume dealingwith magnetostratigraphy (Weeks et al.; Dubuisson et al.) and bio-stratigraphy (Barron and Gladenkov; Basov; Beaufort and Olafsson;Gladenkov and Barron; Morley and Nigrini; Olafsson and Beaufort;Pak and Miller; Shilov). High-resolution chronology for the Pleis-tocene and Pliocene is not treated here, and the reader is referred tothe orbital time scale developed by Tiedemann and Haug (this vol-ume) for Site 882 and oxygen isotope studies by Keigwin (this vol-ume) for Site 883, as well as radiolarian studies by Morley et al. (thisvolume) for Sites 881, 883, and 887. In general, the biostratigraphiczonation of the individual sites is not discussed because only the dia-tom zonation (Barron and Gladenkov, this volume) is widely accept-ed and widely applicable in the North Pacific. Similarly, age vs. depthmodels for Sites 885 and 886 are not treated here because they arethoroughly presented by Dickens et al. (this volume) and Snoeckx etal. (this volume).
METHODS
Time Scale/Chronological Framework
Leg 145 studies employs the Cande and Kent (1992) geomagnet-ic-polarity time scale, and the ages for biostratigraphic datums, whichwere previously calibrated to the Berggren et al. (1985a, 1985b) timescale, are updated accordingly. This recalculation involves reinterpo-lation of the age of a particular datum level within a given magneticsubchron using the Cande and Kent (1992) ages for the onset and ter-mination of that subchron. The correlation of the geologic epochs andperiods used follows that of Berggren et al. (1985a, 1985b) with theexception of the Miocene/Pliocene boundary, where the calibrationof Zijderveld et al. (1986) was used. Whenever possible, ages are as-signed to sediments based on magnetostratigraphy. Other sedimentsare dated by employing previously published calibration of biostrati-
559

J.A. BARRON ET AL.
60"JN
50c
Bering Sea
^Site884c^^
.' ;:Site882lJ
Site 881
. .
ChinookTrough
150°E 180°
Figure 1. Location of sites cored in the North Pacific.
graphic datum levels to magnetostratigraphy in the North Pacific andelsewhere (see individual chapters).
In most cases, ages listed in the tables of this chapter come direct-ly from the individual chapters and are those that the individual au-thors believe to be most reliable. However, the ages of Pak and Miller(this volume) were updated to correspond with the Cande and Kent(1992) time scale because they were referenced to the Berggren et al.(1985a) time scale. In addition, selected Paleogene planktonic fora-miniferal datum levels recorded by Basov (this volume) were as-signed ages according to Berggren et al. (1985a) and Aubry et al.(1988) and then updated to the Cande and Kent (1992) time scale.
RESULTS
Site 881
An apparently continuous sequence of upper Miocene throughQuaternary sediments was recovered at Site 881 (47°6.1'N,161°29.5'E; 5531 m water depth). The 364-m-thick section consistsof diatom ooze overlain by clayey diatom ooze, the latter containingnumerous ash layers. Diatoms and radiolarians generally are com-mon to abundant and moderately well to well preserved throughoutthe section recovered at Site 881, whereas calcareous nannofossilsand foraminifers occur at only a few intervals and at very low abun-dances in sediments from the four holes drilled at this site.
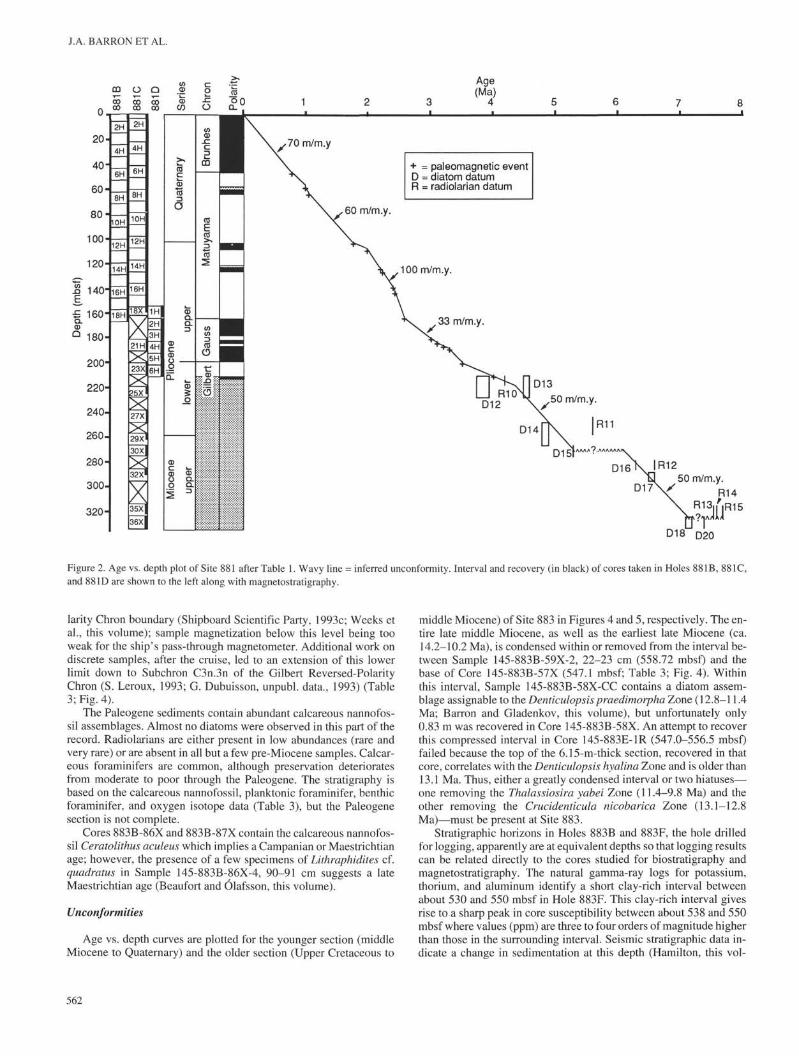
The magnetostratigraphy at Site 881 extends down to the upper-most part of the Gilbert Reversed-Polarity Chron (Shipboard Scien-tific Party, 1993a; Weeks et al., this volume). Table 1 gives theestimated ages of paleomagnetic polarity boundaries and microfossildatum levels recorded in Holes 881B, 881C, and 881D, the intervalover which they occur, and the depth of this interval in meters belowsea floor (mbsf).
An age vs. depth curve for Site 881 is shown in Figure 2 using pa-leomagnetic polarity boundaries (above 210 mbsf) and diatom andradiolarian datum levels (Table 1) that have previously been calibrat-ed to magnetostratigraphy. At about 165 mbsf the sediment accumu-lation rate decreases abruptly down section from 100 m/m.y. to 33 m/
m.y., coinciding with an abrupt change in lithology from clayey dia-tom ooze rich in ashes above to diatom ooze below (Shipboard Sci-entific Party, 1993a). The change in physical and magnetic propertiesat this level is likewise abrupt (Shipboard Scientific Party, 1993a),and seismic stratigraphy recognizes a region-wide reflector at this ho-rizon (Hamilton, this volume).
Unconformities
An unconformity is suggested by the age vs. depth curve (Fig. 2)at about 270 mbsf, where the latest Miocene interval, correspondingto about 6.1 to 5.3 Ma, may be missing. A widespread reflector is ev-ident at this depth from seismic reflection data (Hamilton, this vol-ume); however, no lithologic change or major change in the naturalgamma ray or other logs is apparent at this depth (Shipboard Scien-tific Party, 1993a).
A second possible unconformity is inferred at about 325 mbsf inthe unrecovered interval between Cores 881C-35X and 881C-36X,based on the coincidence of diatom and radiolarian datum levels thatrange in age from ca. 7.7 to 7.1 Ma (Table 1; Fig. 2). Abrupt down-hole increase in the weight-percent of potassium, thorium, and urani-um are indicated by the natural gamma-ray log, and the Gamma-rayattenuation porosity evaluator (GRAPE) recorded an abrupt increasein bulk density at this depth (Shipboard Scientific Party, 1993a).
Site 882
An apparently continuous sequence of upper Miocene throughQuaternary sediments was recovered at Site 882 on the top of the De-troit Seamount at 50°21.8'N; 167°36.0/E (3255 m water depth). Thetwo holes that were drilled penetrated a 398-m-thick section consist-ing of diatom ooze with intervals of clayey diatom ooze and minorcarbonate materials.
Moderately well-preserved calcareous nannofossils occur inabundances ranging from rare to abundant but with little diversityamong species, down to approximately 227 mbsf, where they becomerare. Moderately well-preserved calcareous foraminifers are present
560

BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHIC SUMMARY
Table 1. Age, interval, and depth of magnetostratigraphic polarity boundaries and biostratigraphic datum levels in Holes 881B, 881C, and 881D.
RlDlR2R3R4
R5
D2R6
1)31)4R7D5
D6RS
D7
1)81)9R9
D10
D111)12RIOD131)141)15RMD16R121)17D181)191)20R13R14R15
LOLOLOLOLOBLOTLOLOBLOLCOLOFOTBLOF 0TBLOTBT
TLCOFOFOBTLOBTBTFOFOLOLOFOFOFOLCOFOFOFOFOLO
LOFO
Datum
Lychnocanoma niponica sakaiiSimonseniella curvirostrisStyiacontarium acquiloniumStylatractus universusAnthocyrtella (?) callopsmaC l n . l nLamprocyrtis neoheteroporosClr.lnRhizosolenia matuyamaiEucyrtidium matuyamaiC 1 r. 1 rRhizosolenia matuyamaiActinocyclus oculatusSphaeropyle robustaSimonseniella curvirostrisC2n.lnC2n.lnNeodenticula koizumiiEucyrtidium matuyamaiC2n.2nC2n.2nThalassiosira convexa"X" anomaly"X" anomalyC2An.ln
C2An.lnNeodenticula kamtschaticaNeodenticula seminaeCycladophora davisiana davisianaC2An.lnC2An.2nThalassiosira marujamicaC2An.2nC2An.3nC2An.3nC3n.lnNeodenticula koizumiiActinocyclus oculatusDictyophimus bullatusThalassiosira jacksonii (plicate)Thalassiosira latimarginataThalassiosira oestrupiiDictyophimus bullatusRouxia californicaLamprocyrtis heteroporosThalassiosira jacksoniiNeodenticula kamtschaticaNitzschia reinholdiiThalassionema schraderiS. delmontensis > S. peregrinaPrunopyle hayesiStyiacontarium acquilonium
Age(Ma)
0.050.300.400.550.620.780.85
0.9840.91-1.04
1.001.049
0.98-1.12LO
1.5-1.71.58
1.7571.9832.02.0
2.1972.229
2.42.42
2.4412.6
2.62.63-2.7
2.72.90
3.0543.127
3.1-3.23.221.3.3253.5534.033
3.51-3.853.65-3.85
4.24.5^.64.8^1.9
5.35.606.36.6
6.5-6.67.1-7.27.2-7.3
7.47.557.67.7
Core, section,interval (cm)
145-881B-
Above 1H-CC3H-CC/4H-CC2H-CC/3H-CC2H-CC/3H-CC
6H-3, 07H-CC/8H-CC7H-2, 55
7H-CC/8H-CC7H-6, 120
7H-CC/8H-CC
10H-CC/11H-CC12H-3, 1012H-6, 9511H-CC/12H-CC12H-CC/13H-CC13H-6, 3014H-2, 50?15H-CC/16H-CC?16H-1.016H-4, 7018H-5, 130145-881D-1H-6, 5/2H-l,01H-CC/2H-CC1H-CC/2H-CC2H-CC/3H-CC3H-3, 1203H-5, 1503H-5, 125/3H-CC4H-1, 704H-2, 1205H-4, 40/5H-6, 1006H-6, 80Below 6H-CCBelow 6H-CCBelow 6H-CC
Depth(mbsf)
/5.524.5/34.015.0/24.515.0/24.5
46.5062.5/72.0
55.05
62.5/72.061.70
62.5/72.0
91.0/100.5103.60108.95
100.5/110.0110.0/119.5117.8/119.8
121.5?138.5/148.0
138.50143.70164.80
163.0/164.5164.75/174.0164.75/174.0174.0/183.5
178.2181.5
181.25/183.5184.2186.2
197.9/201.5210.8
Below 212.0Below 212.0Below 212.0
Core, section,interval (cm)
145-881C-
1H-L21-22/1H-CC3H-CC/4H-CC, 23H-CC/4H-CC, 23H-CC/4H-CC, 24H-CC/5H-CC6H-5, 305H-CC/6H-CC7H-5, 307H-CC/8H-CC6H-CC/7H-CC8H-2, 20 or 8H-4, 1408H-CC/9H-CC7H-CC/8H-CC10H-CC/11H-CC11H-CC/12H-CC12H-5, 20?13H-2, 10?11H-CC/12H-CC13H-CC/14H-CC14H-4, 7514H-7, 70
16H-2, 12016H-6, 10
18X-CC/21H-L2518X-CC/21H-L2518X-CC/21H-CC
18X-CC/21H-L25
23X-CC/25X-CC23X-CC/25X-1,20-2123X-CC/25X-CC27X-CC/29X-CC29X-CC/30X-CC27X-5, 20-21/29X-1, 20-2130X-CC/32X-L4630X-7, 22-23/32X-1,20-2132X-3, 46/32X-CC35X-CC/36X-CC32X-CC/35X-CC35X-CC/36X-CC35X-L22-23/36X-1,29-3035X-L22-23/36X-1,29-3035X-L22-23/36X-1,29-30
Depth(mbsf)
0.2/3.82.8/32.32.8/32.32.8/32.3
32.3/41.848.1
41.8/51.357.6
60.8/70.351.3/60.8
62.5 or 66.770.3/79.860.8/70.389.3/98.8
98.8/108.3105?
109.9?98.9/108.3117.8/127.3
123.05127.50
139.50144.40
162.3/182.0162.3/182.0162.3/191.2
162.3/182.0
209.7/228.7209.7/219.2209.7/228.7248.0/267.2267.2/276.9244.5/257.8
276.9/286.96276.42/286.7
288.46/296.20325.1/334.8
296.2/325.10325.1/334.8
315.62/325.39315.62/325.39315.62/325.39
Notes: R = radiolarian, D = diatom, B = base, T = top, FO = first occurrence, LO = last occurrence, LCO = last common occurrence, and > = transition.
in low abundance at several intervals, whereas diatoms are commonto abundant, and are well preserved throughout the entire sequence.Radiolarians occur throughout the section where they are common toabundant in most Quaternary sediments, and rare in Miocene andPliocene sequences. Only a few samples contained minor amounts ofreworked fauna.
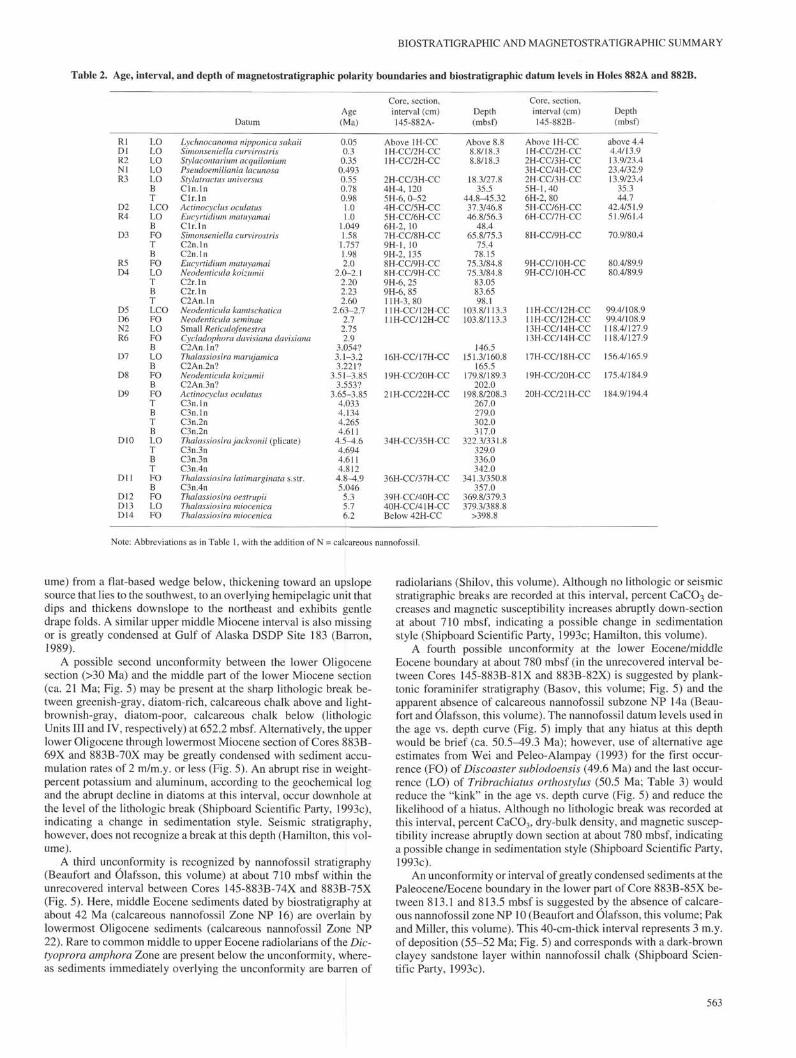
The magnetostratigraphy at Site 882 (Shipboard Scientific Party,1993b; Weeks et al., this volume) extends down through the GilbertReversed-Polarity Chron, although the separate polarity boundariesof the Gauss Normal-Polarity Chron and the upper normal event ofthe Gilbert Reversed-Polarity Chron are only tentatively identified.The estimated ages of paleomagnetic polarity boundaries and diatom,radiolarian, and calcareous nannofossil datum levels recorded in sed-iments recovered from Holes 882A and 882B, the interval over whichthey occur, and the depth of this interval in mbsf are given in Table 2.
An age vs. depth curve was plotted in Figure 3 based primarily onmagnetostratigraphic polarity boundaries (Table 2). However, thereader is referred to Tiedemann and Haug (this volume) where a de-tailed astronomically-tuned time scale is presented. The presence ofthe diatom Thalassiosira miocenica in Sample 145-882A-42H-CC,
the stratigraphically lowest sample taken at Site 882, implies that thebase of Hole 882A is younger than 6.3 Ma (latest Miocene), consis-tent with the magnetic polarity interpretation shown in Figure 3.
Site 883
Site 883 was drilled on the top of Detroit Seamount (5FI 1.9'N,167°46. l'E; 2396 m water depth), 49 nautical miles (nmi) to the northof Site 882. Drilling at Site 883 penetrated an 820-m-thick sequenceof Cenozoic sediments overlying a thin sequence of Cretaceous sed-iments before reaching basement at about 840 mbsf. The 655-m-thicksection of Miocene to Pleistocene sediments is rich in well-preservedsiliceous microfossils, diatoms being the most abundant. The calcar-eous microfossil abundance is highly variable. The diversity of thecalcareous nannofossils is low, and few biostratigraphic datums wereidentified in the Pleistocene. The Miocene to Pleistocene biostratig-raphy is essentially based on siliceous microfossils, which are muchmore diverse than the calcareous microfossils (Table 3).
The magnetostratigraphy, obtained on board ship at Site 883, ex-tends down to the Gauss Normal-Polarity Matuyama Reversed-Po-
561
J.A. BARRON ET AL.
O D O Q
0 .
20-
40"
60-
80-
100-
120-
(mb
sf)
£ 160-Φ
Q 180-
200-
220-
240-
260-
280-
300-
320-
0000
?H
4H
6H
8H
tflH
14H
1RH
18H
0000
2H
4H
6H
8H
10H
12H
U H
16H
18X
X21H
X23X
X25X
X27X
x29X
30 X
32X
X35X
36X
I1H
2H
3H
4H
5H
6H
Se
na
ryΦfδ3σ
upp
er
Φ
lioce
P
we
r
Φ
α> Φü Q.J =
X- O Q
is
(0
o
Age(Ma)
3 4
+ paleomagnetic eventD = diatom datumR = radiolarian datum
D18
Figure 2. Age vs. depth plot of Site 881 after Table 1. Wavy line = inferred unconformity. Interval and recovery (in black) of cores taken in Holes 88IB, 88IC,and 88ID are shown to the left along with magnetostratigraphy.
larity Chron boundary (Shipboard Scientific Party, 1993c; Weeks etal., this volume); sample magnetization below this level being tooweak for the ship's pass-through magnetometer. Additional work ondiscrete samples, after the cruise, led to an extension of this lowerlimit down to Subchron C3n.3n of the Gilbert Reversed-PolarityChron (S. Leroux, 1993; G. Dubuisson, unpubl. data., 1993) (Table3; Fig. 4).
The Paleogene sediments contain abundant calcareous nannofos-sil assemblages. Almost no diatoms were observed in this part of therecord. Radiolarians are either present in low abundances (rare andvery rare) or are absent in all but a few pre-Miocene samples. Calcar-eous foraminifers are common, although preservation deterioratesfrom moderate to poor through the Paleogene. The stratigraphy isbased on the calcareous nannofossil, planktonic foraminifer, benthicforaminifer, and oxygen isotope data (Table 3), but the Paleogenesection is not complete.
Cores 883B-86X and 883B-87X contain the calcareous nannofos-sil Ceratolithus aculeus which implies a Campanian or Maestrichtianage; however, the presence of a few specimens of Lithraphidites cf.quadratus in Sample 145-883B-86X-4, 90-91 cm suggests a lateMaestrichtian age (Beaufort and ólafsson, this volume).
unconformities
Age vs. depth curves are plotted for the younger section (middleMiocene to Quaternary) and the older section (Upper Cretaceous to
middle Miocene) of Site 883 in Figures 4 and 5, respectively. The en-tire late middle Miocene, as well as the earliest late Miocene (ca.14.2-10.2 Ma), is condensed within or removed from the interval be-tween Sample 145-883B-59X-2, 22-23 cm (558.72 mbsf) and thebase of Core 145-883B-57X (547.1 mbsf; Table 3; Fig. 4). Withinthis interval, Sample 145-883B-58X-CC contains a diatom assem-blage assignable to the Denticulopsispraedimorpha Zone (12.8-11.4Ma; Barron and Gladenkov, this volume), but unfortunately only0.83 m was recovered in Core 145-883B-58X. An attempt to recoverthis compressed interval in Core 145-883E-1R (547.0-556.5 mbsf)failed because the top of the 6.15-m-thick section, recovered in thatcore, correlates with the Denticulopsis hyalina Zone and is older than13.1 Ma. Thus, either a greatly condensed interval or two hiatuses—one removing the Thalassiosira yabei Zone (11.4-9.8 Ma) and theother removing the Crucidenticula nicobarica Zone (13.1-12.8Ma)—must be present at Site 883.
Stratigraphic horizons in Holes 883B and 883F, the hole drilledfor logging, apparently are at equivalent depths so that logging resultscan be related directly to the cores studied for biostratigraphy andmagnetostratigraphy. The natural gamma-ray logs for potassium,thorium, and aluminum identify a short clay-rich interval betweenabout 530 and 550 mbsf in Hole 883F. This clay-rich interval givesrise to a sharp peak in core susceptibility between about 538 and 550mbsf where values (ppm) are three to four orders of magnitude higherthan those in the surrounding interval. Seismic stratigraphic data in-dicate a change in sedimentation at this depth (Hamilton, this vol-
BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHIC SUMMARY
Table 2. Age, interval, and depth of magnetostratigraphic polarity boundaries and biostratigraphic datum levels in Holes 882A and 882B.
RlDlR2NIR3
D2R4
D3
R5D4
D5D6N2R6
D7
D8
D9
D10
D11
D12D13D14
Note:
LOLOLOLOLOBTLCOLOBFOTBF0LOTBTLCOF0LOFOBLOBFOBFOTBTBLOTBTF0BF0LOF0
Datum
Lychnocanoma nipponica sakaiiSimonseniella curvirostrisStylacontarium acquiloniumPseudoemiliania lacunosaStylatractus universusCln.lnClr.lnActinocyclus oculatusEucyrtidium matuyamaiClr.lnSimonseniella curvirostrisC2n.lnC2n.lnEucyrtidium matuyamaiNeodenticula koizumiiC2r.lnC2r.lnC2An.lnNeodenticula kamtschaticaNeodenticula seminaeSmall ReticulofenestraCycladophora davisiana davisianaC2An.ln?Thalassiosira marujamicaC2An.2n?Neodenticula koizumiiC2An.3n?Actinocyclus oculatusC3n.lnC3n.lnC3n.2nC3n.2nThalassiosira jacksonii (plicate)C3n.3nC3n.3nC3n.4nThalassiosira latimarginata s.str.C3n.4nThalassiosira oestrupiiThalassiosira miocenicaThalassiosira miocenica
Abbreviations as in Table 1, with the addition of N
Age(Ma)
0.050.3
0.350.4930.550.780.981.01.0
1.0491.58
1.7571.982.0
2.0-2.12.202.232.60
2.63-2.72.7
2.752.9
3.054?3.1-3.23.221?
3.51-3.853.553?
3.65-3.854.0334.1344.2654.611
4.5^1.64.6944.6114.8124.8-4.95.046
5.35.76.2
= cal
Core, section,interval (cm)145-882A-
Above 1H-CC1H-CC/2H-CC1H-CC/2H-CC
2H-CC/3H-CC4H-4, 1205H-6, 0-524H-CC/5H-CC5H-CC/6H-CC6H-2, 107H-CC/8H-CC9H-1, 109H-2, 1358H-CC/9H-CC8H-CC/9H-CC9H-6, 259H-6, 8511H-3, 8011H-CC/12H-CC11H-CC/12H-CC
16H-CC/17H-CC
19H-CC/20H-CC
21H-CC/22H-CC
34H-CC/35H-CC
36H-CC/37H-CC
39H-CC/40H-CC40H-CC/41H-CCBelow 42H-CC
careous nannofossil.
Depth(mbsf)
Above 8.88.8/18.38.8/18.3
18.3/27.835.5
44.8^5.3237.3/46.846.8/56.3
48.465.8/75.3
75.478.15
75.3/84.875.3/84.8
83.0583.6598.1
103.8/113.3103.8/113.3
146.5151.3/160.8
165.5179.8/189.3
202.0198.8/208.3
267.0279.0302.0317.0
322.3/331.8329.0336.0342.0
341.3/350.8357.0
369.8/379.3379.3/388.8
>398.8
Core, section,interval (cm)145-882B-
Above 1H-CC1H-CC/2H-CC2H-CC/3H-CC3H-CC/4H-CC2H-CC/3H-CC5H-l,406H-2, 805H-CC/6H-CC6H-CC/7H-CC
8H-CC/9H-CC
9H-CC/10H-CC9H-CC/10H-CC
11H-CC/12H-CC11H-CC/12H-CC13H-CC/14H-CC13H-CC/14H-CC
17H-CC/18H-CC
19H-CC/20H-CC
20H-CC/21H-CC
Depth(mbsf)
above 4.44.4/13.913.9/23.423.4/32.913.9/23.4
35.344.7
42.4/51.951.9/61.4
70.9/80.4
80.4/89.980.4/89.9
99.4/108.999.4/108.9118.4/127.9118.4/127.9
156.4/165.9
175.4/184.9
184.9/194.4
ume) from a flat-based wedge below, thickening toward an upslopesource that lies to the southwest, to an overlying hemipelagic unit thatdips and thickens downslope to the northeast and exhibits gentledrape folds. A similar upper middle Miocene interval is also missingor is greatly condensed at Gulf of Alaska DSDP Site 183 (Barron,1989).
A possible second unconformity between the lower Oligocenesection (>30 Ma) and the middle part of the lower Miocene section(ca. 21 Ma; Fig. 5) may be present at the sharp lithologic break be-tween greenish-gray, diatom-rich, calcareous chalk above and light-brownish-gray, diatom-poor, calcareous chalk below (lithologicUnits III and IV, respectively) at 652.2 mbsf. Alternatively, the upperlower Oligocene through lowermost Miocene section of Cores 883B-69X and 883B-70X may be greatly condensed with sediment accu-mulation rates of 2 m/m.y. or less (Fig. 5). An abrupt rise in weight-percent potassium and aluminum, according to the geochemical logand the abrupt decline in diatoms at this interval, occur downhole atthe level of the lithologic break (Shipboard Scientific Party, 1993c),indicating a change in sedimentation style. Seismic stratigraphy,however, does not recognize a break at this depth (Hamilton, this vol-ume).
A third unconformity is recognized by nannofossil stratigraphy(Beaufort and Olafsson, this volume) at about 710 mbsf within theunrecovered interval between Cores 145-883B-74X and 883B-75X(Fig. 5). Here, middle Eocene sediments dated by biostratigraphy atabout 42 Ma (calcareous nannofossil Zone NP 16) are overlain bylowermost Oligocene sediments (calcareous nannofossil Zone NP22). Rare to common middle to upper Eocene radiolarians of the Dic-tyoprora amphora Zone are present below the unconformity, where-as sediments immediately overlying the unconformity are barren of
radiolarians (Shilov, this volume). Although no lithologic or seismicstratigraphic breaks are recorded at this interval, percent CaCO3 de-creases and magnetic susceptibility increases abruptly down-sectionat about 710 mbsf, indicating a possible change in sedimentationstyle (Shipboard Scientific Party, 1993c; Hamilton, this volume).
A fourth possible unconformity at the lower Eocene/middleEocene boundary at about 780 mbsf (in the unrecovered interval be-tween Cores 145-883B-81X and 883B-82X) is suggested by plank-tonic foraminifer stratigraphy (Basov, this volume; Fig. 5) and theapparent absence of calcareous nannofossil subzone NP 14a (Beau-fort and Olafsson, this volume). The nannofossil datum levels used inthe age vs. depth curve (Fig. 5) imply that any hiatus at this depthwould be brief (ca. 50.5-49.3 Ma); however, use of alternative ageestimates from Wei and Peleo-Alampay (1993) for the first occur-rence (FO) of Discoaster sublodoensis (49.6 Ma) and the last occur-rence (LO) of Tribrachiatus orthostylus (50.5 Ma; Table 3) wouldreduce the "kink" in the age vs. depth curve (Fig. 5) and reduce thelikelihood of a hiatus. Although no lithologic break was recorded atthis interval, percent CaCO3, dry-bulk density, and magnetic suscep-tibility increase abruptly down section at about 780 mbsf, indicatinga possible change in sedimentation style (Shipboard Scientific Party,1993c).
An unconformity or interval of greatly condensed sediments at thePaleocene/Eocene boundary in the lower part of Core 883B-85X be-tween 813.1 and 813.5 mbsf is suggested by the absence of calcare-ous nannofossil zone NP 10 (Beaufort and Olafsson, this volume; Pakand Miller, this volume). This 40-cm-thick interval represents 3 m.y.of deposition (55-52 Ma; Fig. 5) and corresponds with a dark-brownclayey sandstone layer within nannofossil chalk (Shipboard Scien-tific Party, 1993c).

J.A. BARRON ET AL.
0 .
20-
4 0 -
60-
8 0 -
100-
120-
140"
160"
=fc 180-
E^ 200-Q.Φ21 220-
240-
260-
280-
300-
320-
340-
Jbü
380-
ΛDÙ-
1882B
5H
10H
15H
20H
25H
30H
35H
40H
1882C
5H
1OH
15H
20H
25H
29H
y S
erie
ser
nar
αS3
up
pe
r
oc
en
e
Φ
O
Mio
ce
ne
up
pe
r
oszü
m
COP
k3CO
w
COC5
Ë(1
LPola
rio
•—
-
[
Age(Ma)
3 v ' 4
,46 m/m.y+ = paleomagnetic eventD = diatom datum
,35 m/m.y.
,110 m/m.y.
,140 m/m.y.
^ 6 6 m/m.y.
D11
^,48 m/m.y.
D 1 2 \ j D 1 3D14T
Figure 3. Age vs. depth plot of Site 882 after Table 2.
A major unconformity between Maestrichtian and uppermost Pa-leocene sediments is present in the unrecovered interval between thebase of Core 145-883B-85X and the top of Core 145-883B-84X(814-818.5 mbsf; Fig. 5). This break coincides with the contact be-tween lithologic Unit V, yellow green and yellow brown to reddish-brown ashes, and lithologic Unit IV, nannofossil ash and chalk (Ship-board Scientific Party, 1993c). Abrupt downhole increases in the nat-ural gamma-ray logs for potassium and uranium are recorded at thisinterval (Shipboard Scientific Party, 1993c). Seismic stratigraphicdata indicate that lithologic Unit V fills a local depression that is 3.4km across on the top of the basalt of Detroit Seamount (Hamilton,this volume).
Site 884
Drilling at Site 884 on the east flank of Detroit Seamount(51°27'N, 168°20.2'E; 3836 m water depth) recovered an 850-m-thick sequence of upper Paleocene(?) through Pleistocene sedimentsin five holes (Shipboard Scientific Party, 1993d). The Neogene andPleistocene are represented by a 640-m-thick section of diatomite,
claystone, and clayey diatom ooze. Calcareous microfossil abun-dance is highly variable throughout this interval. A limited nannofos-sil zonation was possible for the Quaternary part of the section.
In general, diatoms are abundant to common and well preservedto moderately well preserved throughout the upper Oligocenethrough Quaternary section cored above 658 mbsf at Site 884 (seealso Gladenkov and Barron, this volume). Radiolarians are abundantonly in the middle Miocene part of the section and in three isolatedsamples from the upper Miocene, upper Pliocene, and lower Pleis-tocene parts of the section.
An added bonus of drilling at Site 884 was the retrieval of an ex-cellent paleomagnetic stratigraphy in the upper 560 m of Hole 884B,which extends from the Pleistocene back to at least the early middleMiocene or more (Table 4; Fig. 6). In addition, Dubuisson et al. (thisvolume) used down-hole magnetic logs to recognize magnetic polar-ity boundaries in the upper 440 m of Hole 884E (Table 6).
Recognition of magnetic-polarity subchrons Cln through C5n (0-10.8 Ma) is straightforward, and good agreement exists between theages suggested by diatoms and radiolarians (Barron and Gladenkov,this volume; Morley and Nigrini, this volume) and the paleomagnetic

BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHIC SUMMARY
Table 3. Age, interval, and depth of magnetostratigraphic polarity boundaries and biostratigraphic datum levels in Hole 883B.
R lNIDlR2R3N2
1)2
N31)3N4
1)4
R4
D5D6R5
D7
N5D8
1)9
1)10D11D12D131)14D15D16R6D17D181)19R7R8D20R9
D211)22D23RIOR H
D24D25N6D26D27R121)281)29N7D30N8D31R13D32N9D33M ON i lN12ISN13N14N15N16N17PIP2NI 8N19
LOF0LOLOLOLOBTLCOLOBLOFOFOTLOBFOTLCOFOFOBTLOBTLOF 0BFOTBTBTLOF 0F 0LOFOF 0F 0FOFOFOLOFO
LCOLO
LOF 0LCOLOFO
F 0FCOLOFOFOF 0FOFOFOF 0OF 0FO0LOF 0LOLOLO
LOLOF 0LOLOLOF 0LOFO
Datum
Lychnocanoma nipponica sakaiiEmiliania huxleyiSimonseniella curvirostrisStylacontarium acquiloniumStylatractus universusPseudoemiliania lacunosaCln.lnClr.lnActinocydus oculatusEucyrtidium matuyamaiClrlnCalcidiscus macintyreiSimonseniella curvirostrisGephyrocapsa oceanicaC2n.lnNeodenticula koizumiiC2n.lnEucyrtidium matuyamaiC2An.lnNeodenticula kamtschaticaNeodenticula seminaeCycladophora davisiana davisianaC2An.ln *C2An.2n *Thalassiosira marujamicaC2An.2n *C2An.3n *Reticulofenestra pseudoumbilicaNeodenticula koizumiiC2An.3n *Actinocydus oculatusC3n.ln*C3n.ln *C3n.2n? *C3n.2n? *C3n.3n *Thalassiosira jacksonii (plicate)Thalassiosira latimarginataThalassiosira oestrupiiThalassiosira miocenicaThalassiosira praeoestrupiiThalassiosira miocenicaThalassiosira jacksoniiLamprocyrtis heteroporosNeodenticula kamtschaticaNitzschia reinholdiiThalassionema schraderiStylacontarium acquiloniumS. delmontensis > S. peregrinaDenticulopsis hustedtiiLychnocanoma nipponica
magnacornutaDenticulopsis dimorphaDenticulopsis dimorphaDenticulopsis praedimorphaCyrtocapsella cornutaLychnocanoma nipponica
magnacornutaDenticulopsis praedimorphaDenticulopsis hustedtiiSphenolithus heteromorphusDenticulopsis hustedtiiDenticulopsis hyalinaEucyrtidium inflatumDenticulopsis lautaDenticulopsis praelautaSphenolithus heteromorphusCrucidenticula sawamuraeSphenolithus belemnosThalassiosira fragaCyrtocapsella tetraperaLisitzinia ornataDictyococcites bisectusCavitatus miocenicaReticulofenestra umbilicaIsthmolithus recurvusEricsonia formosaOxygen isotope increaseDiscoaster saipanensisDiscoaster barbadiensisIsthmolithus recurvusChiasmolithus grandisChiasmolithus solitusAcarinina bullbrookiGlobigerinatheka indexDiscoaster distinctusDiscoaster bifax
Aizc(Ma)
0.050.294
0.30.4
0.550.4930.780.984
1.0LO
1.0491.5381.581.6741.757
1.9-2.01.9832.0
2.602.63-2.7
2.72.9
3.0543.127
3.1-3.23.2213.3253.713
3.51-3.853.553
3.65-3.854.0334.1344.2654.4324.611
4.5-4.64.9-5.0
5.35.7
5.956.2
6.5-6.66.6
7.1-7.27.2-7.3
7.47.77.558.48.8
9.09.811.411.712.5
12.813.1
13.5114.214.')15.315.916.3
18.1818.4
18.54-19.1920.122.6
24.3-27.923.8130.6
31.72(32.4)32.5
32.7 (32.9)34.3
34.98 (34.2)34.98 (34.2)35.08 (36.2)
(37.1)39.7240.542.9
(43.7)
Core, section,interval (cm)
145-883B-
Above 1H-CC1H-5, 130-131/1H-CC1H-CC/2H-CC1H-CC/2H-CC2H-CC/3H-CC3H-5, 46^7/3H-6, 46-474H-5, 505H-5, 1204H-CC/5H-CC5H-CC/6H-CC6H-1, 1007H-7, 46-47/7H-CC6H-CC/7H-CC7H-CC/8H-1,46^78H-2, 706H-CC/7H-CC8H-4, 607H-CC/8H-CC9H-7to 10H-19H-CC/10H-CC9H-CC/10H-CC12H-CC/13H-CC
15H-2, 91/15H-2, 135
16H-2, 46-47/16H-3, 4 6 ^ 718H-CC/19H-CC
20H-CC/21H-CC
3OH-CC/31H-CC31H-CC/32X-CC33X-CC/34X-CC34X-CC/35X-CC34X-CC/35X-CC44X-CC/45X-CC46X-CC/47X-CC33X-CC/34X-CC?49X-CC/50X-CC50X-CC/51X-CC45X-CC/46X-CC46X-CC/47X-CC47X-CC/48H-CC53X-CC/54X-2, 10454X-CC/55X-CC
57X-4, 104/57X-CC57X-CC/58X-CC58X-CC/59X-CC58X-CC/59X-CC57X-CC/58X-CC
58X-CC/59X-2, 2258X-CC/59X-2, 2262X-6, 46^7/62X-7, 46-4758X-CC/59X-2, 2260X-CC/61X-2, 10461X-CC/62X-CC62X-CC/63X-2, 10463X-2, 104/63X-4, 10465X-2, 46-47/65X-3, 46-4765X-CC/66X-2, 10466X-2, 46-4768-4. 22/68X-CC, 367X-CC/68X-CC68X-CC69X-2, 46-47/69X-3, 46-4769X-CC/70X-CC71X-1, 46-47/71X-2, 46-4771X-7, 46-47/71X-CC74X-CC/75X-1, 45-4674X-2, 95-100/75X-1, 30-3574X-CC/75X-1,45-4674X-CC/75X-1,45-4674X-CC/75X-1, 46-4774X-CC/75X-1,46-4774X-CC/75X-1,46-4774X-2, 120-122/75X-1. 120-12275X-1, 120-122/75X-2, 4 5 ^ 775X-5, 46-47/76X-1, 46^1776X-4, 46-47/76X-5, 46-47
Depth(mbsf)
above 7.97.30/7.97.9/17.47.9/17.417.4/26.9
23.86/25.3633.3043.50
36.4/45.945.9/55.4
47.0064.86/64.955.4/64.9
64.9/65.3667.1
55.4/64.970
64.9/74.483.7-84.383.9/93.483.9/93.4
112.4/121.9125.15133.9
133.81/134.25141.45154.9
142.86/144.36159.9/168.4170.4(180)178.9/188.4
231.65(219)241.45(226)
266.2277.95287.45
283.4/292.9292.9/302.5312.2/321.9321.9/331.5321.9/331.5419.2/429.0439.3/448.7312.2/321.9?468.1/477.9477.9/487.9429.0/438.9439.3/458.2448.7/458.2507.6/510.14517.4/527.2
542.64/547.1547.1/557.0557.0/567.0557.0/567.0547.1/557.0
557.0/558.72557.0/558.72594.96/596.46557.0/558.72577.0/579.54587.0/597.0597.0/599.54599.54/602.54618.66/620.06626.3/628.84
628.26649.87/652.13645.1/654.8
654.8656.76/658.26
664.4/674.1674.56/676.06683.56/683.7
705.99/713.06705.45/712.80705.99/713.06705.99/713.06705.99/713.06705.99/713.06705.99/713.06
705.7/713.8713.8/714.55719.06/721.56725.06/726.56
565

J.A. BARRON ET AL.
Table 3 (continued).
N20P3N21N22N23N24X25N26P4N27P5X28N29BFN30N31N32N33N34
OF0LOLOFOLOFOLOFOFOF0F0LO
FOFOF0FOLO
Datum
Chiasmolithus gigasAcarinina bullbrookiDiscoaster sublodoensisNannotetrina fulgensNannotetrina cristataDiscoaster lodoensisDiscoaster sublodoensisTribrachiatus orthostylusAcarinina pentacamerataDiscoaster lodoensisMorozovella aragonensisTribrachiatus orthostylusFasciculithus tympaniformisBenthic extinction eventDiscoaster multiradiatusDiscoaster mohleriHeliolithus kleinpelliiFasciculithus tympaniformisMicula spp.
Age(Ma)
(44.6-46.3)45.2
47.23(47.3)47.847.9
49.29 (49.6)50.78 (50.5)
50.852.04 (52.5)
52.353.3254.0855.3
(56.5)57.79 (57.9)58.57 (59.5)60.63 (60.0)65.0(66.1)
Core, section,interval (cm)
145-883B-
76X-6, 46-4778X-2, 120-122/78X-5, 123-12578X-CC/79X-1,46-4779-CC/80-CC80X-CC/81X-CC80X-CC/81X-CC81X-CC/82X-1,46-4782X-3, 46-47/82X-4, 4 6 ^ 783X-3, 120-122/83X4, 122-12485X-2, 140-141/85X-3, 20-2183X-CC/84X-1,60-6285X-3, 120-121/85X-3, 140-14185X-3, 140-141/85X4,4-585X-3, 129-134/85X4, 10-1585X4,7-8/85X4, 10-1185X-CC/86X-1, 11085X-CC/86X-1, 11085X-CC/86X-1, 11085X-CC/86X-1,110
Depth(mbsf)
728.06740.4/746.93749.5/749.96754.5/769.2769.2/779.2769.2/779.2
779.2/779.66782.66/784.16793.4/794.92
811.80/812.10795.27/799.8
813.10/813.30813.30/813.44813.19/813.50813.47/813.50
814.1/819.6814.1/819.6814.1/819.6814.1/819.6
Notes: Abbreviations as in Table 1, with the addition of N = calcareous nannofossil, P = planktonic foraminifer, IS = oxygen isotope, BF = benthic foraminifer, O = occurrence, andFCO = first common occurrence. Asterisk (*) = after Leroux (1993). Ages in parentheses after Wei and Peleo-Alampay (1993).
Age(Ma) 10
+ = paleomagnetic eventD = diatom datumR = radiolarian datumN = calcareous nannofossil datum
100
200-
300
400
500-
600
.95 m / m.y.JR5
12 14
|R6?
D23,D221 ^ΛΛΛΛΛΛΛΛΛΛΛΛΛΛΛΛΛ•ΛΛΛΛΛΛΛΛ
R11k/
R10D24 D25 D26
28 m / m.y.
D28
D27R121
Figure 4. Age vs. depth plot of the middle Miocene through Quaternary section of Site 883 after Table 3.
interpretation of Weeks et al. (this volume; Table 4). Diatom and ra-diolarian biostratigraphy, however, suggest that sediment accumula-tion rates decrease markedly beginning down-section in the middleMiocene part of the section (ca. 500 mbsf; Fig. 6), and the magneto-stratigraphic interpretation favored by the biostratigraphy for themiddle Miocene sequence of Hole 884B is "Interpretation 2" of the
Shipboard Scientific Party (1993d) and Weeks et al. (this volume). Afew minor modifications, however, are suggested by diatom and ra-diolarian stratigraphy to "Interpretation 2" (Table 4; Fig. 6). The nor-mal-polarity interval recognized between 535.0 and 542.3 mbsf isinferred to represent combined Subchrons C5AA and C5AB, where-as the normal interval recognized between 550 and 559 mbsf is sug-
566

BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHIC SUMMARY
560 A
600-
800-
840 ̂
60X
65)
30
Age(Ma)
40 50 60
h (m
bs
ep
t
Q700-
•
70X
75X1
1
-
80X
5R•
10R
- j1
15R1
VΛΛΛ/
D26
N6
28 m/m.y.
-10 m/m.y.
D311 \ J R 1 3 ?A ^ D32AA\
Olig. (?) N9 \
2 m/m.y. ?
D = diatom datumR = radiolarian datumN = calcareous nannofossil datumP = planktonic foraminifer datum
BF = benthic foraminifer datumIS = isotope event
N12
m/m.y.
-upper Paleocene
16 m/m.y.
Hiatus?
N30 N33N28 / BFN l-4 I
N29 N31 N32 N34
Figure 5. Age vs. depth plot of the Maestrichtian through lower Miocene section of Site 883 after Table 3.
gested to be Subchron C5AD. These modifications are based oncalibration of diatom and radiolarian datum levels with magneto-stratigraphy at Site 887 (Barron and Gladenkov, this volume; Morleyand Nigrini, this volume), where the magnetostratigraphic record iseasier to interpret.
Shipboard measurement of inclination from the pass-throughcryogenic magnetometer (after 15 mT demagnetization) suggestedthe possibility of three normal-polarity intervals beginning at 580.0,594.2, and 602.8 mbsf; however, this part of the record was not inter-preted by Weeks et al. (this volume) because of poor data quality.Nevertheless, diatom and radiolarian datum biostratigraphy at Site887 (Barron and Gladenkov, this volume; Morley and Nigrini, thisvolume) suggests a tentative correlation of these three possible polar-ity boundaries with the tops of Chrons C5C, C5D, and C5E, respec-tively (Table 4; Fig. 6).
On Figure 6, magnetostratigraphic polarity boundaries and bio-stratigraphic datum levels (Table 4) are used to construct an age vs.depth curve for the middle Miocene to the Quaternary of Site 884. Al-though the curve is mainly plotted according to magnetostratigraphicpolarity boundaries, diatom and radiolarian datum levels in the upper
lower Miocene to middle Miocene interval between 600 and 500mbsf (Table 4) proved to be critical in identifying the various paleo-magnetic chrons. These microfossil events can be identified at Site887 where they are constrained by an excellent magnetostratigraphicrecord (Barron and Gladenkov, this volume; Morley and Nigrini, thisvolume; Weeks et al., this volume).
Calcareous nannofossil, planktonic foraminiferal, diatom, and ra-diolarian datum levels from Table 4, as well as the distinctive lateEocene oxygen isotope increase, are used in Figure 7 to plot an agevs. depth curve for the middle Eocene through lower Miocene sectionof Site 884. Sediment accumulation rates are relatively low for theolder interval of Site 884, with the exception of the upper Eocene partof the section (ca. 769 to 694 mbsf) where rates of 70 m/m.y. are sug-gested. This upper Eocene interval coincides with lithologic SubunitHB, a unit of chalk, claystone, and ash, which contains evidence oflarge-scale redeposition, including slumping and diamictic conglom-erates (Shipboard Scientific Party, 1993d). Calcareous nannofossilsand planktonic foraminifer biostratigraphy suggest that the intervalfrom Core 145-884B-87X through the greater part of Core 145-884B-83X (814-769 mbsf) lies in the middle Eocene (Beaufort and
567

J.A. BARRON ET AL.
Table 4. Age, interval, and depth of magnetostratigraphic polarity boundaries and biostratigraphic datum levels in Holes 884B, 884C, and 884E.
RlDlR2R3
D2R4R5
D3R6
D4
D5D6R7
D7
D<SD9
DIO
D11
D12
D13
D14
D15
D16
R8R9
D17
RIO
Dl<S
D19RII
1)20
R12R13
D21R14
LOLOLOLOBTLCOLOFOBFOLOTBLOTBTLCOFOFOBTLOBTBF0FOTBTBLOTBTFOBFOTFOBTF0BTBTBF0TBLCOTLOF0TBBLCOTLOBLOTTTBTBTFOLOBTLCOBLOLOTBTBF0F0TB
Datum
Lychnocanoma nipponica sakaiiSimonseniella curvirostrisStylacontarium acquiloniumStvlatractus universusCln.lnClr.lnActinocyclus oculatusEucyrtidium matuyamaiEucyrtidium matu yamaiClrlnSimonseniella curvirostrisSphaeropyle robustaC2n.lnC2n.lnNeodenticula koizumiiC2r.lnC2r.lnC2An.lnNeodenticula kamtschaticaNeodenticula seminaeCvdadophora davisianaC2An.lnC2An.2nThalassiosira marujamicaC2An.2nC2An.3nC2An.3nNeodenticula koizumiiActinocyclus oculatusC3n.ln'C3n.lnC3n.2nC3n.2nThalassiosira jacksonii (plicate)C3n.3nC3n.3nC3n.4nThalassiosira latimarginata s.str.C3n.4nThalassiosira oestrupiiC3An.lnThalassiosira praeoestrupiiC3An.lnC3An.2nThalassiosira miocenicaC3An.2nC3BnC3BnC3Br.lnC3Br.lnNeodenticula kamtschaticaC4n.lnC4n.lnThalassionema schraderiC4n.2nPrunopyle havesiStylacontarium acquiloniumC4n.2nC4r.lnC4r.lnDenticulopsis katayamaeC4An.lnLychnocanoma nipponica magnacomutaC4An.lnDenticulopsis dimorphaC4Ar.lnC4Ar.lnC4r.2nC4r.2nC5n.lnC5n.lnC5n.2nDenticulopsis dimorphaCyrtocapsella japonicaC5n.2nC5r.2nDenticulopsis praedimorphaC5r.2nCyrtocapsella cornutaLithopera renzaeC5An.lnC5An.lnC5An.2nC5An.2nSimonseniella barboiLychnocanoma nipponica magnacomutaC5Ar.lnC5Ar.ln
Age(Ma)
0.050.30.4
0.550.78
0.9841.01.02.0
1.0491.58
1.5-1.71.7571.983
1.9-2.12.1972.2292.60
2.63-2.72.72.9
3.0543.127
3.1-3.23.2213.3253.553
3.51-3.853.65-3.85
4.0334.1344.2654.432
4.5-4.64.6114.6944.812
4.8-1.95.046
5.35.7055.955.9466.078
6.26.3766.7446.9016.9466.981
7.1-7.27.2457.3767.4
7.4647.67.7
7.8928.0478.0798.4
8.5298.8
8.8619.0
9.0699.1499.4289.4919.5929.7359.7779.810.0
10.83411.37811.4
11.43411.711.7
11.85212.00012.10812.33312.412.5
12.61812.649
Core, section,interval (cm)
145-884B-
Above 1H-CC1H-CC/2H-CC2H-5, 135-136/3H-5, 135-1363H-5, 135-136/4H-5, 135-136
5H-CC/6H-CC6H-5, 135-136/7H-5, 135-13612X-1, 68-69/13X-3, 134-135
11H-CC/12X-CC10H-CC/12X-1,68-69
13X-4, 54/14X-4, 47
15X-CC/16X-CC15X-CC/16X-CC17X-5, 135-136/18X-5, 135-136
18X-CC/19X-CC
23X-5, 25/23X-CC24X-5, 25/24X-CC
29X-CC/30X-2, 25
31X-2, 25/31X-5,25
33X-2, 25/33X-5, 25
36X-5, 25/36X-CC
38X-2, 25/38X-5, 25
41X-CC/42X-2, 26
42X-5, 25/42X-CC
43X-5, 135-136/45X-5, 135-13643X-5, 135-136/45X-5, 135-136
46X-3, 25/46X-4, 25
47X-5, 135-136/48X-5, 135-136
48X-6, 25/48X-CC
50X-CC/51-2. 2751X-5, 135-136/52X-5, 135-136
56X-2, 25/56X-4, 25
56X-5, 135-136/57X-5, 137-13956X-5, 135-136/57X-5, 137-139
57X-CC/58X-2, 2557X-5, 137-139/58X-5, 135-136
Depth(mbsf)
Above 6.56.5/16.0
13.85/23.3523.35/32.85
42.554.00-55.10
44.5/54.051.85/61.3
87.98/97.6459.4
87.3/93.386.3/87.98
9
96.998.34/108.97
107.1108.2
121.30-122.50122.2/131.8122.2/131.8
139.15/148.85148.1152.35
151.0/160.8159.5-159.7
179.2196.05/199.4205.65/209.1
219.65224.2
252.4/259.05252.1?258.4?
266.3-267.2268.75/273.25276.5-276.9
288.05/292.55311.55
321.55/325.03324.8-325.2
334.5336.05/340.55
348.00364.0-364.6
367.6369.1
370.25372.7/374.46
374.2379.65
378.95/382.4381.5
389.9/409.1389.9/409.1
400.2
414.45/416.05418.45
428.3/438.0436.15
438.35/440.2440.65445.2451.1?453.1?453.5?457.3?
458.0-459.5?459.3/461.07466.7/476.2498.0-507.0
523.00509.15/512.15
526.00514.8/524.5514.8/524.5
520.70522.10523.00525.50
526.7/528.45524.5/534.1
528.50530.00
Hole 884Cdepth(mbsf)
2.4/11.911.9/21.411.9/21.221.4/30.9
41.155.0
49.9/59.4559.4/68.978.4/88.2
59.788.2/97.8
9101.9
97.8/107.4
118.50-127.00136.3/146.0136.3/146.0136.3/146.0
147.00-147.80154.50-156.00
155.7/165.3158.5-159.5
164.4178.0
194.3/204.0204.0/213.7
218.45224.5235.9244.25
261.8/271.4251.5-252.4
256.9261.8-261.9271.4/281.1
275.23281.1/290.7
311.4319.7/329.2
325.7332.4
338.7/348.2348.2
458.0
Hole 884Edepth(mbsf)
97.14102.78
107.52108.42123.98
149.57154.61
160.26167.87179.59
218.77224.71237.77246.49
254.28258.23262.96
324.37333.07
348.76366.6369.79370.87372.28
376.81381.98
385.95
398.02404.7405.14
424.97
438.75

BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHIC SUMMARY
Table 4 (continued).
D22
D23
D24D25R15RI6R17R18
D26D27R19R20R21D28
R22D29
D30D3IR23NID32D33D34N2N3ISN4N5N6N7N8PIN9P2N10NilN12
TBF0TFCOBTTFOFOFOF0LOF0TF0F0F0FOLOF0TBTF0F0BFOLOF0LOFOFCOFOLOLO
LOLOFOLOLOLOFOF0F0LOFO
Datum
C5Ar.2nC5Ar.2nDenticulopsis praedimorphaC5AAn.lnDenticulopsis hustedtiiC5ABn.ln#C5Dn.ln#C5Dn.ln#Denticulopsis hustedtiiDenticulopsis hyalinaDictyophimus splendensEucyrtidium inflatumCorythospyrisl sp.Eucyrtidium asanoiC5Cn.ln?#Denticulopsis lautaDenticulopsis praelautaTheocorys redondoensisCorythospyrisl sp.Cenosphaera sp.Crucidenticula kanavaeC5Dn.ln?#C5Dn.ln?#C5En.ln?#Cycladophora cosma cosmaCrucidenticula sawamuraeC6n.ln?#Thalassiosira fragaRocella gelidaCryptocapsella tetraperaDictyococcites bisectusThalassiosira preafragaRocella gelidaRocella vigilansReticulofenestra umbilicaIsthmolithus recurvusOxygen isotope increaseDiscoaster saipanensisDiscoaster barbadiensisIsthmolithus recurvusChiasmolithus grandisChiasmolithus solitusAcarinina bulbrookiDiscoaster bifaxAcarinina bulbrookiNannotetrina fulgensDiscoaster sublodoensisNannotetrina cristata
Age(Ma)
12.71812.764
12.812.941
13.113.47614.16414.608
14.2-14.614.9-15.1
15.315.315.315.8
16.03515.916.2
16.2516.716.716.9
17.3117.6518.31717.2518.4
20.16220.122.422.623.8
24.0-24.326.2-26.4
30.231.732.534.335.035.035.137.139.740.5
(43.7)45.2
47.0 (47.3)47.247.8
Core, section,interval (cm)
145-884B-
58X-5, 25/58X-6, 25
58X-7, 25/58X-CC
60X-CC/61X-CC62X-4, 25/62X-6, 2561X-5, 135-136/62X-5, 135-13661X-5. 135-136/62X-5, 135-13661X-5, 135-136/62X-5, 135-13662X-5, 135-136/63X-5, 135-136
63X-3, 25/63X-4, 2563X-CC/64X-1.2563X-5, 135-136/64X-4, 135-13663X-5, 135-136/64X-4, 135-13663X-5, 135-136/64X-4, 135-13664X-4, 28/64X-5, 25
64X-4, 135-135/65X-5, 135-13665X-CC/66X-1,25-26
66X-5. 25-26/66X-5, 25-2667X-3, 25-26/67X-4, 25-2665X-5, 135-136/66X-5, 134-13566X-CC/69X-5, 18-1967X-CC/68X-1,25-2669X-5, 25-26/69X-6, 25-2671X-7, 25/71X-CC70X-CC/71X-CC73X-4, 75-80/73X-5,140-14574X-7, 10-15/75X-3, 128-13374X-1, 0-5/74X-7, 10-1579X-3, 114-119/79X-5, 113-11882X-3, 107-112/82X-4, 79-8483X-1, 72-77/83X-2, 26-3183X-1.72-77/83X-2, 26-3182X-6, 60-63/86X-CC, 28-3183X-3,10-15/83X-4, 51-5686X-3, 28-33/86X-4, 25-3086X-3, 20-25/86X-4, 20-2586X-5, 65-70 /87X-4, 93-98Below 87X-6, 70-75
Depth(mbsf)
53 1.00532.50
532.75/534.25535.00
535.75/536.4542.30550.00559.00
555.6/565.3567.05/570.05
563/572.7563/572.7563/572.7
572.7/582.2580
578.05/579.55584.4/584.65582.2/590.3582.2/590.3582.2/590.3
589.15/590.65594.2599
602.8590.3/601.5
603.8/604.05610
610.05/611.55616.54/618.04601.45/611.14612.3/638.58622.8/623.05
638.65/640.15660.95/661.3651.7/661.3
676.25/678.40689.7/694.48680.6/689.7
732.94/734.43761.77/762.99768.12/769.16768.12/769.16765.8/766.98770.5/772.41799.68/801.65
799.6/801.6803.05/811.53Below 814.3
Hole 884Cdepth(mbsf)
Hole 884Edepth(mbsf)
Note: Abbreviations and ages as in Tables 1 and 3. # = paleomagnetic boundaries identified by biostratigraphic correlation with Site 887.
Olafsson, this volume; Basov, this volume). The age of the lower-most sediment interval recovered at Site 884 (Cores 145-884B-91Xthrough 145-884B-88X) is uncertain, although the presence of a re-worked specimen of Discoaster mohleri in Sample 145-884B-85X-CC suggests that upper Paleocene sediments may be either present atthe site or located nearby (Shipboard Scientific Party, 1993d).
Unconformities
A possible unconformity in the middle part of the Oligocene maybe present at about 661 mbsf in the lowermost part of Core 145-884B-7IX based on the coincidence of the LO of the calcareous nan-nofossil Reticulofenestra umbilica (31.7 Ma) and the F0 of the dia-tom Rocella vigilans (30.2 Ma; Table 4; Fig. 7). A sharp lithologiccontact is present at the 24-cm interval of Section 145-884B-71X-7,separating brown claystone above from underlying white nannofossilchalk. Diatoms decline down section in abundance and preservationat this horizon (Gladenkov and Barron, this volume). Seismic reflec-tion data reveal a reflector of moderate amplitude and continuity atthis horizon (Hamilton, this volume), but major differences in physi-cal properties are not apparent (Shipboard Scientific Party, 1993d).Unfortunately, the geochemical logging tool string did not penetrateto this depth because a bridge developed in Hole 884E at 654 mbsf.The magnetic susceptibility log, however, shows a peak betweenabout 660 and 650 mbsf (Shipboard Scientific Party, 1993d), sug-gesting a disruption in the sedimentation style.
Benthic foraminifers suggest the possibility of an unconformity atthe Eocene/Oligocene boundary (Pak and Miller, this volume) or atabout 695 mbsf, coinciding with the break between lithologic Sub-units IIA and HB (Shipboard Scientific Party, 1993d). Although theselithologic units are both claystone mixed with nannofossil chalk, ev-idence for soft-sediment deformation and large-scale redeposition iscommon in Subunit HB, while such evidence is much less common-place in Subunit IIA (Shipboard Scientific Party, 1993d). Althoughno hiatus is apparent in the age vs. depth plot (Fig. 7), an abrupt up-section decrease in the sediment accumulation rate from 70 m/m.y. to12 m/m.y. is present at the Eocene/Oligocene boundary that maymark the onset of the Meiji drift sediments (Rea et al., this volume).
A third possible unconformity at about 769 mbsf in the middlepart of Core 884B-83X separates middle Eocene sediments from up-per Eocene sediments (ca. 42-35.5 Ma; Fig. 7). It is marked by theboundary between lithologic Subunit HC, claystone, altered clayeyash, and ash, and overlying Subunit HB, which is made up of chalk,claystone, and minor ash (Shipboard Scientific Party, 1993d). Soft-sediment deformation and evidence of large-scale redeposition,which typifies Subunit HB, decreases abruptly down section in Sub-unit HC, suggesting a change in the style of sedimentation (ShipboardScientific Party, 1993d). At the depth of this contact, seismic reflec-tion data reveal a reflector that separates a unit of flat-lying reflectorswith moderate amplitudes from an overlying unit characterized bydipping, generally discontinuous reflectors with moderate to strongamplitudes (Hamilton, this volume). The age and duration of this un-

J.A. BARRON ET AL.
2 3
Age (Ma)
7 8 10 12 13 15 16
+ = paleomagnetic eventD = diatom datumR = radiolarian datum
R15,R16,R17 D26
D27
R18 R19
Figure 6. Age vs. depth plot of the middle Miocene through Quaternary section of Site 884 after Table 4.
conformity closely matches that of an unconformity at 710 mbsf inHole 883B (42-34 Ma) and seems to indicate a major change in sed-imentation style.
Site 887
Four holes were drilled at Site 887 (54°21.9'N, 148°26.8'W, 3645m water) on the Patton-Murray Seamount in the northeast Pacific(Shipboard Scientific Party, 1993e). At Site 887, about 285 m of low-er Miocene to Quaternary sediments overlie basalt encountered inCore 145-887D-4R. Below this interval a thin layer of sediment isfound between basalt flows in Sample 145-887D-6R-1, 10-15 cm,which contains calcareous nannofossils indicative of an undifferenti-ated Oligocene age. Calcareous microfossils (foraminifers and cal-careous nannofossils) range from barren to abundant and aremoderately well to poorly preserved. The low diversity of the calcar-eous microfossil assemblages prevents the assignment of biostrati-graphic zones to the samples. Siliceous microfossils (diatoms,radiolarians, and silicoflagellates), on the other hand, are more com-mon in the sediments from Site 887 and are typically well preservedto moderately well preserved, but persistent reworking of middle Mi-ocene diatoms and radiolarians is present in the uppermost Mioceneand Pliocene part of the section.
High-quality paleomagnetic stratigraphy at Site 887 extends with-out interruption back to magnetic-polarity Subchron C5En (Table 5,Fig. 8), and the identification of the various paleomagnetic chrons isin good agreement between the biostratigraphers (Barron and Glad-
enkov, this volume; Morley and Nigrini, this volume) and the inde-pendent suggestions of the paleomagneticists (Weeks et al., thisvolume). The interval and subbottom depths of magnetostratigraphicpolarity boundaries and biostratigraphic events are listed for Holes887A and 887C in Table 5. An age vs. depth curve for Site 887 is con-structed in Figure 8, mainly from magnetostratigraphy.
Unconformity
The middle Miocene Crucidenticula nicobarica Zone may bemissing at an unconformity near 245 mbsf in Hole 887A. Sample145-887A-27H-3, 20 cm (244.4 mbsf) is assignable to the D. praed-imorpha Zone (11.4-12.8 Ma), whereas Sample 145-887A-27H-3,124 cm (245.44 mbsf) belongs to the Denticulopsis hyalina Zone(13.1-15.1 Ma). No lithologic evidence, however, is apparent for anunconformity in this interval (Shipboard Scientific Party, 1993e). Ifpresent, such a hiatus would coincide with the early part of a majorhiatus at Site 883 (Fig. 4).
CONCLUSIONS
Biochronology
The Pliocene and Pleistocene magnetostratigraphic records ofSites 881-884 and 887 (Figs. 2-4, 6, 8) provide a framework for eval-uation of the isochroneity and diachroneity of microfossil events. Asmany as eleven diatom, seven radiolarian, and five calcareous nanno-
570

BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHIC SUMMARY
Age (Ma)
600-
650-
700
Q.03Q
750-
800-
30
+ = paleomagnetic event
D = diatom datum
R = radiolarian datum
N = calcareous nannofossil datumP = planktonic foraminifer datumIS = isotope event
12 m/m.y.
40 50
70 m/m.y.?
P1
AAAVVAAAAΛ^ΛAAAAΛ.
N7 N8 " N9
IP2
'7 m/m.y.
\N10
N12
Figure 7. Age vs. depth plot of the middle Eocene through lower Miocene section of Site 884 after Table 4.
571

J.A. BARRON ET AL.
Table 5. Age, interval, and depth of magnetostratigraphic polarity boundaries and biostratigraphic datum levels in Holes 887A and 887C.
DatumAge(Ma)
Core, section,interval (cm)145-887A-
Depth(mbsf)
Core, section,interval (cm)
145-887C-Depth(mbsf)
RlNIDlR2R3
R41)2
D3N2R5
D4
D5D6R6
D7
D8D9
D10
D11
D12
D13
D14D15DI6
R7R8R9
D17
RIO
D18
DI9RH
D20R12
D21RI3
D22D23
LOF0LOLOLOBTLOLCOBF0FOLOTBLOFOTLCOF0FOBTBLOTBF0F0TBBTBLOTFOBFOTBTFOBFOFOLCOTFOLOFOBTLCOTBTLO
BLOTBTBTFOLOBTLCOLOTBTBFOFO
TBTBFOFCOBTB
Lychnocanoma nipponica sakaiiEmiliania huxleyiSimonseniella curvirostrisStylacontarium acquiloniumStvlatractus universusCln.lnClr.lnEucyrtidium matuyamaiActinocyclus oculatusC l r . l nSimonseniella curvirostrisGephyrocapsa oceanicaSphaeropyle robustaC2n.lnC2n.lnNeodenticula koizumiiEucvrtidium matuyamaiC2An.lnNeodenticula kamtschaticaNeodenticula seminaeCycladophora davisiana davisianaC2An.lnC2An.2nC2An.2nThalassiosira marujamicaC2An.3nC2An.3nActinocyclus oculatusNeodenticula koizumiiC3n.lnC3n.lnC3n.2nC3n.3nC3n.3nThalassiosira jacksonii (plicate)C3n.4nThalassiosira latimarginataC3n.4nThalassiosira oestrupiiC3An.lnC3An.lnC3An.2nThalassiosira miocenicaC4n.lnNeodenticula kamtschaticaNitzschia reinholdiiThalassionema schraderiC4n.2nS. delmontensis > S. peregrinaPrunopyle hayesiStylacontarium acquiloniumC4n.2nC4r.lnDenticulopsis hustedtiiC4r.2r-1C4r.2r-1C4An.lnLychnocanoma nipponica
Hl&Q'HCIC'OfflUtCl
C4An.lnDenticulopsis dimorphaC4r.2nC4Ar.2nC5n.lnC5n.lnC5n.2nDenticulopsis dimorphaCyrtocapsella japonicaC5n.2nC5r.2nDenticulopsis praedimorphaCyrtocapsella cornutaC5An.lnC5An.lnC5An.2nC5An.2nSimonseniella barboiLychnocanoma nipponica
magnacornutaC5Ar.lnC5Ar.lnC5Ar.2nC5Ar.2nDenticulopsis praedimorphaDenticulopsis hustedtiiC5ABnC5ACn.lnC5ACn.ln
0.050.2940.30.40.550.78
0.9841.0LO
1.0491.581.674
1.5-1.71.7571.9832.02.02.60
2.63-2.72.72.9
3.0543.1273.221
3.1-3.23.3253.553
3.65-3.853.51-3.85
4.0334.1344.4324.6114.694
4.5^1.64.812
4.8^.95.046
5.35.7055.9466.0786.2
7.3767.1-7.27.2-7.3
7.47.4647.557.67.7
7.8928.0478.4
8.4638.4798.529
8.8
8.8619.0
9.4289.4919.5929.7359.7779.810.0
10.83411.378
11.411.7
11.85212.00
12.10812.333
12.412.5
12.61812.64912.71812.764
12.813.1
13.47613.67414.059
lH-1, 12-13/1H-3, 52-531H-3, 110-111/1H-4, 110-1112H-CC/3H-CC3H-3, 132-133/4H-1, 12-134H-1, 12-13/4H-3, 132-133
7H-3, 54-55/7H-6, 54-556H-CC/7H-CC
7H-CC/8H-CC5H-2, 110-111/5H-6, 110-1118H-3, 54-55/8H-6, 54-55
8H-CC/9H-CC9H-3, 54-55/1OH-3, 54-55
9H-CC/10H-CC9H-CC/10H-CC11H-3, 54-55/12H-3, 54-55
12H-2, 135/12H-3, 5
13H-1, 21/13H-2. 2113H-2, 21/13H-3, 21
16X-CC/17X-1,21
17X-3, 21/17X-4, 21
17X-CC/18H-1,21
21X-1, 22/21X-2, 22
22H-4, 21/22H-5. 2122H-5, 21/22H-6, 2122H-6, 21/22H-CC
22H-6, 54-55/23H-6, 54-5522H-6. 54-55/23H-6, 54-5522H-6, 54-55/23H-6, 54-55
23H-7, 21/23H-CC
23H-6, 54-55/24H-3, 54-55
24H-4, 21/24H-5, 21
25H-3, 133/25H-4, 13324H-6, 54-55/25H-3, 54-55
25H-CC/27H-l,6026H-6, 55-56/27H-3, 55-56
25H-CC/27H-1.60
27H-3, 20/27H-3, 12427H-3, 20/27H-3, 124
0.12/3.524.10/5.6016.2/25.7
20.52/25.8225.82/30.02
45.5555.6
57.74/62.2454.2/63.7
58.9563.7/73.2
37.80/43.8067.24/71.74
73.2/82.776.74/86.24
88.982.7/92.282.7/92.2
95.74/105.24101.1
101.85104.2
104.55/104.75106.55110.35
111.4/112.9112.9/114.4
121.1125.2134.7
142.2144.37/145.3
145.8148.3/149.8
149.95154.8/155.0
183.9/185.4
198.1/199.6199.6/201.1201.1/202.9
201.44/210.94201.44/210.94201.44/210.94
212.1/212.4
210.94/215.94
217.1/218.6
226.23/227.73220.44/225.44
231.4/241.82234.35/239.35
231.4/241.82
244.4/245.44244.4/245.442
6H-6, 21/26H-7, 21
7H-3, 21/27H-6, 2127H-3, 55-56/27H-6, 55-56
27H-6, 21/27H-CC7H-6, 21/27H-CC
42.752.6
56.25
75.3578.95
87.3
98.3100.55102.7
103.4105.9
120.35
134.8139.65
143.55
148.65
169-169.3177.1
181.75
199.2
199.8
205.45206.8
211.25211.65211.8
215.95
219.75220.65221.3
222.85223.05
231.45235.15
234.0/235.5
238.25239.5
240.05241.65
239.0/243.5239.35/243.85
243.05243.25243.45243.75
243.57245.324.57245.3
247.25248.45250.85
572

BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHY SUMMARY
Table 5 (continued).
D24
R14R15R16R17
R1SR19R20D25R21
D26D27
TTBF0TBF0F0LOFOTF0F0LOF0FOTBTFOF0
Datum
C5ADn.lnC5Bn.lnC5Bn.lnDenticulopsis hyalinaC5Bn.2nC5Bn.2nDictyophimus splendensEucyrtidium inflatumCorythospyrisl sp.Eucyrtidium asanoiC5Cn.lnTheocorys redondoensisCorythospyrisl sp.Cenosphaera sp.Crucidenticula kanayaeCycladophora cosma cosmaC5Dn.lnC5Dn.lnC5En. 1 nCrucidenticula sawamuraeThalassiosirafraga
Age(Ma)
14.16414.8
14.8914.9
15.03815.162
15.315.315.315.8
16.03516.2516.716.716.9
17.2517.3117.65
18.31718.420.1
Core, section,interval (cm)
145-887A-
29H-1.51/29H-2, 21
28H-6, 55-56/29H-3, 54-5528H-6, 55-56/29H-3, 54-5528H-6, 55-56/29H-3, 54-5528H-6, 55-56/29H-3, 54-55
28H-6, 55-56/29H-3 54-55Below 29H-3, 54-55Below 29H-3, 54-55Below 29H-3, 54-55Below 29H-3, 54-55
Below 30X-CC
Depth(mbsf)
260.7/261.9
258.75/263.74258.75/263.74258.75/263.74258.75/263.74
258.75/263.74Below 263.74Below 263.74Below 263.74Below 263.74
Below 279.4
Core, section,interval (cm)
145-887C-
Above 3OH-3, 54-55Above 3OH-3, 54-5530H-1.21/30H-2, 21Above 3OH-3, 54-55
30H-6, 21/30H-7, 21Below 30H-CC
Depth(mbsf)
251.75255.6
256.25
257.05257.55
262.1
Above 267.84Above 267.84264.51/266.01Above 267.84
266.75268.4
271.95272.0/273.7Below 273.8
Note: Abbreviations as in Tables I and 3.
fossil events appear to be isochronous for correlation of Pliocene andPleistocene sediments across the middle to high latitudes in the NorthPacific (Tables 1-5). Calcareous nannofossils are useful for bio-stratigraphy only at the sites with shallower water depths (Site 882,883, and 887), and they are very sparse within the thick diatomaceoussections that characterize the lower and lower upper Pliocene. Plank-tonic foraminiferal biostratigraphic studies on the Pliocene and Pleis-tocene sediments of Leg 145 are limited to a study of a very shortinterval of the middle part of the Pliocene of Sites 883 and 887 byDowsett and Ishman (this volume).
A major contribution of Leg 145 is the recovery of the first con-tinuous magnetostratigraphic records in the North Pacific for the up-per lower Miocene through the upper Miocene (magnetic polaritySubchrons C5En through C3An) section at Sites 884 and 887 (Figs.6, 8). These magnetostratigraphic records allow the first direct cali-bration of numerous diatom and radiolarian datum levels to magne-tostratigraphy and establish the isochroneity (or diachroneity) ofthese microfossil events between the northwest and northeast Pacific(Barron and Gladenkov, this volume; Morley and Nigrini, this vol-ume). Within the limits of available stratigraphy, at least fifteen dia-tom and fifteen radiolarian datum levels appear to be isochronous forMiocene correlations between Sites 884 and 887 (Tables 4, 5). Un-fortunately, neither calcareous nannofossils nor planktonic foramini-fers are present in sufficient numbers or diversity in the Miocenesections of Sites 884 and 887 to allow calibration to magnetic stratig-raphy (ólafsson and Beaufort, this volume; Shipboard Scientific Par-ty, 1993d, 1993e).
Below the upper part of the lower Miocene section, microfossilsprovide the only means of assigning ages to the sediment record ofLeg 145. Calcareous nannofossils are the most useful microfossil forcorrelations, with as many as 24 biostratigraphically important datumlevels recognized in the Upper Cretaceous through upper lower Mi-ocene section of Site 883 (Table 3; ólafsson and Beaufort, this vol-ume; Beaufort and ólafsson, this volume). Planktonic foraminifersprovide valuable correlations in the lower and middle Eocene sec-tions, especially of Site 883 (Basov, this volume). Gladenkov andBarron (this volume) propose an Oligocene through lower Miocenediatom zonation based on their study of Site 884, whereas Shilov (thisvolume) documents the Eocene through early Miocene radiolarianassemblages at Sites 883 and 884 and suggests a preliminary zona-tion.
The Sediment Record
Although the age vs. depth plots reveal that the Pliocene andPleistocene record is complete at Sites 881-884 and 887 (Figs. 2-4,6, 8), sediment accumulation rates vary considerably. Although thicksequences of diatomaceous sediment with sedimentation rates in ex-cess of 100 m/m.y. characterize much of the Pliocene (ca. 4.4 to 2.6Ma) of Sites 882 and 883 (Figs. 3, 4), this same interval is marked byrates (ca. 33 m/m.y.) that are reduced compared to those of the post-2.6-Ma Pliocene and Pleistocene record at Site 881 (Fig. 2). Thesedifferences clearly reflect both the better preservation of carbonateand smaller, more finely silicified diatom taxa at Sites 882 and 883on the top of Detroit Seamount (Haug et al., this volume) and an in-crease in the amount of abyssal transport in the younger record ofdeeper Site 881 (Rea et al., this volume).
The upper Miocene section is generally complete at Sites 883,884, and 887, but two unconformities, (ca. 7.7-7.1 Ma) and (6.1-5.3Ma), appear to be present at Site 881. The older of these two hiatusesappears to correlate with hiatus NH6 of Keller and Barron (1987)(7.7-6.5 Ma updated age), which is widespread in the North Pacificand appears to reflect an event of intensification of bottom currents.The younger hiatus, on the other hand, is slightly older than hiatusNH7 of Keller and Barron (1987), which has an updated age of 5.5 to5.0 Ma.
With the exception of a compressed interval of the Denticulopsispraedimorpha diatom Zone (12.8-11.4 Ma), the lowermost part ofthe upper Miocene section and the entire upper middle Miocene sec-tion (14.2-10.2 Ma) is removed at one or more unconformities be-tween about 556 and 547 mbsf at Site 883. At Site 884 on the eastflank of Detroit Seamount, this 14.2-10.2-Ma interval is largely rep-resented by lithologic Subunit IC, a dark-gray to gray claystone,which contains reduced numbers of diatoms and radiolarians (>20%opal) (Shipboard Scientific Party, 1993d). A series of high-frequencydiscontinuous seismic reflectors in this interval of Site 884 (Hamil-ton, this volume) may be suggestive of the intensification of bottomcurrents. At Site 887 in the northwest Pacific, this late middle Mi-ocene unconformity, if present, is restricted to the Crucidenticulanicobarica diatom Zone or from about 13.1-12.8 Ma (Fig. 8). Kellerand Barron (1987) note the widespread occurrence of hiatuses NH3and NH4 in the North Pacific, which correspond, respectively, withthe intervals ranging from 13.0 to 12.1 Ma and 10.9-10.1 Ma.
573

J.A. BARRON ET AL.
+ = paleomagnetic eventD = diatom datumR = radiolarian datum
possibleunconformity13.1-12.8 Ma
300-
* = 887D
Figure 8. Age vs. depth plot of Site 887 after Table 5.
Extensive records of the Paleogene and lower Miocene are re-stricted to Sites 883 and 884. Whereas Site 884 appears to have themore complete upper Eocene through lower Miocene record of thetwo sites (Fig. 7), the lower Eocene through middle Eocene recordappears to be more complete at Site 883 (Fig. 5), which also is char-acterized by better carbonate preservation. In addition, much of theEocene record of Site 884 is chaotic and apparently redeposited(Shipboard Scientific Party, 1993d; Beaufort and Olafsson, this vol-ume). Both Sites 883 and 884 contain major unconformities at themiddle Eocene/upper Eocene boundary, which appear to commenceat about 42 Ma, the age of the major change in the motion of the Pa-cific plate that is associated with the bend in the Emperor SeamountChain. This major hiatus continues to about 34 Ma (latest Eocene) inage at Site 883 (Fig. 5), but apparently ceases slightly earlier (at ca.35.5 Ma; late Eocene) at Site 884.
Paleocene sediments are limited to a thin section in the lower partof Core 145-883B-85X and the upper part of Core 145-883E-19R(Beaufort and Olafsson, this volume; Pak and Miller, this volume)where a compressed section of upper Paleocene sediments uncon-formably overlies a thin sequence of nannofossil-bearing ashes ofLate Cretaceous age at the base of the Site 883 sediment section. Cal-careous nannofossils in Cores 145-883B-86X and 145-883B-87X ap-pear to be Maestrichtian in age (Beaufort and Olafsson, this volume).
ACKNOWLEDGMENTS
This manuscript benefited from the reviews of Bill Sliter, KrisMcDougall, Tim Bralower, and David Harwood and we are gratefulfor their suggestions. The research of various scientists was funded
574

BIOSTRATIGRAPHIC AND MAGNETOSTRATIGRAPHY SUMMARY
by the various programs and grants listed in the acknowledgments oftheir individual chapters. We are grateful to David Rea and the otherscientists and crew members of Leg 145 of the JOIDES Resolutionfor their support and encouragement.
REFERENCES*
Aubry, M.-P., Berggren, W.A., Kent, D.V., Flynn, J.J., Klitgord, K.D., Obra-dovich, J.D., and Prothero, D.R., 1988. Paleogene geochronology: anintegrated approach. Paleoceanography, 3:707-742.
Barron, J.A., 1989. The late Cenozoic stratigraphic record and hiatuses of thenortheast Pacific: results from the Deep Sea Drilling Project. In Winterer,E.L., Hussong, D.M., and Decker, R.W. (Eds.), The Geology of NorthAmerica (Vol. N): The Eastern Pacific Ocean and Hawaii. Geol. Soc.Am., Geol. of North America Ser., 311-322.
Berggren, W.A., Kent, D.V., and Flynn, J J., 1985a. Jurassic to Paleogene,Part 2. Paleogene geochronology and chronostratigraphy. In Snelling,NJ. (Ed.), The Chronology of the Geological Record. Geol. Soc. LondonMem., 10:141-195.
Berggren, W.A., Kent, D.V., and Van Couvering, J.A., 1985b. The Neogene,Part 2. Neogene geochronology and chronostratigraphy. In Snelling, NJ.(Ed.), The Chronology of the Geological Record. Geol. Soc. LondonMem., 10:211-260.
Cande, S.C., and Kent, D.V., 1992. A new geomagnetic polarity time scalefor the Late Cretaceous and Cenozoic. J. Geophys. Res., 97:13917—13951.
Keller, G., and Barron, J.A., 1987. Paleodepth distribution of Neogene deep-sea hiatuses. Paleoceanography, 2:697-713.
Leroux, S., 1993. Susceptibilité magnétique de sediments: comparaison entreles diagraphies et les mesures sur carotte-intérët. Magnétostratigraphiesur carottes de forages (ODP Leg 145, Puits 883B). Mém. DEA, EcoleNormale Supérieure.
Rea, D.K., Basov, LA., Janecek, T.R., Palmer-Julson, A., et al., 1993. Proc.ODP, Init. Repts., 145: College Station, TX (Ocean Drilling Program).
Shipboard Scientific Party, 1993a. Site 881. In Rea, D.K., Basov, I.A., Jan-ecek, T.R., Palmer-Julson, A., et al., Proc. ODP, Init. Repts., 145: Col-lege Station, TX (Ocean Drilling Program), 37-83.
, 1993b. Site 882. In Rea, D.K., Basov, LA., Janecek, T.R.,Palmer-Julson, A., et al., Proc. ODP, Init. Repts., 145: College Station,TX (Ocean Drilling Program), 85-119.
-, 1993c. Site 883. In Rea, D.K., Basov, I.A., Janecek, T.R., Palmer-Julson, A., et al., Proc. ODP, Init. Repts., 145: College Station, TX(Ocean Drilling Program), 121-208.
-, 1993d. Site 884. In Rea, D.K., Basov, I.A., Janecek, T.R.,Palmer-Julson, A., et al., Proc. ODP, Init. Repts., 145: College Station,TX (Ocean Drilling Program), 209-302.
-, 1993e. Site 887. In Rea, D.K., Basov, I.A., Janecek, T.R., Palmer-Julson, A., et al., Proc. ODP, Init. Repts., 145: College Station, TX(Ocean Drilling Program), 335-391.
Wei, W., and Peleo-Alampay, A., 1993. Updated Cenozoic nannofossil mag-netobiochronology. INA Newsl., 15:15-21.
Zijderveld, J.D.A., Zachariasse, W.J., Verhallen, P.J.J.M., and Hilgen, FJ.,1986. The age of the Miocene-Pliocene boundary. Newsl. Stratigr.,16:169-181.
*Abbreviations for names of organizations and publications in ODP reference lists fol-low the style given in Chemical Abstracts Service Source Index (published byAmerican Chemical Society).
Date of initial receipt: 29 September 1994Date of acceptance: 23 January 1995Ms 145SR-145
575





























